КОМЕНТАРИ: Както по-късните проучвания ще разкрият, DeltaFosB е често срещаният молекулярен превключвател както за наркомании, така и за поведенчески зависимости. Това е транскрипционен фактор, което означава, че влияе върху това кои гени са включени или изключени. Както е посочено другаде, пристрастяващите наркотици отвличат само нормалните механизми. Ето защо е глупаво да се предполага, че поведенческите зависимости не могат да съществуват.
Proc Natl Acad Sci US A. 2001 септември 25; 98 (20): 11042-11046.
doi: 10.1073 / pnas.191352698.
Ерик Дж. Нестлер *, Мишел Баро и Дейвид У. Себе
Катедра по психиатрия и център за основни невронауки, Югозападен медицински център на Университета на Тексас, 5323 булевард Хари Хайнс, Далас, TX 75390-9070
абстрактен
Дълготрайността на някои от поведенческите аномалии, които характеризират наркоманията, предполага, че регулирането на нервната генна експресия може да бъде включено в процеса, чрез който наркотичните вещества причиняват пристрастяване. азУвеличените доказателства предполагат, че транскрипционният фактор ΔFosB представлява един механизъм, чрез който лекарствата за злоупотреба произвеждат относително стабилни промени в мозъка, които допринасят за фенотипа на зависимостта. ΔFosB, член на семейството на Fos на транскрипционни фактори, се натрупва в рамките на подгрупа от неврони на nucleus accumbens и дорзална стриатум (области на мозъка, важни за пристрастяване) след многократно приложение на много видове наркотици. Подобно натрупване на ΔFosB настъпва след компулсивно протичане, което предполага, че ΔFosB може да се натрупва в отговор на много видове компулсивно поведение., Важно е да се отбележи, че ΔFosB се запазва в невроните за относително дълги периоди от време поради своята изключителна стабилност. Следователно, ΔFosB представлява молекулен механизъм, който може да инициира и след това да поддържа промени в генната експресия, които продължават дълго след прекратяване на експозицията на лекарството., Проучвания в индуцируеми трансгенни мишки, които свръхекспресират или ΔFosB, или доминиращ отрицателен инхибитор на протеина, осигуряват директно доказателство, че ΔFosB причинява повишена чувствителност към поведенческите ефекти на лекарства от злоупотреба и, евентуално, повишено поведение при търсене на наркотици. Тази работа подкрепя виждането, че ΔFosB функционира като вид устойчив „молекулен превключвател“, който постепенно превръща острата реакция на лекарството в относително стабилни адаптации, които допринасят за дългосрочната нервна и поведенческа пластичност, която е в основата на зависимостта.
Изследването на зависимостта се фокусира върху разбирането на сложните начини, по които лекарствата за злоупотреба променят мозъка, за да предизвикат поведенчески аномалии, които характеризират зависимостта. Едно от най-важните предизвикателства в тази област е да се идентифицират относително стабилни лекарствени промени в мозъка, за да се отчетат тези поведенчески аномалии, които са особено продължителни. Например, човешкият наркоман може да е изложен на повишен риск от рецидив дори след години въздържание.
Стабилността на тези поведенчески аномалии е довела до предположението, че те могат да бъдат медиирани, поне частично, чрез промени в генната експресия (1-3). Според това схващане, повтарящото се излагане на наркотици на злоупотреба многократно нарушава предаването на определени синапси в мозъка, които са чувствителни към лекарството. Такива смущения в крайна сметка сигнализират чрез вътреклетъчни пратеници каскади към ядрото, където те първо инициират и след това поддържат промени в експресията на специфични гени. Първичен механизъм, чрез който сигналните трансдукционни пътища влияят върху генната експресия, е регулирането на транскрипционни фактори, протеини, които се свързват с регулаторни области на гените и модифицират тяхната транскрипция.
Една от целите на изследванията за пристрастяване, следователно, е да се идентифицират транскрипционни фактори, които се променят в участъците на мозъка, замесени в пристрастяването след хронично приложение на наркотици. Няколко такива транскрипционни фактора са идентифицирани през последното десетилетие (1 – 6). Фокусът на този преглед е върху един специфичен транскрипционен фактор, наречен ΔFosB.
Индукция на ΔFosB по лекарства от злоупотреба
ΔFosB, кодиран от гена fosB, е член на Fos фамилията на транскрипционни фактори, които също включват c-Fos, FosB, Fra1 и Fra2 (7). Тези Fos фамилни протеини хетеродимеризират с протеини от семейство Jun (c-Jun, JunB или JunD), за да образуват активни AP-1 (активатор протеин-1) транскрипционни фактори, които се свързват към AP-1 сайтове (консенсусна последователност: TGAC / GTCA) промоторите на определени гени да регулират тяхната транскрипция.
Тези Fos фамилни протеини се индуцират бързо и преходно в специфични участъци на мозъка след остро администриране на много лекарства за злоупотреба (фиг. 1) (8-11). Изтъкнати области са nucleus accumbens и dorsal striatum, които са важни медиатори на поведенческите реакции към лекарствата, по-специално техните благоприятни и локомоторно-активиращи ефекти (12, 13). Тези протеини се връщат до базовите нива в рамките на часове на прилагане на лекарството.
Фигура 1
Схема, показваща постепенното натрупване на ΔFosB в сравнение с бързата и преходна индукция на други протеини на семейството Fos в отговор на злоупотреба с лекарства. (А) Авторадиограмата илюстрира диференциалната индукция на тези различни протеини чрез остра стимулация (1-2 hr след единична експозиция на лекарство) в сравнение с хронична стимулация (1 ден след повторно излагане на лекарството). (B) Няколко вълни на Fos-подобни протеини [състоящи се от c-Fos (52-до 58-kDa изоформи), FosB (46-до 50-kDa изоформи), ΔFosB (33-kDa изоформа), и Fra1 или Fra2 ( 40 kDa)] се индуцират в neurous accumbens и dorsal striatal неврони чрез остро приложение на лекарство за злоупотреба. Също така са индуцирани биохимично модифицирани изоформи на АFosB (35-37 kDa); те също се индуцират (макар и на ниски нива) след острата употреба на лекарството, но продължават в мозъка за дълъг период от време поради тяхната стабилност. (В) При повторно прилагане (например, два пъти дневно), всеки остър стимул индуцира ниско ниво на стабилните ΔFosB изоформи, което е показано от по-ниската група от припокриващи се линии, които показват, че FOSB е индуциран от всеки остър стимул. Резултатът е постепенно увеличаване на общите нива на ΔFosB с повтарящи се стимули по време на хронично лечение, което е показано от нарастващата степенна линия в графиката.
Много различни реакции се наблюдават след хронично приложение на лекарства за злоупотреба (фиг. 1), Биохимично модифицираните изоформи на ΔFosB (молекулна маса 35-37 kDa) се натрупват в същите участъци на мозъка след повторно излагане на лекарството, докато всички други членове на семейството на Fos показват толерантност (т.е. намалена индукция в сравнение с първоначалната експозиция на лекарството). Такова натрупване на ΔFosB е наблюдавано при кокаин, морфин, амфетамин, алкохол, никотин и фенциклидин.e (11, 14-18). Има някои доказателства, че тази индукция е селективна за динорфин / субстанция, съдържаща P подгрупа от средни бодливи неврони, разположени в тези участъци на мозъка (15, 17), въпреки че е необходима повече работа, за да се установи това със сигурност. 35- до 37-kDa изоформите на ΔFosB димеризират предимно с JunD за образуване на активен и дълготраен AP-1 комплекс в тези мозъчни участъци (19, 20). Тези ΔFosB изоформи се натрупват с хронична експозиция на лекарство поради извънредно дългия им полуживот (21) и следователно остават в невроните в продължение на поне няколко седмици след спиране на администрирането на лекарството. Интересно е да се отбележи, че тези ΔFosB изоформи са високо стабилни продукти на непосредствен ранен ген (fosB). Стабилността на ΔFosB изоформите осигурява нов молекулен механизъм, чрез който медикаментозните промени в генната експресия могат да продължат въпреки относително дългите периоди на отнемане на лекарството.
Въпреки че nucleus accumbens играе решаваща роля в благоприятното въздействие на наркотиците на злоупотреба, се смята, че функционира нормално чрез регулиране на реакциите към естествени подсилващи, като храна, напитки, пол и социални взаимодействия (12, 13). В резултат на това е налице значителен интерес към възможна роля на този мозъчен регион при други натрапчиви поведения (например, патологично преяждане, хазарт, упражнения и др.). Поради тази причина изследвахме дали ΔFosB се регулира в животински модел на компулсивно бягане. Действително, стабилните 35- до 37-kDa изоформи на ΔFosB се индуцират селективно в nucleus accumbens при плъхове, които показват компулсивно поведение при движение. †
Биохимична идентичност на стабилни ΔFosB изоформи
Както бе споменато по-горе, изоформите на ΔFosB, които се натрупват след хронично прилагане на лекарство за злоупотреба или компулсивно бягане, показват молекулна маса на 35-37 kDa. Те могат да бъдат диференцирани от 33-kDa изоформата на ΔFosB, която е индуцирана бързо, но временно след еднократна експозиция на лекарство (Фиг. 1) (14, 19, 22). Настоящите доказателства предполагат, че изоформата на 33-kDa е нативна форма на протеина, който е променен, за да образува по-стабилните продукти 35-37-kDa (19, 21). Обаче, природата на биохимичната модификация, която превръща нестабилната 33-kDa изоформа в стабилните 35- до 37-kDa изоформи остава неясна. Предполага се, че фосфорилирането може да бъде отговорно (11). Например, индуцирането на ΔFosB е отслабено при мишки без DARPP-32, обогатен с стриатал протеин (23, 24). Тъй като DARPP-32 регулира каталитичната активност на протеинов фосфатаза-1 и протеин киназа А (25, 26), изискването за този протеин за нормалното натрупване на стабилните ΔFosB изоформи предполага възможна роля за фосфорилирането в генерирането на тези стабилни продукти.
Ролята на ΔFosB в поведенческата пластичност спрямо наркотиците от злоупотреба
Вникването в ролята на ΔFosB в пристрастяването към наркотици до голяма степен се дължи на изследването на трансгенни мишки, при които ΔFosB може да бъде индуциран селективно в рамките на nucleus accumbens и други стритални области на възрастни животни (27, 28). Важно е, че тези мишки свръхекспресират ΔFosB селективно в динорфин / вещество, съдържащо средни неврони, където се смята, че лекарствата индуцират протеина. Поведенческият фенотип на ΔFosB-свръхекспресиращи мишки, които в много отношения приличат на животни след хронична експозиция на лекарство, е обобщен в Таблица 1. Мишките показват увеличени локомоторни отговори на кокаин след остро и хронично приложение (28). Те също така показват повишена чувствителност към благоприятните ефекти на кокаина и морфина в тестовете за кондициониране на място (11, 28) и ще прилагат самостоятелно по-ниски дози кокаин, отколкото тези, които не прекомерно експресират ΔFosB. чувствителност към кокаин и нормално пространствено учене в водния лабиринт на Morris (28). Tтези данни показват, че ΔFosB повишава чувствителността на животното към кокаин и може би други наркотици и може да представлява механизъм за относително продължителна сенсибилизация към лекарствата.
 стриатума
стриатума
| Увеличено локомоторно активиране в отговор на остра и многократна употреба на кокаин. |
| Повишени отговори на кокаина и морфина в тестовете за кондициониране на място. |
| Повишено самостоятелно приложение на ниски дози кокаин. |
| Повишена мотивация за кокаин при прогресивни съотношения. |
| Повишен анксиолитичен отговор към алкохола. |
| Увеличено натрапчиво поведение при движение. |
Въз основа на данни в справки. 28 намлява 29.† ‡ §¶
Поведенческа пластичност, медиирана от ΔFosB в nucleus accumbens-dorsal striatum
IОсвен това, има предварителни доказателства, че ефектите на ΔFosB могат да се простират далеч отвъд регулирането на чувствителността на лекарството per se към по-сложни поведения, свързани с процеса на пристрастяване. Мишки, експресиращи ΔFosB, работят по-усилено за самостоятелно прилагане на кокаин в прогресивно съотношение самостоятелно приложение, \ tтвърдят, че ΔFosB може да повиши чувствителността на животните към стимулиращите мотивационни свойства на кокаина и по този начин да доведе до склонност към рецидив след отнемане на лекарствотоl. ‡ ΔFosB-експресиращите мишки също показват повишени анксиолитични ефекти на алкохола, фенотип, който е свързан с повишен прием на алкохол при хора. Заедно тези ранни открития показват, че ΔFosB, в допълнение към повишаването на чувствителността към наркотици, предизвиква качествени промени в поведението, които насърчават търсенето на наркотици. По този начин, ΔFosB може да функционира като непрекъснат "молекулен превключвател", който помага да се инициира и след това да се поддържат важни аспекти на пристрастеното състояние. Важен въпрос при настоящото изследване е дали натрупването на ΔFosB по време на експозицията на наркотици насърчава поведението на търсещите лекарство след удължаване на периода на оттегляне, дори след като нивата на ΔFosB се нормализират (виж по-долу).
Възрастен Мишките, които свръхекспресират ΔFosB селективно в рамките на nucleus accumbens и дорзалния стриатум, също показват по-голямо компулсивно протичане в сравнение с контролните littermates. поведение, може би чрез засилване на ефикасността на невронните вериги, в които функционират тези неврони.
ΔFosB се натрупва в някои мозъчни участъци извън nucleus accumbens и дорзалния стриатум след хронично излагане на кокаин. Изтъкнати сред тях области са амигдалата и медиалната префронтална кора (15). Основна цел на настоящите изследвания е да се разбере приноса на индукцията на ΔFosB в тези региони към фенотипа на зависимостта.
По-ранната работа върху нокаутиращи мишки fosB разкрива, че тези животни не успяват да развият сенсибилизация към двигателните ефекти на кокаина, което е в съответствие с констатациите на ΔFosB-свръхекспресиращите мишки, споменати по-горе (22). Мутантите fosB обаче показаха повишена чувствителност към острите ефекти на кокаина, което е в противоречие с тези други открития. Тълкуването на откритията с мутантите fosB обаче се усложнява от факта, че на тези животни липсва не само ΔFosB, но и FosB с пълна дължина. Освен това на мутантите липсват както протеини в мозъка, така и от най-ранните етапи на развитие. В действителност, по-скорошна работа подкрепя заключения от мишките свръхекспресиращи ΔFosB: индуцируема свръхекспресия на пресечен мутант на c-Jun, който действа като доминиращ отрицателен антагонист на ΔFosB, селективно в nucleus accumbens и гръбния стриатум показва намалена чувствителност към въздействащите ефекти на кокаина .¶ Тези открития подчертават предпазливостта, която трябва да се използва при интерпретиране на резултати от мишки с конститутивни мутации, и илюстрират значението на мишките с индуцируеми и специфични за клетъчния тип мутации при изследвания на пластичност в мозъка на възрастни.
Целеви гени за ΔFosB
Тъй като ΔFosB е транскрипционен фактор, вероятно протеинът причинява поведенческа пластичност чрез промени в експресията на други гени. ΔFosB се генерира чрез алтернативно заплитане на fosB гена и липсва част от С-терминалния трансактивиращ домен, присъстващ в FosB с пълна дължина. В резултат първоначално беше предложено, че ΔFosB функционира като транскрипционен репресор (29). Въпреки това, работата в клетъчната култура ясно показа, че ΔFosB може да предизвика или потисне AP-1-медиирана транскрипция в зависимост от конкретния използван AP-1 сайт (21, 29-31). FosB с пълна дължина оказва същите ефекти като ΔFosB върху някои фрагменти на промотора, но с различни ефекти върху другите. Необходима е по-нататъшна работа, за да се разберат механизмите, залегнали в основата на тези различни действия на ΔFosB и FosB.
Нашата група използва два подхода за идентифициране на целеви гени за ΔFosB. Единият е кандидат генният подход. Първоначално разгледахме глутаматните рецептори на а-амино-3-хидрокси-5-метил-4-изоксазолпропионова киселина (AMPA) като предполагаеми мишени, като се има предвид важната роля на глутаматергичното предаване в ядрото акумен. Досегашната работа показва, че една конкретна субединица на AMPA глутаматния рецептор, GluR2, може да бъде добросъвестна цел за ΔFosB (фиг. 2). Експресията на GluR2, но не и експресията на други AMPA рецепторни субединици, се увеличава в nucleus accumbens (но не и дорзален стриатум) при свръхекспресия на ΔFosB (28), а експресията на доминиращ отрицателен мутант отслабва способността на кокаина да индуцира протеина. В допълнение, промоторът на гена GluR2 съдържа консенсусно AP-1 място, което свързва ΔFosB (28). Свръхекспресията на GluR2 в nucleus accumbens, чрез използване на вирусно-медииран генен трансфер, повишава чувствителността на животното към възнаграждаващите ефекти на кокаина, като по този начин имитира част от фенотипа, наблюдаван при мишките, експресиращи ΔFosB (28). Индукцията на GluR2 може да обясни намалената електрофизиологична чувствителност на невроните на nucleus accumbens към агонистите на AMPA рецепторите след хронично приложение на кокаин (32), тъй като AMPA рецепторите, съдържащи GluR2, показват намалена обща проводимост и намалена пропускливост на Ca2 +. Намалената отзивчивост на тези неврони към възбуждащи входове може след това да подобри реакциите към злоупотреба с наркотици. Въпреки това, начините, по които допаминергичните и глутаматергичните сигнали в ядрото акумбени регулират пристрастяващото поведение, остават неизвестни; това ще изисква ниво на разбиране на невронна верига, което все още не е налично.
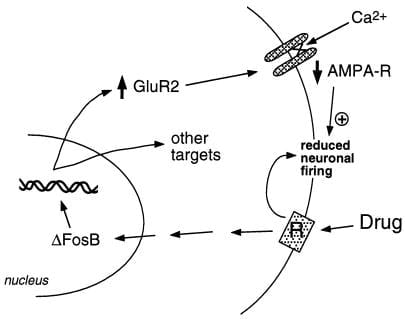
Фигура 2
Субединицата на глутаматния рецептор на AMPA, GluR2, е предполагаема цел за ΔFosB. Показано е как ΔFosB-медиираната индукция на GluR2 може да промени физиологичната реакция на невроните на nucleus accumbens и да доведе до сенсибилизиран отговор на злоупотреба с наркотици. Според тази схема злоупотребите с наркотици произвеждат своите остри подсилващи ефекти чрез инхибиране на ядрените неврони. При многократна експозиция лекарствата индуцират ΔFosB, който регулира множество целеви гени, включително GluR2. Това увеличава дела на AMPA рецепторите (AMPA-R) върху невроните на nucleus accumbens, които съдържат GluR2 субединицата, което причинява намален общ AMPA ток и намален Ca2 + ток. Тази намалена възбудимост може да направи невроните по-чувствителни към острите инхибиторни ефекти на лекарствата и по този начин към усилващите ефекти на лекарствата.
Друга предполагаема цел за ΔFosB е генът, кодиращ динорфин. Както беше посочено по-рано, динорфинът се експресира в подгрупата на nucleus accumbens средни бодливи неврони, които показват индукция на ΔFosB. Динорфинът функционира в междуклетъчната обратна връзка: неговото освобождаване инхибира допаминергичните неврони, които иннервират невроните в средата, чрез κ опиоидни рецептори, присъстващи в допаминергичните нервни терминали в nucleus accumbens, както и върху клетъчни тела и дендрити в коремната област (Фиг. 3) (33-35). Тази идея е в съответствие със способността на κ рецепторния агонист, при администриране в която и да е от тези две области на мозъка, за намаляване на лекарственото възстановяване.d (35).
RПървата работа показва, че ΔFosB намалява експресията на динорфин, което може да допринесе за повишаване на механизмите за възнаграждение, наблюдавани при индукция на ΔFosB. Интересно е, че друг лекарствено-регулиран транскрипционен фактор, CREB (cAMP отговор елемент свързващ протеин) (2, 3), проявява обратния ефект: той индуцира експресия на динорфин в nucleus accumbens и намалява полезните свойства на кокаина и морфина (4). **
Bтъй като индуцираната с лекарство активация на CREB бързо се разсейва след администриране на лекарството, такава реципрочна регулация на динорфина от CREB и ΔFosB може да обясни реципрочните промени в поведението, които настъпват по време на ранните и късни фази на отнемане, с преобладаващи негативни емоционални симптоми и намалена чувствителност към лекарства в ранните фази оттегляне и повишаване на чувствителността към възнаграждаващите и стимулиращите мотивационни ефекти на наркотици, преобладаващи в по-късни времеви моменти.
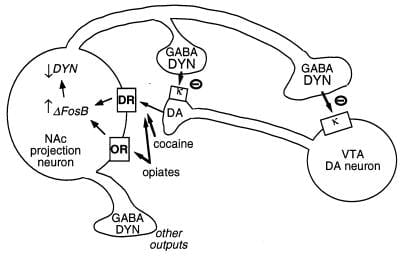
Фигура 3
Динорфинът е предполагаема цел за ΔFosB. Показан е вентрален тегментален участък (VTA) допаминов (DA) неврон, инервиращ клас на nucleus accumbens (NAc) GABAergic проекционен неврон, който експресира динорфин (DYN). Динорфинът служи за механизъм за обратна връзка в тази верига: динорфин, освободен от терминалите на NAc невроните, действа върху κ опиоидните рецептори, разположени на нервните крайници и клетъчните тела на DA невроните, за да инхибират тяхното функциониране. ΔFosB, чрез инхибиране на експресията на динорфин, може да регулира обратната верига на обратната връзка и да подобри полезните свойства на лекарствените средства. Не е показано реципрочния ефект на CREB върху тази система: CREB усилва експресията на динорфин и по този начин намалява полезните свойства на лекарствата, които се злоупотребяват (4). GABA, у-аминомаслена киселина; DR, допаминов рецептор; OR, опиоиден рецептор.
Вторият подход, използван за идентифициране на целеви гени за ΔFosB, включва анализ на ДНК микрочипове. Индуцируемата свръхекспресия на ΔFosB увеличава или намалява експресията на многобройни гени в nucleus accumbens (36). Въпреки че сега е необходима значителна работа за валидиране на всеки от тези гени като физиологични мишени на ΔFosB и за разбиране на техния принос към фенотипа на пристрастяване, една важна цел изглежда е Cdk5 (циклин-зависима киназа-5). По този начин Cdk5 първоначално е идентифициран като ΔFosB-регулиран чрез използване на микрочипове и по-късно е доказано, че се индуцира в nucleus accumbens и дорзален стриатум след хронично приложение на кокаин (37). ΔFosB активира cdk5 гена чрез AP-1 място, присъстващо в промотора на гена (36). Заедно тези данни поддържат схема, при която кокаинът индуцира експресията на Cdk5 в тези мозъчни области чрез ΔFosB. Индукцията на Cdk5 изглежда променя допаминергичното сигнализиране поне отчасти чрез повишено фосфорилиране на DARPP-32 (37), което се превръща от инхибитор на протеин фосфатаза-1 в инхибитор на протеин киназа А при неговото фосфорилиране от Cdk5 (26).
Ролята на ΔFosB в медиирането на “постоянната” пластичност спрямо наркотиците от злоупотреба
Въпреки че ΔFosB сигнала е относително дълготраен, той не е постоянен. ΔFosB се разгражда постепенно и вече не може да се открие в мозъка след 1 – 2 месеца на отнемане на лекарството, въпреки че някои поведенчески аномалии продължават за много по-дълги периоди от време. Следователно ΔFosB per se не изглежда да е в състояние да медиира тези полупостоянни поведенчески аномалии. Трудността да се намерят молекулярните адаптации, които са в основата на изключително стабилните промени в поведението, свързани с пристрастяването, е аналогична на предизвикателствата, пред които е изправена в областта на обучението и паметта. Въпреки че съществуват елегантни клетъчни и молекулярни модели на учене и памет, досега не е било възможно да се идентифицират молекулярни и клетъчни адаптации, които са достатъчно продължителни, за да се вземат предвид високо стабилните поведенчески спомени. Всъщност, ΔFosB е най-дълготрайната адаптация, за която е известно, че се среща в мозъка на възрастни, не само в отговор на злоупотреба с наркотици, но и на всякакви други смущения (които не включват лезии). Две предложения се развиха, както в пристрастяването, така и в областта на ученето и паметта, за да се обясни това несъответствие.
Една възможност е, че по-преходни промени в генната експресия, като тези, медиирани чрез ΔFosB или други транскрипционни фактори (например, CREB), може да медиира по-продължителни промени в невроналната морфология и синаптична структура. Например, увеличаване на плътността на дендритните бодли (особено увеличаването на двуглавите бодли) повишена ефикасност на глутаматергичните синапси при хипокампални пирамидални неврони по време на дългосрочно усилване (38-40) и паралели на повишената поведенческа чувствителност към кокаин, медиирана на нивото на средни бодливи неврони на nucleus accumbens (41). Не е известно дали такива структурни промени са достатъчно дълготрайни, за да се вземат предвид високо стабилните промени в поведението, въпреки че последните продължават поне за 1 месец отнемане на лекарството. Последните доказателства повдигат възможността, че ΔFosB и неговата индукция на Cdk5, е един медиатор на индуцирани от лекарството промени в синаптичната структура в nucleus accumbens (Фиг. 4). ‡ Така инфузията на Cdk5 инхибитор в nucleus accumbens предотвратява способността за повтаряща се експозиция на кокаин за увеличаване на плътността на дендритни гръбначни стълбове в този регион. Това е в съгласие с мнението, че Cdk5, който е обогатен в мозъка, регулира невронната структура и растежа (виж реферите 36 и 37). Възможно е, макар и в никакъв случай да не се докаже, че такива промени в невроналната морфология могат да надживеят самият ΔFosB сигнал.
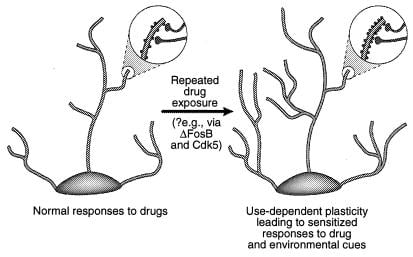
Фигура 4
Регулиране на дендритната структура чрез злоупотреба с наркотици. Показано е разширяването на дендритното дърво на неврона след хронично излагане на злоупотреба с наркотици, както е наблюдавано при кокаин в nucleus accumbens и префронталната кора (41). Областите на увеличение показват увеличение на дендритните бодли, което се предполага, че се случва заедно с активирани нервни терминали. Това увеличаване на дендритната плътност на гръбначния стълб може да бъде медиирано чрез ΔFosB и последващата индукция на Cdk5 (виж текста). Такива промени в дендритната структура, които са подобни на тези, наблюдавани в някои модели на обучение (напр. Дългосрочно усилване), биха могли да медиират дълготрайни сенсибилизирани реакции на злоупотреба с наркотици или сигнали от околната среда. [Възпроизведено с разрешение от реф. 3 (Copyright 2001, Macmillian Magazines Ltd.)].
Друга възможност е, че преходната индукция на транскрипционен фактор (например, ΔFosB, CREB) води до по-постоянни промени в генната експресия чрез модифициране на хроматип. Смята се, че тези и много други транскрипционни фактори активират или потискат транскрипцията на прицелен ген чрез насърчаване съответно на ацетилиране или деацетилиране на хистони в близост до гена (42). Въпреки че такова ацетилиране и деацетилиране на хистони очевидно може да се случи много бързо, е възможно ΔFosB или CREB да произвеждат по-дълготрайни адаптации в ензимната машина, която контролира ацетилирането на хистони. ΔFosB или CREB могат също така да насърчават дълготрайни промени в генната експресия чрез регулиране на други модификации на хроматин (например, метилиране на ДНК или хистон), които са замесени в постоянните промени в генната транскрипция, които се проявяват по време на развитието (виж референции 42 и 43) , Въпреки че тези възможности остават спекулативни, те могат да осигурят механизъм, чрез който преходните адаптации към наркотици на злоупотреба (или някакво друго смущение) водят до по същество поведенчески последици.
Препратки
- ↵
- Nestler EJ,
- Надежда БТ,
- Widnell KL
(1993) Neuron 11: 995 – 1006.
- ↵
- Berke JD,
- Hyman SE
(2000) Neuron 25: 515 – 532.
- ↵
- Nestler EJ
(2001) Nat Rev Neurosci 2: 119 – 128.
- ↵
- Carlezon WA Jr,
- Том Й,
- Olson VG,
- Lane-Ladd SB,
- Brodkin ES,
- Hiroi N,
- Думан РС,
- Neve RL,
- Nestler EJ
(1998) Наука 282: 2272 – 2275.
Резюме / БЕЗПЛАТЕН Пълен текст
- О'Донован KJ,
- WG на Tourtellotte,
- Millbrandt J,
- Baraban JM
(1999) Тенденции Neurosci 22: 167 – 173.
- ↵
- Mackler SA,
- Korutla L,
- Ча XY,
- Koebbe MJ,
- Фурние КМ,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210 – 6217.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Morgan JI,
- Curran T
(1995) Тенденции Neurosci 18: 66 – 67.
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci САЩ 88: 1291 – 1295.
Резюме / БЕЗПЛАТЕН Пълен текст
- Graybiel AM,
- Moratalla R,
- Robertson HA
(1990) Proc Natl Acad Sci САЩ 87: 6912 – 6916.
Резюме / БЕЗПЛАТЕН Пълен текст
- Надежда Б,
- Kosofsky B,
- Hyman SE,
- Nestler EJ
(1992) Proc Natl Acad Sci САЩ 89: 5764 – 5768.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Kelz MB,
- Nestler EJ
(2000) Curr Opin Neurol 13: 715 – 720.
- ↵
- Koob GF,
- Sanna PP,
- Блум ФЕ
(1998) Neuron 21: 467 – 476.
- ↵
- Wise RA
(1998) Зависимост от алкохол на наркотици 51: 13 – 22.
- ↵
- Надежда БТ,
- Nye HE,
- Kelz MB,
- Self DW,
- Iadarola MJ,
- Nakabeppu Y,
- Думан РС,
- Nestler EJ
(1994) Neuron 13: 1235 – 1244.
- ↵
- Nye H,
- Надежда БТ,
- Kelz M,
- Iadarola M,
- Nestler EJ
(1995) J Pharmacol Exp Ther 275: 1671-1680.
Резюме / БЕЗПЛАТЕН Пълен текст
- Nye HE,
- Nestler EJ
(1996) Mol Pharmacol 49: 636-645.
- ↵
- Moratalla R,
- Елибол Б,
- Валехо М,
- Грейбиел АМ
(1996) Neuron 17: 147 – 156.
- ↵
- Pich EM,
- Pagliusi SR,
- Тесари М,
- Talabot-Ayer D,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) Наука 275: 83 – 86.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Hiroi N,
- Nakabeppu Y,
- Надежда БТ,
- Nestler EJ
(1995) Mol Pharmacol 48: 880-889.
- ↵
- Hiroi N,
- Brown J,
- Е,
- Saudou F,
- Vaidya VA,
- Думан РС,
- Greenberg ME,
- Nestler EJ
(1998) J Neurosci 18: 6952 – 6962.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Chen J,
- Kelz MB,
- Надежда БТ,
- Nakabeppu Y,
- Nestler EJ
(1997) J Neurosci 17: 4933 – 4941.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Hiroi N,
- Brown J,
- Haile C,
- Е,
- Greenberg ME,
- Nestler EJ
(1997) Proc Natl Acad Sci САЩ 94: 10397 – 10402.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Fienberg AA,
- Hiroi N,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Ниши А,
- Cheramy A,
- O'Callaghan JP,
- Miller D,
- Cole DG,
- и др.
(1998) Наука 281: 838 – 842.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Hiroi N,
- Фейнберг А,
- Haile C,
- Greengard P,
- Nestler EJ
(1999) Eur J Neurosci 11: 1114-1118.
- ↵
- Greengard P,
- Allen PB,
- Nairn AC
(1999) Neuron 23: 435 – 447.
- ↵
- Bibb JA,
- Snyder GL,
- Ниши А,
- Ян З,
- Meijer L,
- Fienberg AA,
- Цай ЛХ,
- Kwon YT,
- Girault JA,
- Czernik AJ,
- и др.
(1999) Nature (Лондон) 402: 669 – 671.
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Sakai N,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Думан РС,
- Nestler EJ
(1998) Mol Pharmacol 54: 495-503.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Whisler K,
- Gilden L,
- Beckmann AM,
- Steffen C,
- Zhang YJ,
- Marotti L,
- Self SW,
- и др.
(1999) Nature (Лондон) 401: 272 – 276.
- ↵
- Dobrazanski P,
- Ногучи Т,
- Kovary K,
- Rizzo CA,
- Lazo PS,
- Браво R
(1991) Mol Cell Biol 11: 5470-5478.
Резюме / БЕЗПЛАТЕН Пълен текст
- Nakabeppu Y,
- Nathans D
(1991) Клетка 64: 751 – 759.
- ↵
- Йен Й,
- Мъдрост РМ,
- Tratner I,
- Verma IM
(1991) Proc Natl Acad Sci САЩ 88: 5077 – 5081.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Бял FJ,
- Hu XT,
- Zhang XF,
- Wolf ME
(1995) J Pharmacol Exp Ther 273: 445-454.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Hyman SE
(1996) Neuron 16: 901 – 904.
- Kreek MJ
(1997) Pharmacol Biochem Behav 57: 551 – 569.
- ↵
- Shippenberg TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449 – 455.
- ↵
- Chen JS,
- Zhang YJ,
- Kelz MB,
- Steffen C,
- Ang ES,
- Zeng L,
- Nestler EJ
(2000) J Neurosci 20: 8965 – 8971.
Резюме / БЕЗПЛАТЕН Пълен текст
- ↵
- Bibb JA,
- Chen JS,
- Тейлър Дж.
- Svenningsson P,
- Ниши А,
- Snyder GL,
- Ян З,
- Сагава ЗК,
- Nairn AC,
- Nestler EJ,
- и др.
(2001) Nature (Лондон) 410: 376 – 380.
- ↵
- Luscher C,
- Nicoll RA,
- Malenka RC,
- Мюлер Д
(2000) Nat Neurosci 3: 545 – 550.
- Malinow R,
- Mainen ZF,
- Hayashi Y
(2000) Curr Opin Neurobiol 10: 352 – 357.
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133 – 141.
Робинсън, TE и Колб, Б. (1999) (1997) Евро. J. Neurosci.11, 1598 1604.
- ↵
- Кери М,
- Smale ST
(2000) Регулиране на транскрипцията в еукариотите (Cold Spring Harbor Lab. Press, Plainview, NY).
- ↵
- Спенсър VA,
- Дейви Дж
(1999) Джин 240: 1 – 12.
 Facebook
Facebook- Twitter
- Google+
- CiteULike
- Много вкусен
 Digg
Digg- Mendeley
Статиите в HighWire Press са цитирани в тази статия
- Закон за природните и лекарствените възнаграждения относно механизмите за обща нервна пластичност с {Delta} FosB като ключов посредник J. Neurosci. 2013 33 (8) 3434-3442
- Наркотици, престъпност и епигенетика на хедоничната алостаза Вестник на съвременното наказателно правосъдие 2012 28 (3) 314-328
- абстрактен
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- абстрактен
- Пълен текст (HTML)
- Пълен текст (PDF)
- Морфинът активира E двадесет и шест-подобен транскрипционен фактор-1 / път на серумния реакционен фактор чрез екстрацелуларни кинази, регулирани от сигнала 1 / 2 в клетки F11, получени от ганглии неврони на дорзалния корен J. Pharmacol. Exp. Ther. 2012 342 (1) 41-52
- Молекулярният механизъм за лекарство на врата: Епигенетични промени, инициирани от експресия на никотинов първичен ген чрез кокаин Sci Transl Med 2011 3 (107) 107ra109
- Подобрена захароза и кокаин, самоуправление и предизвикани от Cue лекарства, търсещи загуба на VGLUT2 в невроните на средния мозъчен допамин в мишки J. Neurosci. 2011 31 (35) 12593-12603
- Хронична интермитентна хипоксия повишава кръвното налягане и експресията на FosB / {Delta} FosB в централните автономни области Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011 301 (1) R131-R139
- Липсата на GPR37 / PAEL рецептор влошава фосфорилирането на striatal Akt и ERK2, експресията на {Delta} FosB и обуславя предпочитанията към мястото на амфетамина и кокаина FASEB J. 2011 25 (6) 2071-2081
- Връзката между продължителността на първоначалното излагане на алкохол и устойчивостта на молекулярната толерантност е нелинейна. J. Neurosci. 2011 31 (7) 2436-2446
- In vivo биолуминисцентно изобразяване разкрива активаторния протеин-1, активиран при редокс, при паравентрикуларно ядро на мишки с реноваскуларна хипертония Хипертония 2011 57 (2) 289-297
- Стриатална свръхекспресия на {делта} FosB възпроизвежда хронична индуцирана от леводопа движения J. Neurosci. 2010 30 (21) 7335-7343
- Епигенетична медиация на въздействията върху околната среда при големи психотични нарушения Шизофър бик 2009 35 (6) 1045-1056
- ДНК-базирани МРТ сонди за специфично откриване на хронична експозиция на амфетамин в живи мозъци J. Neurosci. 2009 29 (34) 10663-10670
- Променена пластичност на дендритната гръбнака при извадени от кокаин плъхове J. Neurosci. 2009 29 (9) 2876-2884
- Екранът на свръхекспресия при дрозофила идентифицира невроналните роли на GSK-3 {бета} / рошак като регулатор на AP-1-зависима развитие на пластичността Генетика 2008 180 (4) 2057-2071
- MRI на транскрипция: нов изглед на живия мозък Невролог 2008 14 (5) 503-520
- {Делта} FosB индукция в орбитофронталната кора, медиира толерантността към кокаин-индуцираната когнитивна дисфункция J. Neurosci. 2007 27 (39) 10497-10507
- Продължителна уязвимост за възстановяване на търсенето на метамфетамин в митогенни митанти на невротрофичен фактор, получена от глиални клетки FASEB J. 2007 21 (9) 1994-2004
- {Delta} FosB в Nucleus Accumbens регулира усилването на инструменталното поведение и мотивацията J. Neurosci. 2006 26 (36) 9196-9204
- Регулиране на стабилността на фосфорилирането. J. Neurosci. 2006 26 (19) 5131-5142
- Експресия на мутантни NMDA рецептори в клетки, съдържащи рецептор на допамин D1, предотвратява сенсибилизирането на кокаина и намалява предпочитанията за кокаин J. Neurosci. 2005 25 (28) 6651-6657
- D1 Допаминови рецептори Модулират {делта} FosB индукция в щраума на плъх след интермитентен прием на морфин J. Pharmacol. Exp. Ther. 2005 314 (1) 148-154
- Невробиология на мишки, избрана за висока доброволна активност при движение на колела Integr. Comp. Biol. 2005 45 (3) 438-455
- Ефекти на лишаване от вода и рехидратация върху оцветяването с c-Fos и FosB в областта на надоптичното ядро на плъх и областта на ламиналните терминали Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005 288 (1) R311-R321
- Транскрипционна индукция на FosB / {Delta} FosB гена чрез механичен стрес в остеобластите J Biol Chem 2004 279 (48) 49795-49803
- Индукция на {делта} FosB в свързаните с възнаграждение мозъчни структури след хроничен стрес J. Neurosci. 2004 24 (47) 10594-10602
- Дозировката на Sim1 гена модулира хомеостатичния отговор на храненето към увеличените диетични мазнини при мишки Am. J. Physiol. Ендокринолог. Metab. 2004 287 (1) E105-E113
- Анализ на ДНК микрочипове на генната експресия в астроцитите на главата на човешкия оптичен нерв в отговор на хидростатичното налягане Physiol. Геномика 2004 17 (2) 157-169
- Супероксидът участва в активирането на централната нервна система и симпатичното възбуждане на сърдечната недостатъчност, предизвикана от инфаркт на миокарда Circ. Res. 2004 94 (3) 402-409
- Аденозинови А2А рецептори при невроадаптация към повтаряща се допаминергична стимулация: Последици за лечението на дискинезии при болестта на Паркинсон Неврология 2003 61 (90116) S74-81
- Цитоплазмена срещу ядрена локализация на Fos-свързани протеини в жабата, Rana esculenta, тестис: In Vivo и пряк ин витро ефект на гонадотропин-освобождаващ хормонален агонист Biol. Reprod. 2003 68 (3) 954-960
- Periadolescent мишките показват повишена регулация на Delta FosB в отговор на кокаин и амфетамин J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB регулира движението на колелата J. Neurosci. 2002 22 (18) 8133-8138
- Дейността на CREB в черупката nucleus accumbens контролира възникването на поведенчески реакции към емоционални стимули Proc. Natl. Акад. Sci. САЩ 2002 99 (17) 11435-11440
- Психогеномика: възможности за разбиране на зависимостта J. Neurosci. 2001 21 (21) 8324-8327