- 1Centro Interdisciplinario de Neurociencias de Valparaísoคณะวิทยาศาสตร์มหาวิทยาลัย Universidad de Valparaíso, Valparaíso, ชิลี
- 2Departamento de Neurociencia, Facultad de Medicina, มหาวิทยาลัยชิลี, ซันติอาโก, ชิลี
- 3Núcleo Milenio NUMIND ชีววิทยาของความผิดปกติทางประสาทจิต, มหาวิทยาลัยเดอบัลปาราอี, บัลปาราอี, ชิลี
- 4ภาควิชาชีววิทยาเซลล์และโมเลกุลคณะวิทยาศาสตร์ชีวภาพ, Pontificia Universidad Católica de Chile, Santiago, Chile
- 5ภาควิชาเภสัชศาสตร์และศูนย์สหวิทยาการประสาทคณะเคมี, Pontificia Universidad Católica de Chile, Santiago, ชิลี
ความแข็งแรงของพฤติกรรมที่มุ่งเน้นเป้าหมายถูกควบคุมโดยเซลล์ประสาทโดปามีนในสมองส่วนกลาง ความผิดปกติของวงจร dopaminergic เกิดจากการติดยาและความผิดปกติที่ครอบงำ พฤติกรรมบีบบังคับเป็นคุณสมบัติที่ความผิดปกติทั้งสองร่วมกันซึ่งเกี่ยวข้องกับสารสื่อประสาทโดปามีนที่มีความคิดริเริ่ม กิจกรรมของเซลล์ประสาทโดปามีนในสมองส่วนกลางได้รับการควบคุมโดยหลักจากการกระทำของโดพามีนในบ้านผ่าน D2 receptors (D2R) ซึ่งช่วยลดการยิงของเซลล์ประสาทรวมถึงการสังเคราะห์และปล่อยโดปามีน การส่งโดปามีนยังถูกควบคุมโดยระบบสารสื่อประสาทต่างกันเช่นระบบคัปปาโอปิออยและอื่น ๆ ความรู้ในปัจจุบันของเราส่วนใหญ่เกี่ยวกับระบบ kappa opioid และอิทธิพลที่มีต่อการส่งโดปามีนมาจากแบบจำลองสัตว์พรีคลินิกของโรคสมอง ในปี 1988 โดยใช้สมอง microdialysis ก็แสดงให้เห็นว่าการเปิดใช้งานเฉียบพลันของ Kappa Opioid Receptors (KOR) ลดระดับ synaptic ของโดปามีนใน striatum ผลยับยั้งนี้ของ KOR ตรงข้ามกับการอำนวยความสะดวกของยาเสพติดในการปล่อยโดปามีนที่นำไปสู่ข้อเสนอของการใช้งานของ KON agonists เป็นยารักษาทางเภสัชวิทยาสำหรับการบริโภคยาบังคับ น่าแปลกใจที่ 30 ปีต่อมาคู่อริของคู่อริถูกเสนอให้รักษายาเสพติดแทน มีอะไรเกิดขึ้นบ้างในช่วงหลายปีที่สร้างกระบวนทัศน์การเปลี่ยนแปลงครั้งใหญ่นี้? หลักฐานที่รวบรวมได้ชี้ให้เห็นว่าผลของ KOR ต่อระดับโดปามีนใน synaptic นั้นซับซ้อนขึ้นอยู่กับความถี่ของการเปิดใช้งาน KOR และเวลากับสิ่งเร้าอื่น ๆ ที่เข้ามาสู่โดปามีนเซลล์ประสาท ตรงกันข้ามกับผลกระทบเฉียบพลันการเปิดใช้งาน KOR เรื้อรังดูเหมือนว่าจะช่วยให้สารสื่อประสาทโดปามีนและโดพามีนเป็นสื่อกลาง การกระทำของฝ่ายตรงข้ามที่กระทำโดยการเปิดใช้งาน KOR แบบเฉียบพลันและแบบเรื้อรังนั้นมีความเกี่ยวข้องกับการหลีกเลี่ยงเบื้องต้นและผลการให้รางวัลล่าช้าในระหว่างการสัมผัสกับยาเสพติด พฤติกรรมบีบบังคับที่เกิดจากการเปิดใช้งานซ้ำของ D2R ยังมีศักยภาพโดยการเปิดใช้งานร่วมอย่างต่อเนื่องของ KOR ซึ่งสัมพันธ์กับระดับซินดิแคปที่ลดลงของโดปามีนและ D2R ที่ไวต่อแสง ดังนั้นการเปิดใช้งานขึ้นอยู่กับเวลาของ KOR ส่งผลกระทบโดยตรงต่อระดับโดพามีนที่มีผลต่อการปรับพฤติกรรมแรงจูงใจ การตรวจสอบนี้วิเคราะห์การมีส่วนร่วมของระบบ kappa opioid ที่สัมพันธ์กับโดปามิเนอร์จิคสัมพันธ์กับพฤติกรรมบีบบังคับ
บทนำ
ระบบ Dopaminergic ในพฤติกรรมบีบบังคับ
การบังคับเป็นสิ่งที่เป็นไปไม่ได้ของการหยุดตัวเองเพื่อดำเนินการตามปกติกับผลลัพธ์ที่ทราบแม้ว่าจะมีผลเสีย (Robbins และคณะ, 2012) พฤติกรรมบีบบังคับเป็นเครื่องหมายรับประกันคุณภาพของความผิดปกติที่ครอบงำ (OCD) และการติดยาเสพติดในบรรดาโรคทางจิตเวชอื่น ๆ การตรวจสอบพฤติกรรมเป็นเรื่องธรรมดามากในความผิดปกติของคลื่นความถี่ครอบงำซึ่งถูกกำหนดโดยการทำซ้ำอย่างสม่ำเสมอของกิจวัตรบางอย่างในทางที่ตายตัวหรือพิธีกรรม (วิลเลียมส์และอัล 2013) พฤติกรรมปกติที่หลากหลาย (เช่นการตรวจการทำความสะอาดการล้างมือและอื่น ๆ ) สามารถเปลี่ยนเป็นอาการบีบบังคับในผู้ป่วย OCD และโดยทั่วไปเกิดขึ้นในการตอบสนองต่อความคิดครอบงำและความทุกข์ที่ทำให้เกิดความวิตกกังวล ในทำนองเดียวกันการค้นหาและบริโภคยาเสพติดเป็นการละเมิดกลายเป็นสิ่งจำเป็นในการติดยาเช่นเดียวกับใน OCD ความวิตกกังวลมีบทบาทสำคัญในการกระตุ้นการบริโภคยาเสพติดในผู้เสพยาเสพติดที่มีประสบการณ์ ความบกพร่องเดียวกันในการให้รางวัลและการดำเนินการลงโทษมีการปฏิบัติในทั้งสองเงื่อนไข (ฟิกี et al., 2016) ซึ่งได้นำผู้เขียนบางคนเพื่อหารือเกี่ยวกับ OCD เป็นการติดพฤติกรรม (โฮลเดน 2001).
กลไกหนึ่งที่เป็นไปได้ที่จะนำไปสู่พฤติกรรมที่บีบบังคับคือกรอบภายในทฤษฎีการกระตุ้นให้เกิดอาการแพ้ซึ่งเป็นแรงจูงใจที่ขยายเพิ่มขึ้น (“ ต้องการ”) สำหรับยาที่พัฒนาขึ้นในระหว่างการติดยาเสพติดโดยไม่ต้องพัฒนาผลที่น่าพอใจBerridge และคณะ, 1989; Berridge และ Robinson, 2016) การทำให้แพ้อย่างต่อเนื่องของวงจรรางวัล / แรงจูงใจนั้นเกี่ยวข้องกับการเหนี่ยวนำให้เกิดอาการแพ้ที่เกี่ยวข้องกับการแสวงหายาเสพติด วงจรรางวัล / แรงจูงใจประกอบด้วยโดปามีนเซลล์ประสาทส่วนกลางของ substantia nigra (SN) และ ventral tegmental area (VTA) ซึ่งกำหนดเป้าหมายไปที่ชั้นหลังและ ventral ของ striatum ตามลำดับ เซลล์โดพามีนที่โครงการ ventral striatum หรือนิวเคลียส accumbens (NAc) มีความเกี่ยวข้องกับพฤติกรรมที่มุ่งเน้นเป้าหมายในขณะที่โดปามีนเซลล์ประสาทที่โครงการไปยัง striatum หลังมีความสัมพันธ์กับพฤติกรรมการซื้อ (Everitt และ Robbins, 2005; ปรีชาญาณ 2009; เยเกอร์ et al., 2015; Volkow et al., 2017).
การทำให้แพ้ของวงจรรางวัล / แรงจูงใจนั้นพบได้ในหนูเมื่อการเคลื่อนไหวของหัวรถจักรเพิ่มขึ้นอย่างค่อยเป็นค่อยไปเนื่องจากการให้ยาที่คงที่ซึ่งอาจทำให้ติดยาได้ (เพียร์ซและคาลิวาส 1997; Robinson และ Berridge, 2001) การทำให้ไวต่ออาการเคลื่อนไหวของหัวรถจักรเป็นปรากฏการณ์ที่สามารถทนทานได้เนื่องจากสามารถสังเกตได้หลังจากสัปดาห์, เดือนและแม้กระทั่งหนึ่งปีหลังจากการถอนยาRobinson และ Berridge, 1993) มันเป็นข้อเสนอแนะในช่วงต้นว่าการทำให้แพ้ของวงจรรางวัล / แรงจูงใจก่อให้เกิดการแสวงหาสิ่งเสพติด (Robinson และ Berridge, 1993) ดังนั้นการกระตุ้นให้หัวรถจักรอำนวยความสะดวกให้โคเคนการดูแลตนเองเพื่อค้นหาสถานะ (De Vries และคณะ, 2002) ยิ่งไปกว่านั้นหนูที่มีการเข้าถึงโคเคนด้วยตนเองอย่างต่อเนื่องแสดงให้เห็นว่าการตอบสนองของโคเคนในโคเคนนั้นสูงกว่าหนูที่มีการ จำกัด การเข้าถึง (Ferrario et al., 2005) นอกจากนี้ยังมีการสังเกตการเปลี่ยนแปลงทางเคมีประสาทที่เป็นรากฐานของการเคลื่อนไหวของสารกระตุ้นประสาทต่อยาจิตเวชSteketee และ Kalivas, 2011; Giuliano และคณะ, 2019) ข้อมูลเหล่านี้สนับสนุนการโต้ตอบที่เสนอล่วงหน้าระหว่างการกระตุ้นอาการแพ้จากหัวรถจักรและการแสวงหายาเสพติดซึ่งพบในมนุษย์Robinson และ Berridge, 1993; Vanderschuren และ Kalivas, 2000) กลไกการบริหารยาเสพติดซ้ำแล้วซ้ำอีกก่อให้เกิดความไวต่อวงจรโดปามีน mesolimbic ทำให้เพิ่มสารสื่อประสาทโดปามีน Psychostimulants เช่นโคเคนหรือแอมเฟตามีนที่กั้นพลาสมาเมมเบรนโดปามีนขนย้าย (DAT) ทำให้เกิดการเพิ่มขึ้นของโดปามีนในพื้นที่ synaptic ใน striatum และ NAc จึงกระตุ้นการเคลื่อนไหว (Steketee และ Kalivas, 2011) เช่นเดียวกับการติดยาเสพติดการทำให้แพ้ของโดปามีน / วงจรแรงจูงใจทำให้เกิดพฤติกรรมบีบบังคับที่เห็นใน OCD แท้จริงแล้วการเปิดใช้งานซ้ำของตัวรับ dopamine D2 (D2Rs) ก็เพียงพอที่จะชักนำให้เกิดอาการแพ้จากหัวรถจักรและพฤติกรรมการตรวจสอบทั้งในหนูและหนู (Szechtman et al., 1998; Szechtman et al., 1999; Sun et al., 2019) การบริหาร quinpirole ซ้ำเป็นตัวเอก D2R / D3R เป็นรูปแบบที่เป็นที่ยอมรับของ OCD เมื่อมันสรุปความถูกต้องของใบหน้าผ่านการเพิ่มขึ้นของการตรวจสอบเชิงบังคับและพฤติกรรม stereotyped ความถูกต้องเชิงทำนายเมื่อเห็นการลดลงของพฤติกรรมบีบบังคับ inhibitors (SRI) และสร้างความถูกต้องเป็นโครงสร้างสมองที่เกี่ยวข้องในแบบจำลองนี้มีการแบ่งปันกับผู้ที่อยู่ในพยาธิวิทยา (Stuchlik และคณะ, 2016; Szechtman et al., 2017). โดยสรุปการเปิดใช้งานการส่งโดปามีนซ้ำ ๆ ไม่ว่าจะโดย pre-synaptic (การปลดปล่อยโดปามีน) หรือการโพสต์ซินแคปิก (การเปิดใช้งาน D2R) กลไกนำไปสู่การก่อให้เกิดอาการแพ้
ระบบ kappa opioid เป็นหนึ่งในระบบที่สำคัญที่สุดในการควบคุมการส่งโดปามีนในวงจรรางวัล / แรงจูงใจ หลักฐานแสดงให้เห็นว่าการส่งคัปปา - opioid ตรงข้ามกับผลกระทบของโดปามีน; การเปิดใช้งานอย่างฉับพลันของตัวรับ Kappa opioid (KORs) ต่อต้านกิจกรรมของหัวรถจักรที่เกิดจาก psychostimulants (สีเทาและคณะ, 1999) ในทางกลับกันการเปิดใช้งาน KOR ซ้ำรักษาและปรับปรุงการค้นหายาเสพติดซึ่งต้องกระทำและเป็นนิสัย (Koob, 2013) การบริโภคยาเสพติดในทางที่ผิดก่อให้เกิดการเพิ่มการส่งผ่านของแคปปา opioid แบบ homeostatic อาจช่วยให้เกิดสภาวะทางอารมณ์เชิงลบของ dysphoria (Koob, 2013) ก่อให้เกิดการใช้ยาเสพติดซึ่งบีบบังคับChavkin และ Koob, 2016) ในความเป็นจริงการปิดล้อมของ KOR ป้องกันความเครียด - แต่ไม่ใช่ยากระตุ้นการคืนสถานะของนิโคติน (Jackson et al., 2013) โคเคน (Beardsley et al., 2005) และเอทานอล (สเปอร์ลิง และคณะ, 2010) สอดคล้องกับการค้นพบนี้การปิดล้อม KOR ย้อนกลับการเปลี่ยนแปลงโดปามีนในแถบ dorsolateral striatum ของหนูแอมเฟตามีนที่ได้รับไวอาโซคาร์ และคณะ 2019). ดังนั้นระบบ KOR ดูเหมือนว่าจะเพิ่มการเสริมแรงเชิงลบเพิ่มมูลค่ายาเสพติด ใน OCD การเสริมแรงเชิงลบจะถูกกระตุ้นโดยความหลงไหลซึ่งเสริมความแข็งแกร่งให้กับการบังคับเพื่อหลีกเลี่ยงความหลงใหลนั้น แม้ว่าจะไม่ได้รับการทดสอบโดยตรง แต่การเสริมแรงเชิงลบอาจมีบทบาทในการทำให้แพ้ quinpirole อันที่จริง D2R มีส่วนร่วมในการสร้างการเสริมแรงเชิงลบ ตัวอย่างเช่นการหลีกเลี่ยงสถานที่ไปยังพื้นที่ที่จับคู่มอร์ฟีนถูกถอนไม่ได้พัฒนาขึ้นในหนูที่ขาดไอโซฟอร์มที่มีความยาวของ D2R (Smith และคณะ, 2002) และการรักษาด้วย quinpirole ซ้ำ ๆ ในช่วงที่งดเว้นจะเรียกคืนโคเคนและเฮโรอีนในกระบวนทัศน์การบริหารอัตโนมัติผลที่เกี่ยวข้องกับการเคลื่อนไหวที่ไวต่อการ quinpirole (De Vries และคณะ, 2002) เสนอแนะกลไกที่ใช้ร่วมกันระหว่างการกระตุ้นอาการจิตประสาทและการกระตุ้นประสาทตาขาวที่เกิดจาก quinpirole ยิ่งไปกว่านั้นการแนะนำของกรงบ้าน แต่ไม่ใช่กรงที่แปลกใหม่ในเวทีเปิดโล่งลดความรู้สึกไวของหัวรถจักรและพฤติกรรมการตรวจสอบที่ต้องกระทำSzechtman et al., 2001) แสดงว่าความปลอดภัย / ตัวชี้นำที่คุ้นเคยอาจแข่งขันกับตัวชี้นำด้านสิ่งแวดล้อมเชิงลบที่สนับสนุนการแพ้ ในทำนองเดียวกันกับการกระตุ้นให้เกิดอาการกระตุ้นจิตประสาทการเปิดใช้งานซ้ำ ๆ ของ KOR ช่วยให้เกิดอาการแพ้ locomotor (เอสโกบาร์ และคณะ, 2017) และพฤติกรรมการตรวจสอบซึ่งบีบบังคับ (Perreault et al., 2007) เกิดจากการบริหาร quinpirole ซ้ำ ๆ ไม่ว่าจะเป็นผลโพเทนทิโอมิเตอร์นี้เป็นผลมาจากการเสริมแรงเชิงลบที่ได้รับการปรับปรุงให้ดีขึ้น
การวิเคราะห์อย่างละเอียดที่ดำเนินการเมื่อเร็ว ๆ นี้แสดงให้เห็นว่าผลของระบบ kappa-opioid ต่อการส่ง dopaminergic นั้นซับซ้อน: ขึ้นอยู่กับ dopamine pathway ที่เกี่ยวข้อง (Margolis และคณะ, 2006; Margolis และคณะ, 2008) และระยะเวลาระหว่างการเปิดใช้งานตัวรับ KOR และการเปิดใช้งานตัวรับโดปามีน (Chartoff และคณะ, 2016) ด้วยความซับซ้อนนี้จึงได้มีการพูดคุยกันอย่างกว้างขวางเกี่ยวกับการใช้ลิแกนด์ KOR ในการรักษา. มันได้รับการเสนอว่าตัวเอก KOR อาจมีประโยชน์ทางคลินิกในระหว่างขั้นตอนการใช้ยาลดทอนยากระตุ้น hyperdopaminergia (Shippenberg และคณะ, 2007) ในทางกลับกัน KOR อาจมีประโยชน์ในการรักษาอาการถอนที่เกิดจากการเพิ่มขึ้นของการแสดงออกของ dynorphin หลังจากการบริโภคยาซ้ำ ๆ (วีและคูบ 2010) ดังนั้นจึงมีการเสนอว่า KOR ตัวเอกบางส่วน (เบกวิน และคณะ 2012) อาจเป็นตัวเลือกในการรักษาเพื่อรักษาทั้งการบริโภคยาเสพติดและอาการถอนในผู้ที่ติดยา (Chartoff และคณะ, 2016; Callaghan et al., 2018) ในการตรวจสอบนี้เราวิเคราะห์การปรับขึ้นอยู่กับบริบท / เวลาของ dopaminergic correlates ของ sensitization พฤติกรรมและ compulsivity
Crosstalk ทางกายวิภาคและหน้าที่ระหว่างคัปปา Opioid และระบบ Dopaminergic ในภูมิภาค Striatal และ Midbrain
ภูมิภาค Striatal
KORs เป็นตัวรับโปรตีนคู่ / Gi แสดงอย่างสูงในระบบโดปามีน midbrain (Mansour et al., 1996) ตัวรับเหล่านี้อยู่ในตระกูล opioid receptors ที่ประกอบด้วย mu (MOR), delta (DOR) และ kappa (KORs) agonists ภายนอกสำหรับผู้รับเหล่านี้คือ endorphins, enkephalin และ dynorphin ตามลำดับ ใน striatum, dynorphin ถูกสังเคราะห์โดย dopamine D1receptor (D1R) - ควบคุมเซลล์ประสาทขนาดกลาง (MSNs) ที่มีแอกซอนที่กระตุ้นการทำงานของ KORs จากนิวเคลียสเดียวกัน (Mansour et al., 1995) ภาพกล้องจุลทรรศน์อิเล็กตรอนของหนู NAc แสดงให้เห็นว่า KORs จะพบส่วนใหญ่ในโครงสร้าง presynaptic ที่มี DAT ในขณะที่สัดส่วนเล็กน้อยของ KORs จำกัด ใน dendrites ในภาคผนวกกับ DAT (Svingos et al., 2001; คีเวลล์ และคณะ 2014) การศึกษาเกี่ยวกับอิมมูโนฟลูออเรสเซนต์ที่มีลักษณะเฉพาะการเตรียม presynaptic-synaptosomal จาก NAc แสดงให้เห็นว่า KORs และ D2Rs อยู่ร่วมกันเป็นพิเศษใน synaptosomes ที่มีเอนไซม์การสังเคราะห์โดพามีนไทโรซีนไฮดรอกซิส (TH) (เอสโกบาร์ และคณะ, 2017) ยิ่งไปกว่านั้น KORs มีมากมายในเซลล์ของ NAc และ striatum และรวมตัวกับ D2Rs ในเซลล์ย่อย (เอสโกบาร์ และคณะ, 2017) ด้วยข้อมูลเชิงลึกทางพันธุกรรมและโมเลกุลเราแนะนำว่า 20% ของ KOR รวมใน striatum นั้นสังเกตได้จากขั้ว DA (Van't Veer et al., 2013) ยิ่งไปกว่านั้น Tejeda และคณะ (2017) แสดงให้เห็นว่าทั้ง D1R และ D2R MSNs แสดง KOR ด้วยความต้องการที่สูงขึ้นสำหรับ D1R ที่มี MSNs (Tejeda et al., 2017 โดย) ข้อมูลเชิงกายวิภาคนี้บ่งชี้ว่า KORs มีอยู่ก่อนและหลังการเปลี่ยนแปลงการควบคุมสารสื่อประสาทโดปามีนในวงจรรางวัล / แรงจูงใจ
หลายวิธีการทดลองแสดงให้เห็นว่าการเปิดใช้งานของ KORs ยับยั้งโดปามีน releasอี การกระตุ้นแบบเฉียบพลันของ KORs โดยการฉีดอย่างเป็นระบบหรือการฉีดแบบ agonists ในท้องถิ่นลดระดับโดปามีนในเซลล์นอกเซลล์ใน NAc (Di Chiara และ Imperato, 1988; Spanagel et al., 1992; Fuentealba และคณะ, 2006) และหลัง striatum (เกร์เคอ และคณะ 2008) สนับสนุนการยับยั้งการกระทำของ KORs กับสารสื่อประสาทโดปามีนซึ่งเป็นยาโดยตรงของคู่อริ KOR และ nor-binaltorphimine (หรือ BNI)Broadbear และคณะ, 1994) เพิ่มระดับฐานของโดปามีนใน NAc (Spanagel et al., 1992) และการหลั่งโดปามีนใน dorsal striatum (อาโซคาร์ และคณะ 2019) หลักฐานขั้นสุดท้ายของการยับยั้งยาชูกำลังของ KOR ในโดปามีนแสดงให้เห็นในหนูที่น่าพิศวง KOR ซึ่งแสดงระดับโดปามีนที่เพิ่มขึ้นในเซลล์ extracellular ใน striatum และ NAc (Chefer et al., 2005). กลไกที่รับผิดชอบในการยับยั้ง KOR ของการปลดปล่อยโดปามีนไม่ได้รับการอธิบายอย่างสมบูรณ์ อย่างไรก็ตามเป็นที่ทราบกันดีว่าการกระตุ้น KORs นำไปสู่การเพิ่มขึ้นของ K + และการลดลงของ Ca2 + conductances จึงทำให้เกิด hyperpolarization ของเซลล์และการปิดกั้นการปลดปล่อยสารสื่อประสาท vesicular (Bruchas และ Chavkin, 2010; Margolis และ Karkhanis, 2019).
นอกจากนี้, ในหลอดทดลอง และ ในร่างกาย ข้อมูลการทำงานแสดงให้เห็นว่า KORs ปรับระดับโดปามีนนอกเซลล์โดยปรับกิจกรรมของ DAT. ตัวอย่างเช่นการเปิดใช้งาน KORs ในเซลล์ EM4 ที่ร่วมแสดง KORs และ DAT นำไปสู่การเพิ่มขึ้นของการดูดซับโดปามีนที่วัดโดย voltammetry (คีเวลล์ และคณะ 2014) ex vivo การวิเคราะห์ด้วยการใช้โวลแทมเมทรีในเนื้อเยื่อที่ไม่ได้แยกแยะแสดงให้เห็นว่าการฉีด KOR agonist U-69593 เป็นระบบที่เพิ่มการดูดโดปามีนใน NAcThompson และคณะ, 2000) บทความล่าสุดที่คล้ายกันแสดงให้เห็นว่า nor-BNI บล็อกการเพิ่มขึ้นของการดูดซับโดปามีนใน striatum หน้าท้องและหลังซึ่งเกิดจากการฉีดแบบเฉียบพลันของ MP1104 ซึ่งเป็นตัวรับแบบผสม Kappa / Delta opioid agonist (อาทิการิ และคณะ 2019) อย่างไรก็ตามผลของการเปิดใช้งาน KOR ต่อการดูดซับโดพามีนยังไม่ได้รับการอธิบายอย่างสมบูรณ์ การบริหารระบบของ KOR บางส่วน agonist nalmefene ลดปริมาณโดปามีนที่ได้รับการดูดซึมอย่างรวดเร็วในเชิงปริมาณวัดโดยการสแกนแบบเร็วแบบไซคลิกโวลแทมเมทรี (FSCV) (Rose et al., 2016) การใช้ microdialysis ฟลักซ์แบบไม่มีตาข่ายในหนูตัวผู้ผู้ใหญ่การปิดกั้น KOR นั้นมาพร้อมกับการเพิ่มขึ้นของส่วนการสกัด (Ed) ซึ่งเป็นการวัดทางอ้อมของการดูดซึมโดปามีน (Chefer et al., 2006; อาโซคาร์ และคณะ 2019) แนะนำว่าการกระตุ้นโทนิคของ KOR จะเป็นการควบคุมการยับยั้งกิจกรรม DAT (การดูดโดปามีน) ผลลัพธ์เหล่านี้เน้นถึงบทบาทที่ซับซ้อนของกิจกรรม KOR ภายนอกที่มีต่อการดูดซับโดปามีนเพื่อควบคุมระดับโดปามีนนอกเซลล์ วิธีการแก้ปัญหาชั่วคราวที่สูงขึ้นเช่น FSCV ล้มเหลวในการแสดงผลของ KOR ต่อการดูดซึมของโดพามีน (เอบเนอร์ และคณะ, 2010; เอริช และคณะ 2015; Hoffman และคณะ 2016) แนะนำว่า KOR ที่เสริมกิจกรรม DAT ในภูมิภาคที่ต้องการการฟักตัวของระยะเวลาการฟักตัว, การเสริม KOR-mediated ของกิจกรรม DAT นั้นสามารถอธิบายได้โดยการเพิ่มจำนวนของ DAT ในเยื่อหุ้มเซลล์ที่เกิดจากการกระตุ้นด้วย KOR ดังที่รายงานใน synaptosomes และเซลล์ เส้น (Kivell และคณะ, 2014).
ภูมิภาค Midbrain
การตรวจวิเคราะห์อัตชีวประวัติอัตโนมัติในหนูกลางแสดงให้เห็นถึงการผูกพันอย่างมีนัยสำคัญสำหรับ KORs บนแกน rostrocaudal ของ SN และ VTA (ผู้เชี่ยวชาญและคณะ 1993) ในอีกทางหนึ่งข้อมูลกล้องจุลทรรศน์อิเล็กตรอนแสดงให้เห็นว่าขั้วที่มี dynorphin synapse โดยตรงใน TH บวก dendrites ใน SN และ VTA (Sesack และ Pickel, 1992) แนะนำว่า KORs จำกัด วงในช่อง somatodendritic ของเซลล์ประสาทโดพามีน MSN ที่มี Striatal D1R นั้นเป็นหนึ่งใน dynorphin ที่ป้อนเข้าสู่โดปามีนเซลล์สมองส่วนกลาง สิ่งที่น่าสนใจคือการอุดตันของ KORs ไม่ได้แก้ไขผลการยับยั้งของ D1R-MSNs ให้เป็นเซลล์ประสาทโดปามีน VTA ของ VTA ซึ่งบ่งชี้ว่าการยับยั้งนี้เป็นสื่อกลางโดย GABA (Edwards และคณะ, 2017) KORs ปรับการตอบสนอง somatodendritic ของเซลล์ประสาทโดปามีน midbrain การศึกษาทางไฟฟ้าวิทยาแสดงให้เห็นว่าการกระตุ้น KORs ใน VTA hyperpolarizes และลดอัตราการเผา spontaneus ของเซลล์ประสาทโดปามีน (Margolis และคณะ, 2003) ดังนั้นการแช่ของ KOR agonists ลด somatodendritic dopamine efflux (Smith และคณะ, 1992; Dalman และ O'Malley, 1999) อย่างไรก็ตามการยับยั้งผลของ KORs ต่อเซลล์ประสาทโดปามีนดูเหมือนว่าจะขึ้นอยู่กับวงจร การแช่ของตัวเอก kappa-opioid ใน VTA ลดการปลดปล่อยโดปามีนในเยื่อหุ้มสมอง prefrontal cortex (mPFC) (Margolis และคณะ, 2006) แต่ไม่ใช่ใน NAc (Devine et al., 1993; Margolis และคณะ, 2006) ยิ่งไปกว่านั้น Margolis และคณะ (2006) พบว่า KORs ยับยั้งเซลล์ประสาทโดพามีน VTA ที่ฉายไปที่ mPFC และ amygdala ที่เป็น basolateral แต่ไม่ใช่พวกที่ไปยัง NAc ในปีเดียวกันนั้น ฟอร์ดและคณะ (2006) แสดงให้เห็นว่าการประยุกต์ใช้อาบน้ำของ KOR agonists ในหนู VTA ชิ้นเหนี่ยวนำให้เกิดกระแสออกไปข้างนอกที่สูงขึ้นในเซลล์ประสาทโดปามีนที่โครงการเพื่อ NAc เมื่อเทียบกับที่โครงการเพื่อ amygdala basolateral แสดงให้เห็นว่า KORs ออกแรงยับยั้งมากขึ้นของเซลล์โดพามีน เพื่อ amygdala นอกจากนี้การกระตุ้น KOR ยังลดความกว้างของ excitatory (Margolis และคณะ, 2005) และยับยั้ง (ฟอร์ดและอัล 2007) กระแส postsynaptic เป็นเซลล์ประสาทโดปามีนสมองส่วนกลาง ความแตกต่างระหว่างสปีชีส์กับ proerection efferents ที่ซับซ้อนของ VTA ถึง mPFC และ NAc (Van Bockstaele และ Pickel, 1995; Carr และ Sesack, 2000) ทำให้มันยากที่จะพิสูจน์ว่า KORs ยับยั้งการเลือกโดพามีนเส้นประสาทบางส่วนใน VTA หรือไม่ อย่างไรก็ตามข้อมูลสรุปที่นี่แสดงให้เห็นว่า KORs อยู่ในโสมและขั้วของโดปามีนเซลล์ประสาทเช่นเดียวกับในอินพุตที่ควบคุมพวกเขาจึงอยู่ในตำแหน่งที่ควบคุมกิจกรรม synaptic ของเซลล์ประสาทโดปามีนในสมอง
บทบาทของ KORs ในการควบคุม Dopamine Neurotransmission ใน Psychostimulants-Induced Sensitization และพฤติกรรมบีบบังคับ
การติดยาเสพติดเป็นกระบวนการที่เกี่ยวข้องกับการแสวงหายาเสพติดในระยะเริ่มต้นซึ่งเกี่ยวข้องกับผลในเชิงบวก ในทางกลับกัน compulsivity เป็นลักษณะบุคลิกภาพที่สังเกตได้ในผู้ติดยาเสพติด มีการเสนอ neuroadaptations ในเส้นทางโดปามีนเพื่อบัญชีสำหรับการแสวงหายาเสพติดและการบริโภคตามการสัมผัสซ้ำ ๆ กับยาเสพติด (Everitt และ Robbins, 2005; Koob และ Volkow, 2016) หนึ่งในสมมติฐานที่นำเสนอในการผลักดันการบริโภคยาเสพติดซึ่งต้องกระทำคือการทำให้ไวต่อผลกระทบเชิงลบ (Koob, 2013) การควบคุมการยับยั้งของระบบ Kappa opioid ในการปล่อยโดปามีนอาจส่งผลให้เกิดคุณสมบัติเชิงลบของยาเสพติด อย่างไรก็ตามผลที่ตามมาของการเปิดใช้งาน KOR ต่อสารสื่อประสาทโดปามีนและการค้นหายาเสพติดซึ่งดูเหมือนจะซับซ้อนและขัดแย้งกันอย่างเห็นได้ชัด อันที่จริงการปล่อยโดปามีนที่เกิดจากแอมเฟตามีนและโคเคนนั้นถูกลดทอนโดยการบริหารของนักประพันธ์ KOR ด้วยกัน (Heidbreder และ Shippenberg, 1994; เมซงเนิฟ et al., 1994; Thompson และคณะ, 2000) และลดการจัดการโคเคนด้วยตนเอง (เนกัส และคณะ 1997) ยิ่งไปกว่านั้น KORs ยังแสดงความคิดเห็นต่อการยับยั้งการปล่อยโดปามีนในทางเดิน mesolimbic เพื่อตอบสนองต่อการกระตุ้นการทำงานของ D1R หลังการรักษาอย่างต่อเนื่องเช่นเดียวกับการสัมผัสกับยาจิตCole et al., 1995; Nestler, 2001) ความขัดแย้งการเปิดใช้งาน KORs ยังสามารถอำนวยความสะดวกในการปลดปล่อยโดปามีนในเส้นทางของรางวัล / แรงบันดาลใจ (Fuentealba และคณะ, 2006; Fuentealba และคณะ, 2007) และการบริโภค psychostimulants (Wee et al., 2009). Fuentealba และคณะ (2007) แสดงให้เห็นว่าหลังจากสี่วันในการบริหาร U69593 นักอะโกนิสต์ KOR ได้เพิ่มการปลดปล่อยโดปามีนจากแอมเฟตามีนใน NAc เมื่อเร็ว ๆ นี้ก็แสดงให้เห็นว่าการปิดกั้น KORs ย้อนกลับการเปลี่ยนแปลงในการปล่อยโดปามีนและการดูดซึมใน dorsal striatum ที่เกิดขึ้นในช่วงอาการแพ้หัวรถจักรที่เกิดจากแอมเฟตามีน (อาโซคาร์ และคณะ 2019) ข้อมูลเหล่านี้ชี้ให้เห็นว่าการเปิดใช้งาน KORs อาจช่วยเสริมคุณสมบัติในเชิงบวกของยาเสพติดChartoff และคณะ, 2016).
นอกจากนี้การเปิดใช้งาน KORs ก็ดูเหมือนจะมีส่วนช่วยในการแสวงหายาเสพติด การปิดล้อม KORs ช่วยลดโคเคน (Wee et al., 2009) เฮโรอีนSchlosburg และคณะ, 2013) และยาบ้า (วิทฟิลด์ และคณะ, 2015) การบริโภคในหนูที่ไม่ จำกัด การเข้าถึงยา (Wee et al, 2009) ผลกระทบนี้ยังเป็นหลักฐานในการค้นหายาที่ทำให้เกิดความเครียด ตัวอย่างเช่นหนูที่น่าพิศวง KOR ไม่ได้แสดงการตั้งค่าสถานที่โคเคนหลังจากความเครียดว่ายน้ำบังคับ (แมคลาฟลิน และคณะ, 2006ก) การปิดกั้นของ KORs ลดทอนการตั้งค่าสถานที่นิโคตินที่เกิดจากการสัมผัสความเครียดว่ายน้ำบังคับ (Smith และคณะ, 2012) ที่น่าสนใจคือการปิดกั้น KOR ลดทอนโคเคนและนิโคตินที่เกิดจากความเครียด แต่ไม่ส่งผลกระทบต่อการแสวงหาการกระตุ้นด้วยการใช้ยาBeardsley et al., 2005; Jackson et al., 2013) KOR เอฟเฟกต์ที่เกิดจากความเครียดนี้ดูเหมือนจะเป็นสื่อกลางโดยวงจรรางวัล / แรงจูงใจ (Shippenberg และคณะ, 2007; วีและคูบ 2010) ในการศึกษาที่สง่างามที่ดำเนินการโดยดร. คาเออร์และกลุ่มของเธอก็แสดงให้เห็นว่าการปิดกั้น KORs ใน VTA ไม่ว่าก่อนหน้านี้หรือหลังความเครียดเฉียบพลันยับยั้งยับยั้งการแสวงหาโคเคนคืนสถานะผลกระทบที่เกี่ยวข้องกับการช่วยเหลือระยะยาว - ศักยภาพของการยับยั้ง synapses ในเซลล์ประสาทโดพามีน (Graziane et al., 2013 โดย; โพลเตอร์ และคณะ, 2014).
การอำนวยความสะดวกของการบริโภค psychostimulants กระทำโดย KORs ดูเหมือนจะขึ้นอยู่กับเวลาหน้าต่างเกี่ยวกับการสัมผัสยาเสพติด การบริหารของ KOR agonist U50488 1 ชั่วโมงก่อนการสัมผัสโคเคน potentiates ทั้งการตั้งค่าสถานที่โคเคนและการปลดปล่อยโดปามีนที่สัมพันธ์กันปรากฏโดยโคเคนใน NAc ในขณะที่ผลตรงข้ามจะได้รับเมื่อ 15 นาทีก่อน (แมคลาฟลิน และคณะ, 2006ก; เอริช และคณะ 2014) ใช้การกระตุ้นสมองด้วยตนเอง Chartoff และคณะ (2016) ตั้งข้อสังเกตว่า KOR ตัวเอก Salvinorin A มีผลเริ่มต้นและผลตอบแทนล่าช้าพร้อมกับการลดลงและเพิ่มขึ้นในการกระตุ้นโดปามีนกระตุ้นใน NAc ตามลำดับ ข้อมูลทั้งหมดเหล่านี้บ่งบอกถึงผลกระทบที่ขึ้นอยู่กับเวลาของการเปิดใช้งาน KOR ต่อคุณสมบัติที่ได้รับรางวัลของโคเคนและชี้ไปที่การเปิดใช้งาน KOR ที่เป็นสื่อกลางในฐานะผู้เล่นหลักในการพัฒนาการแสวงหายาเสพติด
การทำให้เกิดอาการแพ้จากการเคลื่อนไหวของหัวรถจักร Quinpirole และพฤติกรรมบีบบังคับ
ข้อเท็จจริงที่ว่าระบบโดปามีนมีส่วนร่วมในการสร้างความรู้สึกไวและการบังคับใช้มีความเข้มแข็งโดยพฤติกรรมที่สังเกตได้ในสัตว์ฟันแทะที่ได้รับการรักษาด้วย D2R agonist, quinpirole สั้น ๆ D2Rs เป็น Gi คู่ตัวรับแสดงอย่างกว้างขวางในวงจรรางวัล / แรงจูงใจ พวกเขาจะแสดง somatodendritically และบนซอนซอนของโดปามีนเซลล์ประสาท (Sesack และคณะ, 1994) และการเปิดใช้งานจะลดระดับสารกระตุ้นโดปามีน (Imperato และ Di Chiara, 1988) ใน striatum, D2Rs ยังตั้งอยู่ในตำแหน่งที่มีการเปลี่ยนแปลงในเซลล์ประสาทสปินปานกลาง (Sesack และคณะ, 1994) และการเปิดใช้งานจะยับยั้งเส้นทางที่อ้อมที่อนุญาตให้กิจกรรมของหัวรถจักร
Dr. Henry Szechtman เริ่มศึกษาผลของ quinpirole ที่มีต่อพฤติกรรมของหนูในช่วงปลายทศวรรษ 1980 ผลการวิจัยเบื้องต้นแสดงให้เห็นว่าการให้ quinpirole แบบเฉียบพลันมีผลต่อการออกฤทธิ์ของขมิ้นอ้อย ในปริมาณที่ต่ำ (0.03 มก. / กก.) จะลดการทำงานของขมิ้นอ้อยในขณะที่ปริมาณที่สูงขึ้น (> 0.5 มก. / กก.) จะเพิ่มขึ้น (Eilam และ Szechtman, 1989) เอฟเฟกต์เหล่านี้เกี่ยวข้องกับการเปิดใช้งาน D2Rs presynaptic ที่มีความสัมพันธ์สูงและโพสต์ Naptynaptic D2Rs ที่มีความสัมพันธ์ต่ำตามลำดับ (Usiello และคณะ, 2000) โดยไม่คาดคิดการจัดการ quinpirole ซ้ำทุก ๆ วันทำให้เกิดการเคลื่อนไหวที่เพิ่มขึ้นอย่างต่อเนื่องและค่อยเป็นค่อยไปซึ่งคล้ายกับการกระตุ้นการเคลื่อนไหวของหัวรถจักรที่เกิดจาก psychostimulants (Szechtman et al., 1993; Szechtman et al., 1994) ผลกระทบที่ทำให้เกิดอาการแพ้ในหัวรถจักรแสดงให้เห็นว่าขึ้นอยู่กับ D2Rs เนื่องจากหนูบกพร่องสำหรับตัวรับสัญญาณนี้ไม่ได้พัฒนาอาการแพ้หัวรถจักรไปเป็น quinpirole (เอสโกบาร์ และคณะ, 2015).
ในตอนต้นของทศวรรษที่ 90 Szechtman และ Eilam รายงานว่าพร้อมกับความไวของหัวจักรทำให้หนูมีพฤติกรรมที่ตายตัวซึ่งได้รับการเสริมด้วยการบริหาร quinpirole แต่ละครั้ง (Eilam และ Szechtman, 1989; Szechtman et al., 1993) วันนี้การบริหารซ้ำของ quinpirole เป็นรูปแบบที่ผ่านการตรวจสอบสำหรับ OCD (Szechtman et al., 1999; Szechtman et al., 2001; Eilam และ Szechtman, 2005; Stuchlik และคณะ, 2016; Szechtman et al., 2017) จากการสังเกตว่าพฤติกรรมของหนูมีโครงสร้างมากขึ้นและไม่ยืดหยุ่นมากขึ้นทำให้ระลึกถึงลักษณะของพฤติกรรมพิธีกรรมของพฤติกรรมการตรวจสอบแบบบังคับ (Szechtman et al., 1998; Szechtman et al., 2017) การศึกษาล่าสุดแสดงให้เห็นว่า quinpirole ซ้ำยังก่อให้เกิดพฤติกรรมบีบบังคับในหนูเช่นการตรวจสอบบังคับ (Sun et al., 2019) ความยืดหยุ่นด้านพฤติกรรมและการเคี้ยวอโศก et al., 2019) การย้อนกลับโดยการปิดล้อมของ D2Rs ใน striatum สนับสนุนเพิ่มเติมว่าจำเป็นต้องมีการเปิดใช้งาน D2Rs ซ้ำ ๆ เพื่อกระตุ้นพฤติกรรมที่ต้องกระทำ ร่วมกันชี้ข้อมูลไปยังบทบาทที่สำคัญของ D2Rs ภายในเส้นทางโดพามีนสมองส่วนกลางเพื่อกระตุ้นให้เกิดการแพ้และการบังคับ การบริหาร quinpirole ซ้ำ ๆ ซ้ำซากพฤติกรรมโคเคนที่เกิดขึ้น (Thompson และคณะ, 2010) และผลของหัวรถจักรของแอมเฟตามีน (จัดการและอัล 2010) เสริมสร้างความคิดที่ว่าการเปิดใช้งาน D2Rs ทำให้เกิดอาการแพ้ที่กระตุ้นให้เกิดอาการทางจิตและแนะนำกลไกที่ใช้ร่วมกันระหว่าง quinpirole และ psychostimulants-sensitization น่าสนใจผลการกระตุ้นของการกระตุ้นด้วย D2R ซ้ำหลายครั้งดูเหมือนว่าจะแข็งแกร่งกว่าการกระตุ้นโดย psychostimulants เนื่องจากหนูทุกคนที่รับการรักษาด้วย quinpirole จะพัฒนาความไวต่อการเคลื่อนไหวของ locomotorเอสโกบาร์ และคณะ, 2015) ในขณะที่ประมาณร้อยละหกสิบของหนูไวต่อแอมเฟตามีน (เอสโกบาร์ และคณะ, 2012; คาสโนว่าและคณะ, 2013).
ความไวต่อพฤติกรรมที่เกิดจากการเปิดใช้งาน D2R ซ้ำ ๆ จะมาพร้อมกับการดัดแปลงในวงจรรางวัล / แรงจูงใจ หนูที่ไวต่อยา quinpirole จะมีปริมาณโดปามินอิกต่ำใน NAc ซึ่งสังเกตได้ว่าเป็นพื้นฐานที่ลดลง (Koeltzow และคณะ, 2003) และการกระตุ้นโดพามีนโทนิคและการกระตุ้น phasic (เอสโกบาร์ และคณะ, 2015) แสดงถึงความสามารถในการปลดปล่อยโดปามีนลดลงของวงจรโดปามีนมิดเบรน ระดับโดปามีน Synaptic ใน NAc ถูกควบคุมโดยกิจกรรมของทั้ง DAT และโดปามีนเซลล์ประสาทกิจกรรม (ไปที่และเกรซ 2008) ซึ่ง ในร่างกาย ประกอบด้วยยาชูกำลังและการยิงระเบิด (Wilson และคณะ, 1977; เกรซและบันนีย์ 1980) รายงานก่อนหน้านี้แสดงให้เห็นว่าหนูที่ไวต่อความรู้สึก quinpirole แสดงจำนวนโดปามีนเซลล์ประสาทจำนวนน้อยลงในยาชูกำลังและการเผาไหม้ใน VTA (Sesia et al., 2013 โดย) ข้อมูลเหล่านี้ร่วมกันบ่งชี้ว่าการลดลงของการหลั่งโดปามีนที่เห็นหลังจากการแพ้ quinpirole เป็นผลมาจากการลดลงของกิจกรรมโดยรวมของเซลล์ประสาทโดปามีน พฤติกรรมบีบบังคับและกิจกรรมกระตุ้นประสาทสัมผัสที่เกิดจากการรักษาซ้ำด้วย quinpirole อาจเป็นผลมาจากการไวต่อยา D2Rs เนื่องจากการลดลงของ dopaminergic ใน NAc อันที่จริงหนู quinpirole ที่ไวต่อการแสดงเพิ่มขึ้นในการผูกพันของโดปามีน D2R (คัลเวอร์และคณะ, 2008) และการเพิ่มขึ้นของสถานะความสัมพันธ์ของตัวรับเหล่านี้ (Perreault et al., 2007) สนับสนุนสมมติฐานนี้
KOR-Dopamine ปฏิกิริยาในพฤติกรรมการชักนำให้เกิด Quinpirole
การศึกษาเบื้องต้นเกี่ยวกับบทบาทของ KOR ในพฤติกรรมบีบบังคับที่เกิดจาก D2R ก็มาจากห้องปฏิบัติการของ Szechtman กลุ่มนี้ตรวจสอบการบริหารร่วมกันของ KOR agonist U69593 ร่วมกับ quinpirole ในการทำงานของขมิ้นอ้อย โดยเฉพาะผู้เขียนได้ฉีดยาเข้าใต้ผิวหนังให้กับหนูที่มีส่วนผสมของ U69593 และ quinpirole จนกระทั่งการฉีดเสร็จสิ้น 8 ถึง 10 ครั้ง ตรงกันข้ามกับผล hypolocomotor ของ U69593 เพียงอย่างเดียวพบว่ามีภาวะ hyperlocomotion เมื่อให้ยาร่วมกับ quinpirole ในปริมาณต่ำ (presynaptic) และสูง (postynaptic) U69593 เปลี่ยนผล hypolocomotor ของ quinpirole ขนาด presynaptic เป็น hyperlocomotion และเพิ่มผลของ hyperlocomotor ของ quinpirole ขนาด postynaptic (Perreault et al., 2006) การกระตุ้น Co-activation ของ KORs ยังเร่งการเหนี่ยวนำของการกระตุ้นการเคลื่อนไหวและกระตุ้นผลของการกระตุ้นด้วย D2Rs เนื่องจากการเคลื่อนไหวสูงสุดที่ทำได้โดยการรักษาสองครั้งซ้ำซ้อนกับผลของหัวรถจักรที่เกิดจาก quinpirole เพียงอย่างเดียว (Perreault et al., 2006; เอสโกบาร์ และคณะ, 2017) การเปิดใช้งานร่วมของ KORs ยังช่วยเร่งการได้มาซึ่งพฤติกรรมการตรวจสอบแบบบังคับ (Perreault et al., 2007) ผลโพแทสเซียมเหล่านี้ของ KORs ต่อพฤติกรรมที่เหนี่ยวนำให้เกิด quinpirole จำเป็นต้องมีการกระตุ้น KORs ซ้ำ ๆ ในความเป็นจริงการฉีดยาแบบเฉียบพลันของ KOR agonist U69593 ไม่ได้ปรับเปลี่ยนกิจกรรมของ locomotor ในหนูที่ไวต่อยา quinpirole (เอสโกบาร์ และคณะ, 2017) กลไกการเกิดอาการแพ้ KOR potentiating D2R ไม่เป็นที่รู้จัก สิ่งหนึ่งที่เป็นไปได้คือระบบภายนอกคัปปาโอปิออยด์เองนั้นคือการทำให้เกิดอาการแพ้แบบพึ่งพา D2R อย่างไรก็ตามความเป็นไปได้นี้ถูกยกเลิกโดยแสดงให้เห็นว่าการดูแลก่อนหน้าของ norBNI ไม่ได้ปรับเปลี่ยนการกระตุ้นความรู้สึกของ locomotor ไปเป็น quinpirole โดยบอกว่า dynorphin ไม่ปล่อยการกระตุ้น D2R แบบดาวน์สตรีม (เอสโกบาร์ และคณะ, 2017) ข้อมูลนี้ไม่ได้ออกกฎว่า dynorphin อาจมีบทบาทในการกระตุ้นพฤติกรรมที่ต้องกระทำเช่นความเครียดทำให้เกิดการปล่อย dynorphin และการเปิดใช้งาน KORs ซึ่งช่วยให้พฤติกรรมที่ต้องกระทำ (McLaughlin และคณะ, 2003; แมคลาฟลิน และคณะ, 2006ก; McLaughlin และคณะ 2006b).
crosstalk ระหว่าง D2Rs และ KORs นั้นซับซ้อนและดูเหมือนว่าจะขึ้นอยู่กับว่าการเปิดใช้งานของตัวรับทั้งสองเกิดขึ้นพร้อมกันหรือแยกจากกันชั่วคราว ข้อมูลทางกายวิภาคระบุว่า crosstalk ระหว่าง D2Rs และ KORs สามารถเกิดขึ้นได้ล่วงหน้าในซอนและโซม่าของเซลล์โดปามีนเช่นเดียวกับ postsynaptically ใน MSNs ของ striatum แม้ว่ามันจะไม่ได้ออกกฎสำหรับ KORs ที่ตั้งอยู่บนซอนของระบบประสาทเคมีอื่น ๆ แต่ข้อมูลทางกายวิภาคชี้ให้เห็นถึงบทบาทโดยตรงของ KORs ที่ควบคุม D2Rs ไม่ว่าจะเป็นแบบเฉียบพลันหรือแบบซ้ำการกระตุ้น KORs จะลดการทำงานของ D2Rs ในเซลล์ประสาทโดปามีน การศึกษาทางไฟฟ้าวิทยาแสดงให้เห็นว่าการเปิดใช้งานเฉียบพลันของ KOR ในเซลล์ประสาทโดพามีนของ VTA และ SN ยับยั้งการยับยั้ง D2N-mediated ขัดขวางปัจจุบัน postynaptic ขัดขวางผลกระทบไกล่เกลี่ยโดยกลไกก่อนและ Postynaptic เป็น KOR ลดการปล่อยโดปามีนและ Dynorphin บล็อกฟอร์ดและอัล 2007) การศึกษาทางประสาทวิทยาแสดงให้เห็นว่าการเปิดใช้งานซ้ำของ KORs ยับยั้งการเหนี่ยวนำที่เกิดจากโดปามีน D2R ในการ NAC (Fuentealba และคณะ, 2006) ยิ่งไปกว่านั้น D2Rs และ KORs ที่เกิดขึ้นพร้อมกันแบบเฉียบพลันจะช่วยลดการยับยั้งการปล่อยโดปามีนใน NAc เมื่อเทียบกับผลของตัวรับแต่ละตัวเพียงอย่างเดียว (เอสโกบาร์ และคณะ, 2017) ดังนั้น presynaptic KORs ไม่กระทำการเสริมหรือทำงานร่วมกันกับ presynaptic D2Rs ในทางกลับกัน KORs จะยับยั้งหรือขัดขวางการยับยั้ง D2R กลไกนี้สามารถอธิบายผลของการเคลื่อนไหวของ locomotor ที่มีขนาดเฉียบพลันของ KOR agonists พร้อมกับ quinpirole ขนาดต่ำ (Perreault et al., 2006).
การศึกษาล่าสุดแสดงให้เห็นว่าการเปิดใช้งาน KOR ใน VTA ไกล่เกลี่ยพฤติกรรมบังคับซึ่งวัดเป็นการยับยั้งพฤติกรรมและฝังหินอ่อน (อับราฮัมและคณะ 2017) การตอกย้ำความคิดที่ว่าการเปิดใช้งาน KORs นั้นเป็นตัวกระตุ้นให้เกิดความจำเป็น ข้อมูลเผยแพร่โดย Margolis และคณะ (2006; 2008) ระบุว่าการมีปฏิสัมพันธ์ KORs และ D2R ควรเกิดขึ้นกับโดปามีนเซลล์ประสาทที่กำหนดเป้าหมายไปที่ mPFC (Margolis และคณะ, 2006; Margolis และคณะ, 2008) แต่อย่างไรก็ตาม ฟอร์ดและคณะ (2006; 2007) พบว่าการยับยั้ง KORs ของ D2R ที่ใช้สื่อกลาง IPSC เกิดขึ้นในเซลล์ประสาทโดปามีนซึ่งมีเป้าหมายไปที่ NAc (ฟอร์ดและอัล 2006; ฟอร์ดและอัล 2007) ข้อมูลเหล่านี้ร่วมกันแสดงให้เห็นว่าการมีปฏิสัมพันธ์ KOR กับ D2R ที่ช่อง somatodendritic ของเซลล์ประสาทโดปามีนอาจเกิดขึ้นอันเป็นผลมาจากการ crosstalk ในเซลล์ประสาทโดปามีนเดียวกัน ไม่ว่าจะเกิดขึ้นในการประมาณการ mesolimbic หรือ mesocortical ยังคงเป็นที่ถกเถียงกัน
อย่างน่าทึ่ง KOR พบใน MSNs ของ NAc (เอสโกบาร์ และคณะ, 2017; Tejeda et al., 2017 โดย) ดังนั้นจึงแสดงให้เห็นว่าศักยภาพของพฤติกรรมการบีบอัดที่เกิดจาก D2R ยังสามารถเกิดขึ้นได้โดยการกระทำโดยตรงกับเซลล์เป้าหมายของเซลล์ประสาทโดปามีน ในเรื่องนี้มันเป็นมูลค่าการกล่าวขวัญว่าการบริหารซ้ำของ U69593 เพิ่มปริมาณของ D2Rs ในสถานะความสัมพันธ์สูง (Perreault et al., 2007) ข้อมูลทางประสาทวิทยาระบุว่าการลดลงของระดับโดปามีนนอกเซลล์เกี่ยวข้องกับการแพ้ยา D2R การเปิดใช้งานร่วม KORs ไม่ได้ลดระดับของโดปามีนนอกเซลล์ใน NAc ลงอีกแล้วโดยการเปิดใช้งาน D2Rs ซ้ำ ๆ (เอสโกบาร์ และคณะ, 2017) พิจารณาบทบาทของ KOR แบบ presynaptic ที่เร่งหรือทำให้เกิดการแพ้ของ D2Rs ใน NAc ผ่านกลไกนี้ ดังนั้น KORs จึงกระตุ้นกลไกของโมเลกุลที่ช้าซึ่งส่งผลต่อประสาทและพฤติกรรมของ D2Rs ยิ่งขึ้นซึ่งชี้ให้เห็นว่าการเพิ่มความไวในการเคลื่อนที่นั้นอาจเกิดจากการปรับโพสท์ซินแน็ปทิคมากกว่าแบบ presynaptic ในเรื่องนี้การเปิดใช้งานซ้ำ ๆ ของ KORs สามารถทำให้เกิดการยับยั้ง D2R ทางอ้อม striatal pathway switching D1R / D2R balance ไปเป็น D1R inducing compulsivity (รูป 1).
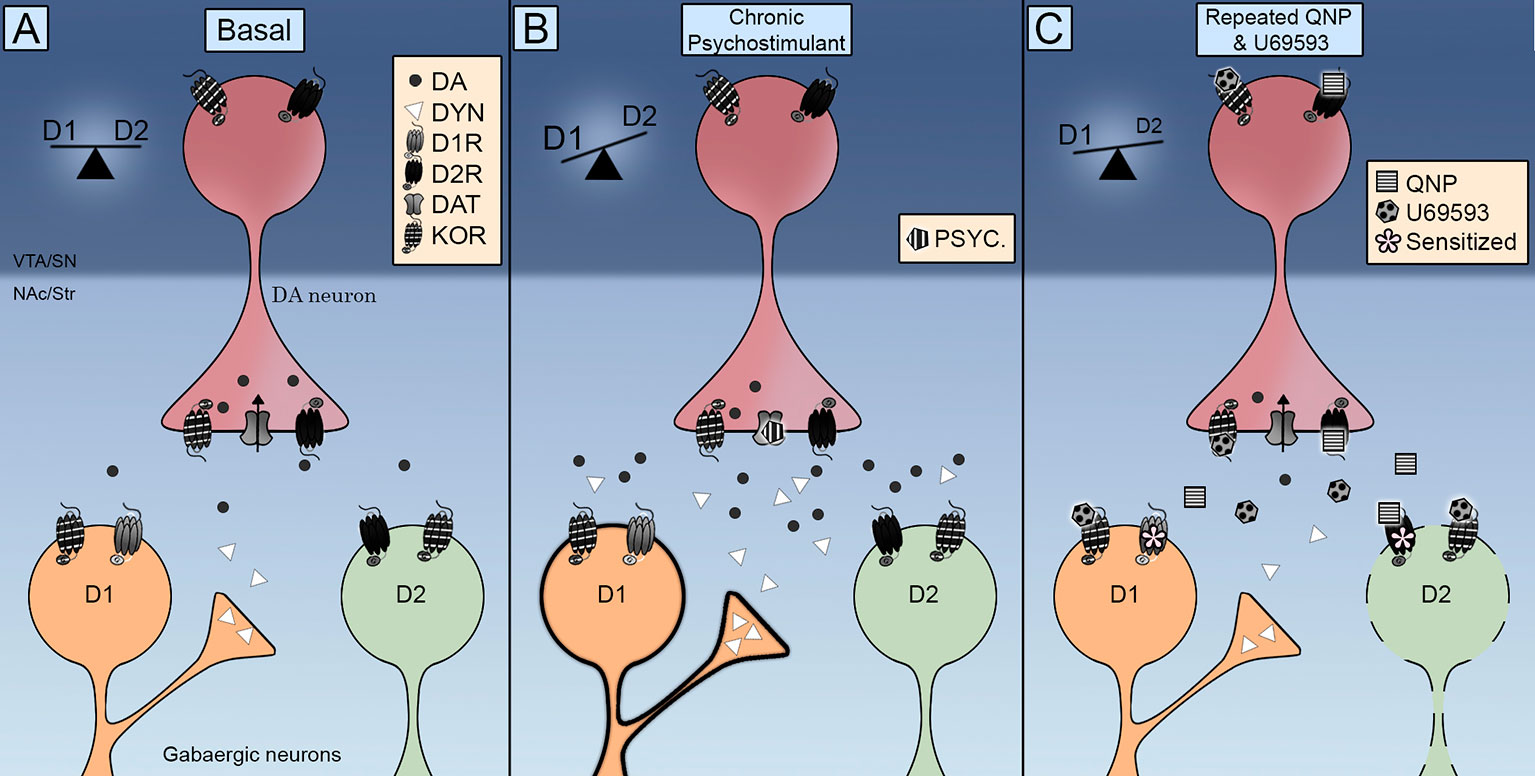 รูป 1 รูปแบบการผสมผสานของการควบคุม Kappa Opioid Receptors (KOR) บน phatways ทางตรง (D1R) และทางอ้อม (D2R) (A) KOR ตั้งอยู่ล่วงหน้าด้วยความเจ็บปวดบนขั้วโดปามีนและโพสต์แบบไซนัสในเซลล์ประสาทขนาดกลาง (MSN) การเปิดใช้งานควบคุมระดับสารกระตุ้นโดปามีนและการโลคัลไลซ์เซชันของมันส่งเสริมการมีปฏิสัมพันธ์กับโดพามีนขนย้าย (DAT) และตัวรับโดปามีน D2 (B) การได้รับสารกระตุ้นทางจิตซ้ำ ๆ จะเพิ่มขึ้นทั้งในระดับโดปามีนและเซลฟลูออโรนิกที่เพิ่มขึ้น การเปิดใช้งานตัวรับ D1 และ D2 สลับความสมดุลเป็นเส้นทางเดินตรง D1R เพื่อส่งเสริมการกระตุ้นการเคลื่อนไหวของหัวรถจักร (C) การบริหารร่วมของ quinpirole และ U69593 นั้นมาพร้อมกับการลดลงของระดับโดพามีนนอกเซลล์ การเปิดใช้งานร่วมกันของตัวรับ KOR และ D2 จะทำให้เส้นทางทางอ้อมทางอ้อมของ D2 อ่อนแอลงทำให้เกิดพฤติกรรมการบีบบังคับ
รูป 1 รูปแบบการผสมผสานของการควบคุม Kappa Opioid Receptors (KOR) บน phatways ทางตรง (D1R) และทางอ้อม (D2R) (A) KOR ตั้งอยู่ล่วงหน้าด้วยความเจ็บปวดบนขั้วโดปามีนและโพสต์แบบไซนัสในเซลล์ประสาทขนาดกลาง (MSN) การเปิดใช้งานควบคุมระดับสารกระตุ้นโดปามีนและการโลคัลไลซ์เซชันของมันส่งเสริมการมีปฏิสัมพันธ์กับโดพามีนขนย้าย (DAT) และตัวรับโดปามีน D2 (B) การได้รับสารกระตุ้นทางจิตซ้ำ ๆ จะเพิ่มขึ้นทั้งในระดับโดปามีนและเซลฟลูออโรนิกที่เพิ่มขึ้น การเปิดใช้งานตัวรับ D1 และ D2 สลับความสมดุลเป็นเส้นทางเดินตรง D1R เพื่อส่งเสริมการกระตุ้นการเคลื่อนไหวของหัวรถจักร (C) การบริหารร่วมของ quinpirole และ U69593 นั้นมาพร้อมกับการลดลงของระดับโดพามีนนอกเซลล์ การเปิดใช้งานร่วมกันของตัวรับ KOR และ D2 จะทำให้เส้นทางทางอ้อมทางอ้อมของ D2 อ่อนแอลงทำให้เกิดพฤติกรรมการบีบบังคับ
ความแตกต่างระหว่างเพศของ KOR-Dopamine ในพฤติกรรมบีบบังคับ
การศึกษาทางคลินิกแสดงให้เห็นความแตกต่างทางเพศในพฤติกรรมบีบบังคับรวมถึงการแสวงหายาเสพติด อาการเริ่มแรกของโรค OCD นั้นพบได้ในผู้ชายเมื่อเทียบกับผู้หญิงมาติส และคณะ, 2011) กับผู้หญิงที่แสดงความชุกของการปนเปื้อนและอาการทำความสะอาดมากขึ้น (Labad et al., 2008) เกี่ยวกับความแตกต่างทางเพศในการติดยาเสพติดหลักฐานทางคลินิกบ่งชี้ว่าในขณะที่การใช้ยามีความแพร่หลายมากขึ้นในผู้ชายผู้หญิงแสดงความก้าวหน้าเร็วกว่าผู้ชายในการค้นหายาเสพติดซึ่งต้องกระทำHernandez-Avila และคณะ, 2004; Fattore และ Melis, 2016).
เมื่อเร็ว ๆ นี้หลักฐานทางคลินิกก่อนได้เน้นฐาน neurobiological อย่างยิ่งหนุนความแตกต่างทางเพศในยาเสพติดสังเกตในการศึกษาทางคลินิก (เบกเกอร์และ Chartoff, 2019) การสังเกตในช่วงต้นโดยใช้ microdialysis ฟลักซ์แบบไม่มีตาข่ายแสดงให้เห็นว่าความเข้มข้นของโดปามีนนอกเซลล์ใน striatum หลังแตกต่างกันไปในระหว่างรอบการเป็นสัดที่มีระดับสูงกว่าใน proestrus และ estrus เมื่อเทียบกับ diestrus ยิ่งไปกว่านั้นในขณะที่การตัดรังไข่จะลดความเข้มข้นของสาร dopamine extracellular ในทารกในครรภ์ แต่การตัดอัณฑะของหนูเพศผู้จะไม่เปลี่ยนแปลงความเข้มข้นของสาร dopamine striatal extracellular (เสี่ยวและเบกเกอร์ 1994) แนะนำบทบาทสำคัญของฮอร์โมนรังไข่ต่อกิจกรรมโดปามีน นอกจากนี้ฮอร์โมนเพศหญิงยังควบคุมการตอบสนองต่อยาจิตเวช ตอนต้น ในหลอดทดลอง การทดลองแสดงให้เห็นว่า estradiol บวกโปรเจสเตอโรนเรียกคืนการปลดปล่อยโดปามีนจากแอมเฟตามีนที่เกิดจากเนื้อเยื่อของทารกแรกเกิดที่ได้รับจากหนูเพศเมียรังไข่เบกเกอร์และรามิเรซ 1981) เมื่อเร็ว ๆ นี้การศึกษาวัฏจักรของวัฏจักรการสแกนอย่างรวดเร็วแสดงให้เห็นว่าเพศหญิงมีการปลดปล่อยโดปามีนกระตุ้นด้วยไฟฟ้าและการดูดซึมที่ดีกว่าเมื่อเปรียบเทียบกับเพศชาย (Walker และคณะ, 2000) ความแตกต่างทางเพศเหล่านี้ในสารสื่อประสาทโดปามีนสามารถอธิบายได้ว่ามีโคเคนและยาบ้าในระดับสูง (Roberts et al., 1989; Cox และคณะ, 2013).
กฎระเบียบของ KOR ในระดับสารกระตุ้นโดปามีนยังแสดงให้เห็นถึงความแตกต่างทางเพศ (Chartoff และ Mavrikaki, 2015) ใช้การกระตุ้นด้วยตนเองในสมองและแรงดันไฟฟ้าแบบวัฏจักร คอนเวย์และคณะ (2019) แสดงให้เห็นว่าความไวที่ลดลงต่อผล anhedonic เฉียบพลันของ KON agonist ที่สังเกตได้ในหนูตัวเมียเมื่อเปรียบเทียบกับหนูเพศผู้พร้อมกับการยับยั้งการปล่อยสารกระตุ้นโดปามีนที่กระตุ้นใน NAcคอนเวย์และอัล 2019) มันได้รับการแนะนำว่า estradiol มีส่วนช่วยในการยับยั้งทื่อโดปามีนที่ปล่อยออกมาในหนูตัวเมียหลังการกระตุ้นด้วย KORอับราฮัมและคณะ 2018) ในขณะที่ crosstalk ระหว่าง KORs และการส่งสัญญาณ dopamine ได้รับการศึกษาในเพศชาย (Tejeda และ Bonci, 2019) การวิจัยเกี่ยวกับปฏิสัมพันธ์นี้และผลกระทบต่อกระบวนการติดยาเสพติดในเพศหญิงยังไม่เพียงพอ (Chartoff และ Mavrikaki, 2015) ในหนูตัวเมียการบริหารแบบเฉียบพลันของ KOR agonist U69593 ทำให้เกิดภาวะ hyperlocomotion แบบโคเคนที่ทำให้เกิดโคเคนลดลงทั้งในการควบคุมและในหนูที่ถูกตัดรังไข่ ที่น่าสนใจ U69593 บริหารซ้ำซ้ำลด hyperlocomotion โคเคนที่เกิดขึ้นในลักษณะที่ขึ้นอยู่กับ estradiol (Puig-Ramos และคณะ, 2008) ข้อมูลเหล่านี้ชี้ให้เห็นว่า estradiol primes การกระทำ KOR ในหนูตัวเมียผลที่อาจเกี่ยวข้องกับความแตกต่างทางเพศในการตอบสนองต่อความเครียด (Puig-Ramos และคณะ, 2008) ไม่ว่าจะเป็นหนูเพศเมียการเปิดใช้งานซ้ำของ KORs ช่วยให้มีการปลดปล่อยโดปามีนแบบ striatal ตามที่สังเกตในเพศชายเป็นคำถามที่ไม่ได้รับคำตอบ
ถึงแม้ว่าการอำนวยความสะดวกในการปลดปล่อยโดปามีนที่กระตุ้นให้เกิดจิตนั้นจะสังเกตได้ในเพศหญิงเมื่อเทียบกับหนูเพศชาย แต่ความแตกต่างระหว่างเพศในกลไกโดปามีนที่แฝงอยู่ในแอมเฟตามีนเบกเกอร์ 1999) การได้รับยาบ้าซ้ำ ๆ ทำให้เกิดการเคลื่อนไหวของหัวรถจักรมากขึ้นทั้งในวัยรุ่น (แมทธิวส์และแมคคอร์มิค 2007) และหนูตัวเมียผู้ใหญ่ (มิเลซี-ฮัลเล et al., 2007) กับหนูวัยรุ่นหญิงแสดงอาการแพ้หัวรถจักรที่แข็งแกร่งมากขึ้นหลังจากได้รับยาบ้าซ้ำ ๆ การเปิดใช้งานทารกแรกเกิดของตัวรับ D2 มีฤทธิ์กระตุ้นให้เกิดอาการแพ้ทางแอมเฟตามีนเฉพาะในหนูตัวเมียเท่านั้น (Brown และคณะ, 2011) ดังที่ได้กล่าวไว้ก่อนหน้านี้มันถูกพบในหนูตัวผู้การสัมผัสซ้ำ ๆ กับ agonist D2 ทำให้เกิดอาการแพ้ทางประสาทสัมผัสและพฤติกรรมคล้ายบังคับDvorkin และคณะ, 2006) ยิ่งไปกว่านั้นการเปิดใช้งานร่วมของ KOR โพเทนชิเซททำให้เกิดอาการแพ้หัวรถจักรที่เกิดจากการสัมผัสกับ quinpirole ซ้ำ ๆ ช่วยอำนวยความสะดวกในการควบคุมการยับยั้งตัวรับ D2 ในการปลดปล่อย DA ใน NAc (เอสโกบาร์ และคณะ, 2017) ความแตกต่างทางเพศเช่นความไวที่ลดลงต่อการยับยั้ง KOR ต่อการปลดปล่อยโดปามีนในเพศหญิงคอนเวย์และอัล 2019) อาจอธิบายถึงการมีส่วนร่วมที่แตกต่างกันของ KOR ในการแสวงหายาเสพติด
สรุป
KORs จะปรับสัญญาณโดปามีนอย่างไรเพื่ออธิบายพฤติกรรมที่มีแรงจูงใจอย่างละเอียดและเมื่อไหร่ที่มันจะส่งผลให้เกิดพฤติกรรมที่ต้องกระทำโดยไว ข้อมูลทางกายวิภาคแสดงให้เห็นว่า KORs อยู่ในตำแหน่งที่สามารถควบคุมกิจกรรม synaptic ของเซลล์ประสาทโดปามีนในสมองส่วนกลางได้ ข้อมูลการทำงานระบุว่า KORs ควบคุมการทำงานของ DAT และ D2R เช่นเดียวกับอัตราการยิงของเซลล์ประสาทโดปามีน หลักฐานเริ่มต้นแสดงให้เห็นว่าการเปิดใช้งานเฉียบพลันของ KORs ลดการปล่อยโดปามีนที่เกิดจากยาเสพติดได้รับการเสริมด้วยข้อมูลบ่งชี้ว่าการเปิดใช้งานซ้ำของ KOR อำนวยความสะดวกในการปล่อยโดปามีนและการค้นหายาเสพติด โดปามีนส่งสัญญาณสมดุลเส้นทางการส่งออกโดยตรงและโดยอ้อมจากพื้นที่ striatal (รูปที่ 1A) ไม่ว่าจะเป็นการกระตุ้นเรื้อรังด้วย psychostimulants ที่เพิ่มการปลดปล่อยโดปามีนทั้ง D1R และ D2Rรูปที่ 1B) หรือ quinpirole ที่เปิดใช้งานเฉพาะ D2R (รูป 1C) ส่งผลให้เกิดอาการแพ้หัวรถจักรและพฤติกรรมการบีบบังคับโดยเส้นทางอ้อมทางอ้อมที่ทำให้ร่างกายทรุดโทรมของ D2R ดังนั้นการเปลี่ยนสมดุลให้เป็นเส้นทางเดินตรง D1R ส่ง KOR จะเพิ่มขึ้นในระหว่างการบริโภค psychostimulant เรื้อรังโดยการเพิ่มขึ้นของ dynorphin ในเซลล์ D1 striatalรูปที่ 1B) การส่ง KOR ที่ปรับปรุงแล้วถูกจำลองในรูปแบบทางเภสัชวิทยาของ OCD โดยการบริหาร U69593 การเปิดใช้งาน KOR ร่วมกันนี้จะทำให้เส้นทางเส้นทางอ้อมทางอ้อม D2 อ่อนลง (รูป 1C) การวิจัยในอนาคตควรดำเนินการเพื่ออธิบายผลของการเปิดใช้งาน KOR อย่างเต็มที่เกี่ยวกับกิจกรรม DAT เข้าใจบทบาทของระบบ KOR endogeous ในการชักนำให้เกิดการกระตุ้นด้วย quinpirole และพิจารณาการมีส่วนร่วมของระบบ KOR กับความแตกต่างทางเพศ
ผลงานของผู้เขียน
AE, MA, และ JF มีส่วนทำให้ความคิดของต้นฉบับ AE และ JF เขียนร่างแรกของต้นฉบับด้วยข้อมูลจาก MA แม่และ JC มีส่วนร่วมในการทบทวนและแก้ไขต้นฉบับ ผู้เขียนทั้งหมดอนุมัติให้ตีพิมพ์
การฝากและถอนเงิน
งานของผู้เขียนที่อ้างถึงในรีวิวนี้ได้รับการสนับสนุนโดย FONDECYT ให้หมายเลข: 1110352 และ 1150200 ถึง MA; 1141088 ถึง JF; DIPOG มอบ 391340281 ให้ JF FONDECYT นักแสดงหลังปริญญาเอก 3170497 ถึง JC และ 3190843 ถึง AE
ขัดผลประโยชน์
ผู้เขียนประกาศว่าการวิจัยได้ดำเนินการในกรณีที่ไม่มีความสัมพันธ์ทางการค้าหรือทางการเงินใด ๆ ที่อาจตีความได้ว่าเป็นความขัดแย้งทางผลประโยชน์ที่อาจเกิดขึ้น
ขณะนี้บรรณาธิการจัดการกำลังจัดการหัวข้อการวิจัยกับหนึ่งในผู้เขียนเจเอฟและยืนยันว่าไม่มีความร่วมมืออื่นใด
อ้างอิง
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, แบร์ด, MA, Kieffer, BL, et al. (2017) การเปิดใช้งานตัวรับ Kappa opioid ในเซลล์ประสาทโดพามีนขัดขวางการยับยั้งพฤติกรรม Neuropsychopharmacology 43 (2), 362–372 ดอย: 10.1038 / npp.2017.133
Abraham, AD, Schattauer, SS, Reichard, KL, โคเฮน, JH, Fontaine, HM, Song, AJ, et al. (2018) การควบคุมเอสโตรเจนของ GRK2 ยับยั้งการรับสัญญาณคัปปาโอปิอิดส่งสัญญาณไกล่เกลี่ยความเจ็บปวด แต่ไม่ใช่ความเกลียดชัง J. Neurosci 38 (37), 8031–8043 ดอย: 10.1523 / JNEUROSCI.0653-18.2018
Asaoka, N. , Nishitani, N. , Kinoshita, H. , Nagai, Y. , Hatakama, H. , Nagayasu, K. , et al. (2019) adenosine A2A receptor antagonist จะช่วยปรับปรุงอาการของโรคจิต quinpirole ที่เกิดซ้ำหลายครั้ง eNeuro 6 (1), 1–16 ENEURO.0366-18.2019 ดอย: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R. , Pasternak, GW, Majumdar, S. , Kivell, BM (2019) MP1104 ซึ่งเป็นตัวรับออปติคัลแคปปาเดลตา opioid ผสมมีคุณสมบัติต่อต้านโคเคนและลดผลข้างเคียงในหนู Neuropharmacology 150, 217 – 228 ดอย: 10.1016 / j.neuropharm.2019.02.010
Azocar, VH, Sepúlveda, G. , Ruiz, C. , Aguilera, C. , Andrés, ME, Fuentealba, JA (2019) การปิดกั้นของตัวรับ kappa-opioid จะทำให้เกิดการเปลี่ยนแปลงในการเคลื่อนไหวของโดปามีน datolateral striatum datamine ในระหว่างการรับสารแอมเฟตามีน J. Neurochem 148, 348–358 ดอย: 10.1111 / jnc.14612
Béguin, C. , Potuzak, J. , Xu, W. , Liu-Chen, LY, Streicher, JM, Groer, CE, et al. (2012) คุณสมบัติการส่งสัญญาณที่แตกต่างกันที่ตัวรับ kappa opioid ของ 12-epi-salvinorin A และ analogues Bioorg Med Chem เลทท์ 15; 22 (2), 1023–1026 doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005) ผลกระทบที่แตกต่างของนวนิยายคู่แคปปา opioid ตัวรับ, JDTic, ในการคืนสถานะของการแสวงหาโคเคนที่เกิดจากแรงกดดัน footshock เทียบกับช่วงเวลาโคเคนและผลกระทบเหมือนยากล่อมประสาทในหนู Psychopharmacol (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
เบกเกอร์, JB, Chartoff, E. (2019) ความแตกต่างทางเพศในกลไกของระบบประสาทเป็นสื่อกลางในการให้รางวัลและการเสพติด Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
เบกเกอร์, JB, รามิเรซ, VD (1981) ความแตกต่างทางเพศในแอมเฟตามีนกระตุ้นให้มีการปล่อย catecholamines ออกจากเนื้อเยื่อของ striatal ในหลอดทดลอง สมอง Res 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
เบกเกอร์, JB (1999) ความแตกต่างระหว่างเพศในการทำงานของโดปามีนในแถบ striatum และนิวเคลียส Pharmacol Biochem Behav 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Berridge, KC, Robinson, TE (2016) ความชอบความอยากและทฤษฎีการกระตุ้นให้ติดไว am จิตวิทยา 71, 670–679 ดอย: 10.1037 / amp0000059
Berridge, KC, Venier, IL, Robinson, TE (1989) การวิเคราะห์การรับรู้รสชาติของ 6-hydroxydopamine-aphagia aphagia: ผลกระทบต่อการกระตุ้นและ anhedonia สมมติฐานของฟังก์ชั่นโดพามีน Behav Neurosci 103, 36 – 45 ดอย: 10.1037 / 0735-7044.103.1.36
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, วูดส์, JH (1994) ผลกระทบที่แตกต่างกันของระบบยา binaltorphimine (หรือ BNI) ที่มีระบบต่อ agonists kappa-opioid ในหนูทดลอง Psychopharmacology 115, 311 – 319 doi: 10.1007 / BF02245071
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J. , Smith, ML (2011) การกระตุ้นอาการยาบ้าของแอมเฟตามีนและความชอบในสถานที่ปรับอากาศในหนูขาววัยรุ่นและหนูเพศหญิงที่ได้รับการรักษาด้วยควินพิโร่ Behav Pharmacol 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Bruchas, MR, Chavkin, C. (2010) Kinase ลดหลั่นและการส่งสัญญาณกำกับแกนด์ที่ตัวรับ kappa opioid Psychopharmacol (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J. , O'Mara, SM (2018). บทบาทที่เป็นไปได้สำหรับผู้รับ opioid ในแรงจูงใจและโรคซึมเศร้าที่สำคัญ Prog ความต้านทานของสมอง 239, 89–119 ดอย: 10.1016 / bs.pbr.2018.07.009
Carr, DB, Sesack, SR (2000) เซลล์ประสาทที่มี GABA ในโครงการพื้นที่หน้าท้องหนูหนูไปยังเยื่อหุ้มสมองส่วนหน้า ไซแนปส์ 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Casanova, JP, Velis, GP, Fuentealba, JA (2013) อาการแพ้ที่เกิดจากแอมเฟตามีนนั้นมาพร้อมกับการปลดปล่อยโดปามีน K + ที่เพิ่มขึ้นสูงในคอร์เท็กซ์ prefrontal หนูที่อยู่ตรงกลาง Behav สมอง Res 237, 313 – 317 ดอย: 10.1016 / j.bbr.2012.09.052
Chartoff, EH, Mavrikaki, M. (2015) ความแตกต่างทางเพศในฟังก์ชั่นตัวรับแคปปา opioid และผลกระทบที่อาจเกิดขึ้นกับการติด ด้านหน้า Neurosci 9, 466. ดอย: 10.3389 / fnins.2015.00466
Chartoff, EH, Ebner, SR, Sparrow, A. , Potter, D. , Baker, PM, Ragozzino, ME, et al. (2016) เวลาสัมพัทธ์ระหว่างการเปิดใช้งานตัวรับแคปปา opioid และโคเคนเป็นตัวกำหนดผลกระทบต่อการให้รางวัลและการปล่อยโดปามีน Neuropsychopharmacology 41, 989 – 1002 doi: 10.1038 / npp.2015.226
Chavkin, C. , Koob, GF (2016) Dynorphin, dysphoria และการพึ่งพาอาศัยกัน: ความเครียดของการติดยาเสพติด Neuropsychopharmacology 41, 373 – 374 doi: 10.1038 / npp.2015.258
Chefer, VI, Czyzyk, T. , Bolan, EA, Moron, J. , Pintar, JE, Shippenberg, T. S. (2005) ระบบตัวรับแคปปา - โอปิออยด์ภายนอกควบคุมพลวัตของโดปามีน mesoaccumbal และความเสี่ยงต่อโคเคน J. Neurosci 25, 5029 – 5037 doi: 10.1523 / JNEUROSCI.0854-05.2005
Chefer, VI, Zapata, A. , Shippenberg, TS, Bungay, PM (2006) ปริมาณ microdialysis no-net-flux เชิงปริมาณช่วยให้ตรวจจับการเพิ่มขึ้นและลดลงของการดูดซับโดปามีนในนิวเคลียสของหนู J. Neurosci วิธีการ 155, 187 – 193 doi: 10.1016 / j.jneumeth.2005.12.018
Cole, RL, Konradi, C. , Douglass, J. , Hyman, SE (1995) การปรับตัวของเซลล์ประสาทกับแอมเฟตามีนและโดปามีน: กลไกระดับโมเลกุลของการควบคุมยีน prodynorphin ในหนู striatum เซลล์ประสาท 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Conway, SM, Puttick, D. , Russell, S. , Potter, D. , Roitman, MF, Chartoff, E. H. (2019). ผู้หญิงมีความไวน้อยกว่าเพศชายต่อผลที่สร้างแรงบันดาลใจและการยับยั้งโดปามีนของการกระตุ้นตัวรับ kappa opioid Neuropharmacology 146, 231 – 241 ดอย: 10.1016 / j.neuropharm.2018.12.002
รับมือ, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, Brown, RW (2010) การรักษาทารกแรกเกิด quinpirole ช่วยกระตุ้นการเคลื่อนไหวของหัวรถจักรและโดปามีนในนิวเคลียส accumbens ในการตอบสนองต่อการรักษาแอมเฟตามีนในวัยผู้ใหญ่ ไซแนปส์ 64, 289 – 300 doi: 10.1002 / syn.20729
Cox, BM, Young, AB, See, RE, Reichel, CM (2013) ความแตกต่างระหว่างเพศในการหายาบ้าในหนู: ผลกระทบของออกซิโตซิน Psychoneuroendocrinology 38, 2343 – 2353 doi: 10.1016 / j.psyneuen.2013.05.005
คัลเวอร์, KE, Szechtman, H. , Levant, B. (2008) เปลี่ยนแปลงตัวรับโดปามีนที่มีลักษณะคล้าย D2 ในหนูด้วยการทำให้ไวต่อพฤติกรรมต่อ quinpirole: ผลของการรักษาล่วงหน้าด้วย Ro 41-1049 Eur J. Pharmacol 592, 67 – 72 doi: 10.1016 / j.ejphar.2008.06.101
Dalman, FC, O'Malley, KL (1999). kappa-Opioid ความอดทนและการพึ่งพาในวัฒนธรรมของเซลล์ประสาทส่วนกลาง dopaminergic J. Neurosci 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
De Vries, TJ, Schoffelmeer, AN, Binnekade, R. , Raasø, H. , Vanderschuren, LJ (2002) การกำเริบของพฤติกรรมการค้นหาโคเคนและเฮโรอีนที่สื่อกลางโดยตัวรับโดปามีน D2 นั้นขึ้นอยู่กับเวลาและเกี่ยวข้องกับการทำให้ไวต่อพฤติกรรม Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Devine, DP, Leone, P. , Pocock, D. , Wise, RA (1993) การมีส่วนร่วมที่แตกต่างกันของตัวรับ tegmental mu, เดลต้าและตัวรับแคปปา opioid ในการปรับการปลดปล่อยโดปามีนพื้นฐาน mesolimbic: ในการศึกษา microdialysis ในร่างกาย J. Pharmacol ประสบการณ์ Ther 266, 1236-1246
Di Chiara, G. , Imperato, A. (1988) ผลตรงข้ามของ mu และ kappa opiate agonists ต่อการปลดปล่อยโดปามีนในนิวเคลียส accumbens และในหางด้านหลังของหนูที่เคลื่อนไหวอย่างอิสระ J. Pharmacol ประสบการณ์ Ther 244, 1067 – 1080 doi: 10.1073 / pnas.85.14.5274
Dvorkin, A. , Perreault, ML, Szechtman, H. (2006) การพัฒนาและการจัดระเบียบชั่วคราวของการตรวจสอบบังคับซึ่งเกิดจากการฉีดซ้ำของโดปามีน agonist quinpirole ในรูปแบบสัตว์ของโรคย้ำคิดย้ำคิดย้ำทำ Behav สมอง Res 169, 303 – 311 ดอย: 10.1016 / j.bbr.2006.01.024
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010) อาการซึมเศร้าของ kappa opioid receptor agonist salvinorin A มีความสัมพันธ์กับการลดลงของการปล่อยสาร dopamine ในระบบประสาทในนิวเคลียส accumbens Psychopharmacol (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M. , Zhang, S. , McDevitt, RA, Wu, J. , et al. (2017) ความจำเพาะของวงจรในสถาปัตยกรรมการยับยั้งของ VTA ควบคุมพฤติกรรมที่เกิดจากโคเคน ชัยนาท Neurosci 20 (3), 438–448 ดอย: 10.1038 / nn.4482
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014) Kappa opioid receptor ยืนยันการใช้งาน potentiates การเพิ่มขึ้นของโคเคนที่เกิดขึ้นในการปลดปล่อยโดพามีนปรากฏในร่างกายในนิวเคลียสของหนู Neuropsychopharmacology 39, 3036 – 3048 doi: 10.1038 / npp.2014.157
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, et al. (2015) Kappa Opioid Receptor-Induced Aversion ต้องใช้ p38 MAPK การเปิดใช้งานในเซลล์ประสาทโดปามีน VTA J. Neurosci 35, 12917 – 12931 doi: 10.1523 / JNEUROSCI.2444-15.2015
Eilam, D. , Szechtman, H. (1989) Biphasic effect ของ D-2 agonist quinpirole ต่อการเคลื่อนไหวและการเคลื่อนไหว Eur J. Pharmacol 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Eilam, D. , Szechtman, H. (2005) Psychostimulant-induced พฤติกรรมเป็นแบบจำลองสัตว์ของโรค obsessive-compulsive: วิธีการเชิงจริยธรรมกับรูปแบบของพิธีกรรมบังคับ ระบบประสาทส่วนกลาง 10, 191–202 ดอย: 10.1017 / S109285290001004X
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012) การรักษาซ้ำด้วยตัวรับแคปปา opioid ตัวเอก U69593 กลับเพิ่ม K + เหนี่ยวนำให้เกิดการปลดปล่อยโดปามีนในนิวเคลียส accumbens แต่ไม่ใช่การแสดงออกของความไวในการเคลื่อนไหวของหนูในกลุ่มแอมเฟตามีน Neurochem int 60 (4), 344–349 doi: 10.1016 / j.neuint.2012.01.014
Escobar, AP, Cornejo, FA, Olivares-Costa, M. , González, M. , Fuentealba, JA, Gysling, K. , et al. (2015) โดพามีนและกลูตาเมตลดการรับสารสื่อประสาทในนิวเคลียสของหนูที่ได้รับไวจาก quinpirole ไวต่อการยับยั้งการทำงานของ D2 autoreceptor J. Neurochem 134, 1081–1090 ดอย: 10.1111 / jnc.13209
Escobar, AP, González, MP, Meza, RC, Noches, V. , Henny, P. , Gysling, K. , et al. (2017) กลไกการทำงานของตัวรับแคปปา opioid potentiation ของฟังก์ชั่นตัวรับโดปามีน D2 ในการกระตุ้นอาการชักแบบควินพิโรเล่ที่เกิดขึ้นในหนูขาว int J. Neuropsychopharmacol 20, 660–669 ดอย: 10.1093 / ijnp / pyx042
Everitt, BJ, Robbins, TW (2005) ระบบประสาทของการเสริมแรงสำหรับการติดยาเสพติด: จากการกระทำไปจนถึงนิสัยการบังคับ ชัยนาท Neurosci 8, 1481 – 1489 ดอย: 10.1038 / nn1579
Fattore, L. , Melis, M. (2016) ความแตกต่างทางเพศในพฤติกรรมหุนหันพลันแล่นและบีบบังคับ: มุ่งเน้นไปที่การติดยาเสพติด ผู้เสพติด Biol 21 (5), 1043–1051 ดอย: 10.1111 / adb.12381
Ferrario, CR, Gorny, G. , Crombag, HS, Li, Y. , Kolb, B. , Robinson, TE (2005) ความหนาแน่นของระบบประสาทและพฤติกรรมที่เกี่ยวข้องกับการเปลี่ยนจากการควบคุมการใช้โคเคนที่เพิ่มขึ้น Biol Psy 58 (9), 751–9
Figee, M. , Pattij, T. , Willuhn, I. , Luigjes, J. , van den Brink, W. , Goudriaan, A. , et al. (2016) ความบีบบังคับในความผิดปกติที่ครอบงำและการเสพติด Eur Neuropsychopharmacol 26, 856 – 868 doi: 10.1016 / j.euroneuro.2015.12.003
ฟอร์ด, CP, Mark, GP, Williams, JT (2006) คุณสมบัติและการยับยั้ง opioid ของเซลล์ประสาทโดปามีน mesolimbic แตกต่างกันไปตามสถานที่เป้าหมาย J. Neurosci 26, 2788 – 2797 doi: 10.1523 / JNEUROSCI.4331-05.2006
ฟอร์ด, CP, Beckstead, MJ, วิลเลียมส์, JT (2007) การยับยั้ง Kappa opioid ของการยับยั้งโดปามีน somatodendritic J. Neurophysiol 97, 883 – 891 doi: 10.1152 / jn.00963.2006
Fuentealba, JA, Gysling, K. , Magendzo, K. , Andrés, ME (2006) การบริหารซ้ำของตัวรับตัวเลือกแคปปา - opioid ตัวเอก U-69593 เพิ่มระดับโดปามีนกระตุ้นเซลล์นอกเซลล์ในนิวเคลียสหนู accumbens J. Neurosci Res 84, 450 – 459 doi: 10.1002 / jnr.20890
Fuentealba, JA, Gysling, K. , Andrés, ME (2007) การตอบสนองของหัวรถจักรที่เพิ่มขึ้นต่อแอมเฟตามีนที่เกิดขึ้นจากการบริหารซ้ำของ agonist ตัวเลือก kappa-opioid ตัวเลือก U-69593 ไซแนปส์ 61, 771 – 777 doi: 10.1002 / syn.20424
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008) ผลของการให้ salvinorin A แบบเฉียบพลันและแบบซ้ำ ๆ ต่อการทำงานของโดปามีนใน striatum หนูหลัง Psychopharmacol (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C. , Belin, D. , Everitt, BJ (2019) การดื่มเครื่องดื่มที่มีแอลกอฮอล์เป็นผลมาจากการล้มเหลวในการควบคุมพฤติกรรมการแสดงออกของ dorsolateral striatal J. Neurosci 39 (9), 1744–1754 ดอย: 10.1523 / JNEUROSCI.2615-18.2018
Goto, Y. , Grace, AA (2008) การประมวลผลข้อมูล Limbic และเยื่อหุ้มสมองในนิวเคลียส accumbens Trends Neurosci 31, 552 – 558 doi: 10.1016 / j.tins.2008.08.002
เกรซ, AA, Bunney, BS (1980) เซลล์ประสาทโดปามีนแบบ Nigral: การบันทึกและการระบุภายในเซลล์ด้วยการฉีด L-dopa และฮิสโตฟลูออเรสเซนต์ วิทยาศาสตร์ 210, 654 – 656 ดอย: 10.1126 / วิทยาศาสตร์. 7433992
สีเทา, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999) Agonist κ-opioid, U-69593, ลดพฤติกรรมแอมเฟตามีนเฉียบพลันและระดับ dialysate ที่ขึ้นกับแคลเซียมของโดปามีนและกลูตาเมตในระดับท้อง J. Neurochem 73, 1066 – 1074 doi: 10.1046 / j.1471-4159.1999.0731066.x
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013) ตัวรับแคปปา opioid ควบคุมการหาโคเคนที่เกิดจากความเครียดและความเป็นพลาสติก เซลล์ประสาท 77, 942 – 954 doi: 10.1016 / j.neuron.2012.12.034
Heidbreder, CA, Shippenberg, TS (1994) U-69593 ป้องกันไม่ให้เกิดอาการแพ้โคเคนโดยการทำให้ปกติโดปามีนเป็นมาตรฐาน Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Hernandez-Avila, CA 1. , Rounsaville, BJ, Kranzler, HR (2004) Opioid- ผู้หญิงที่ติดกัญชาและแอลกอฮอล์แสดงให้เห็นถึงความก้าวหน้าอย่างรวดเร็วของการใช้สารเสพติด ยาเสพติดแอลกอฮอล์ขึ้นอยู่กับ 74 (3), 265–272 doi: 10.1016 / j.drugalcdep.2004.02.001
Hoffman, AF, Spivak, CE, Lupica, CR (2016) การปลดปล่อยโดปามีนที่ปรับปรุงแล้วโดยตัวยับยั้งการขนส่งโดพามีนซึ่งอธิบายโดยรูปแบบการแพร่แบบ จำกัด และการวัดโวลแทมเมทรีแบบวงจรที่สแกนอย่างรวดเร็ว ACS Chem Neurosci 7, 700–709 ดอย: 10.1021 / acschemneuro.5b00277
โฮลเดน, C. (2001) “ พฤติกรรม” การเสพติด: พวกมันมีอยู่จริงหรือไม่? วิทยาศาสตร์ 294, 980 – 982 ดอย: 10.1126 / วิทยาศาสตร์. 294.5544.980
Imperato, A. , Di Chiara, G. (1988) ผลของการใช้ตัวรับ D-1 และ D-2 agonists และคู่อริที่ศึกษาด้วยการล้างไตทางสมอง Eur J. Pharmacol 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013) ผลของคูปปา opioid ตัวรับศัตรู, norbinaltorphimine, ความเครียดและการคืนสถานะยาเสพติดของการตั้งค่าสถานที่นิโคตินในหนู. Psychopharmacol (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B. , Uzelac, Z. , Sundaramurthy, S. , Rajamanickam, J. , Ewald, A. , Chefer, V. , et al. (2014) Salvinorin A ควบคุมฟังก์ชั่นการขนส่งโดพามีนผ่านทางตัวรับ kappa opioid และกลไกการพึ่งพา ERK1 / 2 Neuropharmacology 86, 228 – 240 ดอย: 10.1016 / j.neuropharm.2014.07.016
Koeltzow, TE, Austin, JD, Vezina, P. (2003) ความไวต่อพฤติกรรมต่อ quinpirole ไม่เกี่ยวข้องกับการเพิ่มขึ้นของนิวเคลียส accumbens โดพามีนล้น Neuropharmacology 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Koob, GF, Volkow, ND (2016) ชีววิทยาของการเสพติด: การวิเคราะห์ระบบประสาท มีดหมอจิตเวช 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Koob, GF (2013) ติดยาเสพติดคือการขาดดุลผลตอบแทนและความเครียดที่ผิดปกติ ด้านหน้า จิตเวช 4, 72. doi: 10.3389 / fpsyt.2013.00072
Labad, J. 1. , Menchon, JM, Alonso, P. , Segalas, C. , Jimenez, S. , Jaurrieta, N. , et al. (2008) ความแตกต่างระหว่างเพศในมิติอาการครอบงำ กดดันความวิตกกังวล 25 (10), 832–838 ดอย: 10.1002 / da.20332
Maisonneuve, IM, Archer, S. , Glick, SD (1994) U50,488 ซึ่งเป็นตัวรับแคปปา opioid ตัวเอกลดการเพิ่มขึ้นของโคเคนที่เกิดขึ้นในโดปามีนนอกเซลล์ในนิวเคลียส accumbens ของหนู Neurosci เลทท์ 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Mansour, A. , Fox, CA, Akil, H. , Watson, SJ (1995) Opioid-receptor แสดงออก mRNA ในหนู CNS: ผลกระทบทางกายวิภาคและการทำงาน Trends Neurosci 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Mansour, A. , Burke, S. , Pavlic, RJ, Akil, H. , Watson, SJ (1996) การหาตำแหน่งทางอิมมูโนวิทยาทางเคมีของตัวรับคัปปาโคลน 1 ในระบบประสาทส่วนกลางของหนูและต่อมใต้สมอง Neuroscience 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Margolis, EB, Karkhanis, AN (2019) Dopaminergic การมีส่วนร่วมของเซลล์และวงจรเพื่อความเกลียดชังผู้รับ Kappa opioid Neurochem int 129, 104504. ดอย: 10.1016 / j.neuint.2019.104504
Margolis, EB, Hjelmstad, GO, Bonci, A. , ทุ่ง, HL (2003) ผู้ชำนาญการ Kappa-opioid ยับยั้งเซลล์ประสาท dopaminergic midbrain โดยตรง J. Neurosci 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Margolis, EB, Hjelmstad, GO, Bonci, A. , ทุ่ง, HL (2005) Agonists ทั้ง Kappa และ Mu Opioid ยับยั้งการป้อน Glutamatergic ไปยัง Neurons Area Ventral Tegmental J. Neurophysiol 93, 3086 – 3093 doi: 10.1152 / jn.00855.2004
Margolis, EB, Lock, H. , Chefer, VI, Shippenberg, TS, Hjelmstad, GO, ทุ่งนา, HL (2006) Kappa opioids คัดเลือกควบคุมเซลล์ประสาทโดปามีนที่ฉายไปที่เยื่อหุ้มสมองส่วนหน้า พร Natl Acad วิทย์ สหรัฐอเมริกา 103, 2938 – 2942 doi: 10.1073 / pnas.0511159103
Margolis, EB, Mitchell, JM, Ishikawa, J. , Hjelmstad, GO, Fields, HL (2008) เซลล์ประสาทโดปามีนในสมองส่วนกลาง: เป้าหมายการฉายกำหนดระยะเวลาการกระทำที่อาจเกิดขึ้นและการยับยั้งโดปามีน D (2) J. Neurosci 28, 8908 – 8913 doi: 10.1523 / JNEUROSCI.1526-08.2008
Mathews, IZ, McCormick, CM (2007) หนูตัวเมียและตัวผู้ในวัยรุ่นตอนปลายนั้นแตกต่างจากผู้ใหญ่ในการเคลื่อนไหวของแอมเฟตามีนที่เกิดจากแอมเฟตามีน Behav Pharmacol 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Mathis, MA 1. , Pd, A. , Funaro, G. , RC, T. , Moraes, I. , AR, T. , et al. (2011) ความแตกต่างระหว่างเพศในโรคที่ครอบงำ - บังคับ: การทบทวนวรรณกรรม Braz เจจิตเวช 33 (4), 390–399 ดอย: 10.1590 / S1516-44462011000400014
McLaughlin, JP, Marton-Popovici, M. , Chavkin, C. (2003) Kappa opioid ตัวรับการเป็นปรปักษ์กันและการหยุดชะงักของยีน prodynorphin บล็อกการตอบสนองพฤติกรรมที่เกิดความเครียด J. Neurosci 23 (13), 5674–5683 ดอย: 10.1523 / JNEUROSCI.23-13-05674.2003
McLaughlin, JP, Land, BB, Li, S. , Pintar, JE, Chavkin, C. (2006a) การเปิดใช้งานก่อนหน้าของผู้รับ Kappa opioid โดย U50,488 เลียนแบบบังคับซ้ำความเครียดว่ายน้ำเพื่อปรับสภาพเครื่องโคเคนที่มีศักยภาพ Neuropsychopharmacology 31, 787 – 794 doi: 10.1038 / sj.npp.1300860
McLaughlin, JP, Li, S. , วาลเดซ, เจ, Chavkin, TA, Chavkin, C. (2006b) การตอบสนองเชิงพฤติกรรมที่เกิดจากความเครียดจากการพ่ายแพ้ทางสังคมนั้นถูกสื่อกลางโดยระบบ kappa opioid จากภายนอก Neuropsychopharmacology 31 (6), 1241–1248 ดอย: 10.1038 / sj.npp.1300872
Milesi-Hallé, A. , McMillan, DE, Laurenzana, EM, เบิร์นส์ - เบลค, KA, Owens, SM (2007) ความแตกต่างทางเพศใน (+) - แอมเฟตามีน - และ (+) - การตอบสนองเชิงพฤติกรรมของแอมเฟตตามินในหนูที่เป็นเพศชายและเพศเมีย Pharmacol Biochem Behav 86, 140 – 149 doi: 10.1016 / j.pbb.2006.12.018
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997) ผลของคัปปาโอปิออยด์ต่อการจัดการตนเองของโคเคนโดยลิงจำพวก J. Pharmacol ประสบการณ์ Ther 282, 44-55
Nestler, EJ (2001) พื้นฐานระดับโมเลกุลของการติดอยู่กับพลาสติกในระยะยาว ชัยนาท รายได้ Neurosci 2, 119 – 128 doi: 10.1038 / 35053570
Perreault, ML, Graham, D. , Bisnaire, L. , Simms, J. , Hayton, S. , Szechtman, H. (2006) Kappa-opioid agonist U69593 การเคลื่อนที่ของโพเทนชิโอโพเทนซีสของการทำให้ไวต่อยา D2 / D3 agonist quinpirole: กลไกแบบ pre- และ postynaptic Neuropsychopharmacology 31, 1967 – 1981 doi: 10.1038 / sj.npp.1300938
Perreault, ML, Seeman, P. , Szechtman, H. (2007) การกระตุ้นตัวรับ Kappa-opioid ช่วยเร่งการเกิดโรคของการตรวจสอบแบบบังคับในรูปแบบการกระตุ้นอาการแพ้ของ quinpirole ของโรค obsessive-compulsive Behav Neurosci 121, 976 – 991 ดอย: 10.1037 / 0735-7044.121.5.976
Pierce, RC, Kalivas, PW (1997) แบบจำลองวงจรของการแสดงออกของความไวต่อพฤติกรรมต่อแอมเฟตามีนที่มีลักษณะคล้ายจิต ความต้านทานของสมอง ความต้านทานของสมอง รายได้ 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Polter, AM, Bishop, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014) บล็อกหลังคลอดของผู้รับสารคัปปาโอปิอิดช่วยเพิ่มศักยภาพในระยะยาวของการยับยั้งและยับยั้งการคืนสถานะการแสวงหาโคเคน Biol จิตเวช 76, 785 – 793 doi: 10.1016 / j.biopsych.2014.04.019
Puig-Ramos, A. , Santiago, GS, Segarra, AC (2008) U-69593 ซึ่งเป็นตัวรับตัวรับแคปปา opioid ลดอาการแพ้ที่เกิดจากพฤติกรรมโคเคนในหนูตัวเมีย Behav Neurosci 122, 151 – 160 ดอย: 10.1037 / 0735-7044.122.1.151
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S. , Ersche, KD (2012) เอนโดฟีโนไทป์ของระบบประสาทและแรงกระตุ้น: ต่อจิตเวชมิติ Trends Cognit วิทย์ 16, 81 – 91 doi: 10.1016 / j.tics.2011.11.009
โรเบิร์ตส์ DCS เบนเน็ตต์ขายวิคเกอร์ GJ (1989) วงจรการเป็นสัดส่งผลกระทบต่อการจัดการตนเองของโคเคนในตารางอัตราความก้าวหน้าในหนู Psychopharmacol (Berl) 98, 408 – 411 doi: 10.1007 / BF00451696
โรบินสัน, TE, Berridge, KC (1993) พื้นฐานทางประสาทของความอยากติดยา: ทฤษฎีการกระตุ้นให้ติดยาเสพติด ความต้านทานของสมอง ความต้านทานของสมอง รายได้ 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
โรบินสัน, TE, Berridge, KC (2001) การกระตุ้นและการเสพติด ติดยาเสพติด 96, 103 – 114 doi: 10.1046 / j.1360-0443.2001.9611038.x
Rose, JH, Karkhanis, AN, Steiniger-Brach, B. , Jones, SR (2016) ผลกระทบที่แตกต่างของ Nalmefene ต่ออัตราการดูดโดปามีนและกิจกรรมของตัวรับแคปปาโอปิออยด์ในนิวเคลียส int J. Mol. วิทย์ 17, 1216. ดอย: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr. , Park, PE, Crawford, EF, George, O. , Vendruscolo, LF, et al. (2013) การเป็นปรปักษ์กันในระยะยาวของตัวรับκ opioid ป้องกันการเพิ่มและแรงจูงใจที่เพิ่มขึ้นสำหรับการบริโภคเฮโรอีน J. Neurosci 33 (49), 19384–19392 ดอย: 10.1523 / JNEUROSCI.1979-13.2013
Sesack, SR, Pickel, VM (1992) ultrastructural การแปลคู่ของ enkephalin และ tyrosine hydroxylase immunoreactivity ในพื้นที่หน้าท้องหนู tegmental: สารตั้งต้นหลายสำหรับปฏิกิริยา opiate-dopamine J. Neurosci 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Sesack, SR, Aoki, C. , Pickel, VM (1994) การแปลโครงสร้างของ ultrasructural เหมือน immunoreactivity รับ D2 ในเซลล์ประสาทโดปามีนในสมองส่วนกลางและเป้าหมายของพวกเขา J. Neurosci 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Sesia, T. , Bizup, B. , Grace, AA (2013) การประเมินรูปแบบสัตว์ของโรคย้ำคิดย้ำทำ: ความสัมพันธ์กับกิจกรรมของเซลล์ประสาทโดปามีน int J. Neuropsychopharmacol 16, 1295–1307 ดอย: 10.1017 / S146114571200154X
Shippenberg, TS, Zapata, A. , Chefer, VI (2007) Dynorphin และพยาธิสรีรวิทยาของการติดยา Pharmacol Ther 116, 306–321 ดอย: 10.1016 / j.pharmthera.2007.06.011
สมิ ธ , JA, Loughlin, SE, เลสลี่, FM (1992) การยับยั้ง kappa-Opioid ของการปล่อยโดปามีน [3H] จากการเพาะเลี้ยงเซลล์ที่แยกตัวออกจากเซลล์ mesencephalic mesencephalic mol Pharmacol 42.
Smith, JW, Fetsko, LA, Xu, R. , Wang, Y. (2002) หนูที่ได้รับ Dopamine D2L สิ่งที่น่าพิศวงแสดงการขาดดุลในคุณสมบัติเสริมแรงเชิงบวกและเชิงลบของมอร์ฟีนและในการเรียนรู้การหลีกเลี่ยง Neuroscience 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Smith, JS, Schindler, AG, Martinelli, E. , Gustin, RM, Bruchas, MR, Chavkin, C. (2012) การเปิดใช้งานที่เกิดจากความเครียดของระบบตัวรับ dynorphin / κ-opioid ใน amygdala potentiates การตั้งค่าสถานที่นิโคติน J. Neurosci 32, 1488 – 1495 doi: 10.1523 / JNEUROSCI.2980-11.2012
Spanagel, R. , Herz, A. , Shippenberg, TS (1992) ซึ่งตรงข้ามกับระบบ opioid ภายนอกที่ใช้งานอยู่ในปัจจุบันจะปรับเส้นทางของโดปามีน พร Natl Acad วิทย์ 89, 2046 – 2050 doi: 10.1073 / pnas.89.6.2046
พิเศษ SG, Manaye, KF, Sadeq, M. , เยอรมัน, DC (1993) ตัวรับ Opioid ในบริเวณโดปามีนในสมองส่วนกลางของหนู ครั้งที่สอง อัตชีวประวัติแคปปาและเดลต้ารับ J. Neural Transm พลเอก 91, 53 – 66 doi: 10.1007 / BF01244918
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010) การไกล่เกลี่ย kappa-opioid จากภายนอกของศักยภาพที่เกิดจากความเครียดของการตั้งค่าสถานที่เอทานอลปรับอากาศและการบริหารตนเอง Psychopharmacol (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011) ยาที่ต้องการ: อาการไวต่อพฤติกรรมและการกำเริบของพฤติกรรมการแสวงหาสิ่งเสพติด DR, ed. Pharmacol รายได้ 63, 348–365 ดอย: 10.1124 / pr.109.001933
Stuchlik, A. , Radostová, D. , Hatalova, H. , Vales, K. , Nekovarova, T. , Koprivova, J. , et al. (2016) ความถูกต้องของแบบจำลองการก่อให้เกิดอาการแพ้ของ quinpirole ของ OCD: การเชื่อมโยงหลักฐานจากการศึกษาสัตว์และคลินิก ด้านหน้า Behav Neurosci 10, 209. ดอย: 10.3389 / fnbeh.2016.00209
Sun, T. , Song, Z. , Tian, Y. , Tian, W. , Zhu, C. , Ji, G. , et al. (2019) basalateral amygdala ที่ป้อนไปยังเยื่อหุ้มสมอง prefrontal อยู่ตรงกลางควบคุมพฤติกรรมการตรวจสอบความผิดปกติเหมือนครอบงำครอบงำ พร Natl Acad วิทย์ สหรัฐอเมริกา 116, 3799 – 3804 doi: 10.1073 / pnas.1814292116
Svingos, AL, Chavkin, C. , Colago, EEO, Pickel, VM (2001) การแสดงออกที่สำคัญของตัวรับ opioid และตัวขนส่ง dopamine ในนิวเคลียส accumbens รูปโปรไฟล์ axonal ไซแนปส์ 42, 185 – 192 doi: 10.1002 / syn.10005
Szechtman, H. , Talangbayan, H. , Eilam, D. (1993) องค์ประกอบด้านสิ่งแวดล้อมและพฤติกรรมของการกระตุ้นอาการแพ้ที่เกิดจากโดปามีน quonpirole ตัวเอก Behav Pharmacol 4, 405–410. doi: 10.1097/00008877-199308000-00014
Szechtman, H. , Talangbayan, H. , Canaran, G. , Dai, H. , Eilam, D. (1994) การเปลี่ยนแปลงของความไวต่อพฤติกรรมที่เกิดจากโดปามีนตัวเอก quinpirole และกลไกการควบคุมพลังงานกลางที่เสนอ Psychopharmacol (Berl) 115, 95 – 104 doi: 10.1007 / BF02244757
Szechtman, H. , Sulis, W. , Eilam, D. (1998) Quinpirole ชักนำให้เกิดพฤติกรรมการตรวจสอบบีบบังคับในหนู: แบบจำลองสัตว์ที่มีศักยภาพของโรค obsessive-compulsive (OCD) Behav Neurosci 112, 1475 – 1485 ดอย: 10.1037 / 0735-7044.112.6.1475
Szechtman, H. , Culver, K. , Eilam, D. (1999) บทบาทของระบบโดปามีนในความผิดปกติที่ครอบงำ (OCD): ผลกระทบจากแบบจำลองสัตว์ที่กระตุ้นด้วยสมอง Pol เจ Pharmacol 51, 55-61
Szechtman, H. , Eckert, MJ, Tse, WS, Boersma, JT, Bonura, C a, JZ, M. , et al. (2001) พฤติกรรมการตรวจสอบบีบบังคับของหนูที่ได้รับไว quinpirole ในรูปแบบสัตว์ของ Obsessive-Compulsive Disorder (OCD): รูปแบบและการควบคุม BMC Neurosci 2, 4. doi: 10.1186/1471-2202-2-4
Szechtman, H. , Ahmari, SE, Beninger, RJ, Eilam, D. , Harvey, BH, Edemann-Callesen, H. , et al. (2017) ความผิดปกติที่ครอบงำ - บังคับ: ข้อมูลเชิงลึกจากแบบจำลองสัตว์ Neurosci Biobehav รายได้ 76, 254 – 279 doi: 10.1016 / j.neubiorev.2016.04.019
Tejeda, HA, Bonci, A. (2019) การควบคุมตัวรับไดปามีน / คัปปา - opioid ของพลศาสตร์โดปามีน: ผลกระทบต่อสภาวะอารมณ์เชิงลบและความผิดปกติทางจิตเวช สมอง Res 1713, 91 – 101 doi: 10.1016 / j.brainres.2018.09.023
Tejeda, HA, Wu, J. , Kornspun, AR, Pignatelli, M. , Kashtelyan, V. , Krashes, M. J. , et al. (2017). การปรับตัวรับคัปปา - โอปิออยด์เฉพาะทางและเซลล์ของการปรับสมดุลการยับยั้งการกระตุ้นและการกระตุ้นที่แตกต่างกันประตู d1 และ d2 ทำให้เกิดการทำงานของเซลล์ประสาท เซลล์ประสาท 93 (1), 147–163 ดอย: 10.1016 / j.neuron.2016.12.005
Thompson, AC, Zapata, A. , Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000) การเปิดใช้งานตัวรับ Kappa-opioid จะปรับเปลี่ยนการดูดซึมโดปามีนในนิวเคลียส accumbens และต่อต้านผลกระทบของโคเคน J. Neurosci 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Thompson, D. , Martini, L. , Whistler, JL (2010) อัตราส่วนการเปลี่ยนแปลงของตัวรับโดปามีน D1 และ D2 ใน striatum ของเมาส์นั้นสัมพันธ์กับการตอบสนองต่อพฤติกรรมที่ไวต่อโคเคน PloS One 5, e11038 doi: 10.1371 / journal.pone.0011038
Usiello, A. , Baik, J.-H. , Rougé-Pont, F. , Picetti, R. , Dierich, A. , LeMeur, M. , et al. (2000) ฟังก์ชั่นที่แตกต่างของไอโซฟอร์มสองตัวของตัวรับโดปามีน D2 ธรรมชาติ 408, 199 – 203 doi: 10.1038 / 35041572
Van Bockstaele, EJ, Pickel, VM (1995) เซลล์ประสาทที่มี GABA ในโครงการพื้นที่หน้าท้องเพื่อนิวเคลียส accumbens ในสมองหนู สมอง Res 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Van't Veer, A. , Bechtholt, AJ, Onvani, S. , Potter, D. , Wang, Y. , Liu-Chen, LY และอื่น ๆ (2013). การระเหยของตัวรับแคปปา - โอปิออยด์จากเซลล์ประสาทโดปามีนในสมองมีฤทธิ์คล้ายกับโรควิตกกังวลและช่วยเพิ่มความเป็นพลาสติกที่เกิดจากโคเคน Neuropsychopharmacology 38 (8), 1585–1597 ดอย: 10.1038 / npp.2013.58
Vanderschuren, LJ, Kalivas, PW (2000) การเปลี่ยนแปลงในการส่งโดปามินเนอร์จิคและกลูตามาเทอจิคในการเหนี่ยวนำและการแสดงออกของการกระตุ้นให้เกิดพฤติกรรม: การทบทวนที่สำคัญของการศึกษาพรีคลินิก Psychopharmacol (Berl) 151 (2-3), 99–120 ดอย: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017) ระบบแรงกระตุ้นโดปามีน: ผลกระทบของการติดยาและอาหาร ชัยนาท รายได้ Neurosci 18, 741–752 ดอย: 10.1038 / nrn.2017.130
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000) โดปามีนที่ปล่อยออกมาและการดูดซึมจะดีกว่าในตัวเมียมากกว่าตัวผู้หนู striatum ซึ่งวัดโดยโวลแทมเมทรีแบบรวดเร็ว Neuroscience 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Wee, S. , Koob, GF (2010) บทบาทของระบบ dynorphin – κ opioid ในการเสริมผลกระทบของยาเสพติด Psychopharmacol (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S. , Orio, L. , Ghirmai, S. , Cashman, JR, Koob, GF (2009) การยับยั้งตัวรับแคปปา opioid ลดปริมาณโคเคนที่เพิ่มขึ้นในหนูที่มีการเข้าถึงโคเคนเพิ่มขึ้น Psychopharmacol (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr. , Schlosburg, JE, Wee, S. , Gould, A. , George, O. , Grant, Y. , et al. (2015) κตัวรับ Opioid ในนิวเคลียส accumbens เปลือกเป็นสื่อกลางในการเพิ่มการบริโภคยาบ้า J. Neurosci 35 (10), 4296–4305 ดอย: 10.1523 / JNEUROSCI.1978-13.2015
Williams, MT, Mugno, B. , Franklin, M. , Faber, S. (2013) มิติอาการของโรคที่เกิดจากการครอบงำ: ปรากฏการณ์และผลการรักษาด้วยการเปิดรับและการป้องกันพิธีกรรม พยาธิวิทยา 46, 365 – 376 doi: 10.1159 / 000348582
วิลสัน, CJ, หนุ่มสาว, SJ, โกรฟ, PM (1977) คุณสมบัติทางสถิติของเซลล์ประสาทเส้นประสาทรถไฟใน substantia nigra: ชนิดของเซลล์และปฏิกิริยาระหว่างกัน สมอง Res 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
ปรีชาญาณ RA (2009) บทบาทของ nigrostriatal - ไม่ใช่แค่ mesocorticolimbic - โดปามีนในการให้รางวัลและการเสพติด Trends Neurosci 32, 517 – 524 doi: 10.1016 / j.tins.2009.06.004
Xiao, L. , Becker, JB (1994) การตรวจวิเคราะห์เชิงปริมาณของความเข้มข้นของสารโดปามีนนอกเซลล์นอกเซลล์ในหนูขาวและหนูเพศเมีย: ผลกระทบของรอบการเป็นสัดและการทำศัลยกรรมอวัยวะสืบพันธุ์. Neurosci เลทท์ 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
คำสำคัญ: ตัวรับแคปปา opioid, โดปามีน, ยา, แอมเฟตามีน, quinpirole, อาการแพ้หัวรถจักร
การอ้างอิง: Escobar AdP, Casanova JP, Andrés ME และ Fuentealba JA (2020) Crosstalk ระหว่าง Kappa Opioid และระบบ Dopamine ในพฤติกรรมบีบบังคับ ด้านหน้า Pharmacol 11: 57 doi: 10.3389 / fphar.2020.00057
ได้รับ: 16 ตุลาคม 2019; ยอมรับ: 22 มกราคม 2020;
เผยแพร่: 18 กุมภาพันธ์ 2020
แก้ไขโดย:
กอนซาโล่อี. อีเว่นมหาวิทยาลัยคอนเซปชั่นชิลี
บทวิจารณ์โดย:
หลุยส์ เจราร์โด อกัวโยมหาวิทยาลัยคอนเซปชั่นชิลี
ฮูโก้ เตเจด้า, สถาบันยาเสพติดแห่งชาติ (NIDA), สหรัฐอเมริกา
เซซิเลีย สกอร์ซ่า, Instituto de Investigaciones Biológicas Clemente Estable (IIBCE), อุรุกวัย
ลิขสิทธิ์© 2020 Escobar, Casanova, Andrésและ Fuentealba นี่เป็นบทความแบบเปิดที่เผยแพร่ภายใต้เงื่อนไขของ ใบอนุญาตแสดงที่มาของครีเอทีฟคอมมอนส์ (CC BY). อนุญาตให้ใช้งานแจกจ่ายหรือทำซ้ำในฟอรัมอื่นโดยผู้แต่งดั้งเดิมและเจ้าของลิขสิทธิ์จะได้รับเครดิตและมีการอ้างถึงการตีพิมพ์ต้นฉบับในวารสารนี้ตามแนวทางปฏิบัติทางวิชาการที่ได้รับการยอมรับ ไม่อนุญาตให้ใช้แจกจ่ายหรือทำซ้ำซึ่งไม่เป็นไปตามข้อกำหนดเหล่านี้
* จดหมายโต้ตอบ: José Antonio Fuentealba, [ป้องกันอีเมล]
†ORCID: José Antonio Fuentealba ออร์ซิด.org/0000-0003-0775-0675