Depan. Behav. Neurosci., 23 Maret 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Institut Penelitian Translasional dan Institut Inovasi Biomedis dan Kesehatan, Universitas Teknologi Queensland, Brisbane, QLD, Australia
- 2Fakultas Ilmu Biomedis, Universitas Queensland, Brisbane, QLD, Australia
Diet modern telah menjadi sangat manis, menghasilkan tingkat konsumsi gula yang belum pernah terjadi sebelumnya, terutama di kalangan remaja. Sementara asupan gula jangka panjang yang kronis diketahui berkontribusi pada perkembangan gangguan metabolisme termasuk obesitas dan diabetes tipe II, hanya sedikit yang diketahui mengenai konsekuensi langsung dari konsumsi gula jangka panjang, seperti pesta di otak. BKarena gula dapat menyebabkan pelepasan dopamin dalam nukleus accumbens (NAc) yang serupa dengan obat pelecehan, kami menyelidiki perubahan dalam morfologi neuron di wilayah otak ini setelah jangka pendek (minggu 4) dan jangka panjang (minggu 12). seperti konsumsi sukrosa menggunakan paradigma pilihan dua botol yang terputus-putus. Kami menggunakan pewarnaan Golgi-Cox untuk menghamili neuron berduri sedang (MSN) dari inti NAc dan cangkang tikus yang mengkonsumsi sukrosa jangka pendek dan jangka panjang dan membandingkannya dengan kontrol air yang sesuai dengan usia. Kami menunjukkan bahwa konsumsi sukrosa jangka panjang seperti pesta secara signifikan menurunkan panjang total dendritik MSN shell NAc dibandingkan dengan tikus kontrol yang disesuaikan dengan usia. Kami juga menemukan bahwa restrukturisasi neuron ini terutama disebabkan oleh berkurangnya kompleksitas dendritik distal. Sebaliknya, kami mengamati peningkatan kepadatan tulang belakang pada pesanan cabang distal MSN cangkang NAc dari tikus yang mengonsumsi sukrosa jangka panjang. Dikombinasikan, hasil ini menyoroti efek neuron dari asupan sukrosa seperti pesta berlebihan pada morfologi MSC shell NAc.
Pengantar
Selama 40 tahun terakhir, telah terjadi peningkatan konsumsi minuman dan makanan yang dimaniskan dengan gula yang mengandung gula tambahan (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), dengan laporan yang memperkirakan bahwa hingga 75% dari semua makanan dan minuman mengandung jumlah gula tambahan yang tinggi (Ford dan Dietz, 2013; Bray dan Popkin, 2014). Selama periode ini, ada juga peningkatan bersamaan dalam prevalensi obesitas dan diabetes tipe II, terutama di kalangan remaja (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Studi terbaru menunjukkan bahwa anak-anak yang kelebihan berat badan dan obesitas sering mengonsumsi gula tambahan dalam jumlah tinggi, namun kontribusi diet tinggi gula terhadap peningkatan insiden anak-anak yang kelebihan berat badan dan obesitas tetap kontroversial (Hu, 2013; Bray dan Popkin, 2014; Bucher Della Torre et al., 2015).
Sementara semakin banyak bukti menunjukkan bahwa konsumsi diet tinggi gula dapat, sebagian, berkontribusi pada kenaikan berat badan di antara anak-anak dan remaja (Malik et al., 2010; Te Morenga et al., 2013; Bray dan Popkin, 2014), kurang perhatian telah diberikan pada konsekuensi non-metabolik yang merugikan yang timbul dari asupan gula yang berlebihan. Menariknya, beberapa pola perilaku dan psikologis umum sering muncul di antara sebagian dari mereka yang makan berlebihan dan mempertahankan diet tinggi gula. Yang paling menonjol adalah perkembangan gangguan makan termasuk pesta makan, dikombinasikan dengan timbulnya gejala psikologis termasuk kurangnya motivasi dan depresi (ditinjau dalam Sheehan dan Herman, 2015). Selain itu, karena individu yang makan berlebihan sering menunjukkan kehilangan kontrol dan ketidakmampuan untuk membatasi asupan gula mereka, ada kemungkinan bahwa perilaku ini muncul sebagai akibat dari adaptasi neurologis di daerah otak yang mengevaluasi nilai hedonis dari makanan yang sangat enak. (Saper et al., 2002; Lutter dan Nestler, 2009; Kenny, 2011). Alasan ini juga didukung oleh bukti pada manusia yang menunjukkan bahwa gula dan rasa manis dapat menyebabkan ngidam yang mirip dengan yang diinduksi oleh obat adiktif seperti alkohol dan nikotin (Volkow et al., 2012).
Meskipun sifat adiktif gula masih spekulatif, pengamatan ini dikombinasikan dengan penelitian yang menunjukkan kontribusi asupan gula yang berlebihan terhadap perubahan sirkuit hadiah dan pengembangan perilaku seperti kecanduan dan keadaan emosional dalam model hewan. (Avena et al., 2008; Benton, 2010; Ventura et al., 2014), menjamin perlunya penyelidikan lebih lanjut. Penelitian sebelumnya pada tikus telah menunjukkan bahwa akses intermiten ke sukrosa mengubah aktivitas beberapa neurotransmiter dalam sistem mesolimbik termasuk dopamin, opioid dan asetilkolin (diulas di Avena et al., 2008). Konsumsi sukrosa seperti pesta telah terbukti memfasilitasi pelepasan dopamin dalam nucleus accumbens (NAc), serupa dengan obat-obatan pelecehan (Avena et al., 2008). Selain itu, kami telah menunjukkan bahwa konsumsi sukrosa jangka panjang menggunakan 24 jam akses terputus dua-pilihan paradigma pilihan (Simms et al., 2008) memodulasi ekspresi reseptor asetilkolin nikotinat (nAChR) dalam NAc (Shariff et al., Dalam pers). Menariknya, kami juga telah mengamati bahwa senyawa nAChR yang diketahui memodulasi aktivitas dopamin dan asetilkolin dalam NAc, memiliki efek yang berbeda pada konsumsi sukrosa setelah asupan jangka pendek dan jangka panjang (Shariff et al., Dalam pers).
Sementara penelitian ini telah menunjukkan kesamaan dalam perubahan perilaku dan neurokimia yang disebabkan oleh akses intermiten terhadap gula dan obat-obatan pelecehan, tidak diketahui apakah efek ini memfasilitasi perubahan dalam morfologi neuronal di NAc. Tini berbeda dengan zat pelecehan termasuk kokain, amfetamin dan nikotin yang menghasilkan perubahan yang ditandai dengan baik dalam morfologi neuron berduri menengah (MSNs) di NAc, termasuk peningkatan kepadatan tulang belakang dan perubahan kompleksitas dendritik. (Robinson dan Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Karena kami sebelumnya telah menunjukkan bahwa paparan jangka panjang (minggu 12) terhadap alkohol dan sukrosa menggunakan paradigma pilihan dua botol yang intermiten menghasilkan respons yang berbeda terhadap intervensi farmakoterapi dibandingkan dengan asupan jangka pendek (4 minggu; Steensland et al., 2007; Shariff et al., Dalam pers), kami menilai efek dari konsumsi sukrosa jangka pendek dan jangka panjang pada morfologi MSN di NAc. Kami mengizinkan tikus remaja untuk mengkonsumsi sukrosa dengan cara seperti pesta untuk 4 (jangka pendek) atau 12 (jangka panjang) minggu dan kemudian menganalisis morfologi MSN MSC dari tikus yang mengkonsumsi sukrosa jangka pendek dan jangka panjang dan membandingkannya dengan kontrol sesuai usia yang hanya diberi akses ke air. Hasil kami menunjukkan bahwa MSN dari cangkang NAc diubah mengikuti konsumsi sukrosa jangka panjang tetapi tidak jangka pendek, setelah mengurangi panjang dendritik, tetapi meningkatkan kepadatan tulang belakang dendritik distal. Selain itu, kami menemukan morfologi MSN dari inti NAc tetap relatif utuh setelah konsumsi sukrosa jangka pendek dan jangka panjang. Hasil ini menyoroti konsekuensi neurologis langsung dari konsumsi sukrosa jangka panjang dengan cara seperti pesta. Selain itu, data ini menunjukkan perlunya penelitian lebih lanjut yang bertujuan untuk menjelaskan perubahan molekuler dan neurokimia yang menyertai restrukturisasi morfologis MSN cangkang NAcs yang disebabkan oleh asupan sukrosa yang berkepanjangan seperti binge-like.
Bahan dan Metode
Pernyataan etika
Semua prosedur eksperimental dilakukan sesuai dengan Kode Australia untuk Perawatan dan Penggunaan Hewan untuk Tujuan Ilmiah, Edisi 8th (Dewan Riset Kesehatan dan Medis Nasional, 2013). Protokol disetujui oleh Komite Etika Hewan Universitas Queensland dan Komite Etika Hewan Universitas Queensland.
Hewan dan Perumahan
Tikus Wistar jantan berusia lima minggu (remaja) (Kontrol: 176.4 ± 4.8 g; Sukrosa: 178.3 ± 5.0 g) (ARC, WA, Australia), masing-masing ditempatkan di Plexiglas berventilasi ganda.® kandang. Tikus-tikus tersebut menyesuaikan diri dengan kondisi rumah masing-masing, penanganan, dan siklus cahaya-terbalik 5 hari sebelum dimulainya percobaan. Semua tikus ditempatkan di ruang siklus cahaya / gelap terbalik yang dikendalikan oleh 12-jam (dimatikan pukul 9 pagi) dengan ruang chow dan air tikus standar tersedia ad libitum.
Paradigma Minum Pilihan Dua-Botol Pilihan-Terpisah
Paradigma minum 5% sukrosa pilihan terputus-putusSimms et al., 2008) diadaptasi dari Bijaksana (1973). Semua cairan disajikan dalam botol plastik bertingkat 300 ml dengan cerat stainless steel yang dimasukkan melalui dua grommet di bagian depan sangkar setelah dimulainya siklus cahaya gelap. Bobot masing-masing botol dicatat sebelum presentasi botol. Dua botol disajikan secara bersamaan: satu botol berisi air; botol kedua mengandung sukrosa 5% (b / v). Penempatan botol sukrosa 5% (b / v) diselingi dengan masing-masing paparan untuk mengontrol preferensi sisi. Botol ditimbang 24 h setelah cairan disajikan, dan pengukuran dilakukan ke 0.1 terdekat g. Berat masing-masing tikus juga diukur untuk menghitung gram asupan sukrosa per kilogram berat badan. Pada hari 1 periode minum, tikus (n = 6 – 9) diberi akses ke satu botol 5% (b / v) sukrosa dan satu botol air. Setelah 24 jam, botol sukrosa diganti dengan botol air kedua yang tersedia untuk 24 jam berikutnya. Pola ini diulang pada hari Rabu dan Jumat. Tikus memiliki akses tak terbatas ke air pada hari-hari lainnya. Konsumsi sukrosa seperti pesta menghasilkan peningkatan total asupan sukrosa (ml) dari waktu ke waktu (Gambar Tambahan 1) dan disertai dengan tingkat minum baseline stabil berdasarkan berat badan [20 ± 5 g / kg dari 5% (b / v)] selama jangka pendek [~ 4 minggu (sesi minum 13)] dan jangka panjang [ ~ 12 minggu (sesi minum 37)] periode minum. Kelompok tikus kontrol yang terpisah (n = 6 – 9) diberi akses ke air di kedua botol (yaitu, tidak ada sukrosa) di bawah kondisi yang sama yang dijelaskan di atas. Berat badan rata-rata dari kontrol dan tikus yang mengkonsumsi sukrosa pada akhir paparan jangka pendek adalah masing-masing 405.7 ± 40.8 g dan 426.4 ± 31.2 g. Pada akhir paparan jangka panjang, berat badan rata-rata untuk kelompok kontrol dan sukrosa adalah 578.8 ± 53.4 g dan 600.2 ± 45.2 g.
Pewarnaan Golgi-Cox
Setelah sesi minum terakhir, tikus dipindahkan dari fasilitas hewan untuk memungkinkan pemrosesan sampel otak di fasilitas histologi di School of Biomedical Sciences, Universitas Queensland (St Lucia, Australia). Semua tindakan yang disetujui diambil untuk mengurangi stres selama transportasi, setelah itu, tikus dibiarkan pulih dalam semalam. Keesokan harinya, tikus dikorbankan oleh overdosis natrium pentobarbital (60-80 mg / kg, ip Vetcare, Brisbane, Australia) dan perfusi intrakardial dengan ~ 300 ml cairan serebro-spinal buatan yang berisi, (dalam mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, dan 10 D-glukosa. Setiap hewan kemudian dipenggal dan otak dikeluarkan dan diinkubasi dalam gelap dalam larutan Golgi-Cox yang mengandung 5% kalium dikromat, 5% kalium kromat, dan 5% merkuri klorida (semua bahan kimia dari Sigma-Aldrich) yang dibuat 3 hari segar sebelum berkorban seperti yang dijelaskan sebelumnya (Rutledge et al., 1969). Inkubasi pewarnaan Golgi-Cox dan metode pasca-pemrosesan dimodifikasi Ranjan dan Mallick (2010). Otak dari hewan yang mengonsumsi sukrosa jangka pendek diinkubasi selama 6 hari di 37 ° C, sementara otak dari hewan yang mengonsumsi sukrosa jangka panjang diinkubasi selama 10 hari, dengan satu perubahan ke solusi Golgi-Cox segar setelah 4 hari inkubasi.
Setelah inkubasi, bagian koronal 300 μm dipotong menggunakan mikrotom Zeiss Hyrax V50 yang bergetar (Carl Zeiss, Jerman). Irisan kemudian ditempatkan secara berurutan dalam pelat sumur 24 yang diisi dengan sukrosa 30% (b / v) dalam saline buffered 0.1 M fosfat dan diproses sebagaimana diuraikan dalam (Ranjan dan Mallick, 2010). Secara singkat, bagian didehidrasi dalam 50% etanol selama 5 min, kemudian ditempatkan di 0.1 M NH4Solusi OH untuk 30 min, dibilas dua kali dengan air suling selama 5 min dan ditempatkan di fixer film Fujihunt (Fujifilm, Singapura) untuk 30 min dalam gelap. Irisan kemudian dibilas dua kali dalam air suling untuk masing-masing 2 min dan didehidrasi dalam 70, 90, 95, dan 100% etanol dua kali untuk masing-masing 5 min. Bagian-bagian tersebut kemudian dibersihkan dalam larutan CXA (1: 1: 1 chloroform: xylene: alcohol) untuk 10 min dan dipasang di DPX (Sigma-Aldrich) pada slide Superfrost Plus (Menzel-Glaser, Lomb Scientific, Australia) dan penutup-selip (Menzel-Glaser, Jerman). Slide dibiarkan dalam gelap hingga kering pada suhu kamar semalaman.
Seleksi dan Penelusuran Neuronal dalam Nucleus Accumbens
Irisan koral antara bregma + 2.8 dan + 1.7 disurvei untuk MSN dalam inti dan kulit NAc, menggunakan ventrikel lateral dan commissure anterior sebagai landmark dengan bantuan atlas otak tikus (Paxinos dan Watson, 2007) (Gambar 1). Fungsi kontur di Neurolucida 7 (MBF Bioscience, VT, USA) digunakan untuk membatasi inti NAc dan shell NAc di setiap irisan (Gambar 2). Antara neuron 2 dan 9 per wilayah per hewan dilacak untuk parameter panjang dendritik menggunakan tujuan 63x atau untuk kepadatan tulang belakang (dilaporkan sebagai duri per 100 μm) menggunakan tujuan 100x pada Zeiss Axioskop II (Carl Zeiss, Jerman) menggunakan otomatis xyz tahap didorong oleh Neurolucida® Perangkat lunak 7 (MBF Biosciences, VT, USA). Semua pelacakan dilakukan dengan cara blinded sehubungan dengan perawatan. Parameter morfologis neuron diresapi Golgi-Cox dianalisis dengan cara yang mirip dengan laporan sebelumnya (Klenowski et al., 2015).
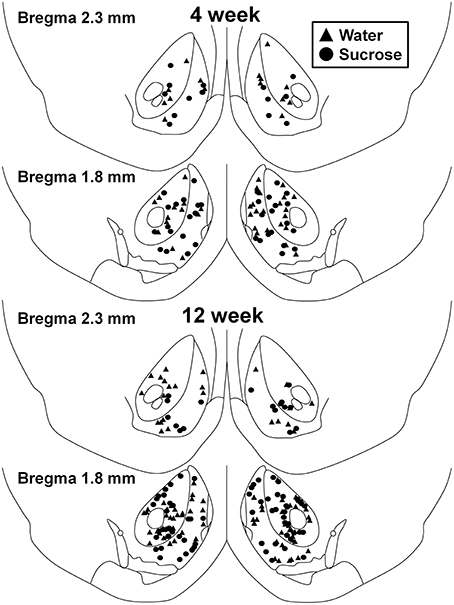
Gambar 1. Peta yang menunjukkan lokasi neuron berduri sedang yang diambil sampel dari nukleus accumbens core dan cangkang tikus yang mengkonsumsi sukrosa 4 dan 12 minggu dan kontrol yang sesuai umur. Dua panel teratas menunjukkan lokasi neuron yang diambil dari nukleus accumbens core dan cangkang kontrol minggu 4 (segitiga) dan sukrosa (lingkaran) hewan. Dua panel bawah menunjukkan posisi neuron sampel dari kontrol minggu 12 (segitiga) dan sukrosa (lingkaran) hewan.
Analisis Statistik
Mean dan standard error of mean (SEM) dihitung untuk setiap set data dengan hewan sebagai n, menggunakan data morfometri rata-rata dari semua MSN inti atau shelln = 7 untuk shell NAc dan n = 6 untuk NAc core 4-minggu, n = 9 untuk kelompok 12 minggu). Dimana ditunjukkan, Student dua ekor tidak berpasangan tuji atau ANOVA dua arah dengan post-test Bonferroni dilakukan untuk semua analisis yang melibatkan perbandingan cara kelompok, menggunakan GraphPad Prism versi 6.02 (GraphPad Software, San Diego, CA). Signifikansi statistik diterima di P <0.05. Semua data di bagian hasil disajikan sebagai sarana ± SEM. Perubahan persentase dihitung sebagai relatif terhadap nilai kontrol.
Hasil
Neuron Berduri Sedang dari Nucleus Accumbens Shell telah Menurunkan Panjang Dendritik, Menurunkan Kompleksitas Dendritik tetapi Meningkatkan Kepadatan Tulang Belakang Rata-rata di Cabang Cabang Berikutnya, Mengikuti Konsumsi Sukrosa Jangka Panjang tetapi tidak Jangka Pendek
Setelah konsumsi sukrosa jangka pendek (4 minggu), tidak ada perbedaan signifikan dalam parameter morfometrik MSC shell NAN (Tabel 1). Juga tidak ada perbedaan signifikan antara konsumsi sukrosa jangka pendek dan MSN cangkang NAc kontrol air dalam analisis yang terkait dengan urutan cabang sentrifugal. Yaitu, segmen dendritik per pesanan cabang (P = 0.4111), berarti panjang dendritik per pesanan cabang (P = 0.5581) dan kepadatan tulang belakang rata-rata per pesanan cabang (P = 0.2977, ANOVA dua arah) tidak berbeda secara signifikan antar kelompok. Peta lokasi yang menunjukkan perkiraan posisi neuron sampel ditunjukkan pada Gambar 1.
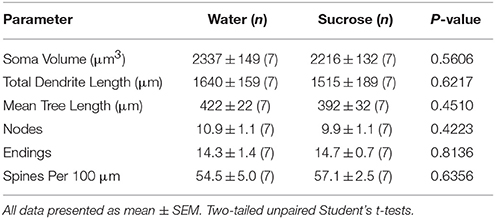
Tabel 1. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens shell dari tikus yang mengkonsumsi sukrosa jangka pendek dan kontrol air yang sesuai umur.
Setelah konsumsi sukrosa jangka panjang (12 minggu), panjang total dendritik pangkalan MSC kulit NAc mengalami penurunan sebesar 21% dibandingkan dengan kontrol yang mengonsumsi air (Air: 1827 ± 148 μm, n = 9; Sukrosa 1449 ± 78 μm, n = 9, *P = 0.0384, Mahasiswa tidak berpasangan dua sisi t-test, Gambar 2, Meja 2). Perbandingan jumlah rata-rata bifurkasi dendritik (node) dan ujung dendritik antara air dan kelompok sukrosa menunjukkan tingkat kompleksitas dendritik yang berkurang (walaupun tidak signifikan) dalam MSC kulit NAc (node: Air 12.9 ± 1.4 n = 9, Sucrose 10.1 ± 0.8 n = 9, P = 0.0879; ujung: Air 17.9 ± 1.4 n = 9, Sucrose 14.8 ± 0.7 n = 9, P = 0.0657, Mahasiswa tidak berpasangan dua sisi t-test, Tabel 2). Tidak ada perubahan dalam volume soma (P = 0.9400), berarti panjang pohon dendritik (P = 0.1646) atau kepadatan tulang belakang total (P = 0.3662) dalam cangkang NAc MSN dari tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol air. Parameter morfometrik ini dirinci dalam Tabel 2.
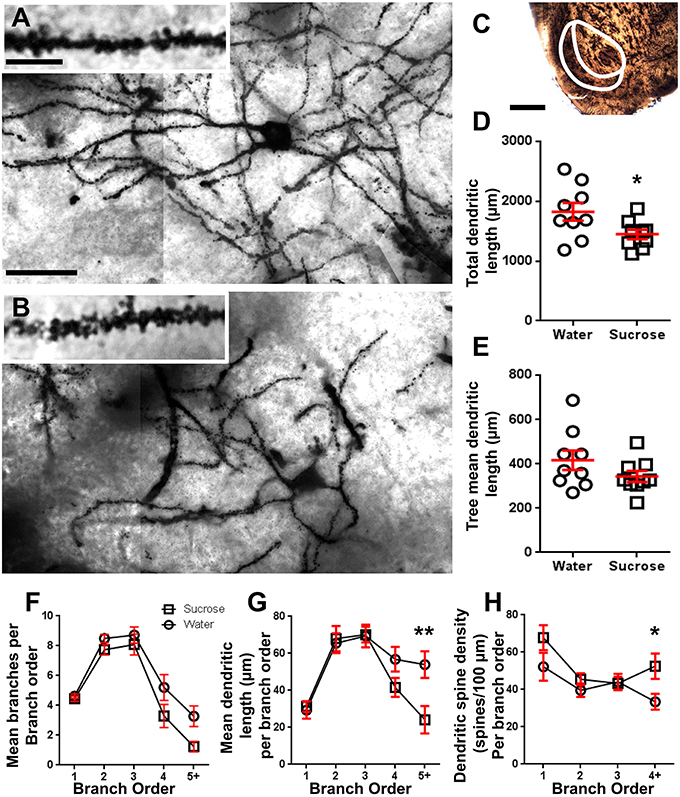 Gambar 2. Mengurangi panjang punjung dendritik dan meningkatkan kepadatan tulang belakang dendritik diston neuron berduri sedang (MSNs) dari cangkang nucleus accumbens (NAc) dari tikus yang diperlakukan dengan sukrosa jangka panjang dibandingkan dengan tikus kontrol. (A, B) menunjukkan representasi kontrol (atas) dan jangka panjang (12 minggu) sukrosa (bawah) diperlakukan brightfield zmosaik tumpukan MSN diresapi Golgi dari shell NAc (pembesaran 63x). Awal dari (A, B) menunjukkan kontrol dan gambar jangka panjang sukrosa yang diobati dari dendrit MSN yang diresapi Golgi dan dendritik duri dari cangkang NAc (pembesaran 100x). (C) menunjukkan daerah anatomi yang diambil sampel MSN dalam penelitian ini. (D) menunjukkan sebar-plot penurunan total dendritik punjung MSN (rata-rata ± SEM) dari cangkang NAc pada hewan sukrosa jangka panjang (kotak) dibandingkan dengan kontrol (lingkaran), siswa tidak berpasangan t-uji, *P <0.05, n = 9; kontrol dan n = 9; Sukrosa minggu 12. (E) menunjukkan sebaran-sebaran panjang pohon dendritik MSN rata-rata yang tidak berubah (rata-rata ± SEM) dari cangkang NAc pada hewan sukrosa jangka panjang (kotak) dibandingkan dengan kontrol (lingkaran), siswa tidak berpasangan t-uji, P > 0.05, n = 9; kontrol dan n = 9; Sukrosa minggu 12. Analisis urutan cabang (rata-rata ± SEM) dari nomor segmen dendritik per pesanan cabang (F), berarti panjang dendritik per pesanan cabang (G) dan kepadatan tulang belakang dendritik per pesanan cabang (H). Konsumsi sukrosa jangka panjang menurunkan panjang dendritik pada pesanan cabang distal (5 +) dan meningkatkan kepadatan tulang belakang dendritik pada pesanan cabang distal (4 +) dibandingkan dengan kontrol (G, H), ANOVA dua arah dengan post-test Bonferroni, *P <0.05, **P <0.01, n = 9; kontrol dan n = 9; sukrosa jangka panjang. Skala Bar: (A, B) = 20 μm; sisipan dari (A, B) = 10 μm; (C) = 1 mm.
Gambar 2. Mengurangi panjang punjung dendritik dan meningkatkan kepadatan tulang belakang dendritik diston neuron berduri sedang (MSNs) dari cangkang nucleus accumbens (NAc) dari tikus yang diperlakukan dengan sukrosa jangka panjang dibandingkan dengan tikus kontrol. (A, B) menunjukkan representasi kontrol (atas) dan jangka panjang (12 minggu) sukrosa (bawah) diperlakukan brightfield zmosaik tumpukan MSN diresapi Golgi dari shell NAc (pembesaran 63x). Awal dari (A, B) menunjukkan kontrol dan gambar jangka panjang sukrosa yang diobati dari dendrit MSN yang diresapi Golgi dan dendritik duri dari cangkang NAc (pembesaran 100x). (C) menunjukkan daerah anatomi yang diambil sampel MSN dalam penelitian ini. (D) menunjukkan sebar-plot penurunan total dendritik punjung MSN (rata-rata ± SEM) dari cangkang NAc pada hewan sukrosa jangka panjang (kotak) dibandingkan dengan kontrol (lingkaran), siswa tidak berpasangan t-uji, *P <0.05, n = 9; kontrol dan n = 9; Sukrosa minggu 12. (E) menunjukkan sebaran-sebaran panjang pohon dendritik MSN rata-rata yang tidak berubah (rata-rata ± SEM) dari cangkang NAc pada hewan sukrosa jangka panjang (kotak) dibandingkan dengan kontrol (lingkaran), siswa tidak berpasangan t-uji, P > 0.05, n = 9; kontrol dan n = 9; Sukrosa minggu 12. Analisis urutan cabang (rata-rata ± SEM) dari nomor segmen dendritik per pesanan cabang (F), berarti panjang dendritik per pesanan cabang (G) dan kepadatan tulang belakang dendritik per pesanan cabang (H). Konsumsi sukrosa jangka panjang menurunkan panjang dendritik pada pesanan cabang distal (5 +) dan meningkatkan kepadatan tulang belakang dendritik pada pesanan cabang distal (4 +) dibandingkan dengan kontrol (G, H), ANOVA dua arah dengan post-test Bonferroni, *P <0.05, **P <0.01, n = 9; kontrol dan n = 9; sukrosa jangka panjang. Skala Bar: (A, B) = 20 μm; sisipan dari (A, B) = 10 μm; (C) = 1 mm.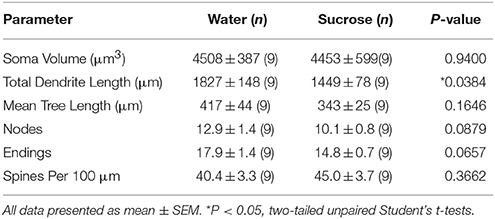
Tabel 2. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens shell dari tikus yang mengkonsumsi sukrosa jangka panjang dan kontrol air yang sesuai usia.
Mengikuti karakterisasi morfologi dendritik umum dari sukrosa MSN cangkang NAc jangka panjang, kami menganalisis arendisasi dendritik dan kepadatan tulang belakang sehubungan dengan karakteristik pesanan cabang mereka. Penilaian komprehensif kami terhadap pohon dendritik mengkuantifikasi jumlah segmen dendritik per pesanan cabang, panjang rata-rata segmen dendritik per pesanan cabang dan rerata kepadatan tulang belakang per pesanan cabang dari MSC kulit MSN kontrol air dan tikus yang mengonsumsi sukrosa jangka panjang. Ringkasan data pesanan cabang dan analisis disajikan pada Tabel 3.
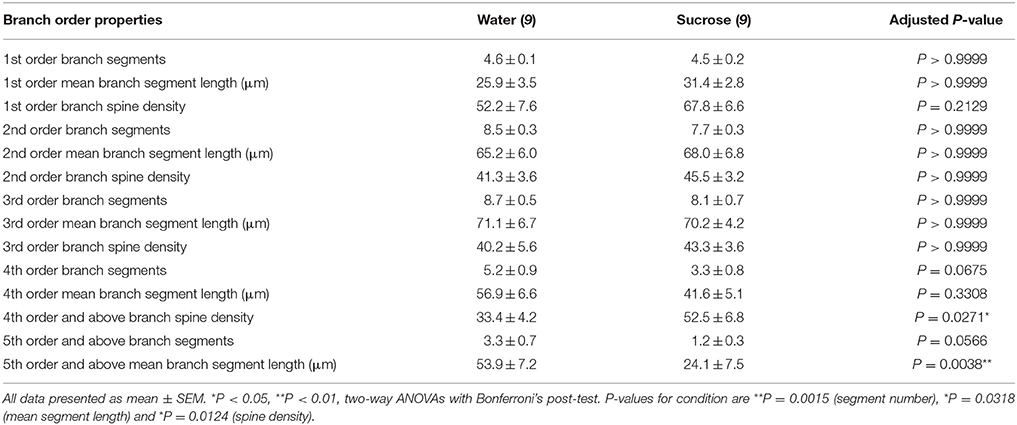 Tabel 3. Karakteristik urutan cabang dari neuron berduri sedang dari sukrosa jangka panjang dan tikus yang minum air.
Tabel 3. Karakteristik urutan cabang dari neuron berduri sedang dari sukrosa jangka panjang dan tikus yang minum air.Jumlah rerata cabang dendritik rata-rata per pesanan cabang dari MSN cangkang NAc berkurang secara signifikan pada tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol air (**P = 0.0015, ANOVA dua arah). Bonferroni post-test mengungkapkan tren penurunan jumlah segmen cabang di 4th (Air: 5.2 ± 0.9, n = 9; Sukrosa 3.3 ± 0.8, n = 9, P = 0.0675, Gambar 2F, Meja 3), dan pesanan 5th dan pesanan cabang di atas (Air: 3.3 ± 0.7, n = 9; Sukrosa 1.2 ± 0.3, n = 9, P = 0.0566, Gambar 2F, Meja 3). Rata-rata panjang segmen dendritik per pesanan cabang MSC shell NAc juga berkurang secara signifikan pada tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol air (*P = 0.0444, ANOVA dua arah). Bonferroni post-test menunjukkan pengurangan 55% di cabang 5 dan seterusnya (Air: 53.9 ± 7.2 μm, n = 9; Sukrosa 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Gambar 2G, Meja 3).
Analisis urutan cabang menunjukkan peningkatan yang signifikan dalam kepadatan tulang belakang dendritik MSC shell NANs dari tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol (*P = 0.0124, ANOVA dua arah). Bonferroni post-tes menunjukkan peningkatan kepadatan tulang belakang 57% di cabang urutan 4 ke-distal dan seterusnya (Air: 33.4 ± 4.2, n = 9; Sukrosa 52.5 ± 6.8, n = 9, P = 0.0271 *, inset Angka 2A, B, H, Meja 3). Gambar representatif dari keseluruhan arsitektur MSN dan kepadatan tulang belakang (inset) digambarkan dalam Gambar 2A, B.
Secara keseluruhan, hasil ini menunjukkan bahwa konsumsi sukrosa jangka pendek memiliki sedikit efek pada parameter morfologi MSN dalam cangkang NAc. Namun, setelah konsumsi berkepanjangan, ada penurunan yang signifikan dalam panjang dan kompleksitas punjung neuronal, terutama di cabang dendritik distal. Peningkatan kerapatan tulang belakang bagian distal yang bersamaan juga terlihat pada MSN shell NACs dari tikus yang mengkonsumsi sukrosa jangka panjang.
Neuron Berduri Sedang dari Inti Nukleus telah Mengurangi Kompleksitas Percabangan setelah Konsumsi Sukrosa Jangka Pendek tetapi tidak Jangka Pendek
Mengikuti konsumsi sukrosa jangka pendek, tidak ada perbedaan signifikan dalam parameter morfometrik inti MSC (Tabel 1) 4). Juga tidak ada perbedaan yang signifikan antara konsumsi sukrosa 4-minggu dan inti kontrol air MSN dalam analisis yang terkait dengan urutan cabang sentrifugal. Yaitu, segmen dendritik per pesanan cabang (P = 0.7717), berarti panjang dendritik per pesanan cabang (P = 0.2096), dan kepadatan tulang belakang rata-rata per pesanan cabang (P = 0.3521, ANOVA dua arah) tidak berbeda antar grup.
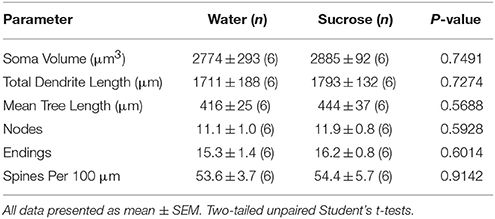 Tabel 4. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens inti tikus pemakan sukrosa jangka pendek dan kontrol air yang sesuai umur.
Tabel 4. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens inti tikus pemakan sukrosa jangka pendek dan kontrol air yang sesuai umur.Konsumsi sukrosa yang berkepanjangan juga tidak signifikan pada parameter morfometrik inti MSC (Tabel 1) 5). Jumlah segmen cabang dendritik rata-rata per pesanan cabang dari MSN inti NAc berkurang secara signifikan pada tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol air (*P = 0.0416, ANOVA dua arah), namun tidak ada perbedaan signifikan dalam panjang rata-rata dendritik per pesanan cabang (P = 0.0995) dan kepadatan tulang belakang rata-rata per pesanan cabang (P = 0.4888, ANOVA dua arah) antara MSNs di inti NAc dari tikus yang mengkonsumsi sukrosa jangka panjang dibandingkan dengan kontrol air. Secara keseluruhan, data kami menunjukkan inti NAc tidak responsif terhadap konsumsi sukrosa jangka panjang dibandingkan dengan MSN dari wilayah shell NAc.
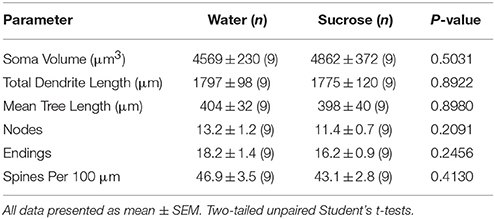 Tabel 5. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens inti tikus pemakan sukrosa jangka panjang dan kontrol air yang sesuai umur.
Tabel 5. Parameter morfologis umum neuron berduri sedang dari nukleus accumbens inti tikus pemakan sukrosa jangka panjang dan kontrol air yang sesuai umur.Diskusi
Meningkatnya ketersediaan makanan yang sangat manis dalam makanan Barat tidak hanya berkontribusi pada peningkatan prevalensi dan beban ekonomi obesitas dan diabetes tipe II, tetapi juga menyebabkan timbulnya gangguan makan seperti pesta makan (Swanson et al., 2011; Kessler et al., 2013; Davis, 2015). Meskipun sifat adiktif gula termasuk fruktosa dan sukrosa tetap spekulatif, ada kesamaan yang mencolok dalam perilaku dan korelasi saraf yang bermanifestasi sebagai akibat dari makan berlebihan dan penggunaan narkoba yang berkepanjangan. (Avena et al., 2008, 2011). Selain itu, gula mengaktifkan sirkuit reward otak dengan cara yang mirip dengan penyalahgunaan obat (Volkow et al., 2012), dan hasil dari penelitian pada manusia menunjukkan bahwa gula dan rasa manis dapat menyebabkan ngidam yang sebanding dengan yang diinduksi oleh obat adiktif seperti alkohol dan nikotin (Volkow et al., 2012). Oleh karena itu kami menggunakan model konsumsi pesta-sukrosa pada tikus untuk menentukan efek pendek (4 minggu) dan konsumsi sukrosa jangka panjang (12 minggu) pada morfologi neuron MSN di NAc, komponen kunci dari sirkuit hadiah yang tumpang tindih yang dimodulasi oleh gula dan obat-obatan adiktif. Kami menunjukkan bahwa MSNs dari cangkang NAc tikus kronis yang mengonsumsi sukrosa jangka panjang telah secara signifikan menurunkan panjang dan kompleksitas dendritik, tetapi meningkatkan kepadatan tulang belakang dendritik. Konsumsi sukrosa jangka panjang tidak berpengaruh pada morfologi MSNs dari inti NAc, sementara konsumsi sukrosa jangka pendek juga tidak berpengaruh signifikan terhadap morfologi MSN dari inti atau cangkang NAc. Hasil ini tidak hanya menunjukkan efek langsung dari asupan sukrosa yang berlebihan seperti binge pada morfologi neuron MSN cangkang NAc, tetapi mereka juga menyoroti potensi konsekuensi berbahaya dari konsumsi jangka panjang dari diet tinggi gula yang mengandung.
NAc, yang merupakan bagian dari ventral striatum, terutama terdiri dari MSN, yang secara morfologis ditandai sebagai neuron berukuran sedang dengan arborisasi dendritik yang luas dan kepadatan tulang belakang yang tinggi (Kemp dan Powell, 1971; Graveland dan DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). Neuron glutamatergik dan dopaminergik adalah dua input aferen utama ke NAc, terutama yang berhubungan dengan poros dendritik dan duri MSN (Groves, 1980; Kaiya dan Namba, 1981; Groves et al., 1994). Secara khusus, cangkang NAc dan inti menerima input glutamatergik dari area kortikal yang berbeda secara fungsional (Brog et al., 1993). Shell NAc juga dipersarafi oleh aferen rangsang dari daerah subkortikal seperti hippocampus, thalamus dan amigdala basolateral (Brog et al., 1993; Wright dan Groenewegen, 1995). Studi sebelumnya telah menunjukkan bahwa input glutamatergik ini memainkan peran penting dalam motivasi dan perilaku yang diarahkan pada tujuan seperti makanan dan pencarian hadiah (Maldonado-Irizarry et al., 1995; Kelley dan Swanson, 1997; Reynolds dan Berridge, 2003; Richard dan Berridge, 2011). Input dominan lainnya ke NAN MSNs adalah dari aferen dopaminergik yang diproyeksikan dari daerah tegmental ventral (Lindvall dan Björklund, 1978; Veening et al., 1980; Kalivas dan Miller, 1984). Menariknya, penelitian sebelumnya yang menggunakan model serupa dari akses gula intermiten telah menunjukkan bahwa hasil pesta seperti yang dihasilkan menghasilkan peningkatan dopamin ekstraseluler dalam NAc yang sama (meskipun pada tingkat lebih rendah) untuk obat penyalahgunaan (Rada et al., 2005; Avena et al., 2006), dan dapat memodulasi ekspresi reseptor dopamin (Colantuoni et al., 2001, 2002) dalam inti dan shell NAc. Menariknya, konsumsi sukrosa seperti pesta menyebabkan peningkatan asupan dari waktu ke waktu sama dengan pemberian sendiri penyalahgunaan obat-obatan seperti kokain dan heroN (Ahmed dan Koob, 1998; Ahmed et al., 2000, 2003) yang terkait dengan perkembangan kondisi "kecanduan".
Analisis kami tentang morfometri pesanan cabang menunjukkan bahwa pengurangan keseluruhan panjang dendritik MSN cangkang NAc yang disebabkan oleh asupan sukrosa jangka panjang, terutama dihasilkan dari pengurangan kompleksitas pesanan cabang di daerah jauh. Kami mengamati berkurangnya percabangan distal (urutan 4th dan 5th dan pesanan cabang di atas) dan secara signifikan mengurangi panjang rata-rata pada pesanan 5th dan di atas dendrit, dikombinasikan dengan peningkatan kepadatan tulang belakang pada pesanan cabang ini. Faktor umum yang kemungkinan mempengaruhi jenis restrukturisasi dendritik ini termasuk perubahan konektivitas dan / atau fungsi sinaptik (Russo et al., 2010). Studi sebelumnya telah menunjukkan bahwa sinapsis glutamatergik pada MSN terbentuk terutama pada spina, terutama pada dendrit distal (Groenewegen et al., 1999). Selain itu, co-lokalisasi input dopamin dan glutamatergik dari korteks prefrontal (Sesack dan Pickel, 1992), unduk-unduk (Totterdell dan Smith, 1989; Sesack dan Pickel, 1990), dan amigdala (Johnson et al., 1994) telah diamati pada duri MSN dendritik. Pengamatan ini dikombinasikan dengan peningkatan kepadatan tulang belakang setelah konsumsi sukrosa jangka panjang yang terlihat dalam penelitian kami, mendukung pembentukan input rangsang yang meningkat. Oleh karena itu, kemungkinan muncul di mana efek persisten yang disebabkan oleh asupan sukrosa seperti pesta panjang dapat memfasilitasi peningkatan aktivitas sinaptik rangsang pada dendrit distal MSNs di cangkang NAc. Akibatnya, pengurangan dan / atau pencabutan dendrit distal dapat terjadi melalui mekanisme homeostatis sinaptik (Reissner dan Kalivas, 2010), namun ini masih harus ditentukan.
Sangat menarik untuk dicatat bahwa Crombag dan rekannya menunjukkan bahwa tidak ada peningkatan kepadatan tulang belakang pada cangkang NAc setelah konsumsi sukrosa 4-minggu melalui paradigma administrasi-hidung-sodok meskipun akuisisi lebih kuat dan tingkat respons yang lebih tinggi terhadap sukrosa bila dibandingkan dengan amfetamin (Crombag et al., 2005). Pengamatan mereka tentang tidak adanya perubahan kepadatan tulang belakang pada minggu 4 mencerminkan temuan kami. Sebaliknya, penelitian kami menunjukkan bahwa setelah paparan jangka panjang (minggu 12) untuk konsumsi sukrosa kronis, ada peningkatan yang signifikan dalam kepadatan tulang belakang pada MSN pada tikus yang mengalami sukrosa. Selain itu, laboratorium kami sebelumnya telah menunjukkan bahwa konsumsi sukrosa jangka panjang (minggu 12) memfasilitasi respons farmakologis diferensial terhadap farmakoterapi yang telah terbukti memodulasi respons dopamin dan asetilkolin pada tingkat NAc (Shariff et al., Dalam pers). Secara keseluruhan, ini menunjukkan bahwa paparan sukrosa jangka panjang (12 minggu dan setelahnya), yang lebih akurat mencerminkan skenario dunia nyata, menghasilkan adaptasi morfologi pada tingkat NAc.
Dalam hal penyalahgunaan obat, paparan berulang terhadap berbagai obat menghasilkan perubahan jangka panjang dalam struktur dendrit dan duri dendritik. Sebagai contoh, amfetamin dan kokain meningkatkan kepadatan tulang belakang pada NAc pada kulit dan inti (Robinson dan Kolb, 2004). Paparan nikotin juga telah terbukti meningkatkan kepadatan tulang belakang di cangkang NAc. Sebaliknya, paparan morfin menyebabkan penurunan kepadatan tulang belakang dan kompleksitas cabang dendritik (Robinson dan Kolb, 2004). Dalam hal konsumsi sukrosa jangka panjang, kami mengamati peningkatan kepadatan tulang belakang yang mirip dengan amfetamin, kokain, dan nikotin dan berlawanan dengan efek morfin. Namun, tidak seperti amfetamin dan kokain, tetapi mirip dengan nikotin, peningkatan kepadatan tulang belakang pada paparan jangka panjang sukrosa terbatas pada cangkang NAc. Sangat menarik bahwa perubahan pada kedua percabangan dendritik (Robinson dan Kolb, 1999) dan kepadatan tulang belakang (Li et al., 2003) yang diproduksi oleh amfetamin atau kokain terbatas pada dendrit distal MSN di NAc, yang mencerminkan temuan dalam penelitian kami.. Lebih lanjut, dan sesuai dengan perubahan yang dijelaskan di atas, konsumsi sukrosa juga sebelumnya telah terbukti meningkatkan kekuatan sinaptik rangsang ke neuron dopamin akumbal (Stuber et al., 2008b) serta komponen lain dari jalur hadiah mesolimbik (Stuber et al., 2008a; Chen et al., 2010). Secara bersama-sama, ini menempatkan sukrosa sebagai modulator potensial morfologi neuron setelah penggunaan jangka panjang, yang mirip dengan efek yang diamati dari obat penyalahgunaan.
Meskipun investigasi lebih lanjut diperlukan untuk mengungkap mekanisme seluler dan sinaptik yang berkontribusi terhadap perubahan morfologis yang terlihat dalam penelitian ini, hasil kami menunjukkan efek neuron yang signifikan yang ditimbulkan oleh konsumsi sukrosa jangka panjang. Secara khusus, pertimbangan yang tidak diperiksa dalam penelitian kami adalah apakah efek morfologis yang diamati dari sukrosa juga dapat diperoleh dengan pemanis non-kalori seperti sakarin. Dalam hal ini, penting untuk dicatat bahwa Lenoir dan rekannya telah menunjukkan bahwa rasa manis yang intens melampaui hadiah kokain, baik itu dihasilkan oleh sakarin atau sukrosa (Lenoir et al., 2007). Selanjutnya, sebuah penelitian terbaru yang diterbitkan oleh lab kami (Shariff et al., Dalam pers) menunjukkan bahwa varenicline, agonis parsial reseptor asetilkolin nikotinat mengurangi baik sukrosa maupun asupan sakarin pada tikus mengikuti rejimen akses intermiten jangka panjang yang sama yang digunakan dalam penelitian ini. Menariknya, penelitian sebelumnya telah menunjukkan kesamaan dalam efek akut dari pemanis non-kalori seperti sakarin dan sukrosa pada tingkat NAc (Scheggi et al., 2013; Tukey et al., 2013; Carelli dan Barat, 2014). Namun, studi lebih lanjut diperlukan untuk menentukan apakah pemanis non-kalori dapat menyebabkan efek jangka panjang yang mirip dengan perubahan dalam morfologi cangkang NAc MSN yang disebabkan oleh konsumsi sukrosa jangka panjang yang dilaporkan di sini.
Kurangnya efek pada morfologi NAN MSN setelah konsumsi sukrosa jangka pendek, menyoroti pentingnya menerapkan penelitian jangka panjang untuk menilai dampak penyalahgunaan obat yang lama atau imbalan alami seperti sukrosa. Dalam hal ketergantungan, tidak hanya siklus berulang asupan pesta dan komponen kunci pantang dari siklus kecanduan, semakin banyak bukti mengungkapkan bahwa transisi menuju ketergantungan adalah proses progresif yang sering terjadi selama periode waktu yang panjang. Meskipun sifat adiktif gula tetap tidak pasti, masuk akal kecanduan hadiah non-narkoba lainnya seperti seks, perjudian dan makanan semakin diselidiki. Hasil dari penelitian ini menambah nilai hipotesis bahwa gula seperti sukrosa berpotensi memiliki sifat adiktif setelah konsumsi jangka panjang, seperti pesta. Hasil kami juga memiliki implikasi untuk meningkatnya jumlah anak-anak dan remaja yang mempertahankan kebiasaan makan yang tidak sehat (konsumsi gula tinggi dan makan berlebihan) hingga dewasa. Sejalan dengan peningkatan risiko terkena efek metabolik, ada kemungkinan juga bahwa konsekuensi neurologis dan psikiatris yang memengaruhi suasana hati dan motivasi juga dapat dihasilkan dari perilaku ini.
Kontribusi Penulis
Berpartisipasi dalam desain penelitian: PK, SB. Percobaan yang dilakukan: PK, MS, AB, MF, EM. Analisis data: PK, MF, MS. Menafsirkan data dan berkontribusi pada penulisan naskah: PK, MS, MF, EM, MB, SB. Semua penulis membaca dan menyetujui naskah akhir untuk diserahkan.
Pernyataan Benturan Kepentingan
Para penulis menyatakan bahwa penelitian ini dilakukan tanpa adanya hubungan komersial atau keuangan yang dapat ditafsirkan sebagai potensi konflik kepentingan.
Peninjau SC, SA dan Editor penanganan menyatakan afiliasi mereka bersama, dan Editor penanganan menyatakan bahwa proses tersebut tetap memenuhi standar peninjauan yang adil dan obyektif.
Ucapan Terima Kasih
Pekerjaan ini didukung oleh dana dari hibah dari Australian Research Council (FT1110884) ke SB dan Dewan Riset Kesehatan dan Medis Nasional (1061979) ke SB dan MB.
Materi tambahan
Bahan Pelengkap untuk artikel ini dapat ditemukan online di: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Gambar Tambahan 1. Asupan sukrosa dan preferensi dari tikus mengkonsumsi sukrosa 4 dan 12 minggu. (A, B) menunjukkan peningkatan asupan sukrosa total (ml) selama paparan 4 dan 12 minggu. (CD) menunjukkan preferensi tinggi untuk sukrosa daripada air selama periode presentasi sukrosa.
Referensi
Ahmed, SH, dan Koob, GF (1998). Transisi dari asupan obat moderat ke berlebihan: perubahan set point hedonis. Ilmu 282, 298 – 300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF, dan Parsons, LH (2003). Eskalasi pemberian kokain sendiri tidak tergantung pada perubahan nukleus yang diinduksi kokain meningkatkan kadar dopamin. J. Neurochem. 86, 102 – 113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR, dan Koob, GF (2000). Peningkatan terus-menerus dalam motivasi untuk memakai heroin pada tikus dengan riwayat peningkatan obat. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Diabetes tipe 2 pada anak-anak: aspek klinis dan faktor risiko. Horm Res 57 (Suppl. 1), 19 – 28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG, dan Gold, MS (2011). Tumpang tindih dalam nosologi penyalahgunaan zat dan makan berlebihan: implikasi translasi dari "kecanduan makanan". Curr. Rev. Penyalahgunaan Narkoba 4, 133 – 139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P., dan Hoebel, BG (2008). Bukti untuk kecanduan gula: efek perilaku dan neurokimiawi dari asupan gula yang intermiten dan berlebihan. Neurosci. Biobehav. Putaran. 32, 20 – 39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N., dan Hoebel, BG (2006). Sukrosa semu memberi makan pada jadwal pesta melepaskan accumbens dopamine berulang kali dan menghilangkan respon kenyang asetilkolin. Neuroscience 139, 813 – 820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Masuk akal kecanduan gula dan perannya dalam obesitas dan gangguan makan. Clin. Nutr. 29, 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA, dan Popkin, BM (2014). Gula makanan dan berat badan: sudahkah kita mencapai krisis epidemi obesitas dan diabetes ?: kesehatan terkutuk! Tuang gula. Perawatan diabetes 37, 950 – 956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY, dan Zahm, DS (1993). Pola persarafan aferen dari inti dan tempurung di bagian “accumbens” dari ventral striatum tikus: deteksi imunohistokimiawi dari fluoro-gold yang diangkut secara retrogradely. J. Comp. Neurol. 338, 255 – 278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., dan Kruseman, M. (2015). Minuman manis dan risiko obesitas pada anak-anak dan remaja: analisis sistematis tentang bagaimana kualitas metodologis dapat memengaruhi kesimpulan. J. Acad. Nutr. Diet. [Epub depan cetak]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM, dan Barat, EA (2014). Ketika selera yang baik berubah buruk: mekanisme saraf yang mendasari munculnya pengaruh negatif dan devaluasi hadiah alami yang terkait dengan kokain. Neurofarmakologi 76 (Pt B), 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW, dan Bonci, A. (2010). Plastisitas sinaptik dalam sistem mesolimbik: implikasi terapeutik untuk penyalahgunaan zat. Ann. NY Acad. Sci. 1187, 129 – 139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Bukti bahwa asupan gula berlebihan yang intermiten menyebabkan ketergantungan opioid endogen. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Kadet, JL, et al. (2001). Asupan gula berlebihan mengubah ikatan pada reseptor dopamin dan mu-opioid di otak. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B., dan Robinson, TE (2005). Efek berlawanan dari pengalaman administrasi-diri amfetamin pada duri dendritik di medial dan prefrontal cortex. Cereb. Cortex 15, 341 – 348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Penyelam, J., et al. (2014). Prevalensi tipe 1 dan diabetes tipe 2 di antara anak-anak dan remaja dari 2001 ke 2009. JAMA 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Epidemiologi dan genetika gangguan pesta makan (BED). CNS Spectr. 20, 522 – 529. doi: 10.1017 / s1092852915000462
Ford, ES, dan Dietz, WH (2013). Tren asupan energi di kalangan orang dewasa di Amerika Serikat: temuan dari NHANES. Saya. J. Clin. Nutr. 97, 848 – 853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD, dan Ogden, CL (2014). Prevalensi Kelebihan Berat Badan, Obesitas, dan Obesitas Ekstrem Di Antara Orang Dewasa: Amerika Serikat, 1960 – 1962 hingga 2011 – 2012. Atlanta, GA: CDC.
Graveland, GA, dan DiFiglia, M. (1985). Frekuensi dan distribusi neuron berukuran sedang dengan nukleus indentasi pada primata dan tikus neostriatum. Res otak. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV, dan Voorn, P. (1999). Konvergensi dan pemisahan input dan output striatal ventral. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Akhir sinaptik dan target postinaptiknya dalam neostriatum: spesialisasi sinaptik yang terungkap dari analisis bagian serial. Proc Natl. Acad. Sci. Amerika Serikat 77, 6926 – 6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC, dan Young, SJ (1994). Akson dopaminergik berlabel 5-hidroksidopamin: rekonstruksi akson tiga dimensi, sinapsis dan target pascasinaps pada tikus neostriatum. Neuroscience 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Diselesaikan: ada bukti ilmiah yang cukup bahwa mengurangi konsumsi minuman yang dimaniskan dengan gula akan mengurangi prevalensi obesitas dan penyakit terkait obesitas. Obes. Putaran. 14, 606 – 619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z., dan Totterdell, S. (1994). Masukan dari amigdala ke nukleus accumbens: hubungannya dengan imunoreaktivitas tirosin hidroksilase dan neuron yang teridentifikasi. Neuroscience 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H., dan Namba, M. (1981). Dua jenis terminal saraf dopaminergik di neostriatum tikus. Studi ultrastruktural. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW, dan Miller, JS (1984). Neurotensin neuron dalam proyek area ventral tegmental ke nukleus medial accumbens. Res otak. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ, dan Emson, PC (1990). Subtipe proyeksi sel-sel matriks neostriatal tikus diungkapkan dengan injeksi biocytin intraseluler. J. Neurosci. 10, 3421-3438.
Kelley, AE, dan Swanson, CJ (1997). Pemberian makan diinduksi oleh blokade reseptor AMPA dan kainate dalam ventral striatum: studi pemetaan mikroinfusi. Behav. Res otak. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM, dan Powell, TP (1971). Organisasi sinaptik dari nukleus kaudat. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403 – 412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mekanisme hadiah dalam obesitas: wawasan baru dan arah masa depan. Neuron 69, 664 – 679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Prevalensi dan korelasi gangguan pesta makan dalam Survei Kesehatan Mental Dunia Organisasi Kesehatan Dunia. Biol. Psikiatri 73, 904 – 914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, dan Bartlett, SE (2015). Karakterisasi struktural dan fungsional dari dendritik arbor dan input sinaptik GABAergik pada interneuron dan sel-sel utama dalam amigdala basolateral tikus. J. Neurophysiol. 114, 942 – 957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., dan Ahmed, SH (2007). Rasa manis yang intens melampaui hadiah kokain. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B., dan Robinson, TE (2003). Lokasi perubahan yang disebabkan oleh amfetamin persisten dalam kepadatan duri dendritik pada neuron berduri sedang dalam nukleus accumbens dan caudate-putamen. Neuropsychopharmacology 28, 1082 – 1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O., dan Björklund, A. (1978). Anatomi sistem neuron dopaminergik di otak tikus. Adv. Biokem. Psychopharmacol. 19, 1-23.
Lutter, M., dan Nestler, EJ (2009). Sinyal homeostatik dan hedonis berinteraksi dalam pengaturan asupan makanan. J. Nutr. 139, 629 – 632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ, dan Kelley, AE (1995). Reseptor glutamat dalam nukleus accumbens mengontrol perilaku makan melalui hipotalamus lateral. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP, dan Hu, FB (2010). Minuman manis, obesitas, diabetes melitus tipe 2, dan risiko penyakit kardiovaskular. Sirkulasi 121, 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM, dan Popkin, BM (2012). Penggunaan pemanis kalori dan non-kalori dalam makanan kemasan konsumen AS, 2005-2009. J. Acad. Nutr. Diet. 112, 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM, dan Popkin, BM (2002). Tren asupan energi di AS antara 1977 dan 1996: perubahan serupa terlihat di seluruh kelompok umur. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
Paxinos, G., dan Watson, C. (2007). Otak Tikus dalam Koordinasi Stereotaxic. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Apa Salahnya Pendekatan AS terhadap Obesitas? Mentor Virtual 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM, dan Hoebel, BG (2005). Makan setiap hari dengan gula berulang kali melepaskan dopamin dalam cangkang accumbens. Neuroscience 134, 737 – 744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW, dan McNeill, TH (1989). Penelitian Golgi tikus striatum: perubahan dendritik terkait usia pada populasi neuron yang berbeda. J. Comp. Neurol. 279, 212 – 227. doi: 10.1002 / cne.902790205
Ranjan, A., dan Mallick, BN (2010). Metode yang dimodifikasi untuk pewarnaan Golgi-cox yang konsisten dan andal dalam waktu yang sangat berkurang. Depan. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Tipe 2 diabetes mellitus pada anak-anak dan remaja. Dunia J. Diabetes 4, 270 – 281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ, dan Kalivas, PW (2010). Menggunakan homeostasis glutamat sebagai target untuk mengobati gangguan kecanduan. Behav. Farmakol 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM, dan Berridge, KC (2003). Ansambel motivasi glutamat dalam nucleus accumbens: gradien kulit rostrocaudal dari rasa takut dan makan. Eur. J. Neurosci. 17, 2187 – 2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM, dan Berridge, KC (2011). Nucleus accumbens interaksi dopamin / glutamat mengubah mode untuk menghasilkan hasrat versus ketakutan: D (1) saja untuk makan selera tetapi D (1) dan D (2) bersama karena rasa takut. J. Neurosci. 31, 12866 – 12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE, dan Kolb, B. (1999). Perubahan morfologi dendrit dan duri dendritik pada nukleus accumbens dan korteks prefrontal setelah perawatan berulang dengan amfetamin atau kokain. Eur. J. Neurosci. 11, 1598 – 1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE, dan Kolb, B. (2004). Plastisitas struktural terkait dengan pajanan terhadap penyalahgunaan obat. Neurofarmakologi 47 (Suppl. 1), 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC, dan Nestler, EJ (2010). Sinaps kecanduan: mekanisme plastisitas sinaptik dan struktural dalam nucleus accumbens. Tren Neurosci. 33, 267 – 276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J., dan Beatty, N. (1969). Sebuah studi tentang agunan akson sel piramidal di korteks serebral dewasa utuh dan sebagian terisolasi. Res otak. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC, dan Elmquist, JK (2002). Kebutuhan makan: kontrol makan homeostatis dan hedonis. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, dan Gambarana, C. (2013). Pengaruh kelezatan pada motivasi untuk beroperasi untuk makanan kalori dan non-kalori pada tikus yang kekurangan makanan dan kekurangan makanan. Neuroscience 236, 320 – 331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR, dan Pickel, VM (1990). Dalam nukleus medial nucleus accumbens, terminal hippocampal dan katekolaminergik bertemu pada neuron berduri dan saling bersesuaian. Res otak. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR, dan Pickel, VM (1992). Eferen kortikal prefrontal pada sinapsis tikus pada target neuron berlabel terminal katekolamin dalam nukleus accumbens septi dan pada neuron dopamin di daerah tegmental ventral. J. Comp. Neurol. 320, 145 – 160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (dalam pers). Modulator reseptor asetilkolin neuronal nikotinat mengurangi asupan gula. PLoS ONE.
Sheehan, DV, dan Herman, BK (2015). Faktor psikologis dan medis yang terkait dengan gangguan makan pesta yang tidak diobati. Formal. Peduli Pendamping Gangguan CNS. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Akses intermiten ke etanol 20% menginduksi konsumsi etanol yang tinggi pada tikus Long-Evans dan Wistar. Alkohol. Clin. Exp. Res. 32, 1816 – 1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, dan Bartlett, SE (2007). Varenicline, agonis parsial reseptor nikotin asetilkolin alfa4beta2, secara selektif mengurangi konsumsi dan pencarian etanol. Proc Natl. Acad. Sci. Amerika Serikat 104, 12518 – 12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., dan Bonci, A. (2008a). Asupan etanol sukarela meningkatkan kekuatan sinaptik rangsang di daerah tegmental ventral. Alkohol. Clin. Exp. Res. 32, 1714 – 1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, dkk. (2008b). Isyarat prediktif penghargaan meningkatkan kekuatan sinaptik rangsang ke neuron dopamin otak tengah. Ilmu 321, 1690 – 1692. doi: 10.1126 / science.1160873
Swanson, SA, Gagak, SJ, Le Grange, D., Swendsen, J., dan Merikangas, KR (2011). Prevalensi dan korelasi gangguan makan pada remaja. Hasil dari suplemen remaja replikasi survei komorbiditas nasional. Lengkungan. Jenderal Psikiatri 68, 714 – 723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., dan Mann, J. (2013). Gula makanan dan berat badan: tinjauan sistematis dan meta-analisis uji coba terkontrol secara acak dan studi kohort. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S., dan Smith, AD (1989). Konvergensi input hippocampal dan dopaminergik ke neuron yang diidentifikasi dalam nucleus accumbens tikus. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., dkk. (2013). Konsumsi sukrosa menginduksi perdagangan reseptor AMPA dengan cepat. J. Neurosci. 33, 6123 – 6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM, dan Lieven, PA (1980). Organisasi topikal aferen ke caudatoputamen tikus. Sebuah studi peroksidase lobak. Neuroscience 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R., dan Contreras, AM (2014). Dasar neurobiologis dari keinginan untuk karbohidrat. makanan 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., dan Baler, R. (2012). Hadiah makanan dan obat-obatan: sirkuit yang tumpang tindih dalam obesitas dan kecanduan manusia. Curr. Teratas. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Bijaksana, RA (1973). Asupan etanol sukarela pada tikus setelah paparan etanol pada berbagai jadwal. Psikofarmakologia 29, 203 – 210. doi: 10.1007 / BF00414034
Wright, CI, dan Groenewegen, HJ (1995). Pola konvergensi dan segregasi pada nukleus medial accumbens tikus: hubungan kortikal prefrontal, garis tengah thalamic, dan aferen amigdaloid basal. J. Comp. Neurol. 361, 383 – 403. doi: 10.1002 / cne.903610304
Kata kunci: konsumsi binge-like, jangka panjang, neuron berduri sedang, nucleus accumbens, sukrosa
Kutipan: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC dan Bartlett SE (2016) Konsumsi Berkepanjangan dari Sukrosa dalam Cara Binge-Seperti, Mengubah Morfologi Neuron Berduri Sedang di Shell Nucleus Accumbens. Depan. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Diterima: 03 Desember 2015; Diterima: 07 Maret 2016;
Diterbitkan: 23 Maret 2016.
Diedit oleh:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Prancis
Diulas oleh:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Prancis
Stéphanie Caille, Centre National de la Recherche Scientifique, Prancis
Hak Cipta © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham dan Bartlett. Ini adalah artikel akses terbuka yang didistribusikan di bawah ketentuan Lisensi Atribusi Creative Commons (CC BY). Penggunaan, distribusi atau reproduksi di forum lain diizinkan, asalkan penulis asli atau pemberi lisensi dikreditkan dan bahwa publikasi asli dalam jurnal ini dikutip, sesuai dengan praktik akademik yang diterima. Dilarang menggunakan, mendistribusikan, atau mereproduksi, yang tidak mematuhi ketentuan ini.
* Korespondensi: Selena E. Bartlett, [email dilindungi]