КОМЕНТАРИ на YBOP: Ефектот „Кулиџ“ стои зад моќта на порно Интернет. Ефектот Кулиџ е феномен што се забележува кај видовите цицачи при што машките (и во помала мерка женките) покажуваат обновен сексуален интерес доколку се запознаат со нови приемчиви сексуални партнери, дури и по одбивање на секс од претходни, но сепак достапни сексуални партнери. Сексуалната новина ја надминува оваа навика со обновена возбуда предизвикана од повисок допамин. Континуиран прилив на новини е она што го прави Интернет порно-толот толку различно од порно од минатото.
Оригинална статија, со графикони
- Денис Ф. Фиорино,
- Ариан Кури, и
- Ентони Г. Филипс
- Весник на невронски мрежи, 15 јуни 1997 година, 17 (12): 4849-4855;
Апстракт
Ефектот Coolidge ја опишува повторната употреба на сексуалното однесување во "сексуално засиленото" животно како одговор на новиот рецептивен партнер. Со оглед на улогата на мезолимбичниот допамин (ДА) систем во иницирањето и одржувањето на мотивираното однесување, микродиализата се користеше за следење на преносот на нуклеус акмунсенс (NAC) DA во текот на копулацијата, сексуалната ситост и повторното ставање во сексуално однесување. Во согласност со претходните извештаи, презентацијата на естропатична женка зад екран и копулација беа поврзани со значително зголемување на NAC DA efflux. Враќањето на NAC DA концентрациите во основните вредности се совпадна со период на сексуална ситост, иако концентрациите на метаболитите на ДА, дихидроксифенилацетичната киселина и хомованичната киселина останале покачени. Презентацијата на новата рецептивна женка зад екран резултираше со мало зголемување на NAC DA, што беше значително зголемено за време на обновената копулација со новата женка. Податоците покажуваат дека стимулативните својства на новата рецептивна женка може да послужат за зголемување на NAC DA трансмисијата кај сексуално засилен машки стаорец, а тоа, пак, може да биде поврзано со повторно воведување на сексуално однесување.
Вовед
Машкиот стаорец кој се припишува на ситост може да биде поттикнат повторно да се резимира ако почетната женка се замени со нова рецептивна женка. Ова е познато како ефект Кулиџ и е забележано кај голем број видови на цицачи (Wilson et al., 1963). Општите фактори како што се замор или моторна депресија не се доволни за да се објасни очигледната состојба на сексуална наслада, бидејќи стимулациите од роман женка сè уште може да предизвикаат копулација. Сексуалната ситост, исто така, може да биде "обратна" фармаколошки, во значителен степен, со администрација на различни лекови кои можат да дејствуваат на различни невротрансмитерски системи. Овие лекови вклучуваат јохимбин, 8-OH-DPAT (Родригез-Манцо и Фернандез-Густи, 1994, 1995a), налаксон (Pfaus и Gorzalka, 1987; Родригез-Манцо и Фернандез-Густи, 1995a,b), и апоморфин (Mas et al., 1995c). Иако периферното дејство на овие лекови не може да се отфрли (на пример, адренергични ефекти врз еректилната функција), ефектите врз централните механизми за сексуална ситост се предложени врз основа на селективни централни норадренергични лезии (Родригез-Манцо и Фернандез-Густи, 1995a) и експерименти со микродиализа кои го следеа допаминергичниот метаболизам во медијалната преоптична област (Mas et al., 1995a,b).
Со оглед на тоа што централните механизми можат да посредуваат во повторното ставање на сексуалното однесување кое е карактеристично за ефектот Coolidge, најверојатен кандидат е мезолимбичниот допамин (DA) систем, проектиран од вентралната тегенментална област до НАК. Мезолимбиќ Д.А. се чини дека делува како примарен модулатор во сложените интегративни процеси кои вклучуваат евалуација на стимули од животната средина, како што се знаци од сексуално приемчивата женка и организацијата на однесување насочено кон цели, вклучувајќи го и кополирањето (Фибигер и Филипс, 1986; Блекбурн и сор., 1992; Филипс и сор., 1992; Лемоал, 1995; Саламон, 1996).
Иако средниот мозок DA невроните реагираат на примарни награди и знаци кои предвидуваат награда, новите или непредвидливи еколошки стимули, најсилно предизвикуваат невронална активација во текот на повторните тренинзи (Фабре и сор., 1983; Шулц, 1992; Миренович и Шулц, 1994). Постојат голем број на докази кои ја поддржуваат важната олеснувачка улога за мезолимбичен ДА во иницирањето и одржувањето на сексуалното однесување на стаорци (Pfaus и Everitt, 1995), а голем број на студии за микродијализа покажуваат зголемување на NAC DA efflux за време на апетитивните и консумирачките фази на машкото сексуално однесување (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Меѓутоа, постојат релативно малку податоци за неврохемиските корелати на сексуалната наслада и за повторно воспоставување на сексуалното однесување. Примената на ин виво Микродијализата за следење на мезолимбичната DA невротрансмисија за време на ефектот Coolidge обезбедува единствена можност да ја испита улогата на NAC DA во копулација, сексуална ситост и повторно воспоставување на копулација.
Експеримент за микродијализа беше спроведен за да се утврди следново: (1) дали појавата на сексуална ситост е придружена со враќање на екстрацелуларните DA концентрации во НАК до вредности на преципитација или подолу, и (2) дали повторно враќање на копулаторното однесување во " сексуално заситен "машки стаорец со нова рецептивна женка е во корелација со зголемувањето на NAC DA efflux.
МАТЕРИЈАЛИ И МЕТОДИ
Предмети. Машките Спрајг Даули стаорци, добиени од Центарот за грижа за животни (на Универзитетот во Британска Колумбија) и женските стаорци Лонг-Еванс, добиени од Чарлс Ривер Канада (Св Констанс, Квебек, Канада), биле сместени во жичени мрежни кафези (18 × 25 × 65 см, пет за кафез) во посебни соби во колонија. Колонија соби се одржуваше на температура од ~20 ° C на обратен 12 час светло / темно циклус. Стаорците имаа неограничен пристап до храна (Purina Rat Chow) и вода.
Хирургија и тестирање на однесувањето пред микродиализа на мозокот.Женските стаорци биле била ovariectomized билатерално под халотанска гасна анестезија (Fluothane, Ayerst Laboratories) најмалку 4 недели пред тестирање. Сексуалната рецептивност кај стимулирачките жени беше индуцирана со поткожни инјекции на естрадиол бензоат (10 μg) и прогестерон (500 μg), 48 и 4 ч, соодветно, пред секоја сесија за тестирање. Машките стаорци беа прикажани за сексуално однесување во два наврати, 4 дислоцирани, во коморите од плексиглас (35 × 35 × 40 cm) со подови со жица. Само машките стаорци кои достигнаа критериум на изведба, кои вклучуваа интромизација во рамките на 5 мин од презентацијата на женскиот и ејакулацијата во рамките на 15 мин од првата интромизација, за време на двата скрининг тестови беа имплантирани со канали за испитување на микродијализа.
Машки стаорци (n = 5) биле анестезирани со кетамински хидрохлорид (100 mg / kg, ip) и ксилазин (10 mg / kg, ip) пред стереотаксична хирургија. Микродијализата канал сонда за мерење (19 мерач) била имплантирана билатерално над NAC (координати од брегма: предна страна, + 1.7 mm, медијален, ± 1.1 mm, вентрален, -1.0 mm, рамна черепа) и беа обезбедени во черепот со стоматолошки акрилни и завртки на златар. Билатералните импланти на канули за вода беа искористени за да се зголеми можноста за успешен експеримент за микродиализа. За среќа, во овој експеримент беше потребна само една канила за секој стаорец. Машките стаорци биле сместени индивидуално во големи пластични кафези со постелнина од плунка за остатокот од експериментот. Една недела по операцијата, стаорците биле тестирани за сексуално однесување. За време на овој дел од обуката, комората за тестирање била опремена со лизгачки екран од плексиглас кој ја поделил комората во големи и мали прегради. Машките стаорци беа воведени во големиот дел и 15 мин подоцна, позади екранот беше поставена женка. По подготвителниот период на 15 min, екранот беше отстранет, а стаорците им беше дозволено да копулираат за 30 мин. Беа спроведени три сесии за обука, по еден 4 d. Сите стаорци го достигнаа критериумот за изведба во текот на секоја сесија.
Експеримент со ефект Coolidge. Стаорците беа имплантирани еднострано со сонди за микродијализа 12-18 час пред ефектот на Coolidge ефект и се ставија во големиот дел од комората за тестирање со слободен пристап до храна и вода. Утрото на експериментот, примероци од микродијализа беа собрани секој 15 мин. Експериментот се состоеше од следните седум последователни фази: (1) основната линија (најмалку 60 мин); (2) женски 1 зад екранот (15 мин); (3) со женски 1 додека 30 min период не помина без монтирање; (4) реинтродукција на женскиот 1 зад екранот (15 мин); (5) пристапот до женскиот 1 за 15 мин период под услов да нема монтирање (ако се случи монтажа, оваа фаза беше третирана како фаза 3); (6) воведување на женски 2 зад екранот (15 мин); 7) копулација со женски 2 за 60 мин.
Однесувањето беше снимено под слаба осветленост користејќи видео систем JVC и забележано на видео монитор лоциран надвор од просторијата за тестирање. Стандардни мерки на сексуално однесување беа снимени со користење на компјутер и соодветен софтвер (Холмс и сор., 1987).
По експериментот со микродиализа, на животните им се даваше предозирање со хлорален хидрат и перфузирани интракардично со солен и формалин (4%). Мозоците беа исечени и замрзнати, а потоа, короналните делови беа обоени со кресливи виолетови за да се утврди поставувањето на микробиолошките сонди. Само стаорци со пласирање на сонди во рамките на НАК беа користени за однесувањето и неврохемиските анализи.
Микродијализа и HPLC-електрохемиска детекција. Микродијализа сондите беа концентрични во дизајнот со полупропустлива шуплива влакна мембрана (2 mm мембрана изложена, надворешен дијаметар 340 μm, 65000 отсекување на молекуларна тежина, Фитрал 12, Хоспал) на дисталниот крај. Пробоните беа перфузирани на 1.0 μl / min со модифициран Ringer-овиот раствор (0.01 m натриум фосфат пуфер, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) со шприц за гас (Хамилтон, Рено, НВ) и пумпа за шприц (модел 22, Харвард Аппарат, Јужна Натик, М.А.). Приклучокот со микродијализа сондата се користеше за да се обезбеди сондата за микродијализа во внатрешноста на водечката канила. Челична калем, прицврстена на течно навртување (Instech 375s), која беше монтирана на врвот на комората за тестирање, беше искористена за заштита на цевката на сондата (Фиорино и сор., 1993).
Микродијализатните анализи, во кои беа вклучени ДА и неговите метаболити дихироксифенилацетична киселина (DOPAC) и хомованилна киселина (HVA), беа одделени со хроматографија со обратна фаза (колона Ултрасфера, Бекман, Фулвертон, CA, ODS 5 μm, 15 cm, 4.6 mm, внатрешен дијаметар ) со користење на 0.083m натриум ацетатен пуфер, pH 3.5 (5% метанол). Концентрациите на анализата се квантифицирани со електрохемиска (ЕК) детекција. Апаратот се состоеше од пумпа Bio-Rad (Richmond, CA), Valco Instruments (Хјустон, TX) EC10W двостран инјектор, ESA (Бедфорд, MA) Coulochem II EC детектор и двоканален рекордер на табели (Kipp и Зонен, Бохемија, Њујорк). Електрохемиските параметри за детектори беа следните: електрода 1, + 450 mV; електрода 2, -300 mV; и заштитна ќелија, -450 mV. Типични закрепнувања на сондата, спроведени ин витро и на собна температура, биле 22% за DA, 18% за DOPAC и 18% за HVA.
РЕЗУЛТАТИ
Однесување
Бихевиоралните мерки од експериментот ефект Coolidge се претставени во Табела 1. Латенциите за монтирање, интроматит и ејакулација, како и постејкулаторниот интервал по првата ејакулација беа слични на оние во претходната обука (податоци не прикажани). Ова укажува на тоа дека процедурата на микродијализа не го изменило нормалното сексуално однесување. Развојот на сексуалната наслада, како што е прикажано со средниот број на ејакулации пред да се исполни критериумот (7.8 ± 0.5), прогресивно намалување на бројот на интромиози кои претходат на секоја ејакулација и прогресивно зголемување на постејакулаторниот интервал (податоци не прикажани) , беше слична на онаа пријавена во претходните студии (Плажа и Јордан, 1956; Фаулер и Вилен, 1961; Фишер, 1962; Bermant et al., 1966; Родригез-Манцо и Фернандез-Густи, 1994; Mas et al., 1995d). Индивидуална варијабилност беше забележана во однос на бројот на ејакулации постигнати кај женскиот 1, времето поминато во копулирање со женскиот 1 и бројот на презентации на женски 1 потребни за постигнување на критериумот за ситирање (Табела 1, долу). Некои стаорци бараа бројни реинтродукции на женскиот 1 додека фазата 5 не заврши (n = 3). Актите на ставање на женскиот 1 зад екранот и отстранувањето на партицијата можеби служеле како примарни апетитивни знаци кои довеле до копулација. Исто така, треба да се забележи дека критериум на ситирање на 30 мин без монтирање, иако претходно се користел (плажа и Јордан, 1965; Mas et al., 1995b), е произволно и не гарантира дека стаорецот нема да биде монтиран со оглед на повеќе време. Дури и така, одложувањата или процедурите за отстранување и замена не резултирале сигурно во обновената копулација со женскиот 1 (на пр. Фази 4 и 5).

Однесување во текот на експериментот ефект Coolidge
Сите стаорци го изложија ефектот Coolidge. Активноста поврзана со поставување на женскиот 2 зад екранот, а особено отстранувањето на партицијата може да придонесе за овој резултат, но, повторно, овие настани сами по себе не беа доволни за обновување на копулацијата порано во експериментот. Компарациите помеѓу мерките за сексуално однесување со женски 1 и женски 2 беа направени со користење t тестови со корекција на Bonferroni. Иако латенциите на монтирање и интромирање во одговор на женскиот 2 не се разликуваа значително од оние во првиот копулаторски рунда кај женскиот 1, воопшто, сексуалното однесување со женскиот 2 беше помалку робусно, како што е прикажано со значително помалку ејакулации (значи, 0.6 vs 4.2; F = 49.86;p <0.01) и интромисии (просек = 11.2 наспроти 37.0;F = 20.17; p <0.05) во текот на првиот час. Бројот на поставувања во првиот час со жени 1 и 2 не се разликуваше значително.
Важно е да се забележи дека женките кои се користат за време на сатисниот дел од експериментот (т.е. женски 1) сеуште покажуваат силен процептивен (т.е. потскокнување и стрелање) и рецептивното (т.е. лордоза) однесување за целото времетраење на нивниот контакт со машкиот .
Неврохемија
Базалните наномоларни концентрации на DA и неговите метаболити во микродијализатите, претставени како средна ± SEM, од првите три основни примероци беа: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; и HVA, 234.2 ± 49.0 (корегиран за наплата на сондата;n = 5). Овие вредности ги претставуваат резултатите од основната линија на 100%.
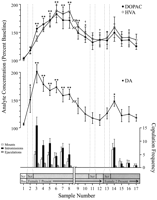
Одредени податоци поени кои одговараат на секоја фаза од експериментот и се заеднички за секој стаорец, се користат за неврохемиски анализи. Тие вклучуваат следново: (1) седум примероци по првото воведување на женски 1, (2) четири примероци придружени со отсуство на копулаторно однесување со женски 1 и (3) пет примероци по презентацијата на женскиот 2. Слика 1 ги илустрира промените во концентрациите на ДА (линиски графикон, средината) и DA метаболити (линиски график, врв) паралелно копулаторно однесување (бар-график, дното) за време на тестот за ефектот Coolidge.
Погледнете поголема верзија:
Сл. 1.
Нуклеус accumbens неврохемиски корели на сексуално однесување за време на Coolidge ефект. Првите осум примероци претставуваат хронолошки континуирани податоци точки од фазите 1 до 3. Примерот 1 е четвртата и последната примерока на основната преопулација (Bas). Примерот 2 претставува воведување на женски 1 зад екранот (Scr). По 15 min, екранот беше отстранет, и на стаорците им беше дозволено да се копулираат (примероци 3-8). Насе скрши на x-аксисот одговара на исклучувањето на податоците од три стаорци кои се подолго време копулирани со почетната женка. Последните девет примероци исто така беа постојани хронолошки. Примероците 9 и 10 одговараат на периодот на ситирање на фазата 3 (т.е. 30 min без монтирање). Женскиот 1 потоа беше повторно вметнат зад екранот (примерок 11) и, 15 мин подоцна, екранот беше отстранет (мостра 12). По 15 min без копулација, женскиот 2 беше поставен зад екранот (мостра 13). Примероците 14-17 одговараат на копулација со женскиот 2. Бројот на монтирања, интроминиуми или ејакулации поврзани со секоја примерока на ХНУМКС мид мидродијализа се прикажани во долен график. Неврохемиските податоци се изразени во однос на процентот на основните концентрации. Промени во NAC DA (затворени квадрати), ДОПАК (затворени кругови), и HVA (отворени кругови) efflux се претставени како линија графики. Беа направени следните споредби: основна мостра 1 наспроти примероци 2-10; нова основна мостра 10 наспроти примероци 11 и 12; нова основна мостра 12 наспроти примероци 13-17 (*p <0.05; ** p <0.01). Независен t беа направени тестови помеѓу основните вредности (примероци 1, 10 и 12). За значајни разлики од првата основа (мостра 1), †p <0.05.
Одделни еднонасочни, повторени мерки ANOVA беа извршени на неврохемиски податоци поврзани со женски 1 (примероци 1-12) и женски 2 (примероци 12-17). А априори споредби беа направени со помош на Dunn's multiple comparison test (Bonferroni t). Следните три главни споредби се направени: (1) почетна основа (примерок 1) наспроти примероци 2-10 (прва изложеност на женски 1), (2) втора основа (примерок 10) наспроти примероци 11 и 12 (повторно изложување на женски 1) , и (3) трета основна линија (мостра 12) наспроти примероци 13-17 (изложеност на женски 2).
Имаше значителна севкупна промена во DA efflux како одговор на женскиот 1 [F (11,44) = 8.48; p <0.001] и жена 2 [F (5,20) = 2.83;p <0.05]. Значително зголемување на приливот на ДА беше откриено кога жената 1 беше присутна зад екранот (+ 44%,p <0.05; примерок 2). За време на копулацијата, концентрациите на ДА се зголемија понатаму, достигнувајќи максимална вредност (+ 95%;p <0.01) за време на првиот меч за копулација (примерок 3). ДА остана воздигнат во текот на копулацијата и се врати само на почетната концентрација во периодот од 30 мин во кој не се случи монтажа (примероци 9 и 10). Ниту повторно воведување на женка 1 зад екранот (примерок 11) ниту можност за физичка интеракција, но без монтирање (примерок 12), зголемени концентрации на DA во однос на втората почетна вредност (примерок 10). Присуството на жена 2 зад екранот (примерок 13) резултираше со мало зголемување на приливот на ДА (12%) од третата почетна вредност (примерок 12) што не достигна статистичко значење. Обновената копулација со жени 2 резултираше со значително (34%) зголемување (p <0.05) во ефлукс на ДА при првиот примерок за копулација (примерок 14). Иако слабото однесување на копулаторот продолжи во текот на следните три примероци, концентрациите на ДА се намалија на почетните вредности (примероци 15-17). Независен t Тестовите спроведени помеѓу "основните" примероци (т.е. 1, 10 и 12) покажаа дека овие вредности не се значително различни.
Во трите стаорци кои продолжиле со копулација кога повторно се внесува женка 1, концентрациите на NAC DA се зголемиле кога женката 1 била присутна зад екранот (опсег, 25-47%) и за време на копулација (опсег, 13-37%), во однос на примерокот пред повторното вградување на женката. Овие зголемувања, сепак, се случиле само кога сексуалното однесување е енергично и доведе до ејакулација.
Значителни севкупни промени кај DOPAC [F (11,44) = 9.57; p <0.001] и ХВА [F (11,44) = 12.47; p Концентрациите на метаболитот се зголемија малку (+ 0% во двата случаи) за време на презентацијата на жената 001 зад екранот (примерок 1), но ова не беше значајно статистички Меѓутоа, имаше значително зголемување на концентрациите на DOPAC и HVA за време на копулацијата (примероци 2-15), достигнувајќи ги максималните вредности (+1 и + 2%, соодветно; p <0.01) по 60 мин (примерок 6 и во двата случаи). Иако концентрациите на метаболит се намалија за време на периодот на сексуална неактивност на крајот од контактот со женката 1 (примероци 9 и 10), концентрациите сепак останаа покачени во однос на почетната линија (p <0.05 и во двата случаи). Повторно воведување на женка 1 зад екранот (примерок 11), пристап до женка 1 по отстранување на екранот (примерок 12) и воведување на жена 2 (примерок 13) не резултираше со какви било промени во концентрацијата на метаболит. Мало, но статистички незначително, зголемување на концентрациите на DOPAC и HVA (+ 23% во двата случаи) во однос на почетната состојба (примерок 12) одговараше на првиот период на копулација со жени 2 (примерок 14). Сепак, ова зголемување траеше кратко и не се согласи на основните вредности за преостанатите три примероци (15-17). Независен tтестовите спроведени помеѓу "основните" примероци (т.е. 1, 10 и 12) покажаа дека втората и третата основна вредност (примероци 10 и 12, соодветно), иако не се разликуваат една од друга, останале значително покачени во споредба со првиот примерок од основната линија за DOPAC и HVA (p <0.05 и во двата случаи).
Хистологија
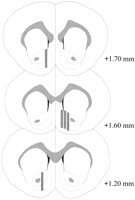
Микродијализа сондите биле лоцирани во НАК (сл.2) во опсег што се протега + 1.20 до + 1.70 mm од брегма (рамен череп). Постоела варијабилност и во медиолатералниот план; податоците одразуваат земање мостри од школка и основни подрегиони на НАК.
Погледнете поголема верзија:
Сл. 2.
Локација на сонди за микродијализа во НАК на машки стаорци кои се користат во експериментот ефект Coolidge. Засенчени правоаголници кореспондираат со изложената мембранска површина на сонди за микродијализа. Серијалните коронални делови на мозокот беа прецртаниПаксинос и Вотсон (1986).
ДИСКУСИЈА
Во согласност со претходните извештаи, сегашните резултати покажуваат зголемен мезолимбичен DA трансмисија поврзана со апетитивните и консумирачките компоненти на сексуалното однесување кај мажите на стаорци, како што е оценето один виво микродиализа (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Покрај тоа, овие резултати обезбедуваат неврохемиска корелација за сексуална заситеност и последователна реиницијација на копулацијата како одговор на новите рецептивни женки (ефектот Coolidge). Податоците покажуваат дека стимулативните својства на новата рецептивна женка може да послужат за зголемување на NAC DA трансмисијата кај сексуално засилен машки стаорец, кој, пак, може да биде поврзан со повторно воведување на сексуално однесување. Ова најпрво се забележува при мало зголемување на NAC DA за време на презентацијата на новата женка зад екранот и се случува најубедливо како понагласен пораст за време на обновената копулација со женскиот 2 (сл.1).
Присуството на првата рецептивна женка зад екранот резултираше со робусно апетитно зголемување на NAC DA efflux (44% од основната линија) слично во големината на она што беше објавено во претходните експерименти користејќи сличен дизајн (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992) Исто така, во согласност со овие студии беше и набудувањето дека NAC DA ефлуксот беше подобрен понатаму за време на копулацијата (до> 95% над основната линија во овој експеримент). Иако можеме да ги сметаме потрошувачките однесувања како поврзани со зголемено ослободување на NAC DA (Wenkstern et al., 1993; Wilson et al., 1995), важно е да се испитаат термините "апетитив" и "консумирање" во контекст на сексуалното однесување. Бидејќи фазата во која женката е присутна зад екранот е исклучиво апетитна или подготвителна, однесувањето во фазата на копулација не може да се смета за чисто консумативна. Бидејќи "апетитивното" може да се користи за опишување на сите однесувања што доведуваат до конзумирање на мотивирано однесување (копулација), примарното однесување на машките експонати додека е активно во "консумирната" фаза најдобро се опишува како апетит; мажот го троши поголемиот дел од своето време и напор што го натера женското да се сопне. Во овој поглед, можеме да корелираме максимален NAC DA пренос со консумативен како и интензивни апетитивни компоненти на сексуалното однесување на машкиот стаорец.
Пристапот до втората, роман женка резултираше со обновена копулација во секој предмет. Претходните истражувања покажаа дека поголемиот дел од стаорците дозволено да се копулираат на ситост, користејќи сличен однесен протокол кон оној што се користи во овој експеримент, не продолжи со парење при тестирање на 24 час подоцнаПлажа и Јордан, 1956). Многу е веројатно дека присуството на новите стимулативни својства на женскиот 2, кој може да вклучува и мирисни, како и визуелни и аудитивни знаци, резултираше со обновена копулација. Интересно прашање, што останува да се одговори, е со каков механизам машкиот стаорец разликува нова женка од женка со која неодамна се соединил. Местото за тој механизам може да лежи во главниот олфакторски систем. Забележано е дека интегритетот на овој систем е од суштинско значење за ефектот Кулиџ во хрчаци (Џонстон и Расмусен, 1984). Сепак, омефариорниот систем на воomeronasal-додаток, во кој неодамна беше опишан феромонален процес на меморија кај глувците (Kaba et al., 1994), исто така е главен кандидат. Во овој поглед, вреди да се забележи дека зголемувањето на NAC DA пренос се мери користејќи ин виво voltammetry кај машки стаорци претставени со постелнина која била изложена на женски стаорци во еструс (Louillot et al., 1991; Мичел и Гратон, 1992). Понатаму, примената на К.+ директно на слојот на венероназалниот нерв на дополнителната олфакторна сијалица, како и на самата дополнителна олфакторна сијалица, била доволна за да се зголеми NAC DA трансмисијата (Мичел и Гратон, 1992).
Првата 15 минијатурна рунда на копулација со женскиот 2 била поврзана со значително зголемување на NAC DA. За разлика од женскиот 1, интеракцијата со женскиот 2 не произведувала зголемување на NAC DA со иста големина за време на апетитивните (12%) или консумирање (34%) фази. Овие мали зголемувања на NAC DA, сепак, добро се поврзуваат со намаленото ниво на сексуално однесување прикажано кај женскиот 2 во споредба со женскиот 1. Концентрациите на метаболитот останале покачени за време на фазата на ситост, што резултирало со нови базични концентрации (примероци 10 и 12) кои биле значително покачени од почетната вредност (примерок 1).
Временскиот застој во зголемувањето на концентрациите на DOPAC и HVA за време на копулирањето е во согласност со нивното формирање како метаболити на родното соединение, ДА. Се претпоставува дека концентрациите на метаболитот на микродијализа, барем за време на природното однесување кое не е фармаколошки управувано, обезбедува корисен индекс на нервната активност (Damsma et al., 1992; Fumero et al., 1994). Фактот дека концентрациите на метаболитот останале покачени дури и за време на периоди на сексуална неактивност во овој експеримент, кога концентрациите на ДА се вратија во основните вредности на предлози, фрла сомнеж врз овој предлог.
Постојаното покачување на концентрациите на DA метаболитот забележано во овој експеримент го отсликува профилот на медијалниот преоптичен регион (mPOA) на DA метаболитите забележани кај стаорци првиот ден откако тие се копитале во насилство (Mas et al., 1995a,b). Одржливи покачувања на концентрациите на DOPAC и HVA во NAC или mPOA не се секогаш забележани кога периодот на парење е фиксно траење, многу пократок од времето потребно за постигнување на ситост. На пример, многу студии покажале дека концентрациите на ДОПАК биле зголемени и останале покачени за време на копулирањето, но одбиле да ги земат основните вредности наскоро откако женката била отстранета (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Во студијата Mas et al. (1995b), базалните екстрацелуларни концентрации на DOPAC и HVA во mPOA останаа покачени над 4 последователни денови соодветни на периодот на сексуална неактивност. До четвртиот ден, непосредно пред животните да продолжат со копулација, базалните концентрации на метаболитите беа блиску до вредностите на пресадија. Авторите ја споредиле моделот на неврохемиски промени со оние забележани по администрацијата на блокаторите на DA рецептор (Zetterström et al., 1984; Императо и ДиКиара, 1985) и сугерираа дека состојбата на сексуална неактивност може да биде посредувана преку ослободување на пролактин, што може да дејствува како "ендогена невролептика" (Mas et al., 1995a,b,d). Јасно е дека неуролептичната администрација е придружена со зголемување на концентрациите на екстрацелуларниот метаболит и DA efflux (Zetterström et al., 1984; Императо и ДиКиара, 1985). За жал, Mas et al. (1995a,b) не беа во можност да ги детектираат концентрациите на mPOA DA. Во оваа студија, концентрациите на DA во NAC се вратија во вредности на претопулација, додека концентрациите на DOPAC и HVA останаа покачени. Овој модел не е во согласност со улогата на ендогени невролептични дејства во НАК за да предизвикаат сексуална ситост.
Со оглед на вклученоста на мезолимбични DA неврони во мотивирано однесување (Фибигер и Филипс, 1986; Блекбурн и сор., 1992; Kalivas et al., 1993; Лемоал, 1995) и нивната чувствителност кон новите еколошки стимули (Фабре и сор., 1983; Шулц, 1992; Миренович и Шулц, 1994), забележаните зголемувања на екстрацелуларните концентрации на NAC DA како одговор на ромската женка се во согласност со хипотезата дека активноста во овој DA систем е важна за повторно воспоставување на сексуалното однесување. Дополнително, извештаите за апетитот и потрошувачката се зголемуваат во пренос на DA (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) и невронска активност (Shimura et al., 1994) кај mPOA на машките стаорци за време на сексуалното однесување укажуваат на тоа дека оваа структура може исто така да придонесе за обновена копулација карактеристика на Coolidge ефект.
Во согласност со општата улога на мезолимбичниот ДА систем во мотивирано однесување, добро е утврдено дека екстрацелуларните концентрации на ДА исто така се покачени пред, за време и веднаш по консумирање на оброк, со враќање на основните вредности ~30 мин подоцна (Wilson et al., 1995). Добро е познато дека ситоста индуцирана од храната е под влијание на неговите сензорни својства. Луѓето и животните ја отфрлаат храната на која се хранат на ситост и внесуваат други видови храна што не се јаде (Ролс, 1986). Ова го покренува прашањето за тоа дали екстрацелуларниот DA efflux во НАК ќе се зголеми селективно со презентација на нов тип на храна, но не со храна што неодамна се консумира на ситост на начин сличен на оној пријавен во оваа студија во контекст на сексуална мотивација. Ако се потврди, овој општ однос меѓу сензорните својства на природните награди, ситост и мезолимбичен DA пренос би значел критична улога за овој нервен систем во регулирањето на мотивациските процеси, нарушување на кои може да доведе до сериозни нарушувања на јадењето и сексуалната функција .
Користена литература
- ↵
- Beach FA,
- Jordan L
(1956) Сексуална исцрпеност и опоравување кај машкиот стаорец. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Андерсон Л
(1966) Временски карактеристики на Coolidge ефектот кај копулаторното однесување кај мажи. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Блекбурн ЈР,
- Pfaus JG,
- Филипс АГ
(1992) Допамин функционира во апетитни и дефанзивни однесувања. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Сексуалното однесување го зголемува допаминскиот пренос во јадрото accumbens и striatum на машки стаорци: споредба со новина и локомотива. Behav Neurosci 106: 181-191.
- ↵
- Фабре М,
- Rolls ET,
- Ештон ЈП,
- Вилијамс Г.
(1983) Активност на невроните во вентралниот тегентален регион на однесувањето мајмун. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Филипс АГ
(1986) Награда, мотивација, познание: психобиологија на мезотелицентните допамински системи. во Прирачникот за физиологија: нервниот систем IV, едс Блум Ф.Е., Гејгер С.Д. (Американско друштво за физиологија, Бетесда, Д.Д.), стр. 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Филипс АГ
(1993) Електрична стимулација на места за награда во вентралната тектонска област го зголемува допаминскиот пренос во јадрото accumbens на стаорецот. Behav Brain Res 55: 131-141.
- ↵
- Фишер А
(1962) Ефекти од стимулираната варијација на сексуалната наслада кај машкиот стаорец. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Фаулер Х,
- Whalen RE
(1961) Варијација на поттик стимул и сексуално однесување кај машкиот стаорец. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Фумеро Б,
- Фернандез-Вера Ј.Р.,
- Гонзалес-Мора Ј.Л.,
- Mas M
(1994) Промени во моноаминскиот промет кај мозочните области поврзани со машко сексуално однесување: студија за микродијализа. Мозокот Res 662: 233-239.
- ↵
- Холмс ГМ,
- Holmes DG,
- Sachs BD
(1987) Систем за собирање на податоци базиран на IBM-PC за снимање на сексуалното однесување на глодарите и за снимање на општ случај. Физиол Behav 44: 825-828.
- ↵
- Hull EM,
- Итон РК,
- Мојсеј Ј,
- Lorrain DS
(1993) Коуплацијата ја зголемува активноста на допамин во медијалната преоптична област на машки стаорци. Life Sci 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Екстрацелуларен допамин во медијалната преоптична област: импликации за сексуална мотивација и хормонална контрола на копулацијата. Ј Neurosci 15: 7465-7471.
- ↵
- Императо А,
- DiChiara G
(1985) ослободување на допаминот и метаболизам кај бубрежни стаорци по системски невролептици како што се испитува преку транс-половата дијализа. Ј Neurosci 5: 297-306.
- ↵
- Џонстон РЕ,
- Расмусен К.
(1984) Индивидуално препознавање на женски хрчаци од машки: улога на хемиски знаци и мирисни и вомороназни системи. Физиол Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Игучи Т,
- Nakanishi S
(1994) Индукција на олфакторна меморија со активирање на метаботропен глутамат рецептор. наука 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Куки MS
(1993) Фармакологија и нервни кола на сензибилизација на психостимуланти. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Мезокортиколимбични допаминергични неврони. Функционални и регулаторни улоги. во Психофармакологија: четвртата генерација на напредок, изданија Блум Ф.Е., Купфер Д.Ј. (Равен, Њујорк), стр. 283-294.
- ↵
- Louillot A,
- Гонзалес-Мора Ј.Л.,
- Гвадалупе Т,
- Mas M
(1991) Олфакторните стимули поврзани со сексот предизвикуваат селективно зголемување на ослободувањето на допамин во јадрото accumbens на машки стаорци. Мозокот Res 553: 313-317.
- ↵
- Mas M,
- Гонзалес-Мора Ј.Л.,
- Louillot A,
- Единствен Ц,
- Гвадалупе Т
(1990) Зголемено ослободување на допамин во јадрото accumbens на копулирачки машки стаорци како што беше потврдено со in vivo voltammetry. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Фумеро Б,
- Фернандез-Вера Ј.Р.,
- Гонзалес-Мора JL
(1995a) Неврохемиски корелати на сексуална исцрпеност и опоравување, како што се проценува со in vivo микродиализа. Мозокот Res 675: 13-19.
- ↵
- Mas M,
- Фумеро Б,
- Гонзалес-Мора JL
(1995b) Волтамметрична и микродиализа следење на ослободување на моноамински невротрансмитер за време на социо-сексуалните интеракции. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Фумеро Б,
- Перез-Родригез I
(1995c) Индукција на однесувањето на парење од апоморфин кај сексуално изладени стаорци. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Фумеро Б,
- Перез-Родригез I,
- Гонзалес-Мора JL
(1995d) Неврохемијата на сексуална ситост. Експериментален модел на инхибирана желба. во фармакологија на сексуалната функција и дисфункција, ед Bancroft J (Raven, Њујорк), pp 115-126.
- ↵
- Миренович Ј,
- Schultz W
(1994) Важноста на непредвидливоста за наградување на одговорите кај примарни допаминергични неврони. J Neurophysiol 72: 1024-1027.
- ↵
- Мичел Ј.Б.,
- Gratton A
(1992) ослободување на месолимбични допамини предизвикани од активирање на додатен олфакторниот систем: брза хроноамперометриска студија. Neurosci Lett 140: 81-84.
- ↵
- Паксинос Г,
- Вотсон Ц
(1986) Мозокот на стаорци во стереотаксични координати (2nd ед). (Академски, Сан Диего).
- ↵
- Pfaus JG,
- Damsma G,
- Номикос Г.Г.,
- Wenkstern D,
- Блаха ЦД,
- Phillips AG,
- Fibiger HC
(1990) Сексуалното однесување го подобрува централниот допамин трансмисија кај машкиот стаорец. Мозокот Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Психофармакологијата на сексуалното однесување. во Психофармакологија: четвртата генерација на напредок, изданија Блум Ф.Е., Купфер Д.Ј. (Равен, Њујорк), стр. 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Опиоиди и сексуално однесување. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Блаха ЦД,
- Pfaus JG,
- Блекбурн Џ
(1992) Невробиолошки корелации на позитивни емоционални состојби: допамин, исчекување и награда. во Меѓународниот преглед на студии за емоции, ед Strongman (Вили, Њујорк), pp 31-50.
- ↵
- Pleim ET,
- Маточик ЈА,
- Barfield RJ,
- Auerbach SB
(1990) Корелација на ослободувањето на допамин во јадрото accumbens со машко сексуално однесување кај стаорци. Мозокот Res 524: 160-163.
- ↵
- Родригез-Манзо Г,
- Фернандез-Густи А
(1994) Пренасочување на сексуална исцрпеност со серотонергични и норадренергични агенси. Behav Brain Res 62: 127-134.
- ↵
- Родригез-Манзо Г,
- Фернандез-Густи А
(1995a) Учество на централниот норадренергичен систем во повторното воспоставување на копулаторно однесување на сексуално исцрпени стаорци од јохимбин, налоксон и 8-OH-DPAT. Мозокот Res Bull 38: 399-404.
- ↵
- Родригез-Манзо Г,
- Фернандез-Густи А
(1995b) опиоидни антагонисти и феномен на сексуална сатисфакција. Психофармацол 122: 131-136.
- ↵
- Ролс БЈ
(1986) Сензорски-специфична ситост. Nutr Rev 44: 93-101.
- ↵
- Саламон JD
(1996) Однесувањето на неврохемијата на мотивацијата: методолошки и концептуални прашања во студиите на динамичната активност на јадрото accumbens допамин. Методи на J Neurosci 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Сузуки Н,
- Shibuya A,
- Adachi H,
- Като Р,
- Цукамото Т,
- Кумамото Ј
(1995) Допамин ослободување во медијалната преоптична област за време на машкото копулаторно однесување кај стаорци. Мозокот Res 692: 66-70.
- ↵
- Schultz W
(1992) Активност на допаминските неврони во однесувањето на приматот. Semin Neurosci 4: 129-138.
- ↵
- Шимура Т,
- Јамамото Т,
- Shimokochi M
(1994) Медијалната преоптична област е вклучена и во сексуалната возбуда и кај перформансите кај машките стаорци: реевалуација на активноста на невроните кај слободно движните животни. Мозокот Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Преносот на допамин се зголемува во јадрото accumbens на машки стаорци за време на нивната прва изложеност на сексуално рецептивни женски стаорци. Мозокот Res 618: 41-46.
- ↵
- Вилсон Ц,
- Номикос Г.Г.,
- Collu M,
- Fibiger HC
(1995) Допаминергиски корелати на мотивирано однесување: важноста на возењето. Ј Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Плажа ФА
(1963) Модификација на сексуалното однесување на машките стаорци произведени со менување на стимулативната женка. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Ефект на невролептични лекови при отстранување на допамин од страна на стреатал и метаболизам во бубрежниот стаорец проучен со интрацеребрална дијализа. Eur J Pharmacol 106: 27-37.
Статии наведувајќи го овој член
- Ендогената невропластичност индуцирана од опиоиди на допаминергичните неврони во централната тегентална област влијае на природна и опијатна награда Весник на невронски мрежи, 25 јуни 2014, 34 (26): 8825-8836
- Нерамнотежа во чувствителноста на различни видови награди во патолошкото коцкање Мозокот, 1 Август 2013, 136 (8): 2527-2538
- Законот за натурални и наркотични награди за заеднички нервни пластични механизми со {Delta} FosB како главен медијатор Весник на невронски мрежи, 20 февруари 2013, 33 (8): 3434-3442
- Невронски реакции во нуклеус акумбенс школка за време на сексуално однесување кај машки стаорци Весник на невронски мрежи, 1 февруари 2012, 32 (5): 1672-1686
- Романтична љубов: систем на цицачи на мозокот за избор на партнер Филозофски трансакции на Кралското друштво Б: Биолошки науки, 29 декември 2006, 361 (1476): 2173-2186
- Хедонични топла места во мозокот Невролог, 1 декември 2006, 12 (6): 500-511
- Модулација од централна и базолатерална амигдална нуклеаза на допаминергични корелации на хранење во сотијата во стаорци нуклеус акумбенс и медијален претходна кортекс Весник на невронски мрежи, 15 декември 2002, 22 (24): 10958-10965
- Фреквенцијата на транзиентите на концентрација на допамин го зголемува дорзалниот и вентралниот стреатум на машкиот стаорци за време на воведувањето на конспектификите Весник на невронски мрежи, 1 декември 2002, 22 (23): 10477-10486
- Преглед на книга: Динамика на екстрацелуларниот допамин во акутните и хроничните дејства на кокаин Невролог, 1 Август 2002, 8 (4): 315-322
- Диференцијално вклучување на NMDA, AMPA / Kainate и допамински рецептори во јадрото на нуклеусот Accumbens во стекнувањето и изведбата на однесување на Pavlovian Approach Весник на невронски мрежи, 1 декември 2001, 21 (23): 9471-9477
- Контрола на однесување кое бара од кокаин од страна на стимулации поврзани со дроги кај стаорци: Ефекти врз обновувањето на изгаснените нивоа на допамин кои реагираат на операција и на екстрацелуларните нивоа на допамин во амигдалата и јадрото accumbens PNAS, 11 април 2000, 97 (8): 4321-4326
- Олеснување на сексуалното однесување и збогатениот ефект на допамин во нуклеус акумбенс на машки стаорци по чувствителност на однесувањето со D-амфетамин Весник на невронски мрежи, 1 јануари 1999, 19 (1): 456-463
- НЕВРОСИЦА: Привлекување внимание на мозокот Наука, 3 Октомври 1997, 278 (5335): 35-37
- Допаминергиски корелации на сензорни-специфични социети во медијалниот предреферен кортекс и нуклеус акумбенс на стаорци Весник на невронски мрежи, 1 октомври 1999, 19 (19): RC29