- 1Centro Interdisciplinario de Neurociencias de Valparaíso, Filozofska fakulteta, Universidad de Valparaíso, Valparaíso, Čile
- 2Departamento de Neurociencia, Facultad de Medicina, Universidad de Chile, Santiago, Čile
- 3Núcleo Milenio NUMIND Biologija nevropsihiatričnih motenj, Universidad de Valparaíso, Valparaíso, Čile
- 4Oddelek za celično in molekularno biologijo, Fakulteta za biološke vede, Pontificia Universidad Católica de Chile, Santiago, Čile
- 5Oddelek za farmacijo in Interdisciplinarni center za nevroznanost, Kemijska fakulteta, Pontificia Universidad Católica de Chile, Santiago, Čile
Moč ciljno usmerjenega vedenja uravnavajo dopaminski nevroni srednjih možganov. Disfunkcije dopaminergičnih vezij opazimo pri odvisnosti od drog in obsesivno-kompulzivne motnje. Prisilno vedenje je lastnost obeh motenj, ki je povezana s povečano nevrotransmisijo dopamina. Dejavnost dopaminskih srednjih možganov v glavnem uravnava homeostatsko delovanje dopamina prek D2 receptorjev (D2R), ki zmanjšujejo sprožitev nevronov, pa tudi sintezo in sproščanje dopamina. Prenos dopamina med drugim uravnavajo tudi heterologni nevrotransmiterji, kot je kappa opioidni sistem. Velik del našega trenutnega poznavanja opioidnega sistema kappa in njegovega vpliva na prenos dopamina izhaja iz predkliničnih živalskih modelov možganskih bolezni. Leta 1988 smo s pomočjo cerebralne mikrodilize pokazali, da akutna aktivacija opioidnih receptorjev Kappa (KOR) zniža sinaptične ravni dopamina v striatumu. Ta zaviralni učinek KOR nasprotuje olajšanju vpliva zlorabe zdravil na sproščanje dopamina, kar vodi k predpostavki uporabe agonistov KOR kot farmakološke terapije za kompulzivni vnos zdravil. Presenetljivo je, da 30 let pozneje antagoniste KOR predlagajo za zdravljenje odvisnosti od drog. Kaj se je lahko zgodilo v teh letih, ki je povzročilo to drastično spremembo paradigme? Zbrani dokazi kažejo, da je vpliv KOR na sinaptične ravni dopamina zapleten, odvisno od pogostosti aktivacije KOR in časovnega razporeda z drugimi dohodnimi dražljaji dopaminskih nevronov, pa tudi zaradi spola in vrst. V nasprotju s svojim akutnim učinkom se zdi, da kronična aktivacija KOR olajša nevrotransmisijo dopamina in vedenje, ki ga posreduje dopamin. Nasprotna dejanja, ki so jih izvajali akutna proti kronični aktivaciji KOR, so bila med izpostavljenostjo zlorabam drog povezana z začetno averzijo in zapoznelim nagrajevalnim učinkom. Obvezno vedenje, ki ga povzroči ponavljajoča se aktivacija D2R, potencira tudi s trajno soaktivacijo KOR, ki je v korelaciji z zmanjšanimi sinaptičnimi ravnmi dopamina in senzibiliziranega D2R. Tako časovno odvisna aktivacija KOR neposredno vpliva na ravni dopamina, ki vplivajo na uglasitev motiviranega vedenja. Ta pregled analizira prispevek kapio opioidnega sistema k dopaminergičnim korelatom kompulzivnega vedenja.
Predstavitev
Dopaminergični sistem v kompulzivnem vedenju
Prisila je nemogoče, da bi se kljub škodljivim posledicam samodejno ustavili, da bi izvedli običajno dejanje z znanim izidom (Robbins et al., 2012). Kompulzivno vedenje je med drugimi psihiatričnimi boleznimi značilnosti obsesivno-kompulzivne motnje (OCD) in odvisnosti od drog. Preverjanje vedenja je zelo pogosto pri obsesivno-kompulzivnih motnjah spektra, za katere je značilno nenehno ponavljanje določene rutine, stereotipno ali ritualno (Williams et al., 2013). Širok spekter običajnega vedenja (npr. Preverjanje, čiščenje, umivanje rok itd.) Se lahko pri bolnikih z OCD spremeni v kompulzivno in na splošno nastane kot odgovor na obsesivne in stisljive misli, ki sprožijo tesnobo. Prav tako iskanje odvisnikov od drog postane zavezujoče kot pri odvisnikih od drog, tako tudi pri OCD igra ključno vlogo tesnoba, ki sproži kompulzivno uživanje drog pri izkušenih uživalcih drog. V obeh pogojih opažamo enake oslabitve pri nagrajevanju in kaznovanju (Figee in sod., 2016), zaradi česar so nekateri avtorji razpravljali o OCD kot vedenjski odvisnosti (Holden, 2001).
Eden od možnih mehanizmov, ki vodi do kompulzivnega vedenja, je uvrščen v teorijo spodbujevalno-senzibilizacijske teorije odvisnosti, to je, da se med zasvojenostjo razvije ojačana motivacija ("želja"), ne da bi pri tem prišlo do povečanega prijetnega ("všečnega") učinka (Berridge et al., 1989; Berridge in Robinson, 2016). Trajna senzibilizacija nagradnega / motivacijskega vezja je vključena v indukcijo spodbudne preobčutljivosti, povezane z iskanjem drog. Nagradno / motivacijsko vezje je sestavljeno iz dopaminskih nevronov srednjega mozga substantia nigra (SN) in ventralno tegmentalno območje (VTA), ki ciljata na hrbtni in ventralni nivo striatuma. Dopaminski nevroni, ki štrlijo v ventralni striatum ali nukleus accumbens (NAc), so tradicionalno povezani s ciljno usmerjenim vedenjem, medtem ko so dopaminski nevroni, ki projicirajo na dorzalni striatum, povezani s pridobivanjem navad (Everitt in Robbins, 2005; Wise, 2009; Yager in sod., 2015; Volkow et al., 2017).
Preobčutljivost kroga nagrajevanja / motivacije pri glodalcih je opaziti kot postopno povečanje lokomotorne aktivnosti, ki ga povzroča večkratna uporaba odmerka, ki lahko povzroči zasvojenost, s fiksnim odmerkom (Pierce in Kalivas, 1997; Robinson in Berridge, 2001). Lokomotorna preobčutljivost je trpežen pojav, saj ga je mogoče opaziti po tednih, mesecih in celo letu po odvzemu zdravila (Robinson in Berridge, 1993). Zgodilo se je, da preobčutljivost vezja nagrad / motivacije prispeva k kompulzivnemu iskanju drog (Robinson in Berridge, 1993). Skladno s tem lokomotorna preobčutljivost olajša kokain, ki sam želi dajati zdravilo, ki želi ponovno vstaviti (De Vries et al., 2002). Poleg tega imajo podgane z razširjenim dostopom do kokaina za samo-administracijo večji lokomotorni odziv na kokain kot podgane z omejenim dostopom (Ferrario et al., 2005). Poleg tega so pri kompulzivnem iskanju drog opažene nevrokemične spremembe, na katerih temelji lokomotorna preobčutljivost za psihostimulante (Steketee in Kalivas, 2011; Giuliano et al., 2019). Ti podatki podpirajo zgodnje predlagano ujemanje med lokomotorno preobčutljivostjo in kompulzivnim iskanjem drog, opaženo pri ljudeh (Robinson in Berridge, 1993; Vanderschuren in Kalivas, 2000). Mehansko gledano večkratno jemanje zdravil za zlorabo senzibilizira mezolimbična dopaminska vezja, kar povečuje dopaminergični nevrotransmisijo. Psihostimulansi, kot kokain ali amfetamini, ki blokirajo transporter dopamina v plazemski membrani (DAT), sprožijo veliko povečanje dopamina v sinaptičnem prostoru v striatumu in NAc in tako aktivirajo lokomocijo (Steketee in Kalivas, 2011). Tako kot pri odvisnosti od drog, tudi preobčutljivost vezja / motivacijskega vezja dopamina prispeva k kompulzivnemu vedenju, ki ga opazimo pri OCD. Dejansko je večkratna aktivacija receptorjev dopamina D2 (D2Rs) dovolj za sprožitev lokomotorne preobčutljivosti in preverjanja vedenja tako pri podganah kot miših (Szechtman in sod., 1998; Szechtman in sod., 1999; Sun et al., 2019). Večkratno dajanje kvinpirola, agonista D2R / D3R, je sprejet model OCD, saj znova povečuje veljavnost obraza, s prirastkom kompulzivnega preverjanja in stereotipnega vedenja, napovedne veljavnosti, kar se kaže v zmanjšanju kompulzivnega vedenja po kroničnem zdravljenju s ponovnim zajemom serotonina. zaviralci (SRI) in konstruirajo veljavnost možganskih struktur, vključenih v ta model, delimo s tistimi v patologiji (Stuchlik et al., 2016; Szechtman in sod., 2017). Če povzamemo, ponavljajoča aktivacija prenosa dopamina bodisi s pomočjo pred-sinaptičnih (sproščanje dopamina) bodisi post-sinaptičnih (aktivacija D2R) mehanizmov vodi v lokomotorno preobčutljivost in kompulzivno vedenje.
Kapio opioidni sistem je eden najpogostejših sistemov, ki nadzira prenos dopamina v vezju nagrajevanja / motivacije. Dokazi kažejo, da prenos kappa-opioidov nasprotuje učinkom dopamina; akutna aktivacija kapio opioidnih receptorjev (KOR) preprečuje lokomotorno aktivnost, ki jo povzročajo psihostimulansi (Gray et al., 1999). Nasprotno, ponavljajoča se aktivacija KOR ohranja in izboljšuje kompulzivno in običajno iskanje drog (Koob, 2013). Uživanje zlorabe drog sproži homeostatski okrepljen prenos opioidov kappa, kar verjetno prispeva k negativnim čustvenim stanjem disforije (Koob, 2013) sprožitev kompulzivne uporabe drog (Chavkin in Koob, 2016). Dejansko je blokada KOR preprečevala ponovno vzpostavitev nikotina zaradi stresa, ne pa tudi z drogami (Jackson in sod., 2013), kokain (Beardsley et al., 2005) in etanol (Sperling in sod., 2010). V skladu s to ugotovitvijo blokada KOR reverzira dopaminergične spremembe v dorsolateralnem striatumu podgan, občutljivih na amfetamin, ne da bi spremenili njihov okrepljeni lokomotorni odziv na zdravilo (Azocar in sod., 2019). Tako se zdi, da sistem KOR povečuje negativno okrepitev in tako povečuje vrednost zdravil. V OCD negativno okrepitev sprožijo obsedenosti, ki krepijo dano prisilo, da se izognejo tej obsedenosti. Čeprav ni bilo neposredno preizkušeno, bi lahko negativna krepitev igrala vlogo pri preobčutljivosti za kvinpirole. Dejansko je D2R vključen v ustvarjanje negativne okrepitve. Na primer, pri miših, ki nimajo dolge izoforme D2R, ni bilo razvitega izogibanja območju, ki je odvzemalo morfine.Smith et al., 2002) in ponavljajoče zdravljenje s kvinpirolom v obdobju abstinence ponovno vzpostavi kokain in heroin, ki iščeta paradigmo o samodejni uporabi, učinek, povezan s preobčutljivim gibanjem na kvinpirol (De Vries et al., 2002), ki predlaga skupne mehanizme med psihostimulantom in preobčutljivostjo, ki jih povzroča kvinpirol. Poleg tega uvedba domače kletke, vendar ne nove kletke, v areno na prostem zmanjšuje lokomotorno preobčutljivost in kompulzivno preverjanje (Szechtman in sod., 2001), kar kaže na to, da lahko varnostni / znani signali tekmujejo z negativnimi okoljskimi namizi, ki podpirajo preobčutljivost. Podobno kot preobčutljivost, ki jo povzroča psihostimulant, večkratna aktivacija KOR olajša lokomotorno preobčutljivost (Escobar in sod., 2017) in kompulzivno preverjanje vedenja (Perreault et al., 2007) povzročeno s ponavljajočim dajanjem kvinpirola. Ali je ta potencialni učinek posledica povečane negativne okrepitve, je treba še razjasniti.
Pred kratkim opravljena temeljita analiza kaže, da je učinek kappa-opioidnega sistema na dopaminergični prenos kompleksen: odvisen je od dopaminove poti (Margolis in sod., 2006; Margolis in sod., 2008) in o času med aktivacijo receptorja KOR in aktivacijo dopaminskega receptorja (Chartoff in sod., 2016). V skladu s to zapletenostjo je bila široka razprava o potencialni terapevtski uporabi ligandov KOR. Predlagano je bilo, da je agonist KOR klinično uporaben med fazo uporabe drog, kar oslabi hiperdopaminergijo, ki jo povzroči zdravilo (Shippenberg et al., 2007). Po drugi strani pa je lahko antagonist KOR uporaben pri zdravljenju odtegnitvenega sindroma, ki ga povzroči povečanje izražanja dinorfin po večkratnem uživanju drog (Wee in Koob, 2010). Skladno s tem se predlaga delni agonist KOR (Béguin in sod., 2012) bi lahko bila terapevtska možnost za zdravljenje kompulzivnega vnosa drog in odtegnitvenih simptomov pri odvisnikih (Chartoff in sod., 2016; Callaghan et al., 2018). V tem pregledu analiziramo časovno / od konteksta odvisno modulacijo dopaminergičnih korelatov vedenjske senzibilizacije in kompulzivnosti.
Anatomske in funkcionalne presledke med opioidnimi in dopaminergičnimi sistemi Kappa v strijtalnih in srednjih možganih
Strijatalne regije
KOR so receptorji, vezani na gi / o beljakovine, močno izraženi v dopaminskem sistemu srednjega mozga (Mansour in sod., 1996). Ti receptorji spadajo v družino opioidnih receptorjev, ki jih sestavljajo mu (MOR), delta (DOR) in kappa (KOR). Endogeni agonisti teh receptorjev so endorfini, enkefalin in dinorfin. V striatumu je dinorfin sintetiziran z dopaminom D1receptor (D1R), ki vsebuje srednje velike nevrone (MSN), ki imajo ponavljajoče se aksone, ki aktivirajo KOR iz istih jeder (Mansour in sod., 1995). Slike elektronske mikroskopije podgana NAc kažejo, da so KOR v glavnem v presinaptičnih strukturah, ki vsebujejo DAT, manjši del KOR pa se nahaja pri dendritih pri imenovanju DAT (Svingos in sod., 2001; Kivell in sod., 2014). Imunofluorescentne študije, ki so značilne za presinaptično-sinaptosomske pripravke iz NAc, kažejo, da KOR in D2Rs prednostno sobivajo v sinaptosomih, ki vsebujejo encim za sintetiziranje dopamina, tirozin hidroksilazo (TH) (Escobar in sod., 2017). Poleg tega so KOR obilne v celičnih telesih NAc in striatumu in se kolokalizirajo z D2R v celični subpopulaciji (Escobar in sod., 2017). Z genetskimi in molekularnimi vpogledi kažejo, da je v terminalih DA opaziti 20% celotne vezave KOR v striatumu (Van't Veer et al., 2013). Poleg tega, Tejeda in sod. (2017) pokazali, da tako D1R kot D2R MSN izražajo KOR z večjo prednostjo za D1R, ki vsebujejo MSN (Tejeda in sod., 2017). Ti anatomski podatki kažejo, da so KOR prisotni pred in postinaptično in uravnavajo nevrotransmisijo dopamina v vezju nagrajevanja / motivacije.
Več eksperimentalnih pristopov kaže, da aktiviranje KOR zavira dopaminske sproščanjee. Akutna aktivacija KOR s sistemsko injekcijo ali lokalno infuzijo agonistov zmanjša zunajcelične ravni dopamina v NAc (Di Chiara in Imperato, 1988; Spanagel et al., 1992; Fuentealba et al., 2006) in hrbtni striatum (Gehrke in sod., 2008). Podpirajo zaviralno delovanje KOR-jev na nevrotransmisijo dopamina, neposredno infuzijo dolgotrajnega in selektivnega antagonista KOR nor-binaltorfimina (nor-BNI) (Broadbear et al., 1994) zvišuje bazalno raven dopamina v NAc (Spanagel et al., 1992) in sproščanje dopamina v dorzalnem striatumu (Azocar in sod., 2019). Končni dokazi o zaviranju tonika KOR dopamina so bili prikazani pri izločilnih miših KOR, ki so pokazale povečano zunajcelično raven dopamina v striatumu in NAc (Chefer et al., 2005). Mehanizmi, ki so odgovorni za zaviranje sproščanja dopamina s KOR, niso popolnoma pojasnjeni. Vendar je dobro znano, da aktivacija KOR vodi do povečanja K + in zmanjšanja prevodnosti Ca2 +, kar povzroči hiperpolarizacijo celic in blokado sproščanja vezikularnega nevrotransmiterja (Bruchas in Chavkin, 2010; Margolis in Karkhanis, 2019).
Poleg tega, in vitro in vivo funkcionalni podatki kažejo, da KOR-ji spreminjajo zunajcelične ravni dopamina z modulacijo aktivnosti DAT. Na primer, aktiviranje KOR-jev v celicah EM4, ki sorazmerno izražajo KOR in DAT, vodi do povečanega vnosa dopamina, merjenega z voltammetrijo (Kivell in sod., 2014). An ex vivo Analiza, ki je uporabila tudi voltammetrijo v razčlenjenih tkivih, je pokazala, da sistemsko injiciranje agonista KOR U-69593 poveča vnos dopamina v NAc (Thompson et al., 2000). Podoben nedavni članek kaže, da nor-BNI blokira povečanje vnosa dopamina v ventralni in dorzalni striatum, ki ga povzroči akutna sistemska injekcija MP1104, mešanega agonista opioidnih receptorjev Kappa / Delta (Atigari in sod., 2019). Kljub temu učinek aktivacije KOR na vnos dopamina še ni povsem razjasnjen. Sistemsko dajanje delnega agonista nalmefena KOR je odvisno od zmanjšanja odmerka striatalnega dopamina, količinsko določeno s hitro skenirano ciklično voltammetrijo (FSCV) (Rose in sod., 2016). Z uporabo mikrodialize neto toka pri odraslih samcih podgan je blokado KOR spremljalo povečanje ekstrakcijske frakcije (Ed), kar je posredno merilo vnosa dopamina (Chefer et al., 2006; Azocar in sod., 2019), kar kaže, da tonično aktiviranje KOR izvaja zaviralno kontrolo aktivnosti DAT (sprejem dopamina). Ti rezultati poudarjajo kompleksno vlogo endogenega delovanja KOR pri zaužitju dopamina za nadzor nadcelične ravni dopamina. Pristopi višje časovne ločljivosti, kot je FSCV, niso pokazali učinka KOR na vnos dopamina (Ebner in sod., 2010; Ehrich in sod., 2015; Hoffman et al., 2016), kar kaže na to, da KOR, ki krepi aktivnost DAT v strijatalnih regijah, potrebuje inkubacijsko obdobje., KOR-posredovano povečanje DAT aktivnosti je mogoče razložiti s povečanjem števila DAT na celičnih membranah, ki jih povzroči aktivacija KOR, kot so poročali v strijatalnih sinaptosomih in celicah vrstice (Kivell in sod., 2014).
Regije srednjih možganov
Avtoradiografski testi, opravljeni v srednjem možganu podgane, kažejo pomembno vezavo za KOR na rostrocaudalni osi SN in VTA (Speciale in sod., 1993). Po drugi strani podatki z elektronsko mikroskopijo kažejo, da terminali, ki vsebujejo dinarfin, sinaptirajo neposredno na TH pozitivne dendrite v SN in VTA (Sesack in Pickel, 1992), kar kaže na to, da se KOR lokalizirajo v somatodendritičnih oddelkih dopaminskih nevronov. Striatalni D1R, ki vsebujejo MSN, so eden od vnosov dinnorfina v nevrone srednjega mozga. Zanimivo je, da blokada KOR ne spremeni zaviralnega učinka D1R-MSN na nevrone Vpam dopamina, kar kaže, da to inhibicijo posreduje GABA (Edwards in sod., 2017). KOR modulirajo somatodendritične odzive dopaminskih nevronov srednjega možganov. Elektrofiziološke študije kažejo, da aktivacija KOR-ov v hipopolarizaciji VTA in zniža hitrost spontanega izgorevanja dopaminskih nevronov (Margolis in sod., 2003). Posledično infuzija agonistov KOR zmanjša somatodendritski dopaminski izliv (Smith et al., 1992; Dalman in O'Malley, 1999). Vendar se zdi, da je ta inhibitorni učinek KOR na dopaminskih nevronih odvisen od vezja. Infuzija kappa-opioidnih agonistov v VTA zmanjšuje sproščanje dopamina v medialni prefrontalni skorji (mPFC) (Margolis in sod., 2006), vendar ne v NAc (Devine et al., 1993; Margolis in sod., 2006). Poleg tega, Margolis in sod. (2006) ugotovili, da KOR zavirajo VTA dopaminske nevrone, ki štrlijo na mPFC in bazolateralno amigdalo, ne pa na tiste, ki štrlijo na NAc. Istega leta je dr. Ford in sod. (2006) je pokazal, da uporaba agonistov KOR v mišjih rezinah VTA povzroča večji zunanji tok v dopaminskih nevronih, ki štrlijo na NAc v primerjavi s tistimi, ki projicirajo na bazolateralno amigdalo, kar kaže, da KOR izvajajo večjo inhibicijo dopaminskih nevronov, ki štrlijo v NAc kot do amigdale. Poleg tega se z aktivacijo KOR zmanjša amplituda vzbujanja (Margolis in sod., 2005) in zaviralno (Ford in sod., 2007) postsinaptični tokovi v nevrone srednjih možganov. Razlike med vrstami in kompleksnimi efektnimi proekcijami VTA na mPFC in NAc (Van Bockstaele in Pickel, 1995; Carr in Sesack, 2000) zahtevno je ugotoviti, ali KOR selektivno zavirajo nekatere populacije nevronskih dopaminov v VTA. Kljub temu tukaj povzeti podatki kažejo, da so KOR v soma in terminalih dopaminskih nevronov, pa tudi v vhodih, ki jih uravnavajo, tako odlično postavljeni za nadzor sinaptične aktivnosti srednjih možganov dopaminskih nevronov.
Vloga KOR-jev pri nadzoru nevrotransmisije dopamina pri preobčutljivosti in kompulzivnem vedenju, ki ga povzročajo psihostimulanti
Zasvojenost z drogami je postopek, ki vključuje prvotno iskanje impulzov, povezanih z njihovimi pozitivnimi učinki. Po drugi strani je kompulzivnost osebnostna lastnost, ki jo je mogoče opaziti pri odvisnikih od drog. Predlagano je bilo več nevroadaptacij na dopaminergičnih poteh, ki bi predstavljale kompulzivno iskanje drog in vnos po večkratni izpostavljenosti zlorabam drog (Everitt in Robbins, 2005; Koob in Volkow, 2016). Ena od predlaganih hipotez, ki spodbuja kompulzivni vnos drog, je preobčutljivost njegovih negativno okrepljivih učinkov (Koob, 2013). Inhibicijski nadzor kapio opioidnega sistema ob sproščanju dopamina lahko prispeva k negativno okrepljenim lastnostim zlorabe zdravil. Vendar se zdi, da so posledice aktivacije KOR na nevrotransmisiji dopamina in kompulzivnih zdravil zapletene in na videz protislovne. Sprostitev sproščanja dopamina, ki ga povzročata amfetamin in kokain, zmanjšuje sočasno dajanje agonistov KOR (Heidbreder in Shippenberg, 1994; Maisonneuve in sod., 1994; Thompson et al., 2000) in celo zmanjšati samoupravljanje kokaina (Negus in sod., 1997). Poleg tega KOR-ji sprožijo zaviralno povratno informacijo o sproščanju dopamina mezolimbične poti kot odgovor na trajno aktivacijo post-sinaptičnega D1R, kot se pojavi pri ponavljajoči se izpostavljenosti psihostimulantom (Cole in sod., 1995; Nestler, 2001). Paradoksalno je, da lahko aktiviranje KOR-ov tudi olajša sproščanje dopamina na poti nagrajevanja / motivacije (Fuentealba et al., 2006; Fuentealba et al., 2007) in porabo psihostimulantov (Wee et al., 2009). Fuentealba in sod. (2007) pokazali, da je po štirih dneh dajanja U69593, agonista KOR, povečano sproščanje dopamina z amfetaminom v NAc. Pred kratkim se je pokazalo, da blokiranje KOR-jev spremeni spremembe v sproščanju in dojemanju dopamina v dorzalnem striatumu, ki se zgodijo med lokomotorno preobčutljivostjo, ki jo povzroča amfetamin (Azocar in sod., 2019). Ti podatki na splošno kažejo, da lahko aktiviranje KOR prispeva tudi k izboljšanju pozitivnih lastnosti zlorabe drog (Chartoff in sod., 2016).
Poleg tega se zdi, da aktiviranje KOR prispeva tudi k kompulzivnemu iskanju drog; Blokada KOR zmanjšuje kokain (Wee et al., 2009), heroin (Schlosburg in sod., 2013) in metamfetamina (Whitfield in sod., 2015) vnos pri podganah z neomejenim dostopom do zdravila (Wee in drugi, 2009). Ta učinek se kaže tudi pri iskanju drog, ki jih povzroča stres. Na primer, izločilne miške KOR po stresnem plavanju niso pokazale kokaina,McLaughlin in sod., 2006a). Blokiranje KOR zmanjšuje prednost nikotinskega mesta, ki jo povzroči prisilna izpostavljenost plavalnemu stresu (Smith et al., 2012). Zanimivo je, da blokiranje KOR zmanjšuje iskanje kokaina in nikotina, ki ga povzroča stres, vendar ni vplivalo na iskanje zaradi izziva zaradi drog (Beardsley et al., 2005; Jackson in sod., 2013). Zdi se, da ta učinek KOR, ki ga povzroča stres, posreduje vezje za nagrajevanje / motivacijo (Shippenberg et al., 2007; Wee in Koob, 2010). V elegantni študiji, ki sta jo opravili dr. Kauer in njena skupina, je bilo dokazano, da blokiranje KOR-ov v VTA bodisi pred ali po akutnem stresu zavira ponovno vnovično iskanje kokaina, kar je učinek, povezan z reševanjem dolgoročnih oz. potenciranje zaviralnih sinapsov v dopaminskih nevronih (Graziane in sod., 2013; Polter in sod., 2014).
Zdi se, da je olajšanje vnosa psihostimulantov, ki jih izvajajo KOR, odvisno od časovnega obdobja glede izpostavljenosti drogam. Uporaba agonista KOR U50488 1 h pred izpostavljenostjo kokainu potencira tako prednost kokainskega mesta kot tudi relativno sproščanje dopamina, ki ga kokain sproži v NAc, medtem ko opazimo nasprotne učinke, če jih damo 15 min pred (McLaughlin in sod., 2006a; Ehrich in sod., 2014). Uporaba intrakranialne samo-stimulacije Chartoff in sod. (2016) opazili, da ima agonist KOR Salvinorin A začetni averzivni učinek in upočasnjen nagrajevalni učinek, ki ga spremljata zmanjšanje in povečanje sproščenega sproščanja dopamina v NAc. Vsi ti podatki kažejo na časovno odvisen vpliv aktivacije KOR na koristne lastnosti kokaina in kažejo na stresno posredovano aktivacijo KOR kot ključnega akterja za razvoj kompulzivnega iskanja drog.
Lokomotorna preobčutljivost in kompulzivno vedenje, ki ga povzroča kvinpirol
Dejstva, da dopaminski sistem sodeluje pri ustvarjanju senzibilizacije in kompulzivnosti, okrepi vedenje pri glodalcih, zdravljenih z agonistom D2R-kinpirola. Na kratko, D2R so receptorji, povezani s Gi, široko izraženi v nagradnem / motivacijskem vezju; izražajo se somatodendritično in na aksonskih terminalih dopaminskih nevronov (Sesack et al., 1994), njegova aktivacija pa zniža zunajcelične ravni dopamina (Imperato in Di Chiara, 1988). V striatumu se D2R-i nahajajo tudi postsinaptično na srednjih bodičavih nevronih (Sesack et al., 1994) in njegova aktivacija zavira indirektno pot, ki omogoča lokomotorno aktivnost.
Dr. Henry Szechtman je začel preučevati učinke kvinpirola na vedenje podgan konec desetletja 1980. Njihove začetne ugotovitve so pokazale, da ima akutno dajanje kinpirola od odmerka odvisen učinek na gibljivo aktivnost. Pri majhnih odmerkih (0.03 mg / kg) zmanjša gibalno aktivnost, pri večjih odmerkih (> 0.5 mg / kg) pa se poveča. (Eilam in Szechtman, 1989). Ti učinki so povezani z aktiviranjem visoko afinitetnih presinaptičnih D2R in z nizkoafinitetnimi postsinaptičnimi D2Rs (Usiello et al., 2000). Nepričakovano ponavljajoče (vsak drugi dan) kinpirol sproži postopno in trajno povečanje lokomocije, podobno lokomotorni preobčutljivosti, ki jo povzročajo psihostimulansi (Szechtman in sod., 1993; Szechtman in sod., 1994). Izkazalo se je, da je učinek preobčutljivosti na lokomotor odvisen od D2R, saj miši, ki imajo pomanjkanje tega receptorja, ne razvijejo lokomotorne preobčutljivosti na kvinpirol (Escobar in sod., 2015).
V začetku 90-ih sta Szechtman in Eilam poročala, da so podgane skupaj z lokomotorno preobčutljivostjo razvile stereotipno vedenje, ki se okrepi z vsakim dajanjem kvinpirola (Eilam in Szechtman, 1989; Szechtman in sod., 1993). Danes je ponavljajoče dajanje kvinpirola validiran model za OCD (Szechtman in sod., 1999; Szechtman in sod., 2001; Eilam in Szechtman, 2005; Stuchlik et al., 2016; Szechtman in sod., 2017), ki temelji na opažanju, da vedenje podgan postaja vse bolj strukturirano in neprožno, kar spominja na obredno vedenje, značilno za kompulzivno preverjanje vedenja (Szechtman in sod., 1998; Szechtman in sod., 2017). Nedavne študije kažejo, da ponavljajoči kvinpirol pri miših sproži tudi kompulzivno vedenje, kot je kompulzivno preverjanje (Sun et al., 2019), vedenjska neprožnost in kompulzivno žvečenje (Asaoka in sod., 2019), slednji je z blokado D2Rs vrnil v striatum in nadalje podpiral, da je za sprožitev kompulzivnega vedenja potrebna večkratna aktivacija D2R. Podatki skupaj kažejo na ključno vlogo D2Rs v srednjih možganskih dopaminskih poteh, da sprožijo lokomotorno preobčutljivost in kompulzivnost. Ponavljajoče jemanje kvinpirola napolni stereotipno vedenje, ki ga povzroča kokain (Thompson et al., 2010) in lokomotorne učinke amfetamina (Cope in sod., 2010), krepitev ideje, da aktivacija D2R temelji na preobčutljivosti, ki jo povzroča psihostimulant, in predlaga skupni mehanizem med preobčutljivostjo, ki jo povzroča kvinpirol in psihostimulansi. Zanimivo je, da se zdi, da je občutljiv učinek ponavljajoče se aktivacije D2R močnejši od učinka psihostimulantov, saj vsaka podgana, ki se zdravi s kvinpirolom, razvije lokomotorno preobčutljivost (Escobar in sod., 2015), medtem ko približno šestdeset odstotkov podgan preobčutljivo na amfetamin (Escobar in sod., 2012; Casanova in sod., 2013).
Vedenjsko preobčutljivost, ki jo povzroči večkratna aktivacija D2R, spremljajo prilagoditve v vezju nagrajevanja / motivacije. Podgane, ki so preobčutljive za kvinpirol, imajo nižji dopaminergični tonus NAc, ki ga opazimo kot zmanjšano bazalno (Koeltzow in sod., 2003) in stimulirano tonično in fazno sproščanje dopamina (Escobar in sod., 2015), kar kaže na zmanjšano zmogljivost sproščanja dopamina v vezju srednjega možganov. Sinaptične ravni dopamina v NAc nadzira aktivnost obeh, DAT in dopaminskih nevronov (Goto in Grace, 2008), ki vivo sestoji iz tonika in porušitve (Wilson et al., 1977; Grace in Bunney, 1980). Prejšnja poročila kažejo, da podgane, preobčutljive za kvinpirol, kažejo manjše število dopaminskih nevronov v toniku in izbruhu v VTA (Sesia in sod., 2013). Ti podatki skupaj kažejo, da je zmanjšanje sproščanja dopamina po preobčutljivosti za kvinpirol posledica zmanjšanja celotne aktivnosti dopaminskih nevronov. Kompulzivno vedenje in senzibilizirana lokomotorna aktivnost, ki sta jo povzročila ponavljajoče zdravljenje s kvinpirolom, sta lahko posledica občutljivosti D2R, zaradi zmanjšanega dopaminergičnega tona NAC. Podgane, občutljive na kvinpirol, podgane kažejo povečanje vezave dopamina D2R (Culver in sod., 2008) in povečanje afinitetnega stanja teh receptorjev (Perreault et al., 2007), ki podpira to hipotezo.
Interakcije KOR-dopamina v kompulzivnem vedenju, ki ga povzroča kvinpirol
Začetne študije o vlogi KOR pri kompulzivnem vedenju, ki ga povzroča D2R, so prišle tudi iz Szechtmanovega laboratorija. Ta skupina je preučevala sočasno uporabo agonista KOR U69593 s kvinpirolom na gibalno aktivnost. Natančneje, avtorji so podganam dajali subkutane injekcije z mešanico U69593 in kvinpirolom, dokler ni bilo dokončanih 8 do 10 injekcij. V nasprotju s hipolokomotornim učinkom samega U69593 so pri sočasni uporabi z nizkimi (presinaptičnimi) in visokimi (postsinaptičnimi) odmerki kinpirola opazili hiperlokomot. U69593 je spremenil hipolokomotorni učinek presinaptičnega odmerka kinpirola v hiperlokomotorijo in povečal hiperlokomotorni učinek postsinaptičnega odmerka kinpirola (Perreault et al., 2006). Koaktivacija KOR-jev je tudi pospešila indukcijo lokomotorne preobčutljivosti in potencirala učinek aktiviranja D2Rs, saj največja lokomotiva, dosežena z dvojnim zdravljenjem, podvaja lokomotorni učinek, ki ga povzroči samo kvinpirol (Perreault et al., 2006; Escobar in sod., 2017). Soaktivacija KOR-jev tudi pospeši pridobivanje kompulzivnega vedenja preverjanja (Perreault et al., 2007). Ti potencirajoči učinki KOR na vedenje, ki ga povzroča kvinpirol, zahtevajo večkratno aktivacijo KOR. Dejansko akutna injekcija agonista KOR U69593 še ni spremenila lokomotorne aktivnosti pri podganah, občutljivih na kvinpirol (Escobar in sod., 2017). Mehanizem KOR, ki potencira D2R-senzibilizacijo, ni znan. Ena od možnosti je, da sam endogeni kappa opioidni sistem posreduje z D2R odvisno senzibilizacijo. Vendar pa je bila ta možnost zavržena, saj so pokazali, da pred-dajanje norBNI-ja ni spremenilo lokomotorne preobčutljivosti na kvinpirol, kar kaže na to, da se dinnorfin ne sprošča po aktivaciji D2R (Escobar in sod., 2017). Ti podatki ne izključujejo, da bi lahko imel dinorfin vlogo pri preobčutljivosti kompulzivnega vedenja, na primer stres sprošča sproščanje dinorfin in aktiviranje KOR, kar olajša kompulzivno vedenje (McLaughlin et al., 2003; McLaughlin in sod., 2006a; McLaughlin in sod., 2006b).
Vmesni pogovor med D2R in KOR je zapleten in zdi se, da je odvisen od tega, ali je aktivacija obeh receptorjev naključna ali časovno ločena. Anatomski podatki kažejo, da se lahko prekrivanje med D2Rs in KOR pojavi presinaptično v aksonih in soma dopaminskih nevronov, pa tudi postinaptično v MSNs striatuma. Čeprav ne izključuje vloge KOR-ov, ki se nahajajo na aksonih drugih nevrokemičnih sistemov, anatomski podatki močno kažejo na neposredno vlogo KOR-jev, ki urejajo D2R. Akutna ali ponavljajoča se z aktivacijo KOR zmanjša inhibitorna D2Rs na dopaminskih nevronih. Elektrofiziološke študije so pokazale, da akutna aktivacija KOR v dopaminskih nevronih VTA in SN zavira zaviralno postsinaptični tok, ki ga posreduje D2R, učinek, ki ga posredujejo pred in postsinaptični mehanizmi, ker KOR zmanjšuje sproščanje dopamina in dinnorfin preprečuje zaviralni učinek dopamina v kopeli (Ford in sod., 2007). Nevrokemijske študije so pokazale, da večkratna aktivacija KOR blokira zaviranje sproščanja dopamina v NAc, ki ga povzroči D2R (Fuentealba et al., 2006). Poleg tega akutna aktivacija D2R in KORs zmanjša inhibicijo sproščanja dopamina v NAc v primerjavi z učinkom posameznega receptorja (Escobar in sod., 2017). Tako presinaptični KOR ne delujejo aditivno ali v sinergiji s presinaptičnimi D2R, nasprotno pa KOR bodisi zavirajo bodisi zavirajo D2R zaviralni učinek. Ta mehanizem lahko razloži lokomotorni učinek akutnega odmerka agonistov KOR, ki je sočasno do majhnega odmerka kinpirola (Perreault et al., 2006).
Nedavna študija kaže, da aktiviranje KOR v VTA posreduje kompulzivno vedenje, merjeno kot vedenje inhibicija in zakopanje marmorja (Abraham in sod., 2017), kar krepi idejo, da je aktivacija KOR-jev res sprožilec kompulzivnosti. Podatke objavil Margolis in sod. (2006); 2008) kažejo, da naj bi potekalo interakcijo KOR in D2R na dopaminskih nevronih, ki ciljajo na mPFC (Margolis in sod., 2006; Margolis in sod., 2008). Ne glede na to, Ford in sod. (2006); 2007) ugotovili, da zaviranje KOR DPS, posredovanega z D2R, poteka na dopaminskih nevronih, ki ciljajo na NAc (Ford in sod., 2006; Ford in sod., 2007). Ti podatki skupaj kažejo, da bi lahko interakcija KOR z D2R na somatodendritičnem oddelku dopaminskih nevronov nastala kot rezultat prekrivanja istega dopaminskega nevrona. Ali se to zgodi v mezolimbični ali mezokortikalni projekciji, je še vedno sporno.
KOR je bilo ugotovljeno v MSN-jih NAc (Escobar in sod., 2017; Tejeda in sod., 2017), kar kaže na to, da potenciranje kompulzivnega vedenja, ki ga povzroča D2R, lahko nastane tudi z neposrednim delovanjem na ciljne celice dopaminskih nevronov. V zvezi s tem je treba omeniti, da večkratno dajanje U69593 poveča količino D2R v stanju z visoko afiniteto (Perreault et al., 2007). Nevrokemični podatki kažejo, da je zmanjšana zunajcelična raven dopamina povezana s preobčutljivostjo za D2R. Koaktivacija KOR-jev se še ne zmanjša, da se je zunajcelična raven dopamina v NAc že zmanjšala s ponavljajočo se aktivacijo D2Rs (Escobar in sod., 2017), ki izključuje vlogo presinaptičnih KOR, ki s tem mehanizmom pospešujejo ali potencirajo preobčutljivost D2R v NAc. Zato KOR sprožijo počasne molekularne mehanizme, ki še dodatno senzibilizirajo nevrokemične in vedenjske učinke D2R, kar kaže na to, da bi lahko bilo povečanje preobčutljivosti za lokomotiranje posledica prilagodljivega postsinaptičnega, ne pa presinaptičnega učinka. V zvezi s tem lahko večkratna aktivacija KOR sproži inhibicijo D2R indirektnega progastega steznega prehoda, ki preklopi ravnovesje D1R / D2R na D1R in povzroči kompulzivnost (Slika 1).
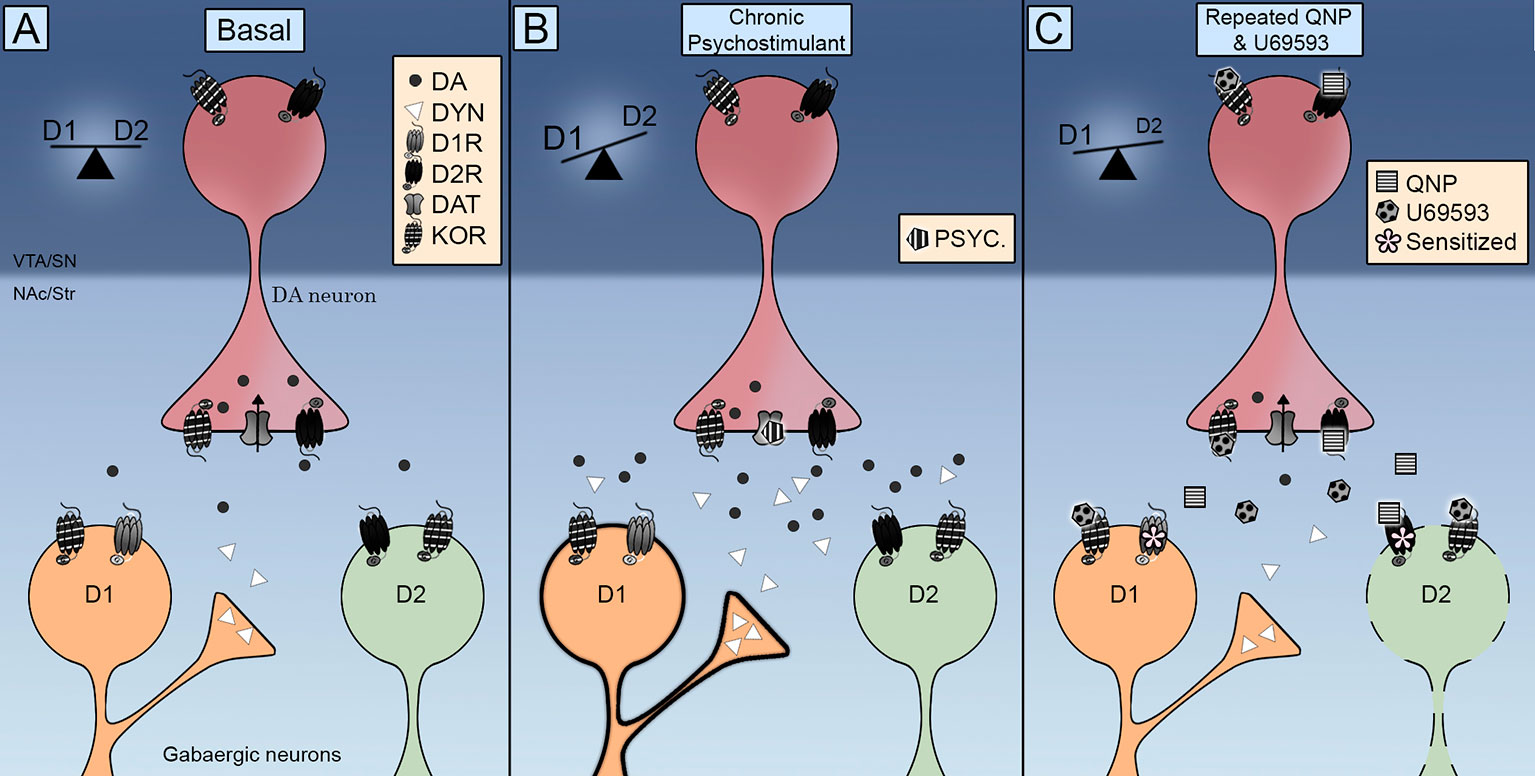 Slika 1 Celostna shema krmiljenja opioidnih receptorjev Kappa (KOR) na neposrednih (D1R) in indirektnih (D2R) progastih poteh. () KOR se nahajajo pred-sinaptično na dopaminskih terminalih in postavijo sinaptično v srednje velikih nevronih (MSN). Njegova aktivacija nadzoruje ravni zunajcelične dopamina, njegova lokalizacija pa spodbuja interakcijo z dopaminskim transporterjem (DAT) in dopamin D2 receptorji. (B) Ponavljajočo se izpostavljenost psihostimulantu spremlja povečanje tako zunajcelične ravni dopamina kot dinorfin. Aktivacija receptorjev D1 in D2 preusmeri ravnotežje na direktno pot D1R, ki spodbuja preobčutljivost lokomotorjev. (C) Sočasna uporaba kinpirola in U69593 spremlja zmanjšanje ravni zunajcelične dopamina. Sočasna aktivacija receptorjev KOR in D2 oslabi indirektno pot D2, kar povzroči kompulzivno vedenje.
Slika 1 Celostna shema krmiljenja opioidnih receptorjev Kappa (KOR) na neposrednih (D1R) in indirektnih (D2R) progastih poteh. () KOR se nahajajo pred-sinaptično na dopaminskih terminalih in postavijo sinaptično v srednje velikih nevronih (MSN). Njegova aktivacija nadzoruje ravni zunajcelične dopamina, njegova lokalizacija pa spodbuja interakcijo z dopaminskim transporterjem (DAT) in dopamin D2 receptorji. (B) Ponavljajočo se izpostavljenost psihostimulantu spremlja povečanje tako zunajcelične ravni dopamina kot dinorfin. Aktivacija receptorjev D1 in D2 preusmeri ravnotežje na direktno pot D1R, ki spodbuja preobčutljivost lokomotorjev. (C) Sočasna uporaba kinpirola in U69593 spremlja zmanjšanje ravni zunajcelične dopamina. Sočasna aktivacija receptorjev KOR in D2 oslabi indirektno pot D2, kar povzroči kompulzivno vedenje.
Spolne razlike med interakcijami KOR-dopamina v kompulzivnem vedenju
Klinične raziskave so pokazale razlike med spoloma v kompulzivnem vedenju, vključno z vsiljevanjem drog. Pri moških v primerjavi z ženskami opazimo zgodnejši pojav simptomov OCD (Mathis in sod., 2011), pri ženskah je večja razširjenost simptomov kontaminacije in čiščenja (Labad in sod., 2008). Kar zadeva razlike med spoloma v odvisnosti od drog, klinični dokazi kažejo, da čeprav je uporaba mamil bolj razširjena pri moških, ženske kažejo hitrejše napredovanje kot moški v kompulzivno iskanje drog (Hernandez-Avila in sod., 2004; Fattore in Melis, 2016).
V zadnjem času so predklinični dokazi močno poudarili nevrobiološke podlage, na katerih temeljijo razlike med spoloma pri zlorabi drog, opažene v kliničnih študijah (Becker in Chartoff, 2019). Zgodnja opazovanja z uporabo mikrodialize fluksa brez mrež so pokazala, da se zunajcelična koncentracija dopamina v dorzalnem striatumu med estrostnim ciklom spreminja z višjimi stopnjami v proestru in estrusu v primerjavi z diestrusom. Še več, medtem ko ovariektomija zmanjšuje zunajcelično koncentracijo dopamina v stripah pri samicah podgan, vendar kastracija samcev podgan ne spremeni stripske celice dopamina v stripu (Xiao in Becker, 1994), kar kaže na pomembno vlogo hormonov jajčnikov pri aktivnosti dopamina. Poleg tega ženski hormoni uravnavajo odziv na psihostimulance. Zgodaj in vitro Poskusi so pokazali, da estradiol in progesteron obnavljata sproščanje dopamina z amfetaminom iz strijatalnega tkiva, pridobljenega pri jajčnikih podganah (Becker in Ramirez, 1981). V zadnjem času so ciklične raziskave ciklične voltammetrije hitro skeniranja pokazale, da imajo ženske večje električno stimulirano sproščanje in absorpcijo dopamina v primerjavi z moškimi (Walker et al., 2000). Te razlike med spoloma v nevrotransmisiji dopamina lahko povzročijo večje iskanje kokaina in amfetamina pri ženskah. (Roberts et al., 1989; Cox et al., 2013).
Ureditev KOR na zunajcelični ravni dopamina kaže tudi spolne razlike (Chartoff in Mavrikaki, 2015). Z uporabo intrakranialne samo-stimulacije in ciklične voltammetrije, Conway in sod. (2019) pokazali, da nižjo občutljivost na akutni anhedonski učinek agonista KOR, opaženo pri samicah podgan, v primerjavi z oslabljeno inhibicijo stimuliranega sproščanja dopamina v NAc (Conway in sod., 2019). Domnevajo, da estradiol prispeva k močnejšemu zaviranju sproščanja dopamina, ki so ga opazili pri podganah po aktivaciji KOR (Abraham in sod., 2018). Medtem ko je bila preiskava navzkrižne povezave med KOR-jem in dopaminsko signalizacijo preučena pri moških (Tejeda in Bonci, 2019), raziskav o tej interakciji in njenem vplivu na proces zasvojenosti pri ženskah primanjkuje (Chartoff in Mavrikaki, 2015). Pri samskih podganah je akutna uporaba agonista KOR U69593 oslabila hiperlokomocijo, povzročeno s kokainom, pri podganah, kontroliranih in ovariektomiranih. Zanimivo je, da je U69593 ponavljajoča uporaba oslabila hiperlokomocijo, ki jo povzroča kokain, na estradiol odvisen način (Puig-Ramos in sod., 2008). Ti podatki kažejo, da estradiol napolni delovanje KOR pri samicah podgan, kar je lahko povezano s spolnimi razlikami v stresnem odzivu (Puig-Ramos in sod., 2008). Ali pri samskih podganah ponavljajoča se aktivacija KOR olajša sproščanje stripam dopamina, opaženo pri samcih, ni vprašanje.
Čeprav pri ženskah opazimo olajšanje sproščanja dopamina, ki ga povzroča psihostimulant, v primerjavi z moškimi podganami, spolne razlike v mehanizmih dopamina, na katerih temelji senzitizacija lokomotorjev na amfetamin, niso bile popolnoma razjasnjene (Becker, 1999). Ponavljajoča izpostavljenost amfetaminu povzroči večjo lokomotorno aktivnost pri obeh, mladostnikih (Mathews in McCormick, 2007) in odrasle podgane (Milesi-Hallé in sod., 2007), pri podganah samic, ki so po večkratni izpostavljenosti amfetaminu pokazale močnejšo lokomotorno preobčutljivost. Neonatalna aktivacija receptorja D2 je potencirala vedenjsko preobčutljivost, ki jo povzroča amfetamin, samo pri samicah podgan (Brown et al., 2011). Kot smo že omenili, so pri samskih podganah opazili, da večkratna izpostavljenost agonistu D2 povzroča lokomotorno preobčutljivost in kompulzivno podobno vedenje (Dvorkin in sod., 2006). Še več, soaktivacija KOR potencira lokomotorno preobčutljivost, ki jo povzroči ponavljajoča se izpostavljenost kvinpirolu, kar olajša inhibicijski nadzor D2 receptorjev ob sproščanju DA v NAc (Escobar in sod., 2017). Razlike med spoloma, kot je opažena manjša občutljivost na zaviralni učinek KOR na sproščanje dopamina pri ženskah (Conway in sod., 2019) lahko predstavljajo različen prispevek KOR k kompulzivnemu iskanju drog.
Sklepi
Kako KOR modulirajo dopaminsko signalizacijo, da razvijejo motivirano vedenje in kdaj povzroči občutljivo kompulzivno vedenje? Anatomski podatki kažejo, da so KOR-ji odlično nameščeni za nadzor sinaptične aktivnosti dopaminskih nevronov srednjega mozga. Funkcionalni podatki kažejo, da KOR nadzirajo delovanje DAT in D2R ter hitrost streljanja dopaminskih nevronov. Prvotni dokazi, ki kažejo, da akutna aktivacija KOR zmanjšuje sproščanje dopamina, ki ga povzročajo zlorabe zdravil, so dopolnili s podatki, ki kažejo, da večkratna aktivacija KOR olajša sproščanje dopamina in kompulzivno iskanje zdravil. Dopaminska signalizacija uravnoteži neposredne in indirektne izhodne poti iz strij (Slika 1A). Bodisi kronična stimulacija s psihostimulansi, ki poveča sproščanje dopamina in aktivira tako D1R kot D2R (Slika 1B) ali kvinpirole, ki aktivirajo samo D2R (Slika 1C) povzroči preobčutljivost lokomotorja in kompulzivno vedenje po izčrpani indirektni poti D2R, s čimer se ravnotežje preusmeri na direktno pot D1R. Prenos KOR se med kroničnim vnosom psihostimulantov poveča s povečanjem dinnorfina v striatalnih D1 nevronih (Slika 1B). Izboljšan prenos KOR se v farmakološkem modelu OCD pospeši z dajanjem U69593. Ta sočasna aktivacija KOR še dodatno oslabi indirektno pot D2 (Slika 1C). Prihodnje raziskave je treba izvesti za popolno razjasnitev posledic aktivacije KOR na aktivnost DAT, razumevanje vloge endogenega sistema KOR v kompulzivnosti, ki ga povzroča kvinpirol, in določitev prispevka sistema KOR k spolnim razlikam, opaženim v kompulzivnem vedenju.
Prispevki avtorjev
AE, MA in JF so prispevali k zasnovi rokopisa. AE in JF sta napisala prvi osnutek rokopisa z vložkom MA. MA in JC sta prispevala k kritičnemu pregledu in urejanju rokopisa. Vsi avtorji so ga odobrili za objavo.
Financiranje
Delo avtorjev, navedenih v tem pregledu, je podprto s številkami donacij FONDECYT: 1110352 in 1150200 MA; 1141088 do JF; Donacija DIPOG 391340281 podjetju JF; FONDECYT podoktorski sodelavci 3170497 za JC in 3190843 za AE.
Navzkrižje interesov
Avtorji izjavljajo, da je bila raziskava izvedena v odsotnosti komercialnih ali finančnih odnosov, ki bi se lahko razumeli kot potencialno navzkrižje interesov.
Urejevalnik obdelave trenutno organizira raziskovalno temo z enim od avtorjev JF in potrjuje odsotnost drugih sodelovanj.
Reference
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL, et al. (2017). Aktivacija Kappa opioidnih receptorjev v dopaminskih nevronih moti vedenjsko inhibicijo. Neuropsychopharmacology 43 (2), 362–372. doi: 10.1038 / npp.2017.133
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ, et al. (2018). Regulacija estrogena GRK2 inaktivira kappa opioidni receptor, ki signalizira posredovanje analgezije, ne pa odvračanja. J. Neurosci. 38 (37), 8031–8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019). Antagonist A2A receptorjev adenozina izboljša številne simptome ponavljajoče psihoze, ki jo povzroča kvinpirol. eNeuro 6 (1), 1–16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, mešani agonist opioidnih receptorjev kappa-delta, ima protikokainske lastnosti z zmanjšanimi stranskimi učinki pri podganah. Neurofarmakologija 150, 217 – 228. doi: 10.1016 / j.neuropharm.2019.02.010
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, ME, Fuentealba, JA (2019). Blokiranje kappa-opioidnega receptorja razveljavi spremembe v dorsolateralni striatumski dinamiki dopamina med preobčutljivostjo za amfetamin. J. Neurochem. 148, 348–358. doi: 10.1111 / jnc.14612
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE, et al. (2012). Diferencialne signalne lastnosti na kapio opioidnem receptorju 12-epi-salvinorina A in njegovih analogih. Bioorg. Med. Kemija. Lett. 15; 22 (2), 1023–1026. doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Diferencialni učinki novega antagonista kapio opioidnih receptorjev, JDTic, na ponovno uvajanje kokaina, ki ga povzročajo stresi stopal proti kokainu, in njegove antidepresivne učinke pri podganah. Psihoparmakol. (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Razlike med spoloma v nevronskih mehanizmih, ki posredujejo med nagrajevanjem in odvisnostjo. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Becker, JB, Ramirez, VD (1981). Razlike med spoloma v amfetaminu so in vitro spodbudile sproščanje kateholaminov iz progastega tkiva podgane. Brain Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Becker, JB (1999). Razlike med spoloma v dopaminergični funkciji v striatumu in jedru. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Berridge, KC, Robinson, TE (2016). Všečkanje, željo in teorija odvisnosti od spodbudne občutljivosti. Am. Psihol. 71, 670–679. doi: 10.1037 / amp0000059
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Berridge, KC, Venier, IL, Robinson, TE (1989). Okusna reaktivnostna analiza afagije, ki jo povzroča 6-hidroksidopamin: posledice za hipotezo o delovanju dopamina in anhedoniji. Behav. Neurosci. 103, 36 – 45. doi: 10.1037 / 0735-7044.103.1.36
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Diferencialni učinki sistemsko danega norbinaltorfimina (nor-BNI) na kappa-opioidne agoniste v testu mišjega krčenja. Psihofarmakologija 115, 311 – 319. doi: 10.1007 / BF02245071
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Preobčutljivost za amfetaminsko lokomotorno motnjo in prednostno pogojeno mesto pri samcih in mladostnih podganah, ki so bili novorojeno zdravljeni s kvinpirolom. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Bruchas, MR, Chavkin, C. (2010). Kinazne kaskade in usmerjanje proti ligandu na kapio opioidnem receptorju. Psihoparmakol. (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Potencialne vloge opioidnih receptorjev pri motivaciji in večji depresivni motnji. Prog. Brain Res. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Carr, DB, Sesack, SR (2000). Nevroni, ki vsebujejo GABA, se v podgralnem tegmentalnem območju podgane projecirajo v predfrontalno skorjo. Synapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Preobčutljivost za amfetaminsko lokomotorno energijo spremlja povečano K + -stimulirano sproščanje dopamina v medialni prefrontalni skorji podgane. Behav. Brain Res. 237, 313 – 317. doi: 10.1016 / j.bbr.2012.09.052
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Chartoff, EH, Mavrikaki, M. (2015). Razlike med spoloma v funkciji kapio opioidnih receptorjev in njihov potencialni vpliv na odvisnost. Spredaj. Nevrosci. 9, 466. doi: 10.3389 / fnins.2015.00466
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME in dr. (2016). Relativni čas med aktivacijo kapio opioidnih receptorjev in kokainom določa vpliv na nagrado in sproščanje dopamina. Neuropsychopharmacology 41, 989 – 1002. doi: 10.1038 / npp.2015.226
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Chavkin, C., Koob, GF (2016). Dinorfin, disforija in odvisnost: stres zasvojenosti. Neuropsychopharmacology 41, 373 – 374. doi: 10.1038 / npp.2015.258
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Endogeni kappa-opioidni receptorski sistemi uravnavajo mezoakumbalno dopaminsko dinamiko in ranljivost za kokain. J. Neurosci. 25, 5029 – 5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Kvantitativna mikrodiliza brez pretoka omogoča odkrivanje povečanja in zmanjšanja vnosa dopamina v mišjih jedrih. J. Neurosci. Metode 155, 187 – 193. doi: 10.1016 / j.jneumeth.2005.12.018
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Cole, RL, Konradi, C., Douglass, J., Hyman, SE (1995). Prilagoditev nevronov amfetaminu in dopaminu: molekularni mehanizmi regulacije gena prodinnorfina v striatumu podgane. Nevron 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). Samice so manj občutljive kot moški na učinke aktivacije kappa opioidnih receptorjev na zaviranje dopamina. Neurofarmakologija 146, 231 – 241. doi: 10.1016 / j.neuropharm.2018.12.002
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Cope, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, Brown, RW (2010). Zdravljenje z neonatalnim kvinpirolom v odzivnosti na zdravljenje z amfetaminom v odrasli dobi poveča aktivacijo lokomotorja in sproščanje dopamina v jedru. Synapse 64, 289 – 300. doi: 10.1002 / syn.20729
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Cox, BM, Young, AB, glej, RE, Reichel, CM (2013). Razlike med spoloma pri iskanju metamfetamina pri podganah: vpliv oksitocina. Psihoneuroendokrinologija 38, 2343 – 2353. doi: 10.1016 / j.psyneuen.2013.05.005
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Culver, KE, Szechtman, H., Levant, B. (2008). Spremenjena dopaminska D2 podobna receptorska vezava pri podganah z vedenjsko preobčutljivostjo za kvinpirol: učinki predhodnega zdravljenja z Ro 41-1049. EUR. J. Pharmacol. 592, 67 – 72. doi: 10.1016 / j.ejphar.2008.06.101
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Dalman, FC, O'Malley, KL (1999). kappa-opioidna toleranca in odvisnost v kulturah dopaminergičnih nevronov srednjega možganov. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Dovoljenje do kokaina in heroina, ki ga posredujejo receptorji dopamina D2, je odvisno od časa in je povezano z vedenjsko preobčutljivostjo. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Diferencialno vključevanje ventralnih tegmentalnih mu, delta in kappa opioidnih receptorjev v modulacijo sproščanja bazalnega mezolimbičnega dopamina: in vivo mikrodializijske študije. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Nasprotni učinki agonisov mu in kappa opiata na sproščanje dopamina v jedru in v hrbtnem kavatu prosto gibajočih se podgan. J. Pharmacol. Exp. Ther. 244, 1067 – 1080. doi: 10.1073 / pnas.85.14.5274
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Razvoj in časovna organizacija kompulzivnega preverjanja, ki ga povzročajo ponavljajoče injekcije kvinpirola agonista dopamina v živalskem modelu obsesivno-kompulzivne motnje. Behav. Brain Res. 169, 303 – 311. doi: 10.1016 / j.bbr.2006.01.024
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Depresivni učinki agonista kappa opioidnega receptorja salvinorina A so povezani z zmanjšanim faznim sproščanjem dopamina v jedrih jeder. Psihoparmakol. (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., et al. (2017). Specifičnost vezja v inhibitorni arhitekturi VTA uravnava vedenje, ki ga povzroča kokain. Nat. Neurosci. 20 (3), 438–448. doi: 10.1038 / nn.4482
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). Aktivacija opioidnih receptorjev Kapa potencira povečanje sproščenega dopamina, ki ga povzroča kokain, zabeleženo in vivo v mišjih jedrih. Neuropsychopharmacology 39, 3036 – 3048. doi: 10.1038 / npp.2014.157
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, et al. (2015). Odpornost proti kapi opioidnim receptorjem zahteva p38 MAPK Aktivacija v VTA dopaminskih nevronih. J. Neurosci. 35, 12917 – 12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Eilam, D., Szechtman, H. (1989). Dvofazni učinek kvinpirola agonista D-2 na gibanje in gibanje. EUR. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Eilam, D., Szechtman, H. (2005). Psihostimulantsko vedenje kot živalski model obsesivno-kompulzivne motnje: etološki pristop k obliki kompulzivnih ritualov. CNS Spectr. 10, 191–202. doi: 10.1017 / S109285290001004X
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012). Ponavljajoče se zdravljenje z agonistom kapio opioidnega receptorja U69593 obrne povečano sproščanje dopamina s K + v okolju jedra, ne pa tudi izraza lokomotorne preobčutljivosti pri podganah, občutljivih na amfetamin. Nevrohem. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). Zmanjšan nevrotransmisija dopamina in glutamata v jedru podgana kvinpirola, občutljivih na podganah, namiguje na zaviralno delovanje autoreceptorjev D2. J. Neurochem. 134, 1081–1090. doi: 10.1111 / jnc.13209
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., et al. (2017). Mehanizmi potenciranja kapio opioidnih receptorjev funkcije receptorja dopamina D2 pri preobčutljivosti lokomotorja, ki jih povzroča kvinpirol. Int. J. Neuropsihofarmakol. 20, 660–669. doi: 10.1093 / ijnp / pyx042
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Everitt, BJ, Robbins, TW (2005). Okrepitev nevronskih sistemov zaradi odvisnosti od drog: od dejanj do navad do prisile. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Fattore, L., Melis, M. (2016). Spolne razlike v impulzivnem in kompulzivnem vedenju: poudarek na odvisnosti od drog. Addict. Biol. 21 (5), 1043–1051. doi: 10.1111 / adb.12381
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Nevronska in vedenjska plastičnost, povezana s prehodom od nadzorovane do stopnjevane uporabe kokaina. Biol. Psy. 58 (9), 751–9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., et al. (2016). Prisilnost pri obsesivno-kompulzivnih motnjah in odvisnostih. EUR. Neuropsihofarmakol. 26, 856 – 868. doi: 10.1016 / j.euroneuro.2015.12.003
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ford, CP, Mark, GP, Williams, JT (2006). Lastnosti in opioidna inhibicija mezolimbičnih dopaminskih nevronov se razlikujejo glede na ciljno lokacijo. J. Neurosci. 26, 2788 – 2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ford, CP, Beckstead, MJ, Williams, JT (2007). Kappa opioidna inhibicija somatodendritičnih dopaminskih inhibitorjev postsinaptičnih tokov. J. Neurophysiol. 97, 883 – 891. doi: 10.1152 / jn.00963.2006
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). Večkratno dajanje selektivnega agonista kappa-opioidnih receptorjev U-69593 poveča stimulirane ravni dopamina v okoliščinah jedra podgane. J. Nevrosci. Res. 84, 450 – 459. doi: 10.1002 / jnr.20890
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Povečan odziv lokomotorja na amfetamin, ki ga povzroči večkratna uporaba selektivnega agonista kappa-opioidnih receptorjev U-69593. Synapse 61, 771 – 777. doi: 10.1002 / syn.20424
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Učinki akutnega in ponavljajočega dajanja salvinorina A na delovanje dopamina v hrbtnem striatumu podgane. Psihoparmakol. (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Obvezno pikanje alkohola je posledica neuspeha izključitve dorsolateralnega strijnega nadzora nad vedenjem. J. Neurosci. 39 (9), 1744–1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Goto, Y., Grace, AA (2008). Limbična in kortikalna obdelava informacij v jedru. Trendi Neurosci. 31, 552 – 558. doi: 10.1016 / j.tins.2008.08.002
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Grace, AA, Bunney, BS (1980). Nigral dopaminski nevroni: medcelično beleženje in identifikacija z injiciranjem L-dope in histofluorescenco. Znanost 210, 654 – 656. doi: 10.1126 / science.7433992
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Grey, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). K-opioidni agonist, U-69593, v ventralnem striatumu zmanjša akutno vedenje, ki ga povzročajo amfetamin, in kalcijev odvisen dializatni nivo dopamina in glutamata. J. Neurochem. 73, 1066 – 1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Kappa opioidni receptorji uravnavajo stres, ki ga povzroča kokain, in sinaptično plastičnost. Nevron 77, 942 – 954. doi: 10.1016 / j.neuron.2012.12.034
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Heidbreder, Kalifornija, Shippenberg, TS (1994). U-69593 preprečuje preobčutljivost za kokain z normalizacijo bazalnih akumulatorjev dopamina. Nevroport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Ženske, odvisne od opioidov, konoplje in alkohola, kažejo hitrejše napredovanje k zdravljenju z odvisnostmi. Od alkohola odvisni. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Izboljšano sproščanje dopamina z zaviralci prenosa dopamina, ki jih opisuje model omejene difuzije in hitro skeniranje ciklične voltammetrije. ACS Chem. Nevrosci. 7, 700–709. doi: 10.1021 / acschemneuro.5b00277
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Holden, C. (2001). "Vedenjske" zasvojenosti: ali obstajajo? Znanost 294, 980 – 982. doi: 10.1126 / science.294.5544.980
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Imperato, A., Di Chiara, G. (1988). Učinki lokalno uporabljenih agonistov in antagonistov receptorjev D-1 in D-2, preučenih z dializo možganov. EUR. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Učinki antagonista kappa opioidnih receptorjev, norbinaltorphimina, na stres in z zdravili povzročeno ponovno vzpostavitev nikotina, ki je pogojen s kraji pri miših. Psihoparmakol. (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., et al. (2014). Salvinorin A uravnava funkcijo prenosa dopamina preko kapio opioidnega receptorja in mehanizma, odvisnega od ERK1 / 2. Neurofarmakologija 86, 228 – 240. doi: 10.1016 / j.neuropharm.2014.07.016
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Vedenjska preobčutljivost za kvinpirol ni povezana s povečanim prekrivanjem dopamina v jedru. Neurofarmakologija 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Koob, GF, Volkow, ND (2016). Nevrobiologija odvisnosti: analiza nevrocircuitry. Lancetova psihiatrija 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Koob, GF (2013). Zasvojenost je primanjkljaj nagrajevanja in stresne bolezni. Spredaj. Psihiatrija 4, 72. doi: 10.3389 / fpsyt.2013.00072
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., et al. (2008). Razlike med spoloma v dimenzijah obsesivno-kompulzivnih simptomov. Stisnite tesnobo. 25 (10), 832–838. doi: 10.1002 / da.20332
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, agonist oppaidnih receptorjev kappa, zmanjšuje povečanje zunajceličnega dopamina, ki ga povzroča kokain, v jedru podgan. Nevrosci. Lett. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Ekspresija mRNA opioidnega receptorja v CNS podgane: anatomske in funkcionalne posledice. Trendi Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Imunohistokemična lokalizacija kloniranega receptorja kappa 1 v CNS in hipofizi pri podganah. Nevroznanost 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Margolis, EB, Karkhanis, AN (2019). Prispevek dopaminergičnega celičnega in vezja k averziji, ki jo posreduje kappa opioidni receptor. Nevrohem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Kappa-opioidni agonisti neposredno zavirajo dopaminergične nevrone srednjih možganov. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Tako Kappa kot Mu opioidni agonisti zavirajo vnos glutamatergike v nevrone ventralnega tegmentalnega območja. J. Neurophysiol. 93, 3086 – 3093. doi: 10.1152 / jn.00855.2004
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Kappa opioidi selektivno nadzorujejo dopaminergične nevrone, ki štrlijo v prefrontalno skorjo. Proc. Natl. Acad. Sci. ZDA 103, 2938 – 2942. doi: 10.1073 / pnas.0511159103
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Srednji možganski dopaminski nevroni: projekcijski cilj določa trajanje akcijskega potenciala in zaviranje receptorjev dopamina D (2). J. Neurosci. 28, 8908 – 8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Mathews, IZ, McCormick, CM (2007). Ženske in samci podgan v pozni adolescenci se od odraslih razlikujejo po lokomotorni aktivnosti, ki jo povzroča amfetamin, vendar ne glede na prednostno pogojene amfetamine na mestu. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T., et al. (2011). Razlike med spoloma pri obsesivno-kompulzivni motnji: pregled literature. Braz. J. Psihiatrija 33 (4), 390–399. doi: 10.1590 / S1516-44462011000400014
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Antagonizem Kappa opioidnih receptorjev in motnja gena prodinnorfina blokirajo vedenjske odzive, ki jih povzročajo stres. J. Neurosci. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Predhodna aktivacija kapio opioidnih receptorjev s pomočjo U50,488 posnema ponavljajoči se stres v plavanju, da se potencira kokainsko mesto. Neuropsychopharmacology 31, 787 – 794. doi: 10.1038 / sj.npp.1300860
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). Vedenjski odzivi, ki jih povzroča stres, povzročajo socialni poraz, posreduje endogeni kappa opioidni sistem. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Razlike med spoloma pri (+) - amfetaminskih in (+) - vedenjskih odzivih na metamfetamin pri samcih in samicah podgan Sprague-Dawley. Pharmacol. Biochem. Behav. 86, 140 – 149. doi: 10.1016 / j.pbb.2006.12.018
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Vplivi kapio opioidov na samokapiranje kokaina s pomočjo opic. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Molekularna osnova dolgoročne plastičnosti, ki je v osnovi zasvojenosti. Nat. Rev. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Kappa-opioidni agonist U69593 potencira lokomotorno preobčutljivost na k2pirole agonista D3 / DXNUMX: pred- in postsinaptične mehanizme. Neuropsychopharmacology 31, 1967 – 1981. doi: 10.1038 / sj.npp.1300938
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Perreault, ML, Seeman, P., Szechtman, H. (2007). Stimulacija kappa-opioidnih receptorjev pospeši patogenezo kompulzivnega preverjanja v modelu senzibilizacije kvinpirola obsesivno-kompulzivne motnje (OCD). Behav. Neurosci. 121, 976 – 991. doi: 10.1037 / 0735-7044.121.5.976
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Pierce, RC, Kalivas, PW (1997). Model vezja izražanja vedenjske preobčutljivosti na psihostimulance, ki so podobni amfetaminom. Brain Res. Brain Res. Rev. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Polter, AM, Bishop, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). Blok za naknadno napetost opioidnih receptorjev Kapa rešuje dolgotrajno potenciranje zaviralnih sinaps in preprečuje ponovno vzpostavitev uživanja kokaina. Biol. Psihiatrija 76, 785 – 793. doi: 10.1016 / j.biopsych.2014.04.019
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). U-69593, agonist kapio opioidnih receptorjev, zmanjšuje vedenjsko preobčutljivost na kokain pri samicah podgan. Behav. Neurosci. 122, 151 – 160. doi: 10.1037 / 0735-7044.122.1.151
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Nevrokognitivni endofenotipi impulzivnosti in kompulzivnosti: proti dimenzijski psihiatriji. Trendi Cognit. Sci. 16, 81 – 91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Evolucijski cikel vpliva na samokokacijo kokaina na urnik progresivnega razmerja pri podganah. Psihoparmakol. (Berl) 98, 408 – 411. doi: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). Nevronska osnova hrepenenja po drogah: spodbujevalno-senzibilizacijska teorija odvisnosti. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Robinson, TE, Berridge, KC (2001). Spodbuda-preobčutljivost in zasvojenost. Odvisnost 96, 103 – 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Različni učinki nalmefena na hitrost vnosa dopamina in aktivnost kappa opioidnih receptorjev v jedru, ki se pojavlja po kronični vmesni izpostavljenosti etanolu. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF in sod. (2013). Dolgotrajni antagonizem κ opioidnih receptorjev preprečuje stopnjevanje in povečanje motivacije za vnos heroina. J. Neurosci. 33 (49), 19384–19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Sesack, SR, Pickel, VM (1992). Dvojna ultrastrukturna lokalizacija imunoreaktivnosti enkefalina in tirozin hidroksilaze v tegmentalnem območju ventralnega podgana: več substratov za interakcije med opiati in dopaminom. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Sesack, SR, Aoki, C., Pickel, VM (1994). Ultrastrukturna lokalizacija D2 receptorju podobne imunoreaktivnosti v nevronih srednjih možganov in njihovih strij. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Sesia, T., Bizup, B., Grace, AA (2013). Vrednotenje živalskih modelov obsesivno-kompulzivne motnje: povezanost s faznim delovanjem dopaminskih nevronov. Int. J. Neuropsihofarmakol. 16, 1295–1307. doi: 10.1017 / S146114571200154X
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dinorfin in patofiziologija odvisnosti od drog. Farmakol. Ther. 116, 306–321. doi: 10.1016 / j.pharmthera.2007.06.011
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Smith, JA, Loughlin, SE, Leslie, FM (1992). inhibicija kappa-opioidov sproščanja [3H] dopamina iz podkožnih celičnih kultur, ki se odvajajo od vencela. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Pri miših, ki izvirajo iz receptorja za dopamin D2L, je prikazan primanjkljaj pozitivnih in negativnih krepitvenih lastnosti morfija in pri izogibanju učenju. Nevroznanost 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Aktivacija sistema dinorfin / κ-opioidnih receptorjev v amigdali zaradi stresa povzroči prednost nikotinskih pogojnih mest. J. Neurosci. 32, 1488 – 1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Spanagel, R., Herz, A., Shippenberg, TS (1992). Nasprotni tonično aktivni endogeni opioidni sistemi modulirajo mezolimbično dopaminergično pot. Proc. Natl. Acad. Sci. 89, 2046 – 2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., nem., DC (1993). Opioidni receptorji v dopaminergičnih regijah srednjega mozga podgane. II. Avtoradiografija kappa in delta receptorjev. J. Nevronski transm. Gen. Sekta 91, 53 – 66. doi: 10.1007 / BF01244918
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Endogeno kappa-opioidno posredovanje potenciranega stresnega potenciala mesta, ki je pogojeno z etanolom, in samo-dajanja. Psihoparmakol. (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Želeli drog: vedenjska preobčutljivost in ponovitev do vedenja, ki išče drogo DR, ed. Farmakol. Rev. 63, 348–365. doi: 10.1124 / pr.109.001933
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., et al. (2016). Veljavnost modela preobčutljivosti za kinpirole podgane OCD: povezovanje dokazov iz živali in kliničnih študij. Spredaj. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Sun, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., et al. (2019). Vhod bazolateralne amigdale v medialno prefrontalno skorjo nadzoruje obsesivno-kompulzivno motnje, podobno preverjanju. Proc. Natl. Acad. Sci. ZDA 116, 3799 – 3804. doi: 10.1073 / pnas.1814292116
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Večja koekspresija ?? - opioidnih receptorjev in prenašalca dopamina v jedru vsebuje aksonske profile. Synapse 42, 185 – 192. doi: 10.1002 / syn.10005
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Okoljski in vedenjski sestavni deli preobčutljivosti, ki jih povzroča dopaminski agonist kvinpirol. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dinamika vedenjske preobčutljivosti, ki jo povzroča kvinpirol dopaminov agonist in predlagani centralni mehanizem za nadzor energije. Psihoparmakol. (Berl) 115, 95 – 104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Quinpirole povzroča kompulzivno preverjanje vedenja pri podganah: potencialni živalski model obsesivno-kompulzivne motnje (OCD). Behav. Neurosci. 112, 1475 – 1485. doi: 10.1037 / 0735-7044.112.6.1475
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Szechtman, H., Culver, K., Eilam, D. (1999). Vloga dopaminskih sistemov pri obsesivno-kompulzivnih motnjah (OCD): posledice novega modela živali, ki jih povzroča psihostimulant. Pol. J. Pharmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, C a, JZ, M., et al. (2001). Obvezno preverjanje vedenja podgan, občutljivih za kvinpirol, kot živalski model obsesivno-kompulzivnega motenja (OCD): oblika in nadzor. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D., Harvey, BH, Edemann-Callesen, H., et al. (2017). Obsesivno-kompulzivna motnja: Vpogled v modele na živalih. Neurosci. Biobehav. Rev. 76, 254 – 279. doi: 10.1016 / j.neubiorev.2016.04.019
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Tejeda, HA, Bonci, A. (2019). Nadzor dinaminskih / kappa-opioidnih receptorjev dinamike dopamina: Posledice za negativna afektivna stanja in psihiatrične motnje. Brain Res. 1713, 91 – 101. doi: 10.1016 / j.brainres.2018.09.023
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., et al. (2017). Modulacija kappa-opioidnega receptorja, značilnega za pot in celice, ravnovesja inhibicije vzbujanja-inhibicije različno vodi dv in d1 akumensko nevronsko aktivnost. Nevron 93 (1), 147–163. doi: 10.1016 / j.neuron.2016.12.005
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Thompson, AC, Zapata, A., Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Aktivacija kappa-opioidnih receptorjev spremeni vnos dopamina v jedro in nasprotuje učinkom kokaina. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Thompson, D., Martini, L., Whistler, JL (2010). Spremenjeno razmerje D1 in D2 dopaminskih receptorjev v mišjem striatumu je povezano z vedenjsko preobčutljivostjo na kokain. PloS One 5, e11038. doi: 10.1371 / journal.pone.0011038
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., et al. (2000). Ločene funkcije obeh izoform dopaminskih receptorjev D2. Narava 408, 199 – 203. doi: 10.1038 / 35041572
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Van Bockstaele, EJ, Pickel, VM (1995). Nevroni, ki vsebujejo GABA v ventralnem tegmentalnem območju, segajo v jedro v možganih podgane. Brain Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY, et al. (2013). Ablacija kappa-opioidnih receptorjev iz možganskih dopaminskih nevronov ima anksiolitične učinke in povečuje plastičnost, ki jo povzroča kokain. Neuropsychopharmacology 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Vanderschuren, LJ, Kalivas, PW (2000). Spremembe prenosa dopaminergičnega in glutamatergičnega v indukciji in izražanju vedenjske preobčutljivosti: kritični pregled predkliničnih študij. Psihoparmakol. (Berl) 151 (2-3), 99–120. doi: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017). Sistem motiv dopamina: posledice za zasvojenost z mamili in hrano. Nat. Rev. Neurosci. 18, 741–752. doi: 10.1038 / nrn.2017.130
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Izpuščanje in vnos dopamina sta pri ženskah podganah striatum večja, merjeno s hitro ciklično voltammetrijo. Nevroznanost 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Wee, S., Koob, GF (2010). Vloga opiridnega sistema dinorfin-κ v krepitvi učinkov zlorabe drog. Psihoparmakol. (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Inhibicija kapio opioidnih receptorjev je pri podganah s podaljšanim dostopom do kokaina zmanjšala vnos kokaina. Psihoparmakol. (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., et al. (2015). κ Opioidni receptorji v jedru nastanejo v lupini in povečujejo vnos metamfetamina. J. Neurosci. 35 (10), 4296–4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Dimenzije simptomov obsesivno-kompulzivne motnje: fenomenologija in rezultati zdravljenja z izpostavljenostjo in ritualnim preprečevanjem. Psihopatologija 46, 365 – 376. doi: 10.1159 / 000348582
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Wilson, CJ, Young, SJ, Groves, PM (1977). Statistične lastnosti vlakov nevronskih konic v substantia nigra: vrste celic in njihove interakcije. Brain Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Wise, RA (2009). Vloge za nigrostriatalni - ne le mezokortikolimbični - dopamin v nagradi in odvisnosti. Trendi Neurosci. 32, 517 – 524. doi: 10.1016 / j.tins.2009.06.004
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Xiao, L., Becker, JB (1994). Kvantitativno določanje mikrodializirane koncentracije zunajceličnega striatalnega dopamina pri samcih in samicah podgan: učinki estrokularnega cikla in gonadektomije. Nevrosci. Lett. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Yager, LM, Garcia, AF, Wunsch, AM, Ferguson, SM (2015). Vloge in izidi striatuma: vloga v odvisnosti od drog. Nevroznanost 301, 529 – 541. doi: 10.1016 / j.neuroscience.2015.06.033
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ključne besede: kapio opioidni receptor, dopamin, kompulzivnost, amfetamin, kvinpirol, lokomotorna preobčutljivost
Citiranje: Escobar AdP, Casanova JP, Andrés ME in Fuentealba JA (2020). Spredaj. Farmakol. 11: 57. doi: 10.3389 / fphar.2020.00057
Prejeto: 16. oktobra 2019; Sprejeto: 22. januarja 2020;
Objavljeno: 18 februar 2020.
Uredil:
Gonzalo E. Yevenes, Univerza v Concepcionu, Čile
Pregledal:
Luis Gerardo Aguayo, Univerza v Concepcionu, Čile
Hugo Tejeda, Nacionalni inštitut za zlorabo drog (NIDA), ZDA
Cecilia Scorza, Instituto de Investigaciones Biológicas Clemente Estable (IIBCE), Urugvaj
Copyright © 2020 Escobar, Casanova, Andrés in Fuentealba. To je članek z odprtim dostopom, ki se distribuira pod pogoji Licenca za priznanje Creative Commons (CC BY). Uporaba, distribucija ali reprodukcija v drugih forumih je dovoljena pod pogojem, da so avtorji avtorjev in lastniki avtorskih pravic priznani in da je navedena originalna objava v tej reviji v skladu s sprejeto akademsko prakso. Uporaba, distribucija ali reprodukcija ni dovoljena, kar ni v skladu s temi pogoji.
* Korespondenca: José Antonio Fuentealba, [e-pošta zaščitena]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675
