Neurosci Biobehav Rev. 2011 јануари; 35 (3): 939-55. doi: 10.1016/j.neubiorev.2010.10.014. Epub 2010 2 ноември.
- 1Бернард Б. Броди Одделот за невронаука, Универзитетот во Каљари, Cittadella Universitaria, 09042 Monserrato, CA, Италија. [заштитена по е-пошта]
АПСТРАКТ
Окситоцинот е моќен поттикнувач на ерекција на пенисот кога се инјектира во централниот нервен систем. Кај машките стаорци, најчувствителната област на мозокот за про-еректилниот ефект на окситоцинот е паравентрикуларното јадро на хипоталамусот. Ова јадро и околните региони ги содржат клеточните тела на сите окситоцинергични неврони кои се проектираат до екстра-хипоталамичните области на мозокот и 'рбетниот мозок. Овој преглед покажува дека окситоцинот предизвикува ерекција на пенисот, исто така, кога се инјектира во некои од овие области (на пр., вентрална тегментална област, вентрален субикулум на хипокампусот, постеромедијално кортикално јадро на амигдалата и торако-лумбалниот 'рбетниот мозок). Студиите за микроинјектирање во комбинација со интрацеребрална микродијализа и студии за двојна имунофлуоресценција сугерираат дека окситоцинот во овие области ги активира директно или индиректно (главно преку глутаминска киселина) мезолимбичните допаминергични неврони. Допаминот ослободен во јадрото accumbens за возврат ги активира нервните патишта кои водат до активирање на инцерто-хипоталамични допаминергични неврони во паравентрикуларното јадро. Ова ги активира не само окситоцинергичните неврони кои проектираат на 'рбетниот мозок и посредуваат во ерекцијата на пенисот, туку и оние кои проектираат на горенаведените екстра-хипоталамични области, модулирајќи директно или индиректно (преку глутаминска киселина) активноста на мезолимбичните допаминергични неврони кои ја контролираат мотивацијата и наградата. Заедно, овие нервни патишта може да сочинуваат сложено хипотетичко коло, кое игра улога не само во консуматорната фаза на сексуалната активност (еректилна функција и копулација), туку и во мотивационите и наградувачките аспекти на антиципиративната фаза на сексуалното однесување.
1.Воведување
Ерекцијата на пенисот е машка сексуална реакција која игра клучна улога во репродукцијата на цицачите, вклучително и човекот, и која може да се набљудува и во контексти различни од оние строго поврзани со репродукцијата. Во зависност од контекстот во кој се јавува ерекцијата на пенисот, различни централни и периферни нервни и/или хуморални механизми учествуваат во нејзината регулација (види Мејсел и Сакс, 1994; Аргиолас и Мелис, 1995, 2004, 2005; Сакс, 2000, 2007; Меккен 2000, Џулијано и Рампин, 2000 година, Андерсон, 2004 година; Меѓу централните невротрансмитери и невропептиди кои ја контролираат ерекцијата на пенисот, најпознати се допамин, серотонин, возбудливи амино киселини, азотен оксид, адренокортикотропин, окситоцин и опиоидни пептиди. Тие можат да ја олеснат или инхибираат ерекцијата на пенисот со дејствување во неколку области на мозокот, т.е. медијална преоптичка област, паравентрикуларното јадро на хипоталамусот, вентралната тегментална област, хипокампусот, амигдалата, јадрото на креветот на stria terminalis, nucleus accumbens, продолжениот мозок и 'рбетниот мозок (Табела 1) (види Meisel and Sachs, 1994; Witt and Insel, 1994; Stancampiano et al., 1994; Argiolas and Melis, 1995, 2005; Argiolas, 1999; Bancila et al., 2002, и Џулиано; Меккена, 2000 година, Хал и сор.
Окситоцинот, неврохипофизеалниот пептид добро познат по својата хормонална улога во лактацијата и породувањето, е присутен кај жените и мажите, не само во невроните со клеточни тела лоцирани во паравентрикуларните и супраоптичките јадра на хипоталамусот кои се проектираат кон неврохипофизата, туку и во невроните кои проектираат од паравентрикуларното јадро и околните структури до екстрахипоталамичните области на мозокот (т.е. септумот, вентралната тегментална област, хипокампусот, амигдалата, продолжениот мозок и 'рбетниот мозок). Се смета дека овие неврони се вклучени во бројни централни функции, како што се меморијата, учењето, афилијативното и социо-сексуалното однесување, вклучително и ерекцијата на пенисот и копулаторното однесување (види Buijs, 1978; Sofroniew, 1983; Argiolas and Gessa, 1991; Pedersen et al. ., 1992 година, Вагнер и Клеменс, 1992 година; Навистина, окситоцинот ја олеснува еректилната функција и машкото сексуално однесување кај глувците, стаорците, зајаците и мајмуните (види Argiolas и Gessa, 1993; Carter, 1995; Pedersen et al., 1997; Argiolas and Argiolas, 1998, 1999). Ова може да се случи и кај луѓето, бидејќи окситоцинот во плазмата се зголемува со сексуални стимули, особено при ејакулација (Carmichael et al., 1991; Murphy et al., 1992) и со манипулација со градите и гениталиите, кои обично се јавуваат за време на сексуалниот сексуален однос (Тиндал, 1992).
Олеснувачкиот ефект на окситоцинот врз машкото сексуално однесување првпат беше докажан со способноста на интравенозниот окситоцин да ја намали латентноста до првата ејакулација и да ја забави сексуалната исцрпеност на машките зајаци спарени со рецептивни женки (Melin и Kihlstrom, 1963). Сепак, сексуалните ефекти на окситоцинот беа дефинитивно препознаени дури во осумдесеттите. Потоа беше откриено дека окситоцинот даден централно во нанограмски количини може да предизвика ерекција на пенисот (Argiolas et al., 1985, 1986) и да го подобри копулаторното однесување (Arletti et al., 1985) кај машките стаорци и да ја зголеми лордозата кај женските стаорци (Arletti и Бертолини, 1985 година; Окситоцинот го подобрува сексуалното однесување не само кај сексуално моќните машки стаорци (Arletti et al., 1986), туку и кај постарите машки стаорци (Arletti et al., 1995), и кај доминантните, но не и кај подредените, машки верверички мајмуни (Winslow и Insel , 2004).
Про-еректилниот ефект на окситоцинот е зависен од тестостерон, бидејќи се укинува со хипофизектомија и кастрација и се обновува со дополнување со тестостерон или неговите метаболити, естрадиол и 5_-дихидро-тестостерон дадени заедно. (Melis et al., 1994a). Најчувствителната област на мозокот за индукција на ерекција на пенисот со окситоцин е паравентрикуларното јадро на хипоталамусот (Melis et al., 1986), од кое потекнуваат сите екстра-хипоталамични окситоцинергични проекции (види погоре). Овде, беше откриено дека окситоцинот може да предизвика ерекција на пенисот (и проѕевање) кога се инјектира во дози до 3 pmol (види Дел 2.1 подолу). Окситоцин индуцирана ерекција на пенисот, исто така, кога се инјектира билатерално во полето CA1 на хипокампусот, но не и во дорзалниот субикулум (види Дел 2.3 подолу), латералниот септум, каудатното јадро, медијалната преоптичка област, вентромедијалното јадро на хипоталамусот и супраоптичко јадро (Melis et al., 1986). Што се однесува до механизмот со кој окситоцинот делува во паравентрикуларното јадро за да го поттикне овој сексуален одговор, бројни студии сугерираат дека окситоцинот ги активира сопствените неврони. Во согласност со оваа хипотеза, сексуалната интеракција го зголемува FOS, генскиот производ на непосредниот ран ген c-fos во паравентрикуларните окситоцинергични неврони кои се проектираат на 'рбетниот мозок, кои се вклучени во контролата на ерекцијата на пенисот (види Witt и Insel, 1994 и референци во него), и сексуалната импотенција (на пр., неможноста на возрасен машки стаорец да се копулира со овариектомизирана естроген-прогестерон-примитирана женка) е поврзана кај машкиот стаорец со ниски нивоа на окситоцин mRNA во паравентрикуларното јадро на хипоталамусот (Arletti et. ал., 1997).
Дали окситоцинот влијае на антиципативната фаза или на консуматорната фаза на сексуалното однесување, во моментов не е јасно. Бидејќи окситоцинот предизвикува ерекција на пенисот и главниот ефект на окситоцинот врз копулаторното однесување е намалувањето на пост-ејакулаторниот интервал кај машките стаорци (Arletti et al., 1985), разумно е да се претпостави дека пептидот ја подобрува сексуалната изведба. Меѓутоа, бидејќи окситоцинот, исто така, ја зголемува социо-сексуалната интеракција (види Педерсен и сор., 1992; Картер и сор., 1997; Ајвел и Расел, 1995), а антагонистите на рецепторот на окситоцин спречуваат бесконтактни ерекции (Melis et al., 1999). се сметаат како индекс на сексуална возбуда (види Sachs, 1997, 2000, 2007; Melis et al., 1998, 1999b и референци таму), можната улога на окситоцинот во сексуалното возбудување и сексуалната мотивација не може да се исклучи.
Овој преглед ги сумира објавените и необјавените резултати од неодамнешните студии, кои покажуваат дека окситоцинот предизвикува ерекција на пенисот не само кога се инјектира во паравентрикуларното јадро на хипоталамусот, туку и во други екстра-хипоталамични области на мозокот, како што е вентралната тегментална област (Melis et al., 2007, 2009a; Succu et al., 2008), ттој вентрален субикулум на хипокампусот и задното јадро на амигдалата (Melis et al., 2009b, 2010), кои се важни конституенти на лимбичкиот систем и се смета дека играат клучна улога во процесите на мотивација и наградување. Овие студии откриваат дека окситоцинот учествува во нервните кола, кои вклучуваат други невротрансмитери, како што се допамин и глутаминска киселина, и други области на мозокот освен паравентрикуларното јадро, на пр., вентралната тегментална област, јадрото accumbens, хипокампусот и областите што допрва треба да се направат. идентификувани. Овие кола најверојатно ќе посредуваат во интеракцијата помеѓу мезолимбичниот и инцерто-хипоталамичниот допаминергичен систем и ќе играат улога не само во консуматорната фаза на машкото сексуално однесување (на пример, ерекција и копулација на пенисот), туку и во сексуалната мотивација и сексуалната возбуда, па оттука обезбедува нервен супстрат за објаснување на мотивационите и наградувачките својства на сексуалната активност.
2. Окситоцинот влијае на ерекцијата на пенисот делувајќи во различни области на мозокот
2.1. Паравентрикуларното јадро на хипоталамусот
Како што се потсетивме погоре, паравентрикуларното јадро на хипоталамусот наскоро беше идентификувано како област на мозокот најчувствителна за про-еректилниот ефект на окситоцинот. Кога се инјектира еднострано во ова јадро, окситоцинот беше пронајден активен во дози од 3 ng (3 pmol) (Melis et al., 1986). Студиите за односот структура-активност открија дека ерекцијата на пенисот индуцирана од окситоцин била посредувана од рецептори на окситоцин од типот на матката, поврзана со прилив на Ca2+ во клеточните тела на окситоцинергични неврони кои проектираат до екстра-хипоталамичните области на мозокот и со активирање на азотен оксид-интаза. Азотниот оксид, пак, делувајќи како интрацелуларен гласник со сè уште непознат механизам (не вклучува гуанилат циклаза) доведува до активирање на окситоцинергични неврони кои проектираат на 'рбетниот мозок и екстра-хипоталамичните области на мозокот, предизвикувајќи ерекција на пенисот (сл. 1) (види подолу и Argiolas и Melis, 1995, 2004, 2005 и референци во него). Способноста на окситоцинот да ги активира сопствените неврони
Сл. 1. (НЕМА) Шематски приказ на окситоцинергични неврони, кои потекнуваат од паравентрикуларното јадро на хипоталамусот и се проектираат во екстра-хипоталамичните области на мозокот, како што се 'рбетниот мозок, VTA, хипокампусот, амигдалата, итн. од овие неврони преку допамин, возбудливи амино киселини, самиот окситоцин, пептиди со аналог на хексарелин и пептиди добиени од VGF доведува до ерекција на пенисот, која може да се намали и/или да се укине со стимулација на GABAergic, опиоидните и канабиноидните CB1 рецептори. Активирањето на окситоцинергичните неврони е секундарно на активирањето на азотен оксид-синтаза присутна во овие неврони. Навистина ендогениот азотен оксид формиран со стимулација на допамин, ексцитаторни аминокиселини или рецептори на окситоцин или егзогени азотен оксид, како оној што произлегува од донаторите на азотен оксид директно во паравентрикуларното јадро, ги активира окситоцинергичните неврони со сè уште неидентификуван механизам, очигледно не поврзан со стимулација на гванилат циклаза. Ова предизвикува ослободување на окситоцин во 'рбетниот мозок и во екстра-хипоталамичните области на мозокот. Некои детали за механизмите со кои окситоцинот предизвикува ерекција на пенисот кога се ослободува во овие области, на пример, VTA, вентралниот субикулум и амигдалата се опишани во соодветните делови од областа на мозокот. Овде, окситоцинот делува на сопствените рецептори и го зголемува производството на NO, што доведува до ерекција на пенисот како што се наоѓа во PVN. Меѓутоа, во варијанса со PVN, во опашката VTA NO ја активира гванилат циклазата. Ова предизвикува зголемување на концентрацијата на cGMP што доведува до активирање на мезолимбичните допаминергични неврони и до ерекција на пенисот. Во VS NO ги активира глутаматергичните неврони кои проектираат во екстра-хипокампалните области, вклучувајќи го и VTA. Глутаминската киселина во VTA за возврат ги активира мезолимбичните допаминергични неврони како што се наоѓаат со окситоцинот. Механизмите слични на оние опишани погоре веројатно ќе функционираат и кога ерекцијата на пенисот се случува во физиолошки контекст, имено како кога машките стаорци се ставаат во присуство на недостапна приемчива женка (на пример, ерекција без контакт) или за време на копулација.
во паравентрикуларното јадро беше поддржано од студии кои покажуваат дека: (1) рецепторите на окситоцин се присутни во ова хипоталамично јадро (Freund-Mercier et al., 1987; Freund-Mercier and Stoeckel, 1995); (2) окситоцинот го олеснува сопственото ослободување ин витро и ин виво (Freund-Mercier и Richard, 1981, 1984; Moos et al., 1984); и (3) окситоцинот ги возбудува сопствените неврони делувајќи во паравентрикуларното јадро (Јамашита и сор., 1987). Покрај тоа, окситоцинергичните синапси кои напаѓаат на клеточните тела на магноцелуларните окситоцинергични неврони се исто така идентификувани во паравентрикуларното и супраоптичкото јадро на хипоталамусот (Теодос, 1985). Конечно, уништувањето на централните окситоцинергични неврони со електролитски или хемиски ексцитотоксични лезии на паравентрикуларното јадро, кое целосно ја осиромашува содржината на окситоцин низ централниот нервен систем и 'рбетниот мозок, го укинува не само про-еректилниот ефект на окситоцинот, туку и го нарушува лекот индуциран ерекција на пенисот и бесконтактни ерекции (види подолу и Argiolas et al., 1987a,b; Liu et al., 1997 и референци таму). Резултати слични на оние пронајдени со лезии на паравентрикуларното јадро се пронајдени со моќни и селективни антагонисти на рецепторот на окситоцин. Навистина, овие соединенија инјектирани во паравентрикуларното јадро во нанограмски количини ја спречија потполно окситоцин-индуцираната ерекција на пенисот, додека кога се даваа во страничните комори спречија не само ерекција на пенисот предизвикана од самиот окситоцин, туку и ерекцијата на пенисот индуцирана од лекови (види Дел 3 подолу и Argiolas и Melis, 1995, 2004, 2005 и референци таму) и бесконтактни ерекции (Melis et al., 1999a), а покрај тоа беа исклучително ефикасни во нарушувањето на копулаторното однесување на сексуално моќните машки стаорци (Argiolas et al., 1988). Понатаму, сексуалната интеракција го зголемува FOS, генскиот производ на непосредниот ран ген c-fos во паравентрикуларните окситоцинергични неврони кои проектираат на 'рбетниот мозок вклучен во контролата на ерекцијата на пенисот (види Witt и Insel, 1994 и референци таму). Конечно, сексуална импотенција (на пр., неможноста на возрасен машки стаорец да се копулира со естроген-прогестерон-примерна рецептивна женка) hкако што е исто така поврзано кај машкиот стаорец со ниски нивоа на окситоцин mRNA и на азотен-оксид синтаза во паравентрикуларното јадро на хипоталамусот (Benelli et al., 1995; Arletti et al., 1997) (за опширен преглед на овие студии види Argiolas, 1999; Argiolas и Melis, 2004, 2005).
2.2. Вентралната тегментална област
Вентралната тегментална област беше откриена неодамна како мозочно место во кое окситоцинот предизвикува ерекција на пенисот. Оваа област содржи окситоцинергични нервни завршетоци кои потекнуваат од паравентрикуларното јадро и рецепторите на окситоцин (Freund-Mercier et al., 1987; Vaccari et al., 1998). Поточно, пронајден е окситоцин способен за ипоттикнување на ерекција на пенисот кога се инјектира еднострано во каудалната, но не и во ростралната вентрална тегментална област на начин зависен од дозата (Melis et al., 2007). Активните дози беа повисоки од потребните кога се инјектираат во паравентрикуларното јадро и слични на оние кои предизвикуваат ерекција на пенисот кога се инјектираат во вентралниот субикулум на хипокампусот или во постеромедијалното кортикално јадро на амигдалата (види подолу). Очигледно, про-еректилниот ефект е посредуван од активирањето на мезолимбичните допаминергични неврони кои се проектираат на обвивката на јадрото accumbens, што за возврат ги активира сè уште непознатите нервни патишта кои проектираат до инцерто-хипоталамичните допаминергични неврони кои напаѓаат на паравентрикуларната окситоцинергична ерекција. (Melis et al., 2007, 2009a).
Што се однесува до механизмот со кој окситоцинот ја активира допаминергичната невротрансмисија во вентралната тегментална област, достапните податоци сугерираат дека окситоцинот ги стимулира окситоцинергичните рецептори лоцирани во клеточните тела на мезолимбичните допаминергични неврони. Ова го зголемува приливот на Ca2+ во клеточните тела на допаминергичните неврони, а со тоа ја активира азотен оксид-синтаза (Succu et al., 2008). Во варијанса со паравентрикуларното јадро (видете го делот 3 подолу), азотен оксид за возврат ја активира гванилат циклазата, со што ја зголемува концентрацијата на цикличниот GMP. Во согласност со овој механизам, или d(CH2)5Tyr(Me)2-Orn8-вазотоцин, моќен антагонист на окситоцин, или S-метил-тио-l-цитрулин, моќен инхибитор на невронскиот азотен оксид-синтаза, инјектиран во опашката вентрална тегментална област пред окситоцин, укината ерекција на пенисот и зголемување на екстра-клеточната концентрација на допамин во обвивката на јадрото accumbens индуцирана од окситоцин. Покрај тоа, 8-бромо-цикличниот GMP, активен цикличен GMP отпорен на фосфодиестераза, предизвикува ерекција на пенисот кога се инјектира во опашката вентрална тегментална област и ја зголемува екстрацелуларната концентрација на допамин во обвивката на јадрото accumbens, како што е откриено со инјектирање на окситоцин. опашката вентрална тегментална област (Succu et al., 2008; Melis et al., 2009a) (види, исто така, Сл. 2).
Секогаш во согласност со овој механизам, халоперидолот, моќен антагонист на рецепторот на допамин Д2, инјектиран во обвивката на јадрото accumbens ја намалува ерекцијата на пенисот индуцирана од окситоцин инјектиран во вентралната тегментална област (Melis et al., 2007). Горенаведениот механизам е поддржан и со двојни имуно-флуоресценциски студии, кои покажуваат дека влакната на окситоцин удираат на клеточните тела на допаминергичните неврони во каудалната вентрална тегментална област, кои претходно биле означени со ретроградниот трагач Флуороголд инјектиран во обвивката на јадрото (акумбен Melis et al., 2007 Succu et al., 2008). Активациите на овие допаминергични неврони и на допаминските рецептори во акумбенс јадрото, пак, доведуваат до активирање на нервните патишта кои допрва треба да се идентификуваат, кои ги стимулираат инцерто-хипоталамичните допаминергични неврони да ослободуваат допамин во паравентрикуларното јадро, а со тоа да го активираат невронски проектирање до 'рбетниот мозок и посредувачката ерекција на пенисот (види погоре и Melis et al., 2007; Succu et al., 2007, 2008). Навистина, окситоцинот инјектиран во опашката вентрална тегментална област во доза што предизвикала ерекција на пенисот, зголемена екстра-клеточна концентрација на допамин во дијализатот добиен не само од јадрото accumbens, туку и од паравентрикуларното јадро (Succu et al., 2007).
2.3. Хипокампусот
Полето CA1 на хипокампусот беше другата област на мозокот богата со окситоцинергични влакна и рецептори идентификувани од претходните студии во кои инјектирањето на окситоцин предизвикало ерекција на пенисот (види Буџис, 1978; Софронив, 1983). Сепак, во варијанса со паравентрикуларното јадро, овде беше откриено дека окситоцинот може да предизвика ерекција на пенисот само кога се инјектира билатерално и во повисоки дози од оние кои се пронајдени активни во паравентрикуларното јадро (Melis et al., 1986; Chen et al., 1992). Инјекциите на окситоцин во субикулумот беа пронајдени неактивни во овие претходни студии. Сепак, неодамнешните и повнимателни студии за микроинјектирање водат до идентификација на а регионот на вентралниот субикулум во кој инјектирањето на окситоцин беше способно да предизвика ерекција на пенисот на начин зависен од дозата (Melis et al., 2009b). Про-еректилниот ефект на окситоцинот кој се инјектира во оваа област на мозокот беше забележан во дози слични на оние кои се пронајдени активни во вентралната тегментална област по еднострано инјектирање (Melis et al., 2007), како што е откриено во паравентрикуларното јадро. Очигледно, окситоцинот инјектиран во вентралниот субикулум предизвикува ерекција на пенисот со активирање на окситоцинергични рецептори во невроните кои содржат азотен оксид-синтаза, предизвикувајќи зголемување на производството на азотен оксид. Азотниот оксид, пак, делувајќи како меѓуклеточен гласник го активира невротрансмисијата на глутаминска киселина, што доведува до ерекција на пенисот, евентуално преку нервни (глутаматергични) еферентни проекции од вентралниот субикулум до екстра-хипокампалните области на мозокот кои ја модулираат активноста на мезолимбичните допаминергични неврони (на пр., вентралната тегментална област, префронталниот кортекс, паравентрикуларното јадро) (види подолу и Melis, 2007, 2009b; Succu et al., 2008).
Овој механизам на дејство е поддржан од експерименти со интрацеребрална микродијализа, кои покажуваат дека окситоцинот инјектиран во вентралниот субикулум во дози кои предизвикуваат ерекција на пенисот, го зголемува производството на азотен оксид и концентрацијата на екстрацелуларната глутаминска киселина во дијализата од вентралниот субикулум (Melis et al. , 2010) и на екстрацелуларниот допамин во јадрото accumbens (Melis et al., 2007). Овие одговори беа антагонизирани не само од антагонистот на рецепторот на окситоцин d(CH2)5Tyr(Me)2-Orn8-вазотоцин, туку и од невронскиот азотен оксид-синтаза инхибитор S-метил-тио-лцитрулин и од азотен оксид даден хемоглобин. во вентралниот субикулум неколку минути пред окситоцинот (Melis et al., 2010).
Покрај тоа, во согласност со овој механизам на дејство, активирањето на глутаматергичната невротрансмисија со NMDA инјектирана во вентралниот субикулум предизвикува ерекција на пенисот (Melis et al., 2010). Во моментов е непознат фенотипот на еферентните проекции од вентралниот субикулум, кои предизвикуваат активирање на мезолимбичните допаминергични неврони и зголемување на екстра-клеточниот допамин во акумбенс јадрото. Сепак, бидејќи ерекцијата на пенисот индуцирана од окситоцин инјектиран во вентралниот субикулум се јавува истовремено со зголемување на екстрацелуларната глутаминска киселина во дијализатот од вентралната тегментална област, но не од јадрото accumbens и е антагонизирана со (+)MK-801, моќен неконкурентен антагонист на рецептори за возбудливи аминокиселини од подтипот NMDA (Woodruff et al., 1987), инјектиран во вентралната тегментална област, но не и во јадрото accumbens (види Сл. 2 и Melis et al., 2009b) , веројатно е дека овие проекции доведуваат до активирање на глутаматергичната невротрансмисија во вентралната тегментална област, што пак ги активира мезолимбичните допаминергични неврони кои се проектираат кон јадрото accumbens. Дали зголемената концентрација на глутаминска киселина пронајдена во вентралната тегментална област по инјектирање на окситоцин во вентралниот субикулум е ослободена од невроните кои потекнуваат од субикулумот или од други области на мозокот (на пример, префронталниот кортекс) во моментов е непознато. Сепак, ова предизвикува активирање на мезолимбичните допаминергични неврони и зголемено ослободување на допамин во акумбенс јадрото. Овде активирањето на допаминските рецептори доведува до активирање на инцерто-хипоталамични допаминергични неврони, ослободувајќи допамин во паравентрикуларното јадро, а со тоа активирајќи ги окситоцинергичните неврони кои проектираат на 'рбетниот мозок и посредуваат во ерекцијата на пенисот. (види погоре и Melis et al., 2007, 2009a; Succu et al., 2008).
2.4. Амигдалата
Амигдалата е уште една мозочна област богата со окситоцин влакна и рецептори (види Freund-Mercier et al., 1987; Vaccari et al., 1998; Uhl-Bronner et al., 2005). Окситоцинот овде се смета дека е вклучен во различни функции, од анксиолиза, социјална меморија и сознание, социјално засилено учење, емоционална емпатија, емоционална обработка на лицето и страв кај луѓето до еректилна функција и сексуално однесување (види Kondo et al., 1998; Dominguez et al. сор., 2001 г., Хубер и соработници, 2005 г.; , 2005). Сепак, способноста на окситоцинот да предизвика ерекција на пенисот кај машките стаорци кога е индуциран во постеромедијалното кортикално јадро на амигдалата беше откриена дури неодамна (Melis et al., 2007b). Овој одговор се случи истовремено со зголемување на екстра-клеточната концентрација на допамин во дијализатот добиен од обвивката на јадрото accumbens, како што е откриено по инјектирање на окситоцин во вентралниот субикулум (Melis et al., 2008b). Механизмот со кој окситоцинот се инјектира во постеромедијалното кортикално јадро на амигдалата предизвикува ерекција на пенисот во моментов е непознат. Достапните податоци покажуваат дека и ерекцијата на пенисот и зголемувањето на екстра-клеточната концентрација на допамин во дијализатот добиен од јадрото accumbens се посредувани од активирањето на окситоцинергичните рецептори, бидејќи и двата одговора беа укинати од антагонистот на рецепторот на окситоцин d(CH2009)2009Tyr( Ме) 2010-Орн2009-вазотоцин инјектиран во јадрото на амигдалата неколку минути пред окситоцинот (Melis et al., 2009b).
Без оглед на механизмот што го активира окситоцинот во постеромедијалното кортикално јадро на амигдалата, сексуалниот одговор индуциран од пептидот се укинува со блокада на сите допаминергични рецептори со цис-флупентиксол инјектиран во обвивката на јадрото accumbens и со блокада на NMDA рецепторите ( +) MK-801 инјектирано во вентралната тегментална област, но не и во јадрото accumbens, како што е откриено за ерекција на пенисот индуцирана од окситоцин инјектиран во вентралниот субикулум (Melis et al., 2009b). Ова сугерира дека окситоцинот инјектиран во постеромедијалното јадро на амигдалата ја активира невротрансмисијата на глутаминска киселина во вентралната тегментална област. Ова предизвикува активирање на мезолимбичните допаминергични неврони, што доведува до ерекција на пенисот. Со оглед на студиите кои покажуваат нервни патишта кои го поврзуваат ова јадро на амигдалата со вентралниот субикулум (Canteras et al., 1995; French and Totterdell, 2003), овие наоди ја зголемуваат можноста дека интеракција може да постои помеѓу овие две области на мозокот, иако директна опишани се патишта од амигдалата или до јадрото accumbens или до вентралната тегментална област (Kelley и Domesick, 1982; Witter, 2006).
2.5. 'Рбетниот мозок
'Рбетниот мозок е друга област на централниот нервен систем што содржи окситоцинергични влакна и рецептори (Freund-Mercier et al., 1987; Uhl-Bronner et al., 2005), во која окситоцинот предизвикува ерекција на пенисот (Tang et al., 1998 Џулијано и Рампин, 1999 година; Како што потсетивме погоре, овие окситоцинергични влакна потекнуваат од паравентрикуларното јадро на хипоталамусот и придонесуваат за опаѓачките патеки кои ги контролираат 'рбетните автономни неврони кои посредуваат во ерекцијата на пенисот. Навистина, овие влакна остваруваат синаптички контакти во дорзалниот рог, предганглионски симпатички и парасимпатички клеточни колони во торако-лумбалниот и лумбо-сакралниот тракт со 'рбетните неврони кои ги инервираат пенилните кавернозни тела (Марсон и Мекена, 1996; Џулијано и Рампин, 2000; , 2001). Овие синаптички контакти беа демонстрирани со означување на 'рбетните неврони кои потекнуваат од пенисот и допираат до' рбетниот мозок со специфични ретроградни трагачи инјектирани во кавернозните тела, комбинирани со двојна имунофлуоресценција и конфокална ласерска микроскопија (Tang et al., 1998; Veronneau; et al., 1999). Во согласност со овие студии, кај анестезирани машки стаорци интратекално инјектирање на кумулативни дози на окситоцин на лумбо-сакрално, но не и на торако-лумбално ниво, предизвиканиот интракавернозен притисок се зголемува на начин зависен од дозата. Овие ефекти беа укинати со блокада на окситоцинергичен рецептор со d(CH2)5Tyr(Me)2-Orn8-вазотоцин и со пресек на карличните нерви (Giuliano и Rampin, 2000; Giuliano et al., 2001). Овие резултати покажуваат дека окситоцинот, делувајќи на лумбо-сакралниот 'рбетниот мозок, го зголемува интракавернозниот притисок и сугерира дека окситоцинот, ослободен за време на физиолошкото активирање на паравентрикуларното јадро е моќен активатор на 'рбетните про-еректилни неврони кои се проектираат на кавернозните тела. Интересно е тоа што овие про-еректилни спинални неврони на кои окситоцинот делува за да го врши својот про-еректилен ефект, исто така добиваат синаптички контакти од серотонинергичните неврони кои потекнуваат од јадрото paragigantocellularis на ретикуларната формација на мозочната медула (Marson и McKenna, 1992; ., 1998). Уништувањето на овие серотонинергични неврони ја олеснува ејакулацијата и пенилните рефлекси кај машките стаорци (Marson и McKenna, 1992; Yells et al., 1992). Бидејќи лековите кои ги стимулираат 5HT2C рецепторите ја олеснуваат ерекцијата на пенисот кога се даваат интрацеребровентрикуларно, но не и во паравентрикуларното јадро, а лековите кои ги блокираат рецепторите на 5HT2C, исто така, го намалуваат агонистот на допаминот и ерекцијата на пенисот индуцирана од окситоцин, додека антагонистите на допаминската индукција не ја намалуваат види Stancampiano et al., 5 и референци таму), исто така е предложено дека окситоцинот го олеснува дејството на про-еректилните 2HT1994C рецептори на ниво на лумбо-сакралниот спинален мозок (Stancampiano et al., 5). Алтернативно, окситоцинот може да влијае на активноста на спинално десцендентните серотонинергични неврони со тоа што дејствува директно во јадрото paragigantocellularis, од каде што потекнуваат овие неврони (види Stancampiano et al., 1994).
3. Интеракции помеѓу окситоцин, допамин и глутаминска киселина во централниот нервен систем и ерекцијата на пенисот
Како што се потсетивме во Дел 1, сите окситоцинергични неврони присутни во централниот нервен систем потекнуваат од паравентрикуларното јадро и околните структури. Активноста на овие неврони е под контрола на различни невротрансмитери и/или невропептиди. Меѓу најпроучените на паравентрикуларно ниво се допамин, глутаминска киселина, гама-аминобутерна киселина (GABA), азотен оксид, ендоканабиноиди, опиоидни пептиди, пептиди кои ослободуваат хормон за раст, пептиди поврзани со VGF и самиот окситоцин. Допамин, глутаминска киселина, пептиди кои ослободуваат хормон за раст, пептиди добиени од VGF и окситоцин се стимулирачки, на пр., овие соединенија и нивните агонисти ја олеснуваат ерекцијата на пенисот кога се инјектираат во паравентрикуларното јадро, додека GABA, опиоидните пептиди и ендоканабиноидите се инхибитни соединенија, или нивните агонисти ја инхибираат ерекцијата на пенисот (види Мејзел и Сакс, 1994; Вит и Инсел, 1994; Аргиолас и Мелис, 1995, 2004, 2005; Џулијано и Рампин, 2000, 2004; Мекена, 2000; Андерсон, 2001, ал., 2002).
Неколку линии на експериментални докази сугерираат дека овие окситоцинергични неврони и горенаведените невротрансмитери и невропептиди се вклучени во контролата на еректилната функција и сексуалното однесување во различни физиолошки контексти. Покрај тоа, окситоцинот ослободен во екстра-хипоталамичните области на мозокот, како што се вентралната тегментална област, хипокампусот и неговите региони, амигдалата и 'рбетниот мозок може да влијае на активноста на оние неврони на кои удираат окситоцинергичните синапси. Во моментот, единствените неврони важни за ерекцијата на пенисот на кои удираат окситоцинергичните синапси, идентификувани со сигурност, се клеточните тела на мезолимбичните допаминергични неврони на каудалната вентрална тегментална област што се проектираат на обвивката на јадрото accumbens (Melis et al., 2007 Succu et al., 2008), и про-еректилните 'рбетни неврони кои се проектираат од лумбо-сакралниот тракт до кавернозните тела (види Giuliano и Rampin, 2000; Giuliano et al., 2001) (види, исто така, Секции 2.2 и 2.5) . Навистина, иако окситоцинергични синапси и рецептори се исто така идентификувани во вентралниот субикулум, амигдалата и 'рбетниот мозок, области кои се сите важни за ерекцијата на пенисот (види погоре), во овие области типот на невротрансмитер/и присутни во невроните на кои окситоцинергични нервни завршетоци се удираат, сè уште се непознати.
Овој дел од прегледот накратко ја сумира неодамнешната литература за механизмите кои лежат во основата на про-еректилниот ефект на окситоцин инјектиран во каудалната вентрална тегментална област, вентралниот субикулум на хипокампусот и во 'рбетниот мозок. Посебно внимание се посветува на интеракцијата на пептидот со допамин и глутаминска киселина во овие области и на улогата што оваа интеракција може да ја игра во централната контрола на еректилната функција. Прво е дадено кратко резиме на ефектите на допаминот и глутаминската киселина врз окситоцинергичните неврони во паравентрикуларното јадро, кои исто така играат клучна улога во еректилната функција, со цел читателот да биде свесен за раната состојба на истражување во оваа област, како овие студии се веќе опширно разгледани (види Argiolas and Melis, 1995, 2004, 2005; Melis and Argiolas, 2003). Исто така, во овој случај, особено внимание се посветува на најновите резултати, кои укажуваат на важна улога и за врската допамин-окситоцин и врската глутаминска киселина-окситоцин не само во сексуалните перформанси (ерекција и копулација на пенисот), туку и во сексуалната возбуда и сексуална мотивација.
3.1. Интеракција на допамин-окситоцин во паравентрикуларното јадро
на способноста на допаминските агонисти да индуцираат ерекција на пенисот преку активирање на централните окситоцинергични неврони беше предложено веднаш по откритието дека апоморфинот индуцира ерекција на пенисот кога се инјектира во паравентрикуларното јадро (Melis et al., 1987) кога беше пронајден антагонистот на рецепторот на окситоцин d(CH2)5Tyr(Me)2-Orn8-вазотоцин интрацеребровентрикуларно способна речиси целосно да ја намали ерекцијата на пенисот предизвикана не само од окситоцин даден icv, туку и од апоморфин, даден субкутано (Argiolas et al., 1987b). Овие резултати беа проследени со оние од други студии кои покажуваат слични резултати кога d(CH2)5Tyr(Me)2-Orn8-вазотоцин беше даден icv и апоморфин беше даден директно во паравентрикуларното јадро (Melis et al., 1989b), lЕдинг сугерира дека агонистите на допамин предизвикуваат ерекција на пенисот со активирање на паравентрикуларните окситоцинергични неврони кои проектираат во екстра-хипоталамичните области на мозокот, а особено на 'рбетниот мозок (види Argiolas и Melis, 1995, 2004, 2005). Во согласност со оваа хипотеза, кај анестезирани стаорци, неодамна беше откриена блокада на лумбо-сакралните окситоцинергични рецептори од не-пептиден антагонист на рецепторот на окситоцин, способна да го укине зголемувањето на интракавернозниот притисок предизвикано од апоморфин, индуцирано од допаминскиот агонист. паравентрикуло-спинален окситоцинергичен пат вклучен во ерекцијата на пенисот (Baskerville et al., 2009).
Студиите насочени кон идентификација на допаминскиот рецептор одговорен за индукција на ерекција на пенисот, откри дека, исто така, во паравентрикуларното јадро на допаминските рецептори агонистите индуцираат ерекција на пенисот делувајќи на допаминските рецептори од семејството D2, како што е откриено со агонисти на допаминските рецептори дадени системски (види Melis et al., 1987; Eaton et al., 1991; Melis and Argiolas, 1995a). Соодветно на тоа, апоморфинот, моќен мешан D1/D2 рецептор агонист, и хинпирол, моќен селективен D2 рецептор агонист, но не и SKF 38393, селективен агонист на рецепторот D1, инјектиран во ова јадро на хипоталамусот, беа пронајдени способни да предизвикаат ерекција зависна од пенисот. , а сексуалниот одговор индуциран од овие агонисти на рецепторот D2 беше укинат од антагонисти на рецепторот D2, како што се халоперидол и l-сулпирид, но не и од SCH 23390, селективен антагонист на рецепторот D1 (Melis et al., 1987). Способноста на апоморфинот да предизвика ерекција на пенисот кога се инјектира во паравентрикуларното јадро беше потврдена и со телеметриски студии кои покажуваат дека агонистот на допамин даден во паравентрикуларното јадро може да го зголеми интракавернозниот притисок кај будните машки стаорци без да го модифицира системскиот крвен притисок (Chen et al. , 1999 Џулијано и Алард, 2001 година), како што е откриено по системска инјекција (Bernabè et al., 1999). Овие студии исто така ја потврди главната улога на D2 рецепторите, бидејќи агонистите на D1 рецепторите обично не се во можност да го зголемат интракавернозниот притисок кога се инјектираат во паравентрикуларното јадро (Чен и сор., 1999).
Тогаш беа достапни неколку линии на експериментални докази кои сугерираат дека паравентрикуларните D2 рецептори, чија стимулација предизвикува ерекција на пенисот, се наоѓаат на клеточните тела на окситоцинергичните неврони. Прво, паравентрикуларното јадро содржи допаминергични нервни терминали кои припаѓаат на таканаречените инцертохипоталамични допаминергични неврони. Клеточните тела на овие неврони се сместени во групата А13 и А14 на Dahlstrom и Fuxe (1964), интензивно арборизираат и инервираат неколку хипоталамични структури, вклучувајќи паравентрикуларни окситоцинергични неврони кои проектираат кон неврохипофизата и/или до екстра-хипоталамичните области на мозокот (Buijs et al., 1984; Lindvall et al., 1984).
Вклучувањето на овие допаминергични неврони на паравентрикуларно ниво во контролата на ерекцијата и копулацијата на пенисот е поддржано од студии за микродијализа кои покажуваат дека концентрациите на екстрацелуларниот допамин и 3,4-дихидроксифенилацетна киселина (DOPAC), нејзиниот главен метаболит, се зголемени во дијализатот. добиено од паравентрикуларното јадро на сексуално потентни машки стаорци кои покажуваат бесконтактна ерекција кога се ставаат во присуство на недостапен овариектомизиран естроген + рецептивна женка подготвена со прогестерон (Melis et al., 2003).
Зголемувањето на концентрациите на допамин и DOPAC беше уште поголемо кога беше дозволена копулација со рецептивната женка (Melis et al., 2003), како што е откриено во медијалната преоптичка област (Hull et al., 1995) и во јадрото accumbens (Pfaus и Еверит, 1995). Второ, неколку студии покажуваат дека ерекцијата на пенисот предизвикана од стимулацијата на паравентрикуларните D2 рецептори, е посредувана од окситоцинот ослободен во овие области. Соодветно, беше откриено дека апоморфинот даден во дози кои предизвикуваат ерекција на пенисот може да ја зголеми концентрацијата на окситоцин, не само во плазмата на стаорци и мајмуни (Melis et al., 1989a; Cameron et al., 1992), туку и во екстра-хипоталамичниот мозок области, како што е хипокампусот (Melis et al., 1990). Во согласност со овие резултати, апоморфинот инјектиран во паравентрикуларното јадро во доза што предизвикува ерекција на пенисот неодамна се покажа дека може да ја зголеми, исто така, екстрацелуларната концентрација на допамин во акумбенс јадрото, ефект намален од антагонистот на рецепторот на окситоцин d(CH2) 5Tyr(Me)2-Orn8-вазотоцин инјектиран во вентралната тегментална област (Succu et al., 2007; Melis et al., 2009a) (види исто така Дел 4). Трето, билатерални електролитски лезии на паравентрикуларното јадро, кои речиси целосно го елиминираат окситоцинот од екстра-хипоталамичните области на мозокот (Hawthorn et al., 1985), ја укинуваат ерекцијата на пенисот индуцирана од апоморфин (Argiolas et al., 1987a) и селективните оксиагонисти дадени во страничните комори, но не и во паравентрикуларното јадро, ја намалуваат дозно-зависната апоморфин-индуцирана ерекција на пенисот со моќ паралелна на онаа на овие соединенија во блокирањето на рецепторите на окситоцин (Melis et al., 1989b). Антагонистите на рецепторот на окситоцин се исто така исклучително моќни во намалувањето на олеснувањето на машкото сексуално однесување индуцирано не само од окситоцинот, туку и од апоморфинот (Argiolas et al., 1988, 1989).
Што се однесува до механизмот со кој D2 рецепторите активирани од допамин или од агонисти на допаминските рецептори, ја зголемуваат активноста на окситоцинергичните неврони, со што се ослободува окситоцин во екстрахипоталамичните области на мозокот и во 'рбетниот мозок, бројни експериментални податоција поддржуваат хипотезата дека стимулацијата на D2 рецепторите ја зголемува концентрацијата на интрацелуларните Ca2+ јони во клеточните тела на окситоцинергичните неврони, што доведува до активирање на азотен оксид-синтаза, ензим зависен од Ca2+-калмодулин, кој е присутен во овие клеточни тела (Vincent и Kimura, 1992; Torres et al., 1993; Sanchez et al., 1994; Sato-Suzuki et al., 1998). Зголеменото производство на азотен оксид предизвикува активирање на окситоцинергични неврони. Соодветно, (1) ерекцијата на пенисот предизвикана од апоморфин беше спречена со органски блокатори на калциумови канали и со_-конотоксин GVIA, моќен и селективен блокатор на напонски зависните Ca2+ канали од N-тип (McCleskey et al., 1987), даден во паравентрикуларната јадро (види Argiolas et al., 1990, и референци таму); (2) ерекцијата на пенисот индуцирана од апоморфин беше спречена со инхибитори на азотен оксид-синтаза дадени во паравентрикуларното јадро (Melis et al., 1994c); и (3) апоморфин и други агонисти на рецепторот D2 дадени во дози кои индуцираат ерекција на пенисот зголемено производство на азотен оксид во паравентрикуларната дијализа добиена со интрацеребрална микродијализа, зголемување што беше намалено со инхибитори на паравентрикуларен азотен оксид-синтаза дадени во дози кои намалуваат Ерекција на пенисот индуцирана од агонист на D2 рецепторот (Melis et al., 1996). Механизмот со кој азотен оксид ги активира паравентрикуларните окситоцинергични неврони сè уште е непознат, иако достапните податоци сугерираат дека азотен оксид делува како интрацелуларен гласник и дека гванилат циклазата не е вклучена. Навистина, активниот фосфодиестеразарезистентен аналог на цикличниот GMP, 8-бромо-цикличниот GMP, беше откриен дека не може да предизвика ерекција на пенисот кога се дава во паравентрикуларното јадро (сл. 2) (види Мелис и Аргиолас, 1995б и референци во него).
Горенаведеното толкување често се сметаше за неубедливо, главно затоа што стимулацијата на рецепторите на допамин D2 обично е поврзана со инхибиција, а не со побудување на клеточните тела на невроните кои ги содржат овие рецептори преку различни механизми поврзани со Г-протеинот. (види Соколоф и Шварц, 1995). Сепак, можно објаснување за ова несовпаѓање, кое е во согласност со директната стимулација на паравентрикуларните окситоцинергични неврони од допамин, неодамна беше предложено со откривањето на Допамин Д4 поврзан со G протеин рецептор, член на семејството на D2 рецептори (D2, D3 и D4), чија стимулација го зголемува приливот на Ca2+ во клеточните препарати кои содржат клонирана верзија на овој подтип на рецептори (Moreland et al., 2004). Уште поважно, селективен агонист на рецепторот D4 (на пример, ABT 724) (N-метил-4-(2-цијанофенил)пиперазинил-3метилбензамид малеат) беше пронајден способен да предизвика ерекција на пенисот кај машки стаорци кога се дава системски (Brioni et al., 2004). Овој ефект не беше пронајден со селективниот подтип на D2 рецептор агонист PNU-95666E (R-5,6-дихидро-N,N-диметил-4Химидазо[4,5,1-i]хинолин-5-амин) (Hsieh et al ., 2004), кој исто така не можеше да го зголеми приливот на Ca2+ во клеточните препарати што ја содржат клонираната верзија на подтипот на рецепторот D4 (Brioni et al., 2004; Moreland et al., 2004). Во согласност со горната хипотеза и наоди, PD 168,077 (N-метил-4-(2-цијанофенил)пиперазинил-3метилбензамид малеат), PIP-3EA (2-[4-(2-метоксифенил) пиперазин-1-илметил]имидазо [1,2-a]пиридин) и други селективни агонисти на рецепторот D4 (Heier et al., 1997; Melis et al., 2006b; Löber et al., 2009), беше откриено дека може да предизвикаат ерекција на пенисот кога се инјектираат системски, icv и во паравентрикуларното јадро, иако помалку ефикасно од апоморфинот. Про-еректилниот ефект на овие агонисти на рецепторот D4 беше спречен со L-745,870 (3-(4-[хлорофенил] пиперазин-1-ил)-метил-1H-пироло[2,3-B]пиридин трихидрохлорид), селективен Антагонист на D4 рецепторот (Patel et al., 1997; Melis et al., 2005, 2006b; Löber et al., 2009).
Конечно, про-еректилниот ефект на горенаведените агонисти на рецепторот D4 исто така беше намален со инхибитори на азотен оксид-синтаза, дадени во паравентрикуларното јадро и со d(CH2)5Tyr(Me)2-Orn8-вазотоцин, селективен антагонист на рецепторот на окситоцин дадена icv, но не и во паравентрикуларното јадро. Овие резултати се во согласност со хипотезата дека агонистите на рецепторот D4, исто така, стимулираат окситоцинергични неврони преку активирање на азотен оксид-синтаза и ослободување на окситоцин во екстрахипоталамичните области на мозокот, што пак ја олеснува ерекцијата на пенисот, како што е прикажано за апоморфин и класичните D2 агонисти (Meliset ., 2005, 2006б., Löber et al., 2009).
Горенаведените наоди, исто така, ја поддржуваат хипотезата дека допаминот предизвикува ерекција на пенисот со дејство на рецепторите D4 лоцирани на клеточните тела на паравентрикуларните окситоцинергични неврони и кои предизвикуваат зголемен прилив на Ca2+ во клеточните тела на окситоцинергични неврони, што доведува до зголемено производство на азотен оксид. Азотниот оксид за возврат ги активира окситоцинергичните неврони за ослободување на окситоцин во екстра-хипоталамичните области на мозокот и во 'рбетниот мозок, како што веќе беше дискутирано. Во таа смисла, Вреди да се забележи дека допаминските рецептори се идентификувани во клеточните тела на окситоцинергичните неврони во паравентрикуларното јадро само неодамна со двојни имунофлуоресцентни студии со високо селективни антитела на рецепторот D2, D3 и D4 и со антитела на окситоцин. Овие студии покажаа изразување на сите три подтипови на D2 рецептори (D2, D3 и D4), кои се ко-локализирани одделно во клеточните тела на окситоцинергични неврони во паравентрикуларното јадро (и исто така во супраоптичкото јадро и медијалната преоптичка област) (Баскервил и Даглас, 2008 година;
Ова обезбедува силна невроанатомска поддршка на можноста агонистите на допамин и допамински рецептори од типот D2 да индуцираат ерекција на пенисот преку активирање директно окситоцинергични неврони кои проектираат на екстрахипоталамичните области на мозокот споменати погоре., на пр., 'рбетниот мозок, вентралната тегментална област, хипокампусот и амигдалата. Сепак, овие наоди не даваат никаква помош за идентификација на подтипот/ите на рецепторот D2, чија стимулација предизвикува еректилен одговор. За жал, не може да се добие помош дури и од студии насочени кон идентификување на окситоцинергични неврони активирани од агонисти на допаминските рецептори во паравентрикуларното јадро. Навистина, и покрај различната активност на различните подтипови на допаминските рецептори, или мешаните агонисти на допаминските рецептори (на пример, апоморфин), или селективни агонисти на рецепторот D2 (на пример, хинпирол, кој делува на сите подтипови на рецепторот Д2) или селективни агонисти на рецепторот Д4 предизвикуваат активирањето на окситоцинергичните неврони, мерено со зголемувањето на FOS протеинот во парвоцелуларните окситоцинергични неврони на паравентрикуларното јадро (Bitner et al., 2006). Сепак, овој наод неодамна беше доведен во прашање, бидејќи зголемувањето на FOS протеинот во паравентрикуларните окситоцинергични неврони беше пронајдено само кога ерекцијата на пенисот беше индуцирана од кинеролан, кој делува главно на подтиповите на D2 и D3 рецептори, но не и од PD 168077, агонист на D4 рецепторот, и покрај способноста на двете соединенија да индуцираат сексуален одговор (Baskerville et al., 2009).
Потоа се неопходни понатамошни експерименти со селективни агонисти на другите подтипови на рецепторот D2 (главно D2 и D3) за да се идентификува точната улога на секој подтип на допамин рецептор во контролата на еректилната функција на паравентрикуларно ниво. Во овој поглед, како што веќе беше споменато претходно, апоморфинот, кој силно делува на сите подтипови на допамински рецептори (види Brioni et al., 2004, и референци во него), е многу поефикасен од агонистите на D4 рецептори во индуцирање на ерекција на пенисот кога се инјектира во паравентрикуларната јадро. Ова може да се објасни со повисок афинитет на апоморфин на D4 рецепторите во споредба со оној на тестираните агонисти на рецепторот D4, или алтернативно, агонистите на рецепторот D4 може да дејствуваат како парцијални агонисти на рецепторот D4, или истовремената активација на различни подтипови на допамински рецептори со апоморфин може да произведуваат поголема активација на окситоцинергични неврони кои посредуваат во ерекцијата на пенисот, отколку активирањето од страна на агонисти на рецепторот D2 само на подтипот на рецепторот D4.
Интеракциите помеѓу рецепторите на допамин D1 и D2 се веќе опишани во контролата на сексуалното однесување на ниво на медијалната преоптичка област (види Hull et al., 1989). Во случај дека неможноста на селективни агонисти на рецепторот D2 да предизвикаат ерекција на пенисот (Hsieh et al., 2004) ќе се потврди (но види Depoortère et al., 2009), на пример, дури и по инјектирање на овие соединенија во паравентрикуларното јадро , главната улога за D3 рецепторите сами или заедно со онаа на D4 рецепторите во активирањето на окситоцинергичните неврони кои посредуваат во ерекцијата на пенисот треба детално да се анализира (види Baskerville et al., 2009). За жал, селективни агонисти на D2 и D3 рецептори (на пр., кои се разликуваат во нивниот афинитет за овие два рецепторски подтипови за најмалку четири/пет реда на големина ин витро) не се достапни во моментот. Поради оваа причина, неодамнешната сугестија дека D3 рецепторите посредуваат во ерекцијата на пенисот индуцирана од класичните агонисти на рецепторот D2, која главно се заснова на способноста на наводните антагонисти на рецепторот D3, карактеристични во in vitro експериментите, да ја намалат ерекцијата на пенисот индуцирана од класичните D2 агонисти, како на пр. апоморфин, хинпирол и прамипексол, кои силно ги активираат сите подтипови на рецептори за допамин Д2 (Колинс и сор., 2009), секако треба да се потврдат со други експерименти. Оваа валидација е неопходна и затоа што во оваа студија не беше пронајден ефект на агонистите на рецепторот D4 врз ерекцијата на пенисот, што е зачудувачки контраст со резултатите од студиите цитирани погоре, кои покажуваат про-еректилен ефект на агонисти на рецепторот D4. Навистина, дури и способноста на апоморфинот да предизвика ерекција на пенисот (и проѕевање) кај глувците со нокаут на рецепторот D4 со моќ идентична на онаа забележана кај глувците со нокаут од дивиот тип на рецепторот D4 и способноста на антагонистите на рецепторот D3 да го укинат одговорот на апоморфин кај овие животни. Collins et al., 2009) не може да се смета како дефинитивен доказ за селективна улога на подтипот на рецепторот D3 во ерекцијата на пенисот индуцирана од агонистот на рецепторот D2. Освен различните видови, студиите со невротрансмитер/невропептид и/или генска аблација на невротрансмитер/невропептиден рецептор (невротрансмитер/невропептид и/или невротрансмитер/невропептиден рецептор нокаут животни) обично додадоа дополнителна конфузија и компликации во потврдувањето на наводната сексуална улога на невротрансмитери и / или невропептиди и нивните рецептори. Имено, аблацијата на генот на окситоцин произведува окситоцин нокаутирани глувци кои се парат и се копулираат нормално, како окситоцинот да е непотребен за парење и копулација. Исто така, хомозиготните женски нокаут глувци со окситоцин покажуваат нормално парење и породување, иако со значително нарушување на слабеењето на млекото (Nishimori et al., 1996; Young et al., 1996). Аблацијата на генот што ја кодира невронската азотен оксид синтаза, исто така, произведува глувци со нокаутирање на азотен оксид синтаза кои се парат и нормално се копулираат (Хуанг и сор., 1993). Сепак, овие наоди веројатно укажуваат на важна карактеристика на репродуктивната физиологија, т.е. вишок на системи вклучени во нејзината контрола на централно и периферно ниво.
Таквата вишок секако има еволутивно потекло, бидејќи гарантира премин на гените до следната генерација за опстанок на видот. Затоа, фактот дека аблацијата на генот на рецепторот D4 не го менува про-еректилниот ефект на апоморфинот, сугерира дека рецепторите D4, како окситоцинот и азотен оксид, се само неколку од посредниците кои работат во системите кои ја контролираат еректилната функција, наместо што сугерира дека нема улога на овие рецептори во контролата на ерекцијата на пенисот и сексуалното однесување. Неуспехот на D4 агонистите да предизвикаат ерекција на пенисот кога се даваат системски на машки стаорци од различни видови неодамна беше пријавен во друга студија (Depoortère et al., 2009). Сепак, за разлика од работата на Колинс и сор. (2009), и за да ја направи сликата за улогата на различните подтипови на рецепторите Д2 во контролата на ерекцијата на пенисот, оваа студија исто така покажува дека наводните селективни антагонисти на рецепторот Д3 дадени системски не биле во можност да ја намалат ерекцијата на пенисот предизвикана од апоморфин кај машките стаорци од сојот почувствителни на про-еректилниот ефект на апоморфинот, додека сексуалниот одговор (и проѕевањето) беше антагонизиран од селективниот D2 антагонист L-741,626 (3-[[4-(4-хлорофенил)-4-хидроксипиперидин -1-ил]метил-1H-индол), што ги наведува авторите да сугерираат дека D2 рецепторите, наместо D3 и D4 рецепторите, се оние кои играат главна улога во ерекцијата на пенисот индуцирана од D2 агонист (Depoortère et al., 2009). Конечно, можноста дека возбудливиот ефект на агонистите на допаминските рецептори врз окситоцинергичните неврони кои посредуваат во ерекцијата на пенисот, барем делумно, е индиректен наместо директен, на пример, посредуван или под влијание на промените во активноста на другите невротрансмитери невропептиди кои можат да ја модулираат активноста окситоцинергичните неврони во паравентрикуларното јадро не можат целосно да се исклучат.
3.2. Интеракција на глутаминска киселина-окситоцин во паравентрикуларното јадро
Паравентрикуларното јадро на хипоталамусот е многу богато со синапси кои содржат возбудлива амино киселина како невротрансмитер (на пример, глутаминска киселина и аспарагинска киселина) (Ван Ден Пол, 1991). Возбудувачките амино киселини во ова јадро се вклучени во бројни функции, вклучувајќи ја ерекцијата на пенисот и сексуалното однесување (Роелинг и сор., 1991; Мелис и сор., 1994б, 2000, 2004б). Според тоа, N-метил-d-аспарагинската киселина (NMDA), селективен агонист на подтипот на рецепторот NMDA, но не (±)-_-амино-3-хидрокси-5-метил-изоксазол-4-пропионска киселина (AMPA) , селективен агонист од подтипот на рецепторот AMPA или (±)-транс(1)-амино-1,3-циклопентан дикарбоксилна киселина (ACPD), селективен агонист на подтипот на метаботропните рецептори, беше пронајден способен да предизвика ерекција на пенисот кога се инјектира во паравентрикуларното јадро на стаорци кои слободно се движат (Melis et al., 1994b). Про-еректилниот ефект на 948 MR Melis, A. Argiolas / Neuroscience and Biobehavioral Reviews 35 (2011) 939-955 NMDA беше спречен од (+)MK-801, неконкурентен антагонист на NMDA рецепторот (Woodruff et al. ), инјектирано во паравентрикуларното јадро (Melis et al., 1987b). Во согласност со овие резултати, во телеметриските студии насочени кон следење на интракавернозниот притисок, NMDA беше откриено многу поактивна од агонистите на другите подтипови на рецептори за возбудливи аминокиселини кога се инјектираат во паравентрикуларното јадро при зголемен интракавернозален притисок кај будни или анестезирани машки стаорци (Zahran et ал., 1994 година, Чен и Чанг, 2000 година).
Како што е предложено погоре за окситоцинот и допаминот, веројатно е дека NMDA рецепторите кои посредуваат во ерекцијата на пенисот се лоцирани во клеточните тела на окситоцинергичните неврони, бидејќи нервните завршетоци на возбудливи аминокиселини удираат на телата на окситоцинергичните клетки во паравентрикуларното јадро. (Ван Ден Пол, 1991). Во аналогија на она што е откриено со агонисти на допаминските рецептори, про-еректилниот ефект на NMDA очигледно е посредуван од активирањето на окситоцинергичната невротрансмисија, укината од антагонистот на окситоцин d(CH2)5Tyr(Me)2-Orn8-вазотоцин, даден icv, но не во паравентрикуларното јадро (види Argiolas и Melis, 1995, 2004, 2005 и референци во него). Исто така, активирањето на окситоцинергичната невротрансмисија предизвикано од NMDA е исто така секундарно на активирањето на азотен оксид-синтаза, бидејќи ерекцијата на пенисот индуцирана од NMDA е спречена со инхибитори на азотен оксид-синтаза (N-Nitro-N-метил-аргинин метилстер метил-тио-l-цитрулин) даден во паравентрикуларното јадро, и NMDA инјектирана во паравентрикуларното јадро во дози кои индуцираат ерекција на пенисот, го зголемува производството на азотен оксид во хипоталамичното јадро (види Argiolas и Melis, 1995, 2004, 2005 и референци таму ). Што се однесува до агонистите на допаминските рецептори, активирањето на азотен оксидсинтаза предизвикано од NMDA може да биде исто така секундарно на зголемениот прилив на Ca2+ во окситоцинергични клеточни тела преку Ca2+ каналот споени NMDA рецептори, како што е прикажано во неколку нервни препарати (за преглед види Снајдер, 1992; Саутам и Гартвејт, 1993 година, Шуман и Медисон, 1994 година и референци во него). Азотниот оксид за возврат ја активира окситоцинергичната невротрансмисија (види погоре). Потеклото на глутаматергичните проекции кои ги активираат паравентрикуларните окситоцинергични неврони кои посредуваат во ерекцијата на пенисот е непознато, иако некои невроанатомски и електрофизиолошки докази сугерираат дека тие може да потекнуваат, барем делумно, од хипокампусот (Saphier и Feldman, 1987; Chen et al., 1992). . Иако е неопходна понатамошна работа за подобро да се карактеризира потеклото на глутаматергичните проекции на паравентрикуларното јадро, вклученоста на глутаминската киселина во паравентрикуларното јадро во контролата на ерекцијата на пенисот и сексуалното однесување е јасно поддржана од студиите за микродијализа. Соодветно на тоа, екстра-клеточните концентрации на глутаминска киселина и аспарагинска киселина беа зголемени во дијализатот добиен од паравентрикуларното јадро на машки стаорци кои покажуваат бесконтактни ерекции кога се ставени во присуство на непристапни приемчиви женски стаорци со естроген + прогестерон (Melis et al., 2004б), пенилните ерекции кои се исто така посредувани од активирањето на централниот окситоцинергичен пренос (Melis et al., 1999a,b). Ваквите зголемувања беа откриени уште повисоки кога беше дозволена копулација со рецептивната женка (Melis et al., 2004a). Во согласност со хипотезата дека зголемена активност на возбудливите аминокиселини се јавува во паравентрикуларното јадро за време на ерекцијата и копулацијата на пенисот, и бесконтактните ерекции и копулаторното однесување (за време на кое во копула се јавуваат ерекции на пенисот) се намалуваат со блокада на NMDA рецепторите во паравентрикуларното јадро. , и ова намалување е проследено со намалување на зголемувањето на производството на азотен оксид што се јавува во ова хипоталамично јадро во овие физиолошки контексти (Melis et al., 2000). Зголемување на концентрацијата на екстрацелуларната глутаминска киселина секундарно на намаленото ослободување на ГАБА од ГАБА-ергичните нервни завршетоци кои напаѓаат на возбудливите аминокиселински синапси спротиставени на телата на окситоцинергичните клетки, исто така беше откриено во паравентрикуларното јадро по блокадата на канабиноидните CB1 рецептори од страна на агонистот CB1R141761, CB2006, се дава во страничните комори или директно во паравентрикуларното јадро во дози кои индуцираат ерекција на пенисот (види Succu et al., 2007; Castelli et al., XNUMX). Ваквото зголемување доведе до активирање на азотен оксид-синтаза во клеточните тела на окситоцинергични неврони, зголемувајќи го производството на азотен оксид. Азотниот оксид за возврат ги активира окситоцинергичните неврони кои посредуваат во ерекцијата на пенисот како што е опишано погоре. Во согласност со таквиот механизам, ерекцијата на пенисот индуцирана од SR 141761A беше намалена со блокада на NMDA рецептори и со инхибитори на азотен оксид-синтаза, но не и со блокада на допамин или окситоцин рецептори во паравентрикуларното јадро, додека беше спречена со блокада на централни окситоцин рецептори од антагонисти на окситоцин рецептори дадени icv
3.3. Интеракција на окситоцин-допамин во вентралната тегментална област
Окситоцинот предизвикува ерекција на пенисот кога се инјектира во каудалниот дел од вентралната тегментална област на начин зависен од дозата (Melis et al., 2007). Најниската активна доза која се инјектира еднострано беше 20 ng, додека највисоката тестирана доза беше 100 ng. Ефектот на окситоцин е посредуван со активирање на окситоцинергични рецептори, бидејќи сексуалниот одговор се укинува со претходно инјектирање на антагонистот на окситоцин d(CH2)5Tyr(Me)2-Orn8-вазотоцин во каудалната вентрална тегментална област. Овие рецептори се локализирани во клеточните тела на допаминергичните неврони, кои се проектираат главно на обвивката на јадрото accumbens. Според тоа, (1) двојните имунофлуоресцентни студии покажуваат дека во опашката вентрална тегментална област окситоцинергичните влакна се во близок контакт со клеточните тела на допаминергичните неврони, од кои повеќето биле позитивно означени за тирозин-хидроксилаза и претходно го содржат ретроградниот трагач Флуороголд инјектирано во обвивката на nucleus accumbens (Melis et al., 2007) и (2) вентралната тегментална област индуцирана од окситоцин ерекција на пенисот се јавува истовремено со зголемување на концентрацијата на екстра-клеточниот допамин во дијализатот добиен од обвивката на јадрото accumbens (Melis et al., 2007). Ерекцијата на пенисот индуцирана од окситоцин, исто така, се јавува истовремено со зголемување на производството на азотен оксид во вентралната тегментална област, што е и двата одговора антагонизирани не само со d(CH2)5Tyr(Me)2-Orn8-вазотоцин и од инхибиторот на синтаза на азотен оксид S- метил-тио-l-цитрулин, но исто така и од _-конотоксин, блокатор на Ca2+ канали зависен од напон и со ODQ (1H-[1,2,4]оксадиазол[4,3-a]хиноксалин-1-он) , моќен инхибитор на гванилат циклаза, сите дадени во каудалната вентрална тегментална област пред окситоцинот (Succu et al., 2008). Како што многу од допаминергични клеточни тела означени со Флуороголд контактирани со окситоцинергични влакна, за кои е откриено дека се позитивни за тирозин хидроксилаза во каудалната вентрална тегментална област, исто така биле позитивно означени за азотен оксид-синтаза и гванилат циклаза (Succu 2008,). -индуцираната ерекција на пенисот може да биде посредувана од следниот механизам. Активирањето на окситоцинергичните рецептори во допаминергичните клеточни тела од страна на пептидот го зголемува приливот на Ca2+ во клеточните тела на допаминергичните неврони. Ова ја активира азотен оксид-синтаза, ензим зависен од Ca2+-калмодулин, а со тоа го зголемува производството на азотен оксид. Азотниот оксид за возврат ја активира гванилат циклазата, што доведува до зголемена концентрација на цикличниот GMP. Цикличниот GMP ги активира допаминергичните неврони кои се проектираат на јадрото accumbens. Улогата на цикличниот GMP во ерекцијата на пенисот, индуцирана од окситоцинот инјектиран во каудалната вентрална тегментална област, исто така е поддржана од способноста на 8-бромо-цикличниот GMP, активен аналог на цикличниот GMP отпорен на фосфодиестераза, да индуцира ерекција на пенисот кога се инјектира во опашката вентрална тегментална област и да се зголеми екстра-клеточниот допамин во дијализатот од јадрото accumbens (Succu et al., 2008; Melis et al., 2009a). Ова е во спротивност со механизмот со кој азотен оксид ги активира окситоцинергичните неврони во паравентрикуларното јадро, бидејќи 8-бромо-цикличниот GMP не може да предизвика ерекција на пенисот кога се инјектира во ова јадро (Melis и Argiolas, 1995b) (Сл. 2). Што се однесува до нервните патишта активирани од допамин во јадрото accumbens што доведува до ерекција на пенисот, тие сè уште се непознати. Сепак, достапните податоци сугерираат дека овие патишта ја активираат невротрансмисијата на допамин во паравентрикуларното јадро на хипоталамусот. Според тоа, ерекцијата на пенисот индуцирана од окситоцин се јавува истовремено со зголемување на екстра-клеточниот допамин не само во nucleus accumbens, но исто така и во паравентрикуларното јадро и е антагонизиран од антагонистот на допаминските рецептори, халоперидол, кој се инјектира во паравентрикуларното јадро (Melis et al., 2007). АСите заедно, овие резултати ја поддржуваат идејата дека окситоцинергичните неврони кои потекнуваат од паравентрикуларното јадро и проектираат кон каудалната вентрална тегментална област, кога се активираат ослободувајќи окситоцин во оваа област, со што се активира NO-цикличниот GMP сигнален систем, што пак ги активира мезолимбичните допаминергични неврони (Melis et al., 2007, 2009a; Succu et al., 2008). Допаминот ослободен во јадрото accumbens за возврат ги активира нервните патишта кои водат до активирање на инцерто-хипоталамични допаминергични неврони, кои ги стимулираат паравентрикуларните окситоцинергични неврони кои проектираат на 'рбетниот мозок посредувајќи ја ерекцијата на пенисот. Во исто време, допаминот ослободен во паравентрикуларното јадро, исто така, може да ги активира окситоцинергичните неврони кои проектираат во екстра-хипоталамичните области на мозокот како што се вентралната тегментална област, хипокампусот, амигдалата и можеби други области на мозокот.
Како што се потсетивме погоре, во согласност со оваа хипотеза, апоморфинот инјектиран во паравентрикуларното јадро во доза што предизвикува ерекција на пенисот, исто така, ја зголемува екстраклеточната концентрација на допамин во јадрото accumbens, ефект намален од антагонистот на рецепторот на окситоцин d(CH2)5Tyr(Me ) 2-Orn8-вазотоцин инјектиран во вентралната тегментална област (Succu et al., 2007; Melis et al., 2009a). Заедно, горенаведените нервни патишта може да сочинуваат хипотетички нервни кола кои вклучуваат допамин, окситоцин и други невротрансмитери (на пример, глутаминска киселина, видете подолу) кои влијаат не само на сексуалните перформанси, туку и на сексуалната мотивација и сексуалното наградување (види дел 4).
3.4. Интеракција окситоцин-глутаминска киселина во вентралниот субикулум на хипокампусот
Окситоцинот предизвикува ерекција на пенисот кога се инјектира во вентралниот, но не и во дорзалниот субикулум, на начин зависен од дозата (Melis et al., 2009b). Сексуалниот одговор е посредуван од стимулацијата на рецепторите на окситоцин, која се укинува со претходна инјекција на d(CH2)5Tyr(Me)2-Orn8-вазотоцин даден во истото место на окситоцин, како што се наоѓа во други области на мозокот (види погоре) . Што се однесува до локализацијата на овие рецептори, достапните податоци сугерираат дека тие се локализирани во клеточните тела на невроните богати со азотен оксид-синтаза.
Соодветно, студиите за микродијализа покажуваат дека ерекцијата на пенисот индуцирана од окситоцин се јавува истовремено со зголемување на производството на азотен оксид во вентралниот субикулум, и ова зголемување е укината не само со претходното инјектирање на инхибиторот на азотен оксидосинтаза S-метил-тио-l-цитрулин. и од хемоглобинот за чистење на азотен оксид, но исто така и од d(CH2)5Tyr(Me)2-Orn8-вазотоцин, сите дадени во истото место на окситоцин во дози кои ја антагонизираат ерекцијата на пенисот (Melis et al., 2010). Уште поважно, ерекцијата на пенисот индуцирана од окситоцин се јавува истовремено со зголемување на концентрацијата на екстра-клеточната глутаминска киселина во вентралниот субикулум, која е само делумно антагонизирана од неконкурентниот антагонист на NMDA рецепторот (+)MK-801 даден во вентрален субикулум (Melis et al., 2010). Заедно, овие резултати сугерираат дека новоформираниот азотен оксид, делувајќи како меѓуклеточен гласник, го активира невротрансмисијата на глутаминска киселина што доведува до ерекција на пенисот, можеби преку нервните еферентни проекции од вентралниот субикулум до екстра-хипокампалните области на мозокот. Во согласност со оваа хипотеза, NMDA инјектирана во вентралниот субикулум индуцира ерекција на пенисот на начин зависен од дозата, и овој ефект е целосно антагонизиран со претходното инјектирање на истото место на (+)MK-801, но не и со S-метил-тио-l-цитрулин , хемоглобин или d(CH2)5Tyr(Me)2-Orn8-вазотоцин (Melis et al., 2010). Што се однесува до нервните еферентни патишта кои се проектираат во екстра-хипокампалните области на мозокот активирани од возбудливите аминокиселини (т.е. глутаминска киселина) во вентралниот субикулум, веројатно е дека тие се глутаматергични, како и поголемиот дел од хипокампалните еферентни проекции. Во моментот, може да се сугерира само дека овие проекции ја модулираат активноста на мезолимбичните допаминергични неврони, кои пак ја модулираат активноста на инцерто-хипоталамичните допаминергични неврони во паравентрикуларното јадро, што доведува до активирање на окситоцинергичните неврони посредување во ерекцијата на пенисот како што веќе беше дискутирано (Види погоре).
Соодветно на тоа, ерекцијата на пенисот предизвикана од вентралниот субикулум окситоцин се јавува истовремено со зголемување на концентрацијата на екстра-клеточниот допамин во обвивката на јадрото accumbens, и ова зголемување, како и ерекцијата на пенисот, се укинува со d(CH2)5Tyr(Me)2- Орн8-вазотоцин даден во вентралниот субикулум пред окситоцинот (Melis et al., 2009b). Покрај тоа, бидејќи ерекцијата на пенисот индуцирана од вентрален субикулум окситоцин е исто така намалена со (+)MK-801 инјектирана во вентралната тегментална област, но не и во акумбенс јадрото (Melis et al., 2009b) и се јавува истовремено со зголемување на екстра-клеточната глутаминска киселина во вентралната тегментална област, но не и во јадрото accumbens, бидејќи двата одговора се укинати со d(CH2)5Tyr(Me)-Orn8-вазотоцин, инјектиран во вентралниот субикулум пред окситоцин (види Сл. 3), веројатно е дека активирањето на мезолимбичните допаминергични неврони е секундарно на зголемената глутаматергична невротрансмисија во вентралната тегментална област. Ова сугерира дека интеракција на глутаминска киселина-допамин што ја контролира ерекцијата на пенисот постои во вентралната тегментална област. Потребни се дополнителни студии за да се утврди дали про-еректилните еферентни глутаметергиски патишта од субикулумот до вентралната тегментална област се директни или индиректни, т.е. преку префронталниот кортекс или други области на мозокот (види Melis et al., 2009b и референци во него). Бидејќи паравентрикуларното јадро исто така добива глутаматергични проекции од хипокампусот (види погоре и Saphier и Feldman, 1987), глутаминската киселина ги активира паравентрикуларните окситоцинергични неврони, вклучително и оние кои се проектираат на вентралната тегментална област (види Argiolas и Melis, 2005 и референци таму), и окситоцинот во вентралната тегментална област предизвикува ерекција на пенисот и ја зголемува активноста на мезолимбичниот допаминергичен невронs (види погоре), примамливо е да се шпекулира дека страравентрикуларните окситоцинергични неврони може исто така да бидат вклучени, барем делумно, во активирањето на мезолимбичните допаминергични неврони со окситоцин инјектиран во вентралниот субикулум (види дел 4).
4. Завршни забелешки
Студиите разгледани погоре ги потврдуваат и продолжуваат раните наоди кои покажуваат дека кај машките стаорци окситоцинот игра клучна улога во централната контрола на ерекцијата на пенисот на ниво на паравентрикуларното јадро на хипоталамусот и на 'рбетниот мозок. Конкретно, најмногу повторноСтудиите покажуваат дека окситоцинот влијае на ерекцијата на пенисот, исто така, делувајќи во други области на мозокот, т.е. вентралната тегментална област, вентралниот субикулум и постеромедијалното кортикално јадро на амигдалата..
На паравентрикуларно ниво, најважното ново откритие е можеби откривањето на експресијата на сите допамински рецептори од семејството D2 (D2, D3 и D4) во клеточните тела на окситоцинергичните неврони во паравентрикуларното јадро (и во супраоптичкото јадро и медијална преоптичка област) (Баскервил и Даглас, 2008; Баскервил и сор., 2009). Ова обезбедува силна невроанатомска поддршка на хипотезата дека допаминот и агонистите на допаминските рецептори можат директно да активираат окситоцинергични неврони вклучени во еректилната функција и проектирање не само на 'рбетниот мозок, туку и на екстра-хипоталамичните области на мозокот. Во овој поглед, исто така е важно откритието дека зголемувањето на интракавернозниот притисок предизвикано од агонисти на допаминските рецептори се намалува со блокадата на окситоцинергичните рецептори во лумбо-сакралниот 'рбетниот мозок (Baskerville et al., 2009). Навистина, иако такви докази се добиени кај анестезирани машки стаорци, наодот го потврдува активирањето на паравентрикуло-спинален окситоцинергичен десцендентен пат вклучен во ерекцијата на пенисот индуцирана од агонисти на допаминските рецептори. Хсепак, сè уште треба да се утврди дали ерекцијата на пенисот предизвикана од стимулацијата на допаминските рецептори присутни во телата на окситоцинергичните клетки е секундарна на активирањето на специфичен подтип на допамински рецептори од семејството D2 (D2, D3 или D4) или ако овие подтипови рецептори соработуваат во модулирањето на еректилниот одговор, можеби на различни начини во зависност од контекстот во кој се јавува ерекцијата на пенисот (види Moreland et al., 2004; Enguehard-Gueiffier et al., 2006; Melis et al., 2006a,b., 2009 година, Колинс и сор., 2009 година;
Друго важно ново откритие е дека окситоцинот индуцира ерекција на пенисот кога се инјектира не само во паравентрикуларното јадро или CA1 полето на хипокампусот, туку и во вентралната тегментална област, вентралниот субикулум и постеромедијалното кортикално јадро на амигдалата. Овие области на мозокот не беа тестирани во претходните студии кои покажаа дека окситоцинот ги зголемува епизодите на спонтана ерекција на пенисот кај машките стаорци, иако тие добиваат како лумбо-сакралните окситоцинергични проекции на 'рбетниот мозок од паравентрикуларното јадро. Навистина беше откриено дека окситоцинот може да ги зголеми епизодите на спонтана ерекција на пенисот, кои се јавуваат кај возрасни машки стаорци во отсуство на какви било сексуални стимули, како што се оние кои потекнуваат од присуството на пристапен или недостапен приемчив (подготвен со естроген-прогестерон) женски стаорец, кој е подложен на јајници или манипулација со гениталиите, кога се инјектира во паравентрикуларното јадро и полето CA1 на хипокампусот, но не и во дорзалниот субикулум, латералниот септум, каудатното јадро, медијалната преоптичка област, вентромедијалното јадро и супраоптичкото јадро (Melis et al. , 1986). Во сите овие студии, ерекцијата на пенисот обично се броела кога пенисот излегувал од обвивката на пенисот од набљудувач кој не бил свесен за дадените третмани директно за време на експериментот или подоцна со набљудување на експериментот снимен на видео лента со апарат за видео камера. Секоја епизода на ерекција на пенисот трае 0.5-1 мин и обично е придружена со чешлање на пенисот и/или свиткување на колкот. Обично не се прави експеримент кај овие стаорци за да се утврди ефектот на сексуалното искуство, возраста или дали овие стаорци може да се поделат на низок или висок одговор на про-еректилниот ефект на окситоцин инјектиран во различни области на мозокот. Ова исто така важи и за повеќето студии за про-еректилниот ефект на други невропептиди и лекови кои ги зголемуваат епизодите на спонтана ерекција на пенисот, вклучувајќи допамински агонисти, возбудливи амино киселини, ACTH-MSH, хексарелин и VGF пептиди. Сепак, про-еректилниот ефект на овие соединенија е постојано потврден со телеметриски методи, кои ја одредуваат појавата на ерекција на пенисот со зголемување на интракавернозниот притисок што се јавува спонтано или по администрација на овие соединенија на различни начини, т.е. системски, интрацеребровентрикуларно или директно во специфичните јадра на мозокот, по имплантирањето на микротрансдуктор на притисок директно во кавернозните тела (види Bernabè et al., 1999). Во вентралната тегментална област, вентралниот субикулум и постеромедијалното јадро на амигдалата, исто така, окситоцинот индуцира ерекција на пенисот делувајќи на окситоцинергичните рецептори. Ова доведува до активирање на мезолимбичните допаминергични неврони кои потекнуваат од вентралната тегментална област и се проектираат до обвивката на јадрото accumbens, мерено со зголемувањето на екстра-клеточната концентрација на допамин во дијализатот добиен од обвивката на nucleus accumbens и со намалувањето на еректилниот одговор предизвикан од пептидот инјектиран во овие екстра-хипоталамични области, пронајден по блокадата на допаминергичните рецептори во nucleus accumbens (види подолу). Што се однесува до механизмите кои се активираат со стимулација на окситоцинергични рецептори во овие области на мозокот, кои доведуваат до активирање на мезолимбичните допаминергични неврони и до ерекција на пенисот, најдобро се разјаснети оние кои се јавуваат во каудалната вентрална тегментална област. Навистина, овде фармаколошките и имуно-флуоресцентните резултати покажуваат дека нервните завршетоци на окситоцин удираат на клеточните тела на допаминергичните неврони кои се проектираат на обвивката на јадрото accumbens (Melis et al., 2007, 2009a; Succu et al., 2008). Многу од овие неврони се богати со азотен оксид синтаза и со гванилат циклаза. Стимулацијата на окситоцинергични рецептори во клеточните тела на овие допаминергични неврони предизвикува активирање на азотен оксид синтаза што доведува до зголемено производство на азотен оксид. Азотниот оксид за возврат ја активира гванилат циклазата, а со тоа ја зголемува концентрацијата на цикличниот GMP, што доведува до активирање на мезолимбичните допаминергични неврони и до ослободување на допамин во акумбенс јадрото, мерено со зголемувањето на екстра-клеточниот допамин во дијализатот од јадрото акумбенс добиен со интрацеребрална микродијализа (Succu et al., 2008 ). Допаминот ослободен во јадрото accumbens за возврат ги активира нервните патишта кои водат до ерекција на пенисот. Ова е поддржано од способноста на антагонистите на допаминските рецептори, халоперидол и/или цис-флупентиксол инјектирани во јадрото accumbens да ја намалат вентралната тегментална област индуцирана од окситоцин ерекцијата на пенисот (Succu et al., 2008). Еден од про-еректилните патишта се чини дека ги активира инцерто-хипоталамичните допаминергични неврони, особено оние кои се проектираат на клеточните тела на паравентрикуларните окситоцинергични неврони. Навистина, окситоцинот кој се инјектира во опашката вентрална тегментална област го зголемува екстрацелуларниот допамин не само во јадрото accumbens, туку и во паравентрикуларното јадро, а блокадата на допаминските рецептори во паравентрикуларното јадро значително ја намалува вентралната тегментална област со намалена окситоцин-индукција. , 2007, 2008 Мелис и сор., 2007, 2009а). Постоењето на овие јадро accumbens допамин-паравентрикуларен допамин-паравентрикуларен окситоцин-вентрален тегментален простор окситоцин-допамин врски се сугерира и со способноста на про-еректилната доза на апоморфин и на агонистот на рецепторот D4 PD 168077 инјектирана во паравентрикуларното јадро да го зголеми екстрацелуларниот допамин во обвивката на јадрото accumbens (Succu et al., 2007), одговор кој е укинат со d(CH2)5Tyr(Me)-Orn8-вазотоцин даден во вентралната тегментална област (Melis et al., 2009a, види исто така подолу). Сепак, потребна е понатамошна работа за да се идентификуваат нервните патишта кои го поврзуваат акумбенс јадрото со инцерто-хипоталамичниот допаминергичен систем.
Механизмот со кој окситоцинот предизвикува ерекција на пенисот и ги активира мезолимбичните допаминергични неврони кога се инјектира во вентралниот субикулум или во постеромедијалното јадро на амигдалата во моментов е само делумно разбран. Во овие области, исто така, окситоцинот ги активира сопствените рецептори кои доведуваат до активирање на азотен оксид-синтаза, а со тоа го зголемува производството на азотен оксид. Азотниот оксид за возврат ги активира непознатите еферентни проекции, кои очигледно ја зголемуваат глутаматергичната невротрансмисија во вентралната тегментална област. Глутаминската киселина потоа ги стимулира мезолимбичните допаминергични неврони што доведува до ерекција на пенисот. Оваа хипотеза е поддржана главно од способноста на окситоцинот инјектиран во вентралниот субикулум да ја зголеми екстра-клеточната глутаминска киселина во вентралната тегментална област (Сл. 3), и на неконкурентниот антагонист на NMDA рецепторот (+)MK-801 инјектиран во вентралната тегментална област, но не и во акумбенс јадрото, за да се намали ерекцијата на пенисот предизвикана од окситоцин инјектиран или во вентралниот субикулум или во постеромедијалното јадро на амигдалата (Melis et al., 2009b). Во моментов, достапни се повеќе детали за ерекцијата на пенисот индуцирана од вентралниот субикулум окситоцин. Овде ерекцијата на пенисот индуцирана од окситоцин се чини секундарна во однос на активирањето на окситоцинергичните рецептори лоцирани во клеточните тела на невроните што содржат азотен оксид-синтаза. Ова предизвикува зголемување на производството на азотен оксид, кој ја активира глутаматергичната невротрансмисија делувајќи како меѓуклеточен гласник со механизам сличен на оној опишан за долгорочно потенцирање (види Снајдер, 1992; Саутам и Гартвејт, 1993; Шуман и Медисон, 2004). . Во согласност со оваа хипотеза, ерекцијата на пенисот индуцирана од окситоцин се јавува истовремено со зголемување на екстрацелуларната глутаминска киселина во дијализатот од вентралниот субикулум и стимулацијата на рецепторите на возбудливи аминокиселини во вентралниот субикулум од NMDA, индуцира ерекција на пенисот. Глутаминската киселина за возврат ги активира нервните еферентни проекции, кои доведуваат до активирање на мезолимбичните допаминергични неврони во вентралната тегментална област, како што е објавено погоре. Ако овие механизми се оперативни, исто така во постеромедијалното јадро на амигдалата, засега е непознато. Понатаму, потребни се дополнителни студии за да се докаже дека (1) окситоцинергични нервни завршетоци и рецептори во вентралниот субикулум и во постеромедијалното јадро на амигдалата се локализирани во клеточните тела на неврони кои содржат азотен оксид-синтаза, (2) ако овие неврони на крајот се под контрола на возбудливите аминокиселински (глутаматергиски) синапси и (3) да се идентификува сигналниот систем активиран од азотен оксид. Во овој поглед, важно е да се забележи дека способноста на окситоцинот да ја зголеми активноста на азотен оксид-синтаза во клеточните тела на допаминергичните неврони во каудалната вентрална тегментална област и на сè уште непознатите неврони во вентралниот субикулум и постеромедијалното јадро на амигдалата, потсетува способноста на окситоцинот да ја активира азотен оксид-синтаза во клеточните тела на окситоцинергичните неврони во паравентрикуларното јадро (Melis et al., 1997). Меѓутоа, додека во вентралната тегментална област зголеменото производство на азотен оксид во клеточните тела на допаминергичните неврони доведува до активирање на овие неврони со активирање на гванилат циклазата и зголемување на цикличниот GMP, тоа не се случува во паравентрикуларното јадро. Соодветно на тоа, 8-бромо-цикличниот GMP инјектиран во паравентрикуларното јадро не предизвикува ерекција на пенисот, додека тоа го прави кога се инјектира во вентралната тегментална област. Друга сигнална патека различна од системот на азотен оксидцикличен GMP потоа е вклучена на паравентрикуларно ниво во активирањето на окситоцинергични неврони кои посредуваат во ерекцијата на пенисот со ендоген и/или егзоген азотен оксид (Melis и Argiolas, 1995b; Melis et al., 1997) (Сл. . 1). Од друга страна, цикличниот GMP во вентралната тегментална област се чини дека игра исто така клучна улога во активирањето на мезолимбичните допаминергични неврони и во зголемувањето на екстра-клеточниот допамин што се јавува во дијализатот добиен од обвивката на јадрото акумбенс на машките стаорци. избрано за прикажување или неприкажување на неконтактна ерекција на пенисот кога се става во присуство на недостапен рецептивен јајник (третиран со естроген + прогестерон) женски стаорец. Во овие експериментални услови, кај машките стаорци кои покажуваат бесконтактна ерекција на пенисот, се забележува зголемување на екстра-клеточната концентрација на допамин како што се очекуваше, и ова зголемување дополнително се зголемува, иако само скромно, со инхибитори на фосфодиестераза дадени во опашката вентрална тегментална област (Sanna et ал., 2009).
Способноста на окситоцинот инјектиран во вентралната тегментална област, во вентралниот субикулум и во постеромедијалното кортикално јадро на амигдалата, заедно со онаа на допаминските агонисти инјектирани во паравентрикуларното јадро, да предизвикаат ерекција на пенисот и да активираат некои мезолимични невросервиси. . Прво, механизмите слични на оние споменати погоре може да бидат оперативни кога ерекцијата на пенисот се јавува во физиолошки контекст, како на пример за време на копулација (кога во копула се јавуваат ерекции на пенисот) или за време на бесконтактна ерекција на пенисот. Овие ерекции се феромонски посредувани ерекции на пенисот, кои не се разликуваат од оние предизвикани од лекови или окситоцин, кои се јавуваат кога сексуално потентни машки стаорци се ставени во присуство на недостапен приемчив (овариектомизиран естроген + прогестерон) женски стаорец и се сметаат за индекс на сексуални возбуда (Sachs, 1997, 2007). Навистина, иако овие резултати не покажуваат дека окситоцинот во овие области игра улога во ерекцијата на пенисот што се јавува во физиолошки контекст или по администрацијата на лекот, тие додаваат дополнителна сила на раните наоди што сугерираат дека овие области на мозокот припаѓаат на оние каде што окситоцинот даден централно делува на зголемување на не само спонтани епизоди на ерекција на пенисот забележани по про-еректилни лекови, туку и за подобрување на машкото (и женското) сексуално однесување (види Argiolas и Melis, 2004 и референци таму). Според тоа, концентрацијата на окситоцин се зголемува во хипокампусот на машки стаорци третирани со про-еректилна доза на апоморфин, класичен допамински агонист (Melis et al., 1990) и d(CH2)5Tyr(Me)-Orn8-вазотоцин, кој го блокира окситоцинот рецептори, е исклучително ефикасен не само во нарушување на копулаторното однесување (Argiolas et al., 1987a), туку и олеснувачкиот ефект на апоморфинот врз машкиот копулаторно однесување (Argiolas et al., 1987b) кај сексуално потентни машки стаорци, при што кај копула ерекција на пенисот се јавуваат. d(CH2)5Tyr(Me)-Orn8-вазотоцинот е исто така исклучително моќен во намалувањето на бесконтактните ерекции кај сексуално потентни машки стаорци, кога се дава во нанограмски количини во страничните комори, но не и во PVN (Melis et al., 1999a).
Второ, мезолимбичните допаминергични неврони играат клучна улога во мотивационите и наградувачките својства на природните зајакнувачки стимули, како што се храната, водата и сексуалната активност (Фибигер и Филипс, 1988; Вајс и Ромпре, 1989; Еверит, 1990). Конкретно, се смета дека допаминот ослободен од овие неврони посредува во транспонирањето на мотивациските аспекти на природните стимули во однесувања насочени кон целта, на пример во случај на сексуална активност, барање сексуален партнер и сексуален однос за да се постигне награда и задоволство. (Гото и Грејс, 2005). Соодветно на тоа, екстрацелуларната концентрација на допамин се зголемува во дијализатот од јадрото на сексуално потентни машки стаорци за време на изложеност на недостапен овариектомизиран естроген + рецептивен женски стаорец подготвен со прогестерон, а таквото зголемување беше уште поголемо кога на машкиот стаорец му беше дозволено да се копулира со приемчивата женка (Пфаус и Еверит, 1995).
Трето, сегашните резултати ја поддржуваат хипотезата дека невронското коло го поврзува паравентрикуларното јадро со вентралната тегментална област директно или индиректно (преку вентралниот субикулум и/или постеромедијалното кортикално јадро на амигдалата) и јадрото accumbens, и од тука преку непознати патеките назад до паравентрикуларното јадро за да ја контролираат активноста на окситоцинергичните неврони кои проектираат на 'рбетниот мозок што посредува во ерекцијата на пенисот и на окситоцинергичните неврони кои проектираат кон вентралната тегментална област, вентралниот субикулум и постеромедијалното кортикално јадро на амигдалата што ја модулира на овој начин. активност на мезолимбичните допаминергични неврони (сл. 4). Ова сложено невронско коло може да игра улога во интеграцијата на нервните активности вклучени во контролата на конзуматорните (еректилно-ејакулаторни) и антиципативни (мотивациски и наградувачки) аспекти на машкото сексуално однесување во физиолошки контекст. Навистина, екстрацелуларниот допамин се зголемува во јадрото accumbens (Пфаус и Еверит, 1995) и во паравентрикуларното јадро на сексуално потентни машки стаорци за време на изложеност на недостапни приемчиви женски стаорци, кога се јавуваат ерекции без контакт, а уште повеќе кога е дозволена копулација. на пр., кога во копула се случуваат ерекции на пенисот (Melis et al., 2003). Така, иако се неопходни дополнителни студии за да се разјасни улогата на ендогениот окситоцин во вентралната тегментална област, вентралниот субикулум и амигдалата за време на бесконтактни ерекции и сексуално однесување, тогаш може да биде разумно да се претпостави дека ова хипотетичко нервно коло, истовремено придонесувајќи за консумативните аспекти на сексуалното однесување, во исто време, исто така, може да го активираат мезолимбичкиот допаминергичен систем обезбедувајќи нервен супстрат за објаснување на наградувачките својства на сексуалната активност (Еверит, 1990; Пфаус и Еверит, 1995). Во овој поглед, вреди да се забележи дека мезолимбичниот допаминергичен систем активиран со окситоцин инјектиран во вентралната тегментална област е ист активиран од лекови за злоупотреба како што се опијати, канабиноиди, амфетамин, кокаин и алкохол (Tanda et al., 1997) и Откриено е дека окситоцинот може да ја намали толеранцијата и зависноста од кокаин, морфин, алкохол и канабиноиди (Kovacs et al., 1998; Cui et al., 2001). Како заклучок, се чини дека окситоцинот ослободен не само во вентралната тегментална област, туку и во вентралниот субикулум и постеромедијалното кортикално јадро на амигдалата, може да ги активира мезолимбичните допаминергични неврони, кои можат да бидат вклучени во апетитивните и наградувачките ефекти на сексуалната активност. . Активирањето на мезолимбичните допаминергични неврони може да биде директно, преку окситоцинергични рецептори во клеточните тела на мезолимбичните допаминергични неврони, или индиректно преку активирање на невротрансмисијата на глутаминска киселина во вентралната тегментална област.
Допаминот ослободен во обвивката на nucleus accumbens, пак, ја модулира активноста на инцерто-хипоталамичните допаминергични неврони во паравентрикуларното јадро, предизвикувајќи или ерекција на пенисот (преку активирање на окситоцинергични неврони кои се проектираат на 'рбетниот мозок), или сексуална мотивација и награда на оксиактивност неврони кои се проектираат кон вентралната тегментална област, вентралниот субикулум или постеромедијалното кортикално јадро на амигдалата). Бидејќи допаминот се ослободува и во обвивката на nucleus accumbens и во паравентрикуларното јадро кога ерекцијата на пенисот се јавува во физиолошки контекст (на пример, бесконтактни ерекции и копулации) (Succu et al., 2007; Melis et al., 2003, 2007), тоа е веројатно дека централните окситоцинергични неврони учествуваат во нервните кола кои посредуваат во интеракцијата помеѓу мезолимбичниот и инцерто-хипоталамичниот допаминергичен систем. Овие нервни кола може да играат улога не само во завршната фаза на сексуалното однесување (на пример, ерекција и копулација на пенисот), туку и во сексуалната мотивација, сексуалната возбуда и сексуалната награда.
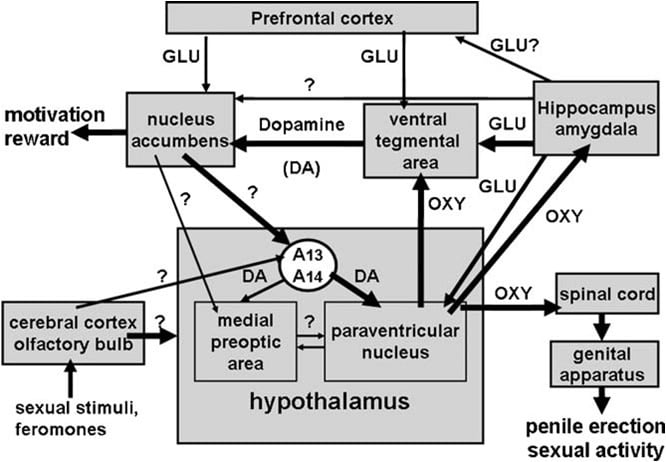
Шематски приказ на хипотетички нервни кола кои вклучуваат окситоцин што влијае на сексуалната мотивација, наградувањето и сексуалните перформанси, како што е предложено од резултатите од ова поглавје и претходните извештаи. Окситоцинергичните неврони кои потекнуваат од паравентрикуларното јадро и проектираат до 'рбетниот мозок кога се активираат на пример со допамин и глутаминска киселина (но и од други невротрансмитери и/или невропептиди), ја олеснуваат ерекцијата на пенисот и сексуалните перформанси со активирање на окситоцинергичните неврони кои се проектираат на' рбетниот столб. Допаминот и глутаминската киселина (но, исто така, невротрансмитери и невропептиди) во паравентрикуларното јадро, исто така, ги активираат окситоцинергичните неврони кои се проектираат во вентралната тегментална област, а со тоа ги активираат мезолимбичните допаминергични неврони кои се проектираат на јадрото акумбенс, модулирање на сексуалната мотивација и награда. Допаминот ослободен во nucleus accumbens (NAs) за возврат ги активира сè уште непознатите нервни патишта, кои ја зголемуваат активноста на инцерто-хипоталамични допаминергични неврони (кои потекнуваат од A13-A14 групите на Dahlstrom и Fuxe) кои напаѓаат меѓу другите на окситоцинергичните неврони, вклучувајќи ги и оние што се проектираат 'рбетниот мозок, што доведува до ерекција на пенисот. Ова коло може да се активира и со окситоцин кој се инјектира не само во опашката вентрална тегментална област, туку и во вентралниот субикулум и во амигдалата, кои исто така добиваат окситоцинергична инервација од паравентрикуларното јадро, можеби преку директен или индиректен глутаматергичен еферент. тегменталната област, што доведува до модулација и на сексуалната мотивација и на ерекцијата на пенисот. Конечно, горенаведените кола може да се активираат и со сексуални стимули и феромони, бидејќи екстра-клеточниот допамин и глутаминската киселина се зголемуваат во паравентрикуларното јадро (и во медијалната преоптичка област) за време на бесконтактни ерекции и копулации посредувани од феромони (за соодветни референци види Список со референци).
Благодарност Оваа работа беше делумно поддржана со грант од италијанското Министерство за Универзитетот и за истражување на АА и МРМ
Референци
Андерсон, КЕ, 2001 година. Фармакологија на ерекција на пенисот. Фармакол. Свештеникот 53, 417-450. Аргиолас, А., 1994 година. Азотниот оксид е централен медијатор на ерекцијата на пенисот. Неврофармакологија 33, 1339-1344. Аргиолас, А., 1999 година. Невропептиди и сексуално однесување. Невроси. Biobehav. Свештеникот 23, 1127-1142. Аргиолас, А., Геса, ГЛ, 1991 година. Централните функции на окситоцинот. Невроси. Biobehav. Свештеникот 15, 217-231. Аргиолас, А., Мелис, М.Р., 1995 година. Неуромодулација на ерекцијата на пенисот: преглед на улогата на невротрансмитери и невропептиди. Прог. Невробиол. 47, 235-255. Аргиолас, А., Мелис, М.Р., 2004 година. Улогата на окситоцинот и паравентрикуларното јадро во сексуалното однесување на машките цицачи. Физиол. Однесување. 83, 309-317. Аргиолас, А., Мелис, М.Р., 2005 година. Централна контрола на ерекцијата на пенисот: улогата на паравентрикуларното јадро на хипоталамусот. Прог. Невробиол. 76, 1-21. Аргиолас, А., Колу, М., Геса, ГЛ, Мелис, МР, Сера, Г., 1988 година. Антагонистот на окситоцинот d(CH2)5Tyr(Me)-Orn8-вазотоцин го антагонизира машкото копулаторно однесување кај стаорци. Е. J. Фармакол. 149, 389-392. Аргиолас, А., Колу, М., Д'Аквила, П., Геса, ГЛ, Мелис, МР, Сера, Г., 1989 година. Апоморфинската стимулација на машкото копулаторно однесување е спречена од антагонистот на окситоцин d(CH2)5Tyr(Me)-Orn8-вазотоцин кај стаорци. Фармакол. Биохемиски. Однесување. 33, 81-83. Аргиолас, А., Мелис, М.Р., Геса, ГЛ, 1985 година. Интравентрикуларниот окситоцин предизвикува зевање и ерекција на пенисот кај стаорци. Е. J. Фармакол. 117, 395-396. Аргиолас, А., Мелис, М.Р., Геса, ГЛ, 1986 година. Окситоцин: исклучително моќен индуктор на ерекција на пенисот и проѕевање кај машки стаорци. Е. J. Фармакол. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Лезијата на паравентрикуларното јадро го спречува прозевањето и ерекцијата на пенисот предизвикани од апоморфин и окситоцин, но не и од ACTH кај стаорци. Мозокот Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d(CH2)5Tyr(Me)-Orn8-вазотоцин, моќен антагонист на окситоцин, ја антагонизира ерекцијата на пенисот и проѕевањето предизвикани од окситоцинот и апоморфинот, но не и од ACTH 1-24. Е. J. Фармакол. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990 година. _-Конотоксинот ја спречува ерекцијата на пенисот и проѕевањето предизвикана од апоморфин и окситоцин кај машките стаорци. Фармакол. Биохемиски. Однесување. 37, 253-257. Арлети, Р., Бертолини, А., 1985 година. Окситоцинот го стимулира однесувањето на лордоза кај женските стаорци. Невропептиди 6, 247-255. Арлети, Р., Бацани, Ц., Кастели, М., Бертолини, А., 1985 година. Окситоцинот ги подобрува машките копулаторни перформанси кај стаорци. Хорм. Однесување. 19, 14-20. Арлети, Р., Бенели, А., Бертолини, А., 1990 година. Сексуалното однесување на постарите машки стаорци е стимулирано со окситоцин. Е. J. Фармакол. 179, 377-382. Арлети, Р., Калза, Л., Џардино, Л., Бенели, А., Кавацути, Е., Бертолини, А., 1997 година. Сексуалната импотенција е поврзана со намалено производство на окситоцин и зголемено производство на опиоидни пептиди во паравентрикуларното јадро на хипоталамусот. Невроси. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Доказ за директна проекција од паравентрикуларното јадро на хипоталамусот до наводните серотонинергични неврони на јадрото paragigantocellularis вклучени во контролата на ерекцијата кај стаорци. Е. J. Невроси. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992 година. Азотен оксид: физиолошки медијатор на ерекцијата на пенисот. Наука 257, 401-403. Баскервил, ТА, Даглас, Еј Џеј, 2008 година. Интеракција помеѓу допамин и окситоцин во контролата на сексуалното однесување. Прог. Мозокот Res. 170, 277-289. Баскервил, Т.А., Алард, Џ., Вејман, Ц., Даглас, Еј Џеј, 2009 година. Интеракција на допамин-окситоцин во ерекцијата на пенисот. Е. J. Невроси. 30, 2151-2164. Бенели, А., Бертолини, А., Поџоли, Р., Кавацути, Е., Калза, Л., Џардино, Л., Арлети, Р., 1995 година. Азотниот оксид е вклучен во машкото сексуално однесување на стаорци. Е. J. Фармакол. 294, 505-510. Бернабе, Ј., Рампин, О., Сакс, БД, Џулијано, Ф., 1999 година. Интракавернозен притисок за време на ерекцијата кај стаорци: интегративен пристап заснован на телеметриско снимање. Am. J. Физиол. 276, R441-R449. Битнер, РС, Никел, АЛ, Оте, С., Мартино, Б., Барлоу, ЕХ, Бхатија, П., Стјуарт, АО, Бриони, ЈД, Декер, МВ, Мореланд, РБ, 2006 година. Сигнализирање на рецепторот на допамин D4 во паравентрикуларното хипоталамично јадро на стаорец: доказ за природно спојување кое вклучува непосредна рана генска индукција и фосфорилација на протеин киназа активирана од митоген. Неврофармакологија 50, 521-531. Бриони, Џ. , Ј.С., м-р Осински, Марш, К., Андерсон, КЕ, Саливан, ЈП, 3 година. Активирањето на рецепторите за допамин D4 од ABT-724 предизвикува ерекција на пенисот кај стаорци. Proc. Natl. Acad. Sci. Соединетите Американски Држави 101, 6758-6763. Бујс, РМ, 1978 година. Интра- и екстра-хипоталамични патишта на вазопресин и окситоцин кај стаорци. Cell Tissue Res. 192, 423-435. Buijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984 година. Допаминергичната инервација на супраоптичкото и паравентрикуларното јадро. Светлосна и електронска микроскопска студија. Мозокот Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986 година. Окситоцинот ја олеснува сексуалната приемчивост на женските стаорци третирани со естроген. Невропептиди 7, 175-189. Камерон, Џ. Допаминергична стимулација на концентрациите на окситоцин во плазмата на машките и женските мајмуни од апоморфин и агонист на рецепторот D2. J. Clin. Ендокринол. Метаб. 75, 855-860. Кантерас, НС, Симерли, РБ, Свансон, ЛВ, 1995 година. Организација на проекции од медијалното јадро на амигдалата: PHAL студија кај стаорци. J. Comp. Неврол. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987 година. Плазма окситоцинот го зголемува човечкиот сексуален одговор. J. Clin. Ендокринол. Метаб. 64, 27-31. Картер, ЦС, 1992 година. Окситоцин и сексуално однесување. Невроси. Biobehav. Свештеникот 16, 131-144. Картер, ЦС, Ледерхендлер, II, Киркпатрик, Б., 1997 година. The Interactive Neurobiology of Affiliation, Анали на Њујоршката академија на науките, кн. 807. Академијата на науките во Њујорк, Њујорк. Кастели, пратеник, Пирас, А.П., Мелис, Т., Суку, С., Сана, Ф., Мелис, МР, Колу, С., Енас, МГ, Дијаз, Г., Маки, К., Аргиолас, А. , 2007 година. Канабиноидни CB1 рецептори во паравентрикуларното јадро и централна контрола на ерекцијата на пенисот: имуноцитохемија, авторрадиографија и бихејвиорални студии. Невронаука 147, 197-206. Чен, КК, Чанг, ЛС, 2003 година. Ефект на агонисти на рецептори за возбудливи амино киселини врз ерекцијата на пенисот по администрација во паравентрикуларното јадро на хипоталамусот кај стаорци. J. Урол. 62, 575-580. Чен, КК, Чан, ЈЈХ, Чанг, ЛС, Чен, МТ, Чанг, ШХ, 1992 година. Елицитација на ерекција на пенисот по активирање на формирањето на хипокампусот кај стаорецот. Невроси. Lett. 141, 218-222. Чен, КК, Чан, ЈЈХ, Чанг, ЛС, 1999 година. Допаминергична невротрансмисија во паравентрикуларното јадро на хипоталамусот во централна регулација на ерекцијата на пенисот кај стаорец. J. Урол. 162, 237-242. Колинс, Г.Т., Труконе, А., Хаџи-Абди, Ф., Њуман, А.Х., Грунд, П., Рајс, К.Ц. , Ј., Ванг, С., Кац, Џ.Л., Гранди, ДК, Сунахара, РК, Вудс, ЈХ, 2009 година. Про-еректилните ефекти на агонисти слични на допамин Д2 се посредувани од рецепторот Д3 кај стаорци и глувци. J. Фармакол. Екстензии. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004 година. Централна регулација на ејакулација. Физиол. Однесување. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001 година. Превенција на синдром на повлекување на канабиноид со литиум: вклучување на окситоцинергична невронска активација. J. Невроси. 21, 9867-9876. Далстром, А., Фукс, К., 1964 година. Доказ за постоење на неврони кои содржат моноамин во централниот нервен систем. I. Демонстрација на моноамини во клеточните тела на невроните на мозочното стебло. Acta Physiol. Сканда. 62 (Suppl. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Ерекција на пенисот и зевање индуцирани од агонисти на рецептори слични на допамин D2 кај стаорци: влијание на напрегањето и придонес на допамин D2, но не и D3 и D4 рецептори. Однесување. Фармакол. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Окситоцинот ги ублажува одговорите на амигдалата на емоционалните лица без оглед на валентноста. Biol. Psychiatry 62, 11871190. Домингез, Ј., Риоло, Ј.В., Ксу, З., Хул, МЕ, 2001 година. Регулирање од страна на медијалната амигдала на копулација и медијално преоптичко ослободување на допамин. J. Невроси. 21, 349-355. Доналдсон, З.Р., Јанг, ЛЈ, 2009 година. Окситоцин, вазопресин и неврогенетиката на социјалноста. Наука 322, 900-904. Итон, Р.Г., Марковски, ВФ, Ламли, Лос Анџелес, Томпсон, Џ.Т., Мозес, Ј., Хал, ЕМ, 1991 година. D2 рецепторите во паравентрикуларното јадро ги регулираат гениталните одговори и копулацијата кај машките стаорци. Фармакол. Биохемиски. Однесување. 39, 177-181. Ebner, K., Bosch, OJ, Krömer, SA, Singewald, N., Neumann, ID, 2005 година. Ослободувањето на окситоцин во централната амигдала на стаорци го модулира однесувањето за справување со стресот и ослободувањето на возбудливи амино киселини. Невропсихофармакологија 30, 223-230. Enguehard-Gueifier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006 година. 2-[(4-Фенилпиперазин-1-ил)метил]имидазо(ди)азини како селективни D4-лиганди. Индукција на ерекција на пенисот со 2-[4-(2-метоксифенил)пиперазин-1-илметил]имидазо[1,2-a]пиридин (PIP3EA), моќен и селективен D4 агонист. J. Мед. Chem. 49, 3938-3947. Еверит, БЈ, 1990 година. Сексуална мотивација: нервна и бихејвиорална анализа на механизмите кои лежат во основата на апетитивните и копулаторните одговори машки стаорци. Невроси. Biobehav. Свештеникот 14, 217-232. Фибигер, HC, Филипс, АГ, 1988 година. Мезокортиколимбичен допамински систем и награда. Ен. N. Y. Acad. Sci. 5, 206-215. Француски, СЈ, Тотердел, С., 2003 година. Индивидуалните неврони на проекцијата на јадрото accumbens добиваат и базолатерална амигдала и вентрални субикуларни аференти кај стаорци. Невронаука 119, 19-31. 954 г Мелис, А. Argiolas / Невронаука и биобихејвиорални прегледи 35 (2011) 939-955 Фројнд-Мерсиер, МЈ, Ричард, П., 1981 година. Возбудливи ефекти на интравентрикуларни инјекции на окситоцин врз рефлексот на исфрлање млеко кај стаорец. Невроси. Lett. 23, 193-198. Фројнд-Мерсиер, МЈ, Ричард, П., 1984 година. Електрофизиолошки докази за олеснувачка контрола на окситоцин неврони со окситоцин за време на цицање кај стаорец. J. Физиол. (Лонд.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995 година. Соматодендритични авторецептори на окситоцин неврони. Во: Ајвел, Р., Расел, Џеј (Eds.), Окситоцин, клеточни и молекуларни пристапи во медицината и истражувањето. Adv. Екстензии. Мед. Биол., 365. Пленум Прес, Њујорк и Лондон, стр. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Фармаколошки карактеристики и анатомска дистрибуција на местата за врзување на 3H окситоцин во мозокот на стаорци Вистар, проучувани со авторрадиографија. Невронаука 20, 599-614. Џулијано, Ф., Рапин, О., 2000 година. Централна контрола на ерекцијата на пенисот. Невроси. Biobehav. Свештеникот 24, 517-533. Џулијано, Ф., Алард, Ј., 2001 година. Допамин и сексуална функција. Инт. J. Impotence Res. 13 (Suppl. 3), 18-28. Џулијано, Ф., Рапин, О., 2004 година. Неврална контрола на ерекцијата. Физиол. Однесување. 83, 189-201. Џулијано, Ф., Бернабе, Ј., Мекена, КЕ, Лонгвил, Ф., Рампин, О., 2001 година. Спинален проеректилен ефект на окситоцин кај анестезирани стаорци. Дали сум Џ. Физиол. Регуларно. Интег. Comp. Физиол. 280, R1870-R1877. Гото, Ј., Грејс, АА, 2005 година. Допаминергична модулација на лимбичкиот и кортикалниот погон на јадрото accumbens во однесување насочено кон целта. Nat. Невроси. 8, 805-812. Глог, Ј., Анг, ВТ, Џенкинс, ЈС, 1985 година. Ефекти од лезии во хипоталамичните паравентрикуларни, супраоптични и супрахијазматични јадра на вазопресин и окситоцин кај мозокот и спиналниот мозок на стаорци. Мозокот Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Moon, MW, 1997 година. Синтеза и биолошки активности на (R)-5,6-дихидро-N,N-диметил-4H-имидазо[4,5,1-ij]хинолин-5-амин) и неговите метаболити. J. Мед. Chem. 40, 639-646. Хсиех, ГЦ, Холингсворт, ПР, Мартино, Б., Чанг, Р., Теранова, м-р, О'Нил, АБ, Линч, Џеј Џеј, Мореланд, Р.Б., Донели-Робертс, Д.Л., Коласа, Т., Микуса, Ј.П. , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Централни механизми што ја регулираат ерекцијата на пенисот кај свесни стаорци: допаминергичните системи поврзани со проеректилниот ефект на апоморфин. J. Фармакол. Екстензии. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993 година. Насочено нарушување на генот на синтезата на неуроналниот азотен оксид. Ќелија 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005 година. Вазопресинот и окситоцинот возбудуваат различни невронски популации во централната амигдала. Наука 308, 245-248. Хал, ЕМ, Ворнер, РК, Базет, Ти Џеј, Итон, РЦ, Томпсон, ЈТ, 1989 година. Односот D2/D1 во медијалната преоптичка област влијае на копулацијата на машките стаорци. J. Фармакол. Екстензии. Ther. 251, 422-427. Хал, ЕМ, Ду, Ј., Лорен, ДС, Матушевич, Л., 1995 година. Екстра-клеточен допамин во медијалната преоптичка област: импликации за сексуалната мотивација и хормоналната контрола на копулацијата. J. Невроси. 15, 7465-7471. Hull, EM, Meisel, RL, Sachs, BD, 2002 година. Машко сексуално однесување. Во: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Eds.), Hormones, Brain and Behavior. Академски печат, Њујорк, стр. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, КМ, 2010 година. Окситоцинот го подобрува учењето зависно од амигдалата, социјално засилено и емоционалната емпатија кај луѓето. J. Невроси. 30, 4999-5007. Ајвел, Р., Расел, ЈА, 1995 година. Окситоцин: клеточни и молекуларни пристапи во медицината и истражувањето. Напредокот во експерименталната медицина и биологијата, кн. 395. Пленум прес, Њујорк. Кели, АЕ, Домесик, ВБ, 1982 година. Распределбата на проекцијата од формацијата на хипокампусот до јадрото на акумбенс кај стаорецот: студија за антероградна и ретроградна пероксидаза на рен. Невронаука 7, 2321-2335. Кимура, Ј., Наиту, Ј., Ванибучи, Ф., Јамагучи, Т., 2008 година. Активирањето на 5-HT(2C) рецепторот е вообичаен механизам на проеректилните ефекти на апоморфин, окситоцин и меланотан-II кај стаорци. Е. J. Фармакол. 589, 157-162. Кондо, Ј., Сакс, БД, Сакума, Ј., 1998 година. Важноста на медијалната амигдала во ерекцијата на пенисот на стаорци предизвикана од далечни стимули од женките во еструсот. Однесување. Мозокот Res. 91, 215-222. Ковач, Г.Л., Сарњаи, З., Сабо, Г., 1998 година. Окситоцин и зависност: преглед. Психоневроендокринологија 23, 945-962. Ли, ХЈ, Макбет, АХ, Пагани, ЈХ, Скот Јанг 3-ти, В., 2009 година. Окситоцин: големиот олеснувач на животот. Прог. Невробиол. 88, 127-151. Линдвал, О., Бјорклунд, А., Скагерберг, Г., 1984 година. Селективна истохемиска демонстрација на допамински терминални системи во ди- и теленцефалон на стаорци: нов доказ за допаминергична инервација на хипоталамични невросекреторни јадра. Мозокот Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997 година. Нарушен сексуален одговор по лезии на паравентрикуларното јадро на хипоталамусот кај машки стаорци. Однесување. Невроси. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Рамката на азулен како нов биоизостер: дизајн на потентни допамински рецепторни лиганди D4 кои поттикнуваат ерекција на пенисот. Chem. Мед. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. _-Конотоксин: директна и постојана блокада на специфични типови на калциумови канали во невроните, но не и во мускулите. Прот. Nat. Acad. Sci. Соединетите Американски Држави 84, 4327-4331. Мекена, КЕ, 2000 година. Некои предлози во врска со организацијата на централниот нервен систем за контрола на ерекцијата на пенисот. Невроси. Biobehav. Свештеникот 24, 535-540. Марсон, Л., Мекена, КЕ, 1992 година. Улога за 5-хидрокситриптамин во опаѓачка инхибиција на спиналните сексуални рефлекси. Екстензии. Мозокот Res. 88, 313-318. Марсон, Л., Мекена, КЕ, 1996 година. Групи на клетки на ЦНС вклучени во контролата на мускулите на ишиокавернозните и булбоспонгиозните: студија за трасирање на трансневроните користејќи вирус на псевдорабија. J. Comp. Неврол. 374, 161-179. Meisel, RL, Sachs, BD, 1994 година. Физиологија на машкото сексуално однесување. Во: Knobil, E., Neil, J. (Уред.), Физиологија на репродукцијата, кн. 2, второ издание. Raven Press, Њујорк, стр. 3-96. Мелин, П., Килстром, Ј.Е., 1963 година. Влијание на окситоцинот врз сексуалното однесување кај машките зајаци. Ендокринологија 73, 433-435. Мелис, М.Р., Аргиолас, А., 1995а. Допамин и сексуално однесување. Невроси. Biobehav. Свештеникот 19, 19-38. Мелис, М.Р., Аргиолас, А., 1995б. Донаторите на азотен оксид предизвикуваат ерекција на пенисот и зевање кога се инјектира во централниот нервен систем на машки стаорци. Е. J. Фармакол. 294, 1-9. Мелис, М.Р., Аргиолас, А., 2003 година. Централна окситоцинергична невротрансмисија: цел на лекот за терапија на психогена еректилна дисфункција. Curr. Цели на дрога 4, 55-66. Мелис, М.Р., Аргиолас, А., Геса, ГЛ, 1986 година. Зевање и ерекција на пенисот индуцирано од окситоцин: место на дејство во мозокот. Мозокот Res. 398, 259-265. Мелис, М.Р., Аргиолас, А., Геса, ГЛ, 1987 година. Зевање и ерекција на пенисот индуцирано од апоморфин: место на дејство во мозокот. Мозокот Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Апоморфинот го зголемува нивото на окситоцин во плазмата кај стаорци. Невроси. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Доказите дека апоморфинот предизвикува ерекција на пенисот и зевање со ослободување на окситоцинот во централниот нервен систем. Е. J. Фамакол. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990 година. Ефект на апоморфин врз концентрациите на окситоцин во различни области на мозокот и плазмата на машки стаорци. Е. J. Фармакол. 182, 101-107. Мелис, М.Р., Маури, А., Аргиолас, А., 1994а. Ерекција на пенисот и зевање индуцирана од апоморфин и окситоцин кај недопрени и кастрирани машки стаорци: ефект на сексуални стероиди. Невроендокринологија 59, 349-354. Мелис, М.Р., Станкампијано, Р., Аргиолас, А., 1994б. Ерекцијата на пенисот и проѕевањето предизвикани од паравентрикуларните NMDA инјекции се посредувани од окситоцин. Фармакол. Биохемиски. Однесување. 48, 203-207. Мелис, М.Р., Станкампијано, Р., Аргиолас, А., 1994в. Превенција со NG-нитро-l-аргинин метил естер на ерекција на пенисот и зевање индуцирана од апоморфин и окситоцин: место на дејство во мозокот. Фармакол. Биохемиски. Однесување. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Агонистите на допамин го зголемуваат производството на азотен оксид во паравентрикуларното јадро на хипоталамусот: корелација со ерекцијата на пенисот и проѕевањето. Е. J. Невроси. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Окситоцинот го зголемува производството на азотен оксид во паравентрикуларното јадро на хипоталамусот: корелација со ерекцијата на пенисот и проѕевање. Рег. Пептиди 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Производството на азотен оксид се зголемува во паравентрикуларното јадро на хипоталамусот кај машките стаорци за време на бесконтаксните перилни ерекции и копулација. Е. J. Невроси. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Антагонистот на окситоцин d(CH2)5Tyr(Me)2-Orn8-вазотоцин ја намалува бесконтактната ерекција на пенисот кај машките стаорци. Невроси. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Морфинот кој се инјектира во паравентрикуларното јадро на хипоталамусот спречува бесконтактни ерекции и ја нарушува копулацијата: вклучување на азотен оксид. Е. J. Невроси. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Ефект на возбудлива амино киселина, допамин и антагонисти на рецепторот на окситоцин врз бесконтактната ерекција на пенисот и производството на паравентрикуларен азотен оксид кај машките стаорци. Однесување. Невроси. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Екстра-клеточниот допамин се зголемува во паравентрикуларното јадро на машките стаорци за време на сексуалната активност. Е. J. Невроси. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Антагонизмот на канабиноидните CB1 рецептори во паравентрикуларното јадро на машките стаорци предизвикува ерекција на пенисот. Невроси. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Екстра-клеточните возбудливи амино киселини се зголемуваат во паравентрикуларното јадро на машките стаорци за време на сексуалната активност: главната улога на NMDAрецепторите во еректилната функција. Е. J. Невроси. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, селективен агонист на рецепторот на допамин D4, предизвикува ерекција на пенисот кога се инјектира во паравентрикуларното јадро на машките стаорци. Невроси. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. SR 141716A-индуцирана ерекција на пенисот кај машки стаорци: вклучување на паравентрикуларна глутаминска киселина и азотен оксид. Неврофармакологија 50, 219-228. Мелис, М.Р., Суку, С., Сана, Ф., Масија, МС, Мелис, Т., Енгуехард-Гајфиер, Ц., Хубнер, Х., Гмениер, П., Гуифиер, А., Аргиолас, А., 2006б. PIP3EA и PD168077, два селективни агонисти на рецепторот на допамин D4, предизвикуваат ерекција на пенисот кај машките стаорци: место и механизам на дејство во мозокот. Е. J. Невроси. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Окситоцинот кој се инјектира во вентралната тегментална област предизвикува ерекција на пенисот и го зголемува екстрацелуларниот допамин во јадрото accumbens MR Мелис, А. Argiolas / Невронаука и биохејвиорални осврти 35 (2011) 939-955 955 и паравентрикуларно јадро на хипоталамусот на машки стаорци. Е. J. Невроси. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Улогата на окситоцинот во антиципаторните и консумативните фази на сексуалното однесување на машките стаорци. Во: Јастроу, Х., Фојербах, Д. (Уред.), Прирачник за истражување на окситоцин: синтеза, складирање и ослободување, дејства и форми на лекови. Nova Publishers Inc, Њујорк, САД, стр. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Окситоцинот кој се инјектира во вентралниот субикулум или постеромедијалното кортикално јадро на амигдалата предизвикува ерекција на пенисот и го зголемува екстрацелуларниот допамин во јадрото акумбенс кај машките стаорци. Е. J. Невроси. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010 година. Окситоцинот предизвикува ерекција на пенисот кога се инјектира во вентралниот субикулум: улога на азотен оксид и глутаминска киселина. Неврофармакологија 58, 1153-1160. Мореленд, Р.Б., Накане, М., Донели-Робертс, ДЛ, Милер, Л.Н., Чанг, Р., Учиќ, МЕ, Теранова, м-р, Губинс, ЕЈ, Хелфрич, Р.Ј., Намовиќ, М.Т., Ел-Кухен, ОФ, Мајстори, Ј.Н., Бриони, ЈД, 2004 година. Компаративна фармакологија на стабилни клеточни линии на рецептор слични на човечки допамин Д(2) споени со флукс на калциум преку Галфа(qo5). Биохемиски. Фармакол. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984 година. Ослободување на окситоцин и вазопресин од магноцелуларните јадра ин витро: специфичен олеснувачки ефект на окситоцинот врз сопственото ослободување. J. Ендокринол. 102, 63-72. Марфи, МР, Секл, ЈР, Бартон, С., Чекли, СА, Лајтман, СЛ, 1987 година. Промени во секрецијата на окситоцин и вазопресин за време на сексуалната активност кај мажите. J. Clin. Ендокринол. Метаб. 65, 738-741. Нишимори, К., Јанг, ЛЈ, Гуо, К., Ванг, З., Инсел, ТР, Мацук, ММ, 1996 година. Окситоцинот е потребен за доење, но не е суштински за породување или репродуктивно однесување. Proc. Natl. Acad. Sci. Соединетите Американски Држави 93, 11699-11704. Пател, С., Фридман, С., Чепмен, КЛ, Емс, Ф., Флечер, АЕ, Ноулс, М., Марвуд, Р., Мекалистер, Г., Мајерс, Џ., Кертис, Ј., Кулаговски, Џеј Џеј, Лисон, П.Д., Ридгил, М., Греам, М., Метсон, С., Ретбоун, Д., Ват, А.П., Бристу, ЛЈ, Рупњак, Н.М., Баскин, Е., Линч, Џеј Џеј, Раган, ЦИ , 1997 година. Биолошки профил на L 745,870, селективен антагонист со висок афинитет за допамин D4 рецепторот. J. Фармакол. Екстензии. Ther. 283, 636-647. Педерсен, Калифорнија, Калдвел, ЈД, Јириковски, ГФ, Инсел, ТР, 1992 година. Окситоцинот во мајките, сексуалните и социјалните однесувања, Анали на Академијата на науките во Њујорк, кн. 652. Академијата на науките во Њујорк, Њујорк. Петровиќ, П., Калиш, Р., Сингер, Т., Долан, Р.Ј., 2008 година. Окситоцинот ги намалува афективните проценки на условените лица и активноста на амигдалата. J. Невроси. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995 година. Психофармакологија на сексуалното однесување. Во: Knobil, FE, Kupfer, DJ (Уред.), Психофармакологија: Четвртата генерација на напредок. Raven Press, Њујорк, стр. 742-758. Рајфер, Ј., Аронсон, В.Џ., Буш, ПА, Дори, ФЈ, Игнаро, ЛЈ, 1992 година. Азотниот оксид како посредник за релаксација на кавернозното тело како одговор на неадренергична, нехолинергична невротрансмисија. N. Engl. J. Мед. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991 година. Ефекти на однесувањето на NMDA инјектирано во хипоталамичното паравентрикуларно јадро на стаорецот. Мозокот Res. 550, 220-224. Сакс, БД, 1997 година. Ерекција предизвикана кај машки стаорци со мирис на воздухот од женките во еструсот. Физиол. Однесување. 62, 921-924. Сакс, БД, 2000 година. Контекстуални пристапи кон физиологијата и класификација на еректилната функција, еректилна дисфункција и сексуална возбуда. Невроси. Biobehav. Свештеникот 24, 541-560. Сакс, БД, 2007 година. Контекстуална дефиниција за машката сексуална возбуда. Хорм. Однесување. 51, 569-578. Санчез, Ф., Алонсо, ЈР, Аревало, Р., Бланко, Е., Ајхон, Ј., Васкез, Р., 1994 година. Коегзистенција на NADPH-дијафораза со вазопресин и окситоцин во хипоталамичното магноцелуларно невросекреторно јадра на стаорецот. Cell Tissue Res. 276, 31-34. Сана, Ф., Суку, С., Бои, А., Мелис, М.Р., Аргиолас, А., 2009 година. Инхибиторите на фосфодиестераза тип 5 ја олеснуваат ерекцијата без контакт кај машките стаорци: местото на дејство во мозокот и механизмот на дејство. J. Секс. Мед. 6, 2680-2689. Сафиер, Д., Фелдман, С., 1987 година. Ефекти на септалните и хипокампалните стимули врз невроните на паравентрикуларното јадро. Невронаука 20, 749-755. Сато-Сузуки, И., Кита, И., Огури, М., Арита, Х., 1998 година. Стереотипни одговори на зевање предизвикани од електрична и хемиска стимулација на паравентрикуларното јадро на стаорецот. J. Неврофизиол. 80, 2765-2775. Шуман, Е.М., Медисон, ДВ, 1994 година. Азотен оксид и синаптичка функција. Ен. Свештеникот Невроси. 17, 153-183. Снајдер, Ш, 1992 година. Азотен оксид: прв во новата класа на невротрансмитери? Наука 254, 494-496. Софронију, М.В., 1983 година. Вазопресин и окситоцин во мозокот и 'рбетниот мозок на цицачите. Трендови Невросци. 6, 467-472. Соколоф, П., Шварц, JC, 1995 година. Нови допамински рецептори половина деценија подоцна. Трендови Фармакол. Sci. 16, 270-275. Саутам, Е., Гартвејт, Ј., 1993 година. Патека за сигнализација на азотен оксид-циклична GMP во мозокот на стаорци. Неврофармакологија 32, 1267-1277. Станкампијано, Р., Мелис, М.Р., Аргиолас, А., 1994 година. Ерекција на пенисот и зевање индуцирани од агонисти на 5-HT1c кај машки стаорци: врска со допаминергичен и окситоцинергичен пренос. Е. J. Фармакол. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006 година. Канабиноидниот антагонист на рецепторот CB1 SR 141716A индуцира ерекција на пенисот со зголемување на екстрацелуларната глутаминска киселина во паравентрикуларното јадро на машките стаорци. Однесување. Мозокот Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Стимулацијата на допаминските рецептори во паравентрикуларното јадро на хипоталамусот на машките стаорци предизвикува ерекција на пенисот и го зголемува екстрацелуларниот допамин во јадрото accumbens: вклучување на централниот окситоцин. Неврофармакологија 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Окситоцинот предизвикува ерекција на пенисот кога се инјектира во вентралната тегментална област на машките стаорци: улогата на азотен оксид и цикличниот GMP. Е. J. Невроси. 28, 813-821. Танда, Г., Понтиери, ФЕ, Ди Кјара, Г., 1997 година. Активирање на канабиноиди и хероин на мезолимбичниот пренос на допамин преку заеднички механизам на рецептор на опиоиди mu1. Наука 276, 2048-2050. Танг, Ј., Рампин, О., Калас, А., Факинети, П., Џулијано, Ф., 1998 година. Окситоцинергична и серотонинергична инервација на идентификуваните лумбосакрални јадра кои ја контролираат ерекцијата на пенисот кај машкиот стаорец. Невронаука 82, 241-254. Теодосис, ДТ, 1985 година. Окситоцин-имунореактивни терминали синапсуваат на невроните на окситоцин во супраоптичките јадра. Природа (Лондон) 313, 682-684. Тиндал, Ј.С., 1974 година. Стимулации кои предизвикуваат ослободување на окситоцин. Во: Гајгер, СР, Кнобил, Е., Соер, ВХ, Гриф, Р., Аствуд, И.Б. (Уред.), Прирачник за физиологија. Секта. 7, Ендокринологија, кн. IV. Американско физиолошко друштво, Вашингтон, стр. 257-267. Торес, Г., Ли, С., Ривиер, Ц., 1993 година. Онтогенеза на хипоталамусот азотен оксид синтаза на стаорец и колокализација со невропептиди. Mol. Мобилен. Невроси. 4, 155-163. Ул-Бронер, С., Валтиспергер, Е., Мартинез-Лорензана, Г., Кондес, ЛМ, Фројнд-Мерсиер, МЈ, 2005 година. Сексуално диморфно изразување на местата за врзување на окситоцин во предниот мозок и 'рбетниот мозок на стаорецот. Невронаука 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Компаративна дистрибуција на вазопресин V1b и рибонуклеинските киселини на гласникот на рецепторот на окситоцин во мозокот. Ендокринологија 139, 5015-5033. Ван Ден Пол, А., 1991 година. Имунореактивност на глутамат и аспартат во хипоталамични пресинаптички аксони. J. Невроси. 11, 2087-2101. Вероно-Лонгувил, Ф., Рампин, О., Фројнд-Мерсиер, МЈ, Танг, Ј., Калас, А., Марсон, Л., Мекена, КЕ, Стоекел, МЕ, Беноа, Г., Џулијано, Ф. , 1999 година. Окситоцингична иннервација на автономни јадра кои ја контролираат ерекцијата на пенисот кај стаорци. Невронаука 93, 1437-1447. Винсент, СР, Кимура, Х., 1992 година. Хистохемиско мапирање на синтаза на азотен оксид во мозокот на стаорци. Невронаука 46, 755-784. Вагнер, ЦК, Клеменс, LG, 1993 година. Патека што содржи неврофизин од паравентрикуларното јадро на хипоталамусот до сексуално диморфно моторно јадро во лумбалниот 'рбетниот мозок. J. Comp. Неврол. 336, 106-116. Винслоу, ЈТ, Инсел, ТР, 1991 година. Социјалниот статус кај паровите машки мајмуни верверица го одредува одговорот на однесувањето на централната администрација на окситоцин. J. Невроси. 11, 2032-2038. Wise, RA, Rompre, P.-P., 1989. Мозок допамин и награда. Ен. Свештеникот Психол. 40, 191-225. Witt, DM, Insel, TR, 1994 година. Сексуалното однесување на мажите го активира протеинот сличен на c-fos во невроните на окситоцин во паравентрикуларното јадро на хипоталамусот. J. Невроендокринол. 6, 13-18. Витер, пратеник, 2006 година. Врски на субикулум на стаорец: топографија во однос на колонообразна и ламинарна организација. Behav Brain Res. 174, 251-264. Вудруф, ГН, Фостер, АС, Гил, Р., Кемп, ЈА, Вонг, ЕХ, Иверсен, ЛЛ, 1987 година. Интеракцијата помеѓу MK-801 и рецепторите за N-метил-d-аспартат: функционални последици. Неврофармакологија 26, 903-909. Јамашита, Х., Шигеру, О., Иненага, К., Касаи, М., Уесуги, С., Канан, Х., Канеко, Т., 1987 година. Окситоцинот претежно ги возбудува наводните окситоцин неврони во супраоптичкото јадро на стаорец ин витро. Мозокот Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992 година. Лезии на јадрото парагигантоцелуларис: ефекти врз однесувањето на парење кај машки стаорци. Мозокот Res. 596, 73-79. Јанг, В.С., Шепард, Е., Амико, Ј., Хенигхаузен, Л., ЛаМарка, МЕ, Мекини, Ц., Гинс, ЕИ, 1996 година. Недостатокот на окситоцин од глувци го спречува исфрлањето на млекото, но не и плодноста или породувањето. J. Невроендокринол. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000 година. Зголемување на интракавернозниот притисок на пенисот по инјектирање на агонисти на рецепторите на возбудливи амино киселини во хипоталамичниот паравентрикуларен јадро на анестезирани стаорци. J. Урол.