YBOP COMMENTS: ຜົນກະທົບຂອງ Coolidge ແມ່ນຢູ່ເບື້ອງຫຼັງພະລັງຂອງຄອມອິນເຕີເນັດ. ຜົນກະທົບຂອງ Coolidge ແມ່ນປະກົດການທີ່ເຫັນໄດ້ໃນສັດລ້ຽງລູກດ້ວຍນົມແມ່ເຊິ່ງຜູ້ຊາຍ (ແລະເພດຍິງທີ່ມີ ຈຳ ນວນ ໜ້ອຍ ກວ່າ) ສະແດງຄວາມສົນໃຈທາງເພດ ໃໝ່ ຖ້າ ນຳ ສະ ເໜີ ຄູ່ຮັກຮ່ວມເພດ ໃໝ່, ເຖິງແມ່ນວ່າຫຼັງຈາກປະຕິເສດການຮ່ວມເພດຈາກຄູ່ຮ່ວມເພດກ່ອນແຕ່ກໍ່ຍັງມີຢູ່. ຄວາມແປກ ໃໝ່ ກ່ຽວກັບເພດ ສຳ ພັນຂ້າມນິໄສນີ້ດ້ວຍຄວາມຕື່ນເຕັ້ນ ໃໝ່ ທີ່ເກີດຈາກ dopamine ທີ່ສູງຂື້ນ. ກະແສ ໃໝ່ ຢ່າງຕໍ່ເນື່ອງຂອງສິ່ງ ໃໝ່ໆ ແມ່ນສິ່ງທີ່ເຮັດໃຫ້ຄອມພີວເຕີ້ທາງອິນເຕີເນັດແຕກຕ່າງຈາກຄອມພີວເຕີ້ໃນອະດີດ.
- Dennis F. Fiorino,
- Ariane Coury, ແລະ
- Anthony G Phillips
- The Journal of Neuroscience, 15 ເດືອນມິຖຸນາ 1997, 17 (12): 4849-4855;
ບົດຄັດຫຍໍ້
ຜົນກະທົບຂອງ Coolidge ອະທິບາຍເຖິງການປະຕິບັດຕົວໃຫມ່ຂອງພຶດຕິກໍາທາງເພດໃນສັດທີ່ມີຄວາມສໍາຄັນທາງເພດໃນການຕອບສະຫນອງຕໍ່ການຕອບສະຫນອງຂອງຜູ້ຮັບໃຫມ່. ໃນການເລີ່ມຕົ້ນແລະບໍາລຸງຮັກສາພຶດຕິກໍາທີ່ກະຕຸ້ນ, microdialysis ໄດ້ຖືກນໍາໃຊ້ເພື່ອຕິດຕາມການລະບາດຂອງ Nucleus accumbens (NAC) ໃນໄລຍະການຮ່ວມເພດ, ຄວາມໂປ່ງໃສທາງເພດແລະການກະທໍາທາງເພດໃຫມ່. ໃນການຕົກລົງກັບບົດລາຍງານໃນເບື້ອງຕົ້ນ, ການນໍາສະເຫນີຂອງແມ່ຍິງທີ່ບໍ່ມີຕົວຕົນທີ່ຢູ່ເບື້ອງຫລັງຫນ້າຈໍແລະການຮ່ວມເພດແມ່ນມີຄວາມກ່ຽວຂ້ອງກັບການເພີ່ມຂຶ້ນຂອງ NAC DA efflux. ການກັບຄືນຂອງ NAC DA ສະເລ່ຍໃນແຕ່ລະຈຸດແມ່ນທຽບກັບໄລຍະເວລາຂອງຄວາມໂປ່ງໃສທາງເພດ, ເຖິງແມ່ນວ່າຄວາມເຂັ້ມຂົ້ນຂອງວິຕາມິນ DA, ອາຊິດ dihydroxyphenylacetic ແລະ acid homovanillic, ແມ່ນສູງ. ການນໍາສະເຫນີຂອງແມ່ຍິງທີ່ໄດ້ຮັບຄວາມນິຍົມດ້ານຫລັງຈາກຫນ້າຈໍໄດ້ເຮັດໃຫ້ເກີດການເພີ່ມຂຶ້ນເລັກນ້ອຍໃນ NAC DA, ເຊິ່ງໄດ້ເພີ່ມຂຶ້ນຢ່າງຫຼວງຫຼາຍໃນລະຫວ່າງການຮ່ວມກັບຄົນໃຫມ່. ຂໍ້ມູນໃນປັດຈຸບັນຊີ້ໃຫ້ເຫັນວ່າຄຸນສົມບັດຂອງ stimulus ຂອງແມ່ຍິງທີ່ໄດ້ຮັບຄວາມນິຍົມໃຫມ່ອາດຈະເຮັດໃຫ້ເກີດການຕິດເຊື້ອ NAC DA ໃນຫນູຂອງຜູ້ຊາຍທີ່ມີທາງກອ່ນທາງເພດ, ແລະນີ້ກໍ່ອາດຈະກ່ຽວຂ້ອງກັບການເລີ້ມຄືນໃຫມ່ຂອງພຶດຕິກໍາທາງເພດ.
ການນໍາສະເຫນີ
ຫນູຂອງຜູ້ຊາຍທີ່ມີການພົວພັນກັບຄວາມອີ່ມໃຈສາມາດເຮັດໃຫ້ມີເພດສໍາພັນອີກຄັ້ງຖ້າແມ່ຍິງເລີ່ມຕົ້ນໄດ້ຮັບການແຕ່ງຕັ້ງໂດຍແມ່ຍິງທີ່ໄດ້ຮັບຄວາມນິຍົມໃຫມ່. ນີ້ໄດ້ຖືກເອີ້ນວ່າເປັນຜົນກະທົບຂອງ Coolidge ແລະໄດ້ຖືກສັງເກດເຫັນໃນຈໍານວນສັດລ້ຽງລູກດ້ວຍນົມແມ່ (Wilson et al, 1963) ປັດໄຈທົ່ວໄປເຊັ່ນ: ການຊຶມເສົ້າຫຼືການຊຶມເສົ້າ motoric ບໍ່ພຽງພໍທີ່ຈະອະທິບາຍສະຖານະພາບຂອງການສະທ້ອນທາງເພດ, ເນື່ອງຈາກວ່າກະຕຸ້ນຈາກແມ່ຍິງໃຫມ່ໆຍັງສາມາດເຮັດໃຫ້ການຮ່ວມເພດ. ຄວາມອີ່ມໃຈທາງເພດສໍາພັນຍັງສາມາດ "ຫັນຫນ້າ" ທາງດ້ານ pharmacological, ໃນລະດັບທີ່ສໍາຄັນ, ໂດຍການນໍາໃຊ້ຢາຊະນິດຕ່າງໆທີ່ສາມາດປະຕິບັດໃນລະບົບທາງ neurotransmitter ຕ່າງໆ. ຢາເຫຼົ່ານີ້ລວມມີ yohimbine, 8-OH-DPAT (Rodriguez-Manzo ແລະ Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus and Gorzalka, 1987; Rodriguez-Manzo ແລະ Fernandez-Guasti, 1995a,b), ແລະ apomorphine (Mas et al, 1995c) ເຖິງແມ່ນວ່າການປະຕິບັດພາຍໃນຂອງຢາເຫຼົ່ານີ້ບໍ່ສາມາດຖືກຍົກເລີກໄດ້ (ຕົວຢ່າງ, ຜົນກະທົບຕໍ່ adrenergic ໃນຫນ້າທີ່ erectile), ຜົນກະທົບກ່ຽວກັບກົນໄກກາງທີ່ຢູ່ພາຍໃຕ້ຄວາມໃຈຮ້າຍທາງເພດແມ່ນໄດ້ຖືກນໍາສະເຫນີໂດຍອີງໃສ່ການທົດລອງເລັບທ່ີທ່ີຕັ້ງສູນກາງ (noradrenergic lesion)Rodriguez-Manzo ແລະ Fernandez-Guasti, 1995a) ແລະການທົດລອງ microdialysis ທີ່ຕິດຕາມກວດກາການຍ່ອຍອາຫານຂອງ dopaminergic ໃນພື້ນທີ່ preoptic medial (Mas et al, 1995a,b).
ເນື່ອງຈາກວ່າກົນໄກກາງສາມາດສະທ້ອນໃຫ້ເຫັນເຖິງລັກສະນະການປະຕິບັດທາງເພດຂອງຜົນກະທົບຂອງ Coolidge, ຜູ້ນໍາທີ່ອາດຈະເປັນລະບົບ dopamine mesolimbic (DA) ທີ່ຄາດຄະເນຈາກພື້ນທີ່ທີ່ມີອາການບວມໄປສູ່ NAC. Mesolimbic DA ເບິ່ງຄືວ່າເປັນຕົວປ່ຽນແປງຫຼັກໃນຂະບວນການເຊື່ອມໂຍງທີ່ຊັບຊ້ອນທີ່ກ່ຽວຂ້ອງກັບການປະເມີນຜົນກະຕຸ້ນຂອງສິ່ງແວດລ້ອມ, ເຊັ່ນ: ຄໍາໂຄສະນາຈາກແມ່ຍິງທີ່ໄດ້ຮັບທາງເພດສໍາພັນ, ແລະການຈັດຕັ້ງພຶດຕິກໍາເປົ້າຫມາຍ,Fibiger ແລະ Phillips, 1986; Blackburn et al, 1992; Phillips et al, 1992; LeMoal, 1995; Salamone, 1996).
ເຖິງແມ່ນວ່າ neurons midbrain DA ຕອບສະຫນອງກັບລາງວັນຕົ້ນຕໍແລະການຄາດຄະເນຂອງລາງວັນ, ສິ່ງປະດິດສິ່ງແວດລ້ອມ Novell ຫຼື unpredictable ເຮັດໃຫ້ການເຄື່ອນໄຫວ neuronal ຫຼາຍທີ່ສຸດຢ່າງແຂງແຮງໃນໄລຍະການຝຶກອົບຮົມຊ້ໍາFabre et al, 1983; Schultz, 1992; Mirenowicz and Schultz, 1994) ມີຫຼັກຖານທີ່ຍິ່ງໃຫຍ່ທີ່ສະຫນັບສະຫນູນບົດບາດໃນການປະສານງານທີ່ສໍາຄັນສໍາລັບ DA mesolimbic ໃນການເລີ່ມຕົ້ນແລະບໍາລຸງຮັກສາພຶດຕິກໍາທາງເພດຂອງຫນູ (Pfaus ແລະ Everitt, 1995), ແລະຈໍານວນຂອງການສຶກສາ microdialysis ເພີ່ມຂຶ້ນລາຍງານໃນ NAC DA efflux ໃນລະຫວ່າງຂັ້ນຕອນທີ່ຫນ້າທຶ່ງແລະການບໍລິໂພກຂອງພຶດຕິກໍາທາງເພດຊາຍ (Pfaus et al, 1990; Pleim et al, 1990; Damsma et al, 1992; Wenkstern et al, 1993; Fumero et al, 1994; Mas et al, 1995b) ຢ່າງໃດກໍຕາມ, ມີຂໍ້ມູນຂ້ອນຂ້າງຫນ້ອຍກ່ຽວກັບການເຊື່ອມໂຍງ neurochemical ຂອງ satiation ທາງເພດແລະ reinitiation ຂອງພຶດຕິກໍາທາງເພດສໍາພັນໄດ້. ຄໍາຮ້ອງສະຫມັກຂອງ in vivo microdialysis ເພື່ອຕິດຕາມກວດກາ mesolimbic DA neurotransmission ໃນລະຫວ່າງຜົນກະທົບ Coolidge ມີໂອກາດທີ່ຈະພິຈາລະນາພາລະບົດບາດຂອງ NAC DA ໃນການຮ່ວມເພດ, ຄວາມໂປ່ງໃສທາງເພດແລະ reinitiation ຂອງ copulation.
ການທົດລອງ microdialysis ໄດ້ຖືກປະຕິບັດເພື່ອກໍານົດດັ່ງຕໍ່ໄປນີ້: (1) ວ່າການເລີ່ມຕົ້ນຂອງຄວາມໂປ່ງໃສທາງເພດແມ່ນປະກອບດ້ວຍການກັບຄືນຂອງ extracellular DA concentrations ໃນ NAC ກັບຄ່າ precopulation ຫຼືຂ້າງລຸ່ມນີ້ແລະ (2) ບໍ່ວ່າຈະ reinstatement ຂອງພຶດຕິກໍາ copulatory ໃນ " sexually satiated "ຫນູຊາຍທີ່ມີເພດຍິງທີ່ມີນ້ໍາໃຈໃຫມ່ມີຄວາມກ່ຽວຂ້ອງກັບການເພີ່ມຂຶ້ນຂອງ NAC DA efflux.
ອຸປະກອນແລະວິທີການ
Subjects ເພດຫນູ Sprague Dawley, ທີ່ໄດ້ຈາກສູນການດູແລສັດ (ມະຫາວິທະຍາໄລ British Columbia), ແລະຫນູຫນີເດັກນ້ອຍ Long-Evans, ທີ່ໄດ້ຮັບຈາກ Charles River Canada (St Constant, Quebec, Canada), ໄດ້ຖືກເກັບໄວ້ໃນຖັງຕາຫນ່າງເຫຼັກ (18 25 × 65 cm, ຫ້າໃນ 1 cage) ໃນຫ້ອງກວ້າງແຍກຕ່າງຫາກ. ຫ້ອງການອານານິຄົມໄດ້ຮັບການຮັກສາໄວ້ຢູ່ໃນອຸນຫະພູມຂອງ ~20 ° C ໃນຮອບວຽນ 12 ຊົ່ວໂມງກັບແສງສະຫວ່າງ / ຊ້ໍາ. ຫນູໄດ້ເຂົ້າເຖິງອາຫານບໍ່ຈໍາກັດ (Purina Rat Chow) ແລະນ້ໍາ.
ການຜ່າຕັດແລະການທົດສອບພຶດຕິກໍາກ່ອນທີ່ຈະ microdialysis ຂອງສະຫມອງ.ຫນູເພດຍິງໄດ້ຮັບການຜ່າຕັດທາງດ້ານສອງດ້ານພາຍໃຕ້ການໃຊ້ຢາ Halotane (Fluothane, Ayerst Laboratories) ຢ່າງຫນ້ອຍ 4 ອາທິດກ່ອນການທົດສອບ. ການໄດ້ຮັບທາງເພດໃນເພດຍິງທີ່ຖືກກະຕຸ້ນໂດຍການສັກຢາ estradiol benzoate (10 μg) ແລະ progesterone (500 μg), 48 ແລະ 4 hr, ຕາມລໍາດັບແຕ່ລະຄັ້ງກ່ອນການກວດໃນແຕ່ລະຄັ້ງ. ຫນູຊາຍໄດ້ຖືກກວດສອບສໍາລັບພຶດຕິກໍາທາງເພດສອງຄັ້ງ, 4 d ແຍກ, ຢູ່ໃນຫ້ອງ Plexiglas (35 × 35 × 40 cm) ດ້ວຍຊັ້ນຊັ້ນລວດເຫຼັກ. ພຽງແຕ່ຫນູຜູ້ທີ່ມີມາດຕະຖານການປະຕິບັດ, ເຊິ່ງລວມມີ intromission ພາຍໃນ 5 ນາທີຂອງການນໍາສະເຫນີຂອງເພດຍິງແລະ ejaculation ພາຍໃນ 15 ນາທີຂອງການ intromission ຄັ້ງທໍາອິດ, ໃນລະຫວ່າງການທົດສອບການກວດສອງຄັ້ງໄດ້ຖືກນໍາໄປໃສ່ດ້ວຍ microdialysis probe guide cannulae.
ຊາຍຫນູ (n = 5) ຖືກ anesthetized ດ້ວຍຢາ ketamine hydrochloride (100 mg / kg, ip) ແລະ xylazine (10 mg / kg, ip) ກ່ອນການຜ່າຕັດ stereotaxic. ປື້ມຄູ່ມືສໍາລັບການຄົ້ນຫາ microdialysis ປືນໃຫຍ່ (19 gauge) ໄດ້ຖືກປະສົມສອງຂ້າງໃນລະດັບ NAC (ປະສົມຈາກເບກເກີ: ຂ້າງຫນ້າ, + 1.7 ມມ, medial, 1.1 ມົມ, ventral, -1.0 ມມ, ກະໂຫຼກກະຮອກ) ແລະໄດ້ຮັບການຮັກສາໄວ້ກັບກະໂຫຼກດ້ວຍຢາ acrylic ແຂ້ວແລະ screws ຂອງເພັດ. ການນໍາ implants ຄູ່ມືສອງຄູ່ມືໄດ້ຖືກນໍາໃຊ້ເພື່ອ maximize ໂອກາດສໍາລັບການທົດລອງ microdialysis ສົບຜົນສໍາເລັດ. ໂຊກດີ, ໃນການທົດລອງໃນປະຈຸບັນ, ມີພຽງແຕ່ຫນຶ່ງຫມຶກສໍາລັບແຕ່ລະຫນູ. ຫນູແມນຢູ່ໃນຖັງພາດສະຕິກຂະຫນາດໃຫຍ່ທີ່ມີຝາຂີ້ມູກສໍາລັບສ່ວນທີ່ເຫລືອຂອງການທົດລອງ. ຫນຶ່ງອາທິດຫຼັງຈາກການຜ່າຕັດ, ຫນູໄດ້ຮັບການທົດສອບສໍາລັບພຶດຕິກໍາທາງເພດສໍາພັນ. ໃນລະຫວ່າງການຝຶກອົບຮົມສ່ວນນີ້, ຫ້ອງທົດສອບໄດ້ຖືກຕິດຕັ້ງດ້ວຍຫນ້າຈໍ Plexiglas ທີ່ເລື່ອນລົງເຊິ່ງແບ່ງຫ້ອງອອກເປັນຫ້ອງຂະຫນາດໃຫຍ່ແລະຂະຫນາດນ້ອຍ. ຫນູຊາຍໄດ້ຖືກນໍາເຂົ້າເຂົ້າໄປໃນຫ້ອງຂະຫນາດໃຫຍ່ແລະ 15 min ຕໍ່ມາ, ຍິງຖືກວາງຢູ່ຫລັງຫນ້າຈໍ. ຫຼັງຈາກໄລຍະເວລາກຽມກຽມ 15 ນາທີ, ຫນ້າຈໍໄດ້ຖືກເອົາອອກ, ແລະຫນູໄດ້ຖືກອະນຸຍາດໃຫ້ເຮັດສໍາເນົາສໍາລັບ 30 ນາທີ. ສາມການຝຶກອົບຮົມໄດ້ຖືກດໍາເນີນການ, ຫນຶ່ງໃນທຸກໆ 4 d. ຫມູທັງຫມົດໄດ້ບັນລຸເງື່ອນໄຂການປະຕິບັດໃນລະຫວ່າງທຸກໆພາກ.
Coolidge effect experiment Rats ໄດ້ຖືກຈັດຕັ້ງເປັນ unilaterally ກັບ probes microdialysis 12-18 ຊົ່ວໂມງກ່ອນການທົດລອງຜົນກະທົບ Coolidge ແລະຈັດວາງຢູ່ໃນຫ້ອງຂະຫນາດໃຫຍ່ຂອງຫ້ອງການທົດສອບທີ່ມີການເຂົ້າເຖິງອາຫານແລະນ້ໍາຟຣີ. ໃນຕອນເຊົ້າຂອງການທົດລອງ, ຕົວຢ່າງ microdialysis ໄດ້ຖືກເກັບກໍາທຸກ 15 ນາທີ. ການທົດລອງປະກອບດ້ວຍເຈັດຂັ້ນຕໍ່ໄປຕໍ່ໄປນີ້: (1) ເສັ້ນເບື້ອງຕົ້ນ (ຢ່າງຫນ້ອຍ 60 ນາທີ); (2) ຍິງ 1 ຫລັງຫນ້າຈໍ (15 ນາທີ); (3) ການຮ່ວມເພດກັບແມ່ຍິງ 1 ຈົນກວ່າໄລຍະເວລາ 30 ຜ່ານໂດຍບໍ່ມີການຕິດຕັ້ງ; (4) reintroduction ຂອງແມ່ຍິງ 1 ຫລັງຫນ້າຈໍ (15 ນາທີ); (5) ການເຂົ້າຫາແມ່ຍິງ 1 ສໍາລັບໄລຍະເວລາ 15 ທີ່ບໍ່ມີການຕິດຕັ້ງ (ຖ້າການຕິດຕັ້ງເກີດຂຶ້ນ, ໄລຍະນີ້ໄດ້ຮັບການປິ່ນປົວເປັນຂັ້ນຕອນ 3); (6) ການແນະນໍາຂອງຍິງ 2 ຫລັງຫນ້າຈໍ (15 ນາທີ); 7) ຮ່ວມກັບເພດຍິງ 2 ສໍາລັບ 60 ນາທີ.
ທັດສະນະຄະຕິໄດ້ຖືກຖ່າຍທອດພາຍໃຕ້ແສງສະຫວ່າງຕ່ໍາໂດຍໃຊ້ລະບົບວິດີໂອ JVC ແລະສັງເກດເຫັນຢູ່ໃນຈໍສະແດງວິດີໂອທີ່ຢູ່ນອກຫ້ອງທົດລອງ. ມາດຕະຖານມາດຕະຖານຂອງພຶດຕິກໍາທາງເພດຖືກບັນທຶກໂດຍໃຊ້ຄອມພິວເຕີແລະຊອບແວທີ່ເຫມາະສົມ (Holmes et al, 1987).
ຫຼັງຈາກການທົດລອງ microdialysis, ສັດໄດ້ຮັບນ້ໍາເກີນ chloral hydrate ແລະ perfrosed intracardially ດ້ວຍ saline ແລະ formalin (4%). ສະຫມອງໄດ້ຖືກແບ່ງປັນແລະແຊ່ແຂໍງແລະຫຼັງຈາກນັ້ນ, ສ່ວນ Coronal ແມ່ນມີສີຂີ້ເຖົ່າດ້ວຍ cresyl violet ເພື່ອກໍານົດຕໍາແຫນ່ງຂອງ probes microdialysis. ມີພຽງແຕ່ຫນູທີ່ມີການທົດສອບການກວດສອບພາຍໃນ NAC ໄດ້ຖືກນໍາໃຊ້ສໍາລັບການວິເຄາະພຶດຕິກໍາແລະທາງດ້ານວັດຖຸ.
Microdialysis ແລະການຄົ້ນພົບ HPLC-electrochemical. ການກວດສອບ Microdialysis ມີຈຸດສຸມໃນການອອກແບບທີ່ມີເສັ້ນໃຍເສັ້ນໃຍທີ່ເປັນ semipermeable (ເສັ້ນຜ່າສູນກາງ 2, 340 μmເສັ້ນຜ່າກາງດ້ານນອກ, ການຕັດນ້ໍາຫນັກຕົວຢ່າງ 65000, Filtral 12, Hospal) ຢູ່ປາຍ distal. Probes ໄດ້ຖືກ perfused ຢູ່ 1.0 μl / ນາທີທີ່ມີການແກ້ໄຂ Ringer ຂອງດັດແກ້ (0.01 ມ sodium phosphate buffer, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, NaNUM xNUMX ມມ) ໃຊ້ syringe gastight (Hamilton, Reno, NV) ແລະສູບຢາ syringe (ແບບ 147, ເຄື່ອງມື Harvard, South Natick, MA). ກະເປົາຄູ່ມືການກວດສອບ microdialysis ໄດ້ຖືກນໍາໃຊ້ເພື່ອຮັບປະກັນການກວດສອບ microdialysis ພາຍໃນ cannula ຄູ່ມື. ສາຍທໍ່ເຫຼັກທີ່ຕິດກັບຫມໍ້ນ້ໍາແຫຼວ (Instech 22s) ທີ່ຕິດຢູ່ເທິງຫ້ອງທົດສອບຖືກນໍາໃຊ້ເພື່ອປ້ອງກັນທໍ່ດູດFiorino et al, 1993).
ການວິເຄາະ Microdialysate ເຊິ່ງລວມທັງ DA ແລະອາຫານທີ່ມີການປ່ຽນແປງຂອງທາດອາຫານທີ່ມີປະສິດທິພາບສູງ, ) ໂດຍນໍາໃຊ້ຕົວຖັງໂຊດຽມໂຊດຽມ 5m, pH 15 (4.6% methanol). ຄວາມເຂັ້ມຂົ້ນຂອງການວິເຄາະໄດ້ຖືກປະເມີນໂດຍການກວດຫາໄຟຟ້າ (EC). ອຸປະກອນດັ່ງກ່າວປະກອບດ້ວຍປັ໊ມ Bio-Rad (Richmond, CA), ເຄື່ອງສີດສອງຕໍາແຫນ່ງ Valco Instruments (Houston, TX), ເຄື່ອງກວດຈັບ Coulochem II EC (Bedford, MA), ແລະເຄື່ອງບັນທຶກກາຟສອງຊ່ອງ (Kipp ແລະ Zonen, Bohemia, NY). ຕົວກໍານົດການກວດຈັບໄຟຟ້າແມ່ນດັ່ງຕໍ່ໄປນີ້: electrode 0.083, + 3.5 mV; electrode 5, -10 mV ແລະຫ້ອງກອງ, -1 mV. ການສືບສວນແບບທໍາມະດາຟື້ນຟູ, ດໍາເນີນການ ໃນ vitro ແລະຢູ່ໃນອຸນຫະພູມຫ້ອງ, 22% ສໍາລັບ DA, 18% ສໍາລັບ DOPAC ແລະ 18% ສໍາລັບ HVA.
ຜົນໄດ້ຮັບ
ພຶດຕິກໍາ
ມາດຕະການດ້ານພຶດຕິກໍາຈາກການທົດລອງຜົນກະທົບ Coolidge ແມ່ນສະແດງຢູ່ໃນຕາຕະລາງ 1ທີ່ຢູ່ Latencies ເພື່ອ mount, intromit, ແລະ ejaculate, ເຊັ່ນດຽວກັນກັບໄລຍະ postjaculatory ຫຼັງຈາກການອິດເມື່ອຍຄືກັນກັບທີ່ຢູ່ໃນກອງປະຊຸມຝຶກອົບຮົມທີ່ຜ່ານມາ (ຂໍ້ມູນທີ່ບໍ່ໄດ້ສະແດງໃຫ້ເຫັນ). ນີ້ຊີ້ໃຫ້ເຫັນວ່າຂັ້ນຕອນຂອງ microdialysis ບໍ່ປ່ຽນແປງພຶດຕິກໍາທາງເພດປົກກະຕິ. (7.8 0.5), ການຫຼຸດລົງໃນຈໍານວນການຮ່ວມເພດກ່ອນການອອກກໍາລັງກາຍແຕ່ລະຄັ້ງແລະການເພີ່ມຂື້ນໃນໄລຍະເວລາ postjaculatory (ຂໍ້ມູນທີ່ບໍ່ໄດ້ສະແດງໃຫ້ເຫັນ) , ແມ່ນຄ້າຍຄືກັນກັບການລາຍງານໃນການສຶກສາກ່ອນຫນ້ານີ້ (ຫາດຊາຍແລະຈໍແດນ, 1956; Fowler ແລະ Whalen, 1961; Fisher, 1962; Bermant et al, 1966; Rodriguez-Manzo ແລະ Fernandez-Guasti, 1994; Mas et al, 1995d) ການປ່ຽນແປງບຸກຄົນໄດ້ຖືກສັງເກດເຫັນກ່ຽວກັບຈໍານວນຂອງການປະຕິກິລິຍາທີ່ເກີດຂຶ້ນກັບແມ່ຍິງ 1, ເວລາທີ່ໃຊ້ຮ່ວມກັບເພດຍິງ 1, ແລະຈໍານວນການນໍາສະເຫນີຂອງແມ່ຍິງ 1 ທີ່ຈໍາເປັນເພື່ອບັນລຸເງື່ອນໄຂການອີ່ມຕົວ (ຕາຕະລາງ 1, ດ້ານລຸ່ມ). ບາງຫນູຈໍາເປັນຕ້ອງມີການສົ່ງຫນັງສືຈໍານວນຫລາຍຂອງແມ່ຍິງ 1 ຈົນກ່ວາໄລຍະ 5 ແລ້ວສົມບູນ (n = 3) ການກະທໍາຂອງເພດຍິງ 1 ຢູ່ຫລັງຫນ້າຈໍແລະການປົດຕໍາແຫນ່ງແຜ່ນປ້າຍໄວ້ບ່ອນອາດຈະເປັນຄໍາແນະນໍາທີ່ຫນ້າປະຫລາດຫລັກທີ່ນໍາໄປສູ່ການຮ່ວມເພດ. ມັນຄວນຈະໄດ້ຮັບການສັງເກດເຫັນວ່າມາດຕະຖານຂອງການສະສົມຂອງ 30 min ໂດຍບໍ່ມີການ mount, ເຖິງແມ່ນວ່ານໍາໃຊ້ກ່ອນຫນ້ານີ້ (ຫາດຊາຍແລະຈໍແດນ, 1965; Mas et al, 1995b), ມັນແມ່ນຄວາມຄິດທີ່ບໍ່ດີແລະບໍ່ຮັບປະກັນວ່າຫນູຈະບໍ່ຕິດຕັ້ງເວລາຫຼາຍກວ່າ. ເຖິງຢ່າງໃດກໍ່ຕາມ, ການຊັກຊ້າຫຼືຂັ້ນຕອນການປົດຕໍາແຫນ່ງແລະການທົດແທນບໍ່ໄດ້ມີຜົນຕໍ່ຄວາມຖືກຕ້ອງໃນການຮ່ວມເພດໃຫມ່ກັບແມ່ຍິງ 1 (ເຊົ່ນໄລຍະ 4 ແລະ 5).

ພຶດຕິກໍາໃນໄລຍະທົດລອງຜົນກະທົບ Coolidge
ທຸກໆຫນູສະແດງໃຫ້ເຫັນຜົນກະທົບຂອງ Coolidge. ກິດຈະກໍາທີ່ກ່ຽວຂ້ອງກັບບ່ອນວາງສະແດງຂອງຍິງ 2 ຢູ່ຫລັງຫນ້າຈໍແລະໂດຍສະເພາະການຖອນການແບ່ງປັນອາດຈະມີສ່ວນຮ່ວມໃນຜົນໄດ້ຮັບນີ້ແຕ່ອີກເທື່ອຫນຶ່ງ, ເຫດການເຫຼົ່ານີ້ບໍ່ພຽງພໍຕໍ່ການປ່ຽນແປງກ່ອນຫນ້ານີ້ໃນການທົດລອງ. ການປຽບທຽບລະຫວ່າງມາດຕະການການກະທໍາທາງເພດກັບຍິງ 1 ແລະຍິງ 2 ໄດ້ຖືກນໍາໃຊ້ t ການທົດສອບດ້ວຍການແກ້ໄຂ Bonferroni. ເຖິງຢ່າງໃດກໍ່ຕາມ, ຄວາມສາມາດທາງເພດທີ່ມີເພດຍິງ 2 ມີຄວາມເຂັ້ມແຂງຫນ້ອຍ, ຕາມການສັງເກດເຫັນວ່າມີຈໍານວນຫນ້ອຍທີ່ຈະອອກຈາລະຈອນ (ຫມາຍຄວາມວ່າ, 1 ກັບ 2; F = 49.86p <0.01) ແລະການບຸກລຸກ (ຫມາຍຄວາມວ່າ = 11.2 vs 37.0;F = 20.17 p <0.05) ໃນຊົ່ວໂມງ ທຳ ອິດ. ຕົວເລກຂອງພູເຂົາໃນຊົ່ວໂມງ ທຳ ອິດທີ່ມີເພດຍິງ 1 ແລະ 2 ບໍ່ແຕກຕ່າງກັນຫຼາຍ.
ມັນແມ່ນສິ່ງສໍາຄັນທີ່ຄວນສັງເກດວ່າແມ່ຍິງທີ່ໃຊ້ໃນຊ່ວງທົດລອງຂອງການທົດລອງ (ເຊົ່ນແມ່ຍິງ 1) ຍັງສະແດງທ່າທີທີ່ມີປະໂຫຍດ (ເຊັ່ນ: ການເລັບແລະກ້າມ) ແລະການຕອບສະຫນອງ (ເຊັ່ນ, lordosis) ສໍາລັບໄລຍະເວລາຂອງການສໍາຜັດກັບຊາຍ ທີ່ຢູ່
Neurochemistry
ຄວາມເຂັ້ມຂົ້ນ nanomolar Basal ຂອງ DA ແລະການປ່ຽນແປງຂອງມັນໃນ microdialysates, ສະເຫນີເປັນຄວາມຫມາຍ± SEM, ຂອງສາມຕົວຢ່າງເບື້ອງຕົ້ນທໍາອິດແມ່ນ: DA, 3.0 ± 0.7; DOPAC, 619.1 77.7 ແລະ HVA, 234.2 49.0 (uncorrected for probe recoveryn = 5) ຄ່າເຫຼົ່ານີ້ເປັນຕົວແທນ 100% ຄະແນນຖານ.
ຈຸດຂໍ້ມູນທີ່ຖືກກໍານົດຕາມພຶດຕິກໍາທີ່ສອດຄ້ອງກັນກັບແຕ່ລະຂັ້ນຕອນຂອງການທົດລອງແລະທົ່ວໄປກັບຫນູທຸກ, ໄດ້ຖືກນໍາໃຊ້ສໍາລັບການວິເຄາະທາງ neurochemical. ຕົວຢ່າງເຫຼົ່ານີ້ລວມມີຂໍ້ມູນດັ່ງຕໍ່ໄປນີ້: (1) ຕົວຢ່າງເຈັດຫຼັງຈາກການແນະນໍາຄັ້ງທໍາອິດຂອງແມ່ຍິງ 1, (2) ສີ່ຕົວຢ່າງທີ່ສົ່ງມາພ້ອມກັບບໍ່ມີການປະພຶດຮ່ວມກັບແມ່ຍິງ 1 ແລະ (3) ຫ້າຕົວຢ່າງຫຼັງຈາກການນໍາສະເຫນີຂອງຍິງ 2. ຮູບພາບ 1 ສະແດງໃຫ້ເຫັນການປ່ຽນແປງໃນຄວາມເຂັ້ມຂົ້ນຂອງ DA (ເສັ້ນສະແດງເສັ້ນ, ກາງ) ແລະ metabolites DA (ແຖວເສັ້ນ, top) ກົງກັນຂ້າມກັບພຶດຕິກໍາການປຽບທຽບ (bar graph, ທາງລຸ່ມ) ໃນລະຫວ່າງການທົດສອບສໍາລັບຜົນກະທົບ Coolidge.
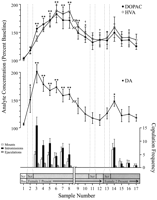
ເບິ່ງເວີຊັນຂະຫນາດໃຫຍ່:
Fig 1
Nucleus accumbens correlates neurochemicals ຂອງພຶດຕິກໍາທາງເພດໃນໄລຍະຜົນກະທົບ Coolidge. ຕົວຢ່າງແປດຕົວທໍາອິດສະແດງໃຫ້ເຫັນຈຸດຂໍ້ມູນຢ່າງຕໍ່ເນື່ອງຂອງໄລຍະເວລາຈາກຂັ້ນ 1 ຫາ 3. ຕົວຢ່າງ 1 ແມ່ນຕົວຢ່າງພື້ນຖານທີ່ສີ່ແລະສຸດທ້າຍທີ່ກໍານົດໄວ້ກ່ອນຫນ້ານີ້ (ເນເທີແລນ) ຕົວຢ່າງ 2 ສະແດງການແນະນໍາຂອງຍິງ 1 ຫລັງຫນ້າຈໍ (Scr) ຫຼັງຈາກທີ່ 15 ນາທີ, ຫນ້າຈໍໄດ້ຖືກລຶບອອກ, ແລະຫນູໄດ້ຖືກອະນຸຍາດໃຫ້ເຮັດສໍາເນົາ (ຕົວຢ່າງ 3-8). Theພັກຜ່ອນ ກ່ຽວກັບ x-axis ເທົ່າກັບການຍົກເວັ້ນຂອງຂໍ້ມູນຈາກສາມຫນູທີ່ໄດ້ຮັບການສະສົມສໍາລັບໄລຍະເວລາທີ່ມີເພດຍິງໃນເບື້ອງຕົ້ນ. ເກົ້າຕົວຢ່າງສຸດທ້າຍແມ່ນຍັງສືບຕໍ່ໃນໄລຍະເວລາ. ຕົວຢ່າງ 9 ແລະ 10 ສອດຄ້ອງກັບໄລຍະເວລາທີ່ satiation ຂອງ phase 3 (ie, 30 min ໂດຍບໍ່ມີການ mount). ຍິງ 1 ຖືກ reinserted ຫລັງຫນ້າຈໍ (ຕົວຢ່າງ 11) ແລະ, 15 min ຕໍ່ມາ, ຫນ້າຈໍໄດ້ຖືກລຶບອອກ (ຕົວຢ່າງ 12). ຫຼັງຈາກ 15 ນາທີບໍ່ມີການຮ່ວມເພດ, ຍິງ 2 ຖືກຢູ່ຫລັງຈໍ (ຕົວຢ່າງ 13). ຕົວຢ່າງ 14-17 ແມ່ນສອດຄ່ອງກັບແມ່ຍິງ 2. ຕົວເລກຂອງການຕິດຕັ້ງ, ການຂັດຂວາງຫຼືການກະຕຸ້ນເຕົ້ານົມທີ່ກ່ຽວຂ້ອງກັບຕົວຢ່າງ microdialysis ທຸກໆ 15 min ແມ່ນສະແດງຢູ່ໃນ bottom bar graphທີ່ຢູ່ ຂໍ້ມູນທາງກາຍະພາບຈະຖືກສະແດງອອກຕາມອັດຕາຮ້ອຍລະຂອງຄວາມເຂັ້ມຂຸ້ນຂອງເສັ້ນສະເລ່ຍ. ການປ່ຽນແປງໃນ NAC DA (ປິດຮຽບຮ້ອຍ), DOPAC (ປິດວົງເລັບ) ແລະ HVA (ເປີດວົງ) efflux ແມ່ນນໍາສະເຫນີເປັນ line graphsທີ່ຢູ່ ການປຽບທຽບດັ່ງຕໍ່ໄປນີ້ໄດ້ຖືກເຮັດໃຫ້: ຕົວຢ່າງຕົ້ນ 1 ທຽບກັບຕົວຢ່າງ 2-10; ຕົວຢ່າງພື້ນຖານໃຫມ່ 10 ທຽບກັບຕົວຢ່າງ 11 ແລະ 12; ຕົວຢ່າງພື້ນຖານໃຫມ່ 12 ທຽບກັບຕົວຢ່າງ 13-17 (*p <0.05; ** p <0.01). ເອກະລາດ t ການທົດສອບໄດ້ຖືກເຮັດໃຫ້ຢູ່ລະຫວ່າງມູນຄ່າພື້ນຖານ (ຕົວຢ່າງ 1, 10, ແລະ 12). ສໍາລັບຄວາມແຕກຕ່າງທີ່ສໍາຄັນຈາກເສັ້ນທໍາອິດ (ຕົວຢ່າງ 1), †p <0.05.
ແຍກ ANOVAs ໄດ້ຖືກປະຕິບັດກ່ຽວກັບຂໍ້ມູນກ່ຽວກັບທາງ neurochemical ທີ່ກ່ຽວຂ້ອງກັບແມ່ຍິງ 1 (ຕົວຢ່າງ 1-12) ແລະແມ່ຍິງ 2 (ຕົວຢ່າງ 12-17). A priori ການປຽບທຽບໄດ້ຖືກນໍາໃຊ້ການທົດສອບການປຽບທຽບຫຼາຍຂອງ Dunn (Bonferroni t) ການປຽບທຽບຕົ້ນຕໍດັ່ງລຸ່ມນີ້: (1) ຈຸດເລີ່ມຕົ້ນ (ຕົວຢ່າງ 1) ທຽບກັບຕົວຢ່າງ 2-10 (XRLX ຄັ້ງທໍາອິດກັບແມ່ຍິງ 1), (2) ຖານສອງ (ຕົວຢ່າງ 10) ທຽບກັບຕົວຢ່າງ 11 ແລະ 12 (ການເປີດເຜີຍຕໍ່ເພດຍິງ 1) , ແລະ (3) ເສັ້ນທີສາມ (ຕົວຢ່າງ 12) ທຽບກັບຕົວຢ່າງ 13-17 (ການຊູນກັບແມ່ຍິງ 2).
ມີການປ່ຽນແປງຢ່າງມີໄນສໍາຄັນໃນ DA efflux ໃນການຕອບສະຫນອງກັບແມ່ຍິງ 1 [F (11,44) = 8.48 p <0.001] ແລະຍິງ 2 [F (5,20) = 2.83p <0.05]. ການເພີ່ມຂື້ນຢ່າງຫຼວງຫຼາຍໃນ DA efflux ໄດ້ຖືກພົບເຫັນໃນເວລາທີ່ແມ່ຍິງ 1 ມີຢູ່ຫລັງຫນ້າຈໍ (+ 44%,p <0.05; ຕົວຢ່າງ 2). ໃນລະຫວ່າງການລະບາຍ, ຄວາມເຂັ້ມຂົ້ນຂອງ DA ເພີ່ມຂື້ນຕື່ມອີກ, ບັນລຸມູນຄ່າສູງສຸດ (+ 95%;p <0.01) ໃນລະຫວ່າງການແຂ່ງຂັນກັບການລະເມີດລິຂະສິດຄັ້ງ ທຳ ອິດ (ຕົວຢ່າງ 3). DA ຍັງຄົງສູງຂື້ນຕະຫຼອດການຮັບມືແລະພຽງແຕ່ກັບຄືນສູ່ຄວາມເຂັ້ມຂົ້ນຂອງພື້ນຖານໃນໄລຍະເວລາ 30 ນາທີເຊິ່ງບໍ່ມີການຕິດຕັ້ງຂຶ້ນ (ຕົວຢ່າງ 9 ແລະ 10). ທັງການເອົາຄືນ ໃໝ່ ຂອງເພດຍິງ 1 ຢູ່ທາງຫລັງຂອງ ໜ້າ ຈໍ (ຕົວຢ່າງ 11) ແລະບໍ່ມີໂອກາດທີ່ຈະພົວພັນກັບຮ່າງກາຍ, ແຕ່ໂດຍບໍ່ມີການເພີ່ມ (ຕົວຢ່າງ 12), ຄວາມເຂັ້ມຂົ້ນຂອງ DA ສູງຂື້ນກັບມູນຄ່າພື້ນຖານທີສອງ (ຕົວຢ່າງ 10). ການປະກົດຕົວຂອງຜູ້ຍິງ 2 ຢູ່ທາງຫລັງຂອງ ໜ້າ ຈໍ (ຕົວຢ່າງ 13) ສົ່ງຜົນໃຫ້ການຂະຫຍາຍຕົວຂອງ DA efflux (12%) ເລັກນ້ອຍຈາກມູນຄ່າພື້ນຖານທີສາມ (ຕົວຢ່າງ 12) ທີ່ບໍ່ສາມາດບັນລຸຄວາມ ສຳ ຄັນທາງສະຖິຕິ. ການປັບ ໃໝ່ ກັບແມ່ຍິງ 2 ເຮັດໃຫ້ມີການເພີ່ມຂື້ນຢ່າງຫຼວງຫຼາຍ (34%) (p <0.05) ໃນ DA efflux ໃນໄລຍະຕົວຢ່າງການລະຫັດຄັ້ງ ທຳ ອິດ (ຕົວຢ່າງ 14). ເຖິງແມ່ນວ່າການປະພຶດທີ່ອ່ອນແອຍັງສືບຕໍ່ໃນສາມຕົວຢ່າງຕໍ່ໄປ, ຄວາມເຂັ້ມຂົ້ນຂອງ DA ໄດ້ຫຼຸດລົງເຖິງຄ່າພື້ນຖານ (ຕົວຢ່າງ 15–17). ເອກະລາດ t ການທົດສອບໄດ້ດໍາເນີນຢູ່ໃນຕົວຢ່າງ "ຕົວຖານ" (ເຊົ່ນ 1, 10, ແລະ 12) ສະແດງໃຫ້ເຫັນວ່າຄ່າເຫຼົ່ານີ້ບໍ່ແຕກຕ່າງກັນຢ່າງຫຼວງຫຼາຍ.
ໃນສາມຫນູທີ່ສືບຕໍ່ການຮ່ວມເພດໃນເວລາຍິງ 1 ໄດ້ຖືກນໍາໃຊ້ໃຫມ່, ຄວາມເຂັ້ມຂົ້ນຂອງ NAC DA ເພີ່ມຂື້ນເມື່ອແມ່ຍິງ 1 ຢູ່ເບື້ອງຫລັງຫນ້າຈໍ (ຊ່ວງ 25-47%) ແລະໃນໄລຍະການປອງກັນ (ຊ່ວງ 13-37%), ທຽບກັບຕົວຢ່າງ ກ່ອນທີ່ຈະກັບຄືນມາຂອງຍິງ. ການເພີ່ມຂຶ້ນເຫຼົ່ານີ້, ແຕ່, ເກີດຂຶ້ນພຽງແຕ່ໃນເວລາທີ່ການກະທໍາທາງເພດມີຄວາມເຂັ້ມແຂງແລະເຮັດໃຫ້ມີການກະຈາຍສຽງ.
ການປ່ຽນແປງທີ່ສໍາຄັນໃນ DOPAC [F (11,44) = 9.57 p <0.001] ແລະ HVA [F (11,44) = 12.47 p <0. 001] ຄວາມເຂັ້ມຂຸ້ນໄດ້ຖືກພົບເຫັນໃນການຕອບສະ ໜອງ ຕໍ່ເພດຍິງ 1, ແຕ່ວ່າບໍ່ແມ່ນເພດຍິງ 2. ຄວາມເຂັ້ມຂົ້ນຂອງເມທາໂບລິນໄດ້ເພີ່ມຂຶ້ນເລັກນ້ອຍ (+ 15% ໃນທັງສອງກໍລະນີ) ໃນລະຫວ່າງການ ນຳ ສະ ເໜີ ແມ່ຍິງ 1 ຢູ່ເບື້ອງຫຼັງ ໜ້າ ຈໍ (ຕົວຢ່າງ 2), ແຕ່ວ່ານີ້ບໍ່ແມ່ນຄວາມ ສຳ ຄັນ ສະຖິຕິ. ເຖິງຢ່າງໃດກໍ່ຕາມ, ການເພີ່ມຂື້ນຢ່າງຫຼວງຫຼາຍໃນຄວາມເຂັ້ມຂົ້ນຂອງ DOPAC ແລະ HVA ໃນໄລຍະການລອກ (ຕົວຢ່າງ 3–8), ບັນລຸມູນຄ່າສູງສຸດ (+80 ແລະ + 86%, ຕາມ ລຳ ດັບ; p <0.01) ຫຼັງຈາກ 60 ນາທີ (ຕົວຢ່າງ 6 ໃນທັງສອງກໍລະນີ). ເຖິງແມ່ນວ່າຄວາມເຂັ້ມຂົ້ນຂອງ metabolite ຫຼຸດລົງໃນຊ່ວງໄລຍະຂອງການບໍ່ມີປະສິດຕິພາບທາງເພດໃນຕອນທ້າຍຂອງການພົວພັນກັບເພດຍິງ 1 (ຕົວຢ່າງ 9 ແລະ 10), ຄວາມເຂັ້ມຂົ້ນຂອງມັນຍັງຄົງສູງຂື້ນກ່ຽວກັບຂໍ້ມູນເບື້ອງຕົ້ນ (p <0.05 ໃນທັງສອງກໍລະນີ). ການອອກ ກຳ ລັງກາຍຂອງເພດຍິງ 1 ຢູ່ທາງຫລັງຂອງ ໜ້າ ຈໍ (ຕົວຢ່າງ 11), ການເຂົ້າເຖິງຜູ້ຍິງ 1 ຫຼັງຈາກຖອດ ໜ້າ ຈໍ (ຕົວຢ່າງ 12), ແລະການແນະ ນຳ ເພດຍິງ 2 (ຕົວຢ່າງ 13) ບໍ່ມີຜົນໃນການປ່ຽນແປງຂອງຄວາມເຂັ້ມຂົ້ນຂອງທາດແປ້ງ. ເລັກນ້ອຍ, ແຕ່ບໍ່ມີຂໍ້ມູນທາງສະຖິຕິ, ການເພີ່ມຂື້ນຂອງຄວາມເຂັ້ມຂົ້ນຂອງ DOPAC ແລະ HVA (+ 23% ໃນທັງສອງກໍລະນີ) ທີ່ກ່ຽວຂ້ອງກັບພື້ນຖານ (ຕົວຢ່າງ 12) ກົງກັບການແຂ່ງຂັນຄັ້ງ ທຳ ອິດກັບແມ່ຍິງ 2 (ຕົວຢ່າງ 14). ເຖິງຢ່າງໃດກໍ່ຕາມ, ການເພີ່ມຂື້ນນີ້ແມ່ນມີອາຍຸສັ້ນ, ແລະປະຕິເສດຄຸນຄ່າພື້ນຖານ ສຳ ລັບສາມຕົວຢ່າງທີ່ຍັງເຫຼືອ (15-17). ເອກະລາດ t(1, 10, ແລະ 12) ສະແດງໃຫ້ເຫັນວ່າຄ່າຕົວຊີ້ວັດທີສອງແລະທີສາມ (ຕົວຢ່າງ 10 ແລະ 12 ຕາມລໍາດັບ) ແຕ່ບໍ່ແຕກຕ່າງຈາກແຕ່ລະອື່ນໆແມ່ນສູງຂຶ້ນເມື່ອທຽບໃສ່ກັບຕົວຢ່າງເບື້ອງຕົ້ນຄັ້ງທໍາອິດ ສໍາລັບ DOPAC ແລະ HVA (p <0.05 ໃນທັງສອງກໍລະນີ).
Histology
ການສືບສວນ Microdialysis ຢູ່ໃນ NAC (Fig.2) ໃນລະດັບການຂະຫຍາຍ + 1.20 ຫາ + 1.70 ມມຈາກ bregma (ກະຮອກກະຮອກ). ມີການປ່ຽນແປງຍັງຢູ່ໃນຍົນ mediolateral; ຂໍ້ມູນສະທ້ອນໃຫ້ເຫັນຕົວຢ່າງຈາກກຸ່ມຫອຍແລະສໍາຄັນຂອງ NAC.
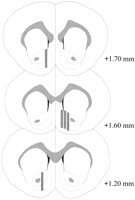
ເບິ່ງເວີຊັນຂະຫນາດໃຫຍ່:
Fig 2
ສະຖານທີ່ຂອງ probes microdialysis ພາຍໃນ NAC ຂອງຫນູເພດທີ່ນໍາໃຊ້ໃນການທົດລອງຜົນກະທົບ Coolidge. ຮູບສີ່ແຈສາກ ສອດຄ້ອງກັບພື້ນທີ່ເຍື່ອຫຸ້ມຂອງການກວດສອບ microdialysis. ພາກສ່ວນຂອງສະຫມອງຂອງຄໍລີນໄດ້ຖືກລວບລວມຈາກPaxinos ແລະ Watson (1986).
ການສົນທະນາ
ໃນການຕົກລົງກັບບົດລາຍງານໃນເບື້ອງຕົ້ນ, ຜົນໄດ້ຮັບໃນປັດຈຸບັນສະແດງໃຫ້ເຫັນເຖິງການສົ່ງເສີມ mesolimbic DA ທີ່ກ່ຽວຂ້ອງກັບອົງປະກອບທີ່ມັກກິນແລະປະສົມປະສານຂອງພຶດຕິກໍາທາງເພດຂອງຫນູທີ່ຖືກປະເມີນໂດຍin vivo microdialysis (Mas et al, 1990; Pfaus et al, 1990;Pleim et al, 1990; Damsma et al, 1992; Wenkstern et al, 1993; Fumero et al, 1994; Mas et al, 1995a,b,d) ນອກຈາກນັ້ນ, ຜົນໄດ້ຮັບເຫຼົ່ານີ້ສະຫນອງການພົວພັນທາງດ້ານ neurochemical ສໍາລັບ satiation ທາງເພດແລະການ reinitiation ຕໍ່ມາຂອງ copulation ໃນການຕອບສະຫນອງກັບແມ່ຍິງທີ່ໄດ້ຮັບການ Novell (ຜົນກະທົບ Coolidge). ຂໍ້ມູນໃນປະຈຸບັນຊີ້ໃຫ້ເຫັນວ່າຄຸນລັກສະນະກະຕຸ້ນຂອງແມ່ຍິງທີ່ໄດ້ຮັບຄວາມນິຍົມໃຫມ່ອາດຈະເຮັດໃຫ້ເກີດການຕິດເຊື້ອ NAC DA ໃນຫນູຜູ້ທີ່ມີຄວາມສໍາຄັນທາງເພດທີ່ອາດຈະກ່ຽວຂ້ອງກັບການເກີດໃຫມ່ຂອງພຶດຕິກໍາທາງເພດ. ນີ້ແມ່ນເຫັນໄດ້ຊັດເຈນໃນການເພີ່ມຂຶ້ນເລັກນ້ອຍໃນ NAC DA ໃນໄລຍະການນໍາສະເຫນີຂອງແມ່ຍິງປະດິດສ້າງຢູ່ຫລັງຈໍແລະພົບເຫັນຫຼາຍທີ່ສຸດເປັນການເພີ່ມຂື້ນຢ່າງຊັດເຈນໃນລະຫວ່າງການປ້ອນໃຫມ່ກັບແມ່ຍິງ 2 (Fig.1).
ການມີແມ່ຍິງທີ່ໄດ້ຮັບການຕອບສະຫນອງຄັ້ງທໍາອິດຫລັງຈາກຫນ້າຈໍໄດ້ເຮັດໃຫ້ການເພີ່ມຂຶ້ນຢ່າງຫນ້າພໍໃຈໃນ NAC DA efflux (44% ຈາກພື້ນຖານ) ທີ່ຄ້າຍຄືກັນກັບສິ່ງທີ່ໄດ້ຖືກລາຍງານໃນການທົດລອງຜ່ານມາໂດຍໃຊ້ການອອກແບບທີ່ຄ້າຍຄືກັນ (30% Pfaus et al, 19901 35%,Damsma et al, 1992). ໃນຂໍ້ຕົກລົງກັບການສຶກສາເຫຼົ່ານີ້ແມ່ນການສັງເກດວ່າ NAC DA efflux ໄດ້ຮັບການປັບປຸງຕື່ມອີກໃນໄລຍະການປະຕິບັດ ((>> 95% ສູງກວ່າພື້ນຖານໃນການທົດລອງໃນປະຈຸບັນ)). ເຖິງແມ່ນວ່າພວກເຮົາສາມາດເບິ່ງພຶດຕິ ກຳ ທີ່ມີປະສິດຕິພາບສູງທີ່ກ່ຽວຂ້ອງກັບການປ່ອຍ NAC DA ທີ່ຖືກປັບປຸງ (Wenkstern et al, 1993; Wilson et al, 1995), ມັນເປັນສິ່ງສໍາຄັນທີ່ຈະກວດເບິ່ງຄໍາວ່າ "ຄວາມຫນ້າພໍໃຈ" ແລະ "ການບໍລິໂພກ" ໃນສະພາບການທາງເພດ. ໃນຂະນະທີ່ຂັ້ນຕອນທີ່ແມ່ຍິງມີຢູ່ຫລັງຈໍແມ່ນເສີຍໆຫຼືກຽມພ້ອມ, ພຶດຕິກໍາໃນຊ່ວງໄລຍະການຮ່ວມເພດບໍ່ສາມາດພິຈາລະນາໄດ້ຢ່າງສົມບູນ. ເນື່ອງຈາກວ່າ "ທີ່ຫນ້າປະຫລາດໃຈ" ສາມາດນໍາໃຊ້ເພື່ອອະທິບາຍເຖິງພຶດຕິກໍາທັງຫມົດທີ່ນໍາໄປສູ່ການປະຕິບັດຕົວຈິງຂອງພຶດຕິກໍາທີ່ກະຕຸ້ນ (ການຮ່ວມເພດ), ການປະພຶດທໍາອິດຂອງຜູ້ຊາຍທີ່ສະແດງໃນຂະນະທີ່ມີການເຄື່ອນໄຫວໃນໄລຍະ "consummatory" ຜູ້ຊາຍໃຊ້ເວລາຫຼາຍທີ່ສຸດແລະຄວາມພະຍາຍາມຂອງແມ່ຍິງທີ່ຈະຫຼອກລວງ. ໃນເລື່ອງນີ້, ພວກເຮົາສາມາດພົວພັນກັບການສົ່ງຜ່ານ NAC DA ສູງສຸດທີ່ມີການບໍລິໂພກ ເຊັ່ນດຽວກັນກັບ ອົງປະກອບທີ່ມີຄວາມຊື່ນຊົມຫຼາຍໃນການປະພຶດຕົວທາງເພດຂອງຜູ້ຊາຍ.
ການເຂົ້າເຖິງແມ່ຍິງທີສອງ, ໃຫມ່ໄດ້ເຮັດໃຫ້ມີການປ່ຽນແປງໃຫມ່ໃນທຸກໆເລື່ອງ. ການສຶກສາກ່ອນຫນ້ານີ້ໄດ້ສະແດງໃຫ້ເຫັນວ່າສ່ວນໃຫຍ່ຂອງຫນູໄດ້ຮັບອະນຸຍາດໃຫ້ນໍາມາຜະສົມກັບຄວາມໂປດປານ, ໂດຍນໍາໃຊ້ໂປໂຕຄອນປະຕິບັດທີ່ຄ້າຍຄືກັນກັບຄົນທີ່ນໍາໃຊ້ໃນການທົດລອງໃນປະຈຸບັນ, ບໍ່ໄດ້ສືບຕໍ່ການຫາຄູ່ເມື່ອທົດລອງ 24 ຊົ່ວໂມງຕໍ່ມາ (ຫາດຊາຍແລະຈໍແດນ, 1956) ມັນອາດຈະເປັນວ່າການມີຄຸນສົມບັດກະຕຸ້ນຂອງ Novell ຂອງແມ່ຍິງ 2, ເຊິ່ງອາດຈະປະກອບມີຄໍາແນະນໍາ olfactory ແລະສາຍຕາແລະສຽງ, ເຮັດໃຫ້ການຮ່ວມມືໃຫມ່. ຄໍາຖາມທີ່ຫນ້າສົນໃຈ, ເຊິ່ງຍັງມີຄໍາຕອບ, ແມ່ນກົນໄກຂອງຫນູຊາຍທີ່ແຕກຕ່າງຈາກເພດຍິງທີ່ມີເພດສໍາພັນທີ່ລາວໄດ້ມາເມື່ອໄວໆມານີ້. ເວັບໄຊສໍາລັບກົນໄກນັ້ນອາດຈະຢູ່ໃນລະບົບ olfactory ຕົ້ນຕໍ. ມັນໄດ້ຖືກລາຍງານວ່າຄວາມສົມບູນຂອງລະບົບນີ້ແມ່ນສໍາຄັນຕໍ່ຜົນກະທົບຂອງ Coolidge ໃນ hamsters (Johnston ແລະ Rasmussen, 1984) ລະບົບ olfactory vomeronasal -operative, ຢ່າງໃດກໍຕາມ, ໃນຂະບວນການຄວາມຊົງຈໍາ pheromonal ໄດ້ຖືກອະທິບາຍເມື່ອໄວໆນີ້ຢູ່ໃນຫນູ (Kaba et al, 1994), ຍັງເປັນຜູ້ສະຫມັກທີ່ສໍາຄັນ. ໃນເລື່ອງນີ້, ມັນຄວນຈະໃຫ້ມີການເພີ່ມຂື້ນໃນການສົ່ງຜ່ານ NAC DA ໂດຍນໍາໃຊ້ in vivo voltammetry ໃນຫນູຂອງຜູ້ຊາຍທີ່ນໍາສະເຫນີທີ່ມີຜ້າຝ້າຍທີ່ໄດ້ຮັບການຊູນກັບຫນູເພດໃນ estrus (Louillot et al, 1991; Mitchell and Gratton, 1992) ຍິ່ງໄປກວ່ານັ້ນ, ການນໍາໃຊ້ຂອງ K ໄດ້+ ໂດຍກົງກັບຊັ້ນຂອງເສັ້ນປະສາດຂອງ vomeronasal ຂອງ bulb olfactory ອຸປະກອນເສີມ, ເຊັ່ນດຽວກັນກັບ bulb olfactory ປະກອບດ້ວຍຕົນເອງ, ແມ່ນພຽງພໍທີ່ຈະເພີ່ມທະວີການສົ່ງຜ່ານ NAC DA (Mitchell and Gratton, 1992).
ການແຂ່ງຂັນ 15 ຄັ້ງທໍາອິດຂອງການຮ່ວມເພດກັບຍິງ 2 ແມ່ນກ່ຽວຂ້ອງກັບການເພີ່ມຂຶ້ນຢ່າງຫຼວງຫຼາຍໃນ NAC DA. ໃນທາງກົງກັນຂ້າມກັບແມ່ຍິງ 1, ການພົວພັນກັບແມ່ຍິງ 2 ບໍ່ໄດ້ຜະລິດເພີ່ມຂຶ້ນໃນ NAC DA ຂອງຂະຫນາດດຽວກັນໃນລະຫວ່າງໄລຍະເວລາພໍສົມຄວນ (12%) ຫຼືປະສົມປະສານ (34%). ການເພີ່ມຂຶ້ນເລັກນ້ອຍເຫຼົ່ານີ້ໃນ NAC DA, ຢ່າງໃດກໍ່ຕາມ, ມີຄວາມສອດຄ່ອງກັບລະດັບການຫຼຸດລົງຂອງພຶດຕິກໍາທາງເພດທີ່ສະແດງດ້ວຍແມ່ຍິງ 2 ເມື່ອທຽບໃສ່ຍິງ 1. ຄວາມເຂັ້ມຂຸ້ນຂອງ Metabolite ຍັງຄົງສູງໃນໄລຍະການ satiety, ເຊິ່ງເຮັດໃຫ້ຄວາມເຂັ້ມຂົ້ນຂອງເສັ້ນສະແດງໃຫມ່ (ຕົວຢ່າງ 10 ແລະ 12) ສູງຂຶ້ນຈາກມູນຄ່າພື້ນຖານເບື້ອງຕົ້ນ (ຕົວຢ່າງ 1).
ການຊັກຊ້າຊົ່ວຄາວໃນການເພີ່ມຂື້ນໃນລະດັບ DOPAC ແລະ HVA ໃນໄລຍະການຮ່ວມກັນແມ່ນສອດຄ່ອງກັບການສ້າງຂອງພວກເຂົາເປັນການປ່ຽນແປງຂອງທາດປະສົມຂອງພໍ່ແມ່ DA. ມັນໄດ້ຖືກແນະນໍາວ່າລະດັບຄວາມເຂັ້ມຂົ້ນຂອງການຍ່ອຍສະຫຼາຍຂອງຈຸລິນຊີ (microdialysis metabolite), ຢ່າງຫນ້ອຍໃນລະຫວ່າງການປະພຶດທໍາມະຊາດທີ່ບໍ່ໄດ້ຮັບຜົນກະທົບທາງດ້ານ pharmacologically,Damsma et al, 1992; Fumero et al, 1994) ຄວາມຈິງທີ່ວ່າຄວາມເຂັ້ມຂຸ້ນຂອງ metabolite ຍັງສູງເຖິງແມ່ນວ່າໃນໄລຍະເວລາທີ່ບໍ່ມີເພດສໍາພັນໃນການທົດລອງນີ້, ໃນເວລາທີ່ຄວາມເຂັ້ມຂົ້ນຂອງ DA ໄດ້ກັບຄືນມາເປັນຄ່າທໍານຽມທົດລອງ, ມັນກໍ່ເປັນຄວາມສົງໃສໃນຄໍາແນະນໍານີ້.
ການເພີ່ມຂື້ນຂອງຄວາມເຂັ້ມຂຸ້ນຂອງ metabolite DA ທີ່ເຫັນໄດ້ໃນການທົດລອງນີ້ສະທ້ອນໃຫ້ເຫັນເຖິງສະພາບຂອງພື້ນທີ່ທີ່ມີຄວາມກ້າວຫນ້າທາງດ້ານຮ່າງກາຍ (mPOA) ຂອງ DA metabolites ສັງເກດເຫັນໃນຫນູໃນມື້ທໍາອິດຫຼັງຈາກພວກມັນໄດ້ປະສົມກັບຄວາມເຄັ່ງຄັດ (Mas et al, 1995a,b) ຄວາມສູງຂອງຄວາມເຂັ້ມຂົ້ນຂອງ DOPAC ແລະ HVA ໃນ NAC ຫຼື mPOA ແມ່ນບໍ່ສັງເກດເຫັນສະເຫມີໃນເວລາທີ່ການຈັບຄູ່ມີໄລຍະເວລາຄົງທີ່, ຫຼາຍກ່ວາທີ່ໃຊ້ເວລາທີ່ຈະຕ້ອງໄດ້ຮັບຄວາມເຄົາລົບ. ຕົວຢ່າງ, ຫລາຍໆການສຶກສາໄດ້ສະແດງໃຫ້ເຫັນວ່າຄວາມເຂັ້ມຂຸ້ນຂອງ DOPAC ໄດ້ເພີ່ມຂຶ້ນແລະຍັງສູງໃນລະຫວ່າງການຮ່ວມກັນແຕ່ໄດ້ຫຼຸດລົງໃນມູນຄ່າເບື້ອງຕົ້ນບໍ່ດົນຫຼັງຈາກແມ່ຍິງຖືກລົບອອກ (Pfaus et al, 1990; Pleim et al, 1990; Damsma et al, 1992;Hull et al, 1993; Wenkstern et al, 1993; Hull et al, 1995) ໃນການສຶກສາໂດຍ Mas et al (1995b), ຄວາມເຂັ້ມຂົ້ນ extracellular ຂອງ DOPAC ແລະ HVA ໃນ mPOA ຍັງຄົງສູງກວ່າ 4 ມື້ຕິດຕໍ່ກັນເຊິ່ງສອດຄ່ອງກັບໄລຍະເວລາຂອງການບໍ່ມີເພດສໍາພັນ. ໃນມື້ທີ່ສີ່, ກ່ອນທີ່ສັດຈະເລີ້ມການຮ່ວມ, ຄວາມເຂັ້ມຂົ້ນຂອງ metabolites ແມ່ນຢູ່ໃກ້ກັບຄ່າຂອງການຮັກສາ. ຜູ້ຂຽນປຽບທຽບຮູບແບບຂອງການປ່ຽນແປງທາງ neurochemical ກັບຜູ້ທີ່ໄດ້ເຫັນຫຼັງຈາກການບໍລິຫານຂອງ DA receptor blockers (Zetterstrm et al, 1984; Imperato and DiChiara, 1985) ແລະໄດ້ແນະນໍາວ່າສະຖານະຂອງການບໍ່ມີເພດສໍາພັນທາງເພດອາດຈະໄດ້ຮັບການໄກ່ເກ່ຍໂດຍຜ່ານການປ່ອຍ prolactin ຊຶ່ງອາດຈະເປັນ "endogenous neuroleptic" (Mas et al, 1995a,b,d) ມັນເປັນທີ່ຊັດເຈນວ່າການບໍລິຫານຂອງ neuroleptic ແມ່ນປະກອບດ້ວຍຄວາມເຂັ້ມຂຸ້ນໃນການປ່ຽນແປງຂອງເມຕາໂບລີນ extracellular ແລະ DA efflux (Zetterstrm et al, 1984; Imperato and DiChiara, 1985) ແຕ່ຫນ້າເສຍດາຍ, Mas et al (1995a,b) ບໍ່ສາມາດກວດພົບຄວາມເຂັ້ມຂົ້ນ MPOA DA. ໃນການສຶກສາໃນປະຈຸບັນ, ຄວາມເຂັ້ມຂົ້ນຂອງ DA ໃນ NAC ກັບຄືນໄປສູ່ຄ່າທີ່ກໍານົດໄວ້ກ່ອນ, ໃນຂະນະທີ່ຄວາມເຂັ້ມຂຸ້ນຂອງ DOPAC ແລະ HVA ຍັງສູງ. ຮູບແບບນີ້ແມ່ນບໍ່ສອດຄ່ອງກັບບົດບາດສໍາລັບການປະຕິບັດທາງ neuroleptic endogenous ໃນ NAC ເພື່ອສົ່ງເສີມຄວາມໂປ່ງໃສທາງເພດ.
ເນື່ອງຈາກການມີສ່ວນຮ່ວມຂອງ neurons DA mesolimbic ໃນພຶດຕິກໍາທີ່ກະຕຸ້ນ (Fibiger ແລະ Phillips, 1986; Blackburn et al, 1992; Kalivas et al, 1993; LeMoal, 1995) ແລະຄວາມອ່ອນໄຫວຂອງພວກເຂົາກັບສິ່ງທ້າທາຍດ້ານສິ່ງແວດລ້ອມໃຫມ່ (Fabre et al, 1983; Schultz, 1992; Mirenowicz and Schultz, 1994), ການເພີ່ມຂື້ນຂອງຄວາມເຂັ້ມຂົ້ນ extracellular ຂອງ NAC DA ໃນການຕອບສະຫນອງກັບແມ່ຍິງ Novell ແມ່ນສອດຄ່ອງກັບສົມມຸດຕິຖານວ່າກິດຈະກໍາໃນລະບົບ DA ນີ້ແມ່ນສໍາຄັນສໍາລັບການ reinitiation ຂອງພຶດຕິກໍາທາງເພດສໍາພັນ. ນອກຈາກນັ້ນ, ບົດລາຍງານຂອງການເພີ່ມຂື້ນແລະຄວາມບໍລິໂພກໃນການສົ່ງໄຟຟ້າ DA (Hull et al, 1993, 1995;Mas et al, 1995b; Sato et al, 1995) ແລະກິດຈະກໍາ neuronal (Shimura et al, 1994) ໃນ mPOA ຂອງຫນູຂອງຜູ້ຊາຍໃນໄລຍະພຶດຕິກໍາທາງເພດຊີ້ໃຫ້ເຫັນວ່າໂຄງສ້າງນີ້ອາດຈະປະກອບສ່ວນກັບລັກສະນະການປອງກັນໃຫມ່ຂອງຜົນກະທົບ Coolidge.
ໃນການຮັກສາສຸຂະພາບຂອງລະບົບ DA ໃນ Mesolimbic DA ໃນການກະທໍາທີ່ມີແຮງກະຕຸ້ນ, ມັນກໍ່ໄດ້ຖືກສ້າງຕັ້ງຂື້ນວ່າຄວາມເຂັ້ມຂົ້ນ extracellular ຂອງ DA ແມ່ນສູງຂຶ້ນກ່ອນ, ໃນໄລຍະແລະທັນທີຫຼັງຈາກການກິນອາຫານ, ກັບຄືນກັບຄ່າເລີ່ມຕົ້ນ ~30 min ຕໍ່ (Wilson et al, 1995) ມັນເປັນທີ່ຮູ້ຈັກດີວ່າຄວາມໃຈຮ້າຍທີ່ເກີດຈາກອາຫານແມ່ນຜົນກະທົບຕໍ່ຄຸນສົມບັດຂອງມັນ. ມະນຸດແລະສັດປະຕິເສດອາຫານທີ່ພວກເຂົາກິນເພື່ອຄວາມອີ່ມໃຈແລະກິນເຂົ້າກິນອາຫານອື່ນໆທີ່ບໍ່ໄດ້ກິນ (Rolls, 1986) ນີ້ຈະເຮັດໃຫ້ຄໍາຖາມກ່ຽວກັບວ່າເອກະສານ efflux extracellular ໃນ NAC ຈະຖືກເພີ່ມຂຶ້ນຢ່າງໃດໂດຍການນໍາສະເຫນີປະເພດອາຫານປະເພດໃຫມ່, ແຕ່ບໍ່ແມ່ນອາຫານທີ່ບໍລິໂພກໃນບໍ່ດົນມານີ້ເພື່ອຄວາມໂປ່ງໃສໃນລັກສະນະທີ່ຄ້າຍຄືກັນກັບການລາຍງານໃນການສຶກສາໃນປະຈຸບັນ ແຮງຈູງໃຈທາງເພດ. ຖ້າໄດ້ຮັບການຢືນຢັນ, ຄວາມສໍາພັນທົ່ວໄປນີ້ລະຫວ່າງຄຸນສົມບັດຂອງ sensory ຂອງຜົນຕອບແທນທໍາມະດາ, satiety ແລະ mesolimbic DA ຈະຫມາຍເຖິງບົດບາດສໍາຄັນສໍາລັບລະບົບປະສາດນີ້ໃນລະບຽບການຂອງຂະບວນການແຮງຈູງໃຈ, ການຂັດແຍ້ງທີ່ອາດນໍາໄປສູ່ຄວາມຜິດປົກກະຕິທີ່ຮ້າຍແຮງຂອງການກິນແລະການເຮັດວຽກທາງເພດ ທີ່ຢູ່
ຂໍ້ມູນອ້າງອິງ
- uf
- Beach FA,
- Jordan L
(1956) ການລ່ວງລະເມີດທາງເພດແລະການຟື້ນຕົວໃນຫນູຂອງຜູ້ຊາຍ. QJ Exp Psychol 8: 121-133.
- uf
- Bermant G,
- Lott DF,
- Anderson L
(1966) ລັກສະນະຊົ່ວຄາວຂອງຜົນກະທົບ Coolidge ໃນການປະພຶດຕົວຂອງຫນູຂອງຜູ້ຊາຍ. J Comp Physiol Psychiatry 65: 447-452.
- uf
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) ຫນ້າທີ່ Dopamine ໃນພຶດຕິກໍາທີ່ຫນ້າທຶ່ງແລະປ້ອງກັນ. Prog Neurobiol 39: 247-279.
- uf
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) ພຶດຕິກໍາທາງເພດເພີ່ມຂຶ້ນການສົ່ງຜ່ານ dopamine ໃນແກນກາງແລະ striatum ຂອງຫນູເພດ: ການປຽບທຽບກັບ novelty ແລະ locomotion. Behav Neurosci 106: 181-191.
- uf
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) ກິດຈະກໍາຂອງ neurons ໃນເຂດ tegmental ventral ຂອງ monkey ປະຕິບັດການ. Behav Brain Res 9: 213-235.
- uf
- Fibiger HC,
- Phillips AG
(1986) ລາງວັນ, ແຮງຈູງໃຈ, ຄວາມຮູ້: ຈິດໃຈຂອງລະບົບ dopamine mesotelencephalic. ໃນປື້ມຄູ່ມືຂອງວິທະຍາສາດ: ລະບົບປະສາດ IV, ED FE, Geiger SD (American Physiology Society, Bethesda, MD), pp. 647-675.
- uf
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) ການກະຕຸ້ນເຕັກໂນໂລຢີຂອງສະຖານທີ່ລາງວັນໃນເຂດທີ່ມີຂະຫນາດໃຫຍ່ໃນລະບົບ ventral ເພີ່ມຄວາມເຂັ້ມຂຸ້ນຂອງ dopamine ໃນແກນຂອງຫນູ. Behav Brain Res 55: 131-141.
- uf
- Fisher A
(1962) ຜົນກະທົບຂອງການປ່ຽນແປງກະຕຸ້ນກ່ຽວກັບຄວາມໂປ່ງໃສທາງເພດໃນຫນູຂອງຜູ້ຊາຍ. J Comp Physiol Psychiatry 55: 614-620.
- uf
- Fowler H,
- Whalen RE
(1961) ການປ່ຽນແປງໃນການກະຕຸ້ນໃຈການກະຕຸ້ນແລະພຶດຕິກໍາທາງເພດໃນຫນູຂອງຜູ້ຊາຍ. J Comp Physiol Psychiatry 54: 68-71.
- uf
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) ການປ່ຽນແປງຂອງລາຍຮັບ monoamine ໃນເຂດ forebrain ທີ່ກ່ຽວຂ້ອງກັບພຶດຕິກໍາທາງເພດຊາຍ: ການສຶກສາ microdialysis. Brain Res 662: 233-239.
- uf
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) ລະບົບການເກັບກໍາຂໍ້ມູນທີ່ອີງໃສ່ IBM-PC ສໍາລັບການບັນທຶກພຶດຕິກໍາທາງເພດສໍາລັບສັດແລະສໍາລັບການບັນທຶກເຫດການທົ່ວໄປ. Physiol Behav 44: 825-828.
- uf
- Hull EM,
- Eaton RC,
- Moses J,
- Lorrain DS
(1993) ການຜ່າຕັດເພີ່ມກິດຈະກໍາ dopamine ຢູ່ໃນພື້ນທີ່ preoptic medial ຂອງຫນູຂອງເພດ. Life Sci 52: 935-940.
- uf
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Extracellular dopamine ໃນພື້ນທີ່ preoptic medial: ຜົນກະທົບຕໍ່ການກະຕຸ້ນທາງເພດແລະຄວບຄຸມຮໍໂມນຂອງການຮ່ວມເພດ. J Neurosci 15: 7465-7471.
- uf
- Imperato A,
- DiChiara G
(1985) ການປ່ອຍຕົວ Dopamine ແລະການຍ່ອຍອາຫານໃນຫນູປຸກຫຼັງຈາກການປະຕິບັດທາງດ້ານວິທະຍາສາດແບບ neuroleptics ຕາມການສຶກສາໂດຍການເປັນ dialysis trans-striatal. J Neurosci 5: 297-306.
- uf
- Johnston RE,
- Rasmussen K
(1984) ການຮັບຮູ້ຕົວຂອງ hamsters ຍິງໂດຍຊາຍ: ບົດບາດຂອງເຄມີສານເຄມີແລະລະບົບ olfactory ແລະ vomeronasal. Physiol Behav 33: 95-104.
- uf
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) ການບົ່ງມະຕິຂອງຄວາມຈໍາເປັນຂອງທາດ olfactory ໂດຍການກະຕຸ້ນຂອງ receptor metabotropic glutamate. ວິທະຍາສາດ 265: 262-264.
- uf
- Kalivas PW,
- Sorg BA,
- Hooks MS
(1993) ເພສັດວິທະຍາແລະວົງຈອນ neural ຂອງ sensitization ກັບ psychostimulants. Behav Pharmacol 4: 315-334.
- uf
- LeMoal M
(1995) Mesocorticolimbic dopaminergic neurons ພາລະບົດບາດແລະກົດລະບຽບ. ໃນ Psychopharmacology: ການຜະລິດສີ່ຂອງຄວາມຄືບຫນ້າ, Eds Bloom FE, Kupfer DJ (Raven, ນິວຢອກ), pp. 283-294.
- uf
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) ການກະຕຸ້ນໃຫ້ມີການຂະຫຍາຍຕົວທາງເລືອກໃນການປົດປ່ອຍ dopamine ໃນກຸ່ມຂອງຫນູເພດ. Brain Res 553: 313-317.
- uf
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) ການເພີ້ມ dopamine ເພີ່ມຂຶ້ນໃນແກນຂອງການຍັບຍັ້ງການເປັນຫນູຂອງຜູ້ຊາຍທີ່ໄດ້ຮັບການພິຈາລະນາໂດຍ volv Neurosci Lett 110: 303-308.
- uf
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Neurochemical ກ່ຽວຂ້ອງຂອງຄວາມປວດລ້າວທາງເພດແລະການຟື້ນຕົວດັ່ງທີ່ໄດ້ຖືກປະເມີນໂດຍ microdialysis in vivo. Brain Res 675: 13-19.
- uf
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) ການຕິດຕາມກວດເລືອດແລະ microdialysis ຂອງການປ່ອຍຈຸລັງ neurotransmitter monoamine ໃນໄລຍະການພົວພັນຮ່ວມເພດສໍາພັນ. Behav Brain Res 71: 69-79.
- uf
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) ການບົ່ງມະຕິຂອງການຈັບຄູ່ໂດຍ apomorphine ໃນຫນູຫນູທາງເພດສໍາພັນ. Eur J Pharmacol 280: 331-334.
- uf
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
(1995d) neurochemistry ຂອງ satiety ທາງເພດ. ຮູບແບບການທົດລອງຂອງຄວາມຕ້ອງການທີ່ຫຍຸ້ງຍາກ. ໃນວິທະຍາສາດຂອງການທໍາງານທາງເພດແລະການຂາດການເຮັດວຽກ, ed Bancroft J (Raven, ນິວຢອກ), pp. 115-126.
- uf
- Mirenowicz J,
- Schultz W
(1994) ຄວາມສໍາຄັນຂອງຄວາມບໍ່ສາມາດຄາດເດົາໄດ້ສໍາລັບການຕອບຮັບຂອງລາງວັນໃນ neuron dopaminergic ສັດຕູ. J Neurophysiol 72: 1024-1027.
- uf
- Mitchell JB,
- Gratton A
(1992) ການປ່ອຍໂມລີເມລິມິກ dopamine ໂດຍການກະຕຸ້ນຂອງລະບົບ olfactory ອຸປະກອນເສີມ: ການຄົ້ນຄວ້າ chronoamperometric ຄວາມໄວສູງ. Neurosci Lett 140: 81-84.
- uf
- Paxinos G,
- Watson C
(1986) ສະຫມອງຫນູໃນປະສານງານ stereotaxic (2nd ed). (Academic, San Diego).
- uf
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) ພຶດຕິກໍາທາງເພດທີ່ເສີມຂະຫຍາຍການສົ່ງຜ່ານ dopamine ກາງໃນຫນູຂອງຜູ້ຊາຍ. Brain Res 530: 345-348.
- uf
- Pfaus JG,
- Everitt BJ
(1995) ການ psychopharmacology ຂອງພຶດຕິກໍາທາງເພດສໍາພັນ. ໃນ Psychopharmacology: ການຜະລິດສີ່ຂອງຄວາມຄືບຫນ້າ, Eds Bloom FE, Kupfer DJ (Raven, ນິວຢອກ), pp. 743-758.
- uf
- Pfaus JG,
- Gorzalka BB
(1987) Opioids ແລະພຶດຕິກໍາທາງເພດ. Neurosci Biobehav Rev 11: 1-34.
- uf
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Neurobiological correlates ຂອງອາລົມທາງບວກ: dopamine, ຄວາມຄາດຫວັງແລະລາງວັນ. ໃນການທົບທວນສາກົນຂອງການສຶກສາກ່ຽວກັບຄວາມຮູ້ສຶກ, ed Strongman (Wiley, ນິວຢອກ), pp. 31-50.
- uf
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) ການພົວພັນຂອງການປ່ອຍ dopamine ໃນນິວຄຽດມີຄວາມສໍາພັນກັບພຶດຕິກໍາທາງເພດຊາຍໃນຫນູ. Brain Res 524: 160-163.
- uf
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) ການປະຕິເສດຂອງການຂາດສານອາຊີບໂດຍຕົວແທນ serotonergic ແລະ noradrenergic. Behav Brain Res 62: 127-134.
- uf
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) ການມີສ່ວນຮ່ວມຂອງລະບົບການສູນກາງຂອງ noradrenergic ໃນການຟື້ນຟູການປະພຶດຮ່ວມຂອງຫນູທີ່ກໍາລັງກະທົບທາງເພດສໍາພັນໂດຍ yohimbine, naloxone, ແລະ 8-OH-DPAT. Brain Res Bull 38: 399-404.
- uf
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) antagonists opioid ແລະປະກົດການທາງເພດ. Psychopharmacol 122: 131-136.
- uf
- Rolls BJ
(1986) Satiety ຄວາມຮູ້ສຶກສະເພາະ. Nutr Rev 44: 93-101.
- uf
- Salamone JD
(1996) ການປະຕິບັດທາງ neurochemistry ຂອງແຮງຈູງໃຈ: ບັນຫາທາງວິຊາການແລະແນວຄິດໃນການສຶກສາຂອງກິດຈະກໍາແບບເຄື່ອນໄຫວຂອງນິວຄລີໂນ accumbens dopamine. J Neurosci Methods 64: 137-149.
- uf
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) ການປ່ອຍ Dopamine ຢູ່ໃນພື້ນທີ່ preoptic medial ໃນໄລຍະການປະພຶດ copulatory ຂອງຜູ້ຊາຍໃນຫນູ. Brain Res 692: 66-70.
- uf
- Schultz W
(1992) ກິດຈະກໍາຂອງ neurones dopamine ໃນສັດປະພຶດຕົວ. Semin Neurosci 4: 129-138.
- uf
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) ເຂດພື້ນທີ່ preoptic medial ແມ່ນມີສ່ວນຮ່ວມໃນການກະຕຸ້ນທາງເພດແລະການປະຕິບັດໃນຫນູຂອງຜູ້ຊາຍ: ການປະເມີນຄືນການເຄື່ອນໄຫວຂອງ neuron ໃນສັດທີ່ເຄື່ອນຍ້າຍ freely. Brain Res 640: 215-222.
- uf
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) ການແຜ່ກະຈາຍຂອງ Dopamine ເພີ່ມຂຶ້ນໃນລະດັບຂອງຫນູເພດໃນລະຫວ່າງການສໍາຜັດກັບຫນູເພດຍິງທີ່ໄດ້ຮັບການຮ່ວມເພດຄັ້ງທໍາອິດ. Brain Res 618: 41-46.
- uf
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Dopaminergic correlates ຂອງພຶດຕິກໍາທີ່ກະຕຸ້ນ: ຄວາມສໍາຄັນຂອງການຂັບລົດ. J Neurosci 15: 5169-5178.
- uf
- Wilson JR,
- Kahn RE,
- Beach FA
(1963) ການປ່ຽນແປງໃນພຶດຕິກໍາທາງເພດຂອງຫນູເພດທີ່ຜະລິດໂດຍການປ່ຽນແປງກະຕຸ້ນເພດຍິງ. J Comp Physiol Psychiatry 56: 636-644.
- uf
- Zetterstrm T,
- Sharp T,
- Ungerstedt U
(1984) ຜົນກະທົບຂອງຢາເສບຕິດ neuroleptic ກ່ຽວກັບການປ່ອຍ dopamine striatal ແລະ metabolism ໃນຫນູປຸກສຶກສາໂດຍການ dialysis intracerebral. Eur J Pharmacol 106: 27-37.
ບົດຄວາມອ້າງອີງບົດຄວາມນີ້
- Neuroplasticity ຂອງໂຣກ neuroplastic dopaminergic endogenous ໂດຍບໍ່ໄດ້ຮັບຜົນກະທົບຈາກແຫຼ່ງຂໍ້ມູນທາງດ້ານການຂະຫຍາຍຕົວທາງດ້ານເສດຖະກິດແລະທໍາມະຊາດ. ວາລະສານ Neuroscience, 25 ເດືອນມິຖຸນາ 2014, 34 (26): 8825-8836
- ຄວາມບໍ່ສົມດຸນໃນຄວາມອ່ອນໄຫວຕໍ່ກັບປະເພດຕ່າງໆຂອງລາງວັນໃນການຫຼີ້ນການພະນັນທາງໂພຊະນາການ ສະຫມອງ, 1 ສິງຫາ 2013, 136 (8): 2527-2538
- ກົດຫມາຍວ່າດ້ວຍທໍາມະຊາດແລະຢາເສບຕິດກ່ຽວກັບກົນໄກການປະສົມປະສານແບບປະສົມປະສານຮ່ວມກັບ {Delta} FosB ເປັນຜູ້ໄກ່ເກ່ຍທີ່ສໍາຄັນ Journal of Neuroscience, 20 February 2013, 33 (8): 3434-3442
- ການຕອບສະຫນອງ Neuronal ໃນ Nucleus Accumbens Shell ໃນລະຫວ່າງການປະພຶດທາງເພດໃນຊາຍຫນູ Journal of Neuroscience, 1 February 2012, 32 (5): 1672-1686
- ຮັກໂລແມນຕິກ: ລະບົບສະຫມອງຂອງສັດລ້ຽງລູກດ້ວຍນົມແມ່ສໍາລັບການເລືອກຄູ່ ທຸລະກິດ Philosophical ຂອງ Royal Society B: ວິທະຍາສາດຊີວະສາດ, 29 ເດືອນທັນວາ 2006, 361 (1476): 2173-2186
- ສະຖານທີ່ຮ້ອນ Hedonic ໃນສະຫມອງ Neuroscientist, 1 ເດືອນທັນວາ 2006, 12 (6): 500-511
- Modulation by Central and Basolateral Amygdalar Nuclei of Dopaminergic Correlates of Feeding to Satiety in the Rat Nucleus Accumbens and Medial Prefrontal Cortex Journal of Neuroscience, 15 December 2002, 22 (24): 10958-10965
- ຄວາມຖີ່ຂອງການປ່ຽນເສັ້ນເລືອດຂອງ Dopamine ເພີ່ມຂຶ້ນຂອງຫນ້າຕາແລະ Ventral Striatum ຂອງຫນູໃນລະຫວ່າງການນໍາສະເຫນີຂອງ Conspecifics Journal of Neuroscience, 1 December 2002, 22 (23): 10477-10486
- ການທົບທວນປື້ມ: Dynamics of Extracellular Dopamine ໃນການປະຕິບັດແບບສ້ວຍແຫຼມແລະຄໍໄລຍະຂອງ Cocaine ນັກວິທະຍາສາດ neuroscientist, 1 ສິງຫາ 2002, 8 (4): 315-322
- ການເຂົ້າຮ່ວມທີ່ແຕກຕ່າງກັນຂອງ NMDA, AMPA / Kainate, ແລະ Receptors Dopamine ໃນ Nucleus accumbens Core ໃນການຊື້ແລະປະສິດທິພາບຂອງ Behavior Approach Pavlovian Journal of Neuroscience, 1 December 2001, 21 (23): 9471-9477
- ການຄວບຄຸມພຶດຕິກໍາການໃຊ້ cocaine ໂດຍການກະຕຸ້ນທີ່ກ່ຽວຂ້ອງກັບຢາເສບຕິດໃນຫນູ: ຜົນກະທົບກ່ຽວກັບການຟື້ນຕົວຂອງລະດັບ dopamine ທີ່ໄດ້ຮັບການທໍາລາຍແລະໃນ extracellular amygdala ແລະ nucleus accumbens PNAS, 11 ເມສາ 2000, 97 (8): 4321-4326
- ການສະຫນັບສະຫນູນການປະພຶດທາງເພດແລະການເພີ່ມຂື້ນ Dopamine Efflux ໃນຄັນຮາກຂອງຮຸກຮານຂອງຜູ້ຊາຍຫຼັງຈາກ D-Amphetamine-Induced Behavioral Sensitization Journal of Neuroscience, 1 January 1999, 19 (1): 456-463
- NEUROSCIENCE: ໄດ້ຮັບຄວາມສົນໃຈຂອງສະ ໝອງ ວິທະຍາສາດ, 3 ເດືອນຕຸລາ 1997, 278 (5335): 35-37
- Dopaminergic Correlates ຂອງ Satiety Sensory-Specific Satisfaction ໃນ Medial Prefrontal Cortex ແລະ Nucleus Accumbens ຂອງຫນູ Journal of Neuroscience, 1 October 1999, 19 (19): RC29