ด้านหน้า จิตเวชศาสตร์, 25 กุมภาพันธ์ 2016 | http://dx.doi.org/10.3389/fpsyt.2016.00024
จริดกู๊ดแมน และ มาร์คกรัมแพคการ์ด*
- ภาควิชาจิตวิทยา Texas A&M Institute for Neuroscience มหาวิทยาลัย Texas A&M College Station รัฐเท็กซัสสหรัฐอเมริกา
มุมมองที่ว่าระบบหน่วยความจำที่แตกต่างทางกายวิภาคมีส่วนช่วยในการพัฒนาของการติดยาเสพติดและการกำเริบของโรคได้รับการสนับสนุนอย่างกว้างขวาง การทบทวนสั้น ๆ ในปัจจุบันทบทวนสมมติฐานนี้ใหม่เมื่อมันถูกเสนอเมื่อ 20 ปีก่อน (1) และไฮไลท์พัฒนาการล่าสุดหลายประการ การวิจัยอย่างกว้างขวางที่ใช้กระบวนทัศน์การเรียนรู้สัตว์ที่หลากหลายบ่งชี้ว่าระบบประสาทที่แบ่งแยกไม่ได้เป็นสื่อกลางในการเรียนรู้และความทรงจำที่แตกต่าง ระบบหน่วยความจำแต่ละระบบอาจก่อให้เกิดองค์ประกอบที่ไม่ซ้ำกับพฤติกรรมที่เรียนรู้ซึ่งสนับสนุนการติดยาเสพติดและการกำเริบของโรค โดยเฉพาะอย่างยิ่งการเปลี่ยนจากการใช้ยาเพื่อการผ่อนคลายไปเป็นการใช้ยาในทางที่ผิดอาจสะท้อนถึงการเปลี่ยนแปลงทางระบบประสาทจากการควบคุมการรับรู้ของพฤติกรรมที่สื่อกลางโดยฮิบโปแคมปัส / dorsomedial striatum ไปสู่การควบคุมพฤติกรรมของพฤติกรรม นอกจากนี้ความเครียด / ความวิตกกังวลอาจเป็นปัจจัยร่วมที่เอื้อต่อหน่วยความจำที่ขึ้นอยู่กับ DLS และสิ่งนี้อาจทำหน้าที่เป็นกลไก neurobehavioral พื้นฐานการใช้ยาที่เพิ่มขึ้นและการกำเริบของโรคในมนุษย์หลังจากเหตุการณ์ชีวิตที่เครียด หลักฐานที่สนับสนุนมุมมองหลายระบบของการติดยาเสพติดมาจากการศึกษาการเรียนรู้และความทรงจำที่ใช้เป็นตัวเสริมสารเสพติดมักพิจารณาในบริบทของการวิจัยติดยาเสพติดรวมถึงโคเคนแอลกอฮอล์และยาบ้า นอกจากนี้หลักฐานล่าสุดแสดงให้เห็นว่าวิธีการระบบหน่วยความจำอาจเป็นประโยชน์สำหรับการทำความเข้าใจแหล่งที่มาเฉพาะของการติดยาเสพติดที่สะท้อนถึงปัญหาสุขภาพที่เกิดขึ้นใหม่รวมถึงการใช้กัญชาอาหารที่มีไขมันสูงและการเล่นวิดีโอเกม
บทนำ
นักวิจัยมักจะมองไปที่กลไกของการเรียนรู้และพฤติกรรมเพื่ออธิบายว่ามนุษย์ได้รับและแสดงออกทางด้านจิตวิทยาอย่างไร ตัวอย่างของแอปพลิเคชันดังกล่าวจัดทำโดยนอร์แมนเอ็มไวท์ผู้ใช้หลักคำสอนของทฤษฎีการเรียนรู้แบบดั้งเดิมและหลักฐานการทดลองที่สนับสนุนการมีอยู่ของระบบหน่วยความจำหลายระบบในสมองเพื่อให้ได้นวนิยาย1) โดยเฉพาะอย่างยิ่งสีขาวระบุว่ายาเสพติดสามารถเล่นเป็นส่วนหนึ่งของ "reinforcers" ที่เช่นอาหารหรือน้ำในงานการเรียนรู้เสริมสร้างความสัมพันธ์ระหว่างสิ่งเร้าที่เกี่ยวข้องกับยาเสพติดบริบทและพฤติกรรมเพื่อส่งเสริมการใช้ยาเสพติดและเมื่อเวลาผ่านไปติดยาเสพติด White ยังได้รวมสมมติฐานที่เกิดขึ้นใหม่ว่ามีหน่วยความจำประเภทต่าง ๆ ที่ถูกสื่อโดยระบบประสาทที่แยกตัวไม่ออก จากมุมมองของนวนิยายยาเสพติดสามารถปรับเปลี่ยนระบบประสาทหลายระบบได้โดยตรงและระบบประสาทเหล่านี้จะเข้าสู่การเข้ารหัสส่วนประกอบที่แตกต่างกันของหน่วยความจำที่เกี่ยวข้องกับยาซึ่งเมื่อแสดงออกแล้วส่งเสริมการใช้ยาต่อไป
ปี 2016 เป็นวันครบรอบปีที่ 20 ของมุมมองระบบหน่วยความจำที่หลากหลายเกี่ยวกับการติดยาเสพติดตามที่อธิบายโดย White การทบทวนในปัจจุบันกลับมาทบทวนสมมติฐานที่มีอิทธิพลนี้ในขณะที่เน้นการพัฒนาล่าสุดที่สำคัญบางอย่างที่ไม่เพียง แต่ยืนยันสมมติฐานดั้งเดิม แต่ยังสร้างความเข้าใจเพิ่มเติมเกี่ยวกับวิธีที่ระบบหน่วยความจำหลายระบบสนับสนุนการติดยาเสพติด
มุมมองระบบหน่วยความจำหลายระบบของการเสพติด
การรวบรวมหลักฐานจากการศึกษาที่ใช้มนุษย์และสัตว์ที่ต่ำกว่าแสดงว่าหน่วยความจำของสัตว์เลี้ยงลูกด้วยนมเป็นสื่อกลางโดยระบบประสาทที่ค่อนข้างอิสระ (2-4)] การทดลองในช่วงแรกที่แยกตัวออกจากระบบหน่วยความจำหลายแห่งนั้นดำเนินการเป็นหลักในเขาวงกตเรเดียลและระบุหน้าที่ช่วยในการจำที่ไม่ซ้ำกันสำหรับฮิบโปแคมตัส, ดอร์ซัล striatum และ amygdala (5, 6) ฮิปโปแคมปัสเป็นสื่อกลางในรูปแบบขององค์ความรู้ / เชิงพื้นที่ในขณะที่หลัง striatum เป็นสื่อกลางในการกระตุ้นการตอบสนอง (S-R) หน่วยความจำนิสัย amygdala ไกล่เกลี่ย Pavlovian และความสัมพันธ์ที่กระตุ้นผลกระทบต่อการเชื่อมโยง (6, 7) ในขณะที่ยังช่วยลดบทบาทเร้าอารมณ์ของอารมณ์เร้าอารมณ์ในหน่วยความจำประเภทอื่น ๆ (8-12).
ภายในบริบทของมุมมองหลายระบบของหน่วยความจำขาว (1) ชี้ให้เห็นว่าฮิบโปคัสตัสหลัง striatum และ amygdala เข้ารหัสองค์ประกอบที่เป็นเอกลักษณ์ของความทรงจำที่เกี่ยวข้องกับยา (ดูรูปที่ 1) ฮิบโปเข้ารหัสความรู้ที่ชัดเจนเกี่ยวกับความสัมพันธ์ระหว่างตัวชี้นำและเหตุการณ์ (เช่นความสัมพันธ์ระหว่างสิ่งเร้าและสิ่งเร้า) ในบริบทของยา ที่สำคัญฮิปโปแคมปัสไม่เข้ารหัสการตอบสนองเชิงพฤติกรรม แต่สามารถใช้ข้อมูลที่ได้รับจากฮิบโปเพื่อสร้างการตอบสนองเชิงพฤติกรรมที่เหมาะสมเพื่อรับการเสริมแรงด้วยยา ในทางกลับกัน striatum หลังเข้ารหัสความสัมพันธ์ระหว่างสิ่งเร้าที่เกี่ยวข้องกับยาและการตอบสนองพฤติกรรม สิ่งนี้อาจช่วยให้การนำเสนอของคิวที่เกี่ยวข้องกับยาเพื่อเปิดใช้งานการตอบสนองพฤติกรรมอัตโนมัติที่ส่งผลให้เกิดการเสพยาเสพติด (เช่นวิ่งเข้าหาหรือกดคันเครื่องมือ) amygdala เข้ารหัสความสัมพันธ์ของ Pavlovian ดังนั้นจึงช่วยให้การชี้นำเป็นกลางในบริบทยาเสพติดกลายเป็นความเกี่ยวข้องกับรางวัลยาเสพติด สัตว์เหล่านี้มีปฏิกิริยาตอบสนองต่อสิ่งที่มีเงื่อนไขเหล่านี้ในทำนองเดียวกันกับวิธีที่พวกมันตอบโต้กับยาในขั้นต้น โดยเฉพาะอย่างยิ่งตัวชี้นำที่มีเงื่อนไขเปิดใช้งานการตอบสนองทางอารมณ์แบบมีเงื่อนไขรวมถึงสภาวะอารมณ์ภายในและแนวทางที่มีเงื่อนไขต่อ (หรือในบางกรณี องค์ประกอบที่สำคัญอีกประการหนึ่งของสมมติฐานของไวท์คือยาสามารถปรับการทำงานของหน่วยความจำของแต่ละส่วนของสมอง ดังนั้นยาเสพติดอาจเพิ่มประสิทธิภาพการบริหารตนเองของตนเองผ่านการรวมการเพิ่มความทรงจำที่เกี่ยวข้องกับยาที่เข้ารหัสโดยฮิปโปแคมปัส, อะมิกดาลาและหลัง striatum (ดูรูปที่ 1).
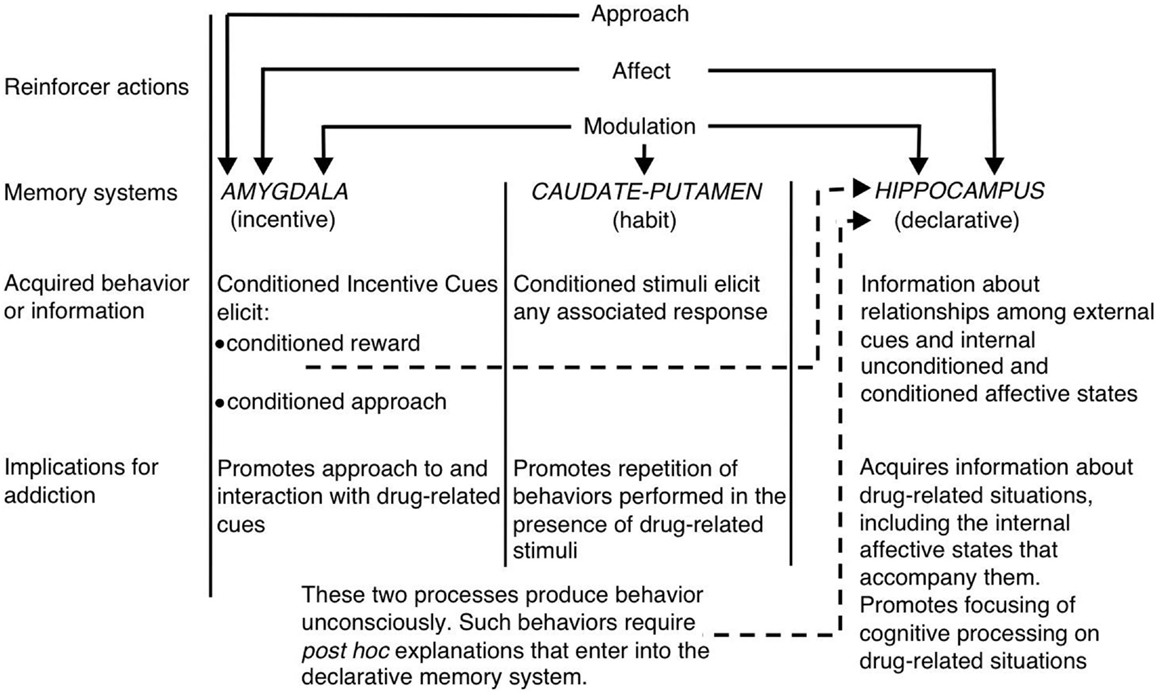
รูปที่ 1 สีขาว (1) ระบบหน่วยความจำหลายมุมมองของการติดยาเสพติด. เช่นเดียวกับสารเสริมแรงตามธรรมชาติยาเสพติดมี“ ฤทธิ์เสริมแรง” หลายอย่างรวมถึงความสามารถในการกระตุ้นให้เกิดผลกระทบเชิงบวก / เชิงลบแนวทางและการปรับระบบความจำ amygdala, caudate – putamen (เช่น dorsal striatum) และ hippocampus เป็นสื่อกลางของระบบความจำที่แยกไม่ออกและระบบความจำแต่ละระบบน่าจะเข้ารหัสส่วนประกอบเฉพาะของความทรงจำที่เกี่ยวข้องกับยา เนื่องจากคุณสมบัติในการปรับเปลี่ยนหน่วยความจำยาเสพติดสามารถเพิ่มประสิทธิภาพการบริหารตนเองของตนเองโดยการเพิ่มการทำงานของระบบเหล่านี้ (พิมพ์ซ้ำจาก White โดยได้รับอนุญาตจาก John Wiley & Sons)
สอดคล้องกับระบบหน่วยความจำหลายมุมมองของการติดยาเสพติดหลักฐานที่กว้างขวางบ่งชี้ถึงบทบาทที่สำคัญสำหรับฮิบโป, ดอร์ซัล striatum และ amygdala ในการติดยาเสพติดและการกำเริบของสารเสพติดที่หลากหลาย (13)] ฮิบโปหลังดูเหมือนจะมีบทบาทในการควบคุมบริบทของการแสวงหายาเสพติดโคเคน (14-16) บริเวณด้านข้างของ dorsal striatum (DLS) ไกล่เกลี่ย S-R คันโยกสำหรับโคเคนและแอลกอฮอล์เป็นนิสัย17, 18) และ amygdala basolateral (BLA) ไกล่เกลี่ยยาเสพติดปรับอากาศที่กำลังมองหาโคเคนแอลกอฮอล์และเฮโรอีน (19-22) ยังสอดคล้องกับสมมติฐานของ White สารเสพติดสามารถปรับฟังก์ชั่นช่วยในการจำของฮิบโป, ดอร์ซัล striatum และ amygdala (23-31).
การศึกษาเมื่อเร็ว ๆ นี้ได้มีส่วนร่วมในการแก้ไขนวนิยายเกี่ยวกับระบบหน่วยความจำหลายวิธีในการติดยาเสพติด คุณสมบัติที่สำคัญของมุมมองร่วมสมัยนี้ ได้แก่ (1) การเปลี่ยนแปลงทางระบบประสาทเมื่อเวลาผ่านไปเป็นความจำนิสัยที่ขึ้นอยู่กับ DLS, (2) การมีปฏิสัมพันธ์ระหว่างระบบหน่วยความจำ (3) บทบาทของความเครียดและความวิตกกังวลในการเสริมสร้างการค้นหายาเสพติดเป็นประจำ การประยุกต์ใช้สมมติฐานนี้กับแหล่งที่มาใหม่ของการติดยาเสพติด
Neuroanatomical Shift จากการรับรู้ถึงนิสัย
ในสถานการณ์การเรียนรู้แบบทดลองโดยทั่วไปแล้วผู้เรียนจะใช้พฤติกรรมที่มีจุดประสงค์เมื่อเริ่มแก้ไขภารกิจ อย่างไรก็ตามหลังจากการฝึกอบรมอย่างกว้างขวางพฤติกรรมจะกลายเป็นอิสระและสามารถดำเนินการได้ด้วยความสนใจความตั้งใจหรือความพยายามทางปัญญาเพียงเล็กน้อยซึ่งถือเป็น“ นิสัย” [สำหรับการตรวจสอบโปรดดูที่การอ้างอิง (32)] ในการสาธิตช่วงต้นของการเปลี่ยนแปลงนี้จากการควบคุมการรับรู้พฤติกรรมเป็นนิสัยหนูได้รับการฝึกฝนโดยใช้รางวัลอาหารในงานสองทางบวก - เขาวงกต33-35) ในงานนี้หนูได้รับการปล่อยตัวจากตำแหน่งเริ่มต้นเดียวกัน (เช่นแขนใต้) และต้องทำการเลี้ยวตัวที่สอดคล้องกันที่ทางแยกเขาวงกตเพื่อรับรางวัลอาหารที่ตั้งอยู่เสมอในแขนเป้าหมายเดียวกัน (เช่นให้ชิดซ้ายเสมอ หันไปหาอาหารที่แขนตะวันตก) หนูสามารถแก้ปัญหานี้ได้โดยการเรียนรู้การตอบสนองด้วยการเลี้ยวตัวที่สอดคล้องกันหรือทำการตอบสนองใด ๆ ก็ตามที่จำเป็นเพื่อไปยังตำแหน่งอวกาศเดียวกัน เพื่อกำหนดกลยุทธ์ที่หนูจ้างนักวิจัยดำเนินการทดสอบการสอบสวนในสัตว์ที่ได้รับการปล่อยตัวจากแขนเริ่มต้นที่ตรงกันข้าม (เช่นแขนเหนือ) หากสัตว์หันตัวไปทางตรงกันข้ามเพื่อไปยังตำแหน่งเป้าหมายดั้งเดิมพวกมันจะถูกระบุว่าเป็นผู้เรียนในสถานที่ หากสัตว์ทำการหันตัวเช่นเดียวกับในระหว่างการฝึก (เช่นไปที่แขนตรงข้ามกับตำแหน่งเป้าหมายดั้งเดิม) สัตว์จะถูกระบุว่าเป็นผู้เรียนที่ตอบสนอง หลักฐานบ่งชี้ว่าหลังจากการฝึกอบรมสัตว์ส่วนใหญ่แสดงการเรียนรู้ในสถานที่ในขณะที่หลังจากการฝึกฝนอย่างกว้างขวางสัตว์ก็เปลี่ยนเป็นการเรียนรู้การตอบสนองตามปกติ (34-36) ที่น่าสนใจการเปลี่ยนจากการเรียนในสถานที่เป็นการเรียนรู้การตอบสนองอาจสะท้อนการเปลี่ยนแปลงทางระบบประสาท การใช้สถานที่เริ่มต้นของการเรียนรู้ในงานนี้เป็นสื่อกลางโดยฮิบโปแคมปัสและ dorsomedial striatum [DMS (36, 37)] ในขณะที่การใช้การเรียนรู้การตอบสนองหลังจากการฝึกอบรมที่ยาวนานนั้นถูกใช้โดย DLS (36).
นอกเหนือจากการสาธิตในช่วงต้นโดยใช้เขาวงกตบวก (34, 35) พฤติกรรมที่เปลี่ยนไปสู่ความจำที่เป็นนิสัยได้แสดงให้เห็นในภายหลังโดยใช้กระบวนการกดคันบังคับ38-42) ในงานการเรียนรู้ที่เป็นเครื่องมือเหล่านี้สัตว์เริ่มกดปุ่มอย่างตั้งใจเพื่อให้ได้ผลลัพธ์และจะหยุดการกดคันเมื่อผลลัพธ์ของอาหารนั้นลดคุณค่าลง อย่างไรก็ตามการติดตามสัตว์ฝึกอย่างกว้างขวางจะเปลี่ยนเป็นการตอบสนองตามปกติและจะดำเนินการกดคันโยกต่อไปแม้ว่าจะได้รับคุณค่าทางอาหารแล้วก็ตาม (40) ตามที่แสดงให้เห็นเดิมในเขาวงกตบวก (36) การเปลี่ยนจากการรับรู้สู่นิสัยในงานการเรียนรู้ด้วยเครื่องมืออาจมีสาเหตุมาจากการเปลี่ยนแปลงทางระบบประสาท การควบคุมการรับรู้เริ่มต้นของพฤติกรรมในงานการเรียนรู้ด้วยเครื่องมือเหล่านี้เป็นสื่อกลางโดยฮิบโปและ DMS (43, 44) ในขณะที่การตอบสนองแบบปกติภายหลังนั้นเป็นสื่อกลางโดย DLS (18, 45, 46).
นักวิจัยหลายคนแนะนำว่าการเปลี่ยนระบบประสาทเป็นความทรงจำของนิสัยที่แสดงให้เห็นในเขาวงกตและงานการเรียนรู้ที่เป็นเครื่องมืออาจรองรับการเปลี่ยนจากการใช้ยาเพื่อการพักผ่อนไปเป็นการใช้ยาในทางที่ผิด13, 47-50) สอดคล้องกับสมมติฐานนี้ผู้วิจัยได้แสดงให้เห็นถึงความหลากหลายของสารที่ถูกทารุณกรรมที่ DMS ไกล่เกลี่ยเป้าหมายการตอบสนองสำหรับการเสริมแรงยาและ DLS ไกล่เกลี่ยนิสัยตอบสนองสำหรับการเสริมแรงยา (18, 31, 51-53).
เมื่อพิจารณาถึงศักยภาพในการใช้ยาในทางที่ผิดผู้วิจัยได้แนะนำว่ายาเสพติดอาจช่วยเพิ่มฟังก์ชั่นหน่วยความจำตามนิสัยของ DLS และเร่งการเปลี่ยนจากองค์ความรู้ไปเป็นการควบคุมพฤติกรรม สอดคล้องกับสมมติฐานนี้การเปิดรับยาบ้าหรือโคเคนซ้ำ ๆ ช่วยอำนวยความสะดวกในการเปลี่ยนจากเป้าหมายที่กำหนดไปเป็นการตอบสนองตามปกติสำหรับการเสริมอาหารในงานเร่งด่วนด้วยเครื่องมือ (31, 54-59) นอกจากนี้การกดคันโยกสำหรับสารเสพติด (เช่นแอลกอฮอล์หรือโคเคน) เมื่อเทียบกับรางวัลอาหารมีความสัมพันธ์กับการตอบสนองที่เป็นนิสัยมากกว่าเมื่อเทียบกับเป้าหมายที่กำหนด (24, 60, 61) ในมนุษย์บุคคลที่ติดเหล้าจะแสดงการตอบสนองที่เป็นนิสัยในงานเรียนรู้เกี่ยวกับอุปกรณ์สัมพันธ์กับบุคคลที่ไม่ได้ขึ้นอยู่กับการควบคุม (62) การเพิ่มประสิทธิภาพของหน่วยความจำตามนิสัย DLS นี้โดยการเสพติดยาเสพติดยังได้รับการปฏิบัติในงานการเรียนรู้เขาวงกตหนู การสัมผัสโคเคนแอมเฟตามีนและแอลกอฮอลล์นั้นเกี่ยวข้องกับการเรียนรู้ที่เพิ่มขึ้นในงานเขาวงกตที่พึ่งพา DLS หรือใช้กลยุทธ์การตอบสนองที่พึ่งพา DLS มากขึ้นในเขาวงกตสองทาง (25, 63, 64) ในมนุษย์การใช้สารที่ถูกทารุณกรรมรวมถึงแอลกอฮอล์และยาสูบมีความสัมพันธ์กับการใช้กลยุทธ์การเดินเรือแบบพึ่งพาอาศัยกันมากขึ้นหลัง striatum ในเขาวงกตเสมือนจริง (65) ดังนั้นยาเสพติดบางชนิดอาจเพิ่มหน่วยความจำนิสัยที่ขึ้นกับ DLS และการมีส่วนร่วมที่สูงขึ้นของระบบหน่วยความจำ DLS นี้อาจเร่งการเปลี่ยนจากการใช้ยาเพื่อการพักผ่อนไปเป็นยาเสพติดเป็นประจำ กลไกที่เสนอนี้สอดคล้องกับสีขาว (1) การโต้เถียงดั้งเดิมที่บางครั้งยาเสพติดอาจทำให้การดูแลตนเองของตนเองแย่ลงโดยการเพิ่มฟังก์ชั่นของระบบหน่วยความจำ
การแข่งขันระหว่างระบบความจำ
แม้ว่ามันจะเป็นไปได้ที่ยาเสพติดจะเพิ่มความจำให้กับนิสัยโดยตรงโดยการเพิ่มฟังก์ชั่นของ DLS [เช่นอ้างอิง (29)] ความเป็นไปได้อีกอย่างคือยาเสพติดเพิ่มความจำนิสัยโดยอ้อมผ่านการปรับระบบหน่วยความจำอื่น กลไกทางเลือกนี้ใช้สมมุติฐานว่าในบางสถานการณ์การเรียนรู้ระบบหน่วยความจำจะแข่งขันกันเพื่อควบคุมการเรียนรู้และโดยการลดการทำงานของระบบหน่วยความจำหนึ่งระบบอาจทำให้การทำงานของระบบอื่นสมบูรณ์ (11, 66) โดยเฉพาะอย่างยิ่งฮิบโปแคมปัสและ DLS บางครั้งอาจแข่งขันกันเพื่อควบคุมการเรียนรู้โดยรอยโรคของฮิปโปแคมปัสจะเพิ่มฟังก์ชั่นหน่วยความจำที่พึ่งพา DLS (5, 6, 67, 68) การทำงานร่วมกันของคู่แข่งยังสามารถแสดงให้เห็นได้ในงานแบบดูอัลโซลูชั่นเมื่อระบบหน่วยความจำหนึ่งที่เสียไปนั้นส่งผลให้เกิดการใช้กลยุทธ์ที่เป็นสื่อกลางโดยระบบอื่นที่ไม่เป็นอันตราย ตัวอย่างเช่นสัตว์ที่ได้รับรอยโรค DMS แสดงการตอบสนองตามนิสัยของ DLS เพื่อรับรางวัลอาหารในงานการเรียนรู้ด้วยเครื่องมือ (44).
เมื่อพิจารณาถึงปฏิสัมพันธ์ที่มีการแข่งขันซึ่งบางครั้งเกิดขึ้นระหว่างระบบหน่วยความจำความเป็นไปได้อย่างหนึ่งคือยาบางชนิดอาจใช้เพิ่มประสิทธิภาพหน่วยความจำตามนิสัยของ DLS โดยอ้อมโดยทำให้กลไกหน่วยความจำความรู้ความเข้าใจบกพร่องโดย DMS ตามที่ระบุไว้ก่อนหน้านี้แอลกอฮอล์มีความเกี่ยวข้องกับการใช้หน่วยความจำนิสัยที่ขึ้นกับ DLS มากขึ้นในทางคดเคี้ยวและคานเร่งด่วน24, 61, 62, 64, 65) หลักฐานบ่งชี้ว่าแอลกอฮอล์ลดการเรียนรู้ในงานความจำเชิงพื้นที่ที่ขึ้นอยู่กับฮิบโปแคมปัส [(64, 69-72); สำหรับการตรวจสอบดูที่อ้างอิง (73)] รวมถึงงานการเรียนรู้การกลับรายการที่พึ่งพา DMS (74-77) สอดคล้องกับการโต้ตอบการแข่งขันระหว่างระบบหน่วยความจำมันได้รับการตั้งสมมติฐานว่าแอลกอฮอล์อาจอำนวยความสะดวกขึ้นอยู่กับหน่วยความจำนิสัย DLS ทางอ้อมผ่านการทำให้เสียความรู้ความเข้าใจกลไกหน่วยความจำ (78).
ควรสังเกตว่านอกเหนือจากแอลกอฮอล์แล้วยังมียาอีกมากมายที่เกี่ยวข้องกับการขาดความจำทางปัญญา การสัมผัสกับมอร์ฟีนเฮโรอีนเมทแอมเฟตามีน MDMA (อี) หรือโคเคนเรื้อรังในทำนองเดียวกันก่อให้เกิดความผิดปกติของความจำเชิงพื้นที่ที่ขึ้นอยู่กับฮิบโปแคมปัส79-89) มันเป็นเรื่องน่าดึงดูดที่จะคาดเดาว่าตามที่แนะนำสำหรับแอลกอฮอล์ความบกพร่องทางความจำทางปัญญาที่ผลิตโดยยาเสพติดอาจเพิ่มความจำนิสัยในทางอ้อมขึ้นอยู่กับ DLS และอาจเป็นกลไกหนึ่งที่อนุญาตให้การบริหารตนเองของยาเสพติดกลายเป็นนิสัย ในทางกลับกันก็เป็นไปได้ว่าการขาดดุลการเรียนรู้เชิงพื้นที่ที่ผลิตโดยยาเสพติดอาจเกิดขึ้นทางอ้อมผ่านการเพิ่มประสิทธิภาพของกระบวนการหน่วยความจำขึ้นอยู่กับ DLS สอดคล้องกับสมมติฐานนี้การกระตุ้นกิจกรรม CREB ใน DLS บั่นทอนความจำเชิงพื้นที่ที่ขึ้นอยู่กับฮิบโปแคมปัส (90) ในขณะที่การยับยั้งกิจกรรม CREB ใน DLS จะลดความผิดปกติของความจำเชิงพื้นที่ที่เกิดจากมอร์ฟีน (91).
บทบาทของความเครียดและความวิตกกังวล
การพิจารณาเพิ่มเติมเกี่ยวกับวิธีการหลายระบบหน่วยความจำในการติดยาเสพติดคือบทบาทของความเครียด หลักฐานชี้ให้เห็นว่าการใช้อารมณ์แปรปรวนทางอารมณ์เอื้อให้หน่วยความจำนิสัย DLS ขึ้นอยู่กับสัตว์ฟันแทะและมนุษย์ (9-12)] การบริหารยา anxiogenic ช่วยเพิ่มการเรียนรู้การตอบสนองขึ้นอยู่กับ DLS ในน้ำบวกเขาวงกต92-97) การเพิ่มประสิทธิภาพของหน่วยความจำนิสัยที่ต้องพึ่งพา DLS นี้ยังได้รับการสังเกตหลังจากได้รับแรงกดดันจากพฤติกรรมที่ไม่มีเงื่อนไขเช่นความยับยั้งชั่งใจเรื้อรังหางช็อคกลิ่นนักล่าเป็นต้น (98-101)] และการเปิดรับสิ่งเร้าที่มีเงื่อนไขที่น่ากลัว102, 103)] แม้ว่าจะแสดงให้เห็นในขั้นต้นในหนู92) การเพิ่มประสิทธิภาพของหน่วยความจำนิสัยที่เกิดจากอารมณ์เร้าอารมณ์ที่แข็งแกร่งยังได้รับการพิสูจน์อย่างกว้างขวางในมนุษย์ (99, 104-110).
กลไกที่ช่วยให้ความเครียด / ความวิตกกังวลเพื่ออำนวยความสะดวกความจำนิสัยส่วนใหญ่ยังไม่ทราบ; แม้กระนั้นหลักฐานบ่งชี้ว่ามีบทบาทที่สำคัญในการควบคุมของ BLA (93-95, 100) สอดคล้องกับการโต้ตอบการแข่งขันระหว่างระบบหน่วยความจำหลักฐานบางอย่างยังแสดงให้เห็นว่าความเครียด / ความวิตกกังวลอาจเพิ่มหน่วยความจำนิสัยขึ้นอยู่กับ DLS ทางอ้อมโดยการลดฟังก์ชั่น hippocampal (94, 95).
การเพิ่มหน่วยความจำนิสัยตามความเครียดหรือความวิตกกังวลอาจเกี่ยวข้องกับการทำความเข้าใจปัจจัยสำคัญบางอย่างที่นำไปสู่การใช้ยาเสพติด กล่าวคือเหตุการณ์ในชีวิตที่ตึงเครียดหรือความเครียด / ความกังวลเรื้อรังที่ยืดเยื้อมานานเกี่ยวข้องกับความอ่อนแอที่เพิ่มขึ้นของการติดยาเสพติดและการกำเริบของโรคในมนุษย์ (111-117) และการสังเกตที่คล้ายกันได้ทำในรูปแบบสัตว์ของการบริหารจัดการยาด้วยตนเอง (118)] นักวิจัยได้แนะนำว่าสอดคล้องกับอิทธิพลของอารมณ์ความรู้สึกในระบบหน่วยความจำหลายระบบ (10) ความเครียดเฉียบพลันหรือเรื้อรังอาจเพิ่มประสิทธิภาพการติดยาเสพติดและการกำเริบของโรคในมนุษย์โดยการมีส่วนร่วมในกระบวนการจำนิสัยขึ้นอยู่กับ DLS (9, 49, 119) สอดคล้องกับข้อเสนอแนะนี้ความเครียดในบุคคลที่ขึ้นอยู่กับโคเคนมีความเกี่ยวข้องกับกิจกรรมลดลง - ขึ้นอยู่กับระดับออกซิเจนออกซิเจน (BOLD) ในฮิบโปและกิจกรรมที่เพิ่มขึ้นใน striatum หลังและการเปลี่ยนแปลงกิจกรรม BOLD เหล่านี้เกี่ยวข้องกับความอยากโคเคนความเครียด (120).
แหล่งที่มาของการเสพติด
นอกเหนือจากยาเสพติดแล้วการใช้ระบบหน่วยความจำหลายข้อก็ถูกนำมาใช้เพื่อทำความเข้าใจกับแหล่งติดยาอื่น ๆ ตัวอย่างเช่นการเพิ่มขึ้นของโรคอ้วนในช่วงสองสามทศวรรษที่ผ่านมานำไปสู่การเพิ่มขึ้นของความสนใจในการทดลองโดยนักวิจัยจำนวนมากวาดแนวระหว่างการติดยาเสพติดและการกินมากเกินไป (121-123)] หลักฐานล่าสุดบางอย่างชี้ให้เห็นว่าเช่นการติดยาเสพติดการติดอาหารอาจมีสาเหตุมาจากความผูกพันที่เพิ่มขึ้นของความจำนิสัยที่ขึ้นกับ DLS ในหนูการบริโภคอาหารที่ดื่มด่ำเหมือนช่วยอำนวยความสะดวกในการเปลี่ยนจากการรับรู้เป็นการควบคุมพฤติกรรมที่เป็นนิสัย (124, 125) ยิ่งกว่านั้นพฤติกรรมที่เป็นนิสัยในสัตว์ที่ดื่มสุรานั้นมีความเกี่ยวข้องกับกิจกรรม DLS ที่เพิ่มขึ้นและอาจป้องกันได้โดยการปิดกั้น AMPA หรือตัวรับ Dopamine D1 ใน DLS (125) โรคอ้วนที่เกิดจากอาหารได้รับการเชื่อมโยงกับการใช้หน่วยความจำนิสัยในงาน Y-maze126).
ความผิดปกติทางพฤติกรรมที่เกิดขึ้นอีกประการหนึ่งที่มีความคล้ายคลึงกับคุณสมบัติบางประการของการติดยาเสพติดคือการเล่นวิดีโอเกมทางพยาธิวิทยาหรือการเสพติดวิดีโอเกม (127)] เช่นเดียวกับการติดยาเสพติดการเล่นวิดีโอเกมที่มากเกินไปในระยะยาวนั้นสัมพันธ์กับการรับสารโดปามีน D2 ซึ่งลดลงในการผูกไว้กับหลัง striatum (128) การเล่นวิดีโอเกมมีความสัมพันธ์กับการเปิดใช้งานของ striorsum ด้านหลังที่เพิ่มขึ้น (129, 130) และปริมาตรของกระดูกสันหลังส่วนหลังที่มากขึ้นทำนายระดับความสามารถของวิดีโอเกมที่สูงขึ้น (131) ผู้ที่เล่นวิดีโอเกมแอคชั่นเป็นประจำมีแนวโน้มที่จะใช้หน่วยความจำนิสัยที่ขึ้นอยู่กับหลังของ striatum ในเขาวงกตเสมือนจริง (132) และการเล่นวิดีโอเกมก่อนการฝึกอบรมนำไปสู่การตอบสนองอย่างเป็นกิจวัตรเหนือเป้าหมายที่ตอบสนองในงานการตัดสินใจสองขั้นตอน (133) ดังนั้นตามที่เสนอเพื่อการละเมิดยาเสพติดการเล่นวิดีโอเกมอาจช่วยเพิ่มการติดวิดีโอเกมผ่านการมีส่วนร่วมกับระบบหน่วยความจำนิสัยแบบพึ่งพา DLS
ในที่สุดวิธีการหลายระบบหน่วยความจำอาจเป็นประโยชน์สำหรับการทำความเข้าใจกับการติดกัญชา แม้ว่ากัญชาอาจมีโอกาสถูกล่วงละเมิดต่ำกว่าสารผิดกฎหมายอื่น ๆ ที่ได้รับการพิจารณาในบริบทของการวิจัยการติดยาเสพติด (เช่นโคเคนมอร์ฟีนเฮโรอีน ฯลฯ ) การใช้กัญชาอย่างหนักสามารถส่งเสริมการพึ่งพายาเสพติดและอาการถอนได้ การละเมิด134-137) เมื่อไม่นานมานี้มีข้อเสนอแนะว่าการติดกัญชาอาจเกิดจากการมีส่วนร่วมของหน่วยความจำนิสัยตาม DLS ที่เพิ่มขึ้น (138) ในขณะที่การสัมผัสของ cannabinoid เฉียบพลันจะทำให้การทำงานของหน่วยความจำขึ้นอยู่กับ DLS (139, 140) การได้รับ cannabinoid ซ้ำ ๆ นำไปสู่การตอบสนองตามนิสัยแบบพึ่งพา DLS มากขึ้นในงานการเรียนรู้ด้วยเครื่องมือ (141) นอกจากนี้ผู้ใช้กัญชาจำนวนมากแสดงการเปิดใช้งานที่ดีกว่าของ striorsum หลังซึ่งสัมพันธ์กับผู้ที่ไม่ใช่ผู้ใช้เมื่อดำเนินการรุ่นกัญชาของงานเชื่อมโยงโดยนัย (142) และผู้เข้าร่วมที่มีประวัติของการใช้กัญชามีแนวโน้มที่จะใช้หน่วยความจำนิสัยหลังขึ้นอยู่กับ striatum หลังในเขาวงกตเสมือน (65).
เมื่อแอปพลิเคชันที่ประสบความสำเร็จของระบบหน่วยความจำเข้าใกล้แหล่งที่มาของการติดยาก็มีเหตุผลที่จะตั้งสมมติฐานว่าระบบหน่วยความจำหลายระบบอาจมีส่วนเกี่ยวข้องในพฤติกรรมเชิงพฤติกรรมอื่น ๆ ที่เกี่ยวข้องกับการติดยาเสพติด แน่นอนว่าแนวทางของระบบหน่วยความจำอาจมีประโยชน์สำหรับการทำความเข้าใจการพนันทางพยาธิวิทยาก็ได้รับความสนใจเช่นกัน (143, 144).
สรุป
หลักฐานการทดลองยี่สิบปีมีหลักฐานส่วนใหญ่ของไวท์ (1) ระบบหน่วยความจำหลายวิธีในการติดยาเสพติด หลักฐานชี้ให้เห็นว่าฮิบโปเป็นสื่อกลางในการควบคุมบริบทของการบริหารตนเองของยาเสพติด, DLS เป็นสื่อกลางในการตอบสนองต่อการเสริมแรงด้วยยา S-R เป็นประจำ นอกจากนี้การวิจัยที่ตามมาได้นำไปสู่ความเข้าใจเพิ่มเติมเกี่ยวกับมุมมองหลายระบบหน่วยความจำของการติดยาเสพติดรวมถึงการเปลี่ยนเป็นความทรงจำที่เป็นนิสัยการแข่งขันระหว่างระบบความจำ
การวิจัยในอนาคตควรพยายามบูรณาการระบบหน่วยความจำเข้ากับทฤษฎีการเสพติดอื่น ๆ เช่นกระบวนการสร้างแรงบันดาลใจของฝ่ายตรงข้าม (145) นอกจากนี้ยังจะเป็นประโยชน์ในการรวมเข้าไปในระบบหน่วยความจำดูคุณสมบัติเพิ่มเติมของการติดเช่นการพึ่งพายาความอดทนและการถอน แม้ว่าการทบทวนในปัจจุบันจะเน้นไปที่บริเวณสมองที่พิจารณาโดย White (เช่น the hippocampus, dorsal striatum และ amygdala) แต่ก็ควรสังเกตว่าบริเวณสมองที่เกี่ยวข้องกับการเรียนรู้และความทรงจำนั้นมีส่วนเกี่ยวข้องอย่างมากในการติดยาและการกำเริบของโรค รวมถึงเยื่อหุ้มสมองด้านหน้า prefrontal และนิวเคลียส accumbens [สำหรับการตรวจสอบดูอ้างอิง (13)] ในที่สุดแม้ว่าจะอยู่นอกเหนือขอบเขตของการทบทวนในปัจจุบันก็ควรได้รับการยอมรับว่ามีหลักฐานมากมายแสดงให้เห็นว่าการเปลี่ยนแปลงระดับเซลล์และโมเลกุลในระบบ dopaminergic midbrain ก็มีส่วนทำให้เกิดการติดยา (146).
แม้ว่าความทรงจำที่เป็นนิสัยอาจควบคุมได้ยากเป็นพิเศษ แต่หลักฐานบางอย่างระบุว่าหน่วยความจำที่ขึ้นกับ DLS ที่ได้มาครั้งหนึ่งสามารถระงับได้ในบางสถานการณ์147) หรือแม้กระทั่งย้อนกลับ (148, 149) ดังนั้นจึงเป็นไปได้ว่าการเปลี่ยนแปลงทางเภสัชวิทยาและขั้นตอนพฤติกรรมที่นำไปสู่การกลับรายการหรือการปราบปรามความจำนิสัยในรูปแบบการเรียนรู้ของสัตว์อาจมีการปรับเพื่อรักษายาเสพติดและการกำเริบของโรคในมนุษย์
ผลงานของผู้เขียน
JG และ MP ต่างก็มีส่วนร่วมในการคิดและการเขียนมินิรีวิวในปัจจุบัน
คำชี้แจงความขัดแย้งทางผลประโยชน์
ผู้เขียนประกาศว่าการวิจัยได้ดำเนินการในกรณีที่ไม่มีความสัมพันธ์ทางการค้าหรือทางการเงินใด ๆ ที่อาจตีความได้ว่าเป็นความขัดแย้งทางผลประโยชน์ที่อาจเกิดขึ้น
อ้างอิง
1. White NM ยาเสพติดที่ใช้เป็นตัวช่วยเสริม: การกระทำบางส่วนหลายอย่างในระบบหน่วยความจำ ติดยาเสพติด (1996) 91(7):921–50. doi: 10.1111/j.1360-0443.1996.tb03586.x
2. White NM, McDonald RJ ระบบหน่วยความจำแบบขนานหลายระบบในสมองของหนู Neurobiol Learn Mem (2002) 77(2):125–84. doi:10.1006/nlme.2001.4008
3. นายพล LR ระบบความจำของสมอง: ประวัติโดยย่อและมุมมองปัจจุบัน Neurobiol Learn Mem (2004) 82(3):171–7. doi:10.1016/j.nlm.2004.06.005
4. White NM, Packard MG, McDonald RJ การแยกจากกันของระบบหน่วยความจำ: เรื่องราวแผ่ออกไป Behav Neurosci (2013) 127(6):813–34. doi:10.1037/a0034859
5. Packard MG, Hirsh R, White NM ผลกระทบที่แตกต่างของฟอร์นิกซ์และรอยแผลนิวเคลียสในงานเขาวงกตเรเดียลสองงาน: หลักฐานสำหรับระบบหน่วยความจำหลายระบบ Neurosci J (1989) 9(5): 1465 72-
6. McDonald RJ, White NM การแยกตัวของระบบหน่วยความจำสามเท่า: ฮิปโปแคมปัส, อะมิกดาลา, และหลัง striatum. Behav Neurosci (1993) 107(1):3–22. doi:10.1037/0735-7044.107.1.3
7. Maren S. Neuroiology ของการปรับความกลัว Pavlovian Annu Rev Neurosci (2001) 24(1):897–931. doi:10.1146/annurev.neuro.24.1.897
8. McGaugh JL อะมิกดาลาปรับเปลี่ยนการรวมความทรงจำของประสบการณ์การปลุกอารมณ์ Annu Rev Neurosci (2004) 27: 1-28 ดอย: 10.1146 / annurev.neuro.27.070203.144157
9. แพคการ์ด MG ความวิตกกังวลความรู้ความเข้าใจและนิสัย: มุมมองของระบบหน่วยความจำที่หลากหลาย สมอง Res (2009) 1293: 121 8- ดอย: 10.1016 / j.brainres.2009.03.029
10. แพคการ์ด MG, กู๊ดแมนเจอารมณ์เร้าอารมณ์และระบบความจำหลายอย่างในสมองของสัตว์เลี้ยงลูกด้วยนม ด้านหน้า Behav Neurosci (2012) 6: 14 ดอย: 10.3389 / fnbeh.2012.00014
11. Packard MG, Goodman J. ปัจจัยที่มีอิทธิพลต่อการใช้ระบบหน่วยความจำหลายระบบแบบสัมพัทธ์ Hippocampus (2013) 23(11):1044–52. doi:10.1002/hipo.22178
12. Schwabe L. ความเครียดและการมีส่วนร่วมของระบบหน่วยความจำหลายอย่าง: การบูรณาการการศึกษาสัตว์และมนุษย์ Hippocampus (2013) 23(11):1035–43. doi:10.1002/hipo.22175
13. Everitt BJ, Robbins TW ระบบประสาทของการเสริมแรงสำหรับการติดยาเสพติด: จากการกระทำไปจนถึงนิสัยการบังคับ Nat Neurosci (2005) 8(11):1481–9. doi:10.1038/nn1579
14. Fuchs RA, Evans KA, Ledford CC, Parker MP, กรณี JM, Mehta RH, และคณะ บทบาทของเยื่อหุ้มสมองส่วนหน้า dorsomedial, basolateral amygdala, และ dorsal hippocampus ในการคืนสถานะตามบริบทของโคเคนที่แสวงหาหนู Neuropsychopharmacology (2005) 30(2):296–309. doi:10.1038/sj.npp.1300579
15. Fuchs RA, Eaddy JL, Su ZI, Bell GH ปฏิกิริยาของ amygdala basolateral กับ dorsal hippocampus และ dorsomedial prefrontal cortex ควบคุมการคืนสถานะของยาเสพติดที่เกิดจากบริบทของการค้นหาโคเคนในหนู Eur J Neurosci (2007) 26(2):487–98. doi:10.1111/j.1460-9568.2007.05674.x
16. Kramar CP, Barbano MF, Medina JH ต้องมีตัวรับ Dopamine D1 / D5 ในฮิปโปแคมปัสที่ด้านหลังสำหรับการได้มาและการแสดงออกของหน่วยความจำโคเคนที่เกี่ยวข้องกับการทดลองเดียว Neurobiol Learn Mem (2014) 116: 172 80- ดอย: 10.1016 / j.nlm.2014.10.004
17. Zapata A, Minney VL, Shippenberg TS เปลี่ยนจากเป้าหมายไปสู่โคเคนเป็นนิสัยเพื่อค้นหาประสบการณ์ที่ยาวนานในหนู Neurosci J (2010) 30(46):15457–63. doi:10.1523/JNEUROSCI.4072-10.2010
18. Corbit LH, Nie H, Janak PH การค้นหาแอลกอฮอล์ที่เป็นนิสัย: หลักสูตรเวลาและการสนับสนุนอนุภูมิภาคของแถบด้านหลัง จิตเวช Biol (2012) 72(5):389–95. doi:10.1016/j.biopsych.2012.02.024
19. Whitelaw RB, Markou A, Robbins TW, Everitt BJ บาดแผลที่เกิดจาก Excitotoxic ของ amygdala basolateral ทำให้การแสวงหาพฤติกรรมการค้นหาโคเคนลดน้อยลงภายใต้กำหนดการเสริมครั้งที่สอง Psychopharmacology (1996) 127(1–2):213–24. doi:10.1007/BF02805996
20. Alderson HL, Robbins TW, Everitt BJ ผลของรอยโรค excitotoxic ของ amygdala basolateral ต่อการได้มาของพฤติกรรมการแสวงหาเฮโรอีนในหนู Psychopharmacology (2000) 153(1):111–9. doi:10.1007/s002130000527
21. Gabriele A ดู RE การย้อนกลับของ amygdala basolateral แต่ไม่ใช่ dorsolateral caudate putamen, attenuates รวมของการเรียนรู้การเชื่อมโยงโคเคน - คิวในรูปแบบสถานะของการแสวงหายาเสพติด Eur J Neurosci (2010) 32(6):1024–9. doi:10.1111/j.1460-9568.2010.07394.x
22. Sciascia JM, Reese RM, Janak PH, Chaudhri N. การค้นหาเครื่องดื่มแอลกอฮอล์ที่ถูกกระตุ้นโดยตัวชี้นำ Pavlovian ที่ไม่ต่อเนื่องนั้นได้รับการฟื้นฟูโดยบริบทแอลกอฮอล์ Neuropsychopharmacology (2015) 40: 2801 12- ดอย: 10.1038 / npp.2015.130
23. Packard MG, Teather LA การปรับ Amygdala ของระบบหน่วยความจำหลายระบบ: hippocampus และ caudate-putamen Neurobiol Learn Mem (1998) 69(2):163–203. doi:10.1006/nlme.1997.3815
24. Dickinson A, Wood N, Smith JW แอลกอฮอล์กำลังค้นหาโดยหนู: การกระทำหรือนิสัย? QJ Exp Psychol B (2002) 55(4):331–48. doi:10.1080/0272499024400016
25. Udo T, Ugalde F, DiPietro N, Eichenbaum HB, Kantak KM ผลของการบริหารตนเองด้วยโคเคนแบบถาวรต่อการเรียนรู้จากการขึ้นอยู่กับ amygdala และหลัง striatum ขึ้นอยู่กับหนู Psychopharmacology (2004) 174(2):237–45. doi:10.1007/s00213-003-1734-1
26. Wood SC, Fay J, Sage JR, Anagnostaras SG การจำกัดความกลัวโคเคนและพาฟโลเวียน: การวิเคราะห์ปริมาณรังสี Behav Brain Res (2007) 176(2):244–50. doi:10.1016/j.bbr.2006.10.008
27. Wood SC, Anagnostaras SG. ความทรงจำและ psychostimulants: การปรับของความกลัว Pavlovian โดยยาบ้าใน C57BL / 6 หนู Psychopharmacology (2009) 202(1–3):197–206. doi:10.1007/s00213-008-1185-9
28. Iñiguez SD, Charntikov S, Baella SA, Herbert MS, Bolaños-Guzmán CA, Crawford CA การเปิดรับโคเคนหลังการฝึกอบรมจะช่วยอำนวยความสะดวกในการรวมหน่วยความจำเชิงพื้นที่ในหนู C57BL / 6 Hippocampus (2012) 22(4):802–13. doi:10.1002/hipo.20941
29. DePoy L, Daut R, Brigman JL, MacPherson K, Crowley N, Gunduz-Cinar O, et al. แอลกอฮอล์เรื้อรังทำให้เกิดการปรับตัวของประสาท Proc Natl Acad Sci สหรัฐอเมริกา (2013) 110(36):14783–8. doi:10.1073/pnas.1308198110
30. Leri F, Nahas E, Henderson K, Limebeer CL, Parker LA, White NM ผลของเฮโรอีนหลังการฝึกอบรมและแอมเฟตามีนต่อการรวมการเรียนรู้แบบ win-stay และการปรับสภาพความกลัว J Psychopharmacol (2013) 27(3):292–301. doi:10.1177/0269881112472566
31. Schmitzer-Torbert N, Apostolidis S, Amoa R, O'Rear C, Kaster M, Stowers J, et al. การบริหารโคเคนหลังการฝึกอบรมอำนวยความสะดวกในการเรียนรู้นิสัยและต้องใช้เยื่อหุ้มสมอง infralimbic และ dorsolateral striatum Neurobiol Learn Mem (2015) 118: 105 12- ดอย: 10.1016 / j.nlm.2014.11.007
32. โนว์ลตันบีเจ ฐานปมประสาท: การสร้างนิสัย ใน: Jaeger D, Jung R, บรรณาธิการ สารานุกรมประสาทวิทยาศาสตร์การคำนวณ. นิวยอร์ก: สปริงเกอร์ (2014) พี 1-17
33. Tolman EC, Ritchie BF, Kalish D. การศึกษาในการเรียนรู้เชิงพื้นที่ IV การถ่ายโอนสถานที่การเรียนรู้ไปยังเส้นทางเริ่มต้นอื่น ๆ J Exp Psychol (1947) 37(1):39–47. doi:10.1037/h0062061
34. Ritchie BF, Aeschliman B, Pierce P. การศึกษาในการเรียนรู้เชิงพื้นที่ VIII ประสิทธิภาพของสถานที่และการได้มาของการจัดการสถานที่ J Comp Physiol Psychol (1950) 43(2):73–85. doi:10.1037/h0055224
35. ฮิกส์ LH ผลของการฝึก overtraining ต่อการได้มาและการพลิกกลับของสถานที่และการเรียนรู้การตอบสนอง ตัวแทน Psychol (1964) 15(2):459–62. doi:10.2466/pr0.1964.15.2.459
36. Packard MG, McGaugh JL การหยุดใช้งานของฮิปโปแคมปัสหรือนิวเคลียส caudate กับ lidocaine มีผลต่อการแสดงออกของสถานที่และการเรียนรู้การตอบสนองที่แตกต่างกัน Neurobiol Learn Mem (1996) 65(1):65–72. doi:10.1006/nlme.1996.0007
37. Yin HH, Knowlton BJ ผลงานของภูมิภาคย่อยเพื่อการเรียนรู้และการตอบสนอง เรียนรู้ Mem (2004) 11(4):459–63. doi:10.1101/lm.81004
38. อดัมส์ซีดี, ดิกคินสัน A. เครื่องมือตอบสนองต่อการลดค่าเงินของ reinforcer QJ Exp Psychol (1981) 33B: 109 12- ดอย: 10.1080 / 14640748108400816
39. อดัมส์ซีดี, ดิกคินสัน A. การกระทำและนิสัย: การเปลี่ยนแปลงในการเชื่อมโยงระหว่างการเรียนรู้ด้วยเครื่องมือ ใน: Spear NE, Miller RR, editors การประมวลผลข้อมูลในสัตว์: กลไกความจำ. Hillsdale, นิวเจอร์ซีย์: Erlbaum (1981) พี 143-65
40. อดัมส์ซีดี ความแปรปรวนของความไวของเครื่องมือตอบสนองต่อการลดค่าเงินของ reinforcer QJ Exp Psychol (1982) 34B: 77 98- ดอย: 10.1080 / 14640748208400878
41. Dickinson A, Nicholas DJ การเรียนรู้เพื่อสร้างแรงจูงใจที่ไม่เกี่ยวข้องระหว่างการใช้เครื่องมือ: บทบาทของความสัมพันธ์ระหว่างผู้ผลักดันและการตอบสนอง QJ Exp Psychol (1983) 35B: 249 63- ดอย: 10.1080 / 14640748308400909
42. Dickinson A, Nicholas DJ, Adams CD ผลกระทบของความไม่แน่นอนของเครื่องมือที่มีต่อความอ่อนไหวต่อการลดค่าเงิน QJ Exp Psychol (1983) 35B: 35 51- ดอย: 10.1080 / 14640748308400912
43. Corbit LH, Balleine BW บทบาทของฮิบโปในการปรับสภาพเครื่องมือ Neurosci J (2000) 20(11): 4233 9-
44. Yin HH, Ostlund SB, Knowlton BJ, Balleine BW บทบาทของ striatum dorsomedial ในการปรับสภาพเครื่องมือ Eur J Neurosci (2005) 22:513–23. doi:10.1111/j.1460-9568.2005.04218.x
45. Yin HH, Knowlton BJ, Balleine BW รอยโรคของ dorsolateral striatum รักษาความคาดหวังผล แต่ขัดขวางการสร้างนิสัยในการเรียนรู้ด้วยเครื่องมือ Eur J Neurosci (2004) 19:181–9. doi:10.1111/j.1460-9568.2004.03095.x
46. Quinn JJ, Pittenger C, Lee AS, Pierson JL, Taylor JR Striatum-based อุปนิสัยไม่รู้สึกตัวทั้งการเพิ่มและการลดลงของค่า reinforcer ในหนู Eur J Neurosci (2013) 37: 1012 21- ดอย: 10.1111 / ejn.12106
47. Yin HH จากการกระทำสู่นิสัย: การปรับระบบประสาทนำไปสู่การพึ่งพา แอลกอฮอล์ที่ดีต่อสุขภาพ (2008) 31(4): 340 4-
48. Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ กระบวนการเรียนรู้แบบขนานและเชิงโต้ตอบภายในฐานปมประสาท: ความเกี่ยวข้องสำหรับความเข้าใจในการเสพติด Behav Brain Res (2009) 199(1):89–102. doi:10.1016/j.bbr.2008.09.027
49. Schwabe L, Dickinson A, Wolf OT ความเครียดนิสัยและการติดยาเสพติด: มุมมองทางจิตประสาทต่อมไร้ท่อ Exp Clin Psychopharmacol (2011) 19(1):53–63. doi:10.1037/a0022212
50. Hogarth L, Balleine BW, Corbit LH, Killcross S. กลไกการเรียนรู้ที่เชื่อมโยงการเปลี่ยนจากการใช้ยาเพื่อการพักผ่อนไปสู่การเสพติด Ann NY Acad Sci (2013) 1282(1):12–24. doi:10.1111/j.1749-6632.2012.06768.x
51. Murray JE, Belin D, Everitt BJ การแยกส่วนสองส่วนของการควบคุมการเกิดโคโลนัยของ dorsomedial และ dorsolateral มากกว่าการได้มาและประสิทธิภาพของการแสวงหาโคเคน Neuropsychopharmacology (2012) 37(11):2456–66. doi:10.1038/npp.2012.104
52. Clemens KJ, Castino MR, Cornish JL, Goodchild AK, Holmes NM พื้นผิวพฤติกรรมและระบบประสาทของการสร้างนิสัยในหนูนิโคตินทางหลอดเลือดดำด้วยตนเองจัดการ Neuropsychopharmacology (2014) 39: 2584 93- ดอย: 10.1038 / npp.2014.111
53. Corbit LH, Nie H, Janak PH การตอบสนองตามนิสัยสำหรับแอลกอฮอล์นั้นขึ้นอยู่กับการส่งสัญญาณของ AMPA และ D2 receptor ใน dorsolateral striatum ด้านหน้า Behav Neurosci (2014) 8: 301 ดอย: 10.3389 / fnbeh.2014.00301
54. Schoenbaum G, Setlow B. Cocaine ทำให้การกระทำไม่รู้สึกตัวกับผลลัพธ์ แต่ไม่สูญพันธุ์: ความหมายของฟังก์ชั่น orbitofrontal-amygdalar ที่มีการเปลี่ยนแปลง Cortex cereb (2005) 15(8):1162–9. doi:10.1093/cercor/bhh216
55. เนลสันเอ, การสัมผัสคิลครอสเอสแอมเฟตามีนช่วยเพิ่มการสร้างนิสัย Neurosci J (2006) 26(14):3805–12. doi:10.1523/JNEUROSCI.4305-05.2006
56. Nordquist RE, Voorn P, De Mooij-van Malsen JG, Joosten RNJMA, Pennartz CMA, Vanderschuren LJMJ เติมสารเสริมคุณค่าและเร่งการสร้างนิสัยหลังจากการรักษายาบ้าซ้ำ Eur Neuropsychopharmacol (2007) 17(8):532–40. doi:10.1016/j.euroneuro.2006.12.005
57. LeBlanc KH, แม่บ้าน NT, Ostlund SB การสัมผัสโคเคนซ้ำ ๆ ช่วยให้เกิดการแสดงออกของแรงจูงใจและกระตุ้นการควบคุมนิสัยในหนู PLoS One (2013) 8: e61355 ดอย: 10.1371 / journal.pone.0061355
58. เนลสัน AJ, Killcross S. การก่อตัวของพฤติกรรมเร่งด่วนหลังจากได้รับแอมเฟตามีนใน D1 แต่ได้รับการปรับปรุงโดย D2 คู่อริตัวรับ ด้านหน้า Neurosci (2013) 7: 76 ดอย: 10.3389 / fnins.2013.00076
59. Corbit LH, Chieng BC, Balleine BW ผลของการได้รับโคเคนซ้ำ ๆ ต่อการเรียนรู้นิสัยและการกลับรายการโดย N-acetylcysteine Neuropsychopharmacology (2014) 39(8):1893–901. doi:10.1038/npp.2014.37
60. ไมล์ FJ, Everitt BJ, ดิกคินสัน A. การค้นหาโคเคนด้วยหนู: การกระทำหรือนิสัย? Behav Neurosci (2003) 117(5):927–38. doi:10.1037/0735-7044.117.5.927
61. Mangieri RA, Cofresí RU, Gonzales RA การค้นหาเอทานอลโดยหนูอีแวนส์ยาวนั้นไม่ใช่พฤติกรรมที่มุ่งเป้าไปที่เป้าหมายเสมอไป PLoS One (2012) 7: e42886 ดอย: 10.1371 / journal.pone.0042886
62. Sjoerds Z, De Wit S, Van Den Brink W, Robbins TW, Beekman ATF, Penninx BWJH, และคณะ หลักฐานเชิงพฤติกรรมและระบบประสาทสำหรับการเรียนรู้นิสัยในผู้ป่วยติดสุรา จิตเวชศาสตร์แปล (2013) 3(12): e337 ดอย: 10.1038 / tp.2013.107
63. Packard MG, McGaugh JL Quinpirole และ d-amphetamine บริหารหลังการฝึกอบรมช่วยเพิ่มความทรงจำเกี่ยวกับการเลือกปฏิบัติเชิงพื้นที่และคิวในเขาวงกตน้ำ Psychobiology (1994) 22(1): 54 60-
64. Matthews DB, Ilgen M, White AM, PJ ที่ดีที่สุด การบริหารเอทานอลแบบเฉียบพลันนั้นทำให้ประสิทธิภาพเชิงพื้นที่ลดลงในขณะที่อำนวยความสะดวกในการทำงานแบบไม่สูบบุหรี่ในหนู Neurobiol Learn Mem (1999) 72(3):169–79. doi:10.1006/nlme.1998.3900
65. Bohbot VD, Balso D, Conrad K, Konishi K, Leyton M. Caudate กลยุทธ์การนำทางที่ขึ้นกับนิวเคลียสสัมพันธ์กับการใช้ยาเสพติดเพิ่มขึ้น Hippocampus (2013) 23(11):973–84. doi:10.1002/hipo.22187
66. Poldrack RA, Packard MG การแข่งขันในระบบหน่วยความจำหลายระบบ: การรวบรวมหลักฐานจากการศึกษาสัตว์และสมองของมนุษย์ Neuropsychologia (2003) 41(3):245–51. doi:10.1016/S0028-3932(02)00157-4
67. Matthews DB, PJ ยอดเยี่ยม รอยโรค Fimbria / fornix ช่วยให้การเรียนรู้ของภารกิจการตอบสนองที่ไม่เชิญชม Psychon Bull Rev (1995) 2(1):113–6. doi:10.3758/BF03214415
68. ชโรเดอร์ JP, Wingard JC, Packard MG โพสต์การฝึกอบรมการยกเลิกการย้อนกลับของฮิบโปแสดงให้เห็นการรบกวนระหว่างระบบหน่วยความจำ Hippocampus (2002) 12(2):280–4. doi:10.1002/hipo.10024
69. Matthews DB, Simson PE, PJ ที่ดีที่สุด เฉียบพลันเอทานอลบั่นทอนหน่วยความจำเชิงพื้นที่ แต่ไม่กระตุ้น / หน่วยความจำการตอบสนองในหนู แอลกอฮอล์ Clin ค่าใช้จ่าย Res (1995) 19(4):902–9. doi:10.1111/j.1530-0277.1995.tb00965.x
70. White AM, Elek TM, Beltz TL, PJ ที่ดีที่สุด ประสิทธิภาพเชิงพื้นที่นั้นมีความอ่อนไหวต่อเอทานอลมากกว่าประสิทธิภาพที่ไม่มีการมองเห็นโดยไม่คำนึงถึงความใกล้ชิดคิว แอลกอฮอล์ Clin ค่าใช้จ่าย Res (1998) 22(9):2102–7. doi:10.1111/j.1530-0277.1998.tb05922.x
71. Matthews DB, Morrow AL, Tokunaga S, McDaniel JR การบริหารเอทานอลแบบเฉียบพลันและการบริหาร allopregnanolone เฉียบพลันทำให้หน่วยความจำในอวกาศสูญเสียไปในน้ำมอร์ริส แอลกอฮอล์ Clin ค่าใช้จ่าย Res (2002) 26(11):1747–51. doi:10.1111/j.1530-0277.2002.tb02479.x
72. Berry RB, Matthews DB การบริหารเอทานอลแบบเฉียบพลันจะช่วยลดความทรงจำเชิงพื้นที่ในหนู C57BL / 6J แอลกอฮอล์ (2004) 32(1):9–18. doi:10.1016/j.alcohol.2003.09.005
73. ซิลเวอร์ JM, Tokunaga S, Berry RB, White AM, Matthews DB การด้อยค่าในการเรียนรู้เชิงพื้นที่และความจำ: เอทานอล, allopregnanolone และฮิบโปแคมปัส การต่อต้านสมอง (2003) 43(3):275–84. doi:10.1016/j.brainresrev.2003.09.002
74. Badanich KA, Becker HC, Woodward JJ ผลของการได้รับเอทานอลแบบเรื้อรังเป็นระยะ ๆ ต่อพฤติกรรมขึ้นอยู่กับเยื่อหุ้มสมอง orbitofrontal และ medial prefrontal cortex ในหนู Behav Neurosci (2011) 125(6):879–91. doi:10.1037/a0025922
75. Coleman LG Jr, He J, Lee J, Styner M, Crews FT การดื่มสุราวัยรุ่นจะเปลี่ยนแปลงการแสดงออกของสารสื่อประสาทในสมองของผู้ใหญ่พฤติกรรมปริมาณของสมองในระดับภูมิภาคและ neurochemistry ในหนู แอลกอฮอล์ Clin ค่าใช้จ่าย Res (2011) 35(4):671–88. doi:10.1111/j.1530-0277.2010.01385.x
76. Kuzmin A, Liljequist S, Meis J, Chefer V, Shippenberg T, Bakalkin G. การใช้เอทานอลในปริมาณปานกลางซ้ำแล้วซ้ำอีกทำให้การทำงานขององค์ความรู้ในหนู Wistar ลดลง ติดยาเสพติด Biol (2012) 17(1):132–40. doi:10.1111/j.1369-1600.2010.00224.x
77. โคลแมน LG, Liu W, Oguz I, Styner M, Crews FT การบำบัดด้วยเอทานอลในวัยรุ่นจะช่วยเปลี่ยนแปลงปริมาณสมองในภูมิภาคผู้ใหญ่โปรตีนเมทริกซ์นอกเซลล์เยื่อหุ้มสมองและความยืดหยุ่นของพฤติกรรม Pharmacol Biochem Behav (2014) 116: 142-51 ดอย: 10.1016 / j.pbb.2013.11.021
78. Matthews DB, Silvers JR การใช้การบริหารเอทานอลเฉียบพลันเป็นเครื่องมือในการตรวจสอบระบบหน่วยความจำหลายระบบ Neurobiol Learn Mem (2004) 82(3):299–308. doi:10.1016/j.nlm.2004.06.007
79. การฝึกอบรม HW, Morford LL, Inman-Wood SL, Fukumura M, Vorhees CV 3, 4-methylenedioxymethamphetamine (ความปีติยินดี) - การเรียนรู้ที่ลดลงและความจำบกพร่องขึ้นอยู่กับอายุของการได้รับสารในช่วงแรกของการพัฒนา Neurosci J (2001) 21(9): 3228 35-
80. Williams MT, Morford LL, Wood SL, Wallace TL, Fukumura M, Broening HW, และคณะ พัฒนาการของการรักษาด้วย d-methamphetamine ทำให้เกิดความบกพร่องในการเดินเรือในหน่วยความจำอ้างอิงในเขาวงกตน้ำมอร์ริสขณะที่ประหยัดหน่วยความจำในการทำงาน ไซแนปส์ (2003) 48(3):138–48. doi:10.1002/syn.10159
81. Vorhees CV, Reed TM, Skelton MR, Williams MT การสัมผัสกับ 3, 4-methylenedioxymethamphetamine (MDMA) ในวันหลังคลอด 11-20 ทำให้เกิดการอ้างอิง แต่ไม่ได้ทำงานขาดความทรงจำในเขาวงกตน้ำมอร์ริสในหนู: ผลกระทบของการเรียนรู้ก่อนหน้า Int J Dev Neurosci (2004) 22(5):247–59. doi:10.1016/j.ijdevneu.2004.06.003
82. โคเฮน MA, Skelton MR, Schaefer TL, Gudelsky GA, Vorhees CV, Williams MT การเรียนรู้และความจำหลังจากได้รับทารกแรกเกิดถึง 3, 4-methylenedioxymethamphetamine (อี) ในหนู: ปฏิสัมพันธ์กับการได้รับสัมผัสในวัยผู้ใหญ่ ไซแนปส์ (2005) 57(3):148–59. doi:10.1002/syn.20166
83. Skelton MR, Williams MT, Vorhees CV การรักษาด้วย MDMA จาก P11-20 ขัดขวางการเรียนรู้เชิงพื้นที่และการเรียนรู้การรวมเส้นทางในหนูวัยรุ่น แต่การเรียนรู้เชิงพื้นที่เฉพาะในหนูที่มีอายุมากกว่า Psychopharmacology (2006) 189(3):307–18. doi:10.1007/s00213-006-0563-4
84. มะ MX, เฉิน YM, เขาเจ, เซง T, วัง JH ผลของมอร์ฟีนและการถอนต่อความจำเชิงพื้นที่ของ Y-maze ในหนู Neuroscience (2007) 147(4):1059–65. doi:10.1016/j.neuroscience.2007.05.020
85. Belcher AM, Feinstein EM, O'Dell SJ, Marshall JF ยาบ้ามีผลต่อความจำ: การเปรียบเทียบการเพิ่มปริมาณและการใช้ยาในหนึ่งวัน Neuropsychopharmacology (2008) 33(6):1453–63. doi:10.1038/sj.npp.1301510
86. Tramullas M, Martínez-Cué C, Hurlé MA การบริหารเฮโรอีนให้กับหนูอย่างเรื้อรังทำให้เกิดโปรตีนที่เกี่ยวข้องกับการตายของสมองและควบคุมการเรียนรู้เชิงพื้นที่และความจำ Neuropharmacology (2008) 54(4):640–52. doi:10.1016/j.neuropharm.2007.11.018
87. North A, Swant J, Salvatore MF, Gamble-George J, Prins P, Butler B, และคณะ การสัมผัสยาบ้าแบบเรื้อรังจะทำให้เกิดการขาดดุลความจำที่ยาวนานและล่าช้า ไซแนปส์ (2013) 67(5):245–57. doi:10.1002/syn.21635
88. Fole A, Martin M, Morales L, Del Olmo N ผลของการรักษาโคเคนเรื้อรังในช่วงวัยรุ่นในหนู Lewis และ Fischer-344: การรับรู้ตำแหน่งที่ผิดปกติและการเปลี่ยนแปลงของความแข็งแรงของ synaptic ในวัยผู้ใหญ่ Neurobiol Learn Mem (2015) 123: 179 86- ดอย: 10.1016 / j.nlm.2015.06.001
89. โจว M, Luo P, Lu Y, Li CJ, วัง DS, Lu Q, และคณะ ความไม่สมดุลของการแสดงออกของ HCN1 และ HCN2 ในพื้นที่ hippocampal CA1 บั่นทอนการเรียนรู้เชิงพื้นที่และความจำในหนูที่สัมผัสมอร์ฟีนเรื้อรัง Prog Neuropsychopharmacol Biol จิตเวชศาสตร์ (2015) 56: 207 14- ดอย: 10.1016 / j.pnpbp.2014.09.010
90. Kathirvelu B, Colombo PJ ผลของการแสดงออกของ CREB lentivirus-mediated ใน dorsolateral striatum: การเพิ่มประสิทธิภาพของหน่วยความจำและหลักฐานสำหรับการโต้ตอบและแข่งขันกับฮิบโปแคมปัส Hippocampus (2013) 23(11):1066–74. doi:10.1002/hipo.22188
91. Baudonnat M, Guillou JL, Husson M, Vandesquille M, Corio M, Decorte L, et al. ผลกระทบของการให้รางวัลยากระตุ้นต่อการเรียนรู้เชิงพื้นที่ แต่ไม่ได้ชี้นำคิว: ความหมายของโปรตีนไคเนส striatal โปรตีน A / cAMP ตอบสนององค์ประกอบทางเดินผูกพันโปรตีนผูกพัน Neurosci J (2011) 31:16517–28. doi:10.1523/JNEUROSCI.1787-11.2011
92. Packard MG, Wingard JC Amygdala และ "อารมณ์" การปรับของการใช้ญาติของระบบหน่วยความจำหลาย ๆ Neurobiol Learn Mem (2004) 82(3):243–52. doi:10.1016/j.nlm.2004.06.008
93. Elliott AE, Packard MG การให้ยา anxiogenic กับผู้ป่วยก่อนการรับลำเอียงเพื่อใช้หน่วยความจำของนิสัย Neurobiol Learn Mem (2008) 90(4):616–23. doi:10.1016/j.nlm.2008.06.012
94. Wingard JC, Packard MG amygdala และการปรับอารมณ์ของการแข่งขันระหว่างความรู้ความเข้าใจและความจำนิสัย Behav Brain Res (2008) 193(1):126–31. doi:10.1016/j.bbr.2008.05.002
95. Packard MG, Gabriele A. การฉีดยาเสพติด anxiogenic รอบนอกส่งผลกระทบต่อความรู้ความเข้าใจและความจำนิสัย: บทบาทของ amygdala basolateral Neuroscience (2009) 164(2):457–62. doi:10.1016/j.neuroscience.2009.07.054
96. Leong KC, Goodman J, Packard MG Buspirone บล็อกผลการเพิ่มประสิทธิภาพของยาเสพติด anxiogenic RS 79948-197 ในการรวมของหน่วยความจำนิสัย Behav Brain Res (2012) 234(2):299–302. doi:10.1016/j.bbr.2012.07.009
97. กู๊ดแมนเจ, เลหลงเคซี, แพคการ์ด MG การเพิ่มประสิทธิภาพ Glucocorticoid ของหน่วยความจำนิสัย dorsolateral ขึ้นอยู่กับ striatum ต้องใช้กิจกรรม noradrenergic พร้อมกัน Neuroscience (2015) 311: 1 8- ดอย: 10.1016 / j.neuroscience.2015.10.014
98. คิม JJ, Lee HJ, Han JS, Packard MG Amygdala มีความสำคัญสำหรับการปรับความเครียดที่เกิดจาก potentiation ระยะยาวและการเรียนรู้ hippocampal Neurosci J (2001) 21(14): 5222 8-
99. Schwabe L, Dalm S, Schächinger H, Oitzl MS ความเครียดเรื้อรังจะปรับเปลี่ยนการใช้กลยุทธ์การเรียนรู้เชิงพื้นที่และการตอบสนองต่อการกระตุ้นในหนูและมนุษย์ Neurobiol Learn Mem (2008) 90(3):495–503. doi:10.1016/j.nlm.2008.07.015
100. Leong KC, Packard MG การสัมผัสกับกลิ่นของนักล่ามีผลต่อการใช้ระบบหน่วยความจำหลายแบบแบบสัมพัทธ์: บทบาทของอะมิกดาลาที่เป็นเบส Neurobiol Learn Mem (2014) 109: 56 61- ดอย: 10.1016 / j.nlm.2013.11.015
101. Taylor SB, Anglin JM, Paode PR, Riggert AG, Olive MF, CD Conrad ความเครียดเรื้อรังอาจช่วยให้การรับสมัครงานของระบบประสาทที่เกี่ยวข้องกับนิสัยและติดยาเสพติดผ่านการปรับโครงสร้างของเส้นประสาทของ striatum Neuroscience (2014) 280: 231 42- ดอย: 10.1016 / j.neuroscience.2014.09.029
102. Leong KC, Goodman J, Packard MG โพสต์การฝึกอบรมการสัมผัสซ้ำเพื่อกลัวสิ่งเร้าที่มีเงื่อนไขช่วยเพิ่มการรวมหน่วยความจำและอคติต่อการใช้งานของการเรียนรู้การตอบสนองขึ้นอยู่กับ dorsolateral striatum Behav Brain Res (2015) 291: 195 200- ดอย: 10.1016 / j.bbr.2015.05.022
103. Goode TE, Leong KC, Goodman J, Maren S, Packard MG การเพิ่มประสิทธิภาพของหน่วยความจำขึ้นอยู่กับ striatum โดยความกลัวปรับอากาศถูกไกล่เกลี่ยโดยผู้รับเบต้า adrenergic ใน amygdala basolateral ชีววิทยาของความเครียด (ในการกด) ดอย: 10.1016 / j.ynstr.2016.02.004
104. Schwabe L, Oitzl MS, Philippsen C, Richter S, Bohringer A, Wippich W, et al. ความเครียดจะปรับการใช้กลยุทธ์การเรียนรู้เชิงพื้นที่กับการตอบสนองต่อสิ่งเร้าในมนุษย์ เรียนรู้ Mem (2007) 14(1–2):109–16. doi:10.1101/lm.435807
105. Schwabe L, Schächinger H, de Kloet ER, Oitzl MS Corticosteroids ทำงานเป็นสวิตช์ระหว่างระบบหน่วยความจำ J Cogn Neurosci (2010) 22(7):1362–72. doi:10.1162/jocn.2009.21278
106. Schwabe L, Tegenthoff M, Höffken O, Wolf OT กิจกรรม glucocorticoid ที่เกิดขึ้นพร้อมกันและ noradrenergic เปลี่ยนพฤติกรรมเครื่องมือจากการควบคุมเป้าหมายไปยังการควบคุมนิสัย Neurosci J (2010) 30(24):8190–6. doi:10.1523/JNEUROSCI.0734-10.2010
107. Schwabe L, Tegenthoff M, Höffken O, Wolf OT การปิดล้อมตัวรับ Mineralocorticoid ช่วยป้องกันการมอดูเลตที่เกิดจากความเครียดของระบบหน่วยความจำหลายระบบในสมองของมนุษย์ จิตเวช Biol (2013) 74(11):801–8. doi:10.1016/j.biopsych.2013.06.001
108. Schwabe L, Wolf OT ความเครียดแจ้งพฤติกรรมพฤติกรรมในมนุษย์ Neurosci J (2009) 29(22):7191–8. doi:10.1523/JNEUROSCI.0979-09.2009
109. Schwabe L, Wolf OT ความเครียดจากความกดดันทางสังคมที่ประเมินโดยสังคมหลังจากการเรียนรู้ผ่านสื่อมีส่วนช่วยสนับสนุนนิสัยมากกว่าการกระทำตามเป้าหมาย Psychoneuroendocrinology (2010) 35(7):977–86. doi:10.1016/j.psyneuen.2009.12.010
110. Guenzel FM, Wolf OT, Schwabe L. Glucocorticoids เพิ่มการสร้างความจำตอบสนองต่อสิ่งเร้าในมนุษย์ Psychoneuroendocrinology (2014) 45: 21-30 ดอย: 10.1016 / j.psyneuen.2014.02.015
111. ฮิกกินส์ RL, Marlatt GA ความกลัวของการประเมินระหว่างบุคคลในฐานะปัจจัยกำหนดปริมาณการดื่มแอลกอฮอล์ในนักดื่มสังคมชาย J Abnorm Psychol (1975) 84(6):644–51. doi:10.1037/0021-843X.84.6.644
112. Marlatt GA, Gordon JR ปัจจัยกำหนดของการกำเริบของโรค: ความหมายสำหรับการบำรุงรักษาการเปลี่ยนแปลงพฤติกรรม ใน: Davidson PO, Davidson SM, บรรณาธิการ ยาพฤติกรรม: การเปลี่ยนแปลงวิถีชีวิต. นิวยอร์ก: Brunne / Mazel (1980) พี 410-52
113. Newcomb MD, Bentler PM ผลกระทบของการใช้ยาวัยรุ่นและแรงสนับสนุนทางสังคมต่อปัญหาของคนหนุ่มสาว: การศึกษาระยะยาว. J Abnorm Psychol (1988) 97:64–75. doi:10.1037/0021-843X.97.1.64
114. วอลเลซก่อนคริสต์ศักราช ปัจจัยทางจิตวิทยาและสิ่งแวดล้อมของการกำเริบของโรคในผู้สูบบุหรี่โคเคนแคร็ก J Subst Abuse Treat (1989) 6(2):95–106. doi:10.1016/0740-5472(89)90047-0
115. Kaplan HB, Johnson RJ ความสัมพันธ์ระหว่างสถานการณ์รอบตัวการใช้ยาผิดกฎหมายเริ่มต้นและการเพิ่มการใช้ยา: ผลการควบคุมของเพศและประสบการณ์วัยรุ่นตอนต้น ใน: Glantz M, Pickens R, บรรณาธิการ ช่องโหว่ต่อยาเสพติด. วอชิงตันดีซี: สมาคมจิตวิทยาอเมริกัน (1992) พี 200-358
116. แฮร์ริสัน PA, Fulkerson JA, Beebe TJ การใช้สารหลายอย่างในผู้ที่ตกเป็นเหยื่อการล่วงละเมิดทางร่างกายและทางเพศของวัยรุ่น การทารุณกรรมเด็ก (1997) 21:529–39. doi:10.1016/S0145-2134(97)00013-6
117. Chilcoat HD, Breslau N. ความผิดปกติของความเครียดหลังถูกทารุณกรรมและความผิดปกติของยา: ทดสอบทางเดินสาเหตุ จิตเวชศาสตร์ Arch Gen (1998) 55(10):913–7. doi:10.1001/archpsyc.55.10.913
118. Piazza PV, Le Moal M. บทบาทของความเครียดในการบริหารจัดการยาด้วยตนเอง แนวโน้ม Pharmacol Sci (1998) 19(2):67–74. doi:10.1016/S0165-6147(97)01115-2
119. กู๊ดแมนเจ, ลีองเคซี, แพคการ์ด MG การปรับอารมณ์ของระบบหน่วยความจำที่หลากหลาย: ผลกระทบต่อระบบประสาทของความผิดปกติของความเครียดโพสต์บาดแผล รายได้ Neurosci (2012) 23(5–6):627–43. doi:10.1515/revneuro-2012-0049
120. Sinha R, Lacadie C, Skudlarski P, Fulbright RK, Rounsaville BJ, Kosten TR, et al. กิจกรรมของระบบประสาทที่เกี่ยวข้องกับความอยากโคเคนที่เกิดจากความเครียด: การศึกษาการถ่ายภาพด้วยคลื่นสนามแม่เหล็ก Psychopharmacology (2005) 183(2):171–80. doi:10.1007/s00213-005-0147-8
121 Avena NM, Rada P, Hoebel BG หลักฐานสำหรับการติดน้ำตาล: ผลกระทบด้านพฤติกรรมและระบบประสาทของการบริโภคน้ำตาลที่มากเกินไป Neurosci Biobehav Rev (2008) 32(1):20–39. doi:10.1016/j.neubiorev.2007.04.019
122. Gearhardt AN, Corbin WR, Brownell KD การติดอาหาร: การตรวจสอบเกณฑ์การวินิจฉัยสำหรับการพึ่งพา J Addict Med (2009) 3(1):1–7. doi:10.1097/ADM.0b013e318193c993
123. สมิ ธ DG, Robbins TW ปัจจัยทางด้านระบบประสาทของโรคอ้วนและการกินมาก: เหตุผลสำหรับการนำรูปแบบการติดอาหาร จิตเวช Biol (2013) 73(9):804–10. doi:10.1016/j.biopsych.2012.08.026
124. de Jong JW, Meijboom KE, Vanderschuren LJ, Adan RA การควบคุมอาหารที่รับประทานได้ในหนูน้อยมีความสัมพันธ์กับพฤติกรรมที่เป็นนิสัยและความอ่อนแอของการกำเริบของโรค: ความแตกต่างระหว่างบุคคล PLoS One (2013) 8(9): e74645 ดอย: 10.1371 / journal.pone.0074645
125. Furlong TM, Jayaweera HK, Balleine BW, Corbit LH การบริโภคอาหารที่น่าพึงพอใจจะช่วยเร่งการควบคุมพฤติกรรมและขึ้นอยู่กับการกระตุ้นของ dorsolateral striatum Neurosci J (2014) 34(14):5012–22. doi:10.1523/JNEUROSCI.3707-13.2014
126. Hargrave SL, Davidson TL, Zheng W, Kinzig KP อาหารตะวันตกทำให้เกิดการรั่วไหลของสิ่งกีดขวางในสมองและเลือดและปรับเปลี่ยนกลยุทธ์เชิงพื้นที่ในหนู Behav Neurosci (2016) 130(1):123–35. doi:10.1037/bne0000110
127. Smith KL, Hummer TA, Hulvershorn LA การเล่นวิดีโอเกมทางพยาธิวิทยาและความสัมพันธ์กับการใช้สารผิดปกติ ตัวแทนติดยาเสพติด (2015) 2(4):302–9. doi:10.1007/s40429-015-0075-6
128. Weinstein AM การติดเกมคอมพิวเตอร์และวิดีโอเกม - การเปรียบเทียบระหว่างผู้ใช้เกมและผู้ใช้ที่ไม่ใช่เกม การเสพสุราของ Am Am Drug (2010) 36(5):268–76. doi:10.3109/00952990.2010.491879
129. Kätsyri J, Hari R, Ravaja N, Nummenmaa L. ฝ่ายตรงข้ามมีความสำคัญ: fMRI ให้รางวัลสูงกว่าการตอบสนองต่อการชนะต่อมนุษย์กับคอมพิวเตอร์ฝ่ายตรงข้ามระหว่างการเล่นวิดีโอเกมแบบโต้ตอบ Cortex cereb (2013) 23(12):2829–39. doi:10.1093/cercor/bhs259
130. Kätsyri J, Hari R, Ravaja N, Nummenmaa L. เพียงแค่ดูเกมยังไม่พอ: fMRI ผู้ให้รางวัลตอบสนองต่อความสำเร็จและความล้มเหลวในวิดีโอเกมระหว่างการเล่นที่กระตือรือร้นและการเป็นตัวแทน Front Hum Neurosci (2013) 7: 278 ดอย: 10.3389 / fnhum.2013.00278
131. Erickson KI, Boot WR, Basak C, Neider MB, Prakash RS, Voss MW และอื่น ๆ ปริมาตร Striatal ทำนายระดับการได้มาซึ่งทักษะวิดีโอเกม Cortex cereb (2010) 20: 2522-30 ดอย: 10.1093 / cercor / bhp293
132. West GL, Drisdelle BL, Konishi K, Jackson J, Jolicoeur P, Bohbot VD การเล่นวิดีโอเกมแบบเป็นนิสัยสัมพันธ์กับกลยุทธ์การนำทางขึ้นอยู่กับนิวเคลียสของหางเสือ Proc R Soc B (2015) 282(1808) ดอย: 10.1098 / rspb.2014.2952
133. Liu S, Schad DJ, Kuschpel MS, Rapp MA, Heinz A. เพลงและวิดีโอเกมในช่วงพัก: อิทธิพลของการตัดสินใจที่เป็นนิสัยเมื่อเทียบกับเป้าหมาย กระดาษนำเสนอในการประชุมประจำปีครั้งที่ 45 ของสมาคมเพื่อประสาทวิทยาศาสตร์. Chicago, IL: สมาคมประสาทวิทยาศาสตร์ (2015)
134. de Fonseca FR, Carrera MRA, Navarro M, Koob GF, Weiss F. การกระตุ้นการทำงานของ corticotropin-factor ที่ปลดปล่อยในระบบ limbic ระหว่างการถอน cannabinoid วิทยาศาสตร์ (1997) 276(5321):2050–4. doi:10.1126/science.276.5321.2050
135. คอร์นีเลียสจูเนียร์ชุงทีมาร์ตินซีวู้ดดีเอสคลาร์กดีบี การถอนกัญชาเป็นเรื่องธรรมดาในหมู่วัยรุ่นที่กำลังมองหาการรักษาด้วยการพึ่งพากัญชาและโรคซึมเศร้าที่สำคัญและมีความสัมพันธ์กับการกำเริบของโรคอย่างรวดเร็ว ติดยาเสพติด Behav (2008) 33(11):1500–5. doi:10.1016/j.addbeh.2008.02.001
136. กรีน MC, Kelly JF ความชุกของการถอนกัญชาและอิทธิพลที่มีต่อการตอบสนองและผลลัพธ์ของการรักษาของวัยรุ่น: การสอบสวนในอนาคต 12 เดือน J Addict Med (2014) 8: 359-67 ดอย: 10.1097 / ADM.0000000000000064
137. แว็กเนอร์ FA, Anthony JC ตั้งแต่การใช้ยาครั้งแรกจนถึงการพึ่งพายา ระยะเวลาการพัฒนาของความเสี่ยงสำหรับการพึ่งพากัญชาโคเคนและแอลกอฮอล์ Neuropsychopharmacology (2002) 26:479–88. doi:10.1016/S0893-133X(01)00367-0
138. กู๊ดแมนเจ, แพคการ์ด MG อิทธิพลของ cannabinoids ต่อกระบวนการเรียนรู้และหน่วยความจำของ striatum หลัง Neurobiol Learn Mem (2015) 125: 1 14- ดอย: 10.1016 / j.nlm.2015.06.008
139. Rueda-Orozco PE, Soria-Gomez E, Montes-Rodriguez CJ, Martínez-Vargas M, Galicia O, Navarro L, et al. ฟังก์ชันที่เป็นไปได้ของ endocannabinoids ในการเลือกกลยุทธ์การนำทางโดยหนู Psychopharmacology (2008) 198(4):565–76. doi:10.1007/s00213-007-0911-z
140. สามีเจ, แพคการ์ด MG อุปกรณ์ต่อพ่วงและการฉีด striatum striatum ภายในและภายในของตัวรับ cannabinoid ตัวเอกชนะ 55,212-2 ทำให้การรวมหน่วยความจำของหน่วยความจำกระตุ้นตอบสนองลดลง Neuroscience (2014) 274: 128 37- ดอย: 10.1016 / j.neuroscience.2014.05.007
141. Nazzaro C, Greco B, Cerovic M, Baxter P, Rubino T, Trusel M, et al. การปรับแชนเนล SK ช่วยชีวิตปั้นพลาสติกและควบคุมนิสัยในการยอมรับ cannabinoid Nat Neurosci (2012) 15: 284 93- ดอย: 10.1038 / nn.3022
142. Ames SL, Grenard JL, Stacy AW, Xiao L, He Q, Wong SW, และคณะ การถ่ายภาพการทำงานของสมาคมกัญชาโดยปริยายในระหว่างการปฏิบัติงานในการทดสอบการเชื่อมโยงโดยนัย (IAT) Behav Brain Res (2013) 256: 494 502- ดอย: 10.1016 / j.bbr.2013.09.013
143. Redish AD, Jensen S, Johnson A. โครงสร้างแบบครบวงจรสำหรับการติดยาเสพติด: ช่องโหว่ในกระบวนการตัดสินใจ Behav Brain Sci (2008) 31(04):415–37. doi:10.1017/S0140525X0800472X
144. Brevers D, Bechara A, Cleeremans A, Noël X. งานการพนันในรัฐไอโอวา (IGT): ยี่สิบปีต่อมา - การพนันที่ผิดปกติและ IGT ด้านหน้า Psychol (2013) 4: 665 ดอย: 10.3389 / fpsyg.2013.00665
145. Koob GF, Le Moal M. กลไกทางระบบประสาทสำหรับกระบวนการสร้างแรงบันดาลใจของคู่ต่อสู้ในการเสพติด Philos Trans R Soc B Biol Sci (2008) 363(1507):3113–23. doi:10.1098/rstb.2008.0094
146. Hyman SE, Malenka RC, Nestler EJ กลไกประสาทของการเสพติด: บทบาทของการเรียนรู้ที่เกี่ยวข้องกับการให้รางวัลและความทรงจำ Annu Rev Neurosci (2006) 29: 565-98 ดอย: 10.1146 / annurev.neuro.29.051605.113009
147. กู๊ดแมนเจ, แพคการ์ดเอ็มระบบหน่วยความจำที่มีส่วนร่วมในระหว่างการครอบครองนั้นเป็นตัวกำหนดประสิทธิภาพของโปรโตคอลการสูญพันธุ์ที่แตกต่างกัน ด้านหน้า Behav Neurosci (2015) 9: 314 ดอย: 10.3389 / fnbeh.2015.00314
148. Palencia CA, Ragozzino ME การมีส่วนร่วมของตัวรับ NMDA ใน dorsolateral striatum เพื่อการเรียนรู้การตอบสนองแบบไร้ศูนย์กลาง Behav Neurosci (2005) 119(4):953–60. doi:10.1037/0735-7044.119.4.953
149. Rueda-Orozco PE, Montes-Rodriguez CJ, Soria-Gomez E, Méndez-Díaz M, Prospéro-García O การด้อยค่าของกิจกรรม endocannabinoids ใน dorsolateral striatum ล่าช้าการสูญพันธุ์ของพฤติกรรมในกระบวนการหน่วยความจำขั้นตอนในหนู Neuropharmacology (2008) 55(1):55–62. doi:10.1016/j.neuropharm.2008.04.013
คำสำคัญ: หน่วยความจำ, ติดยาเสพติด, ฮิปโปแคมปัส, striatum, amygdala, ความเครียด, ความวิตกกังวล
การอ้างอิง: กู๊ดแมนเจและแพคการ์ด MG (2016) ระบบหน่วยความจำและสมองติดยาเสพติด ด้านหน้า จิตเวช 7: 24 doi: 10.3389 / fpsyt.2016.00024
ได้รับ: 01 ธันวาคม 2015; ยอมรับแล้ว: 11 กุมภาพันธ์ 2016;
เผยแพร่เมื่อ: 25 February 2016
แก้ไขโดย:
วินเซนต์เดวิด, ศูนย์แห่งชาติ de la Recherche Scientifique (CNRS), ฝรั่งเศส
บทวิจารณ์โดย:
ฌาค มิโช, มหาวิทยาลัยบอร์โดซ์ 1, ฝรั่งเศส
Roberto Ciccocioppoมหาวิทยาลัยคาเมริโนประเทศอิตาลี
ลิขสิทธิ์: © 2016 Goodman and Packard นี่เป็นบทความแบบเปิดที่เผยแพร่ภายใต้เงื่อนไขของ ใบอนุญาตแสดงที่มาของครีเอทีฟคอมมอนส์ (CC BY). อนุญาตให้ใช้งานแจกจ่ายหรือทำซ้ำในฟอรัมอื่นโดยผู้แต่งหรือผู้ออกใบอนุญาตจะได้รับเครดิตและสิ่งพิมพ์ต้นฉบับในวารสารนี้ได้รับการอ้างอิงตามแนวทางปฏิบัติทางวิชาการที่เป็นที่ยอมรับ ไม่อนุญาตให้ใช้แจกจ่ายหรือทำซ้ำซึ่งไม่เป็นไปตามข้อกำหนดเหล่านี้
* สารบรรณ: Mark G. Packard, [ป้องกันอีเมล]