Behav Neurosci depan. 2015 Mei 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Abstrak
Dalam pengkondisian klasik, stimulus awalnya netral (stimulus terkondisi, CS) menjadi terkait dengan peristiwa yang menonjol secara biologis (stimulus tidak terkondisi, AS), yang mungkin berupa nyeri (pengkondisian permusuhan) atau makanan (pengkondisian nafsu makan). Setelah beberapa asosiasi, CS masing-masing dapat memulai respons defensif atau penyempurnaan. Berlawanan dengan pengkondisian permusuhan, pengkondisian nafsu makan jarang diteliti pada manusia, meskipun pentingnya untuk perilaku normal dan patologis (misalnya, obesitas, kecanduan) tidak dapat disangkal. Penelitian ini bermaksud untuk menerjemahkan temuan hewan tentang pengkondisian nafsu makan ke manusia yang menggunakan makanan sebagai AS. Tiga puluh tiga peserta diperiksa antara pukul 8 dan 10 pagi tanpa sarapan untuk memastikan bahwa mereka merasa lapar. Selama dua fase akuisisi, satu bentuk geometris (avCS +) memprediksi US yang tidak menyenangkan (sengatan listrik yang menyakitkan), bentuk lain (appCS +) memprediksi US selera (coklat atau pretzel asin sesuai dengan preferensi peserta), dan bentuk ketiga (CS– ) tidak memprediksikan AS. Dalam fase kepunahan, tiga bentuk ini ditambah bentuk baru (BARU) dihadirkan kembali tanpa pengiriman AS. Peringkat valensi dan gairah serta respons kejutan dan konduktansi kulit (SCR) dikumpulkan sebagai indeks pembelajaran. Kami menemukan pengkondisian permusuhan dan nafsu makan yang berhasil. Di satu sisi, avCS + dinilai lebih negatif dan lebih membangkitkan gairah daripada CS– dan menginduksi potensi kejutan dan peningkatan SCR. Di sisi lain, appCS + dinilai lebih positif daripada CS- dan menginduksi redaman kejut dan SCR yang lebih besar. Singkatnya, kami berhasil mengkonfirmasi temuan hewan pada manusia (lapar) dengan mendemonstrasikan pembelajaran nafsu makan dan pembelajaran permusuhan normal.
Pengantar
Memprediksi ancaman dan makanan sangat penting untuk kelangsungan hidup organisme apa pun. Dalam pengkondisian klasik (Pavlov, 1927), sebuah isyarat mendahului peristiwa permusuhan, seperti sengatan listrik ringan yang menyakitkan (rangsangan tak berkondisi permusuhan, AS), atau peristiwa nafsu makan, seperti pelet makanan (makanan AS), beberapa kali. Setelah itu, rangsangan ini saja (sekarang diberi label CS, CS +) masing-masing dapat memperoleh respons defensif atau penyempurnaan. Jenis pembelajaran asosiatif yang pertama disebut pengkondisian permusuhan, sedangkan yang terakhir disebut pengkondisian selera.
Meskipun prediksi peristiwa nafsu makan sama pentingnya bagi kelangsungan hidup seperti prediksi kejadian permusuhan, pengkondisian nafsu makan sangat kurang diselidiki pada hewan (Bouton dan Peck, 1989; Koch et al., 1996; McDannald et al., 2011, untuk ulasan lihat Martin-Soelch et al., 2007) serta pada manusia (Klucken et al., 2009, 2013; Austin dan Duka, 2010; Delgado et al., 2011; Levy dan Glimcher, 2011). Kurangnya penelitian ini mungkin disebabkan oleh kompleksitas paradigma nafsu makan dibandingkan dengan paradigma permusuhan. Misalnya, makanan sebagai penguat utama harus dikirimkan ketika organisme lapar agar mendapat imbalan (untuk ulasan baru lihat Dickinson dan Balleine, 1994; Clark et al., 2012). Dalam penelitian manusia, kesulitan ini telah diatasi dengan menggunakan uang (Austin dan Duka, 2010; Delgado et al., 2011; Levy dan Glimcher, 2011) atau gambar erotis (Klucken et al., 2009, 2013). Namun, studi neuro-imaging telah menunjukkan bahwa penguat primer (yaitu, makanan ringan atau minuman) dan sekunder (yaitu, uang) mengaktifkan beberapa daerah otak yang umum (misalnya, striatum) tetapi juga memerlukan pola aktivasi yang berbeda (Delgado et al., 2011; Levy dan Glimcher, 2011).
Sepengetahuan kami, hanya beberapa studi pengkondisian manusia yang meneliti efek dari penguat nafsu makan utama seperti bau (Gottfried et al., 2002), air (Kumar et al., 2008), atau makanan (Prévost et al., 2012). Dalam studi pertama, Gottfried et al. (2002) wajah netral terkait (CSs) dengan bau tidak menyenangkan, menyenangkan, atau netral. Menariknya, mereka menemukan aktivasi yang lebih besar di korteks orbitofrontal (OFC) dan di ventral striatum sebagai respons terhadap CS + nafsu makan versus CS + permusuhan. Mereka menyimpulkan bahwa OFC memproses nilai bau dan terlibat dalam mentransfer nilai afektif dari penciuman (AS) ke sistem visual (CS). Aktivasi striatal (ventro) telah diinterpretasikan sebagai mencerminkan CR nafsu makan yang ditimbulkan oleh CS + nafsu makan. Dalam studi kedua, Kumar et al. (2008) mengundang peserta mereka (kontrol sehat dan pasien dengan depresi berat) ke laboratorium pagi-pagi dan meminta mereka untuk tidak minum pada malam hari untuk memastikan bahwa mereka haus sebelum pemindaian. Gambar fraktal adalah CSs dan 0.1 ml air AS. Menariknya, di antara aktivasi lainnya, peserta yang sehat (tetapi bukan pasien depresi) menunjukkan aktivasi yang lebih besar di ventral striatum ke CS +, menunjukkan bahwa stimulus ini diproses sebagai hadiah. Dalam studi ketiga, Prévost et al. (2012) juga menampilkan gambar fraktal sebagai CS dan snack baik yang manis maupun asin sesuai US sesuai preferensi peserta. CS ditampilkan selama 6 detik dan selama detik terakhir gambar makanan juga ditampilkan. Setiap kali gambar makanan muncul, peneliti meletakkan sepotong makanan di tangan partisipan, yang diizinkan untuk langsung mengonsumsi camilan tersebut. Sayangnya, penulis tidak melaporkan aktivasi otak ke CS + nafsu makan selama fase pengkondisian klasik; Namun, mereka mengamati lebih sedikit perlambatan jantung pada CS + yang diberi penghargaan dibandingkan dengan CS yang tidak diberi penghargaan, yang menunjukkan efek pengkondisian yang berbeda.
Kurangnya penelitian yang menggunakan respon mengejutkan sebagai indeks untuk pengkondisian nafsu makan adalah mengejutkan, terutama mengingat penggunaannya yang luas dalam pengkondisian permusuhan. Respons mengejutkan adalah respons defensif leluhur dan otomatis terhadap peristiwa permusuhan yang mendadak, tidak terduga, dan kuat (Koch, 1999). Respons defensif ini dimediasi oleh jalur neuronal yang relatif sederhana yang melibatkan neuron akar koklea, inti pontine caudal dari formasi reticular (PnC) dan motoneuron spinal (Fendt dan Fanselow, 1999; Koch, 1999). Penelitian pada hewan telah mengungkapkan bahwa potensiasi mengejutkan tergantung pada proyeksi dari amigdala ke PnC (Fendt dan Fanselow, 1999; Koch, 1999), sedangkan redaman kejutan tergantung pada nucleus accumbens yang utuh (NAcc, Koch, 1999). Modulasi respons kejutan semacam itu merupakan ukuran implisit yang berguna untuk valensi rangsangan latar depan yang disajikan. Dengan demikian, potensiasi menunjukkan valensi negatif, sedangkan atenuasi menunjukkan valensi positif, keduanya tanpa sangat dipengaruhi oleh proses kognitif (Hamm dan Weike, 2005; Andreatta et al., 2010). Sepengetahuan kami, hanya satu penelitian pada hewan yang menyelidiki pengkondisian nafsu makan dengan mengukur respons yang mengejutkan sebagai ukuran dependen. Menariknya, tikus yang diperiksa menunjukkan redaman mengejutkan ke CS + (yaitu, cahaya) terkait dengan pengiriman larutan sukrosa. Khususnya, pelemahan seperti itu terganggu pada hewan-hewan dengan lesi NAcc, tetapi tidak pada mereka yang lesi amigdala, menunjukkan bahwa NACC memainkan peran spesifik dalam memunculkan CR nafsu makan dan dalam melemahkan respons mengejutkan (Koch et al., 1996).
Studi saat ini adalah untuk pengetahuan kita yang pertama untuk menerjemahkan paradigma pengkondisian selera ini kepada manusia dengan menggunakan penguat utama seperti AS, yaitu manis (cokelat Smarties).®) atau makanan asin (pretzel asin kecil), dan modulasi mengejutkan sebagai ukuran CRs. Kami berharap appCS + untuk memicu CR selera yang kuat seperti tercermin dalam pelemahan mengejutkan, peningkatan SCR, dan peringkat valensi positif dibandingkan dengan rangsangan lainnya, yaitu, avCS +, dan CS-.
Bahan dan Metode
Peserta
Empat puluh dua sukarelawan diterima untuk berpartisipasi dalam penelitian ini dan menerima kredit kursus. Sembilan peserta dikeluarkan dari analisis, dua karena masalah teknis, tiga karena mereka dikodekan sebagai non-responden (rata-rata amplitudo kejut <5 μV), dan empat karena mereka tidak menunjukkan tanggapan yang cukup mengejutkan per kondisi (minimum = 2; untuk detailnya, lihat Bahan dan Metode). Pada akhirnya, 33 peserta dipertimbangkan dalam analisis (16 laki-laki; usia rata-rata: 22.09 tahun, SD: 2.84; kisaran: 18 – 29 tahun). Empat peserta bukan orang Jerman asli, dan enam orang kidal. Satu peserta tetap tidak mengetahui hubungan CSs-USs selama percobaan (lihat Prosedur), tetapi kami memutuskan untuk tidak mengecualikan peserta ini karena tanggapannya normal dan tidak mempengaruhi hasil.
bahan
Stimuli Tanpa Syarat (AS)
Dua jenis AS digunakan. Sebagai pembenci AS, kami menerapkan sengatan listrik yang agak menyakitkan pada lengan peserta yang tidak dominan. Sengatan listrik disampaikan melalui dua elektroda dengan diameter 9 mm dan jarak 30 cm. Stimulus listrik terdiri dari stimulus pulsa dengan frekuensi 50 Hz dan durasi 200 ms, yang dihasilkan oleh stimulator saat ini (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Inggris, 400 V, maksimum 9.99 mA). Intensitas sengatan listrik ditentukan secara individual melalui prosedur ambang yang dijelaskan sebelumnya (Andreatta et al., 2010). Secara singkat, peserta menjalani dua seri intensitas naik dan turun dalam interval 0.5 mA. Mereka harus menilai setiap stimulus pada skala visual mulai dari 0 (tidak merasakan apa-apa) hingga 10 (sakit sangat intens) dengan 4 sebagai jangkar untuk ambang (hanya rasa sakit yang terlihat). Intensitas rata-rata stimulus listrik adalah 2.12 mA (SD = 0.56) dan dinilai menyakitkan (M = 6.45, SD = 1.73). Makanan pembuka AS terdiri dari cokelat (Smarties®) atau pretzel asin kecil. Pilihan selera AS tergantung pada preferensi individu peserta seperti yang dilaporkan selama wawancara pendahuluan. Yakni, peserta harus melaporkan apakah biasanya mereka makan makanan asin atau manis saat sarapan. Mereka juga dapat dengan bebas memilih apakah mereka lebih suka cokelat atau pretzel asin selama percobaan berlangsung. Pada akhirnya, 22 peserta memilih coklat dan 11 pretzel kecil asin.
Stimuli Terkondisikan (CS)
Bentuk geometris (kotak biru, lingkaran kuning, segitiga hijau, heksagon merah) dengan diagonal 8 cm disajikan sebagai CS. Bentuk disajikan di tengah-tengah layar komputer hitam untuk 8 s. Satu bentuk (avCS +) selalu dikaitkan dengan AS permusuhan (sengatan listrik menyakitkan), satu bentuk (appCS +) selalu dikaitkan dengan selera AS (coklat atau pretzel asin), satu bentuk (CS-) tidak pernah dikaitkan dengan salah satu dari permusuhan atau selera AS, dan bentuk keempat (BARU) disajikan dalam fase kepunahan tetapi tidak selama fase akuisisi untuk memastikan netralitasnya.
Probe mengejutkan
Kebisingan putih 103 dB dengan durasi 50 ms digunakan sebagai probe kejut. Stimulus akustik disajikan secara binaural melalui headphone dan terjadi secara acak 4-6 detik setelah onset bentuk.
Kuesioner
Sebelum dan sesudah percobaan, peserta harus mengisi versi Jerman dari State-Trait Anxiety Inventory (STAI, Laux et al., 1981) dan Jadwal Pengaruh Negatif Positif (PANAS, Krohne et al., 1996). STAI adalah inventaris untuk menilai sifat peserta dan / atau kecemasan negara dan terdiri dari 20 item untuk sifat dan versi negara bagian. Tingkat kecemasan peserta sebelumnya (M = 37.06, SD = 7.80) dan setelah (M = 39.33, SD = 9.16) percobaan tidak berubah secara signifikan [t(32) = 1.61, p = 0.117]. Skor kecemasan sifat dalam sampel saat ini berkisar antara 20 dan 58 (M = 36.6, SD = 8.98), yang sebanding dengan kisaran normal orang dewasa yang dipublikasikan (Laux et al., 1981). PANAS (Krohne et al., 1996) adalah indeks suasana hati positif dan negatif. Individu dengan skor tinggi pada skala pengaruh positif (PAS) rentan terhadap emosi seperti antusiasme, sedangkan individu dengan skor tinggi pada skala pengaruh negatif (NAS) rentan terhadap emosi seperti kesusahan. Setiap item terdiri dari kata sifat, dan peserta menunjukkan pada skala mulai dari 1 (sangat sedikit) hingga 5 (sangat) sejauh mana kata sifat tersebut mencerminkan perasaan mereka pada saat itu. Tidak ada perbedaan signifikan yang ditemukan pada pengaruh negatif peserta sebagai hasil dari percobaan [mulai: M = 11.67, SD = 2.29; akhir: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Entah bagaimana, peserta mengubah suasana hati positif mereka sejak awal (M = 26.72, SD = 4.70) hingga akhir (M = 23.66, SD = 7.02) dari percobaan [t(31) = 3.11, p = 0.004]. Penurunan mood positif peserta mungkin terkait dengan ketidaknyamanan paradigma (sengatan listrik yang menyakitkan serta kebisingan putih permusuhan disajikan).
Prosedur
Setibanya di laboratorium, peserta membaca dan menandatangani formulir informed consent yang disetujui oleh komite etika Departemen Psikologi Universitas Würzburg. Mereka tidak diberitahu tentang kontingensi antara CS dan AS. Setelah mengisi kuesioner, elektroda dilampirkan dan prosedur ambang nyeri dilakukan seperti yang dijelaskan di atas.
Selama fase pembiasaan, empat bentuk geometris disajikan dua kali dengan interval antar-percobaan (ITI) bervariasi antara 18 dan 25 s (rata-rata: 21.5 s). Tidak ada AS atau probe mengejutkan yang dikirimkan selama fase ini.
Sebelum fase akuisisi, tujuh serangan white noise dikirimkan setiap 7 – 15 untuk mengurangi reaktivitas guncangan awal.
Berikut dua fase akuisisi identik (Gambar 1). Setiap fase akuisisi terdiri dari uji coba 24: uji coba 8– uji coba, uji coba 8 avCS +, dan uji coba aplikasi 8 appCS +. Urutan CS adalah pseudorandom dengan satu-satunya batasan adalah bahwa stimulus yang sama tidak dapat disajikan lebih dari dua kali berturut-turut. Khususnya, avCS + disajikan bersama dengan baut petir sebagai simbol untuk sengatan listrik dan AS yang menyakitkan disampaikan pada offset. AppCS + disajikan dalam campuran dengan gambar Smarties atau pretzel asin dan peserta dapat memilih Smarties atau pretzel dari toples. CS– disajikan bersama dengan simbol larangan dan tidak ada AS yang dikirim. Selama tiga presentasi 8 CS dari masing-masing jenis, penyelidikan mengejutkan disampaikan antara 4 dan 6 setelah onset stimulus. Tiga probe kejutan tambahan disajikan selama ITI untuk memastikan ketidakpastian mereka dan untuk mengurangi pembiasaan mengejutkan. ITI, yang terdiri dari layar hitam, bervariasi antara 18 dan 25 dengan rata-rata 21.5.
GAMBAR 1
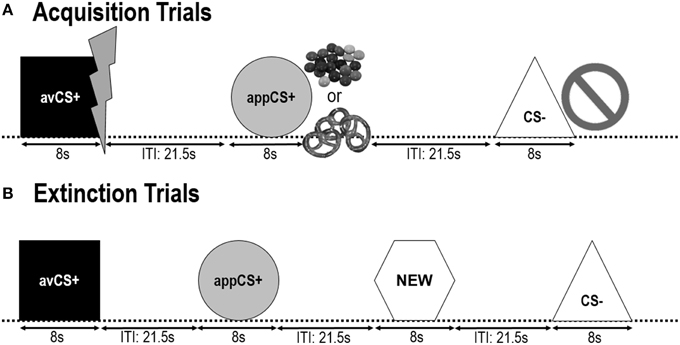
Gambar 1. Uji coba selama dua fase akuisisi (A) dan fase kepunahan (B). Peserta belajar bahwa satu bentuk (avCS +) memprediksi sengatan listrik ringan yang menyakitkan, satu bentuk (appCS +) memprediksi baik sepotong coklat atau pretzel asin kecil (sesuai dengan preferensi mereka), dan bentuk ketiga (CS-) tidak memprediksi adanya acara yang menonjol secara biologis. Setiap bentuk disajikan bersama dengan gambar yang menggambarkan sengatan listrik, smarties / pretzel asin, atau tidak tergantung pada hubungan dengan AS. Selama fase kepunahan, tiga bentuk geometris disajikan sekali lagi, tetapi tidak ada AS yang dikirim. Selain itu, bentuk geometris keempat (BARU) disajikan sebagai kontrol netral.
Selama fase kepunahan, peserta melihat tiga bentuk geometris (yaitu, avCS +, appCS +, dan CS-) lagi ditambah bentuk netral baru (BARU). Tidak ada US dikirim dan bentuk tidak disajikan bersama dengan gambar flash, coklat / asin pretzel, atau larangan. Setiap stimulus disajikan delapan kali dalam urutan pseudorandom (yaitu, stimulus yang sama tidak disajikan lebih dari dua kali berturut-turut), menciptakan uji coba 32. Stimulus probe rangsangan disajikan selama 4 keluar dari presentasi stimulus 8 dari masing-masing jenis CS. Seperti pada fase akuisisi dan pembiasaan, ITI bervariasi antara 18 dan 25, dan 4 probe kejutan tambahan dikirimkan secara tak terduga selama ITI.
Setelah setiap fase, peserta menilai valensi (kesenangan) dan gairah (rangsang) CSs dengan skala analog visual (VAS) mulai dari 1 ke 9. Skala kelambu berkisar dari "1" yang menunjukkan "sangat tidak menyenangkan" hingga "9" yang menunjukkan "sangat menyenangkan;" skala gairah berkisar dari "1" yang menunjukkan "tenang" hingga "9" yang menunjukkan "menarik." Selain itu, peringkat kontingensi adalah dinilai setelah dua fase akuisisi dan fase kepunahan. Peserta melihat bentuk geometris untuk 1 dan kemudian mereka harus menunjukkan apakah bentuk ini dikaitkan dengan sengatan listrik, dengan pretzel cokelat / asin, tanpa apa-apa, atau apakah mereka tidak dapat membuat hubungan apa pun. Khususnya, semua peserta (kecuali satu) menyadari kemungkinan setelah Akuisisi 2.
Pengurangan Data
Respons fisiologis direkam dengan penguat V-Amp 16 dan Software Perekam Visi V-Amp Edition (Versi 1.03.0004, BrainProducts Inc., Munich, Jerman). Tingkat sampling 1000 Hz dan filter takik 50 Hz diterapkan. Analisis offline dilakukan dengan Brain Vision Analyzer (Versi 2.0; BrainProducts Inc., Munich, Jerman).
Respon yang mengejutkan
Respon mengejutkan diukur dengan menggunakan electromyography (EMG) di sebelah kiri Otot kelopak mata otot dengan dua elektroda Ag / AgCl 5 mm. Sesuai dengan pedoman (Blumenthal et al., 2005), satu elektroda diposisikan di bawah pupil dan yang kedua 1 cm lateral. Elektroda ground dan referensi ditempatkan di masing-masing mastoid kanan dan kiri. Sebelum memasang elektroda, kulit dikelupas ringan dan dibersihkan dengan alkohol untuk menjaga impedansi di bawah 10 kΩ. Sinyal electromyographic secara offline difilter dengan filter cutoff rendah 28 Hz dan filter cutoff tinggi 500 Hz. Kemudian, sinyal EMG diperbaiki dan rata-rata bergerak 50 ms diterapkan. Kami menggunakan 50 ms sebelum onset probe awal sebagai baseline (Grillon et al., 2006). Respons terhadap probe mengejutkan diberi skor secara manual, dan uji coba dengan pergeseran baseline yang berlebihan (± 5 μV) atau artefak gerakan dikeluarkan dari analisis lebih lanjut. Respons kejutan lebih rendah dari 5 μV dikodekan sebagai nol dan dipertimbangkan untuk perhitungan besarnya kejutan (Blumenthal et al., 2005). Secara keseluruhan, 10.4% percobaan ditolak, dan minimum 2 dari tanggapan mengejutkan 3 dalam fase akuisisi dan 4 dari tanggapan mengejutkan 8 dalam fase kepunahan untuk setiap kondisi diperlukan untuk menjaga peserta dalam kumpulan analisis. Karena alasan ini, empat peserta dikeluarkan. Amplitudo puncak didefinisikan sebagai puncak maksimum relatif terhadap garis dasar selama jendela waktu 20-120 ms setelah onset probe tiba-tiba. Data mentah kemudian dinormalisasi dalam-mata pelajaran menggunakan zskor dan skor-T untuk mengurangi pengaruh variabilitas individu dan untuk mendeteksi proses psikologis dengan lebih baik. Skor-T rata-rata untuk setiap kondisi (avCS +, appCS +, CS–, BARU, dan ITI). Untuk menyelidiki potensi mengejutkan atau pelemahan mengejutkan, skor untuk respons mengejutkan ITI dikurangi dari respons kejutan untuk setiap kondisi.
Skin Conductance Response (SCR)
Skin conductance response (SCR) direkam menggunakan dua elektroda 5 mm Ag / AgCl yang diletakkan di telapak tangan yang tidak dominan. Respons galvanik offline disaring dengan filter cutoff tinggi 1 Hz. SCR didefinisikan sebagai perbedaan (dalam μS) antara onset respons (1-3 s setelah onset stimulus) dan puncak respons (Tranel dan Damasio, 1994; Delgado et al., 2011). Uji coba yang mengandung probe mengejutkan tidak dipertimbangkan dalam analisis SCR. Respons di bawah 0.02 μS dikodekan sebagai nol. Lima peserta lebih lanjut dikeluarkan dari analisis SCR karena mereka memiliki rata-rata SCR lebih rendah dari 0.02 μS. Data konduktansi kulit mentah ditransformasikan akar kuadrat untuk menormalkan distribusi dan skor dirata-rata untuk setiap kondisi secara terpisah untuk dua fase akuisisi (avCS +, appCS +, CS-) dan fase kepunahan (avCS +, appCS +, CS-, dan NEW ).
Analisis Statistik
Semua data dianalisis dengan SPSS untuk Windows (Versi 20.0, SPSS Inc.). Untuk respon fisiologis, analisis varians multivariat yang terpisah (ANOVA) dihitung untuk dua fase akuisisi dan fase kepunahan. ANOVA untuk fase akuisisi memiliki stimulus (avCS +, appCS +, CS-) dan fase (Akuisisi 1, Akuisisi 2) sebagai faktor dalam mata pelajaran. ANOVA untuk fase kepunahan hanya memiliki stimulus (avCS +, appCS +, CS–, NEW) sebagai faktor dalam mata pelajaran. Peringkat valensi, gairah, dan kontingensi dianalisis dengan ANOVA terpisah yang berisi stimulus faktor dalam-subyek (avCS +, appCS +, CS-, dan BARU) dan fase. Faktor ini memiliki empat level untuk peringkat valensi dan gairah (T1: setelah fase pembiasaan, T2: setelah fase akuisisi pertama, T3: setelah fase akuisisi kedua, T4: setelah fase kepunahan), tetapi tiga level untuk peringkat kontingensi (T1: setelah fase akuisisi pertama, T2: setelah fase akuisisi kedua, T3: setelah fase kepunahan).
Tingkat alpha (α) ditetapkan pada 0.05 untuk semua analisis. Ukuran efek dilaporkan sebagai parsial η2.
Hasil
Peringkat valensi dan gairah untuk setiap fase digambarkan pada Gambar 2; tanggapan mengejutkan dan SCR digambarkan dalam Gambar 3.
GAMBAR 2
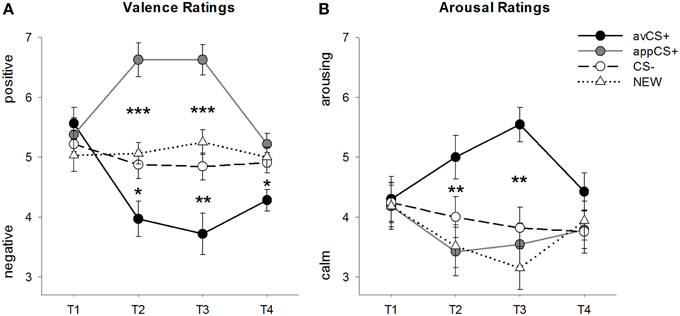
Gambar 2. Peringkat untuk valensi (A) dan gairah (B). Garis (dengan kesalahan standar) menggambarkan peringkat setelah fase habituasi (T1), Akuisisi 1 (T2), Akuisisi 2 (T3), dan fase kepunahan (T4). CS + (garis hitam solid) permusuhan diperoleh valensi negatif dan gairah tinggi setelah dua fase akuisisi dibandingkan dengan CS- (garis putus-putus hitam) dan BARU (garis putus-putus hitam). Yang penting, CS + (garis abu-abu solid) selera makan memperoleh valensi positif dibandingkan dengan CS- dan BARU. *p <0.05, **p > 0.01, ***p <0.001.
GAMBAR 3
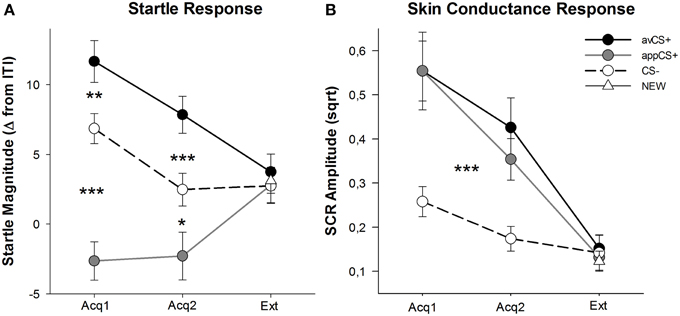
Gambar 3. Respons mengejutkan (A) dan konduktansi kulit (B) (dengan kesalahan standar) selama fase akuisisi pertama (Acq1), fase akuisisi kedua (Acq2), dan fase kepunahan (Ext). Respons yang mengejutkan secara signifikan dipotensiasi ke CS + permusuhan (garis solid hitam) dan secara signifikan dilemahkan ke CS + (garis abu-abu solid) selera dibandingkan dengan CS- (garis putus-putus hitam) selama fase akuisisi. SCR secara signifikan lebih besar pada avCS + dan appCS + dibandingkan dengan CS–. Tidak ada perbedaan yang terungkap selama fase kepunahan. *p <0.05, **p > 0.01, ***p <0.001.
Peringkat
ANOVA untuk peringkat valensi selama akuisisi mengungkapkan efek utama yang signifikan dari stimulus [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, sebagian η2 = 0.358] dan fase [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, parsial η2 = 0.096] serta interaksi yang signifikan antara stimulus dan fase [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, sebagian η2 = 0.235]. Mengikuti t-test menunjukkan bahwa valensi dari empat bentuk geometris pada awal percobaan identik (ps> 0.19), sementara setelah Akuisisi 1 dan Akuisisi 2, avCS + dinilai sebagai negatif khususnya dan appCS + sangat positif. Secara khusus, avCS + dinilai lebih bervalensi negatif dibandingkan dengan CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], BARU [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001], dan appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + dinilai secara signifikan lebih positif daripada CS- [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] dan BARU [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Perbedaan antara CS– dan NEW tidak pernah signifikan (ps> 0.18).
Yang penting, tidak ada perbedaan signifikan yang ditemukan ketika membandingkan peringkat valensi untuk appCS + yang terkait dengan cokelat vs pretzel asin [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Setelah fase kepunahan, avCS + masih dinilai lebih negatif dibandingkan dengan CS- [t(31) = 2.40, p = 0.023], BARU [t(31) = 3.40, p = 0.002], dan appCS + [t(31) = 3.35, p = 0.002]. Sebaliknya, valensi appCS + tidak berbeda lagi dari CS– [t(31) = 1.77, p = 0.086] atau BARU [t(31) = 1.07, p = 0.293].
ANOVA untuk peringkat gairah selama akuisisi mengungkapkan efek utama yang signifikan dari stimulus [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, parsial η2 = 0.181], tetapi tidak dari fase [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, parsial η2 = 0.038], dan interaksi yang signifikan antara stimulus dan fase [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, parsial η2 = 0.124]. Mengikuti t-test menunjukkan tidak ada perbedaan yang signifikan antara rangsangan mengenai gairah awal mereka (ps> 0.74). Namun, setelah dua fase akuisisi, avCS + dinilai lebih menggairahkan daripada yang BARU [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] dan appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001], dan sedikit lebih membangkitkan gairah dibandingkan CS– setelah fase akuisisi pertama [t(32) = 1.96, p = 0.058], tetapi secara signifikan lebih membangkitkan setelah fase akuisisi kedua [t(32) = 3.65, p = 0.001]. Berbeda dengan peringkat valensi, gairah appCS + tidak berbeda dari gairah CS– dan BARU (ps > 0.13) setelah fase akuisisi.
Demikian pula dengan peringkat valensi, CS- dan BARU tidak berbeda dalam gairah mereka (ps> 0.07).
Seperti halnya valensi, tidak ada perbedaan yang terungkap untuk appCS + yang terkait dengan cokelat dan appCS + yang terkait dengan pretzel asin [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Setelah fase kepunahan, tidak ada perbedaan signifikan dalam peringkat gairah yang terungkap (ps> 0.08).
Untuk tambahan post-hoc t- pengujian membandingkan peringkat setelah fase pembiasaan, fase akuisisi pertama dan kedua dan fase kepunahan, lihat Bahan Tambahan.
Respon yang mengejutkan
ANOVA untuk fase akuisisi menghasilkan efek utama dari stimulus [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, sebagian η2 = 0.609], tetapi tidak fase [F(1, 32) = 3.16, p = 0.085, parsial η2 = 0.090], dan interaksi yang signifikan Stimulus × Phase [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, parsial η2 = 0.095]. Mengikuti t-pengujian mengungkapkan potensi mengejutkan signifikan untuk avCS + dibandingkan dengan CS– selama kedua [t(32) = 3.27, p = 0.003] dan yang kedua [t(32) = 4.00, p <0.001] fase akuisisi. Respon mengejutkan terhadap avCS + juga secara signifikan berpotensi dibandingkan dengan appCS +, sekali lagi setelah kedua Akuisisi 1 [t(32) = 8.20, p <0.001] dan Akuisisi 2 [t(32) = 5.74, p <0.001]. Yang penting, besarnya kejut pada appCS + secara signifikan dilemahkan dibandingkan dengan CS– selama kedua Akuisisi 1 [t(32) = 6.34, p <0.001] dan Akuisisi 2 [t(32) = 2.91, p = 0.007]. Sekali lagi dan sesuai dengan peringkat, tidak ada respons kejut diferensial yang terungkap untuk appCS + ketika dikaitkan dengan cokelat atau dengan pretzel asin [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Selama fase kepunahan, tidak ada efek signifikan yang ditemukan [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, parsial η2 = 0.008].
Skin Conductance Response (SCR)
Dari ANOVA untuk SCR selama dua fase akuisisi, efek utama stimulus [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, sebagian η2 = 0.401] dan fase [F(1, 27) = 20.91, p <0.001, sebagian η2 = 0.436] ternyata signifikan, tetapi bukan interaksinya [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, parsial η2 = 0.024]. Pasca-hoc t-percobaan menunjukkan SCR yang lebih besar secara signifikan ke avCS + [t(27) = 6.46, p <0.001] dan ke appCS + [t(27) = 4.84, p <0.001] dibandingkan dengan CS–, sementara peserta menunjukkan SCR yang sebanding dengan avCS + dan appCS + [t(27) = 0.64, p = 0.527]. Khususnya, tidak ada perbedaan dalam SCRs dengan appCS + coklat dan ke appCS + asin asin yang ditemukan [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Seperti yang benar untuk peringkat dan respon mengejutkan, tidak ada efek signifikan yang ditemukan untuk fase kepunahan [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, parsial η2 = 0.010].
Diskusi
Tujuan dari penelitian ini adalah untuk menerjemahkan temuan hewan kepada manusia dengan menggunakan paradigma pengkondisian nafsu makan klasik dengan penguat utama sebagai stimulus tanpa syarat (AS). Untuk melakukannya, peserta datang ke lab pagi-pagi sekali tanpa sarapan untuk memastikan bahwa mereka lapar, dan sesuai dengan preferensi mereka, potongan cokelat atau pretzel asin digunakan sebagai makanan pembuka AS. Selama fase akuisisi, satu bentuk geometris (avCS +) menjadi terkait dengan sengatan listrik yang agak menyakitkan (AS permusuhan), bentuk lain (appCS +) dengan selera AS, dan bentuk ketiga (CS-) dengan tidak ada selera AS atau dengan US benci. Hasil-hasil menunjukkan keberhasilan yang tidak menyenangkan dan pengkondisian nafsu makan pada tingkat verbal eksplisit (yaitu, peringkat), pada tingkat perilaku implisit (yaitu, respons mengejutkan), dan pada tingkat fisiologis (yaitu, SCR). Secara khusus, avCS + dibandingkan dengan CS- memperoleh lebih banyak peringkat valensi negatif, peringkat gairah yang lebih tinggi, potensiasi yang mengejutkan, dan SCR yang lebih besar. Yang paling penting, appCS + dibandingkan dengan CS– memicu peringkat valensi yang lebih positif, redaman yang mengejutkan, dan SCR yang lebih besar. Temuan kami pada pengkondisian permusuhan sejalan dengan harapan, karena penelitian sebelumnya telah menemukan bahwa ancaman prediksi stimulus (avCS +) dinilai sebagai permusuhan, memunculkan respons ketakutan yang lebih besar, dan meningkatkan gairah fisiologis (Fendt dan Fanselow, 1999; Hamm dan Weike, 2005; Andreatta et al., 2010, 2013). Hasil kami pada pengkondisian nafsu makan juga sejalan dengan penelitian manusia dan hewan sebelumnya yang menunjukkan bahwa hadiah prediksi stimulus (appCS +) dinilai sebagai positif, menghambat respons rasa takut, dan meningkatkan gairah fisiologis (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin dan Duka, 2010; Prévost et al., 2012). Sepengetahuan kami, ini adalah studi pertama yang menunjukkan pelemahan mengejutkan yang dikondisikan pada manusia terhadap stimulus yang memprediksi hadiah utama. Yang penting, kami dapat mentransfer dan mengkonfirmasi hasil studi hewan (Koch et al., 1996). Studi hewan ini menunjukkan bahwa pelemahan mengejutkan pada tikus tergantung pada proyeksi dari NACC (bagian dari ventral striatum) ke PnC. Oleh karena itu, pelemahan mengejutkan dalam penelitian kami dapat menyiratkan aktivitas NACC, yang juga sejalan dengan temuan fMRI (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy dan Glimcher, 2011). Oleh karena itu, kami menyimpulkan bahwa paradigma pengkondisian nafsu makan kami berhasil, seperti yang ditunjukkan oleh valensi positif (penilaian) dan implisit (pelemahan mengejutkan).
Selain temuan baru namun agak dapat diprediksi ini, dua hasil menarik lebih lanjut perlu disebutkan. Pertama, respons rangsangan verbal dan fisiologis terhadap appCS + dipisahkan. Kedua, kami menemukan kepunahan yang lebih cepat dari CR selera dibandingkan dengan CR permusuhan.
SCR menunjukkan aktivasi simpatik, yang meningkat baik untuk stimulus yang terkait dengan hadiah (appCS +) dan stimulus yang terkait dengan ancaman (avCS +). Hasil ini sejalan dengan studi pengkondisian sebelumnya, di mana gambar erotis digunakan sebagai selera AS (Klucken et al., 2013), dan sebuah penelitian mengungkapkan SCR sebanding dengan isyarat yang memprediksi uang dan isyarat yang memprediksi kebisingan permusuhan (Austin dan Duka, 2010). Khususnya, SCR adalah respons berorientasi yang terkait dengan aktivasi sistem simpatik. Tanggapan ini telah disarankan untuk mencerminkan persiapan reaksi perilaku terhadap peristiwa yang menonjol secara motivasi (Bradley, 2009). Berdasarkan hal ini, kami berpikir bahwa baik stimuli yang berhubungan dengan ancaman maupun yang terkait dengan hadiah menimbulkan respons persiapan untuk respons perilaku yang berurutan. Dengan kata lain, sinyal makanan dan rasa sakit memulai persiapan untuk pendekatan dan perilaku menghindar, masing-masing. Berbeda dengan rangsangan fisiologis yang tinggi, respons verbal menunjukkan rangsangan rendah terhadap stimulus yang terkait dengan hadiah. Ada kemungkinan bahwa gairah verbal mungkin lebih dipengaruhi oleh sifat membangkitkan dari AS daripada aktivasi fisiologis sendiri. Bahkan, kami berpikir bahwa appCS + vs avCS + dinilai rendah vs sangat membangkitkan gairah karena mereka terkait dengan masing-masing AS yang rendah dan sangat membangkitkan gairah. Sayangnya, kami tidak mengumpulkan peringkat gairah untuk AS, dan oleh karena itu akan tergantung pada studi di masa depan untuk menguji hipotesis ini secara eksplisit.
Selama fase kepunahan, tidak ada AS yang dikirim. Ini mungkin telah menginduksi pembelajaran penghambatan baru yang disebut kepunahan (untuk tinjauan lihat Milad dan Quirk, 2012), dengan menurunnya CR permusuhan dan selera makan sebagai konsekuensinya. Jelasnya, pada perilaku (yaitu, respons yang mengejutkan) dan pada tingkat fisiologis (yaitu, SCR), tidak ada reaksi diferensial terhadap avCS +, appCS +, dan CS- yang terdeteksi lagi. Demikian pula, gairah eksplisit rangsangan terkondisi dinilai setelah fase kepunahan menyamakan pada tingkat rendah, menunjukkan keberhasilan pembelajaran kepunahan. Namun, avCS + masih dinilai secara signifikan lebih negatif daripada CS–, sedangkan valensi appCS + tidak berbeda dari CS-valensi lagi. Kepunahan yang lebih lambat dari respons eksplisit permusuhan mungkin disebabkan oleh konservatisme evolusioner, yang berarti bahwa sinyal ancaman sangat sulit untuk dilupakan karena non-respons terhadap sinyal ancaman mungkin mengancam jiwa.
Masih ada satu pertanyaan: mengapa kemudian respon mengejutkan (yaitu, valensi implisit) padam sepenuhnya selama fase kepunahan? Pertama, tanggapan ini dihitung selama seluruh fase. Oleh karena itu, dapat dibayangkan bahwa CR diskriminatif akan tetap dapat dideteksi selama uji coba pertama fase kepunahan. Secara eksploratif, kami mengikuti hipotesis ini dan melihat tanggapan mengejutkan dan SCR sepanjang fase kepunahan (lihat Bahan Tambahan). Meskipun kami tidak menemukan perbedaan yang signifikan, kami mengamati besarnya kejutan yang sedikit lebih tinggi untuk avCS + dibandingkan dengan CS- dan stimulus kontrol baru. Kami juga mengamati sedikit lebih banyak pelemahan mengejutkan pada appCS + dibandingkan dengan CS- dan stimulus kontrol baru pada awal fase kepunahan, yang, bagaimanapun, menghilang dalam beberapa percobaan. SCR ke appCS + turun sudah setelah percobaan kepunahan 2nd, sementara SCR ke avCS + tetap lebih tinggi untuk hampir semua kepunahan dibandingkan dengan SCR ke BARU. Meskipun kita harus menafsirkan hasil ini dengan sangat hati-hati, tanggapan mengejutkan tampaknya sejajar dengan peringkat valensi. Selain itu, baik tanggapan mengejutkan maupun SCR lebih jauh mendukung gagasan konservatisme evolusioner terhadap rangsangan ancaman.
Akhirnya, kami harus mengakui beberapa keterbatasan penelitian ini. Pertama, karena masalah teknis kami tidak dapat melaporkan peringkat kesenangan (dan gairah) untuk cokelat dan pretzel asin. Namun, CR selera menunjukkan bahwa peserta benar-benar mengalami dua nafsu makan AS. Kedua, durasi AS permusuhan dan selera sangat berbeda. Dengan demikian, sengatan listrik yang menyakitkan disampaikan pada offset dari avCS + dan berlangsung tepat 200 ms, sedangkan coklat dan pretzel asin disajikan kepada peserta sekitar 2 setelah onset appCS +, dan durasinya tidak dapat ditentukan karena bergantung pada seberapa cepat individu memakannya. Pilihan untuk memberikan selera makan AS dengan cara ini didasarkan pada penelitian sebelumnya pada manusia (Prévost et al., 2012). Namun, secara metodologis akan lebih elegan untuk memberikan AS yang lebih enak dibandingkan dengan AS yang permusuhan, seperti seteguk jus atau bir jahe di appCS + offset. Untuk mengimbangi perbedaan persepsi yang besar ini, kami menyajikan bentuk geometris bersama dengan stimulus visual yang melambangkan AS. Ketiga, kita tidak dapat secara pasti mengecualikan kemungkinan bahwa respons fisiologis yang cepat padam pada fase kepunahan disebabkan oleh aspek metodologis. Bahkan, rangsangan visual dalam fase ini tidak disajikan dalam hubungannya dengan simbol AS seperti halnya dalam fase akuisisi. Ada kemungkinan bahwa presentasi CSs saja mungkin telah mempengaruhi CRs dan karena alasan itu tidak ada perbedaan signifikan yang terlihat selama percobaan kepunahan pertama. Namun, tanggapan cepat kami padam sejalan dengan tanggapan padam dalam penelitian sebelumnya di mana CS + (wajah) disajikan dalam gabungan dengan AS (teriakan) selama akuisisi tetapi tidak selama kepunahan (Lissek et al., 2008).
Sebagai kesimpulan, kami menemukan respon terkondisi dan selera makan yang berhasil untuk stimulus yang terkait dengan ancaman dan untuk stimulus yang terkait dengan hadiah, masing-masing. Menariknya, tingkat respons eksplisit (peringkat) dan implisit (refleks kejut) bekerja secara sinergis, karena avCS + dilaporkan sebagai potensiasi negatif yang diinduksi dan appCS + dilaporkan sebagai pelemahan mengejutkan yang diinduksi positif. Selain itu, gairah eksplisit (peringkat) dan fisiologis (SCR) dari appCS + dipisahkan, yang mencerminkan dua proses yang berbeda.
Pernyataan Benturan Kepentingan
Para penulis menyatakan bahwa penelitian ini dilakukan tanpa adanya hubungan komersial atau keuangan yang dapat ditafsirkan sebagai potensi konflik kepentingan.
Ucapan Terima Kasih
Pekerjaan ini didukung oleh Pusat Penelitian Kolaboratif "Ketakutan, Kecemasan, dan Gangguan Kecemasan," SFB-TRR 58, proyek B1.
Materi tambahan
Bahan Pelengkap untuk artikel ini dapat ditemukan online di: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Referensi
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E., dan Pauli, P. (2013). Prediktabilitas nyeri membalikkan peringkat valensi dari stimulus yang terkait dengan pertolongan. Depan. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B., dan Pauli, P. (2010). Perpecahan antara valensi tersirat implisit dan eksplisit setelah pembelajaran pereda nyeri pada manusia. Proc Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Austin, AJ, dan Duka, T. (2010). Mekanisme perhatian untuk hasil nafsu makan dan permusuhan di pengkondisian Pavlovian. Behav. Res Otak. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV, dan van Boxtel, A. (2005). Laporan Komite: pedoman untuk studi electromyographic eyeblink mengejutkan manusia. Psikofisiologi 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Bouton, ME, dan Peck, CA (1989). Efek konteks pada pengkondisian, kepunahan, dan pemulihan dalam persiapan pengkondisian selera. Anim. Belajar. Behav. 17, 188 – 198. doi: 10.3758 / BF03207634
Teks Lengkap CrossRef | Google Scholar
Bradley, MM (2009). Perhatian selektif alami: orientasi dan emosi. Psikofisiologi 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Clark, JJ, Hollon, NG, dan Phillips, PEM (2012). Sistem penilaian Pavlovian dalam pembelajaran dan pengambilan keputusan. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Delgado, MR, Jou, RL, dan Phelps, EA (2011). Sistem saraf yang mendasari pengkondisian permusuhan pada manusia dengan penguat primer dan sekunder. Depan. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Dickinson, A., dan Balleine, B. (1994). Kontrol motivasi tindakan yang diarahkan pada tujuan. Anim. Belajar. Behav. 22, 1 – 18. doi: 10.3758 / BF03199951
Teks Lengkap CrossRef | Google Scholar
Fendt, M., dan Fanselow, MS (1999). Basis neuroanatomik dan neurokimiawi dari ketakutan yang terkondisikan. Neurosci. Biobehav. Putaran. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Gottfried, JA, O'Doherty, J., dan Dolan, RJ (2002). Pembelajaran penciuman nafsu makan dan permusuhan pada manusia dipelajari menggunakan pencitraan resonansi magnetik fungsional terkait peristiwa. J. Neurosci. 22, 10829 – 10837. Tersedia online di: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B., dan Johnson, L. (2006). Pengondisian konteks dan penghindaran perilaku dalam lingkungan realitas virtual: efek dari prediktabilitas. Biol. Psikiatri 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Hamm, AO, dan Weike, AI (2005). Neuropsikologi pembelajaran ketakutan dan regulasi rasa takut. Int. J. Psychophysiol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Aktivasi saraf perolehan gairah seksual terkondisi: efek dari kesadaran kontingensi dan seks. J. Seks. Med. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). Polimorfisme 5-HTTLPR dikaitkan dengan perubahan respons hemodinamik selama pengkondisian selera. Bersenandung. Pemetaan otak. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Koch, M. (1999). Neurobiologi yang mengejutkan. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Koch, M., Schmid, A., dan Schnitzler, H.-U. (1996). Pelemahan-kesenangan dari kejut terganggu oleh lesi nukleus accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W., dan Tausch, A. (1996). Untersuchungen mit einer deutschen versi der “Jadwal Pengaruh Positif dan Negatif” (PANAS). Diagnostika 42, 139-156.
Kumar, P., Pelayan, G., Ahearn, T., Milders, M., Reid, I., dan Steele, JD (2008). Perbedaan temporal yang abnormal sinyal hadiah-belajar pada depresi berat. Otak 131, 2084 – 2093. doi: 10.1093 / otak / awn136
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P., dan Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: Uji Beltz.
Levy, DJ, dan Glimcher, PW (2011). Membandingkan apel dan jeruk: menggunakan representasi nilai subjektif spesifik-hadiah dan umum-umum di otak. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Peningkatan rasa takut pada rangsangan tanpa syarat yang relevan secara sosial dalam gangguan kecemasan sosial. Saya. J. Psikiatri 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Martin-Soelch, C., Linthicum, J., dan Ernst, M. (2007). Pengkondisian nafsu makan: dasar saraf dan implikasi untuk psikopatologi. Neurosci. Biobehav. Putaran. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y., dan Schoenbaum, G. (2011). Ventral striatum dan orbitofrontal cortex keduanya diperlukan untuk pembelajaran penguatan model berbasis, tetapi tidak bebas model. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Milad, MR, dan Quirk, G. (2012). Kepunahan yang ditakuti sebagai model untuk neuroscience translasi: sepuluh tahun kemajuan. Annu. Pendeta Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Pavlov, IP (1927). Reflex Terkondisikan: Investigasi Aktivitas Fisiologis Korteks Serebral. London: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM, dan O'Doherty, JP (2012). Korelasi saraf dari transfer Pavlovian-ke-instrumental yang spesifik dan umum dalam subkawasan amygdalar manusia: studi fMRI resolusi tinggi. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Tranel, D., dan Damasio, H. (1994). Korelasi neuroanatomikal dari respon konduktansi kulit electrodermal. Psikofisiologi 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
Abstrak PubMed | Teks Lengkap CrossRef | Google Scholar
Kata kunci: pengkondisian klasik, imbalan, hukuman, refleks kejut, respons konduktansi kulit
Kutipan: Andreatta M dan Pauli P (2015) Appetitive vs Aversive conditioning pada manusia. Depan. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Diterima: 11 Februari 2015; Diterima: 05 Mei 2015;
Diterbitkan: 19 Mei 2015.
Diedit oleh:
Niels Birbaumer, Universitas Tuebingen, Jerman
Diulas oleh:
Tim Klucken, Justus Liebig University Giessen, Jerman
Alessandro Angrilli, Universitas Padova, Italia
Hak Cipta © 2015 Andreatta dan Pauli. Ini adalah artikel akses terbuka yang didistribusikan di bawah ketentuan Lisensi Atribusi Creative Commons (CC BY). Penggunaan, distribusi atau reproduksi di forum lain diizinkan, asalkan penulis asli atau pemberi lisensi dikreditkan dan bahwa publikasi asli dalam jurnal ini dikutip, sesuai dengan praktik akademik yang diterima. Dilarang menggunakan, mendistribusikan, atau mereproduksi, yang tidak mematuhi ketentuan ini.
* Korespondensi: Marta Andreatta, Departemen Psikologi (Psikologi Biologis, Psikologi Klinis, dan Psikoterapi), Universitas Würzburg, Marcusstraße 9-11, D-97070 Würzburg, Jerman, [email dilindungi]