КОМЕНТАРИ НА YBOP: Ефектът Coolidge стои зад силата на интернет порно. Ефектът на Coolidge е явление, наблюдавано при видове бозайници, при което мъжете (и в по-малка степен женските) проявяват подновен сексуален интерес, ако бъдат представени на нови възприемчиви сексуални партньори, дори след отказ от секс от предишни, но все още налични сексуални партньори. Сексуалната новост отменя това привикване с подновено вълнение, причинено от по-високия допамин. Непрекъснат поток от новости е това, което прави интернет порно толкова различно от порно от миналото.
- Денис Ф. Фиорино,
- Ариана Кури и
- Антъни Г. Филипс
- The Journal of Neuroscience, 15 юни 1997 г., 17 (12): 4849-4855;
абстрактен
Ефектът на Кулидж описва повторното стартиране на сексуално поведение в „сексуално наситено“ животно в отговор на нов рецептивен партньор. Като се има предвид ролята на мезолимбичната система на допамин (DA) при инициирането и поддържането на мотивирано поведение, микродиализата се използва за наблюдение на предаването на nucleus accumbens (NAC) DA по време на копулацията, сексуалното насищане и повторното създаване на сексуално поведение. В съгласие с по-ранните доклади, представянето на естросната женска зад екрана и копулацията са свързани със значително увеличаване на изтичането на NAC DA. Връщането на NAC DA концентрации към изходните стойности съвпадна с период на сексуална ситост, въпреки че концентрациите на DA метаболитите, дихидроксифенилоцетната киселина и хомованиловата киселина остават повишени. Представянето на нова рецептивна жена зад екрана доведе до леко повишаване на NAC DA, което бе значително увеличено по време на повторното копулация с новата жена. Настоящите данни предполагат, че стимулационните свойства на нова рецептивна жена могат да служат за увеличаване на предаването на NAC DA при сексуално наситен мъжки плъх, а това на свой ред може да бъде свързано с повторното иницииране на сексуално поведение.
Въведение
Плъх от мъжки пол, който е копулирал до пресищане, може да бъде индуциран да се чифтоса отново, ако първоначалната жена е заменена с нова рецептивна жена. Това стана известно като ефект на Кулидж и се наблюдава в редица видове бозайници (Wilson et al., 1963). Общи фактори като умора или моторна депресия не са достатъчни, за да обяснят очевидното състояние на сексуално насищане, тъй като стимулите от нова женска все още могат да предизвикат копулация. Сексуалното пресищане също може да бъде "обърнато" фармакологично, до значителна степен, чрез администрирането на различни лекарства, които могат да действат върху различни невротрансмитерни системи. Тези лекарства включват йохимбин, 8-OH-DPAT (Родригес-Манзо и Фернандес-Гуасти, 1994, 1995a), налаксон (Pfaus и Gorzalka, 1987; Родригес-Манзо и Фернандес-Гуасти, 1995a,b) и апоморфин (Mas et al., 1995c). Въпреки че периферното действие на тези лекарства не може да бъде изключено (напр. Адренергични ефекти върху еректилната функция), въздействието върху централните механизми, лежащи в основата на сексуалната ситост, са предложени на базата на селективни експерименти с централна норадренергична лезия (Родригес-Манзо и Фернандес-Гуасти, 1995a) и експерименти с микродиализа, които наблюдават допаминергичния метаболизъм в медиалната преоптична област (Mas et al., 1995a,b).
Като се има предвид, че централните механизми могат да посредничат при повторното иницииране на сексуално поведение, характерно за ефекта на Кулидж, вероятният кандидат е мезолимбичната система на допамин (DA), проектираща се от вентралната част до NAC. Мезолимбичният ДД изглежда действа като основен модулатор в сложни интегративни процеси, които включват оценка на въздействията на околната среда, като сигнали от сексуално възприемчива жена, и организацията на целенасочено поведение, включително копулация (Fibiger и Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Въпреки че невроните на средния мозък DA реагират на първични награди и сигнали за предсказване на наградата, нови или непредсказуеми стимули на околната среда предизвикват най-силно невронално активиране при многократни тренировки (Fabre et al., 1983; Шулц, 1992; Mirenowicz и Schultz, 1994). Има много доказателства, които подкрепят важна улесняваща роля на мезолимбичния DA при започване и поддържане на сексуалното поведение на плъхове (Pfaus и Everitt, 1995), а редица изследвания за микродиализа докладват за увеличаване на изтичането на NAC DA по време на апетитни и консуматорски фази на мъжкото сексуално поведение (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Има, обаче, сравнително малко данни за неврохимичните корелати на сексуалното насилие и повторното въвеждане на сексуалното поведение. Прилагането на ин виво микродиализа за мониторинг на мезолимбичната DA невротрансмисия по време на ефекта на Coolidge предоставя уникална възможност да се изследва ролята на NAC DA в копулацията, сексуалното насищане и повторното въвеждане на копулация.
Извършен е експеримент за микродиализа, за да се определи следното: (1) дали началото на сексуалната ситост е придружено от връщане на екстрацелуларните концентрации на DA в NAC до стойности за пре-пулпулация или по-долу, и (2) дали възстановяването на копулаторно поведение в a сексуално наситен мъжки плъх с нова рецептивна жена корелира с повишаване на изтичането на NAC DA.
МАТЕРИАЛИ И МЕТОДИ
Индивиди. Мъжки плъхове Sprague Dawley, получени от Центъра за грижа за животните (в Университета на Британска Колумбия), и женски плъхове Long-Evans, получени от Charles River Canada (St. Constant, Quebec, Canada), бяха настанени в клетки от телени мрежи (18) × 25 × 65 cm; пет на клетка) в отделни стаи за колонии. Стаите с колонии се поддържат при температура от ~ 20 ° C на обратен цикъл светлина / тъмнина 12 hr. Плъховете имат неограничен достъп до храна (Purina Rat Chow) и вода.
Хирургия и поведенчески тестове преди микродиализа в мозъка.Женски плъхове са били овариектомирани двустранно при галотанова газова анестезия (Fluothane, Ayerst Laboratories) поне 4 седмици преди тестване. Сексуалната възприемчивост в женските стимулира се индуцира чрез подкожни инжекции от естрадиол бензоат (10 μg) и прогестерон (500 μg), 48 и 4 hr, съответно, преди всяко изпитване. Мъжки плъхове се изследват за сексуално поведение два пъти, 4 d, в плексигласови камери (35 × 35 × 40 cm) с телени подове. Само мъжки плъхове, които са достигнали критерий за ефективност, които включват интро-мисия в рамките на 5 min от представянето на женската и еякулацията в рамките на 15 min от първата интронизация, по време на двата скрининг теста са имплантирани с микродиализна сонда с водещи канюли.
Мъжки плъхове (n = 5) се анестезират с кетамин хидрохлорид (100 mg / kg, ip) и ксилазин (10 mg / kg, ip) преди стереотаксична хирургия. Водната канюла с микродиализна сонда (габарит 19) се имплантира двустранно през NAC (координати от bregma: anterior, + 1.7 mm; медиална, ± 1.1 mm; вентрална, -1.0 mm; плосък череп) и се закрепват към черепа с дентален акрил и винтове за бижута. Използват се двустранни импланти с водещи канюли, за да се максимизира възможността за успешен експеримент за микродиализа. За щастие, в настоящия експеримент е необходима само една канюла за всеки плъх. Мъжки плъхове се настаняват индивидуално в големи пластмасови клетки с царевично покритие за остатъка от експеримента. Една седмица след операцията, плъховете бяха тествани за сексуално поведение. По време на тази част от обучението, тестовата камера беше оборудвана с плъзгащ се плексигласов екран, който разделяше камерата на големи и малки отделения. Мъжки плъхове се въвеждат в голямото отделение и 15 min по-късно, женската се поставя зад екрана. След 15 min подготвителен период, екранът се отстранява и плъховете се оставят да копулират за 30 min. Бяха проведени три обучителни сесии, по един на всеки 4 d. Всички плъхове достигат критерия за изпълнение по време на всяка сесия.
Експеримент с ефект на Кулидж. Плъховете се имплантират едностранно с микродиализни сонди 12 – 18 часа преди експеримента с ефекта на Coolidge и се поставят в голямото отделение на изпитвателната камера със свободен достъп до храна и вода. На сутринта на експеримента се събират проби от микродиализа всеки 15 min. Експериментът се състои от следните седем последователни фази: (1) базова линия (поне 60 min); (2) женски 1 зад екрана (15 min); (3) копулация с женски 1 до изтичане на 30 мин. Без монтиране; (4) реинтродукция на женски 1 зад екрана (15 min); (5) достъп до женски 1 за 15 min период, при условие че не е имало монтаж (ако е станало монтиране, тази фаза се третира като фаза 3); (6) въвеждане на женски 2 зад екрана (15 min); 7) копулация с женски 2 за 60 min.
Поведението се заснема при ниска осветеност с помощта на JVC видео система и се наблюдава на видео монитор, разположен извън помещението за тестване. Стандартните мерки за сексуално поведение бяха записани с помощта на компютър и подходящ софтуер (Holmes et al., 1987).
След експеримента за микродиализа, на животните се дава свръхдоза от хлоралхидрат и се перфузират интракардиално с физиологичен разтвор и формалин (4%). Мозъците бяха нарязани и замразени и впоследствие коронарните срезове бяха оцветени с крезил виолет, за да се определи поставянето на микродиализни сонди. Само плъхове с разположение на пробата в NAC бяха използвани за поведенчески и неврохимични анализи.
Микродиализа и HPLC-електрохимично откриване. Микродиализните сонди са концентрични в дизайна с полупропусклива мембрана от кухи влакна (2 mm мембрана, 340 μm външен диаметър, 65000 отрязък на молекулно тегло, Filtral 12, Hospal) на дисталния край. Сондите бяха перфузирани при 1.0 μl / min с модифициран разтвор на Ringer (0.01 m натриев фосфатен буфер, рН 7.4, 1.3 mmCaCl23.0 mm KCl, 1.0 mmMgCl2147 mm NaCl), като се използва газова неръждаема спринцовка (Hamilton, Reno, NV) и помпа за спринцовка (модел 22, Harvard Apparatus, South Natick, МА). Използва се направляващ пръстен с микродиализна сонда за закрепване на микродиализната сонда вътре в водещата канюла. За защита на тръбата на сондата е използвана стоманена бобина, прикрепена към въртяща се течност (Instech 375s), която е монтирана на върха на изпитвателната камера (Fiorino et al., 1993).
Микродиализатни аналити, които включват DA и неговите метаболити дихироксифенилоцетна киселина (DOPAC) и хомованилична киселина (HVA), се разделят чрез хроматография с обратна фаза (колона Ultrasphere; Beckman, Fullerton, СА, ODS 5 μm, 15 cm, 4.6 mm, вътрешен диаметър) ) като се използва 0.083m натриев ацетатен буфер, рН 3.5 (5% метанол). Концентрациите на аналитите се определят количествено чрез електрохимично (EC) откриване. Апаратът се състои от Bio-Rad (Richmond, CA) помпа, Valco Instruments (Хюстън, TX) EC10W двупозиционен инжектор, ESA (Bedford, MA) Coulochem II EC детектор и двуканален рекордер за диаграма (Kipp). и Zonen, Бохемия, Ню Йорк). Параметрите на електрохимичния детектор са следните: електрод 1, + 450 mV; електрод 2, -300 mV; и охранителна клетка, -450 mV. Проведени са типични възстановявания на сонда витро и при стайна температура, бяха 22% за DA, 18% за DOPAC и 18% за HVA.
РЕЗУЛТАТИ
Поведение
Поведенческите мерки от експеримента на Кулидж ефекта са представени в Таблица 1, Закъсненията при монтиране, интромит и еякулат, както и пост-целулиращия интервал след първата еякулация са подобни на тези в предишното обучение (данните не са показани). Това предполага, че процедурата по микродиализа не променя нормалното сексуално поведение. Развитието на сексуално насищане, посочено от средния брой еякулации, преди да бъде изпълнен критерият (7.8 ± 0.5), прогресивно намаляване на броя на интродукциите, предшестващи всяка еякулация, и прогресивно увеличаване на пост-целулиращия интервал (данните не са показани) , е подобен на този, докладван в предишни проучвания (Плаж и Йордания, 1956; Фаулър и Уолън, 1961; Fisher, 1962; Bermant et al., 1966; Родригес-Манзо и Фернандес-Гуасти, 1994; Mas et al., 1995d). Наблюдавана е индивидуална вариабилност по отношение на броя на еякулациите, постигнати с женски 1, времето, прекарано в копулиране с женски 1, и броя на представянето на женски 1, необходими за достигане на критерия за насищане (таблица) 1, отдолу). Някои плъхове изискват многократно повторно въвеждане на женски 1 до приключване на фазата 5 (n = 3). Действията на поставяне на женски 1 зад екрана и премахването на делението може да са служили като основни апетитни сигнали, водещи до копулация. Трябва също да се отбележи, че критерият за насищане на 30 min без планина, въпреки че е използван преди това (Beach и Jordan, 1965; Mas et al., 1995b), е произволно и не гарантира, че плъхът няма да се монтира, ако има повече време. Въпреки това, забавянията или процедурите за отстраняване и замяна не доведоха надеждно при повторно копулиране с женски 1 (например, фази 4 и 5).

Поведение по време на експеримента с ефекта на Coolidge
Всички плъхове показват ефекта на Coolidge. Активността, свързана с поставянето на женски 2 зад екрана и по-специално отстраняването на делението, може да е допринесла за този резултат, но, отново, тези събития не са сами по себе си достатъчни да подновят копулацията по-рано в експеримента. Използвани са сравнения между мерките за сексуално поведение с женски 1 и женски 2 t тестове с корекция на Bonferroni. Въпреки, че латентността при монтиране и интродукция в отговор на женски 2 не се различават значително от тези при първото копулиращо нападение с женски 1, като цяло сексуалното поведение с женски 2 е по-малко стабилно, както се посочва от значително по-малко еякулации (средно, 0.6 срещу 4.2; F = 49.86;p <0.01) и интромисии (средно = 11.2 срещу 37.0;F = 20.17; p <0.05) през първия час. Броят на монтирания през първия час при жени 1 и 2 не се различава значително.
Важно е да се отбележи, че жените, използвани по време на наситената част от експеримента (т.е. женски 1), все още проявяват силно процептивно (т.е. подскачащо и стремително) и рецептивно (т.е. лордозно) поведение за цялата продължителност на техния контакт с мъжа ,
Neuroche-
Базалните наномоларни концентрации на DA и неговите метаболити в микродиализите, представени като средно ± SEM, от първите три проби от изходното ниво са: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; и HVA, 234.2 ± 49.0 (некоригирани за възстановяване на сондата;n = 5). Тези стойности представляват 100% базисни резултати.
Поведенчески определени данни, съответстващи на всяка фаза на експеримента и общи за всеки плъх, бяха използвани за неврохимични анализи. Те включват следното: (1) седем проби след първото въвеждане на женски 1, (2) четири проби, съпровождащи липсата на копулаторно поведение с женски 1, и (3) пет проби след представяне на женски 2. Фигура 1 илюстрира промени в концентрациите на DA (линейна графика, среден) и DA метаболити (линейна графика, връх) паралелно поведението на копулатор (графика, дъно) по време на теста за ефекта на Coolidge.
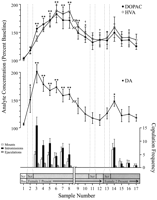
Преглед на по-голямата версия:
Фиг. 1.
Nucleus accumbens неврохимични корелати на сексуално поведение по време на Coolidge ефект. Първите осем проби представляват хронологично непрекъснати данни от фази 1 до 3. Проба 1 е четвъртата и последна извадка от изходната линия на прескопцията (Bas). Примерна 2 представлява въвеждане на женски 1 зад екрана (Scr). След 15 min, екранът беше отстранен и плъховете бяха оставени да копулират (проби 3-8). Най-почивка на x-xis съответства на изключването на данни от три плъха, които са копулирали за продължителни периоди с първоначалната женска. Последните девет проби също са непрекъснати хронологично. Пробите 9 и 10 съответстват на периода на насищане на фаза 3 (т.е. 30 min без опора). Женският 1 беше след това отново поставен зад екрана (проба 11) и 15 min по-късно, екранът беше премахнат (проба 12). След 15 min без копулация, женският 2 се поставя зад екрана (проба 13). Пробите 14 – 17 съответстват на копулация с женски 2. Броят на монтирания, интромисиите или еякулациите, свързани с всяка 15 min микродиализна проба, са показани в долната графика, Неврохимичните данни са изразени като процент от изходните концентрации. Промени в NAC DA (затворени квадрати), DOPAC (затворени кръгове) и HVA (отворени кръгове) изтичането е представено като линейни графики, Бяха направени следните сравнения: изходна извадка 1 срещу проби 2 – 10; нова изходна проба 10 срещу проби 11 и 12; нова извадка 12 спрямо извадки 13 – 17 (*)p <0.05; ** p <0.01). Независим t Извършени са тестове между базовите стойности (образци 1, 10 и 12). За значителни разлики от първата базова линия (проба 1), †p <0.05.
Отделни еднопосочни ANOVAs с многократно измерване бяха проведени на неврохимични данни, свързани с женски 1 (проби 1-12) и женски 2 (проби 12-17). Априори сравненията са направени с помощта на теста за многократно сравнение на Dunn (Bonferroni t). Бяха направени следните три основни сравнения: (1) първоначална базова линия (проба 1) спрямо проби 2 – 10 (първа експозиция на женски 1), (2) втора изходна (образец 10) срещу проби 11 и 12 (преизлагане на женски 1) и (3) трета базова линия (проба 12) спрямо проби 13 – 17 (експозиция на женски 2).
Налице е значителна цялостна промяна в изтичането на DA в отговор на женския 1 [F (11,44) = 8.48; p <0.001] и женски 2 [F (5,20) = 2.83;p <0.05]. Установено е значително увеличение на изтичането на DA, когато жена 1 присъства зад екрана (+ 44%,p <0.05; проба 2). По време на съвкуплението концентрациите на DA се увеличават допълнително, достигайки максимална стойност (+ 95%;p <0.01) по време на първия копулационен двубой (проба 3). DA остава повишен по време на копулация и се връща към изходните концентрации само през периода от 30 минути, в който не е настъпило нарастване (проби 9 и 10). Нито повторно въвеждане на женски 1 зад екрана (проба 11), нито възможността за физическо взаимодействие, но без монтиране (проба 12), повишени концентрации на DA спрямо втората изходна стойност (проба 10). Присъствието на жена 2 зад екрана (проба 13) води до малко увеличение на изтичането на DA (12%) спрямо третата базова стойност (проба 12), което не достига статистическа значимост. Подновеното съвкупление с жена 2 доведе до значително (34%) увеличение (p <0.05) при изтичане на DA по време на първата проба от копулация (проба 14). Въпреки че слабото копулаторно поведение продължава през следващите три проби, концентрациите на DA намаляват до изходни стойности (проби 15-17). Независим t тестовете, проведени между „изходни“ проби (т.е. 1, 10 и 12), показаха, че тези стойности не са били значително различни.
При трите плъха, които са възобновили копулацията при повторно въвеждане на женски 1, концентрациите на NAC DA се повишават, когато женските 1 се намират зад екрана (диапазон, 25 - 47%) и по време на копулация (диапазон, 13 - 37%), по отношение на пробата. преди повторното въвеждане на женската. Тези увеличения, обаче, се случват само когато сексуалното поведение е енергично и води до еякулация.
Значителни цялостни промени в DOPAC [F (11,44) = 9.57; p <0.001] и HVA [F (11,44) = 12.47; p <0. 001] бяха открити концентрации в отговор на женски 1, но не и женски 2. Концентрациите на метаболит се увеличиха леко (+ 15% и в двата случая) по време на представянето на женски 1 зад екрана (проба 2), но това не беше значително статистически. Имаше обаче значително увеличение на концентрациите на DOPAC и HVA по време на копулация (проби 3–8), достигайки максимални стойности (+80 и + 86%, съответно; p <0.01) след 60 минути (проба 6 и в двата случая). Въпреки че концентрациите на метаболит намаляват през периода на сексуално бездействие в края на контакта с жена 1 (проби 9 и 10), концентрациите все още остават повишени по отношение на изходното ниво (p <0.05 и в двата случая). Повторното въвеждане на женска 1 зад екрана (проба 11), достъп до женска 1 след отстраняване на екрана (проба 12) и въвеждането на женска 2 (проба 13) не доведоха до никакви промени в концентрациите на метаболитите. Лекото, но статистически незначително увеличение на концентрациите на DOPAC и HVA (+ 23% и в двата случая) спрямо изходното ниво (проба 12) съответства на първата атака на копулация с жена 2 (проба 14). Това увеличение обаче е краткотрайно и намалява до изходните стойности за останалите три проби (15-17). Независим tтестовете, проведени сред "изходни" проби (т.е. 1, 10 и 12) показват, че втората и третата изходни стойности (проби 10 и 12, съответно), въпреки че не се различават една от друга, остават повишени значително в сравнение с първата извадка. за DOPAC и HVA (p <0.05 и в двата случая).
Хистология
Микродиализните сонди се намират в NAC (Фиг.2) в диапазон + 1.20 до + 1.70 mm от брегма (плосък череп). Съществува и вариабилност в средната равнина; Данните отразяват извадката от черупките и основните подрегиони на НКП.
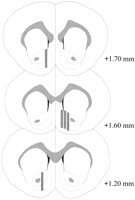
Преглед на по-голямата версия:
Фиг. 2.
Местоположение на микродиализни сонди в NAC на мъжки плъхове, използвани в експеримента на Coolidge ефекта. Защриховани правоъгълници отговарят на откритата мембранна област на микродиализните сонди. Серийни коронални мозъчни секции бяха преначертаниПаксинос и Уотсън (1986).
ДИСКУСИЯ
В съгласие с предишни доклади, настоящите резултати показват засилено мезолимбично предаване на DA, свързано с апетитни и консуматорни компоненти на сексуалното поведение на мъжки плъхове, оценено отин виво микродиализа (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). В допълнение, тези резултати осигуряват неврохимична корелация за сексуално насищане и последващото повторно иницииране на копулация в отговор на нова рецептивна жена (ефектът на Coolidge). Настоящите данни предполагат, че стимулационните свойства на нова рецептивна жена могат да служат за увеличаване на предаването на NAC DA при сексуално наситен мъжки плъх, който от своя страна може да бъде свързан с повторното създаване на сексуално поведение. Това е първото, което се проявява в лекото повишаване на NAC DA по време на представянето на нова женска зад екрана и се появява най-убедително като по-изразено увеличение по време на повторното копулация с женски 2 (Фиг.1).
Присъствието на първата рецептивна женска зад екрана доведе до силно апетитно увеличение на изтичането на NAC DA (44% от изходното ниво), подобно на величината, описана в предишни експерименти с подобен дизайн (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992). Също в съгласие с тези проучвания беше наблюдението, че изтичането на NAC DA се засилва допълнително по време на копулация (до> 95% над изходното ниво в настоящия експеримент). Въпреки че можем да разглеждаме консуматорското поведение като свързано с подобрено освобождаване на NAC DA (Wenkstern et al., 1993; Wilson et al., 1995), важно е да се разгледат понятията „апетит“ и „консуматор“ в контекста на сексуалното поведение. Докато фазата, в която жената присъства зад екрана, е изключително апетитна или подготвителна, поведението по време на фазата на копулация не може да се счита за чисто консуматорско. Тъй като „апетит“ може да се използва за описване на всички поведения, водещи до завършване на мотивирано поведение (копулация), основното поведение на мъжките експонати, докато е активно в „консумативната” фаза, се описва най-добре като апетит; мъжът прекарва по-голямата част от времето и усилията си, преследвайки жената, за да се съчетае. В тази връзка, можем да съпоставим максималната NAC DA предаване с консуматор както и интензивни апетитни компоненти на сексуално поведение на мъжки плъхове.
Достъпът до втората, нова женска жена доведе до ново съживяване във всеки субект. Предишни проучвания показват, че по-голямата част от плъховете, на които е разрешено да копулират до ситост, като използват подобен поведенчески протокол спрямо този, използван в настоящия експеримент, не възобновяват чифтосването, когато се тестват 24 часа по-късно (Плаж и Йордания, 1956). Вероятно наличието на нови стимулиращи свойства на женски 2, които може да включват обонятелни, както и визуални и слухови сигнали, са довели до ново съживяване. Интересен въпрос, на който остава да се отговори, е с какъв механизъм мъжкият плъх различава нова женска от женска, с която наскоро се е родил. Сайт за този механизъм може да се намира в основната обонятелна система. Съобщава се, че целостта на тази система е от съществено значение за ефекта на Coolidge в хамстерите (Джонстън и Расмусен, 1984). Вомероназално-аксесоарната обонятелна система, при която наскоро е описан процес на феромонна памет при мишки (Kaba et al., 1994), също е основен кандидат. В това отношение е важно да се отбележи, че е използвано увеличаване на NAC DA предаването ин виво при мъжки плъхове, представени с легла, които са били изложени на женски плъхове в \ tLouillot et al., 1991; Мичъл и Гратън, 1992). Освен това, прилагането на К+ директно към вомероназалния нервен слой на допълнителната обонятелна крушка, както и към самата аксесоарна обонятелна крушка, е достатъчен за увеличаване на предаването на NAC DA (Мичъл и Гратън, 1992).
Първият 15 min копулация с женски 2 се свързва със значително увеличение на NAC DA. За разлика от женския 1, взаимодействието с женски 2 не води до увеличаване на NAC DA със същата величина по време на фазите на апетит (12%) или на консуматор (34%). Тези малки увеличения на NAC DA, обаче, корелират добре с намаленото ниво на сексуално поведение, показвано с женски 2 в сравнение с женския 1. Концентрациите на метаболитите остават повишени по време на фазата на ситост, което води до нови изходни концентрации (проби 10 и 12), които са повишени значително от първоначалната базова стойност (проба 1).
Временното забавяне на повишаването на концентрациите на DOPAC и HVA по време на копулация е в съответствие с тяхното образуване като метаболити на изходното съединение, DA. Предполага се, че концентрациите на микродиализния метаболит, поне по време на естествено поведение, което не е фармакологично предизвикано, осигуряват полезен индекс на невронната активност (Damsma et al., 1992; Fumero et al., 1994). Фактът, че метаболитните концентрации остават повишени дори по време на периоди на сексуална активност в този експеримент, когато концентрациите на ДА се връщат до предварителните стойности на базовата линия, поставя под съмнение това предложение.
Устойчивото повишаване на концентрациите на метаболита на DA, наблюдавано в този експеримент, отразява медиалния преоптичен профил (mPOA) на DA метаболитите, наблюдавани при плъхове на първия ден след като са се събрали до насищане (Mas et al., 1995a,b). Продължителните повишения на DOPAC и концентрациите на HVA в NAC или mPOA не винаги се наблюдават, когато периодът на чифтосване е с фиксирана продължителност, много по-кратък от времето, необходимо за достигане на насищане. Например, много проучвания показват, че концентрациите на DOPAC са повишени и остават повишени по време на копулацията, но намаляват до изходните стойности скоро след отстраняването на женската (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). В изследването от Mas et al. (1995b)базалните екстрацелуларни концентрации на DOPAC и HVA в mPOA остават повишени за 4 последователни дни, съответстващи на период на сексуална неактивност. До четвъртия ден, точно преди животните да възобновят копулацията, базалните концентрации на метаболитите са близки до стойностите на пренаселване. Авторите сравняват модела на неврохимичните промени с тези, наблюдавани след администрирането на DA рецепторни блокери (Zetterström et al., 1984; Императо и ДиЧиара, 1985) и са предположили, че състоянието на сексуалната активност може да бъде медиирано чрез освобождаване на пролактин, което може да действа като „ендогенен невролептик“ (Mas et al., 1995a,b,d). Ясно е, че прилагането на невролептици е придружено от повишаване на концентрациите на извънклетъчния метаболит и изтичането на DA (Zetterström et al., 1984; Императо и ДиЧиара, 1985). За жалост, Mas et al. (1995a,b) не са в състояние да открият mPOA DA концентрации. В настоящото проучване, концентрациите на DA в NAC се връщат до стойности на пре-населението, докато концентрациите на DOPAC и HVA остават повишени. Този модел е несъвместим с ролята на ендогенния невролептик, действащ в NAC да индуцира сексуална ситост.
Като се има предвид участието на мезолимбични DA неврони в мотивирано поведение (Fibiger и Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) и тяхната чувствителност към нови \ tFabre et al., 1983; Шулц, 1992; Mirenowicz и Schultz, 1994), наблюдаваните повишения на извънклетъчните концентрации на NAC DA в отговор на новата женска са в съответствие с хипотезата, че активността в тази DA система е важна за повторното иницииране на сексуално поведение. В допълнение, доклади за апетитно и консумативно увеличение на предаването на DA (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) и невронна активност (Shimura et al., 1994) в mPOA на мъжки плъхове по време на сексуално поведение предполагат, че тази структура може също да допринесе за обновяване на копулацията, характерна за ефекта на Coolidge.
В съответствие с общата роля на мезолимбичната DA система при мотивирано поведение, е добре установено, че извънклетъчните концентрации на DA също са повишени преди, по време и веднага след консумирането на храна, с връщане към изходните стойности N30 min по-късно (Wilson et al., 1995). Добре известно е, че ситото, предизвикано от храната, се влияе от неговите сензорни свойства. Хората и животните отхвърлят храната, на която са били хранени, за да понасят и да погълнат други храни, които не са били изядени (Ролки, 1986). Това повдига въпроса дали екстрацелуларният изтичане на DA в NAC ще бъде увеличен селективно чрез представяне на нов вид храна, но не и от консумирана наскоро храна за ситост по начин, аналогичен на този, докладван в настоящото проучване в контекста на сексуална мотивация. Ако се потвърди, тази обща връзка между сетивните свойства на естествените награди, ситостта и мезолимбичното предаване на DA би означавало критична роля за тази невронна система при регулирането на мотивационните процеси, нарушаването на които може да доведе до сериозни нарушения на храненето и сексуалната функция. ,
СПРАВКИ
- ↵
- Beach FA,
- Jordan L
(1956) Сексуално изтощение и възстановяване при мъжки плъх. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Времеви характеристики на ефекта на Coolidge при поведенческо поведение на мъжки плъхове. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Блекбърн,
- Pfaus JG,
- Филипс АГ
(1992) Допаминът функционира при апетитно и отбранително поведение. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Филипс АГ,
- Fibiger HC
(1992) Сексуалното поведение увеличава трансмисията на допамин в nucleus accumbens и striatum на мъжките плъхове: сравнение с новост и локомоция. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Уилямс Г.
(1983) Активност на невроните в коремната област на маймуната. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Филипс АГ
(1986) Награда, мотивация, познание: психобиология на мезотенцефаличните допаминови системи. в Наръчник по физиология: нервната система IV, eds Bloom FE, Geiger SD (Американско дружество по физиология, Bethesda, MD), стр. 647 – 675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Филипс АГ
(1993) Електрическото стимулиране на местата за възнаграждение в коремната област увеличава трансмисията на допамин в nucleus accumbens на плъха. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Ефекти от вариация на стимула при сексуално насищане при мъжки плъх. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Промяна в стимулиращия стимул и сексуалното поведение при мъжки плъх. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Промени в обмяната на моноамин в областите на преден мозък, свързани с мъжко сексуално поведение: изследване на микродиализа. Brain Res 662: 233-239.
- ↵
- Холмс ГМ,
- Holmes DG,
- Sachs BD
(1987) Система за събиране на данни на базата на IBM-PC за записване на сексуално поведение на гризачи и за записване на общо събитие. Physiol Behav 44: 825-828.
- ↵
- Hull EM,
- Eaton RC,
- Мойсей,
- Lorrain DS
(1993) Копулацията повишава активността на допамина в медиалната преоптична област на мъжки плъхове. Life Sci 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Екстрацелуларен допамин в медиалната преоптична област: последствия за сексуалната мотивация и хормоналния контрол на копулацията. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Освобождаване и метаболизъм на допамин при будни плъхове след системни невролептици, както е изследвано чрез транс-стриатална диализа. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Расмусен К
(1984) Индивидуално разпознаване на женски хамстери от мъже: роля на химичните сигнали и на обонятелните и вомероназалните системи. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Индукция на обонятелна памет чрез активиране на метаботропен глутаматен рецептор. наука 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Куки MS
(1993) Фармакологията и нервната верига на чувствителността към психостимуланти. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Мезокортиколимбични допаминергични неврони. Функционални и регулаторни роли. в Psychopharmacology: четвъртото поколение на прогреса, eds Bloom FE, Kupfer DJ (Raven, New York), стр. 283 – 294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Гуадалупе Т,
- Mas M
(1991) Свързаните с пола обонятелни стимули индуцират селективно увеличение на освобождаването на допамин в nucleus accumbens на мъжки плъхове. Brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Едноличен С,
- Guadalupe T
(1990) Повишено освобождаване на допамин в nucleus accumbens на копулиращи мъжки плъхове, както е доказано чрез in vivo волтаметрия. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Фернандес-Вера,
- Gonzalez-Mora JL
(1995a) Неврохимични корелати на сексуалното изтощение и възстановяване, оценени чрез микродиализа in vivo. Brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Волтаметричен и микродиализен мониторинг на освобождаването на мозъчен моноаминов невротрансмитер по време на социосексуални взаимодействия. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Перес-Родригес I
(1995c) Индуциране на поведението на чифтосване от апоморфин при сексуално наситени плъхове. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Перес-Родригес I,
- Gonzalez-Mora JL
Неврохимия на сексуалната ситост. Експериментален модел на инхибирано желание. в фармакологията на сексуалната функция и дисфункция, ed Bancroft J (Raven, New York), стр. 1995 – 115.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Значение на непредсказуемостта за отговора на възнагражденията при допаминергичните неврони на приматите. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Мезолимбичното освобождаване на допамин се предизвиква чрез активиране на допълнителната обонятелна система: високоскоростно хроноамперометрично изследване. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Уотсън С
(1986) Мозъкът на плъх в стереотаксични координати (2nd ed). (Академик, Сан Диего).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- CD Блаха,
- Филипс АГ,
- Fibiger HC
(1990) Сексуалното поведение повишава централната трансформация на допамин при мъжки плъх. Brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Психофармакологията на сексуалното поведение. в Psychopharmacology: четвъртото поколение на прогреса, eds Bloom FE, Kupfer DJ (Raven, New York), стр. 743 – 758.
- ↵
- Pfaus JG,
- Горзалка Б.Б.
(1987) Опиоиди и сексуално поведение. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Филипс АГ,
- CD Блаха,
- Pfaus JG,
- Блекбърн Дж
(1992) Невробиологични корелати на позитивни емоционални състояния: допамин, очакване и награда. в Международен преглед на изследвания върху емоциите, ed Strongman (Wiley, New York), стр. 31 – 50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Съотношение на освобождаването на допамин в nucleus accumbens с мъжко сексуално поведение при плъхове. Brain Res 524: 160-163.
- ↵
- Родригес-Манзо Г,
- Фернандес Гуасти А
(1994) Обръщане на сексуалното изтощение от серотонергични и норадренергични средства. Behav Brain Res 62: 127-134.
- ↵
- Родригес-Манзо Г,
- Фернандес Гуасти А
(1995a) Участие на централната норадренергична система при възстановяване на копулаторното поведение на сексуално изчерпани плъхове от йохимбин, налоксон и 8-OH-DPAT. Brain Res Bull 38: 399-404.
- ↵
- Родригес-Манзо Г,
- Фернандес Гуасти А
(1995b) Опиоидни антагонисти и феномен на сексуално насищане. Psycho- 122: 131-136.
- ↵
- Rolls BJ
(1986) Сензорно специфична ситост. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) Поведенческата неврохимия на мотивацията: методологични и концептуални въпроси в изследванията на динамичната активност на nucleus accumbens dopamine. J Neurosci методи 64: 137-149.
- ↵
- Сато Й,
- Wada H,
- Horita H,
- Сузуки N,
- Shibuya A,
- Адачи Х,
- Kato R,
- Tsukamoto T,
- Кумамото Y
(1995) Освобождаването на допамин в медиалната преоптична област по време на мъжко поведение при плъхове. Brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Активност на допаминовите неврони в поведението на приматите. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Ямамото Т,
- Шимокочи М
(1994) Медиалната преоптична област е включена както в сексуалната възбуда, така и в представянето при мъжки плъхове: преоценка на невронната активност в свободно движещи се животни. Brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Допаминовото предаване се увеличава в nucleus accumbens на мъжки плъхове по време на първото им излагане на полово възприемчиви женски плъхове. Brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Допаминергични корелати на мотивираното поведение: значение на задвижването. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Плаж ФА
(1963) Модификация на сексуалното поведение на мъжки плъхове, произведени чрез промяна на стимулатора женски. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Ефект на невролептичните лекарства върху освобождаването на стриматен допамин и метаболизма в събудените плъхове, изследвани чрез интрацеребрална диализа. Eur J Pharmacol 106: 27-37.
Статии, цитиращи тази статия
- Ендогенна опиоидно-индуцирана невропластичност на допаминергични неврони в вентралната зона на тигмалата влияе върху естественото и наградата за опиати Вестник на неврологията, 25 юни 2014, 34 (26): 8825-8836
- Дисбаланс в чувствителността към различни видове награди в патологичния хазарт Мозъчен, 1 август 2013, 136 (8): 2527-2538
- Закон за природните и лекарствените възнаграждения относно механизмите за обща нервна пластичност с {Delta} FosB като ключов посредник Вестник на неврологията, 20 февруари 2013, 33 (8): 3434-3442
- Невронални реакции в обвивката на ядрото при сексуално поведение при мъжки плъхове Вестник на неврологията, 1 февруари 2012, 32 (5): 1672-1686
- Романтична любов: бозайникова мозъчна система за избор на партньор Философски сделки на Кралското общество Б: Биологични науки, 29 Декември 2006, 361 (1476): 2173-2186
- Хедонични горещи петна в мозъка Невролозите, 1 декември 2006, 12 (6): 500-511
- Модулация от централни и базолатерални амигдаларни ядра на допаминергични корелати на хранене към сатиите в ядрените ядра на плъх и медиалния префронтален кортекс Вестник на неврологията, 15 декември 2002, 22 (24): 10958-10965
- Честота на преходните фази на концентрацията на допамин в гръбначния и вентралния стрийатум на мъжките плъхове при въвеждане на конспецифични Вестник на неврологията, 1 декември 2002, 22 (23): 10477-10486
- Преглед на книгата: Динамика на извънклетъчния допамин в остри и хронични действия на кокаина Невролозите, 1 Август 2002, 8 (4): 315-322
- Диференцирано включване на NMDA, АМРА / каинат и допаминови рецептори в ядрото на акумулиращото ядро при придобиване и изпълнение на поведение на Павловския подход Вестник на неврологията, 1 декември 2001, 21 (23): 9471-9477
- Контрол на кокаин-търсещо поведение чрез свързани с лекарството стимули при плъхове: Ефекти върху възстановяването на погасените оперантни и екстрацелуларни нива на допамин в амигдала и nucleus accumbens PNAS, 11 април 2000, 97 (8): 4321-4326
- Улесняване на половото поведение и засилено допаминово изтичане в ядрените обвивки на мъжки плъхове след индуцирана от D-амфетамин поведенческа сенсибилизация Journal of Neuroscience, 1, 1999, 19 (1): 456-463
- НЕВРОУКА: Привличане на вниманието на мозъка Наука, 3 октомври 1997, 278 (5335): 35-37
- Допаминергични корелати на сензорно-специфичната плътност в медиалния префронтален кортекс и ядрените обвивки на плъха Вестник на неврологията, 1 октомври 1999, 19 (19): RC29
