 Fabrizio Sanna1*,
Fabrizio Sanna1*,  Jessica Bratzu1,
Jessica Bratzu1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melis1,
Maria R. Melis1,  Osvaldo Giorgi2 และ
Osvaldo Giorgi2 และ  อันโตนิโอ1,3
อันโตนิโอ1,3- 1ภาควิชาวิทยาศาสตร์ชีวการแพทย์, แผนกประสาทวิทยาและเภสัชวิทยาคลินิก, และศูนย์ความเป็นเลิศด้านชีววิทยาของการเสพติด, มหาวิทยาลัย Cagliari, กาลยารี, อิตาลี
- 2ภาควิชาวิทยาศาสตร์สิ่งมีชีวิตและสิ่งแวดล้อม, แผนกเภสัชศาสตร์, เภสัชวิทยาและวิทยาศาสตร์เภสัชศาสตร์, มหาวิทยาลัย Cagliari, Cagliari, อิตาลี
- 3สถาบันประสาท, สภาวิจัยแห่งชาติ, แผนกกาลยารี, Cittadella Universitaria, กาลยารี, อิตาลี
Roman High- (RHA) และ Low-Avoidance (RLA) outbred rats ซึ่งแตกต่างกันไปตามลำดับอย่างรวดเร็วเมื่อเทียบกับการได้มาซึ่งการตอบสนองต่อการหลีกเลี่ยงการใช้งานที่ไม่ดีในกล่องกระสวยแสดงความแตกต่างในกิจกรรมทางเพศ หนูหญิงเปิดกว้าง อันที่จริงหนู RHA แสดงระดับของแรงจูงใจทางเพศและการมีเพศสัมพันธ์สูงกว่าหนู RLA ซึ่งยังคงมีอยู่หลังจากกิจกรรมทางเพศซ้ำ ๆ ความแตกต่างเหล่านี้มีความสัมพันธ์กับระดับเสียงที่สูงขึ้นของระบบ dopaminergic mesolimbic ของหนู RHA เทียบกับหนู RLA เปิดเผยโดยการเพิ่มขึ้นของโดปามีนที่พบใน dialysate ที่ได้รับจากนิวเคลียส accumbens ของ RHA มากกว่าหนู RLA ในช่วงกิจกรรมทางเพศ งานนี้แสดงให้เห็นว่าโดปามีนนอกเซลล์และนอเรนดาลีน (NA) นอกเซลล์เพิ่มขึ้นใน dialysate จากเยื่อหุ้มสมอง prefrontal cortex (mPFC) ที่อยู่ตรงกลางของหนู RHA และ RLA เพศชายวางต่อหน้าหนูเพศเมียที่ไม่สามารถเข้าถึงได้ การเพิ่มขึ้นของโดปามีน (และเมตาโบไลต์หลัก 3,4-dihydroxyphenylacetic acid, DOPAC) และ NA นั้นพบได้ในสัตว์ที่ไร้สมรรถภาพทางเพศและสัตว์ที่มีประสบการณ์ แต่พวกมันสูงกว่า: (i) ใน RHA มากกว่าหนู RLA; และ (ii) ในหนูที่มีประสบการณ์ทางเพศสัมพันธ์ของ RHA และ RLA มากกว่าหนูที่ไร้เดียงสาของพวกเขา ในที่สุดความแตกต่างของโดปามีนและ NA ใน mPFC นั้นเกิดขึ้นพร้อมกันกับกิจกรรมทางเพศเนื่องจากหนู RHA แสดงระดับของแรงจูงใจทางเพศและการมีเพศสัมพันธ์ในระดับที่สูงกว่าหนู RLA ทั้งในเพศและสภาพที่มีประสบการณ์ ผลลัพธ์เหล่านี้ชี้ให้เห็นว่าเสียงโดปามีนที่สูงขึ้นนั้นเกิดขึ้นใน mPFC พร้อมกับเสียงนอร์ดเรนเนอร์จิคที่เพิ่มขึ้นซึ่งอาจมีส่วนร่วมในรูปแบบ copulatory ต่างๆที่พบในหนู RHA และ RLA
บทนำ
โรมันหนูสูง - (RHA) และเส้นหลีกเลี่ยง (RLA) outbred หนูสายเดิมเลือกอย่างรวดเร็วตามลำดับเมื่อเทียบกับการเข้าซื้อกิจการที่ไม่ดีของการตอบสนองการหลีกเลี่ยงการใช้งานในกล่องกระสวย (Bignami, 1965; Broadhurst และ Bignami, 1965; Driscoll และBättig, 1982; Fernández-Teruel และคณะ, 2002; Giorgi et al., 2007) แสดงความแตกต่างอย่างมีนัยสำคัญในกิจกรรมทางเพศเมื่อมีหนูเพศเมียเปิดกว้าง อันที่จริงแล้วหนู RHA แสดงแรงจูงใจทางเพศในระดับที่สูงกว่าหนู RLA ที่ได้รับการเปิดเผยจากจำนวนอวัยวะเพศชายที่ไม่แสดงการสัมผัสอวัยวะเพศสัมพันธ์ที่แสดงโดยหนูเหล่านี้เมื่อใส่ตัวเมียที่ไม่สามารถเข้าถึงได้และการแสดงร่วมที่ดีกว่าหนู RLA โดยความแตกต่างอย่างมีนัยสำคัญในพารามิเตอร์ copulatory จำนวนมากที่พบส่วนใหญ่ในการทดสอบ copulatory ครั้งแรกระหว่างสองสายหนู แต่ยังหลังจากห้าการทดสอบ copulatory แม้ว่าลดทอน (Sanna และคณะ, 2014a) ในบรรดาความแตกต่างที่เด่นชัดที่สุดที่พบระหว่างหนู RHA และ RLA คือเปอร์เซ็นต์ของหนูที่เริ่มต้นการติดตั้งและ intromitting และอุทานในการทดสอบ copulatory ครั้งแรก, 80% ของหนู RHA เทียบกับหนู 40% ของ RLA ตามลำดับ (Sanna และคณะ, 2014a) หลักฐานการทดลองขนาดใหญ่แสดงให้เห็นว่าน้ำย่อยที่มีหน้าที่แตกต่างกันมีส่วนร่วมในความแตกต่างระหว่าง RHA และ RLA หนู ในความเป็นจริงรูปแบบ copulatory ที่แตกต่างกันของ RHA และ RLA หนูถูกพบว่ามีการแก้ไขแตกต่างกันโดย apomorphine, ผสม D1 / D2 ตัวรับโดปามีน agonist บริหารงานในปริมาณที่อำนวยความสะดวกพฤติกรรมทางเพศและ haloperidol, D2 dopamine receptor antagonist ปริมาณต่ำที่ยับยั้งพฤติกรรมทางเพศ (Sanna และคณะ, 2014b) ดังนั้นพบว่าหนู RLA มีความไวต่อการอำนวยความสะดวกและการยับยั้งพฤติกรรมทางเพศที่เกิดจาก apomorphine และ haloperidol ตามลำดับซึ่งแสดงให้เห็นถึงการปรับเปลี่ยนที่มากขึ้นของยาสองตัวส่วนใหญ่เมื่อได้รับยาในปริมาณต่ำในพารามิเตอร์ copulatory ต่างๆ หนู RLA ด้วยความเคารพหนู RHA (Sanna และคณะ, 2014b) บางทีอาจมีความสำคัญมากกว่าสำหรับการศึกษานี้รูปแบบ copulatory ที่แตกต่างกันของ RHA และ RLA หนูยังเกี่ยวข้องกับความแตกต่างในกิจกรรมของเซลล์ประสาท dopaminergic mesolimbic ซึ่งกิจกรรมเป็นที่รู้จักกันดีในการเพิ่มพฤติกรรมทางเพศ (Pfaus et al., 1990; Pleim et al., 1990; Pfaus และ Phillips, 1991; Damsma et al., 1992; Wenkstern และคณะ, 1993; Balfour et al., 2004; Pitchers และคณะ, 2010, 2013; เบโลอาเตและคณะ 2016) อันที่จริงแล้วทั้งๆที่หนูที่ไร้สมรรถภาพทางเพศและมีประสบการณ์ทางเพศ RHA และ RLA หนูมีค่าฐานที่คล้ายกันของ extracellular dopamine ใน dialysate จากนิวเคลียส accumbens, ความเข้มข้นของ dopamine extracellular และ 3,4-dihydroxyphenylacetic acid (DOPAC) ) พบว่ามีการเพิ่มขึ้นแตกต่างกันใน dialysates ที่ได้รับจากนิวเคลียส accumbens เปลือกของหนูที่ไร้เดียงสาและมีประสบการณ์ทางเพศ RHA และ RLA หนูทั้งในระยะที่คาดการณ์และบริโภคของพฤติกรรมทางเพศ ความแตกต่างดังกล่าวข้างต้นมีความชัดเจนมากขึ้นระหว่างหนูที่มีเชื้อเอชไอวีและหนู RLA แต่ยังคงมีอยู่ระหว่างหนูที่มีประสบการณ์ทางเพศกับหนูอาร์เอชเอและอาร์แอลเอแม้ว่าจะมีแนวโน้มลดลงในหนูเหล่านี้Sanna และคณะ, 2015).
บทบาทการทำงานของการเพิ่มขึ้นของ dopaminergic ในนิวเคลียส accumbens ที่เห็นในระหว่างกิจกรรมทางเพศ (ทั้งในขั้นตอนของพฤติกรรมทางเพศที่น่ากินและการบริโภค) ยังคงเป็นที่ถกเถียงกันเช่นเดียวกับที่เห็นในระหว่างการให้อาหารเช่นกิจกรรมทางเพศ สร้างแรงบันดาลใจจุ ดังนั้นแม้ว่าโดปามีนในนิวเคลียส accumbens มีส่วนร่วมในแรงจูงใจและเซลล์ประสาทโดปามีน mesolimbic มักจะเรียกว่าเซลล์ประสาทรางวัลในช่วง 15 ปีที่ผ่านมาการศึกษาล่าสุดสนับสนุนสมมติฐานที่โดปามีน mesolimbic ไม่ได้เกี่ยวข้องกับการแสดงออกหลักของพฤติกรรม แต่เป็นการเรียนรู้และความทรงจำเกี่ยวกับความสัมพันธ์กับสิ่งกระตุ้นAgmo et al., 1995; Berridge และ Robinson, 1998; Ikemoto และ Panksepp, 1999; Pitchers และคณะ, 2013, 2014; เบโลอาเตและคณะ 2016; ซาลาโมนและคณะ, 2016) สอดคล้องกับสมมติฐานนี้การปิดล้อมของตัวรับโดปามีนในนิวเคลียส accumbens หรือการใช้งานของเซลล์ประสาทโดปามิเนอร์จิคในพื้นที่หน้าท้องเมื่อไม่นานมานี้พบว่าไม่สามารถเปลี่ยนการแสดงออกของความอยากอาหารและการบริโภคของพฤติกรรมร่วมPitchers และคณะ, 2013, 2014; เบโลอาเตและคณะ 2016).
อย่างไรก็ตามบทบาทของพื้นที่สมองอื่น ๆ ที่มีโดปามีนในพฤติกรรมทางเพศที่แตกต่างกันระหว่าง RHA และ RLA ไม่สามารถตัดออกได้ ดังนั้นจึงเป็นที่ทราบกันดีว่าโดปามีนมีผลในการอำนวยความสะดวกต่อขั้นตอนการคาดการณ์และการบริโภคของพฤติกรรมทางเพศในสัตว์ทดลองและในมนุษย์ไม่เพียง แต่ในนิวเคลียส accumbens (Everitt, 1990; Pfaus et al., 1990; ฮัลล์และอัล 1991; Pfaus และ Everitt, 1995; Melis และ Argiolas, 2011) แต่ยังอยู่ในพื้นที่สมองอื่น ๆ เช่นพื้นที่ preoptic อยู่ตรงกลาง, มลรัฐและนิวเคลียสของมัน (เช่นนิวเคลียส paraventricular (PVN); Pfaus และ Phillips, 1991; Argiolas and Melis, 1995, 2005, 2013; ฮัลล์และอัล 1995, 1999; Melis และ Argiolas, 1995; Melis et al., 2003; Succu et al., 2007; Pfaus, 2010) อีกพื้นที่ที่มีโดปามีนและอาจมีบทบาทในพฤติกรรมทางเพศก็คือเยื่อหุ้มสมอง prefrontal cortex (mPFC; Fernández-Guasti และคณะ, 1994; Agmo และ Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama และคณะ, 2003; Balfour et al., 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011) ในความเป็นจริงในพื้นที่สมองนี้จะพบปลายประสาทของเซลล์ประสาทโดปามีน mesocortical กับร่างกายเซลล์ของพวกเขาในพื้นที่ tegmental หน้าท้องเป็นเซลล์ประสาทโดปามีน mesolimbic สำหรับนิวเคลียส accumbens บทบาทที่แน่นอนของพื้นที่สมองนี้ในพฤติกรรมทางเพศอยู่ไกลจากความชัดเจน ที่จริงแล้วรอยโรคของ mPFC นั้นมักจะไม่สามารถเปลี่ยนพฤติกรรมทางเพศของหนูเพศผู้กับเพศหญิงที่รับได้ทางเพศ (Fernández-Guasti และคณะ, 1994; Agmo และ Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama และคณะ, 2003; Balfour et al., 2006; Afonso et al., 2007) และการแสดงออกของการตั้งค่าสถานที่ปรับอากาศสำหรับรางวัลทางเพศ (Davis et al., 2010) อย่างไรก็ตามแผล mPFC ซึ่งไม่ได้เปลี่ยนรูปลักษณ์ของสถานที่ที่กำหนดไว้สำหรับรางวัลทางเพศยกเลิกในสัตว์เดียวกันความสามารถในการสร้างความเกลียดชังปรับอากาศต่อกิจกรรมทางเพศเมื่อจับคู่กับสิ่งเร้า aversive (Davis et al., 2010) และเซลล์ที่เลือกสรรในระหว่างการเข้าใกล้พฤติกรรมของหนูตัวผู้ที่มีต่อเพศหญิงที่ไม่สามารถเข้าถึงได้ถูกวัดใน mPFC ของหนูตัวผู้ (Febo, 2011) การค้นพบนี้นำไปสู่การแนะนำว่าการกระตุ้น mPFC ในระหว่างพฤติกรรมทางเพศมีบทบาทในการบูรณาการข้อมูลภายนอกและภายในสำหรับการดำเนินการและการควบคุมพฤติกรรมมุ่งเป้าหมายมากกว่าการแสดงออกของการตอบสนองโดยธรรมชาติต่อผู้สนับสนุนทางธรรมชาติ ไปที่และเกรซ 2005) ดังนั้นร่วมกับนิวเคลียส accumbens, mPFC เป็นส่วนหนึ่งของระบบประสาทที่ซับซ้อนที่เกี่ยวข้องในการปรับพฤติกรรมกระตุ้น (พฤติกรรมเป้าหมายกำกับ) ซึ่งต้องมีการบูรณาการข้อมูลความรู้จาก mPFC ข้อมูลทางอารมณ์จาก amygdala และบริบท - ข้อมูลที่เกี่ยวข้องจากฮิบโปแคมปัสในนิวเคลียส accumbens (ไปที่และเกรซ 2005).
หลักฐานจากการทดลองชี้ให้เห็นว่าการปล่อยโดปามีนใน mPFC นั้นมีส่วนเกี่ยวข้องกับการปรับพฤติกรรมกระตุ้นและการควบคุมกลไกเหล่านี้มีบทบาทในสภาพพยาธิสภาพหรือการปรับตัวที่ผิดปกติเช่นโรคทางจิตเวชเช่นโรคจิตเภท สมาธิสั้น) ภาวะซึมเศร้า (Dunlop และ Nemeroff, 2007; Masana และคณะ, 2011) หรือการใช้สารเสพติดและพฤติกรรมการพนัน (Everitt และ Robbins, 2005) สิ่งที่น่าสนใจคือกิจกรรมของโดปามีนที่ปล่อยใน mPFC อาจได้รับอิทธิพลจาก noradrenaline (NA) ซึ่งมีอยู่ใน mPFC ในระดับที่สูงกว่าโดปามีนและโดยเฉพาะอย่างยิ่งจาก NA transporter (NET) ซึ่งไม่เพียง แต่อุดมสมบูรณ์กว่า โดปามีนขนย้าย (DAT) ใน mPFC (Carboni และคณะ, 1990, 2006; Gresch et al., 1995; Westernik และคณะ, 1998) แต่ก็แสดงให้เห็นถึงความสัมพันธ์ของโดปามีนที่สูงกว่า NAฮอร์น 1973) เนื่องจากโดปามีนและ NA มักให้ความร่วมมือในฟังก์ชั่น mPFC หลายอย่างตั้งแต่หน่วยความจำในการทำงานและการตั้งค่าแบบตั้งใจและเปลี่ยนเป็นการเรียนรู้แบบกลับด้านการยับยั้งการตอบสนองและการตอบสนองต่อความเครียด Robbins และ Arnsten, 2009) สิ่งนี้ทำให้เกิดความเป็นไปได้ที่โดปามีนเพียงอย่างเดียวหรือร่วมกับ NA ใน mPFC อาจมีบทบาทในความแตกต่างของพฤติกรรมระหว่างหนู RHA และ RLA รวมถึงที่พบในพฤติกรรมทางเพศ
เพื่อทดสอบสมมติฐานนี้กิจกรรมของระบบ dopaminergic mesocortical และของระบบ noradrenergic ใน mPFC ได้ถูกศึกษาใน RHA และ RLA หนูสองสายโดยวิธีการ intracerebral microdialysis โดยย่อ dopamine (และ metabolite หลัก DOPAC) และ NA วัดใน dialysates ที่ได้รับจาก prelimbic (PrL) และ infralimbic (IL) ช่องของ mPFC ของเพศที่ไร้เดียงสา (เช่นไม่เคยสัมผัสกับผู้หญิงที่อ่อนไหว) และ RHA ที่มีเพศสัมพันธ์ หนู RLA (เช่นที่ผ่านการทดสอบการมีเพศสัมพันธ์เบื้องต้นห้าครั้งและแสดงระดับของกิจกรรม copulatory อย่างต่อเนื่อง) เมื่ออยู่ในที่ที่ไม่สามารถเข้าถึงตัวเมียได้และในระหว่างการมีเพศสัมพันธ์โดยตรงโดยโครมาโทกราฟีของเหลวความดันสูง
วัสดุและวิธีการ
สัตว์
RHA และ RLA ชายตัวผู้ (N = 30 สำหรับแต่ละบรรทัดชั่ง≈300 g ที่จุดเริ่มต้นของงานทดลอง) ทั้งหมดมาจากอาณานิคมที่ก่อตั้งขึ้นใน 1998 ที่มหาวิทยาลัย Cagliari อิตาลี (Giorgi et al., 2007) ขั้นตอนที่ใช้ในการคัดเลือกสายพันธุ์ของอาณานิคมซาร์ดิเนียได้รับการอธิบายไว้ในรายละเอียดแล้ว (Giorgi et al., 2005).
Ovariectomized stimulus SD female rats (250 – 300 g ที่จุดเริ่มต้นของงานทดลอง) ที่ใช้ในการทดลองทั้งหมดได้มาจาก Envigo (San Pietro al Natisone, อิตาลี) สัตว์ถูกปรับสภาพสี่ตัวต่อกรง (38 cm × 60 cm × 20 cm) ไปยังที่พักอาศัยของภาควิชาวิทยาศาสตร์ชีวการแพทย์ของมหาวิทยาลัย Cagliari อย่างน้อย 10 วันก่อนเริ่มการทดลองที่ 24 ° C ความชื้น 60%, สลับ 12 h วงจรแสง / ความมืด (ไฟดับจาก 08: 00 h ถึง 20: 00 h, น้ำและอาหารห้องปฏิบัติการมาตรฐาน โฆษณาฟรี. สัตว์ได้รับการดูแลเป็นประจำทุกวันสำหรับ 1 – 2 นาทีตลอดระยะเวลาการทำให้คุ้นเคยเพื่อ จำกัด ความเครียดในการจัดการในระหว่างการทดลอง นอกจากนี้การติดต่อกับเจ้าหน้าที่บำรุงรักษาบ้านสัตว์ถูก จำกัด ให้ผู้ดูแลเพียงคนเดียวและมีเครื่องนอนในกรงที่บ้านไม่เคยเปลี่ยนทั้งวันก่อนหรือในวันของการทดลอง การทดลองทั้งหมดดำเนินการระหว่าง 10: 00 h และ 18: 00 h การศึกษานี้ดำเนินการตามคำแนะนำของแนวทางของประชาคมยุโรปคำสั่งของ 22 กันยายน 2010 (2010 / 63 / EU) และกฎหมายของอิตาลี (DL March 4, 2014, n. 26) โปรโตคอลได้รับการอนุมัติจากคณะกรรมการจริยธรรมสำหรับการทดลองสัตว์ของมหาวิทยาลัยกายารี่ (หมายเลขอนุญาต 361 / 2016-PR, เมษายน 08, 2016 ถึง FS)
กลุ่มทดลอง
มีการใช้หนู RHA และ RLA เพศชายที่ไร้เดียงสาและมีประสบการณ์ทางเพศสัมพันธ์ หนูที่ไร้เดียงสาทางเพศสัมพันธ์เป็นหนูที่ไม่เคยสัมผัสกับเพศที่ไวต่อรังไข่และ estradiol + progesterone ที่มีเพศเมีย หนูที่มีประสบการณ์ทางเพศสัมพันธ์คือหนูที่ผ่านการทดสอบการมีเพศสัมพันธ์ติดต่อกัน 5 ครั้งต่อนาที 60 เป็นระยะเวลา 3 วันกับผู้หญิงที่เปิดกว้าง (Sanna และคณะ, 2014a,b) โดยทำการรักษาด้วยเอสตราไดออลเบนโซเอตใต้ผิวหนัง (200 μg / หนูในน้ำมันถั่วลิสง) และโปรเจสเตอโรน (0.5 mg / หนูในน้ำมันถั่วลิสง), 48 h และ 6 h ก่อนการทดสอบการมีเพศสัมพันธ์ตามลำดับ Oestrus ได้รับการยืนยันโดย May-Grunwald-Giemsa coloration และการตรวจด้วยกล้องจุลทรรศน์ของรอยเปื้อนในช่องคลอด 1 h ก่อนการทดลอง ในข้อตกลงกับการศึกษาก่อนหน้า (Sanna และคณะ, 2014a,b, 2015) การทดสอบการมีเพศสัมพันธ์เบื้องต้นห้าครั้งพบว่าเพียงพอที่จะให้หนูโรมันเพศผู้ทั้งสองเส้นแสดงระดับการคงที่ของกิจกรรมการมีเพศสัมพันธ์อย่างต่อเนื่องเช่นหนูที่มีประสบการณ์ทางเพศ RHA และ RLA พอใจเกณฑ์การหลั่งอย่างน้อยหนึ่งครั้ง (หนู RHA หนึ่งตัวและหนู RLA สองตัวที่ไม่พอใจเกณฑ์นี้ถูกยกเลิกในช่วงนี้) สองวันหลังจากการทดสอบเบื้องต้นของการทำ copulatory หนูโรมันที่มีประสบการณ์ทางเพศได้รับการผ่าตัด stereotaxic สำหรับการฝัง microdialysis probe ใน mPFC ดังที่อธิบายไว้ด้านล่าง (Sanna และคณะ, 2015).
microdialysis ใน mPFC ระหว่างพฤติกรรมทางเพศ
วันก่อน microdialysis, ไร้สมรรถภาพทางเพศหรือหนู RHA และ RLA ที่มีประสบการณ์ถูกจัดวางไว้ในเครื่อง stereotaxic (Stoelting Co. , Wood Dale, IL, USA) ภายใต้การฉีดยาไอโซฟลูรูน (1.5% –2%; และปลูกถ่ายด้วยหัววัดขนาดเล็กแบบโฮมเมดในแนวตั้ง (เมมเบรนล้างไต≈3มม. ของพื้นผิวที่ว่าง; Melis et al., 2003) และชี้นำข้างเดียวที่ช่อง mPFC, PrL และ IL (พิกัด: 3.0 mm ล่วงหน้าและ 0.7 mm ด้านข้างถึง bregma และ 5.5 mm ventral ถึง dura; Paxinos และ Watson, 2004) วันของการทดลองสัตว์ถูกถ่ายโอนในช่วงที่มืดของวงจรไปยังกรงผสมพันธุ์ (45 cm × 30 cm × 24 cm) ซึ่งตั้งอยู่ในห้องกันเสียงที่สว่างด้วยแสงสีแดงสลัวและบรรจุภายในอีก กรงลูกแก้วขนาดเล็ก (15 cm × 15 cm × 15 cm) ที่มีรู 25 (Ø 2 mm) ในผนังแนวตั้งเพื่อให้มองเห็นการดมกลิ่นและเสียง แต่ไม่ต้องทำปฏิกิริยาโดยตรง หลังจากช่วงเวลาแห่งความเคยชินของ 2 h โพรบ microdialysis นั้นเชื่อมต่อกับปั๊ม microinfusion CMA / 100 (เครื่องมือฮาร์วาร์ด, Holliston, MA, USA) ด้วยท่อโพลีเอทิลีนและใช้กับสารละลายของ Ringer (147 mM NaCl, 3 mM KCl แคลเซียมคลอไรด์2, pH 6.5) ที่อัตราการไหลของ 2.5 μl / นาที หลังจากช่วงเวลาการปรับสมดุลของ 2 h ของตัวกลางการแพร่กระจายด้วยของเหลวนอกเซลล์จะทำการรวบรวมส่วนแยก dialysate ของ 37.5 μLทุกนาที 15 ในระหว่างการทดลองในหลอดโพลิเอทิลีนเย็นน้ำแข็งสำหรับการวัดความเข้มข้นของโดปามีน, DOPAC และ NA ด้านล่าง หลังจากการรวบรวมอย่างน้อยสี่ dialysate aliquots หนูตัวเมียที่เปิดกว้างถูกนำเข้าไปในกรงขนาดเล็กที่ตั้งอยู่ภายในกรงผสมพันธุ์เป็นเวลา 30 นาที ในช่วง 30 ขั้นต่ำเหล่านี้จะทำการรวบรวมส่วนลงตัวของ dialysate อีกสองตัว ในเงื่อนไขเหล่านี้หนูตัวผู้ไม่สามารถโต้ตอบกับเพศหญิงได้โดยตรง แต่แสดงการแข็งตัวแบบไม่สัมผัส (ดูด้านล่าง) หลังจากช่วงเวลานี้กรงขนาดเล็กจะถูกลบออกมีส่วนร่วมได้รับอนุญาตสำหรับ 75 นาทีและอื่น ๆ อีกห้ารวบรวม aliquots dialysate ในตอนท้ายของช่วงเวลานี้ผู้หญิงก็ถูกย้ายออกจากกรงผสมพันธุ์และเก็บส่วนขยาย dialysate เพิ่มเติม (Pfaus และ Everitt, 1995; Melis et al., 2003; Sanna และคณะ, 2015) พารามิเตอร์ทางเพศที่เกี่ยวข้องกับขั้นตอนการคาดการณ์และการสิ้นเปลืองของพฤติกรรมทางเพศถูกบันทึกไว้ตลอดการทดลอง (ดูด้านล่าง)
พฤติกรรมทางเพศ
มีการบันทึกพารามิเตอร์หลายอย่างของแรงจูงใจทางเพศและการมีเพศสัมพันธ์ร่วมกับขั้นตอนการคาดการณ์และการบริโภคของพฤติกรรมทางเพศตลอดการทดลองโดยผู้สังเกตการณ์ที่ไม่ได้รับรู้ถึงเงื่อนไขการทดลองที่เฉพาะเจาะจงเช่นผู้ที่ไม่รู้เส้นและระดับประสบการณ์ทางเพศ ของสัตว์ที่ใช้ในการทดสอบ (ดูด้านล่าง) โดยสังเขปเวลาแฝงของการแข็งตัวแบบไม่สัมผัสครั้งแรก (NCPEL หมดเวลาจากการแนะนำตัวเมียที่เปิดกว้างในกรงขนาดเล็กด้านใน) และความถี่ (NCPEF จำนวนการแข็งตัวของอวัยวะเพศชายแบบไม่สัมผัสที่เกิดขึ้นในช่วงเวลาที่ตัวเมียอยู่ บันทึกอยู่ในกรงภายใน) การแข็งตัวของอวัยวะเพศชายฟีโรโมนที่ใช้สื่อกลางซึ่งเกิดขึ้นในหนูเพศผู้ที่มีศักยภาพทางเพศในการปรากฏตัวของเพศหญิงที่ไม่สามารถเข้าถึงได้ถือเป็นดัชนีสำคัญของความเร้าอารมณ์ทางเพศ (Sachs et al., 1994; Sachs, 2000; Melis et al., 2003) เมื่อการมีเพศสัมพันธ์ได้รับอนุญาตเช่นระหว่างการสังวาสความล่าช้าในการติดตั้งและ intromit (ML และ IL หมดเวลาจากการถอนกรงขนาดเล็กด้านในจนกระทั่งการติดตั้งครั้งแรกหรือครั้งแรกตามลำดับ); ความถี่ของการติดตั้งและการเกิดเสียงดัง (MF และ IF, จำนวนของการติดตั้งและการแจ้งเตือนตามลำดับในลำดับแรกของกิจกรรมการมีเพศสัมพันธ์และในช่วงระยะเวลาการมีเพศสัมพันธ์ทั้งหมด); เวลาแฝงในการพุ่งออกมา (EL, หมดเวลาตั้งแต่การบุกรุกครั้งแรกของซีรีส์แรกจนถึงการพุ่งออกมา); ความถี่ของการพุ่งออกมา (EF, จำนวนการหลั่งทั้งหมดระหว่างการทดสอบการมีเพศสัมพันธ์) และช่วงเวลาหลังการหลั่ง (PEI, หมดเวลานับจากการพุ่งออกมาครั้งแรกจนถึงการหายใจครั้งต่อไป) ถูกบันทึก ยิ่งไปกว่านั้นประสิทธิภาพการมีส่วนร่วม (CE; จำนวน intromissions ของซีรีย์ที่กำหนดหารด้วยผลรวมของจำนวนของการเมานต์และอินโมชันในซีรีย์เดียวกัน) และช่วง inter-intromission (III; อัตราส่วนระหว่างเวลาแฝงการหลั่งของอนุกรมที่กำหนด และจำนวน intromissions ในซีรีส์นั้น) ก็ถูกคำนวณสำหรับกิจกรรมชุด copulatory ชุดแรก (Sachs และ Barfield, 1976; Meisel และ Sachs, 1994; Melis et al., 2003; Sanna และคณะ, 2014a,b, 2015).
ความมุ่งมั่นของ Dopamine, DOPAC และ Noradrenaline ความเข้มข้นใน Dialysate จาก mPFC
ความเข้มข้นของโดปามีน DOPAC และ NA ใน 20 μLของ aliquot dialysate เดียวกันจาก mPFC โดยโครมาโตกราฟีของเหลวความดันสูง (HPLC) ควบคู่ไปกับการตรวจจับทางเคมีไฟฟ้าโดยใช้ 4011 dual cell (Coulochem II, ESA, Cambridge, MA, USA) อธิบายไว้แล้ว (Melis et al., 2003) ตรวจจับได้ในโหมดย่อที่ + 350 และ −180 mV HPLC ได้รับการติดตั้งคอลัมน์ Supelcosil C18 (7.5 cm × 3.0 mm id, ขนาดอนุภาค 3 μm; Supelco, Supelchem, มิลาน, อิตาลี), ประกอบด้วย 0.06 M citrate / acetate pH 4.2 mM EDTA, 20 μM triethylamine และ 0.1 mM โซเดียมโดเดซิลซัลเฟตเป็นเฟสเคลื่อนที่ในอัตราการไหลของ 1 mL / นาทีและอุณหภูมิห้อง ความไวของการทดสอบคือ 0.03 pg สำหรับ dopamine, 0.6 pg สำหรับ DOPAC และ 0.125 pg สำหรับ NA
จุลกายวิภาคศาสตร์เนื้อเยื่อ
ในตอนท้ายของการทดลองหนูถูกฆ่าตายโดยการตัดหัวสมองออกจากกะโหลกศีรษะทันทีและแช่ในฟอร์มัลดีไฮด์ในน้ำ 4% สำหรับ 12 – 15 วัน หลังจากช่วงเวลานี้ 40 μmส่วนของสมองโคโรนาถูกเตรียมด้วย microtome แช่แข็งเปื้อนด้วย Neutral Red และตรวจสอบด้วยกล้องจุลทรรศน์คอนทราสต์เฟส ตำแหน่งของปลายโพรบถูกแปลเป็นภาษาท้องถิ่นใน mPFC โดยทำตามทางเดินของโพรบผ่านชุดของส่วนสมอง (ดูรูปที่ 1) พบว่าหนูเท่านั้นที่มีส่วนที่ใช้งานของเยื่อแผ่นกรองอย่างถูกต้องในช่อง PrL และ IL ของ mPFC ได้รับการพิจารณาสำหรับการประเมินผลทางสถิติของผลลัพธ์ (หนูหนึ่งตัวจากแต่ละกลุ่มการทดลองถูกทิ้งในระยะนี้)
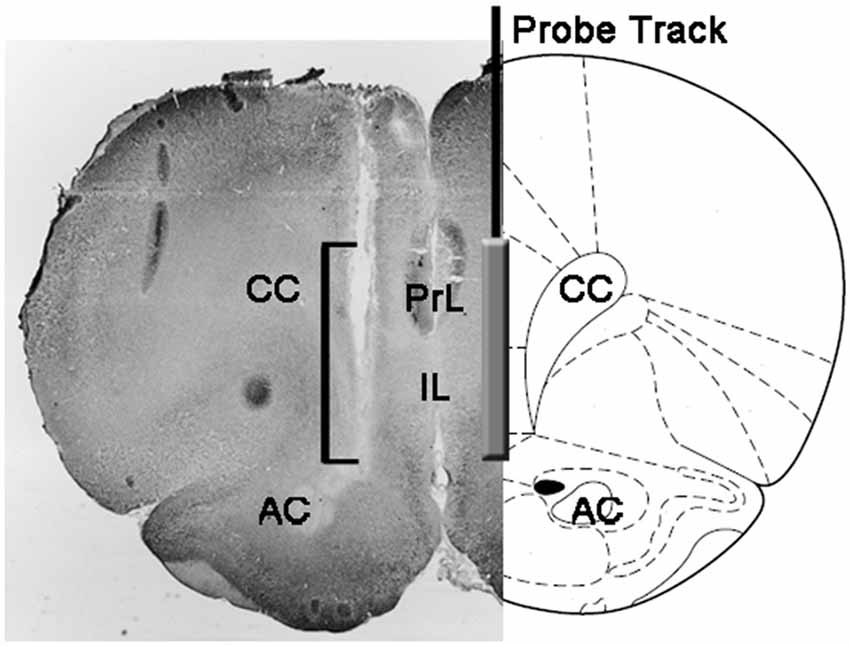 รูป 1. การเป็นตัวแทนแผนผังของส่วนชเวียนของสมองหนูแสดงการติดตามของการสอบสวน microdialysis ในส่วน PrL และ IL ของเยื่อหุ้มสมอง prefrontal อยู่ตรงกลาง (mPFC; Paxinos และ Watson, 2004) กรอบสี่เหลี่ยมจัตุรัสในภาพถ่ายขนาดเล็กแสดงส่วนของส่วนสีแดงกลางที่เป็นกลางซึ่งแสดงส่วนที่ทำงานของเมมเบรนแบบ dialyzing ของโพรบ microdialysis ตัวย่อ: PRL พื้นที่ก่อนกำหนด IL, พื้นที่ infralimbic; AC, commissura ด้านหน้า; CC, callosum คลัง
รูป 1. การเป็นตัวแทนแผนผังของส่วนชเวียนของสมองหนูแสดงการติดตามของการสอบสวน microdialysis ในส่วน PrL และ IL ของเยื่อหุ้มสมอง prefrontal อยู่ตรงกลาง (mPFC; Paxinos และ Watson, 2004) กรอบสี่เหลี่ยมจัตุรัสในภาพถ่ายขนาดเล็กแสดงส่วนของส่วนสีแดงกลางที่เป็นกลางซึ่งแสดงส่วนที่ทำงานของเมมเบรนแบบ dialyzing ของโพรบ microdialysis ตัวย่อ: PRL พื้นที่ก่อนกำหนด IL, พื้นที่ infralimbic; AC, commissura ด้านหน้า; CC, callosum คลัง
สถิติ
การวิเคราะห์ทางสถิติของชีวเคมี (โดปามีน, DOPAC และ NA) และพฤติกรรม (NCPEL และ NCPEF, ML, IL และ EL, MF, IF และ EF และ PEI) ข้อมูลได้ดำเนินการอย่างใดอย่างหนึ่งรวมทั้งวิชาทดลองทั้งหมด พฤติกรรมการมีเพศสัมพันธ์) หรือยกเว้นผู้ที่ไม่ได้มีเพศสัมพันธ์กับอุทานในระหว่างการทดลอง microdialysis ในกรณีแรกเมื่อสัตว์ทั้งหมดรวมอยู่ในการวิเคราะห์สัตว์ที่ไม่ได้แสดง NCPE ภายในเวลาที่ผู้หญิงไม่สามารถเข้าถึงได้หรือไม่ได้ติดตั้งหรือ intromit หรืออุทานกับเพศหญิงที่มีอยู่ได้รับมอบหมายคะแนนเต็มช่วง: 1800 s หาก ตัวผู้ไม่ได้แสดง NCPE ใน 30 ขั้นต่ำที่ไม่สามารถเข้าถึงตัวเมียได้ 900 s สำหรับ ML และ IL หากตัวผู้ไม่ได้เมานต์หรือ intromit กับตัวเมียที่มีอยู่ 1800 s สำหรับ EL หากชายไม่ได้พุ่งออกมาและ 600 s สำหรับ PEI หากชายไม่ได้อยู่ในภาวะ intromit หลังจากพุ่งออกมาครั้งแรก ในกรณีที่สองสัตว์ที่ไม่ได้มีเพศสัมพันธ์กับอุทานถูกแยกออกจากการวิเคราะห์ สิ่งนี้ทำเพื่อประเมินว่าความแตกต่างในพฤติกรรมทางเพศอาจมีความสัมพันธ์กับความแตกต่างของความเข้มข้นของสารกระตุ้นโดปามีนนอกเซลล์, DOPAC และ NA ใน mPFC ของสายหนูหนูโรมันในช่วงระยะที่สองของกิจกรรมทางเพศ ของอาสาสมัครที่ไม่แสดงพฤติกรรมทางเพศโดยกำหนดค่าคงที่สำหรับพารามิเตอร์ทางเพศที่วิเคราะห์แล้ว
มีการวิเคราะห์ทางสถิติหลายครั้งด้วย ANOVAs เพื่อตรวจจับและระบุลักษณะความแตกต่างที่เป็นไปได้ระหว่างเงื่อนไขการทดลองที่ใช้ ก่อนดำเนินการ ANOVAs ชุดข้อมูลของตัวแปรการทดลองแต่ละแบบจะถูกตรวจสอบเพื่อความสม่ำเสมอของความแปรปรวนในกลุ่มการทดลองทั้งสี่กลุ่ม (ทำกับสัตว์ทุกชนิดหรือหนูที่มีการสัมผัสกับการพุ่งออกมาเท่านั้น) ด้วยการทดสอบของ Bartlett หรือ Levine เมื่อพบความแตกต่างอย่างมีนัยสำคัญในความแปรปรวนชุดข้อมูลจะถูกแปลงลอการิทึม (เช่นค่า Y ทดลองเปลี่ยนเป็นค่า Log Y) ตรวจสอบความสม่ำเสมอของความแปรปรวนอีกครั้งแล้ววิเคราะห์โดย ANOVAs สั้น ๆ พารามิเตอร์พฤติกรรมที่ได้รับมาก่อน (เช่นกับผู้หญิงที่ไม่สามารถเข้าถึงได้) และระหว่างกิจกรรมชุด copulatory ชุดแรก (จากการเมานท์ครั้งแรก / การหายใจเข้าสู่การเมานท์ครั้งแรก / การฉีดยาเข้าหลังจากการหลั่งครั้งแรก) ของหนูเพศผู้ ในระหว่างการวิเคราะห์ด้วย microdialysis โดยการวิเคราะห์ความแปรปรวนสองทางโดยใช้สายหนูและระดับประสบการณ์ทางเพศเป็นปัจจัยระหว่างวิชา (ค่าเฉลี่ยของพารามิเตอร์พฤติกรรมมีการรายงานในตาราง 1 และ F ค่าและระดับนัยสำคัญของการวิเคราะห์ความแปรปรวนในตาราง 2).
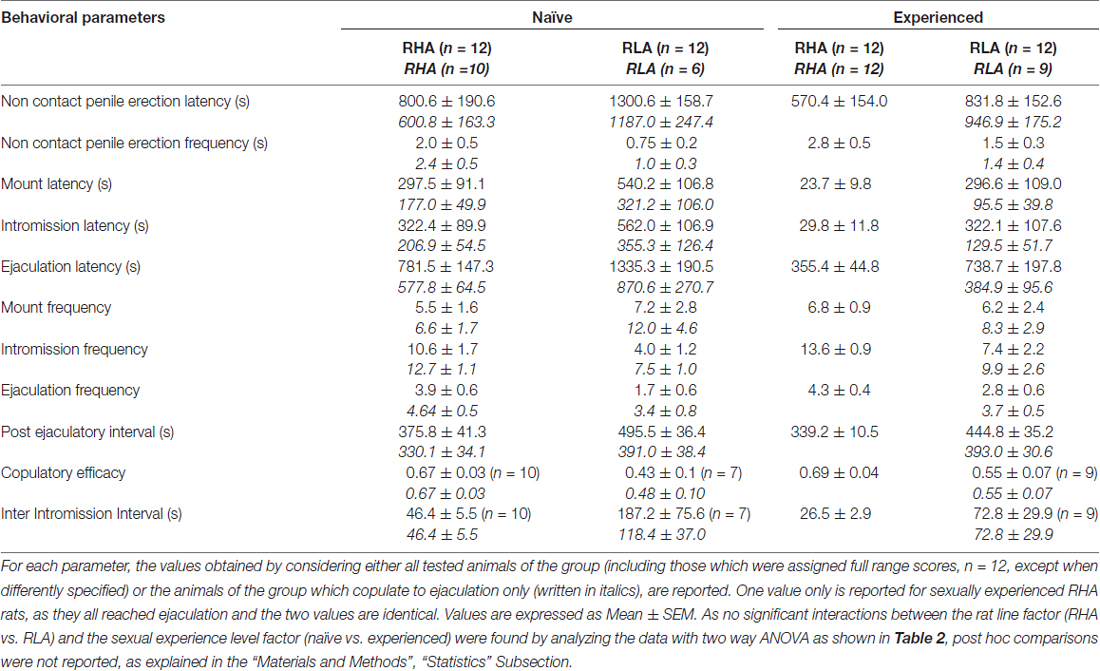 1 ตาราง. พารามิเตอร์ copulatory วัดในชุดแรกของกิจกรรม copulatory (ซึ่งเริ่มต้นด้วยการเมานท์ครั้งแรก / intromission เมื่อเพศหญิงจะทำให้สามารถเข้าถึงชายและสิ้นสุดหลังจากช่วงเวลา ejaculatory โพสต์กับเมาท์ / intrommision แรกของซีรีส์ที่สอง) และ non อวัยวะเพศชาย การแข็งตัวของหนูที่ไร้สมรรถภาพทางเพศและมีประสบการณ์ RHA และ RLA
1 ตาราง. พารามิเตอร์ copulatory วัดในชุดแรกของกิจกรรม copulatory (ซึ่งเริ่มต้นด้วยการเมานท์ครั้งแรก / intromission เมื่อเพศหญิงจะทำให้สามารถเข้าถึงชายและสิ้นสุดหลังจากช่วงเวลา ejaculatory โพสต์กับเมาท์ / intrommision แรกของซีรีส์ที่สอง) และ non อวัยวะเพศชาย การแข็งตัวของหนูที่ไร้สมรรถภาพทางเพศและมีประสบการณ์ RHA และ RLA
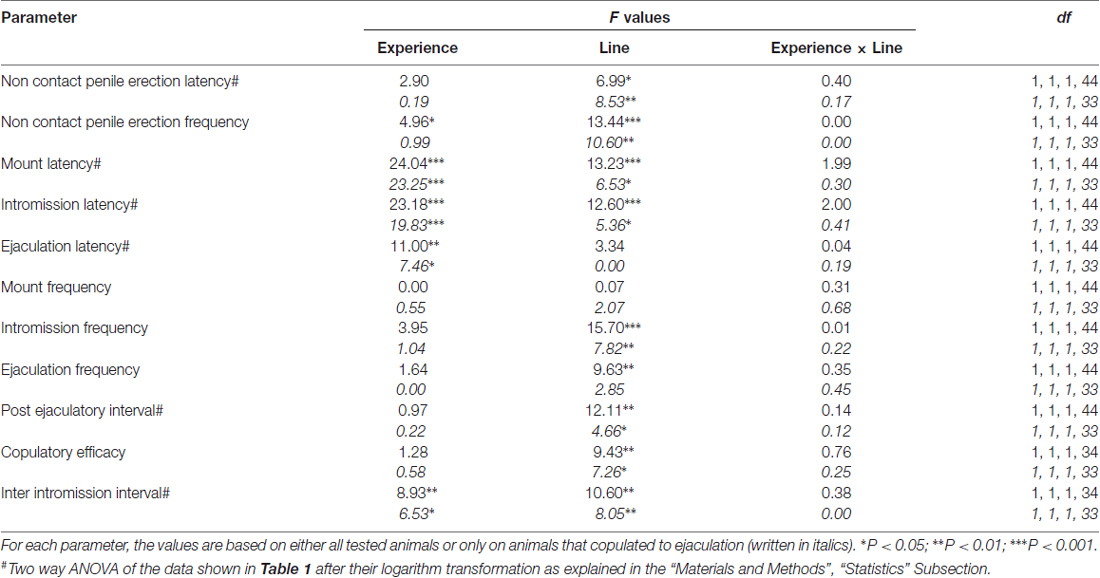 2 ตาราง. F ค่าและระดับนัยสำคัญของ ANOVA แบบสองทางที่ดำเนินการกับข้อมูลที่รายงานในตาราง 1 โดยใช้สายหนู (RHA กับ RLA) และระดับประสบการณ์ทางเพศ (ไร้เดียงสาเทียบกับประสบการณ์) เป็นปัจจัยระหว่างวิชา
2 ตาราง. F ค่าและระดับนัยสำคัญของ ANOVA แบบสองทางที่ดำเนินการกับข้อมูลที่รายงานในตาราง 1 โดยใช้สายหนู (RHA กับ RLA) และระดับประสบการณ์ทางเพศ (ไร้เดียงสาเทียบกับประสบการณ์) เป็นปัจจัยระหว่างวิชา
นอกจากนี้การวิเคราะห์โดยรวมของข้อมูลที่ได้รับจากหนูแต่ละตัวในช่วง microdialysis ทำโดยการคำนวณ AUCs แรกที่ได้รับโดยการวางแผนค่าความเข้มข้นของโดปามีน, DOPAC หรือ NA หรือจำนวนของการแข็งตัวของการสัมผัสไม่ติดต่อ . เวลา (เริ่มต้น 1 ชั่วโมงหลังจากการแนะนำหนูตัวผู้ในกรงผสมพันธุ์จนจบการทดลอง −180 นาทีแบ่งออกเป็นเศษส่วนของ 15 นาทีสำหรับค่าทางประสาท - หรือในเวลาที่ผู้หญิงได้รับการแนะนำในกรงผสมพันธุ์ สำหรับพารามิเตอร์พฤติกรรม) และจากนั้นเปรียบเทียบค่าที่คำนวณได้ด้วย ANOVAs แบบสองทางโดยใช้บรรทัดหนูและระดับประสบการณ์ทางเพศระหว่างปัจจัยของอาสาสมัคร (ค่าเฉลี่ยของ AUCs ของพารามิเตอร์ทางประสาทและพฤติกรรมมีการรายงานในตาราง 4 และ F ค่าและระดับนัยสำคัญของการวิเคราะห์ความแปรปรวนในตาราง 5) ในที่สุดการวิเคราะห์แบบจุดต่อจุดโดยละเอียดยิ่งขึ้นของชุดข้อมูลแต่ละชุด (เช่นค่าความเข้มข้นของโดปามีน, DOPAC, NA และจำนวนการติดตั้งแบบไม่สัมผัส, การติดตั้ง, การแจ้งเตือนและอุทานเทียบกับเวลา) ANOVAs สำหรับการวัดซ้ำโดยใช้สายหนูและระดับของประสบการณ์ทางเพศระหว่างปัจจัยเรื่องและเวลา (เช่นเศษส่วน dialysate) เป็นปัจจัยภายในวิชา ( F ค่าและระดับนัยสำคัญของการวิเคราะห์ความแปรปรวนเหล่านี้ถูกรายงานในตาราง 6) การวิเคราะห์ที่คล้ายกันได้ดำเนินการโดยพิจารณาเฉพาะค่าฐานของพารามิเตอร์ทางเคมีประสาท (aliquots dialysate สี่ครั้งล่าสุดที่รวบรวมก่อนการแนะนำในกรงผสมพันธุ์ของเพศหญิงที่รับได้) ซึ่งมีรายงานค่าเฉลี่ยในตาราง 3. เป็นครั้งแรก แต่ไม่พบการโต้ตอบลำดับที่สองเมื่อดำเนินการ ANOVA แบบแฟกทอเรียลทั่วไปสำหรับการวัดซ้ำโดยเฉพาะในบรรทัด×เวลาและประสบการณ์×เวลาโต้ตอบสำหรับพารามิเตอร์ทางเคมีประสาท (ดูตาราง 6) การวิเคราะห์ความแปรปรวนแบบสองทางกับเส้นหรือระดับประสบการณ์ทางเพศระหว่างปัจจัยของอาสาสมัครและเวลาตามปัจจัยภายในเรื่องถูกดำเนินการในชุดข้อมูลเหล่านี้โดยการเปรียบเทียบโดยตรงทางเพศที่ไร้เดียงสาหรือมีประสบการณ์ RHA กับ RLA หรือ RDA หนู RHA ไร้เดียงสาหรือมีประสบการณ์ทางเพศสัมพันธ์กับหนูไร้เดียงสา RLA ผลของการ โพสต์เฉพาะกิจ จับคู่ความแตกต่างที่ชาญฉลาดโดยใช้การทดสอบ HSD ของ Tukey ในการโต้ตอบที่สำคัญที่เปิดเผยโดย ANOVAs แบบสองทางเหล่านี้จะถูกรายงานในรูปที่ 2. ในกรณีอื่น ๆ ทั้งหมด โพสต์เฉพาะกิจ ไม่มีการเปรียบเทียบเนื่องจาก ANOVAs ไม่สามารถเปิดเผยการมีปฏิสัมพันธ์ที่สำคัญระหว่างสายหนูระดับประสบการณ์ทางเพศและเวลา การวิเคราะห์ทางสถิติได้ดำเนินการทั้งหมดด้วย Graph Pad 5 (PRISM, San Diego, CA, USA) และ STATISTICA 12 (Statsoft, Tulsa, OK, USA) โดยมีระดับนัยสำคัญที่กำหนดไว้ที่ P <0.05.
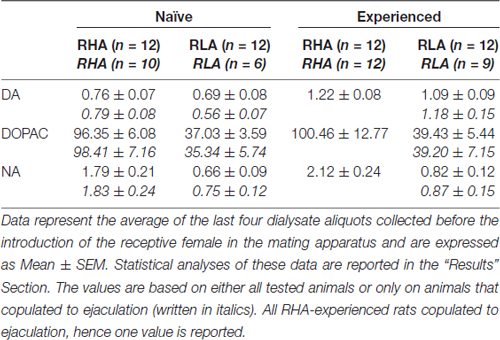 3 ตาราง. Basal dopamine (DA), 3,4-Dihydroxyphenylacetic acid (DOPAC) และ noradrenaline (NA) ความเข้มข้น (nM) ใน dialysate จากเยื่อหุ้มสมอง prefrontal cortex (mPFC) ของหนูที่มีเพศสัมพันธ์และมีประสบการณ์ RHA และ RLA
3 ตาราง. Basal dopamine (DA), 3,4-Dihydroxyphenylacetic acid (DOPAC) และ noradrenaline (NA) ความเข้มข้น (nM) ใน dialysate จากเยื่อหุ้มสมอง prefrontal cortex (mPFC) ของหนูที่มีเพศสัมพันธ์และมีประสบการณ์ RHA และ RLA
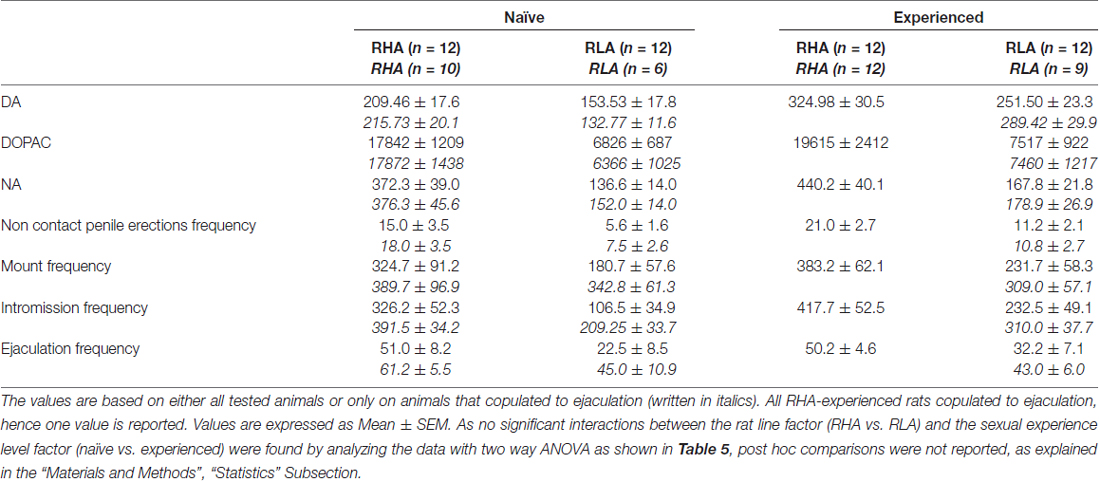 4 ตาราง. การประเมินความแตกต่างโดยรวมของความเข้มข้น DA, DOPAC และ NA และพฤติกรรมทางเพศระหว่างหนูที่ไร้สมรรถภาพทางเพศและหนูที่มีประสบการณ์ RHA และ RLA โดยการวิเคราะห์ AUC เฉลี่ยที่ได้จากผลลัพธ์ที่แสดงในรูป 2 และ 3.
4 ตาราง. การประเมินความแตกต่างโดยรวมของความเข้มข้น DA, DOPAC และ NA และพฤติกรรมทางเพศระหว่างหนูที่ไร้สมรรถภาพทางเพศและหนูที่มีประสบการณ์ RHA และ RLA โดยการวิเคราะห์ AUC เฉลี่ยที่ได้จากผลลัพธ์ที่แสดงในรูป 2 และ 3.
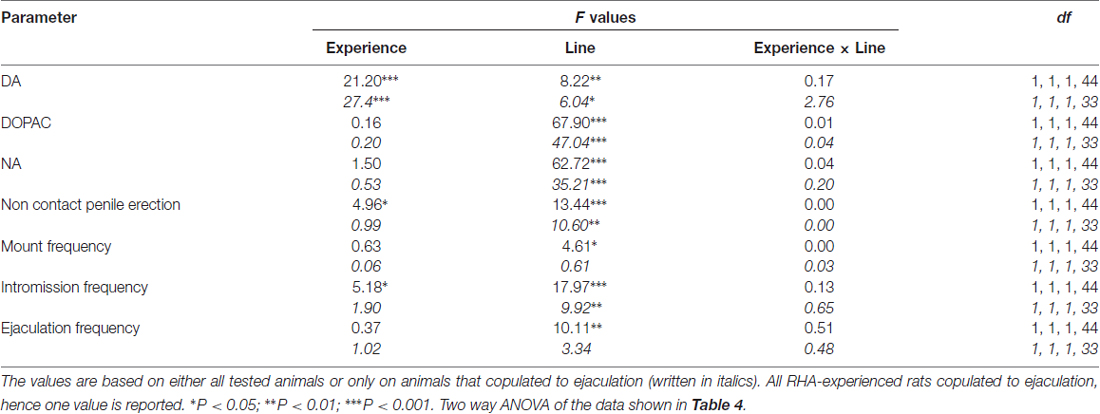 5 ตาราง. F ค่าและระดับนัยสำคัญของ ANOVAs แบบสองทางที่ดำเนินการกับข้อมูลที่รายงานในตาราง 4 โดยใช้สายหนู (RHA กับ RLA) และระดับประสบการณ์ทางเพศ (ไร้เดียงสาเทียบกับประสบการณ์) เป็นปัจจัยระหว่างวิชา
5 ตาราง. F ค่าและระดับนัยสำคัญของ ANOVAs แบบสองทางที่ดำเนินการกับข้อมูลที่รายงานในตาราง 4 โดยใช้สายหนู (RHA กับ RLA) และระดับประสบการณ์ทางเพศ (ไร้เดียงสาเทียบกับประสบการณ์) เป็นปัจจัยระหว่างวิชา
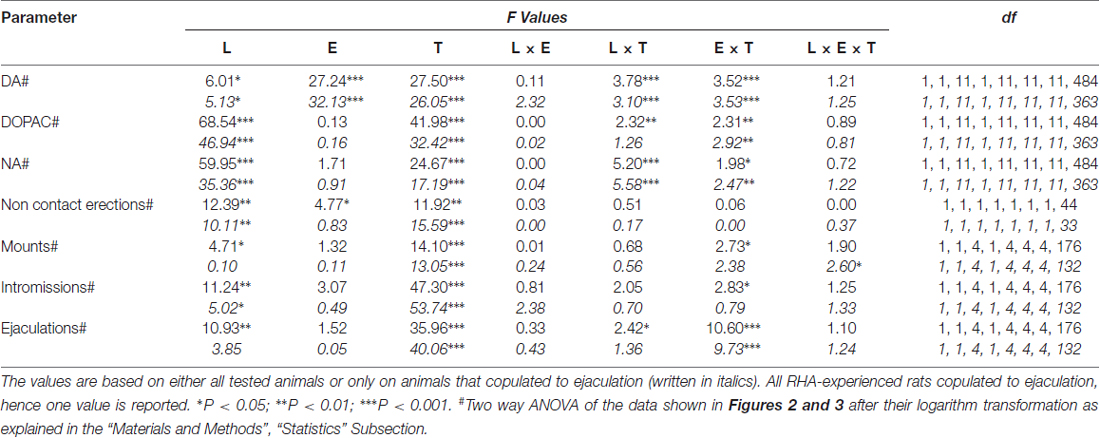 6 ตาราง. F ค่าและระดับความสำคัญของ ANOVA แบบแฟกทอเรียลทั่วไปสำหรับการวัดซ้ำที่ดำเนินการกับผลลัพธ์ที่แสดงในรูป 2 และ 3 โดยใช้เส้นหนู (L; RHA เทียบกับ RLA) และระดับประสบการณ์ทางเพศ (E) (ไร้เดียงสาเทียบกับประสบการณ์) ระหว่างปัจจัยเรื่องและเวลา (T) (เศษส่วน dialysate) เป็นปัจจัยภายในวิชา
6 ตาราง. F ค่าและระดับความสำคัญของ ANOVA แบบแฟกทอเรียลทั่วไปสำหรับการวัดซ้ำที่ดำเนินการกับผลลัพธ์ที่แสดงในรูป 2 และ 3 โดยใช้เส้นหนู (L; RHA เทียบกับ RLA) และระดับประสบการณ์ทางเพศ (E) (ไร้เดียงสาเทียบกับประสบการณ์) ระหว่างปัจจัยเรื่องและเวลา (T) (เศษส่วน dialysate) เป็นปัจจัยภายในวิชา
 รูป 2. Dopamine (DA), DOPAC และ noradrenaline (NA) ความเข้มข้นใน mPFC dialysates จากการมีเพศสัมพันธ์ (A, C, E) และหนูตัวผู้ RHA และ RLA ที่มีประสบการณ์ (B, D, F) ระหว่างทำกิจกรรมทางเพศกับหญิงที่เปิดกว้าง การมีเพศสัมพันธ์ไร้เดียงสา (กล่าวคือไม่เคยสัมผัสกับผู้หญิงที่เปิดรับทางเพศสัมพันธ์มาก่อน) และมีประสบการณ์ทางเพศ (ซึ่งได้รับการทดสอบการมีเพศสัมพันธ์ห้าครั้ง) ของทั้งสองบรรทัดด้วย microdialysis probe stereotaxically ฝังใน mPFC หัววัดแบบไมโครดีซีถูกนำมาใช้ร่วมกับบัฟเฟอร์การล้างไตตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ" หลังจากการรวบรวมของ dialysate aliquots สี่ตัวเพื่อหาค่าฐานแล้วตัวเมียที่รับไม่สามารถเข้าถึงได้จะถูกวางไว้ในกรงขนาดเล็กของอุปกรณ์การผสมพันธุ์ (time = 0, inaccess.fe) หลังจาก 30 นาทีกรงขนาดเล็กจะถูกลบออกและอนุญาตให้มีเพศสัมพันธ์สำหรับ 75 นาที (การมีเพศสัมพันธ์) หลังจากที่หญิงถูกลบออกจากกรงผสมพันธุ์ (fe.remov) ในระหว่างการทดลองจะทำการวัดการแข็งตัวของหน้าสัมผัสแบบไม่สัมผัสและพารามิเตอร์ของ copulatory และทำการแยกส่วนลงตัวของ dialysate ทุกๆ 15 นาทีและวิเคราะห์สำหรับ dopamine, DOPAC และ NA ตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ" ค่าหมายถึง± SEM ของค่าที่ได้รับจากหนู 12 ทั้งหมดต่อกลุ่ม (RHA_all = เส้นสีเขียวที่เป็นของแข็ง, RLA_all = เส้นสีน้ำตาลทึบ) หรือที่เกี่ยวข้องกับการพุ่งออกมาเท่านั้น (RHA_cop = เส้นสีแดงประ; RLA_cop = เส้นสีน้ำเงินประ) ในหนูที่มีประสบการณ์ RHA สัตว์ทุกตัวจะมีเพศสัมพันธ์และค่าเหมือนกัน * * * *P <0.05 เมื่อเทียบกับค่าพื้นฐาน (ไม่มีเพศหญิง) ของกลุ่ม (สีเขียวสำหรับ RHA_all; สีแดงสำหรับ RHA_cop, สีน้ำตาลสำหรับ RLA_all, สีน้ำเงินสำหรับ RLA_cop); #P <0.05 เทียบกับค่าที่สอดคล้องกันของกลุ่ม RLA (สีแดง RHA_all เทียบกับ RLA_all; สีเขียว RHA_cop เทียบกับ RLA_cop); §P <0.05 เมื่อเทียบกับค่าที่ตรงตามเวลาของหนูที่ไร้เดียงสาทางเพศ (สีเขียว RHA_all ที่มีประสบการณ์เทียบกับไร้เดียงสา RHA_all สีแดง RHA_cop ที่มีประสบการณ์เทียบกับไร้เดียงสา RHA_cop สีน้ำตาล RLA_all ที่มีประสบการณ์เทียบกับไร้เดียงสา RLA_all สีฟ้า RLA_cop ที่มีประสบการณ์เทียบกับไร้เดียงสา RLA_cop) (ANOVA แบบสองทางที่ทำกับข้อมูลที่แสดงหลังจากการแปลงลอการิทึมตามที่อธิบายไว้ในส่วนย่อย "วัสดุและวิธีการ" "สถิติ" ตามด้วยการทดสอบ HSD ของ Tukey)
รูป 2. Dopamine (DA), DOPAC และ noradrenaline (NA) ความเข้มข้นใน mPFC dialysates จากการมีเพศสัมพันธ์ (A, C, E) และหนูตัวผู้ RHA และ RLA ที่มีประสบการณ์ (B, D, F) ระหว่างทำกิจกรรมทางเพศกับหญิงที่เปิดกว้าง การมีเพศสัมพันธ์ไร้เดียงสา (กล่าวคือไม่เคยสัมผัสกับผู้หญิงที่เปิดรับทางเพศสัมพันธ์มาก่อน) และมีประสบการณ์ทางเพศ (ซึ่งได้รับการทดสอบการมีเพศสัมพันธ์ห้าครั้ง) ของทั้งสองบรรทัดด้วย microdialysis probe stereotaxically ฝังใน mPFC หัววัดแบบไมโครดีซีถูกนำมาใช้ร่วมกับบัฟเฟอร์การล้างไตตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ" หลังจากการรวบรวมของ dialysate aliquots สี่ตัวเพื่อหาค่าฐานแล้วตัวเมียที่รับไม่สามารถเข้าถึงได้จะถูกวางไว้ในกรงขนาดเล็กของอุปกรณ์การผสมพันธุ์ (time = 0, inaccess.fe) หลังจาก 30 นาทีกรงขนาดเล็กจะถูกลบออกและอนุญาตให้มีเพศสัมพันธ์สำหรับ 75 นาที (การมีเพศสัมพันธ์) หลังจากที่หญิงถูกลบออกจากกรงผสมพันธุ์ (fe.remov) ในระหว่างการทดลองจะทำการวัดการแข็งตัวของหน้าสัมผัสแบบไม่สัมผัสและพารามิเตอร์ของ copulatory และทำการแยกส่วนลงตัวของ dialysate ทุกๆ 15 นาทีและวิเคราะห์สำหรับ dopamine, DOPAC และ NA ตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ" ค่าหมายถึง± SEM ของค่าที่ได้รับจากหนู 12 ทั้งหมดต่อกลุ่ม (RHA_all = เส้นสีเขียวที่เป็นของแข็ง, RLA_all = เส้นสีน้ำตาลทึบ) หรือที่เกี่ยวข้องกับการพุ่งออกมาเท่านั้น (RHA_cop = เส้นสีแดงประ; RLA_cop = เส้นสีน้ำเงินประ) ในหนูที่มีประสบการณ์ RHA สัตว์ทุกตัวจะมีเพศสัมพันธ์และค่าเหมือนกัน * * * *P <0.05 เมื่อเทียบกับค่าพื้นฐาน (ไม่มีเพศหญิง) ของกลุ่ม (สีเขียวสำหรับ RHA_all; สีแดงสำหรับ RHA_cop, สีน้ำตาลสำหรับ RLA_all, สีน้ำเงินสำหรับ RLA_cop); #P <0.05 เทียบกับค่าที่สอดคล้องกันของกลุ่ม RLA (สีแดง RHA_all เทียบกับ RLA_all; สีเขียว RHA_cop เทียบกับ RLA_cop); §P <0.05 เมื่อเทียบกับค่าที่ตรงตามเวลาของหนูที่ไร้เดียงสาทางเพศ (สีเขียว RHA_all ที่มีประสบการณ์เทียบกับไร้เดียงสา RHA_all สีแดง RHA_cop ที่มีประสบการณ์เทียบกับไร้เดียงสา RHA_cop สีน้ำตาล RLA_all ที่มีประสบการณ์เทียบกับไร้เดียงสา RLA_all สีฟ้า RLA_cop ที่มีประสบการณ์เทียบกับไร้เดียงสา RLA_cop) (ANOVA แบบสองทางที่ทำกับข้อมูลที่แสดงหลังจากการแปลงลอการิทึมตามที่อธิบายไว้ในส่วนย่อย "วัสดุและวิธีการ" "สถิติ" ตามด้วยการทดสอบ HSD ของ Tukey)
ผลสอบ
หนู RHA และ RLA แสดงจำนวนของการติดตั้งแบบไม่สัมผัสที่แตกต่างกันและพฤติกรรมการมีเพศสัมพันธ์ที่แตกต่างกัน
สอดคล้องกับการศึกษาก่อนหน้า (Sanna และคณะ, 2014a,b, 2015) หนูเพศผู้ที่ไร้สมรรถภาพทางเพศและหนู RLA จำนวนหนึ่งมีส่วนร่วมในกิจกรรมทางเพศเมื่อใส่กับผู้หญิงที่เปิดกว้างในระหว่างการรวบรวมส่วนแบ่ง dialysate จาก mPFC โดย microdialysis intracerebral โดยสังเขปในการศึกษานี้ 10 จาก 12 ทางเพศnaïveหนู RHA เพศชาย (83%) ที่เกี่ยวข้องกับการพุ่งออกมาในการทดสอบ copulatory ครั้งแรกของพวกเขากับ 6 จาก 12 naïve RLA หนู (50%) ในระหว่างการทดลอง microdialysis ความแตกต่างนี้ยังพบได้ในหนูเพศผู้ RHA และ RLA ที่มีประสบการณ์ทางเพศ (หลังจากการทดสอบ copulatory ห้าครั้ง) แม้ว่าจะถูกลดทอนด้วยหนู 12 RHA ทั้งหมดก็พุ่งออกมาต่อต้าน 9 จากหนู 12 RLA ในระหว่างการ microdialysis เสมอในข้อตกลงกับการศึกษาก่อนหน้านี้ในการศึกษานี้ทั้งสองหนูโรมันสายพันธุ์แสดงความล่าช้าและความถี่ที่แตกต่างกันของอวัยวะเพศชายที่ไม่ติดต่ออวัยวะเพศชายเมื่อสัมผัสกับเพศหญิงที่ไม่สามารถเข้าถึงได้เช่นเดียวกับรูปแบบที่แตกต่างกัน ความแตกต่างเหล่านี้ถูกค้นพบเมื่อพิจารณาข้อมูลของสัตว์ทดลองทั้งหมดของกลุ่มที่กำหนดไม่ว่าพวกมันจะถูกหลั่งออกมาหรือไม่ก็ตาม (เช่นโดยการให้คะแนนเต็มให้กับสัตว์ที่ไม่แสดงพฤติกรรมนั่นคือหนูที่ไม่ได้มีพฤติกรรม การพุ่งออกมา) หรือเมื่อพิจารณาข้อมูลของสัตว์ที่แสดงพฤติกรรมของกลุ่มที่กำหนดเท่านั้น (เช่นสัตว์ที่เกี่ยวข้องกับการพุ่งออกมาตารางที่ 1) ดังนั้นการวิเคราะห์ทางสถิติของค่าของการไม่ติดต่อ (NCPE) และพารามิเตอร์ copulatory วัดในชุดแรกของกิจกรรม copulatory โดย ANOVAs แบบสองทางเปิดเผยความแตกต่างอย่างมีนัยสำคัญระหว่างหนูสองสายทั้งเมื่อพิจารณาหนูทดลองทั้งหมดหรือ Copulating หนูเท่านั้น ในกลุ่มการทดลองสี่กลุ่ม (ดูตาราง 2) ในความเป็นจริงเมื่อพิจารณาข้อมูลที่ได้จากสัตว์ทดลองทั้งหมดจำนวนของการแข็งตัวของการติดต่อไม่สูงขึ้นและ NCPEL, ML, IL และ PEI นั้นสั้นกว่าหนู RHA อย่างมีนัยสำคัญเมื่อเปรียบเทียบกับหนู RLA นอกจากนี้ EF และ CE นั้นสูงขึ้นในขณะที่หนู III สั้นกว่าอย่างมีนัยสำคัญในหนู RHA เมื่อเทียบกับหนู RLA ความแตกต่างดังกล่าวข้างต้นบางส่วนระหว่างสองสายหนูโรมันมีแนวโน้มที่จะลดลงหรือหายไปเมื่อมีการทดสอบ copulatory ซ้ำ อย่างไรก็ตามความแตกต่างเหล่านี้บางส่วนยังคงปรากฏหลังจากการรักษาพฤติกรรมทางเพศโดยการมีประสบการณ์ทางเพศซ้ำแล้วซ้ำอีก ผลลัพธ์ที่คล้ายกันได้รับเมื่อพิจารณาเฉพาะค่าของสัตว์ในกลุ่มทดลองทั้งสี่กลุ่มที่มีการหลั่งออกมายกเว้นกลุ่ม EF (ดูตาราง 1, 2).
ความเข้มข้นพื้นฐานของโดปามีนนอกเซลล์, DOPAC และ Noradrenaline ใน mPFC dialysates จากการมีเพศสัมพันธ์ไร้เดียงสาและ RHA และ RLA ที่มีประสบการณ์
ภายใต้เงื่อนไขการทดลองในปัจจุบันปริมาณโดปามีน, DOPAC และ NA ใน dialysates ที่ได้รับจาก mPFC ของทุกคน (เช่นไม่ว่าพวกมันจะมีเพศสัมพันธ์กับการหลั่งหรือไม่ก็ตาม) ทางเพศหนูที่ไร้เชื้อ RHA และ RLA คือ 2.32 pg และ 2.11 pg สำหรับ dopamine 322.56 pg และ 124.42 pg สำหรับ DOPAC และ 6.05 pg และ 2.24 pg สำหรับ NA ตามลำดับใน 20 μlของ dialysate ปริมาณที่คล้ายกันถูกวัดใน dialysates ที่ได้รับจาก mPFC ของหนูที่มีประสบการณ์ RHA และ RLA (โดปามีน: 3.73 pg และ 3.33 pg, DOPAC: 353.62 pg และ 133.32 pg, NA: 7.17 pg และ 2.78 pg ใน RHA และ RLA ตามลำดับ) . ค่าเหล่านี้บ่งบอกถึงความเข้มข้นของ≅0.8 – 1.2 nM และ≅95 – 100 nM สำหรับ dopamine นอกเซลล์และ DOPAC ตามลำดับและ≅1.8 – 2.2 nM สำหรับ extracellular NA ใน mPFC ของ RHA rats และความเข้มข้น≅0.8 nMX – 1.2 nMX สำหรับ ncx และ≅35 – 40 nM สำหรับโดปามีนนอกเซลล์และ DOPAC ตามลำดับและ≅0.6 – 0.9 nM สำหรับ extracellular NA ใน mPFC ของหนู RLA (ตาราง 3) ค่าดังกล่าวข้างต้นได้มาหลังจากช่วงเวลาสมดุลของ 2 h ของ dialysis buffer ด้วยของเหลวนอกเซลล์ mPFC เนื่องจากการฟื้นตัวของโดปามีนจริง DOPAC และ NA ของโพรบฟอกไตนั้นประมาณว่าใกล้เคียงกับ 20%, โดปามีนนอกเซลล์, ความเข้มข้นของ DOPAC และ NA อาจใกล้เคียงกับ≅4 – 5 nM ทั้งสองบรรทัดสำหรับโดปามีน, ≅500 และ≅ 200 nM สำหรับ DOPAC และ≅10และ≅4 nM สำหรับ NA ใน mPFC ของหนู RHA และ RLA ตามลำดับ ANOVA แบบแฟคทอเรียลสำหรับมาตรการซ้ำ ๆ ที่ทำกับค่าข้างต้นหลังจากการแปลงลอการิทึมเผยให้เห็นความแตกต่างอย่างมีนัยสำคัญในระดับฐานของโดปามีนระหว่างหนูที่มีเพศสัมพันธ์กับหนูที่มีประสบการณ์ทางเพศ (F(1,44,132) = 26.05, P <0.001) และในระดับพื้นฐานของ DOPAC (F(1,44,132) = 63.36, P <0.001) และของ NA (F(1,44,132) = 42.14, P <0.001) ระหว่างหนู RHA และ RLA (เก็บตัวอย่างสี่ตัวสุดท้ายก่อนนำตัวเมียไปขังในกรงเล็ก) พบค่าที่คล้ายกันเมื่อพิจารณาเฉพาะหนูที่มีเพศสัมพันธ์กับการหลั่งในระหว่างการทดลองด้วยเครื่องไตเทียม (ตารางที่ 3) ในกรณีนี้แฟคทอเรียล ANOVAs สำหรับมาตรการซ้ำ ๆ ที่ทำกับค่าหลังจากการแปลงลอการิทึมเปิดเผยความแตกต่างอย่างมีนัยสำคัญในระดับฐานของโดปามีนระหว่างหนูที่มีเพศสัมพันธ์และหนูที่มีประสบการณ์ทางเพศ (F(1,33,99) = 26.97, P <0.001) และในระดับพื้นฐานของ DOPAC (F(1,33,99) = 42.95, P <0.001) และของ NA (F(1,33,99) = 27.63, P <0.001) ระหว่าง RHA และ RLA หนู
ความเข้มข้นของโดปามีนนอกเซลล์, DOPAC และ Noradrenaline ใน mPFC dialysates จากNaïveทางเพศและ RHA และ RLA หนูที่มีประสบการณ์จะเปลี่ยนไปตามกิจกรรมทางเพศที่ต่างกัน
การปรากฏตัวของเพศหญิงที่ไม่สามารถเข้าถึงได้ในกรงขนาดเล็กด้านในและการมีเพศสัมพันธ์โดยตรงต่อมาเพิ่มความเข้มข้นของ dopamine extracellular, DOPAC และ NA ใน mPFC dialysates ที่ได้รับจากทั้งเพศผู้และหนูที่มีประสบการณ์ RHA และ RLA สองบรรทัดหนูและระดับของเงื่อนไขประสบการณ์ไม่ว่าจะเป็นเมื่อพิจารณาสัตว์ทดลองทั้งหมดหรือเฉพาะที่เกี่ยวข้องกับการพุ่งออกมา (รูปที่ 2) ในความเป็นจริงการวิเคราะห์ความแปรปรวนสองทางของ AUCs ของความเข้มข้นของโดปามีน, DOPAC และ NA แสดงให้เห็นว่าเนื้อหาโดยรวมของโดปามีน, DOPAC และ NA ของการทดสอบทั้งหมดนั้นสูงกว่าหนู RHA อย่างมีนัยสำคัญทางเพศ มีประสบการณ์เมื่อเทียบกับหนูไร้เดียงสา (ดูตาราง 4 สำหรับค่า AUCs และตาราง 5 for F ค่านิยมและระดับนัยสำคัญ) ความแตกต่างเหล่านี้ได้รับการยืนยันเพิ่มเติมจากจุดหนึ่งไปยังอีกจุดหนึ่ง ANOVAs สำหรับมาตรการซ้ำ ๆ ของโดปามีน, DOPAC และปริมาณ NA ของทั้งสองเพศที่ไร้เดียงสาและเส้นหนูที่มีประสบการณ์ตลอดการทดลองซึ่งเผยให้เห็นผลกระทบที่สำคัญ และประสบการณ์×การโต้ตอบสั่งซื้อครั้งแรกของเวลา (ดูตาราง 6 for F ค่านิยมและระดับนัยสำคัญ)
ความเข้มข้นของโดปามีนนอกเซลล์, DOPAC และ Noradrenaline ใน mPFC dialysates ที่ได้รับจากการมีเพศสัมพันธ์หนูแรชและหนู RLA เปลี่ยนไปตามกิจกรรมทางเพศที่ต่างกัน
ในเพศที่ไร้เดียงสา RHA และ RLA หนูนอกเซลล์โดพามีน แต่ไม่ใช่ DOPAC เพิ่มขึ้นเป็นครั้งแรกเมื่อวางต่อหน้าหญิงที่ไม่สามารถเข้าถึงได้ในขณะที่หนูเพิ่มขึ้นเฉพาะในหนู RHA ในหนูRïnaïve, โดปามีนนอกเซลล์เพิ่มขึ้นอีกใน 15 นาทีแรกของการมีเพศสัมพันธ์ที่พบกับ DOPAC และ NA การเพิ่มขึ้นของโดปามีนนอกเซลล์, DOPAC และ NA กินเวลาตลอดเวลาของการสังวาส, ด้วยโดปามีน, DOPAC และ NA ถึงค่าสูงสุดที่ 60 นาที, 75 นาทีและ 75 นาทีตามลำดับ, จากนั้นออกมาเป็นเซลล์เสริม DOPAC และ NA ลดลงเป็นค่าที่คล้ายกับค่าฐานหลังจากลบตัวเมีย
ในทางตรงกันข้ามการเพิ่มขึ้นอย่างมีนัยสำคัญครั้งแรกและค่าสูงสุดในโดปามีนนอกเซลล์, DOPAC และ NA เกิดขึ้นในหนูหนูRïnaïve RLA ในช่วง 15 นาทีแรกของการมีเพศสัมพันธ์ อย่างไรก็ตามที่ความแปรปรวนจากหนู RHA ในหนู RLA extracellular dopamine, DOPAC และ NA มีแนวโน้มที่จะกลับไปที่ค่าคล้ายกับฐานแรกภายใน 30 นาทีแรกของการมีปฏิสัมพันธ์โดยตรงกับผู้หญิง (ดูรูปที่ 2A, C, E).
ในที่สุดระหว่างการสังวาสการเพิ่มขึ้นของโดปามีนนอกเซลล์มีขนาดใหญ่ขึ้นและคงอยู่ใน RHA ทางเพศสัมพันธ์มากกว่าหนู RLA (ดูรูปที่ 2A) ในขณะที่ไม่มีตัวเมียความเข้มข้นของพวกมันมีความคล้ายคลึงกันแม้ว่าจะไม่เหมือนกันในสองสายหนู (ดูด้านบนและตาราง 3) ที่น่าสนใจแม้ว่าจะไม่พบความแตกต่างระหว่างสองบรรทัดโรมันในเปอร์เซ็นต์ที่เพิ่มขึ้นของ DOPAC และในระดับที่น้อยกว่าความเข้มข้นของ NA ในระหว่างการปรากฏตัวของหญิงที่เปิดกว้างนั้นมีความแตกต่างมากกว่าสองเท่าในค่าสัมบูรณ์ของความเข้มข้น DOPAC และ NA พบว่าระหว่างสองสายหนูทั้งสองอยู่ในสภาพฐาน (ดูตาราง 3) และตลอดการทดสอบกับหญิงที่เปิดกว้าง (ดูรูปที่ 2C, E).
ความเข้มข้นของโดปามีนนอกเซลล์, DOPAC และ Noradrenaline ใน mPFC Dialysates ที่ได้รับจาก RHA และ RLA หนูที่มีประสบการณ์ทางเพศเปลี่ยนไปในช่วงกิจกรรมทางเพศแตกต่างกัน
ดังที่พบในหนูที่ไร้สมรรถภาพทางเพศ RHA และ RLA การปรากฏตัวของเพศหญิงที่ไม่สามารถเข้าถึงได้และการมีเพศสัมพันธ์โดยตรงต่อมานำไปสู่การเพิ่มขึ้นของ dopamine extracellular และ DOPAC ใน mPFC dialysate ที่ได้รับจาก RHA เพศชายและหนู RLA อย่างไรก็ตามในกรณีนี้ก็พบความแตกต่างที่เกี่ยวข้องกับหนูด้วยเช่นกันเมื่อพิจารณาสัตว์ทดลองทั้งหมดหรือในระดับที่น้อยกว่าเฉพาะสัตว์ที่มีเพศสัมพันธ์กับการหลั่ง 2) ในประสบการณ์ทางเพศของ RHA และ RLA หนูโดปามีนนอกเซลล์เพิ่มขึ้นครั้งแรกที่เกิดขึ้นกับหญิงที่ไม่สามารถเข้าถึงได้ หลังจากนั้นความเข้มข้นโดปามีนเพิ่มขึ้นในระหว่างมีเพศสัมพันธ์ในทั้งสองสายหนู ความเข้มข้นของโดปามีนถึงค่าสูงสุดหลังจาก 45 – 60 นาทีของการมีเพศสัมพันธ์และลดลงอย่างช้า ๆ ไปสู่ค่าฐานในตอนท้ายของการทดสอบแบบมีส่วนร่วม (ดูรูปที่ 2B) ในทำนองเดียวกันกับหนูที่ไร้สมรรถภาพทางเพศความเข้มข้นของโดปามีนนอกเซลล์สูงกว่าในหนูที่มีประสบการณ์ทางเพศสัมพันธ์เมื่อเปรียบเทียบกับหนู RLA ส่วนใหญ่อยู่ในส่วนของ aliquots ของ dialysate ที่รวบรวมในช่วงส่วนกลางของระยะการสังวาส (หลัง 30 – 45 นาที) ความเข้มข้นของ DOPAC เพิ่มขึ้นในระหว่างการปรากฏตัวของตัวเมียที่ไม่สามารถเข้าถึงได้ในหนู RHA ในขณะที่มันเพิ่มขึ้นในระดับที่ใกล้เคียงกันในทั้งสองสายหนูในระหว่างการมีเพศสัมพันธ์ (ค่าสูงสุดที่ 75 – 90 นาที) และมีแนวโน้มที่จะกลับไป การทดสอบ copulatory (ดูรูปที่ 2D) ความเข้มข้นของ NA เพิ่มขึ้นทั้งในหนูที่มีเพศสัมพันธ์หลังจากการแนะนำของหญิงถึงค่าสูงสุดที่ 90 min ใน RHA rats และที่ 45 min ใน RLA rats (ดูรูปที่ 2F) ที่ความแปรปรวนจากโดปามีน (ดูด้านบน) พบความแตกต่างที่คล้ายกันในการเพิ่มความเข้มข้นของ DOPAC และ NA ในหนูที่มีประสบการณ์ทางเพศสัมพันธ์จากทั้งสองสายเมื่อพิจารณาสัตว์ทุกตัวและสัตว์ที่พุ่งออกมาเท่านั้น อย่างไรก็ตามที่พบในหนูหนูที่ไม่มีเพศสัมพันธ์ถึงแม้ว่าเปอร์เซ็นต์ความเข้มข้นของ DOPAC และ NA เพิ่มขึ้นมีความคล้ายคลึงกัน แต่ความแตกต่างอย่างมีนัยสำคัญในค่าสัมบูรณ์ของ DOPAC และ NA พบระหว่างหนูที่มีประสบการณ์ทางเพศ RHA และ RLA ตลอดการทดสอบ (เช่นก่อนระหว่างและหลัง การปรากฏตัวของเพศหญิงที่รับได้) โดยที่หนู RHA แสดงค่าสูงกว่าหนู RLA (ดูตาราง 3 สำหรับค่าฐานและตัวเลข 2D, F สำหรับค่าตลอดการทดสอบ)
ประสบการณ์ทางเพศมีอิทธิพลต่อการเปลี่ยนแปลงของความเข้มข้นของโดพามีนและนอร์ราดีนลีนใน mPFC dialysates ที่ได้รับจาก RHA และ RLA หนูในระหว่างกิจกรรมทางเพศ
การเปรียบเทียบความเข้มข้นของโดปามีนนอกเซลล์และ NA ในหนูที่มีเพศสัมพันธ์และมีประสบการณ์ RHA และ RLA แสดงให้เห็นว่าประสบการณ์ทางเพศเปลี่ยนความเข้มข้นของโดปามีนนอกเซลล์และในระดับที่น้อยกว่าของ NA ในทั้งหนูทดลอง หรือผู้ที่มีเพศสัมพันธ์กับการพุ่งออกมาเท่านั้น (ตัวเลข 2A, B, E, F) ดังนั้นความแตกต่างอย่างมีนัยสำคัญเกิดขึ้นในค่าฐานของโดปามีนระหว่างหนูที่มีเพศสัมพันธ์กับหนูที่มีประสบการณ์ของทั้งสองสาย (ดูตารางที่ 3) และค่าโดปามีนในระหว่างกิจกรรม copulatory โดยทั่วไปสูงกว่าในหนูที่มีประสบการณ์ทางเพศสัมพันธ์ RHA และ RLA เมื่อเปรียบเทียบกับหนูที่ไร้เดียงสาของพวกเขา (ดูรูปที่ 2B) นอกจากนี้ในหนู RHA ที่มีประสบการณ์ทางเพศแม้ว่าพบโดปามีนเพิ่มขึ้นในระหว่างที่มีหญิงที่ไม่สามารถเข้าถึงได้ตามรูปแบบชั่วคราวที่เห็นในหนูหนูเพศสัมพันธ์ แต่พบความแตกต่างในรูปแบบชั่วคราวในระหว่างการสังวาส ในความเป็นจริงในกรณีนี้ที่ความแปรปรวนจากสิ่งที่สังเกตได้ในหนูหนู RHA ทางเพศ (ยอดหนึ่งหลักที่ 60 นาที) พบค่าสูงสุดสองหลักแรกหลังจาก 15 นาทีและครั้งที่สองหลังจาก 45 นาทีของการมีเพศสัมพันธ์ หลังจากการเพิ่มครั้งล่าสุดนี้ค่าโดปามีนมีแนวโน้มที่จะกลับไปเป็นค่าฐาน (ดูรูปที่ 2B).
พบภาพที่คล้ายกันในหนูที่ไร้สมรรถภาพทางเพศกับหนู RLA ที่มีประสบการณ์ ในความเป็นจริงคล้ายกับสิ่งที่พบในหนู RHA เช่นกันในหนู RLA ที่มีประสบการณ์รูปแบบชั่วคราวที่มีลักษณะเพิ่มขึ้นสามหลักในการค้นพบโดปามีนนอกเซลล์: คนแรกที่มีเพศหญิงไม่สามารถเข้าถึงได้และอีกสองคนระหว่างการสังเวยคือ 15 min และ 60 นาทีของการมีเพศสัมพันธ์ (ดูรูปที่ 2A, B) โดยทั่วไปแล้วประสบการณ์ทางเพศดูเหมือนจะทำให้เกิดการเปลี่ยนแปลงที่ยั่งยืนมากขึ้นในรูปแบบชั่วคราวของการปลดปล่อยโดปามีนในทั้งสองหนูเมื่อผ่านจากเพศที่ไร้เดียงสาไปสู่สภาพที่มีประสบการณ์
ดังแสดงในรูปที่ 2C-Fตรวจพบความแตกต่างเพียงเล็กน้อยในความเข้มข้นของ DOPAC และ NA ระหว่างหนูที่ไร้เดียงสากับหนูที่มีประสบการณ์ของทั้งสองสาย อย่างไรก็ตามแนวโน้มที่ไม่สำคัญในนานอกเซลล์ต่อค่าที่สูงขึ้นในประสบการณ์เมื่อเปรียบเทียบกับหนูnaïveถูกพบในฐาน (ก่อนการแนะนำของเพศหญิงที่เปิดกว้างดูตาราง 3) และในจำนวน NA โดยรวมตามที่เปิดเผยโดยค่า AUCs (ดูตาราง 4) คำนวณจากความเข้มข้นนอกเซลล์ที่ได้จากการทดสอบทั้งหมด (ก่อนและระหว่างการปรากฏตัวของหญิงที่เปิดกว้าง) ในที่สุดรูปแบบชั่วคราวของความเข้มข้นของ NA นั้นคล้ายคลึงกันมากเมื่อเปรียบเทียบหนูที่มีประสบการณ์ของทั้งสองสายหนูกับคู่ที่ไร้เดียงสาของพวกเขาแสดงว่าประสบการณ์ทางเพศสามารถนำไปสู่การเพิ่มขึ้นของ extracellular NA มากกว่าการเปลี่ยนแปลงรูปแบบการเพิ่มขึ้น 2E, F).
การเปลี่ยนแปลงในความเข้มข้นของสาร Dopamine และ Noradrenaline ใน mPFC Dialysates ที่ได้รับจาก RHA และ RLA หนูที่มีประสบการณ์ทางเพศและมีประสบการณ์ร่วมกับการเปลี่ยนแปลงพฤติกรรมทางเพศ
ความแตกต่างของความเข้มข้นของโดปามีนนอกเซลล์, DOPAC และ NA ใน dialysates ที่ได้รับจาก mPFC พบได้ทั้งในหนูที่มีอาการทางเพศและไม่มีประสบการณ์ RHA และ RLA ในระหว่างการทดสอบ 2) เกิดขึ้นพร้อมกับการปรับเปลี่ยนพารามิเตอร์ทางเพศที่แตกต่างกันที่วัดได้ระหว่างสองช่วงหลักของการทดลอง (เช่นหญิงที่ไม่สามารถเข้าถึงได้และมีให้สำหรับผู้ชายตามลำดับ) ซึ่งรวมถึงกลุ่มคนอื่น ๆ 3A, B) เมานต์ (ตัวเลข 3C, D) intromissions (ตัวเลข 3E, F) และอุทาน (ตัวเลข) 3G, H) ความแตกต่างเหล่านี้ถูกค้นพบเมื่อพิจารณาจากสัตว์ทดลองทั้งหมดหรือที่มีการหลั่งเพื่อการหลั่งเท่านั้น ในความเป็นจริงการวิเคราะห์เบื้องต้นของค่า AUCs ของพารามิเตอร์เหล่านี้โดยการวิเคราะห์ความแปรปรวนสองทาง (ดูตาราง 4 สำหรับค่า AUCs และตาราง 5 for F ค่าและระดับนัยสำคัญ) ตามด้วยการวิเคราะห์แบบจุดต่อจุดของค่าของพารามิเตอร์เหล่านี้พร้อมการทดสอบโดย ANOVA แบบ factorial สำหรับมาตรการซ้ำ ๆ เผยให้เห็นผลกระทบที่สำคัญของเส้น, ประสบการณ์, เวลา, ลำดับที่สำคัญครั้งแรก×เวลาและประสบการณ์ และการโต้ตอบลำดับที่สองที่สำคัญสำหรับการเมาท์ (ดูตาราง 6 for F ค่านิยมและระดับนัยสำคัญ) สอดคล้องกับการศึกษาก่อนหน้า (Sanna และคณะ, 2015): (i) หนูที่ไร้สมรรถภาพทางเพศหนู RHA แสดงให้เห็นว่าการแข็งตัวของอวัยวะเพศชายไม่ติดต่อ, เมานต์, ความไม่แน่นอนและอุทานมากกว่า RLA (ii) หนูที่มีประสบการณ์ทางเพศสัมพันธ์ RHA แสดงให้เห็นว่ามีจำนวนการแข็งตัวของอวัยวะเพศชายที่ไม่ติดต่อการสัมผัสที่สูงขึ้นและการหลั่งเร็วกว่าคู่ RLA และ (iii) หนูที่มีประสบการณ์ทางเพศของทั้งสองสายและโดยเฉพาะอย่างยิ่งหนู RLA แสดงให้เห็นว่าการแข็งตัวของอวัยวะเพศชายที่ไม่ติดต่อกันมากขึ้น ในความเป็นจริงในขณะที่หนู RHA ส่วนใหญ่ของการเปลี่ยนแปลงที่เกิดจากประสบการณ์ทางเพศเกิดขึ้นในช่วงเวลามากกว่าความถี่ของพารามิเตอร์พฤติกรรม (เช่นหนูที่มีประสบการณ์ copulated ส่วนใหญ่ในช่วงครึ่งแรกของการทดสอบในขณะที่หนูnaïveแสดงให้เห็น แนวโน้มการมีเพศสัมพันธ์ที่คล้ายกันตลอดการทดสอบ) ในหนู RLA การปรับปรุงพฤติกรรมทางเพศโดยทั่วไปที่โดดเด่นมากขึ้นส่วนใหญ่ในการปฏิบัติทางเพศถูกผลิตโดยประสบการณ์ทางเพศตามที่เปิดเผยโดยการเปลี่ยนแปลงของค่าของพารามิเตอร์ copulatory ในหนู RLA ที่มีประสบการณ์ คู่หูไร้เดียงสา
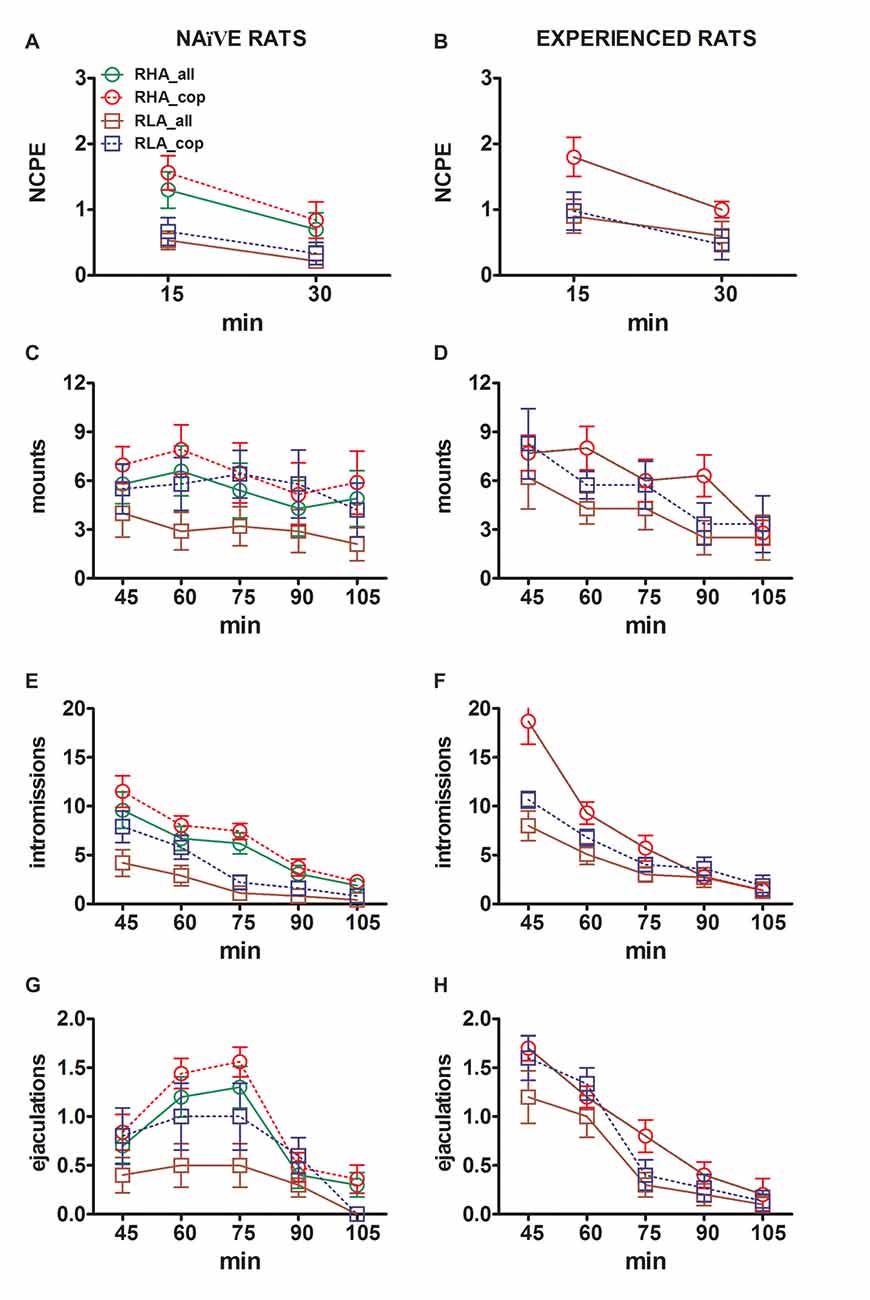 รูป 3. การเปลี่ยนแปลงจำนวนของการแข็งตัวของอวัยวะเพศชายแบบไม่สัมผัส (NCPE), การติดตั้ง, การเกิดอาการมึนงงและการหลั่งจากบันทึกทางเพศ (A, C, E, G) และหนูที่มีประสบการณ์ RHA และ RLA (B, D, F, H) ในระหว่างการทดลอง microdialysis รายงานในรูปที่ 2. เงื่อนไขการทดลองทั้งหมดเหมือนกันกับที่อธิบายไว้ในคำอธิบายแผนภูมิ 2. ค่าหมายถึง± SEM ของค่าที่ได้รับจากหนู 12 ทั้งหมดต่อกลุ่ม (รวมถึงที่ได้รับคะแนนเต็มเวลา RHA = เส้นสีเขียวทึบ, RLA = เส้นสีน้ำตาลทึบ) หรือผู้ที่เกี่ยวข้องกับการพุ่งออกมาเท่านั้น (RHA = เส้นสีแดงประ ; RLA = เส้นสีน้ำเงินเส้นประ) ในหนูที่มีประสบการณ์ RHA สัตว์ทุกตัวจะมีเพศสัมพันธ์และค่าเหมือนกัน เนื่องจากไม่มีการโต้ตอบที่สำคัญระหว่างปัจจัยบรรทัดหนู (RHA กับ RLA) ปัจจัยระดับประสบการณ์ทางเพศ (naïveกับประสบการณ์) และเวลาถูกพบโดยการวิเคราะห์ข้อมูลด้วย ANOVAs สองทางดังที่แสดงในตาราง 6, โพสต์เฉพาะกิจ ไม่มีการรายงานการเปรียบเทียบตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ", "สถิติ"
รูป 3. การเปลี่ยนแปลงจำนวนของการแข็งตัวของอวัยวะเพศชายแบบไม่สัมผัส (NCPE), การติดตั้ง, การเกิดอาการมึนงงและการหลั่งจากบันทึกทางเพศ (A, C, E, G) และหนูที่มีประสบการณ์ RHA และ RLA (B, D, F, H) ในระหว่างการทดลอง microdialysis รายงานในรูปที่ 2. เงื่อนไขการทดลองทั้งหมดเหมือนกันกับที่อธิบายไว้ในคำอธิบายแผนภูมิ 2. ค่าหมายถึง± SEM ของค่าที่ได้รับจากหนู 12 ทั้งหมดต่อกลุ่ม (รวมถึงที่ได้รับคะแนนเต็มเวลา RHA = เส้นสีเขียวทึบ, RLA = เส้นสีน้ำตาลทึบ) หรือผู้ที่เกี่ยวข้องกับการพุ่งออกมาเท่านั้น (RHA = เส้นสีแดงประ ; RLA = เส้นสีน้ำเงินเส้นประ) ในหนูที่มีประสบการณ์ RHA สัตว์ทุกตัวจะมีเพศสัมพันธ์และค่าเหมือนกัน เนื่องจากไม่มีการโต้ตอบที่สำคัญระหว่างปัจจัยบรรทัดหนู (RHA กับ RLA) ปัจจัยระดับประสบการณ์ทางเพศ (naïveกับประสบการณ์) และเวลาถูกพบโดยการวิเคราะห์ข้อมูลด้วย ANOVAs สองทางดังที่แสดงในตาราง 6, โพสต์เฉพาะกิจ ไม่มีการรายงานการเปรียบเทียบตามที่อธิบายไว้ในส่วน "วัสดุและวิธีการ", "สถิติ"
การสนทนา
การศึกษาครั้งนี้ยืนยันและขยายผลการศึกษาก่อนหน้านี้แสดงให้เห็นว่าหนู RHA และ RLA ซึ่งแสดงรูปแบบการเผชิญปัญหาที่แตกต่างกันอย่างเห็นได้ชัดเพื่อตอบสนองต่อสภาวะ aversive (Driscoll และBättig, 1982; Giorgi et al., 2003a; Steimer และ Driscoll, 2003) แสดงรูปแบบพฤติกรรมที่แตกต่างในการปรากฏตัวของเพศหญิงที่ไม่สามารถเข้าถึงได้และในระหว่างการทดสอบ copulatory แบบดั้งเดิม (Sanna และคณะ, 2014a,b, 2015) ดังนั้นการศึกษานี้แสดงให้เห็นว่าหนูที่มีเพศสัมพันธ์แบบไร้ RHA (ไม่เคยสัมผัสกับผู้หญิงที่เปิดกว้าง) มีแรงจูงใจทางเพศที่สูงขึ้นและสมรรถภาพทางเพศที่ดีกว่าเมื่อเปรียบเทียบกับหนูเพศเมีย RLA ที่ไร้สมรรถภาพทางเพศเช่นหนู RHA การแข็งตัวของอวัยวะเพศชายเหนี่ยวนำให้เกิดการพิจารณาดัชนีของความเร้าอารมณ์ทางเพศ; Sachs et al., 1994; Sachs, 2000) และการเปลี่ยนแปลงทางเพศที่แตกต่างกันในพารามิเตอร์ร่วมหลายอย่างที่พบในการศึกษาก่อนหน้า (Sanna และคณะ, 2014a,b, 2015) ความแตกต่างเหล่านี้ยังคงอยู่ในสายหนูโรมันทั้งสองหลังจากได้รับประสบการณ์ทางเพศนั่นคือหลังจากการรักษาเสถียรภาพของพฤติกรรมทางเพศด้วยการเปิดรับห้าครั้งและการทดสอบการมีเพศสัมพันธ์กับผู้หญิงที่เปิดกว้าง การศึกษาครั้งนี้ยังแสดงให้เห็นเป็นครั้งแรกว่าในทั้งสองสาย RHA และ RLA หนูทั้งในสภาพไร้เดียงสาหรือมีประสบการณ์การแข็งตัวของอวัยวะเพศที่ไม่ติดต่อและการมีเพศสัมพันธ์เกิดขึ้นพร้อมกันกับการเพิ่มความเข้มข้นของโดปามีน ) และ NA ใน dialysates ที่ได้รับโดย intracerebral microdialysis จาก mPFC ของทั้งสองสายหนูโรมัน ที่น่าสนใจการเพิ่มขึ้นของโดปามีนนอกเซลล์และ NA ใน mPFC นั้นสัมพันธ์กับความแตกต่างทั้งในฟีโนไทป์ของหนูหรือในระดับประสบการณ์ทางเพศ ดังนั้นตามที่แสดงอย่างกว้างขวางในส่วนผลลัพธ์หนูที่ไร้สมรรถภาพทางเพศและผู้ที่มีประสบการณ์แสดงให้เห็นว่าการเพิ่มขึ้นของโดปามีนนอกเซลล์ในระยะยาวและระยะยาวจะเพิ่มขึ้นและยาวนานขึ้นเมื่อเปรียบเทียบกับคู่ RLA ทั้งในช่วงระยะเวลา ยิ่งไปกว่านั้นหนูที่มีประสบการณ์ทางเพศของทั้งสองสายโรมันแสดงว่าโดปามีนนอกเซลล์เพิ่มขึ้นและติดทนนานใน mPFC ทั้งในระยะของพฤติกรรมทางเพศทั้งสองเมื่อเปรียบเทียบกับคู่ที่ไร้เดียงสาของพวกเขา ในที่สุดความแตกต่างข้างต้นทั้งหมดในโดปามีนนอกเซลล์และ NA ใน mPFC นั้นมีความสัมพันธ์กับความแตกต่างของพารามิเตอร์พฤติกรรมของพฤติกรรมทางเพศเช่นหนูที่มีประสบการณ์ทางเพศของทั้งสองสายโรมันแสดงแรงจูงใจทางเพศที่สูงขึ้น และหนูที่ไร้สมรรถภาพทางเพศและมีประสบการณ์หนู RHA แสดงแรงจูงใจทางเพศที่สูงขึ้นและการแสดงออกที่ดีขึ้นเมื่อเปรียบเทียบกับหนูที่ไร้สมรรถภาพทางเพศ
สำหรับการเพิ่มขึ้นของโดปามีนนอกเซลล์ที่พบใน mPFC dialysate ในระหว่างกิจกรรมทางเพศการเพิ่มขึ้นครั้งแรกเกิดขึ้นในนาทีแรก 15 หลังจากการแนะนำของเพศหญิงที่รับได้ทางเพศในอุปกรณ์จับคู่เมื่อเพศหญิงไม่สามารถเข้าถึงได้ ในสภาพนี้ไม่แข็งตัวอวัยวะเพศชายติดต่อ การเพิ่มขึ้นของโดปามีนนอกเซลล์ที่สองและใหญ่กว่านั้นเกิดขึ้นในช่วง 15 นาทีแรกหลังจากที่เพศเมียสามารถเข้าถึงได้เพื่อการสังวาส ที่น่าสนใจในขณะที่ภาวะโดปามีนนอกเซลล์เพิ่มขึ้นส่วนใหญ่ในส่วนแรกของการทดสอบกับหญิงที่สามารถเข้าถึงได้โดยมีแนวโน้มที่จะกลับไปที่ค่าฐานในส่วนที่สองในโดปามีนสภาพประสบการณ์ทางเพศยังคงเพิ่มขึ้นตลอดการทดสอบ หลังจาก 45 – 60 ขั้นต่ำของการมีเพศสัมพันธ์ในทั้งสองบรรทัดหนูโรมัน
นอกเหนือจากความแตกต่างข้างต้นในโดปามีนนอกเซลล์ในช่วงระยะกิจกรรมทางเพศที่แตกต่างกันระหว่าง RHA และ RLA ในสภาพทางเพศและประสบการณ์พบว่าการศึกษานี้แสดงให้เห็นถึงความแตกต่างที่สำคัญและคาดไม่ถึงในความเข้มข้นของสาร DOPAC ค่าฐาน) และหลังการเปิดตัวของหญิงเปิดกว้างในอุปกรณ์จับคู่ ดังนั้นค่าฐานของ extracellular DOPAC ใน mPFC dialysate พบในหนู RHA จะมีค่า 2.6-fold สูงกว่าที่พบในหนู RLA ความแตกต่างระหว่างหนู RHA และ RLA นี้เกิดขึ้นและคงอยู่ในสภาวะไร้สมรรถภาพทางเพศและมีประสบการณ์ทั้งๆที่มีความแตกต่างเล็กน้อยในค่าฐานของโดปามีนนอกเซลล์ใน mPFC dialysates ที่ได้รับจากหนูโรมันสองสายในเงื่อนไขการทดลองเหล่านี้ เมื่อนำมารวมกันความแตกต่างเหล่านี้อาจบ่งบอกถึงการเปลี่ยนแปลงโดปามีนที่แตกต่างกันที่ระดับของ mPFC ระหว่างสองสายหนูโรมัน ดังนั้นความเข้มข้นของ DOPAC มักจะสะท้อนปริมาณของโดปามีนที่ปล่อยออกมาและกลับคืนมาอีกครั้งโดยขั้วประสาทโดปามีนและเปลี่ยนเป็น DOPAC โดย monoaminooxydase (MAO; Carlsson, 1975) ดังนั้นความเข้มข้นของฐาน DOPAC ที่สูงกว่าที่พบในหนู RHA ชี้ให้เห็นว่าปริมาณโดปามีนในปริมาณที่สูงกว่านั้นถูกปล่อยออกมาและกลับคืนสู่สภาพเดิมโดยปลายประสาทโดปามีเซอร์คิคใน mPFC ของหนู RHA เช่นกิจกรรมพื้นฐานที่สูงขึ้น หนู ระดับโดปามีนที่สูงกว่า mesocortical dopaminergic ในหนู RHA นั้นยังได้รับการสนับสนุนจากความเข้มข้นของโดปามีนที่เพิ่มขึ้นในระหว่างกิจกรรมทางเพศใน mPFC dialysate จากหนู RHA ที่มีความสัมพันธ์ทางเพศ ในเรื่องนี้มันเกี่ยวข้องกับการระลึกว่าน้ำโดปามีนที่สูงกว่านั้นถือเป็นความรับผิดชอบอย่างน้อยก็ในบางส่วนของลักษณะที่แตกต่างและตรงข้ามกับพฤติกรรมหลายอย่างในหนู RHA และ RLA ดังนั้น RHA rats เป็น copers ที่ใช้งานสูงหุนหันพลันแล่นผู้แสวงหาความรู้สึกแปลกใหม่และมีแนวโน้มที่จะบริโภคและใช้ยาเสพติดหลายชั้นของการติดยาเสพติดในขณะที่ RLA rats เป็น copers ปฏิกิริยา hyperemotional และมีแนวโน้มที่จะพัฒนาอาการเหมือนซึมเศร้า (Zeier และคณะ 1978; Giorgi et al., 1994, 2003b, 2007; Corda et al., 1997, 2014; Escorihuela และคณะ, 1999; Steimer และ Driscoll, 2003; Lecca และคณะ, 2004; Giménez-Llort et al., 2005; Carrasco et al., 2008; Fattore et al., 2009; Moreno และคณะ 2010; Coppens et al., 2012; Díaz-Moránและคณะ, 2012; Sabariego และคณะ, 2013; Manzo et al., 2014a,b; Oliveras et al., 2015) รวมถึงแรงจูงใจทางเพศที่สูงขึ้นและสมรรถภาพทางเพศที่ดีขึ้นของหนูที่ไร้สมรรถภาพทางเพศและหนู RHA ที่มีประสบการณ์กับหนู RHA (Sanna และคณะ, 2015) การเพิ่มขึ้นของความเข้มข้นของโดปามีนนอกเซลล์ที่พบใน dialysate จาก mPFC ของหนูที่มีเพศสัมพันธ์และ RHA ที่มีประสบการณ์จะมีลักษณะคล้ายกับความเข้มข้นของโดปามีนนอกเซลล์ที่สูงกว่าที่พบใน dialysate จากนิวเคลียส (Sanna และคณะ, 2015) อย่างไรก็ตามที่ความแปรปรวนจาก mPFC ระดับพื้นฐานของสารกระตุ้นโดปามีนและ DOPAC ในไดอะลอกซิสจากนิวเคลียส accumbens ของหนูทั้งสองที่มีเพศสัมพันธ์และมีประสบการณ์ RHA และ RLA ไม่แตกต่างกันอย่างมีนัยสำคัญ (Sanna และคณะ, 2015) ดังนั้นปริมาณโดปามินอจิกที่มี mesolimbic สูงกว่าในนิวเคลียส accumbens ของหนูที่มีเชื้อเอชไอวีและมีประสบการณ์ RHA อาจถูกสรุปโดยความเข้มข้นของโดปามีนนอกเซลล์ที่สูงกว่าที่พบใน dialysates จากนิวเคลียส accumbens ของ RHA เมื่อเทียบกับคู่ RLA เพศหญิงและไม่ใช่จากกิจกรรม dopaminergic ฐานที่แตกต่างกันพบว่าระดับความเข้มข้นของ DOPAC ในหนู RHA และ RLA ตามลำดับสูงกว่าและต่ำกว่าตามลำดับพบว่าเกิดขึ้นใน mPFC ในการศึกษานี้ นี่ก็สอดคล้องกับผลลัพธ์ของรายงานก่อนหน้านี้แสดงให้เห็นว่าระดับโดปามีนนอกเซลล์ใน dialysate จาก mPFC ของหนู RHA พบสูงกว่าหนู RLA ในขณะที่ระดับโดปามีนที่คล้ายกันพบใน dialysate จากเปลือกของนิวเคลียส accumbens ของหนูทั้ง RHA และ RLA ภายใต้เงื่อนไขการทดลองที่แตกต่างกัน (D'Angio et al., 1988; Scatton และคณะ, 1988; Willig et al., 1991; Giorgi et al., 2003a, 2007) แม้จะมีความแตกต่างข้างต้นระหว่าง mPFC และนิวเคลียส accumbens ประสบการณ์ทางเพศดูเหมือนว่าจะผลิตใน mPFC เพิ่มขึ้นอีกต่อไปในรูปแบบชั่วคราวของการปลดปล่อยโดปามีนในทั้งสองเส้นโรมันหนูเมื่อผ่านจากไร้เดียงสาทางเพศถึงสภาพที่มีประสบการณ์ มักจะสูงกว่าในหนู RHA มากกว่าหนู RLA สิ่งนี้ค่อนข้างแตกต่างจากสิ่งที่พบในนิวเคลียส accumbens ซึ่งประสบการณ์ทางเพศมีแนวโน้มที่จะเปลี่ยนสารกระตุ้นโดปามีนนอกเซลล์เพิ่มขึ้นเป็นส่วนแรกของการทดสอบร่วมในหนู RHA ในขณะที่เพิ่มขึ้นตลอดการทดสอบการมีเพศสัมพันธ์ทั้งหมดในหนู RLASanna และคณะ, 2015).
ตามความรู้ของเราการศึกษานี้ยังแสดงให้เห็นเป็นครั้งแรกที่ไม่เพียง แต่ต้องเพิ่มโดปามีนนอกเซลล์ แต่ยังเพิ่มการทำงานของเซลนอกเซลล์ในระหว่างกิจกรรมทางเพศใน dialysate จาก mPFC ของหนูทั้งสอง RHA และ RLA เช่นเดียวกับโดปามีน ความแตกต่างที่เกี่ยวข้องกับสายหนูโรมันและประสบการณ์ทางเพศ โดยเฉพาะอย่างยิ่งที่ความแปรปรวนจากระดับโดพามีนพื้นฐานที่พบว่าคล้ายกันมากใน dialysate จาก mPFC ของหนูทั้ง RHA และ RLA ระดับพื้นฐานของ extracellular NA ใน dialysate จาก mPFC ของ RHA สูงกว่าอย่างมีนัยสำคัญ (ประมาณ 2,5-fold ) มากกว่าหนู RLA เหตุผลของความแตกต่างเหล่านี้ไม่เป็นที่รู้จัก แต่อย่างที่กล่าวไว้ข้างต้นสำหรับโดปามีนและ DOPAC ผลลัพธ์เหล่านี้อาจบ่งชี้ว่าเสียง noradrenergic พื้นฐานที่สูงกว่ามีอยู่อย่างน้อยใน mPFC cortex ของหนู RHA เมื่อเปรียบเทียบกับหนู RLA อาจมีส่วนร่วมในลักษณะที่แตกต่างและมักจะตรงกันข้ามกับพฤติกรรมที่มีอยู่ในหนู RHA และ RLA ตามที่กล่าวไว้ข้างต้นสำหรับโดปามีน การสนับสนุนเพิ่มเติมสำหรับเสียง noradrenergic ที่สูงขึ้นในหนู RHA มากกว่าหนู RLA ใน mPFC มาจากผลลัพธ์ที่แสดงว่าความแตกต่างในระดับ NA นอกเซลล์พื้นฐานยังคงอยู่ในระหว่างกิจกรรมทางเพศทั้งในสภาพทางเพศและประสบการณ์ที่เพิ่มขึ้น mPFC dialysate ใน RHA หนูเมื่อสัมผัสกับตัวเมียที่ไม่สามารถเข้าถึงได้และในช่วงระยะเวลาการมีเพศสัมพันธ์ทั้งหมดมากกว่าใน RLA หนูซึ่งระดับ NA เพิ่มขึ้นเฉพาะใน 15 นาทีแรกของระยะเวลาการมีเพศสัมพันธ์หลังจากที่ระดับ NA กลับสู่ค่าฐาน . การเปลี่ยนแปลงของระดับ NA ใน extracellular เหล่านี้จะเห็นได้ในสภาพทางเพศและประสบการณ์ แต่การเพิ่มขึ้นของระดับ NA นั้นสูงกว่าในหนูที่มีประสบการณ์ทางเพศสัมพันธ์ RHA และ RLA เมื่อเปรียบเทียบกับคู่ที่ไม่ได้มีเพศสัมพันธ์ ดังนั้นประสบการณ์ทางเพศทำให้เกิดความสามารถในการกระตุ้นกิจกรรม noradrenergic เพิ่มเติมใน mPFC ของ RHA หนูและในระดับที่น้อยกว่าของหนู RLA ตามที่ได้กล่าวถึงโดปามีนแล้ว เสียงประสานที่สูงกว่า noradrenergic และ dopaminergic ร่วมกันใน mPFC ของหนู RHA เมื่อเปรียบเทียบกับหนู RLA สมควรได้รับความคิดเห็นบางส่วน ในความเป็นจริงมีรายงานว่ากิจกรรม noradrenergic ใน mPFC สามารถส่งผลกระทบต่อการปลดปล่อยโดปามีนผ่านกลไกต่าง ๆ (เช่นดู Carboni และคณะ, 1990, 2006; Gresch et al., 1995; Westernik และคณะ, 1998 และการอ้างอิงในนั้น) หนึ่งในสิ่งเหล่านี้เกี่ยวข้องกับกิจกรรมของ NET ซึ่งแสดงให้เห็นถึงการผูกโดปามีนที่มีความสัมพันธ์ใกล้ชิดกับ NA ใน synaptosomes ที่ได้จาก homogenates ของ PFCฮอร์น 1973) หากสิ่งนี้เกิดขึ้นใน mPFC ในร่างกายโดปามีนที่ถูกปล่อยใน mPFC นั้นไม่เพียง แต่สามารถยึดกลับได้โดยโดปามินอจิก แต่ยังสามารถใช้กับขั้วประสาท noradrenergic ที่ความเร็วสูงกว่า NA สิ่งนี้อาจทำให้เกิดการเพิ่มขึ้นของการหลั่งโดปามีนจากขั้วโดปามีน (เช่นเสียงโดปามีนเพิ่มขึ้น) เพื่อชดเชยปริมาณโดปามีนที่ถูกลบออกจากแหว่ง synaptic; ดังนั้นเสียง noradrenergic ที่สูงกว่าที่พบในหนู RHA สามารถมีส่วนร่วมอย่างน้อยก็ในบางส่วนกับเสียงโดปามีนสูงกว่าที่พบใน mPFC ของหนู RHA เมื่อเปรียบเทียบกับหนู RLA การศึกษาเพิ่มเติมจะต้องตรวจสอบความเป็นไปได้นี้ โดยเฉพาะอย่างยิ่งความแตกต่างของกิจกรรม noradrenergic ที่ระบุในการศึกษานี้ใน mPFC ระหว่างหนู RHA และ RLA อาจมีบทบาทในการตอบสนองต่อการรักษาด้วยยากล่อมประสาทที่แตกต่างกัน ในความเป็นจริงหนู RHA ถือเป็นแบบจำลองของความยืดหยุ่นและ RLA หนูเป็นแบบจำลองของความอ่อนแอต่อภาวะซึมเศร้า ดังนั้นการรักษาแบบเฉียบพลันหรือเรื้อรังกับยากล่อมประสาทเช่น desipramine, fluoxetine และ chlorimipramine ทั้งหมดปรับปรุงการตอบสนองเชิงพฤติกรรมอย่างมีนัยสำคัญในการทดสอบว่ายน้ำบังคับในหนู RLA โดยไม่มีผลต่อการตอบสนองของหนู RHA (Piras et al., 2010, 2014).
ในที่สุดการศึกษานี้ยืนยันและขยายผลการวิจัยก่อนหน้านี้แสดงให้เห็นว่า mPFC มีส่วนร่วมในการควบคุมพฤติกรรมทางเพศแม้ว่าบทบาทที่แน่นอนของพื้นที่สมองนี้ในพฤติกรรมทางเพศอยู่ไกลจากความชัดเจน (Fernández-Guasti และคณะ, 1994; Agmo และ Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama และคณะ, 2003; Balfour et al., 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011) เมื่อเร็ว ๆ นี้การคัดเลือกเซลล์ที่ถูกยิงใน mPFC ในระหว่างการเข้าใกล้พฤติกรรมของหนูตัวผู้ที่มีต่อเพศหญิงที่ไม่สามารถเข้าถึงได้ทางเพศนั้นถูกแสดงโดยใช้กระบวนทัศน์การบันทึกเซลล์เดียวFebo, 2011) น่าสนใจเซลล์ประสาทที่ไม่ตอบสนองในระหว่างการแสดงออกครั้งแรกของผู้ชายกับหญิงที่เปิดกว้างไม่สามารถเข้าถึงได้กลายเป็นที่ใช้งานในช่วงการแสดงออกที่สองราวกับว่าประสบการณ์ก่อนหน้านี้สามารถทำให้เกิดการเปลี่ยนแปลงพลาสติกที่นำไปสู่การตอบสนองเชิงพฤติกรรม อย่างไรก็ตามรอยโรคด้วยกรดไอโซเทนิกของ mPFC พบว่าไม่สามารถเปลี่ยนพฤติกรรมทางเพศของหนูเพศผู้กับเพศหญิงที่รับได้ทางเพศ อย่างไรก็ตามความสามารถในการสร้างความเกลียดชังที่มีต่อกิจกรรมทางเพศเมื่อพบว่าถูกจับคู่กับสิ่งเร้า aversive ถูกยกเลิกอย่างสมบูรณ์ในสัตว์ที่มีแผลเหล่านี้ (Davis et al., 2010) สิ่งนี้ชี้ให้เห็นว่าการเปิดใช้งาน mPFC ในระหว่างพฤติกรรมทางเพศมีบทบาทในการบูรณาการข้อมูลภายนอกและภายในสำหรับการดำเนินการและการควบคุมพฤติกรรมที่มุ่งเป้าไปที่เป้าหมายมากกว่าในการแสดงออกของการตอบสนองโดยธรรมชาติต่อผู้สนับสนุนธรรมชาติ ไปที่และเกรซ 2005) มีความสำคัญมากกว่าสำหรับงานนี้การศึกษาหลายชิ้นแสดงให้เห็นว่าสมองมีส่วนเกี่ยวข้องกับแรงจูงใจทางเพศและพฤติกรรมทางเพศเช่นนิวเคลียส accumbens, หน้าท้องพื้นที่ tegmental, พื้นที่ preoptic อยู่ตรงกลาง, นิวเคลียสเตียงของ Stria terminalis, basolateral amygdala และ parvocellular subparafascicular thalamic จาก mPFC ระหว่างกิจกรรมทางเพศ (ดู Balfour et al., 2006) พื้นที่สมองเหล่านี้หลายแห่งมีส่วนร่วมในวงจรประสาทที่ซับซ้อนที่เกี่ยวข้องกับการควบคุมพฤติกรรมทางเพศจากแรงจูงใจทางเพศและการให้รางวัลกับสมรรถภาพทางเพศ วงจรนี้รวมถึงเซลล์ประสาท oxytocinergic ที่เกิดขึ้นใน PVN ของ hypothalamus (PVN) และโครงการไปยังพื้นที่ tegmental หน้าท้อง / mesorticamalcic, นิวเคลียส accumbens, ฮิปโปแคมปัส, amygdala, PFC, นิวเคลียสเตียงของ stria terminal, ไขกระดูก เซลล์ประสาทที่ยื่นออกมาจากพื้นที่หน้าท้องไปสู่นิวเคลียส accumbens และ PFC, incertohypothalamic dopaminergic dopaminergic เซลล์ประสาทและเซลล์ประสาท glutamatergic ที่มีส่วนร่วมในระดับท้องถิ่นและระบบในหลายพื้นที่ของวงจร (ดู Melis และ Argiolas, 1995, 2011; Melis et al., 2003, 2007, 2009, 2010; Succu et al., 2007, 2008, 2011) ดังนั้นการเพิ่มขึ้นของโดปามีนนอกเซลล์ (แต่ยังรวมถึงของ NA) ใน dialysates จาก mPFC ที่เห็นในระหว่างกิจกรรมทางเพศอาจบ่งชี้ว่า mPFC เป็นส่วนหนึ่งของวงจรนี้นั่นคือเซลล์ประสาทโดปามีนที่มีส่วนร่วมในการควบคุมกิจกรรมทางเพศด้วยกัน กับ noradrenergic, mesolimbic dopaminergic, oxytocinergic และ glutamatergic เซลล์ประสาทแม้ว่ามันดูเหมือนจะไม่จำเป็นอย่างยิ่งสำหรับการแสดงออกของพฤติกรรมทางเพศ (Davis et al., 2010) ตามสมมติฐานนี้การศึกษาครั้งนี้แสดงให้เห็นว่าความแตกต่างของโดปามีนและการปลดปล่อย NA ใน mPFC ระหว่างกิจกรรมทางเพศเกิดขึ้นพร้อมกับความแตกต่างของจำนวนการแข็งตัวของอวัยวะเพศชายแบบไม่สัมผัสและการเปลี่ยนแปลงพารามิเตอร์ร่วม ความอยากอาหารและระยะการบริโภคที่เกี่ยวข้องกับแรงจูงใจทางเพศที่สูงขึ้นและการแสดงออกที่ดีขึ้นทั้งในสายหนูโรมันและในสภาวะทางเพศและประสบการณ์
การอภิปรายที่กล่าวมาข้างต้นนั้นขึ้นอยู่กับการเพิ่มขึ้นของสารโดปามีนนอกเซลล์ที่รู้จักกันดีซึ่งเกิดขึ้นในนิวเคลียส accumbens ของหนูเพศผู้ในระหว่างกิจกรรมทางเพศ (Pfaus et al., 1990; Pleim et al., 1990; Pfaus และ Phillips, 1991; Damsma et al., 1992; Wenkstern และคณะ, 1993) อย่างไรก็ตามโดปามีนและสารสื่อประสาทอื่น ๆ อาจถูกปลดปล่อยใน mPFC (และในนิวเคลียส accumbens) ในเงื่อนไขการทดลองอื่น ๆ เช่นในระหว่างความเครียดหรือการเผชิญกับมัน (Thierry et al., 1976; Fadda et al., 1978) ซึ่งไม่เกี่ยวข้องกับพฤติกรรมทางเพศอย่างเคร่งครัด ดังนั้นหนึ่งอาจยืนยันว่าการเพิ่มโดปามีนที่พบในโดปามีนนอกเซลล์ใน mPFC (การศึกษานี้) หรือในนิวเคลียส accumbens (Sanna และคณะ, 2015) มีความสัมพันธ์มากขึ้นกับความเครียดที่สองหรือการเผชิญความเครียดและ / หรือการมีปฏิสัมพันธ์กับผู้หญิงที่เปิดรับทางเพศสัมพันธ์มากกว่ากิจกรรมทางเพศ สิ่งนี้อาจมีความเกี่ยวข้องบางอย่างในหนู RHA และ RLA ซึ่งแสดงรูปแบบการเผชิญปัญหาที่แตกต่างกันอย่างชัดเจนและการกระตุ้นการทำงานของต่อมใต้สมองต่อมหมวกไตต่อมใต้สมองที่แตกต่างกันในการตอบสนองต่อสภาวะ aversive (หนู RLA แสดงการเปิดใช้งานของแกนนี้ Carrasco et al., 2008; Díaz-Moránและคณะ, 2012) สิ่งนี้ยังเพิ่มความเป็นไปได้ที่ฮอร์โมนความเครียด (เช่น corticosteroids) อาจมีผลต่อกิจกรรม dopaminergic ใน mPFC และพื้นที่สมองอื่น ๆ เช่นกัน อันที่จริงหนู RLA นั้นเป็นตำรวจที่ทำปฏิกิริยาและแสดงพฤติกรรม hyperemotional ที่โดดเด่นด้วยความดันโลหิตต่ำและการแช่แข็งในขณะที่หนู RHA แสดงพฤติกรรมการเผชิญปัญหาเชิงรุกที่มุ่งควบคุมความเครียดDriscoll และBättig, 1982; Willig et al., 1991; Escorihuela และคณะ, 1999; Steimer และ Driscoll, 2003; Giorgi et al., 2007). แม้ว่าจะไม่สามารถแยกแยะได้อย่างสมบูรณ์ว่าสิ่งนี้อาจเกิดขึ้นในหนูที่ไร้เดียงสาทางเพศ (ซึ่งไม่เคยมีปฏิสัมพันธ์กับตัวเมียที่เปิดกว้างมาก่อน) และในหนู RLA ที่ไร้เดียงสาทางเพศโดยเฉพาะ (ดูด้านบน) ก็ไม่น่าเป็นไปได้ ในเงื่อนไขการทดลองของเราเป็นไปได้ทั้งหมดเพื่อหลีกเลี่ยงความเครียดทุกประเภท (ดูหัวข้อ "วัสดุและวิธีการ") และการทดลองจัดขึ้นในลักษณะที่ผู้ชายไม่โต้ตอบกับผู้หญิงทันที แต่หลังจากแสดงให้เธอเห็นแล้ว 30 นาที ในระหว่างที่โดพามีนเพิ่มขึ้นที่วัดได้จะต่ำกว่าที่เกิดขึ้นระหว่างการมีปฏิสัมพันธ์ทางเพศมาก หากพบการเพิ่มขึ้นของโดปามีนใน mPFC (และในนิวเคลียสแอคคัมเบน) ของหนูโรมัน (RHA ที่ไร้เดียงสาและ RLA ที่ไม่มีเพศมีระดับของโดพามีนนอกเซลล์พื้นฐานในไดอัลดีเสทจาก mPFC และนิวเคลียสแอคคัมเบนซึ่งคล้ายกันมาก) นั้นเป็นรองจริงๆ ความเครียดเนื่องจากหรือการเผชิญกับการมี / ปฏิสัมพันธ์กับเพศหญิง: (i) การเพิ่มขึ้นของโดปามีนจะพบได้สูงขึ้นมากในช่วงแรกของเวลาที่ตัวผู้ถูกวางต่อหน้าตัวเมียและไม่ช้ากว่านั้น ตามที่พบในระหว่างการมีเพศสัมพันธ์ และ (ii) การเพิ่มขึ้นของโดปามีนคาดว่าจะหายไปหรือลดลงอย่างมากหลังจากได้มาซึ่งกิจกรรมทางเพศที่มั่นคงนั่นคือเมื่อกระบวนการเรียนรู้ (รวมถึงการรับมือกับสิ่งแปลกใหม่) เสร็จสิ้นและมีบทบาทเพียงเล็กน้อยในการดำเนินการทางเพศ กิจกรรมในขณะที่ในทางตรงกันข้ามยังคงเกิดขึ้นเมื่อมีกิจกรรมทางเพศซ้ำ ๆ กล่าวอีกนัยหนึ่งโดปามีนนอกเซลล์ใน mPFC (และในนิวเคลียส accumbens) จะเพิ่มขึ้นเสมอในระหว่างการมีเพศสัมพันธ์แม้ว่าจะได้เรียนรู้พฤติกรรมทางเพศแล้วก็ตาม จำเป็นต้องมีการศึกษาเพิ่มเติมเพื่อชี้แจงบทบาทการทำงานของการเพิ่มขึ้นของกิจกรรมโดปามีนใน mPFC (และในนิวเคลียส accumbens โปรดดูหัวข้อ "บทนำและการอ้างอิง" ในนั้น)
โดยสรุปการศึกษาครั้งนี้แสดงให้เห็นเป็นครั้งแรกว่าการปรากฏตัวของและยิ่งกว่านั้นการมีปฏิสัมพันธ์กับเพศหญิงที่เปิดกว้างทางเพศนำไปสู่การเพิ่มขึ้นของ dopamine extracellular และ NA ใน dialysate จาก mPFC ของnaïveทางเพศ (ไม่เคยสัมผัสกับสิ่งเร้าทางเพศ ) และประสบการณ์ทางเพศ (ซึ่งได้รับการทดสอบการมีเพศสัมพันธ์เบื้องต้นห้าครั้งและแสดงการมีเพศสัมพันธ์ที่มั่นคง) หนู RHA และ RLA ซึ่งแสดงรูปแบบการเผชิญปัญหาที่แตกต่างกันอย่างเห็นได้ชัดเพื่อตอบสนองต่อสภาพ aversive และรูปแบบที่แตกต่างกัน ดังนั้นความเข้มข้นของโดปามีนและ NA จึงสูงกว่าใน dialysate จากหนูที่มีเพศสัมพันธ์และมีประสบการณ์ RHA เมื่อเปรียบเทียบกับ RLA ในช่วงกิจกรรมทางเพศที่คาดการณ์ไว้และสมบูรณ์ ผลลัพธ์เหล่านี้อาจเกิดจากส่วน dopaminergic และ noradrenergic tone ใน mPFC ของ RHA rats สูงกว่า rata rats ตามรายงานสำหรับ dopamine ในนิวเคลียส accumbens (Sanna และคณะ, 2015) นอกจากนี้ยังพบว่าโดปามีน mPFC และ extracellular mPFC ที่สูงกว่าใน RHA ที่มีประสบการณ์ทางเพศและหนู RLA ที่มีประสบการณ์น้อยกว่าเมื่อเปรียบเทียบกับคู่นอนที่ไม่มีการมีเพศสัมพันธ์ในสภาพฐานรวมทั้งในช่วงระยะรับประทานอาหารและบริโภค สิ่งนี้ชี้ให้เห็นว่าประสบการณ์ทางเพศทำให้เกิดกระบวนการพลาสติกที่เพิ่ม dopamine ที่มีศักยภาพและการส่งผ่านสารสื่อประสาทของ NA ใน mPFC ซึ่งพบ dopamine ในนิวเคลียส accumbens ของทั้งสองหนูโรมัน (Sanna และคณะ, 2015) ความแตกต่างข้างต้นในสารสื่อประสาท dopaminergic และ noradrenergic ใน mPFC ของทั้งสองสายโรมันอาจมีบทบาทไม่เพียง แต่ในรูปแบบที่แตกต่างกันของพฤติกรรมทางเพศ แต่ยังอยู่ในลักษณะที่แตกต่างและมักจะตรงข้ามพฤติกรรมที่เป็นลักษณะเส้นหนูทั้งสอง
ผลงานของผู้เขียน
FS, AA, MRM, OG และ MGC ออกแบบโครงการ FS และ JB ออกแบบดำเนินการและวิเคราะห์ข้อมูลจากพฤติกรรมทางเพศและการทดลอง microdialysis MAP, OG และ MGC เลือกและเลี้ยงหนูโรมัน FS, AA, MRM, OG และ MGC ดูแลการศึกษา FS, AA, MRM, OG และ MGC เขียนต้นฉบับ ผู้เขียนทั้งหมดกล่าวถึงผลลัพธ์และแสดงความคิดเห็นต้นฉบับ
การฝากและถอนเงิน
งานนี้ได้รับการสนับสนุนบางส่วนโดยทุนจากกระทรวงการต่างประเทศอิตาลีและการวิจัย (MIUR) (ความเป็นอิสระทางวิทยาศาสตร์ของนักวิจัยรุ่นใหม่, SIR 2014, หมายเลขรหัส RBSI14IUX7) ถึง FS, จากเขตปกครองตนเองของซาร์ดิเนีย (LR 7 / 2007 ,“ การส่งเสริมการวิจัยทางวิทยาศาสตร์และนวัตกรรมทางเทคโนโลยีในซาร์ดิเนีย”, กองทุนเพื่อสังคมแห่งยุโรป, 2007 – 2013, รหัสโครงการ CRP-59842) ถึง OG, และจากมหาวิทยาลัย Cagliari ถึง AA, MRM, FS, OG และ MGC
คำชี้แจงความขัดแย้งทางผลประโยชน์
ผู้เขียนประกาศว่าการวิจัยได้ดำเนินการในกรณีที่ไม่มีความสัมพันธ์ทางการค้าหรือทางการเงินใด ๆ ที่อาจตีความได้ว่าเป็นความขัดแย้งทางผลประโยชน์ที่อาจเกิดขึ้น
กิตติกรรมประกาศ
Dr. Barbara Tuveri มหาวิทยาลัย Cagliari เพื่อที่อยู่อาศัยและการดูแลสัตว์
อ้างอิง
Afonso, VM, Sison, M. , Lovic, V. และ Fleming, AS (2007) แผลเยื่อหุ้มสมอง prefrontal อยู่ตรงกลางในหนูหนูส่งผลกระทบต่อพฤติกรรมทางเพศและมารดาและองค์กรตามลำดับของพวกเขา Behav Neurosci 121, 515 – 526 ดอย: 10.1037 / 0735-7044.121.3.515
Agmo, A. และ Villalpando, A. (1995) สารกระตุ้นประสาทส่วนกลางช่วยให้พฤติกรรมทางเพศในหนูตัวผู้มีรอยโรคเยื่อหุ้มสมอง prefrontal อยู่ตรงกลาง สมอง Res 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A. , Villalpando, A. , ตัวเลือก, Z. และFernández, H. (1995) รอยโรคของเยื่อหุ้มสมอง prefrontal อยู่ตรงกลางและพฤติกรรมทางเพศในหนูผู้ชาย สมอง Res 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A. และ Melis, MR (1995) Neuromodulation ของอวัยวะเพศชายแข็งตัว: ภาพรวมของบทบาทของสารสื่อประสาทและ neuropeptides Prog Neurobiol 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A. และ Melis, MR (2005) การควบคุมส่วนกลางของการแข็งตัวของอวัยวะเพศชาย: บทบาทของนิวเคลียส paraventricular ของ hypothalamus Prog Neurobiol 76, 1 – 21 doi: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A. และ Melis, MR (2013) Neuropeptides และการควบคุมส่วนกลางของพฤติกรรมทางเพศตั้งแต่อดีตจนถึงปัจจุบัน: บทวิจารณ์ Prog Neurobiol 108, 80 – 107 doi: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L. และ Coolen, LM (2006) การมีส่วนร่วมที่เป็นไปได้ของการปล่อยออกมาจากเยื่อหุ้มสมอง prefrontal เยื่อหุ้มสมองกลางถึงการกระตุ้นประสาทตามพฤติกรรมทางเพศในหนูชาย Neuroscience 137, 1259 – 1276 doi: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L. และ Coolen, LM (2004) พฤติกรรมทางเพศและการชี้นำสิ่งแวดล้อมที่เกี่ยวข้องกับเพศเปิดใช้งานระบบ mesolimbic ในหนูตัวผู้ Neuropsychopharmacology 29, 718 – 730 doi: 10.1038 / sj.npp.1300350
Beloate, LN, Omrani, A. , Adan, RA, Webb, IC และ Coolen, LM (2016) การกระตุ้นเซลล์โดปามีนในพื้นที่ท้องน้อยในช่วงที่มีพฤติกรรมทางเพศของหนูเพศชายจะควบคุมระบบประสาทและความไวต่อยา d-amphetamine cross-sensitization J. Neurosci 36, 9949 – 9961 doi: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K. , และ Robinson, T. (1998) อะไรคือบทบาทของโดปามีนในการให้รางวัล: ผลกระทบทางความชอบ, รางวัลการเรียนรู้หรือการกระตุ้นสิ่งจูงใจ? ความต้านทานของสมอง รายได้ 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965) การเลือกอัตราที่สูงและอัตราการปรับสภาพการหลีกเลี่ยงต่ำในหนู Anim Behav 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL และ Bignami, G. (1965) ผลสหสัมพันธ์ของการคัดเลือกทางจิต: การศึกษาของสายพันธุ์โรมันสูงและต่ำหลีกเลี่ยงของหนู Behav Res Ther 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E. , Silvagni, A. , Vacca, C. , และ Di Chiara, G. (2006) ผลสะสมของ norepinephrine และ dopamine carrier blockade ต่อการเพิ่มโดปามีนนอกเซลล์ในนิวเคลียส accumbens shell, นิวเคลียสเตียงของ stria terminalis และเยื่อหุ้มสมองส่วนหน้า. J. Neurochem 96, 473 – 481 doi: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E. , Tanda, GL, Frau, R. , และ Di Chiara, G. (1990) การปิดล้อมของผู้ให้บริการ noradrenaline เพิ่มความเข้มข้นของโดปามีนนอกเซลล์ในเยื่อหุ้มสมองส่วนหน้า: หลักฐานที่แสดงว่าโดพามีนถูกนำขึ้น ในร่างกาย โดยสถานี noradrenergic J. Neurochem 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975) “ การควบคุมตัวรับโดพามีนเป็นสื่อกลาง,” ใน ตัวรับก่อนและหลัง Synaptic, eds E. Usdin และ WE Bunney (นิวยอร์ก, NY: Marcel Deker), 49 – 65
Carrasco, J. , Márquez, C. , Nadal, R. , Tobeña, A. , Fernández-Teruel, A. และ Armario, A. (2008) การจำแนกองค์ประกอบของส่วนกลางและอุปกรณ์ต่อพ่วงของแกน hypothalamus-pituitary-adrenal ในสายพันธุ์หนูหนูโรมัน Psychoneuroendocrinology 33, 437 – 445 doi: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T. และ Koolhaas, JM (2012) ความหุนหันพลันแล่นและพฤติกรรมก้าวร้าวในหนูสูงและต่ำที่โรมันหลีกเลี่ยง: ความแตกต่างพื้นฐานและความเครียดทางสังคมของวัยรุ่นที่เกิดจากการเปลี่ยนแปลง Physiol Behav 105, 1156 – 1160 doi: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D. , Piras, G. , Di Chiara, G. , และ Giorgi, O. (1997) พารามิเตอร์ทางชีวเคมีของโดปามินเนอร์จิคและสารสื่อประสาท GABAergic ในระบบประสาทส่วนกลางของการหลีกเลี่ยงโรมันสูงและหนูที่หลีกเลี่ยงโรมันต่ำ Behav จำพวก 27, 527 – 536 ดอย: 10.1023 / A: 1021452814574
Corda, MG, Piras, G. , Piludu, MA และ Giorgi, O. (2014) ผลกระทบที่แตกต่างกันของการใช้เอทานอลโดยสมัครใจต่อปริมาณโดปามีนในนิวเคลียส accumbens เปลือกของหนูสูง - และ - หลีกเลี่ยงหนูโรมัน: การศึกษาพฤติกรรมและสมอง microdialysis โลก J. Neurosci 4, 279 – 292 doi: 10.4236 / wjns.2014.43031
Dahlstrom, A. และ Fuxe, K. (1964) หลักฐานการมีอยู่ของเซลล์ประสาทที่ประกอบด้วย monoamine ในระบบประสาทส่วนกลาง I. การสาธิต monoamines ในเซลล์ของเซลล์สมองก้านสมอง Acta Physiol Scand suppl 62, 1-54
Damsma, G. , Pfaus, J. , Wenkstern, D. , Phillips, A. , และ Fibiger, H. (1992) พฤติกรรมทางเพศเพิ่มการส่งโดปามีนในนิวเคลียส accumbens และ striatum ของหนูเพศผู้: เปรียบเทียบกับความแปลกใหม่และการเคลื่อนไหว Behav Neurosci 106, 181 – 191 ดอย: 10.1037 / 0735-7044.106.1.181
D'Angio, M. , Serrano, A. , Driscoll, P. , และ Scatton, B. (1988) สิ่งเร้าทางสิ่งแวดล้อมที่ตึงเครียดนั้นเพิ่มระดับ DOPAC นอกเซลล์ในคอร์เทกซ์ prefrontal ของ hypoemotional (Roman high-Avoidance) แต่ไม่ใช่ hyperemotional (Roman low-Avoidance) หนู ในร่างกาย การศึกษาปริมาตร สมอง Res 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M. , Di Sebastiano, AR, Brown, JL, Lehman, MN, และ Coolen, LM (2010) รอยโรคของเยื่อหุ้มสมอง prefrontal อยู่ตรงกลางทำให้เกิดพฤติกรรมทางเพศที่ไม่เหมาะสมในหนูเพศผู้ Biol จิตเวช 67, 1199 – 1204 doi: 10.1016 / j.biopsych.2009.12.029
Díaz-Morán, S. , Palència, M. , Mont-Cardona, C. , Cañete, T. , Blázquez, G. , Martínez-Membrives, E. , et al. (2012) รูปแบบการเผชิญความเครียดและการตอบสนองของฮอร์โมนความเครียดในหนูที่ต่างกันทางพันธุกรรม: เปรียบเทียบกับสายพันธุ์หนูหนูโรมัน Behav สมอง Res 228, 203 – 210 ดอย: 10.1016 / j.bbr.2011.12.002
Driscoll, P. , และBättig, K. (1982) “ รูปแบบพฤติกรรมอารมณ์และ neurochemical ของหนูที่เลือกสำหรับความแตกต่างมากในการใช้งานประสิทธิภาพการหลีกเลี่ยงสองทาง” ใน พันธุศาสตร์ของสมองเอ็ด I. Lieblich (อัมสเตอร์ดัม: Elsevier), 95 – 123
Dunlop, BW และ Nemeroff, CB (2007) บทบาทของโดปามีนในพยาธิสรีรวิทยาของภาวะซึมเศร้า โค้ง. พลศาสตร์จิตเวช 64, 327 – 337 ดอย: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A. , Gil, L. , Aguilar, R. , Tobeña, A. และ Driscoll, P. (1999) หนูพันธุ์สูง - และต่ำ - โรมันที่ผสมพันธุ์ของหนู: ความแตกต่างของความวิตกกังวล, พฤติกรรมแปลกใหม่และพฤติกรรมกระสวย Physiol Behav 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990) แรงจูงใจทางเพศ: การวิเคราะห์ระบบประสาทและพฤติกรรมของกลไกการตอบสนองต่อการกระตุ้นและกระตุ้นความอยากอาหารของหนูเพศผู้ Neurosci Biobehav รายได้ 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ และ Robbins, TW (2005) ระบบประสาทของการเสริมแรงสำหรับการติดยาเสพติด: จากการกระทำไปจนถึงนิสัยการบังคับ ชัยนาท Neurosci 8, 1481 – 1489 ดอย: 10.1038 / nn1579
Fadda, F. , Argiolas, A. , Melis, MR, Tissari, AH, Onali, PL, และ Gessa, GL (1978) การเพิ่มขึ้นของความเครียดในระดับ 3,4-dihydroxyphenylacetic acid (DOPAC) ในเยื่อหุ้มสมองสมองและ n.accumbens: การกลับรายการโดย diazepam ชีวิตวิทย์ 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L. , Piras, G. , Corda, MG และ Giorgi, O. (2009) บรรทัดหนูหนูที่หลีกเลี่ยงไม่ได้สูงและต่ำแตกต่างกันในการได้มาการบำรุงรักษาการสูญพันธุ์และการคืนสถานะของการจัดการโคเคนทางหลอดเลือดดำด้วยตนเอง Neuropsychopharmacology 34, 1091 – 1101 doi: 10.1038 / npp.2008.43
Febo, M. (2011) การยิงเซลล์ Prefrontal ในหนูตัวผู้ระหว่างเข้าหาตัวเมียที่รับทางเพศสัมพันธ์: การมีปฏิสัมพันธ์กับโคเคน ไซแนปส์ 65, 271 – 277 doi: 10.1002 / syn.20843
Fernández-Guasti, A. , Omaña-Zapata, I. , Luján, M. , และCondés-Lara, M. (1994) การกระทำของการผูกเส้นประสาท sciatic ต่อพฤติกรรมทางเพศของหนูเพศผู้ที่มีประสบการณ์ทางเพศและไม่มีประสบการณ์: ผลกระทบจากการลดลงของเสาด้านหน้า Physiol Behav 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A. , Driscoll, P. , Gil, L. , Aguilar, R. , Tobeña, A. และ Escorihuela, RM (2002) ผลกระทบที่ยั่งยืนของการเสริมสร้างสิ่งแวดล้อมต่อการแสวงหาความแปลกใหม่การบริโภคขัณฑสกรและเอทานอลในสองสายหนู (RHA / Verh และ RLA / Verh) แตกต่างกันในพฤติกรรมการแสวงหาแรงจูงใจ Pharmacol Biochem Behav 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L. , Cañete, T. , Guitart-Masip, M. , Fernández-Teruel, A. และTobeña, A. (2005) ฟีโนไทป์ที่เหนี่ยวนำให้เกิด apomorphine ที่โดดเด่นสองประการในหนูสูงและต่ำที่หลีกเลี่ยงจากโรมัน .. Physiol Behav 86, 458 – 466 doi: 10.1016 / j.physbeh.2005.07.021
Giorgi, O. , Lecca, D. , Piras, G. , และ Corda, MG (2005) การกระตุ้นที่แตกต่างกันของการปลดปล่อยโดปามีนในนิวเคลียสและแกนเปลือกหุ้มหลังการฉีดแอมเฟตามีนแบบเฉียบพลันหรือแบบซ้ำ: การศึกษาเปรียบเทียบในสายหนูหนูสูงและโรมันที่หลีกเลี่ยงไม่ได้ Neuroscience 135, 987 – 998 doi: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O. , Lecca, D. , Piras, G. , Driscoll, P. , และ Corda, MG (2003a) การแยกความสัมพันธ์ระหว่างการปลดปล่อยโดปามีนจาก mesocortical และพฤติกรรมที่เกี่ยวข้องกับความกลัวในหนูที่ได้รับการคัดเลือกทางประสาทสัมผัสสองสายพันธุ์ที่แตกต่างกันในกลวิธีการเผชิญปัญหากับสภาวะที่มีชีวิต Eur J. Neurosci 17, 2716 – 2726 doi: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O. , Piras, G. , Lecca, D. , Hansson, S. , คอลล์, P. และ Corda, MG (2003b) คุณสมบัติทางประสาทวิทยาที่แตกต่างกันของการส่ง serotonergic กลางในหนูสูงและต่ำที่หลีกเลี่ยงโรมัน J. Neurochem 86, 422 – 431 doi: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O. , Orlandi, M. , Escorihuela, RM, Driscoll, P. , Lecca, D. , และ Corda, MG (1994) การส่งผ่านของ GABAergic และ dopaminergic ในสมองของหนูที่มีการหลีกเลี่ยงสูงและหนูที่มีการหลีกเลี่ยงต่ำในภาษาโรมัน สมอง Res 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O. , Piras, G. , และ Corda, MG (2007) บรรทัดหนูหนูสูงและหลีกเลี่ยงทางจิตที่ได้รับการออกแบบทางจิตใจ: แบบจำลองเพื่อศึกษาความเปราะบางต่อการติดยาเสพติดของแต่ละบุคคล Neurosci Biobehav รายได้ 31, 148 – 163 doi: 10.1016 / j.neubiorev.2006.07.008
Goto, Y. และ Grace, AA (2005) การปรับ Dopaminergic ของแรงขับของ limbic และ cortical ของนิวเคลียส accumbens ของพฤติกรรมตามเป้าหมาย ชัยนาท Neurosci 8, 805 – 812 ดอย: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ, และ Finlay, JM (1995) อิทธิพลในท้องถิ่นของ norepinephrine ภายนอกที่มีต่อโดปามีนนอกเซลล์ในเยื่อหุ้มสมองส่วนหน้า prefrontal หนู J. Neurochem 65, 111 – 116 doi: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, M. , Guevara, MA, Cervantes, M. , Morali, G. , และ Corsi-Cabrera, M. (1998) แถบความถี่ลักษณะของ EEG คอร์ติโกหน้าผากระหว่างการมีเพศสัมพันธ์ของหนูชายซึ่งเป็นผลมาจากการวิเคราะห์ปัจจัย J. สรีรวิทยา 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M. , Prieto-Beracoechea, แคลิฟอร์เนีย, Arteaga-Silva, M. , และ Guevara, MA (2007) ฟังก์ชั่นที่แตกต่างของเยื่อหุ้มสมอง prefrontal อยู่ตรงกลางและวงโคจรในระหว่างงานที่มีแรงจูงใจทางเพศในหนู Physiol Behav 90, 450 – 458 doi: 10.1016 / j.physbeh.2006.10.006
ฮอร์น, AS (1973) ความสัมพันธ์ระหว่างโครงสร้างและกิจกรรมในการยับยั้ง catecholamine ที่ดูดซึมเข้าสู่ synaptosomes จากเซลล์ประสาท noradrenergic และ dopaminergic ในเซลล์สมองของหนูหนู br J. Pharmacol 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
ฮัลล์, EM, Du, J. , Lorrain, DS และ Matuszewich, L. (1995) โดปามีน extracellular ในพื้นที่ preoptic อยู่ตรงกลาง: ความหมายสำหรับแรงจูงใจทางเพศและการควบคุมฮอร์โมนของการมีเพศสัมพันธ์ J. Neurosci 15, 7465-7471
ฮัลล์, EM, Lorrain, DS, Du, J. , Matuszewich, L. , Lumley, LA, Putnam, SK, et al. (1999) ปฏิกิริยาของฮอร์โมนและสารสื่อประสาทในการควบคุมพฤติกรรมทางเพศ Behav สมอง Res 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
ฮัลล์, EM, Weber, MS, Eaton, RC, Dua, R. , Markowski, VP, Lumley, L. , et al. (1991) ตัวรับโดปามีนในพื้นที่หน้าท้องมีผลต่อมอเตอร์ แต่ไม่ใช่ส่วนประกอบที่สร้างแรงจูงใจหรือแบบสะท้อนแสงซึ่งเป็นส่วนประกอบของการมีเพศสัมพันธ์ในหนูตัวผู้ สมอง Res 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S. และ Panksepp, J. (1999) บทบาทของนิวเคลียส accumbens โดปามีนในพฤติกรรมกระตุ้น: การตีความแบบรวมที่มีการอ้างอิงพิเศษเพื่อการแสวงหารางวัล ความต้านทานของสมอง รายได้ 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, M. , Sone, H. , Miyabara, Y. และ Tohyama, C. (2003) การได้รับปริกำเนิดจาก 2,3,7,8-tetrachlorodibenzo-p-dioxin จะเปลี่ยนแปลงการแสดงออกของ BDNF mRNA ในกิจกรรมของ neocortex และพฤติกรรมทางเพศของหนูเพศชายในวัยผู้ใหญ่ Neurotoxicology 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D. , Piras, G. , Driscoll, P. , Giorgi, O. , และ Corda, MG (2004) การกระตุ้นที่ต่างกันของโดปามีนในเปลือกและแกนกลางของนิวเคลียส accumbens นั้นสัมพันธ์กับการตอบสนองของยาต่อสิ่งเสพติด: การศึกษาการล้างไตทางสมองในหนูที่สูงและต่ำในโรมัน Neuropharmacology 46, 688 – 699 ดอย: 10.1016 / j.neuropharm.2003.11.011
Manzo, L. , Gómez, MJ, Callejas-Aguilera, JE, Donaire, R. , Sabariego, M. , Fernández-Teruel, A. , et al. (2014a) ความสัมพันธ์ระหว่างความชอบของเอทานอลกับการแสวงหาความรู้สึก Physiol Behav 133, 53 – 60 doi: 10.1016 / j.physbeh.2014.05.003
Manzo, L. , Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A. , Papini, MR และ Torres, C. (2014b) ต่อต้านความวิตกกังวลที่เกิดจากการสูญเสียสิ่งจูงใจด้วยตนเองในหนู - ยา Physiol Behav 123, 86 – 92 doi: 10.1016 / j.physbeh.2013.10.002
Masana, M. , Bortolozzi, A. และ Artigas, F. (2011) การเพิ่มประสิทธิภาพการคัดเลือกของการส่ง dopaminergic mesocortical โดยยาเสพติด noradrenergic: โอกาสในการรักษาโรคจิตเภท int J. Neuropsychopharmacol 14, 53 – 68 ดอย: 10.1017 / s1461145710000908
Meisel, RL และ Sachs, BD (1994) “ สรีรวิทยาของพฤติกรรมทางเพศชาย” ใน สรีรวิทยาของการสืบพันธุ์ปีที่ 2, 2 และ Edn., eds E. Knobil และ J. Neil (New York, NY: Raven Press), 3 – 96
Melis, MR และ Argiolas, A. (1995) โดปามีนและพฤติกรรมทางเพศ Neurosci Biobehav รายได้ 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR และ Argiolas, A. (2011) การควบคุมกลางของการแข็งตัวของอวัยวะเพศชาย: การเยี่ยมชมอีกครั้งของบทบาทของอุ้งและการมีปฏิสัมพันธ์กับโดปามีนและกรดกลูตามิกในหนูตัวผู้ Neurosci Biobehav รายได้ 35, 939 – 955 doi: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, T. , Cocco, C. , Succu, S. , Sanna, F. , Pillolla, G. , et al. (2007) ออกซิโตซินฉีดเข้าไปในพื้นที่หน้าท้องทำให้เกิดการแข็งตัวของอวัยวะเพศชายและเพิ่มโดปามีนนอกเซลล์ในนิวเคลียส accumbens และ paraventricularnucleus ของ hypothalamus ของหนูเพศผู้ Eur J. Neurosci 26, 1026 – 1035 doi: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S. , Cocco, C. , Caboni, E. , Sanna, F. , Boi, A. , et al. (2010) ออกซิโตซินก่อให้เกิดการแข็งตัวของอวัยวะเพศชายเมื่อฉีดเข้าไปในอวัยวะย่อยหน้าท้อง: บทบาทของไนตริกออกไซด์และกรดกลูตามิก Neuropharmacology 58, 1153 – 1160 ดอย: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S. , Mascia, MS, Cortis, L. และ Argiolas, A. (2003) โดปามีนเสริมเซลล์เพิ่มขึ้นในนิวเคลียส paraventricular ของหนูเพศผู้ในระหว่างกิจกรรมทางเพศ Eur J. Neurosci 17, 1266 – 1272 doi: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S. , Sanna, F. , Boi, A. , และ Argiolas, A. (2009) ออกซิโตซินฉีดเข้าไปในอวัยวะย่อยหน้าท้องหรือนิวเคลียสเยื่อหุ้มสมอง posteromedial ของ amygdala ทำให้อวัยวะเพศชายแข็งตัวและเพิ่มระดับโดปามีนนอกเซลล์ในนิวเคลียสของหนูตัวผู้ Eur J. Neurosci 30, 1349 – 1357 doi: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M. , Cardona, D. , Gómez, MJ, Sánchez-Santed, F. , Tobeña, A. , Fernández-Teruel, A. , et al. (2010) การศึกษาลักษณะหุนหันพลันแล่นในสายพันธุ์หนูหนูสูงและต่ำที่หลีกเลี่ยง: ความแตกต่างด้านพฤติกรรมและประสาทเคมี Neuropsychopharmacology 35, 1198 – 1208 doi: 10.1038 / npp.2009.224
Oliveras, I. , Río-Álamos, C. , Cañete, T. , Blázquez, G. , Martínez-Membrives, E. , Giorgi, O. , et al. (2015) การยับยั้งการ Prepulse คาดการณ์ประสิทธิภาพการทำงานของหน่วยความจำเชิงพื้นที่ในหนูโรมันสูงและต่ำหลีกเลี่ยงหนูและในหนูที่แตกต่างกันทางพันธุกรรม NIH-HS หนู: ความเกี่ยวข้องสำหรับการศึกษาล่วงหน้าผิดปกติและความรู้ความเข้าใจในโรคจิตเภท ด้านหน้า Behav Neurosci 9: 213 doi: 10.3389 / fnbeh.2015.00213
Pfaus, JG (2010) Dopamine: ช่วยให้ผู้ชายมีเพศสัมพันธ์อย่างน้อย 200 ล้านปี: ความคิดเห็นทางทฤษฎีของ Kleitz-Nelson และคณะ (2010) Behav Neurosci 124, 877 – 880 doi: 10.1037 / a0021823
Pfaus, JG, Damsma, G. , Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG, et al. (1990) พฤติกรรมทางเพศช่วยเพิ่มการส่งโดปามีนส่วนกลางในหนูตัวผู้ สมอง Res 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG และ Everitt, BJ (1995) “ เภสัชวิทยาของพฤติกรรมทางเพศ,” ใน Psychopharmacology: ความก้าวหน้ารุ่นที่สี่, eds FE Bloom และ DJ Kupfer (นิวยอร์ก, NY: Raven Press), 742 – 758
Pfaus, JG และ Phillips, AG (1991) บทบาทของโดปามีนในด้านพฤติกรรมทางเพศแบบคาดการณ์ล่วงหน้าและการบริโภคของหนูเพศผู้ Behav Neurosci 105, 727 – 747 ดอย: 10.1037 / 0735-7044.105.5.727
Piras, G. , Giorgi, O. และ Corda, MG (2010) ผลของยากล่อมประสาทที่มีต่อประสิทธิภาพในการทดสอบการว่ายน้ำแบบบังคับของหนูสองเส้นที่ผ่านการคัดเลือกทางจิตใจที่แตกต่างกันในกลวิธีการเผชิญปัญหาที่มีต่อสภาวะ aversive Psychopharmacology 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G. , Piludu, MA, Giorgi, O. และ Corda, MG (2014) ผลของการรักษาด้วยยาแก้ซึมเศร้าเรื้อรังในรูปแบบทางพันธุกรรมสมมุติของความอ่อนแอ (หนูที่หลีกเลี่ยงโรมันต่ำ) และความต้านทาน (หนูที่หลีกเลี่ยงโรมันสูง) ต่อภาวะซึมเศร้าที่เกิดจากความเครียด Psychopharmacology 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V. , Mouzon, K. , Nestler, EJ, Lehman, MN, et al. (2010) ΔFosBในนิวเคลียส accumbens เป็นสิ่งสำคัญสำหรับการเสริมผลกระทบของรางวัลทางเพศ ยีนสมอง Behav 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J. , Van, S. , Frohmader, KS, et al. (2014) neuroplasticity opioid ที่เกิดจากภายนอกของเซลล์ประสาท dopaminergic ในพื้นที่หน้าท้องมีอิทธิพลต่อธรรมชาติและรางวัลยาเสพติด J. Neurosci 34, 8825 – 8836 doi: 10.1523 / JNEUROSCI.0133-14.2014
Pitchers, KK, Vialou, V. , Nestler, EJ, Laviolette, SR, Lehman, MN, และ Coolen, LM (2013) รางวัลจากธรรมชาติและยาเสพติดทำหน้าที่เกี่ยวกับกลไกพลาสติกปั้นประสาทด้วยΔFosBเป็นสื่อกลางที่สำคัญ J. Neurosci 33, 3434 – 3442 doi: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E. , Matochik, J. , Barfield, R. , และ Auerbach, S. (1990) สหสัมพันธ์ของการปลดปล่อยโดปามีนในนิวเคลียสมีพฤติกรรมทางเพศของผู้ชายในหนู สมอง Res 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW และ Arnsten, AFT (2009) neuropsychopharmacology ของ fronto-executive function: การปรับ monoaminergic แอน รายได้ Neurosci 32, 267 – 287 doi: 10.1146 / annurev.neuro.051508.135535
Sabariego, M. , Morón, I. , Gómez, MJ, Donaire, R. , Tobeña, A. , Fernández-Teruel, A. , et al. (2013) การสูญเสียแรงจูงใจและการแสดงออกของยีนฮิปโปแคมปัสในหนูพันธุ์โรมันพันธุ์สูง (RHA-I) และหนูพันธุ์โรมันต่ำ (RLA-I) หนูหลีกเลี่ยง Behav สมอง Res 257, 62 – 70 ดอย: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000) วิธีการทางบริบททางสรีรวิทยาและการจำแนกประเภทของสมรรถภาพทางเพศ, สมรรถภาพทางเพศและความเร้าอารมณ์ทางเพศ Neurosci Biobehav รายได้ 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K. , Citro, JK, Daniels, SB, และ Natoli, JH (1994) การกระตุ้นแบบไม่สัมผัสจากตัวเมีย estrus ทำให้อวัยวะเพศชายแข็งตัวในหนู Physiol Behav 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD และ Barfield, RJ (1976) การวิเคราะห์การทำงานของพฤติกรรมการมีเพศสัมพันธ์ของผู้ชายในหนู Adv แกน Behav 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M. , Yohn, SE, López-Cruz, L. , San Miguel, N. และ Correa, M. (2016) โดปามีน Mesolimbic และการควบคุมพฤติกรรมที่กระตุ้น ฟี้ ด้านบน Behav Neurosci 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F. , Corda, MG, Melis, MR, Piludu, MA, Giorgi, O. และ Argiolas, A. (2014a) หนูตัวผู้สูงและต่ำหลีกเลี่ยงหนูแสดงพฤติกรรมที่แตกต่างกันของโคพอลิเมอร์: เปรียบเทียบกับหนูแฮมสเตอร์สปราก Physiol Behav 127, 27 – 36 doi: 10.1016 / j.physbeh.2014.01.002
Sanna, F. , Piludu, MA, Corda, MG, Argiolas, A. , Giorgi, O. และ Melis, MR (2014b) โดพามีนมีส่วนร่วมในรูปแบบที่แตกต่างกันของพฤติกรรม copulatory ของหนูสูงและต่ำหลีกเลี่ยงหนู: การศึกษากับ apomorphine และ haloperidol Pharmacol Biochem Behav 124, 211 – 219 doi: 10.1016 / j.pbb.2014.06.012
Sanna, F. , Piludu, MA, Corda, MG, Melis, MR, Giorgi, O. และ Argiolas, A. (2015) การมีส่วนร่วมของโดปามีนในความแตกต่างของพฤติกรรมทางเพศระหว่างหนูสูงและต่ำที่หลีกเลี่ยงหนู: การศึกษา microdialysis ในสมอง Behav สมอง Res 281, 177 – 186 ดอย: 10.1016 / j.bbr.2014.12.009
Scatton, B. , D'Angio, M. , Driscoll, P. , และ Serrano, A. (1988) ในร่างกาย การศึกษา voltammetric ของการตอบสนองของเซลล์ประสาท mesocortical และ mesoaccumbens dopaminergic เพื่อกระตุ้นสิ่งแวดล้อมในสายพันธุ์ของหนูที่มีระดับอารมณ์ที่แตกต่างกัน แอน NY Acad วิทย์ 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T. และ Driscoll, P. (2003) การตอบสนองความเครียดที่แตกต่างกันและรูปแบบการเผชิญปัญหาในหนูที่ได้รับการคัดเลือกในลักษณะโรมันสูง (RHA) และต่ำ (RLA): ลักษณะพฤติกรรม, neuroendocrine และพัฒนาการ ความตึงเครียด 6, 87 – 100 doi: 10.1080 / 1025389031000111320
Succu, S. , Sanna, F. , Argiolas, A. และ Melis, MR (2011) ออกซิโตซินที่ฉีดเข้าไปในอวัยวะย่อยหน้าท้องของฮิปโปแคมปัสทำให้อวัยวะเพศชายแข็งตัวขึ้นในหนูเพศผู้ Neuropharmacology 61, 181 – 188 ดอย: 10.1016 / j.neuropharm.2011.03.026
Succu, S. , Sanna, F. , Cocco, C. , Melis, T. , Boi, A. , Ferri, GL, et al. (2008) ออกซิโตซินก่อให้เกิดการแข็งตัวของอวัยวะเพศชายเมื่อฉีดเข้าไปในบริเวณหน้าท้องของหนูเพศผู้: บทบาทของไนตริกออกไซด์และวงจร GMP Eur J. Neurosci 28, 813 – 821 doi: 10.1111 / j.1460-9568.2008.06385.x
Succu, S. , Sanna, F. , Melis, T. , Boi, A. , Argiolas, A. และ Melis, MR (2007) การกระตุ้นตัวรับโดปามีนในนิวเคลียส paraventricular ของ hypothalamus ของหนูตัวผู้ทำให้อวัยวะเพศชายแข็งตัวและเพิ่มโดปามีนเสริมเซลล์ในนิวเคลียส accumbens: การมีส่วนร่วมของ oxytocin ส่วนกลาง Neuropharmacology 52, 1034 – 1043 ดอย: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G. และ Glowinski, J. (1976) การเปิดใช้งานการเลือกระบบ DA mesocortical โดยความเครียด ธรรมชาติ 263, 242 – 244 doi: 10.1038 / 263242a0
Wenkstern, D. , Pfaus, J. และ Fibiger, H. (1993) การส่งโดปามีนจะเพิ่มขึ้นในนิวเคลียสของหนูตัวผู้ในช่วงแรกที่สัมผัสกับหนูเพศเมียที่เปิดทางเพศได้ สมอง Res 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik, BHC, De Boer, P. , De Vries, JB และ Long, S. (1998) ยารักษาโรคจิตทำให้เกิดผลที่คล้ายกันในการปลดปล่อยโดปามีนและนอร์มารีนไลน์ในคอร์เทกซ์ prefrontal cortex ของสมองหนู Eur J. Pharmacol 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F. , M'Harzi, M. , Bardelay, C. , Viet, D. , และ Delacour, J. (1991) สายพันธุ์โรมันเป็นแบบจำลองทางจิตสำหรับการศึกษาความจำในการทำงาน: ข้อมูลพฤติกรรมและชีวเคมี Pharmacol Biochem Behav 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
คำสำคัญ: พฤติกรรมทางเพศ, โดปามีน, noradrenaline, mPFC, microdialysis, RHA และ RLA หนู
การอ้างอิง: Sanna F, Bratzu J, Piludu MA, Corda MG, Melis MR, Giorgi O และ Argiolas A (2017) โดปามีน, Noradrenaline และความแตกต่างในพฤติกรรมทางเพศระหว่างหนูเพศผู้สูงและต่ำที่หลีกเลี่ยงได้: การศึกษา Microdialysis . ด้านหน้า Behav Neurosci 11: 108 doi: 10.3389 / fnbeh.2017.00108
ได้รับ: 22 ธันวาคม 2016; ยอมรับแล้ว: 19 อาจ 2017;
เผยแพร่: 07 มิถุนายน 2017