 Fabrizio Sanna1*,
Fabrizio Sanna1*,  Jessica Bratzu1,
Jessica Bratzu1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melis1,
Maria R. Melis1,  Osvaldo Giorgi2 ve
Osvaldo Giorgi2 ve  Antonio Argiolas1,3
Antonio Argiolas1,3- 1Biyomedikal Bilimler Bölümü, Sinir Bilimi ve Klinik Farmakoloji Bölümü ve Bağımlılıkların Nörobiyolojisi Mükemmeliyet Merkezi, Cagliari Üniversitesi, Cagliari, İtalya
- 2Yaşam ve Çevre Bilimleri Bölümü, Farmasötik, Farmakolojik ve Beslenme Bilimleri Bölümü, Cagliari Üniversitesi, Cagliari, İtalya
- 3Sinirbilim Enstitüsü, Ulusal Araştırma Konseyi, Cagliari Bölümü, Cittadella Universitaria, Cagliari, İtalya
Mekik kutusundaki aktif kaçınma tepkisinin sırasıyla hızlı ve zayıf edinimi açısından farklılık gösteren Roma Yüksek (RHA) ve Düşük Kaçınma (RLA) aşırı üreme fareleri, cinsel bir varlığın huzuruna konulduklarında cinsel aktivitede farklılıklar sergilerler. alıcı dişi sıçan. Aslında RHA fareleri, RLA farelerine göre daha yüksek düzeyde cinsel motivasyon ve çiftleşme performansı sergiler ve bu durum tekrarlanan cinsel aktiviteden sonra da devam eder. Bu farklılıklar, RHA sıçanlarına kıyasla RLA sıçanlarının mezolimbik dopaminerjik sisteminin daha yüksek tonuyla ilişkilendirilmiştir; bu durum, cinsel aktivite sırasında RHA sıçanlarına kıyasla RHA'nın akümbens çekirdeğinden elde edilen diyalizatta bulunan dopaminin daha yüksek artışıyla ortaya çıkar. Bu çalışma, hücre dışı dopamin ve noradrenalinin (NA), erişilemeyen bir dişi sıçanın varlığında ve daha belirgin olarak doğrudan cinsel etkileşim sırasında erkek RHA ve RLA sıçanlarının medial prefrontal korteksinden (mPFC) diyalizatta da artış olduğunu göstermektedir. Dopamin (ve onun ana metaboliti 3,4-dihidroksifenilasetik asit, DOPAC) ve NA'daki bu tür artışlar hem cinsel açıdan deneyimsiz hem de deneyimli hayvanlarda bulundu, ancak bunlar daha yüksekti: (i) RHA'da RLA sıçanlarına göre; ve (ii) cinsel açıdan deneyimli RHA ve RLA sıçanlarında, saf muadillerine göre. Son olarak, mPFC'deki dopamin ve NA'daki farklılıklar, cinsel aktivitede olanlarla eşzamanlı olarak ortaya çıktı; çünkü RHA sıçanları, hem cinsel açıdan saf hem de deneyimli koşullarda RLA sıçanlarına göre daha yüksek düzeyde cinsel motivasyon ve çiftleşme performansı sergiledi. Bu sonuçlar, mezolimbik dopaminerjik sistem için önerildiği gibi, RHA ve RLA sıçanlarında bulunan farklı çiftleşme modellerinde rol oynayabilecek artan noradrenerjik tonla birlikte mPFC'de daha yüksek bir dopaminerjik tonun da meydana geldiğini göstermektedir.
Giriş
Roma Yüksek (RHA) ve Düşük Kaçınma (RLA) soyları aşan sıçan soyları, orijinal olarak mekik kutusundaki aktif kaçınma tepkisinin sırasıyla hızlı ve zayıf edinimi için seçilmiştir (Bignami, 1965; Broadhurst ve Bignami, 1965; Driscoll ve Bättig, 1982; Fernández-Teruel ve diğerleri, 2002; Giorgi ve diğerleri, 2007) cinsel açıdan alıcı bir dişi sıçanın huzuruna konulduğunda cinsel aktivitede önemli farklılıklar gösterir. Gerçekte, RHA sıçanları, RLA sıçanlarına göre daha yüksek düzeyde cinsel motivasyon sergiler; bu, erişilemeyen alıcı bir dişinin huzuruna konulduklarında bu sıçanların gösterdiği daha yüksek sayıda temassız penil ereksiyon ve RLA sıçanlarından daha iyi çiftleşme performanslarının ortaya çıkardığı gibi ortaya çıkar. esas olarak iki sıçan soyu arasındaki ilk çiftleşme testinde bulunan ve aynı zamanda zayıflatılmış olmasına rağmen beş çiftleşme testinden sonra da bulunan çok sayıda çiftleşme parametresindeki önemli farklılıklarla (Sanna ve diğerleri, 2014a). RHA ve RLA sıçanları arasında bulunan en belirgin farklar arasında, bağlanmayı ve içe girmeyi başlatan ve ilk çiftleşme testinde boşalan sıçanların yüzdesi vardır; sırasıyla RHA sıçanlarının %80'i, RLA sıçanlarının %40'ı (sırasıyla).Sanna ve diğerleri, 2014a). Çok sayıda deneysel kanıt, RHA ve RLA sıçanları arasındaki çiftleşme farklılıklarında işlevsel olarak farklı bir dopaminerjik tonun rol oynadığını göstermektedir. Aslında, RHA ve RLA sıçanlarının farklı çiftleşme modellerinin, cinsel davranışı kolaylaştıran dozlarda uygulanan karma bir D1/D2 dopamin reseptör agonisti olan apomorfin ve aynı dozda uygulanan bir D2 dopamin reseptör antagonisti olan haloperidol tarafından farklı şekilde modifiye edildiği bulunmuştur. cinsel davranışı engelleyen düşük dozlar (Sanna ve diğerleri, 2014b). Buna göre, RLA sıçanlarının, esasen farklı çiftleşme parametrelerinde daha düşük dozlarda uygulandığında iki ilacın neden olduğu daha büyük modifikasyonların gösterdiği gibi, apomorfin ve haloperidol tarafından indüklenen cinsel davranışın kolaylaştırılmasına ve inhibisyonuna karşı sırasıyla daha duyarlı olduğu bulunmuştur. RHA sıçanlarına göre RLA sıçanları (Sanna ve diğerleri, 2014b). Bu çalışma için belki de daha önemli olan, RHA ve RLA sıçanlarının farklı çiftleşme kalıplarının, cinsel davranış sırasında aktivitesinin arttığı iyi bilinen mezolimbik dopaminerjik nöronların aktivitesindeki farklılıklarla da ilişkili olmasıdır.Pfaus ve diğerleri, 1990; Pleim ve diğerleri, 1990; Pfaus ve Phillips, 1991; Damsma ve diğerleri, 1992; Wenkstern ve arkadaşları, 1993; Balfour ve arkadaşları, 2004; Pitchers ve arkadaşları, 2010, 2013; Beloate ve diğerleri, 2016). Gerçekten de, cinsel açıdan naif ve cinsel açıdan deneyimli RHA ve RLA sıçanlarının, akümbens çekirdeğinden gelen diyalizattaki hücre dışı dopaminin bazal bazal değerlerine sahip olmasına rağmen, hücre dışı dopamin ve 3,4-dihidroksifenilasetik asit (DOPAC, bunlardan biri) konsantrasyonları ana metabolitler), cinsel davranışın hem önceden hem de tüketim aşamasında, saf ve cinsel açıdan deneyimli RHA ve RLA sıçanlarının nükleus accumbens kabuğundan elde edilen diyalizatlarda farklı şekilde arttığı bulunmuştur. Yukarıdaki farklılıklar cinsel açıdan deneyimsiz RHA ve RLA sıçanları arasında daha belirgindi, ancak cinsel açıdan deneyimli RHA ve RLA sıçanları arasında devam etti, ancak cinsel davranıştaki farklılıklarda olduğu gibi bu sıçanlarda azalma eğilimindeydi (XNUMX).Sanna ve diğerleri, 2015).
Cinsel aktivite sırasında (cinsel davranışın iştah açıcı ve tüketici aşamalarında) görülen çekirdek akümbensteki dopaminerjik aktivitedeki artışların fonksiyonel rolü, cinsel aktivite gibi güçlü bir etkiye sahip olan beslenme sırasında görülenlerin yanı sıra hala tartışma konusudur. motivasyon değeri. Bu nedenle, akümbens çekirdeğindeki dopamin motivasyonla ilgili olmasına ve mezolimbik dopaminerjik nöronlara genellikle ödüllendirici nöronlar olarak atıfta bulunulmasına rağmen, son 15 yılda yapılan son çalışmalar, mezolimbik dopaminin motive edici veya ödüllendirici davranışların birincil ifadesinde yer almadığı hipotezini desteklemektedir. daha ziyade uyaran-ödül ilişkilerinin öğrenilmesi ve hatırlanmasıyla (Agmo ve diğerleri, 1995; Berridge ve Robinson, 1998; Ikemoto ve Panksepp, 1999; Pitchers ve arkadaşları, 2013, 2014; Beloate ve diğerleri, 2016; Salamone ve diğerleri, 2016). Bu hipoteze uygun olarak, çekirdek akümülatördeki dopamin reseptörlerinin blokajının veya ventral tegmental alandaki dopaminerjik nöronların etkisizleştirilmesinin, yakın zamanda erkek sıçanlarda çiftleşme davranışının iştah açıcı ve tüketici yönlerinin ifadesini değiştiremediği bulunmuştur.Pitchers ve arkadaşları, 2013, 2014; Beloate ve diğerleri, 2016).
Bununla birlikte, RHA ve RLA sıçanları arasındaki cinsel davranıştaki yukarıdaki farklılıklarda dopamin içeren diğer beyin alanlarının rolü göz ardı edilemez. Buna göre, dopaminin sadece akümbens çekirdeğinde değil, laboratuvar hayvanlarında ve insanlarda cinsel davranışın öngörücü ve tamamlayıcı aşamaları üzerinde kolaylaştırıcı etkiler gösterdiği iyi bilinmektedir (1).Everitt, 1990; Pfaus ve diğerleri, 1990; Hull ve diğerleri, 1991; Pfaus ve Everitt, 1995; Melis ve Argiolas, 2011), fakat aynı zamanda medial preoptik alan, hipotalamus ve onun çekirdekleri (yani paraventriküler çekirdek (PVN) gibi diğer beyin alanlarında da; Pfaus ve Phillips, 1991; Argiolas ve Melis, 1995, 2005, 2013; Hull ve diğerleri, 1995, 1999; Melis ve Argiolas, 1995; Melis ve diğerleri, 2003; Succu ve diğerleri, 2007; Pfaus, 2010). Dopamin içeren ve cinsel davranışta rol oynayabilecek bir diğer alan ise medial prefrontal kortekstir (mPFC; Fernández-Guasti ve diğerleri, 1994; Agmo ve Villalpando, 1995; Agmo ve diğerleri, 1995; Hernández-Gonzáles ve diğerleri, 1998, 2007; Kakeyama ve diğerleri, 2003; Balfour ve arkadaşları, 2006; Afonso ve diğerleri, 2007; Davis ve diğerleri, 2010; Şubat, 2011). Aslında, bu beyin bölgesinde mezokortikal dopamin nöronlarının sinir uçları bulunur ve hücre gövdeleri mezolimbik dopamin nöronları olarak ventral tegmental alanda lokalize olur. Çekirdek accumbens'e gelince, bu beyin bölgesinin cinsel davranıştaki kesin rolü açık olmaktan uzaktır. Aslında, mPFC lezyonlarının genellikle erkek sıçanların cinsel açıdan alıcı dişilerle cinsel davranışlarını değiştiremediği bulunmuştur.Fernández-Guasti ve diğerleri, 1994; Agmo ve Villalpando, 1995; Agmo ve diğerleri, 1995; Hernández-Gonzáles ve diğerleri, 1998, 2007; Kakeyama ve diğerleri, 2003; Balfour ve arkadaşları, 2006; Afonso ve diğerleri, 2007), ne de cinsel ödül için koşullu yer tercihinin ifadesi (Davis ve diğerleri, 2010). Bununla birlikte, cinsel ödül için koşullu yer tercihinin görünümünü değiştirmeyen mPFC lezyonları, aynı hayvanlarda, caydırıcı uyaranlarla eşleştirildiğinde cinsel aktiviteye karşı koşullu tiksinti oluşturma yeteneğini ortadan kaldırmıştır.Davis ve diğerleri, 2010) ve erkek bir sıçanın erişilemeyen, cinsel açıdan alıcı bir dişiye yaklaşma davranışları sırasındaki seçici hücre ateşlemesi, erkek sıçanların mPFC'sinde ölçülmüştür (Şubat, 2011). Bu bulgular, cinsel davranış sırasında mPFC aktivasyonunun, doğal pekiştiricilere verilen doğuştan gelen tepkilerin ifadesinden ziyade, hedefe yönelik davranışların yürütülmesi ve kontrolü için dış ve iç bilgilerin entegrasyonunda bir rol oynadığını öne sürmeye yol açtı (bkz. Goto ve Grace, 2005). Buna göre, Accumbens çekirdeğiyle birlikte mPFC, mPFC'den gelen bilişsel bilgilerin, amigdaladan gelen duygusal bilgilerin ve bağlamdan gelen duygusal bilgilerin entegrasyonunu gerektiren motive edilmiş davranışın (hedefe yönelik davranış) modülasyonunda yer alan karmaşık bir sinir sisteminin parçasıdır. İlgili bilgiler, akümbens çekirdeğindeki hipokampustan gelir (Goto ve Grace, 2005).
Deneysel kanıtlar, mPFC'deki dopamin salınımının, motive olmuş davranışın uyarlanabilir düzenlenmesinde yer aldığını ve bu mekanizmaların kuralsızlaştırılmasının, şizofreni, dikkat eksikliği ve hiperaktif bozukluklar gibi psikiyatrik bozukluklar gibi patolojik veya uyumsuz durumlarda rol oynadığının düşünüldüğünü göstermektedir. DEHB), depresyon (Dunlop ve Nemeroff, 2007; Masana ve diğerleri, 2011) veya madde bağımlılığı ve kumar davranışı (Everitt ve Robbins, 2005). İlginç bir şekilde, mPFC'de salınan dopaminin aktivitesi, mPFC'de dopaminden daha yüksek seviyelerde bulunan noradrenalinden (NA) ve özellikle sadece dopaminden daha bol miktarda bulunan NA taşıyıcısından (NET) etkilenebilir. mPFC'deki dopamin taşıyıcısı (DAT) (Carboni ve diğerleri, 1990, 2006; Gresch ve diğerleri, 1995; Westernik ve diğerleri, 1998), fakat aynı zamanda dopamin için NA'dan bile daha yüksek bir afinite gösterir (Boynuz, 1973). Dopamin ve NA çoğu zaman çalışma belleği ve dikkat seti oluşumundan tersine öğrenmeye, yanıt inhibisyonuna ve strese yanıta kadar birçok mPFC işlevinde işbirliği yaptığından (bkz. Robbins ve Arnsten, 2009), bu, mPFC'deki dopaminin tek başına veya NA ile birlikte, cinsel davranışta bulunanlar da dahil olmak üzere RHA ve RLA sıçanları arasındaki davranışsal farklılıklarda da rol oynayabileceği olasılığını artırır.
Bu hipotezi test etmek amacıyla, mezokortikal dopaminerjik sistemin ve mPFC'deki noradrenerjik sistemin aktivitesi, intraserebral mikrodiyaliz yoluyla iki RHA ve RLA sıçan soyunda incelenmiştir. Kısaca, dopamin (ve onun ana metaboliti DOPAC) ve NA, cinsel açıdan saf (örneğin, hiçbir zaman alıcı bir kadına maruz kalmamış) ve cinsel açıdan deneyimli RHA ve mPFC'nin prelimbik (PrL) ve infralimbik (IL) bölümlerinden elde edilen diyalizatlarda ölçüldü. Erişilemeyen alıcı bir dişinin huzuruna konulduklarında ve elektrokimyasal tespit (HPLC-ECD) ile birleştirilmiş yüksek basınçlı sıvı kromatografisi ile doğrudan cinsel etkileşim sırasında RLA fareleri (örneğin, beş ön çiftleşme testinden geçmiş ve sabit düzeyde çiftleşme aktivitesi gösteren).
Malzemeler ve yöntemler
Hayvanlar
Aşırı yetiştirilmiş RHA ve RLA erkek sıçanları (N = 30, deneysel çalışmanın başlangıcında ağırlığı ≈300 g olan) hepsi 1998 yılında İtalya'nın Cagliari Üniversitesi'nde kurulan kolonidendi (Giorgi ve diğerleri, 2007). Sardunya kolonisinin seçici ıslahı için kullanılan prosedürler daha önce ayrıntılı olarak anlatılmıştır (Giorgi ve diğerleri, 2005).
Tüm deneylerde kullanılan yumurtalıkları alınmış uyarıcı SD dişi sıçanlar (deneysel çalışmanın başlangıcında 250-300 g), Envigo'dan (San Pietro al Natisone, İtalya) elde edildi. Hayvanlar, deneylerin başlamasından en az 38 gün önce, 60°C'de, nemde, Cagliari Üniversitesi Biyomedikal Bilimler Bölümü'nün barındırma tesislerine kafes başına dört adet (20 cm x 10 cm x 24 cm) alıştırıldı. %60, ters 12 saatlik aydınlık/karanlık döngüsü (08:00'dan 20:00'ye kadar ışıklar kapalı), su ve standart laboratuvar gıdası ad libitum. Deneyler sırasında manipülasyon stresini sınırlamak için hayvanlara alışma süresi boyunca günlük olarak 1-2 dakika süreyle muamele edildi; Ayrıca, hayvan barınağı bakım personeli ile temas tek bir görevli ile sınırlandırılmış ve deneylerin yapıldığı gün ve öncesinde ev kafeslerindeki yataklar hiç değiştirilmemiştir. Tüm deneyler saat 10:00 ile 18:00 arasında gerçekleştirildi. Bu çalışma, Avrupa Toplulukları yönergeleri, 22 Eylül 2010 Direktifi (2010/63/EU) ve İtalyan Mevzuatı (DL 4 Mart 2014, n. 26) önerilerine uygun olarak gerçekleştirilmiştir. Protokol, Cagliari Üniversitesi Hayvan Deneyleri Etik Kurulu tarafından onaylandı (Yetki No. 361/2016-PR, 08 Nisan 2016, FS).
Deney Grupları
Cinsel açıdan saf ve cinsel açıdan deneyimli erkek RHA ve RLA sıçanları kullanıldı. Cinsel açıdan naif sıçanlar, hiçbir zaman cinsel açıdan alıcı-yumurtalıkları alınmış ve estradiol + progesteronla hazırlanmış dişiye maruz bırakılmamış sıçanlardı; cinsel açıdan deneyimli sıçanlar, alıcı bir dişiyle 60 gün aralıklarla 3 dakikalık beş ardışık çiftleşme testine tabi tutulmuş sıçanlardı (Sanna ve diğerleri, 2014a,b). Dişiler, çiftleşme testlerinden sırasıyla 200 saat ve 0.5 saat önce deri altı estradiol benzoat (yer fıstığı yağında 48 μg/sıçan) ve progesteron (yer fıstığı yağında 6 mg/sıçan) ile muamele edilerek östrusa getirildi. Östrus, May-Grunwald-Giemsa renklenmesi ve deneylerden 1 saat önce vajinal smearlerin mikroskobik incelenmesiyle doğrulandı. Önceki çalışmalarla uyumlu olarak (Sanna ve diğerleri, 2014a,b, 2015) beş ön çiftleşme testinin, her iki soydaki erkek Romalı sıçanların sabit düzeyde çiftleşme aktivitesi göstermesi için yeterli olduğu bulunmuştur: örneğin, cinsel açıdan deneyimli RHA ve RLA sıçanları, son iki testin her birinde elde edilen en az bir boşalma kriterini karşılamıştır (Bu kriteri karşılamayan bir RHA sıçanı ve iki RLA sıçanı bu aşamada atılmıştır). Bu ön çiftleşme testlerinden iki gün sonra, cinsel açıdan deneyimli Romalı sıçanlara, aşağıda açıklandığı gibi mPFC'ye bir mikrodiyaliz probunun implantasyonu için stereotaksik ameliyat uygulandı (Sanna ve diğerleri, 2015).
Cinsel Davranış Sırasında mPFC'de Mikrodiyaliz
Mikrodiyalizden bir gün önce, cinsel açıdan tecrübesiz veya deneyimli RHA ve RLA sıçanları, izofluran anestezisi (%1.5-%2; Harvard Apparatus, Holliston, MA, ABD) altında stereotaksik bir aparata (Stoelting Co., Wood Dale, IL, ABD) yerleştirildi. ve dikey bir ev yapımı mikrodiyaliz probu implante edildi (diyaliz zarı ≈3 mm serbest yüzey; Melis ve diğerleri, 2003) ve tek taraflı olarak mPFC, PrL ve IL bölmelerine yönlendirilir (koordinatlar: bregmaya 3.0 mm anterior ve 0.7 mm lateral ve duraya 5.5 mm ventral); Paxinos ve Watson, 2004). Deney günü hayvanlar, döngünün karanlık aşamasında, loş kırmızı ışıkla aydınlatılan ve içinde başka bir kafesin bulunduğu ses geçirmez bir odada bulunan çiftleşme kafesine (45 cm x 30 cm x 24 cm) transfer edildi. Görsel, kokusal ve akustik etkileşime izin vermek, ancak doğrudan etkileşime izin vermemek için dikey duvarlarda 15 delikli (Ø 15 mm) küçük Pleksiglas kafes (15 cm × 25 cm × 2 cm). 2 saatlik bir alışma süresinden sonra mikrodiyaliz probu, polietilen tüplü bir CMA/100 mikroinfüzyon pompasına (Harvard Apparatus, Holliston, MA, ABD) bağlandı ve bir Ringer çözeltisi (147 mM NaCl, 3 mM KCl ve 1.2 mM) ile perfüze edildi. CaClXNUMX2, pH 6.5), 2.5 ul/dakika akış hızında. Perfüzyon ortamının hücre dışı sıvı ile 2 saatlik bir dengelenme süresinden sonra, tarif edildiği gibi dopamin, DOPAC ve NA konsantrasyonunun ölçümü için deney sırasında her 37.5 dakikada bir 15 μL'lik diyalizat alikotları buzla soğutulmuş polietilen tüplerde toplandı. altında. En az dört diyalizat alikotunun toplanmasından sonra, çiftleşme kafesinin içindeki küçük kafese 30 dakika boyunca alıcı bir dişi sıçan yerleştirildi. Bu 30 dakika boyunca diğer iki diyalizat alikotları toplandı. Bu koşullarda erkek sıçanlar dişilerle doğrudan etkileşime giremez ancak temassız ereksiyonlar gösterirler (aşağıya bakınız). Bu sürenin ardından küçük kafes çıkarıldı, çiftleşmeye 75 dakika izin verildi ve diğer beş diyalizat alikuotu toplandı. Bu sürenin sonunda dişi çiftleşme kafesinden çıkarıldı ve ek bir diyalizat kısmı toplandı (Pfaus ve Everitt, 1995; Melis ve diğerleri, 2003; Sanna ve diğerleri, 2015). Deney boyunca cinsel davranışın öngörücü ve tamamlayıcı aşamalarıyla ilgili cinsel parametreler kaydedildi (aşağıya bakınız).
Cinsel Davranış
Cinsel davranışın öngörücü ve tamamlayıcı aşamalarıyla ilgili cinsel motivasyon ve çiftleşme performansının çeşitli parametreleri, deney boyunca, belirli deneysel koşulların farkında olmayan, örneğin cinsel deneyimin çizgisini ve düzeyini bilmeyen bir gözlemci tarafından kaydedildi. Bu deneyde kullanılan hayvanların sayısı (aşağıya bakın). Kısaca, ilk temassız ereksiyonun gecikmesi (NCPEL, alıcı dişinin iç küçük kafese girmesinden itibaren zamanlanmıştır) ve bunların sıklığı (NCPEF, dişinin bulunduğu dönemde meydana gelen temassız penis ereksiyonlarının sayısı) iç kafeste mevcut) kaydedildi. Cinsel açıdan güçlü erkek sıçanlarda, erişilemeyen alıcı bir dişinin varlığında meydana gelen bu feromon aracılı penis ereksiyonları, cinsel uyarılmanın önemli bir göstergesi olarak kabul edilir.Sachs ve diğerleri, 1994; Sachs, 2000; Melis ve diğerleri, 2003). Cinsel etkileşime izin verildiğinde, örneğin çiftleşme sırasında, binme ve içeri girme gecikmesi (ML ve IL, içteki küçük kafesin çıkarılmasından sırasıyla ilk binmeye veya ilk girişe kadar zamanlanmıştır); montaj ve giriş sıklığı (MF ve IF, ilk çiftleşme faaliyeti serisinde ve tüm çiftleşme dönemi boyunca sırasıyla montaj ve giriş sayısı); boşalma gecikmesi (EL, ilk serinin ilk girişinden boşalmaya kadar zamanlanmıştır); boşalma sıklığı (EF, çiftleşme testi sırasındaki toplam boşalma sayısı) ve boşalma sonrası aralık (PEI, ilk boşalmadan bir sonraki girişe kadar zamanlanmış) kaydedildi. Ayrıca, çiftleşme etkinliği (CE; belirli bir serideki giriş sayısının aynı serideki montaj ve giriş sayısının toplamına bölümü) ve girişler arası aralık (III; belirli bir serinin boşalma gecikmesi arasındaki oran) ve bu serideki girişlerin sayısı) aynı zamanda ilk çiftleşme faaliyeti serisi için de hesaplandı (Sachs ve Barfield, 1976; Meisel ve Sachs, 1994; Melis ve diğerleri, 2003; Sanna ve diğerleri, 2014a,b, 2015).
mPFC'den Diyalizattaki Dopamin, DOPAC ve Noradrenalin Konsantrasyonlarının Belirlenmesi
Dopamin, DOPAC ve NA konsantrasyonları, mPFC'den aynı diyalizat alikotunun 20 μL'sinde, bir 4011 çift hücre (Coulochem II, ESA, Cambridge, MA, ABD) kullanılarak elektrokimyasal tespitle birleştirilmiş yüksek basınçlı sıvı kromatografisi (HPLC) ile ölçüldü. zaten anlatılmıştı (Melis ve diğerleri, 2003). Tespit, +350 ve −180 mV'de azaltma modunda gerçekleştirildi. HPLC, %18 v/v, 7.5 metanol içeren 3.0 M sitrat/asetat pH 3 ile elüt edilen bir Supelcosil C0.06 kolonu (4.2 cm x 20 mm iç çap, 0.1 μm parçacık boyutu; Supelco, Supelchem, Milan, İtalya) ile donatıldı. Mobil faz olarak mM EDTA, 1 μM trietilamin ve 0.03 mM sodyum dodesil sülfat, 0.6 mL/dk akış hızında ve oda sıcaklığında. Testin duyarlılığı dopamin için 0.125 pg, DOPAC için 0.1 pg ve NA için 0.2 pg idi.
histoloji
Deneylerin sonunda sıçanlar başları kesilerek öldürüldü, beyinleri hemen kafatasından çıkarıldı ve 4-12 gün boyunca %15'lük sulu formaldehit içerisine daldırıldı. Bu sürenin sonunda dondurucu mikrotomla 40 µm'lik koronal beyin kesitleri hazırlandı, Neutral Red ile boyandı ve faz kontrast mikroskobunda incelendi. Daha sonra probun ucunun konumu, bir dizi beyin bölümü boyunca probun yolu takip edilerek mPFC'de lokalize edildi (bkz. 1). Sonuçların istatistiksel değerlendirmesi için yalnızca diyaliz membranının aktif kısmının mPFC'nin PrL ve IL bölmelerinde doğru şekilde konumlandırıldığı bulunan sıçanlar dikkate alındı (bu aşamada her deney grubundan bir sıçan atıldı).
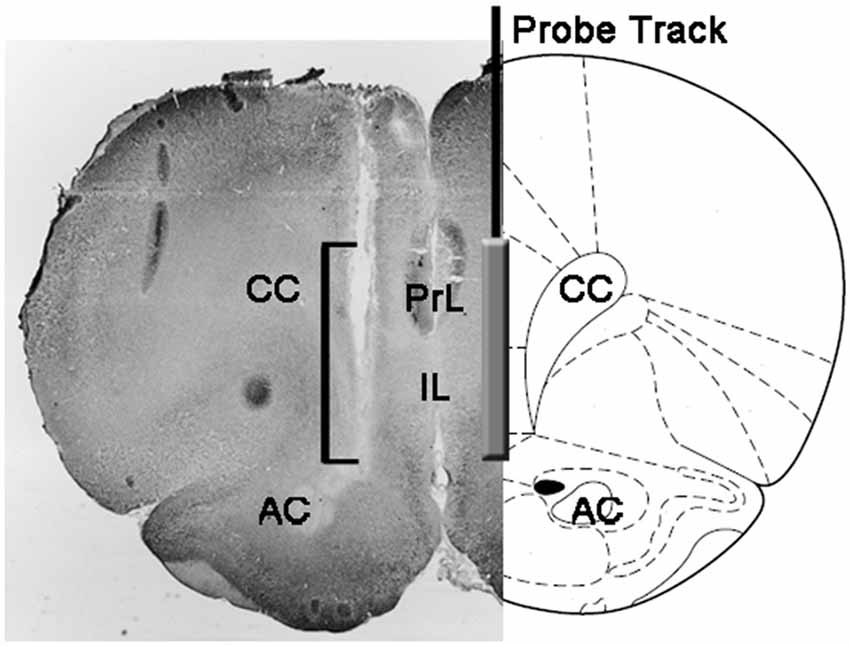 Şekil 1. Sıçan beyninin koronal bölümünün şematik gösterimi, mikrodiyaliz probunun medial prefrontal korteksin PrL ve IL kısımlarındaki izini gösterir (mPFC; Paxinos ve Watson, 2004). Mikro fotoğraftaki köşeli parantez, Nötr Kırmızı lekeli bölümün, mikrodiyaliz probunun diyaliz membranının aktif kısmını gösteren kısmını gösterir. Kısaltmalar: PrL, prelimbik alan; IL, infralimbik alan; AC, ön komissura; CC, korpus kallosum.
Şekil 1. Sıçan beyninin koronal bölümünün şematik gösterimi, mikrodiyaliz probunun medial prefrontal korteksin PrL ve IL kısımlarındaki izini gösterir (mPFC; Paxinos ve Watson, 2004). Mikro fotoğraftaki köşeli parantez, Nötr Kırmızı lekeli bölümün, mikrodiyaliz probunun diyaliz membranının aktif kısmını gösteren kısmını gösterir. Kısaltmalar: PrL, prelimbik alan; IL, infralimbik alan; AC, ön komissura; CC, korpus kallosum.
İstatistikler
Biyokimyasal (dopamin, DOPAC ve NA) ve davranışsal (NCPEL ve NCPEF, ML, IL ve EL, MF, IF ve EF ve PEI) verilerin istatistiksel analizleri, tüm deney deneklerini kapsayacak şekilde (geleneksel olarak erkek sıçan çalışmalarında yapıldığı gibi) gerçekleştirildi. çiftleşme davranışı) veya mikrodiyaliz deneyi sırasında boşalmayla çiftleşmeyen deneklerin hariç tutulması. İlk durumda, tüm hayvanlar analizlere dahil edildiğinde, dişinin erişilemez olduğu süre içinde NCPE göstermeyen veya mevcut dişiyle birlikte binmediği, içeri girmediği veya boşalmadığı hayvanlara ilgili tam aralık puanları verildi: 1800 s erkek, dişinin erişilemez olduğu 30 dakika içinde NCPE göstermedi; Eğer erkek mevcut dişiye binmemiş veya giriş yapmamışsa ML ve IL için 900 s; Erkek boşalmaya ulaşamazsa EL için 1800 saniye, erkek ilk boşalmadan sonra içeri girmezse PEI için 600 saniye. İkinci durumda, boşalmayla çiftleşmeyen hayvanlar analizlerin dışında tutuldu. Bu, cinsel davranıştaki farklılıkların, cinsel aktivitenin iki aşaması sırasında Roma sıçan soylarının mPFC'sindeki hücre dışı dopamin, DOPAC ve NA konsantrasyonlarındaki farklılıklar ile ilişkili olup olmadığını değerlendirmek için yapıldı; bu, dahil edilmeden kaynaklanan olası kafa karıştırıcı müdahaleleri ortadan kaldırır. Analiz edilen cinsel parametreler için onlara sabit değerler atayarak cinsel davranış göstermeyen deneklerin sayısı.
Kullanılan deney koşulları arasındaki olası farklılıkları tespit etmek ve daha iyi karakterize etmek için ANOVA'larla çeşitli istatistiksel analizler yapıldı. ANOVA'ları gerçekleştirmeden önce, farklı deney değişkenlerinin her birinin veri setleri, duruma bağlı olarak Bartlett veya Levine testi ile dört deney grubu arasındaki varyansların homojenliği açısından incelendi (tüm hayvanlarla veya yalnızca boşalmayla çiftleşen sıçanlarla yapıldı). Varyanslarda önemli farklılıklar bulunduğunda, veri setleri logaritmaya dönüştürüldü (yani deneysel Y değerleri, Log Y değerlerine değiştirildi), varyansların homojenliği yeniden kontrol edildi ve ardından ANOVA'larla analiz edildi. Kısaca, saf ve cinsel açıdan deneyimli erkek RHA ve RLA sıçanlarının öncesinde (örneğin, erişilemeyen dişiyle) ve ilk çiftleşme aktivitesi sırasında (ilk montajdan/girişten ilk boşalmadan sonraki ilk montaja/girişe kadar) elde edilen davranış parametreleri mikrodiyaliz sırasında sıçan çizgisi ve denekler arası faktörler arasındaki cinsel deneyim düzeyi kullanılarak iki yönlü ANOVA'larla analiz edilmiştir (davranışsal parametrelerin ortalama değerleri Tablo'da rapor edilmiştir). 1 ve F ANOVA analizlerinin değerleri ve anlamlılık düzeyleri Tabloda 2).
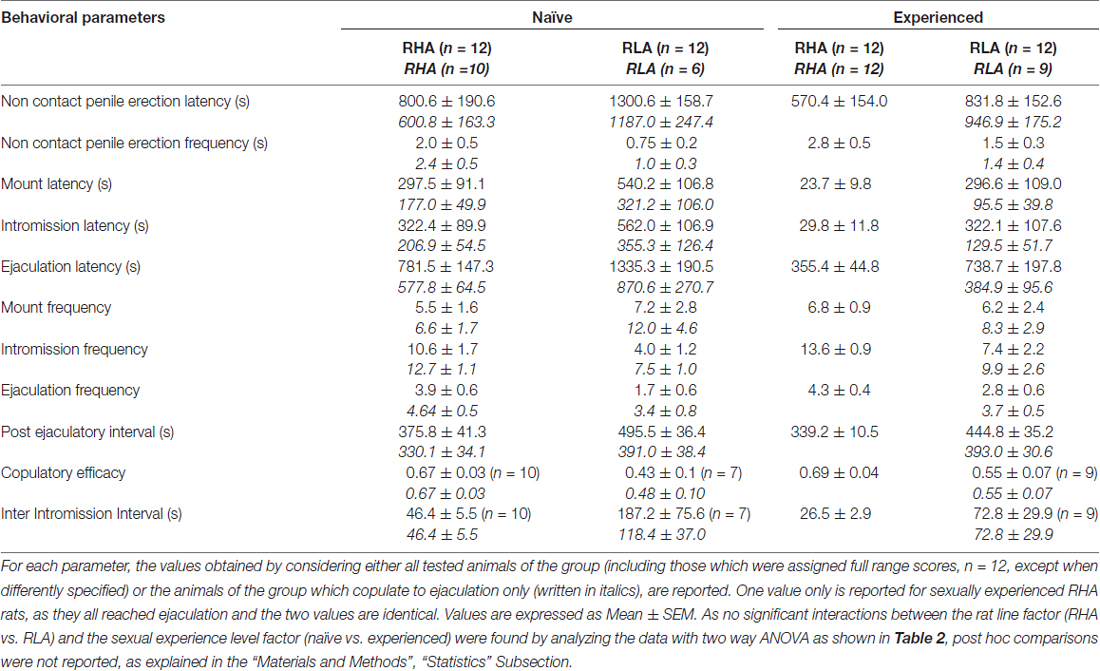 Tablo 1. Birinci çiftleşme aktivitesi serisinde ölçülen çiftleşme parametreleri (dişinin erkeğe erişilebilir hale getirildiği ilk montaj/giriş ile başlar ve ikinci serinin ilk montaj/giriş ile boşalma sonrası aralıktan sonra sona erer) ve temassız penis cinsel açıdan saf ve deneyimli RHA ve RLA sıçanlarının ereksiyonları.
Tablo 1. Birinci çiftleşme aktivitesi serisinde ölçülen çiftleşme parametreleri (dişinin erkeğe erişilebilir hale getirildiği ilk montaj/giriş ile başlar ve ikinci serinin ilk montaj/giriş ile boşalma sonrası aralıktan sonra sona erer) ve temassız penis cinsel açıdan saf ve deneyimli RHA ve RLA sıçanlarının ereksiyonları.
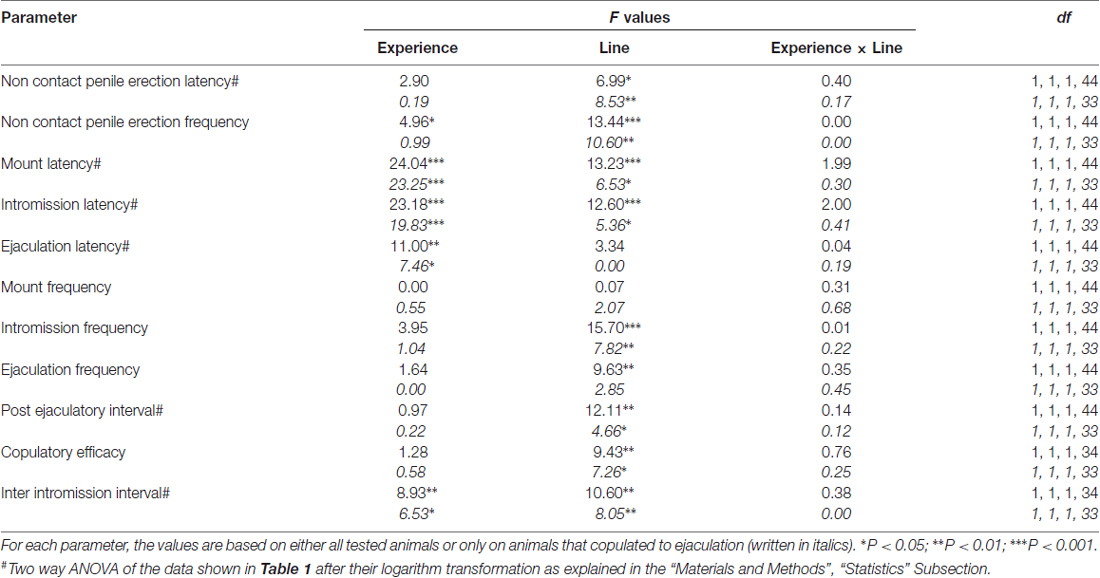 Tablo 2. F Tabloda rapor edilen veriler üzerinde gerçekleştirilen iki yönlü ANOVA'nın değerleri ve anlamlılık düzeyleri 1 denek faktörleri arasında sıçan çizgisi (RHA vs. RLA) ve cinsel deneyim düzeyi (naif vs. deneyimli) kullanılarak.
Tablo 2. F Tabloda rapor edilen veriler üzerinde gerçekleştirilen iki yönlü ANOVA'nın değerleri ve anlamlılık düzeyleri 1 denek faktörleri arasında sıçan çizgisi (RHA vs. RLA) ve cinsel deneyim düzeyi (naif vs. deneyimli) kullanılarak.
Ek olarak, mikrodiyaliz sırasında her bir sıçandan elde edilen verilerin genel bir analizi, ilk önce dopamin, DOPAC veya NA konsantrasyonlarının değerlerinin veya temassız ereksiyon, montaj, intromisyon ve ejakülasyon sayısının vs. grafiğinin çizilmesiyle elde edilen AUC'lerin hesaplanmasıyla yapıldı. . süre (erkek sıçanın çiftleşme kafesine yerleştirilmesinden 1 saat sonra başlayarak deneyin sonuna kadar -180 dakika, nörokimyasal değerler için 15 dakikalık bölümlere bölünmüş olarak- veya dişinin çiftleşme kafesine sokulduğu zamana kadar) davranışsal parametreler için) ve daha sonra hesaplanan değerlerin sıçan çizgisi ve denek faktörleri arasındaki cinsel deneyim düzeyi kullanılarak iki yönlü ANOVA'larla karşılaştırılmasıyla (nörokimyasal ve davranışsal parametrelerin AUC'lerinin ortalama değerleri Tablo'da rapor edilmiştir) 4 ve F ANOVA analizlerinin değerleri ve anlamlılık düzeyleri Tabloda 5). Son olarak, her bir veri kümesinin (yani dopamin, DOPAC, NA konsantrasyonlarının değerleri ve temassız ereksiyon, montaj, intromisyon ve boşalma sayılarına karşı zamana göre) daha ayrıntılı bir noktadan noktaya analizi faktöriyel yöntemle gerçekleştirildi. Tekrarlanan ölçümler için ANOVA'lar, sıçan çizgisi ve denek faktörleri ile zaman (yani diyalizat fraksiyonları) arasında denek faktörü (diyalizat fraksiyonları) arasındaki cinsel deneyim düzeyi kullanılarak yapılır. F Bu ANOVA analizlerinin değerleri ve anlamlılık düzeyleri Tablo'da rapor edilmiştir. 6). Ortalama değerleri Tablo 1'de rapor edilen nörokimyasal parametrelerin yalnızca bazal değerleri (alıcı dişinin çiftleşme kafesine yerleştirilmeden önce toplanan son dört diyalizat bölümü) dikkate alınarak benzer analizler yapıldı. 3. Tekrarlanan ölçümler için genel faktöriyel ANOVA'lar gerçekleştirilirken, özellikle nörokimyasal parametreler için Çizgi × Zaman ve Deneyim × Zaman etkileşimleri yapılırken birinci dereceden etkileşimler bulundu, ancak ikinci dereceden etkileşimler bulunamadı (bkz. Tablo 6), denek faktörü ve zaman arasındaki cinsel deneyim çizgisi veya düzeyi ile denek faktörü içindeki iki yönlü ANOVA'lar, cinsel açıdan saf veya deneyimli RHA ve RLA fareleri veya cinsel açıdan deneyimli ve cinsel açıdan deneyimli fareleri doğrudan karşılaştırarak bu veri kümeleri üzerinde gerçekleştirildi. saf RHA fareleri veya cinsel açıdan deneyimli ve saf RLA fareleri. Sonuçları Post hoc Bu iki yönlü ANOVA'ların ortaya çıkardığı önemli etkileşimler üzerinde Tukey HSD testi kullanılarak gerçekleştirilen ikili kontrastlar Şekilde rapor edilmektedir. 2. Diğer tüm durumlarda, Post hoc ANOVA'lar sıçan çizgisi, cinsel deneyim düzeyi ve zaman arasında herhangi bir anlamlı etkileşimi ortaya çıkaramadığından karşılaştırmalar bildirilmedi. İstatistiksel analizlerin tümü Graph Pad 5 (PRISM, San Diego, CA, ABD) ve STATISTICA 12 (Statsoft, Tulsa, OK, ABD) ile anlamlılık düzeyi şu şekilde ayarlanarak gerçekleştirildi: P <0.05.
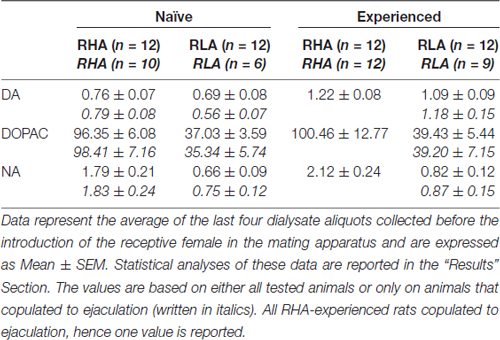 Tablo 3. Cinsel açıdan saf ve deneyimli RHA ve RLA sıçanlarının medial prefrontal korteksinden (mPFC) alınan diyalizattaki bazal dopamin (DA), 3,4-Dihidroksifenilasetik asit (DOPAC) ve noradrenalin (NA) konsantrasyonları (nM).
Tablo 3. Cinsel açıdan saf ve deneyimli RHA ve RLA sıçanlarının medial prefrontal korteksinden (mPFC) alınan diyalizattaki bazal dopamin (DA), 3,4-Dihidroksifenilasetik asit (DOPAC) ve noradrenalin (NA) konsantrasyonları (nM).
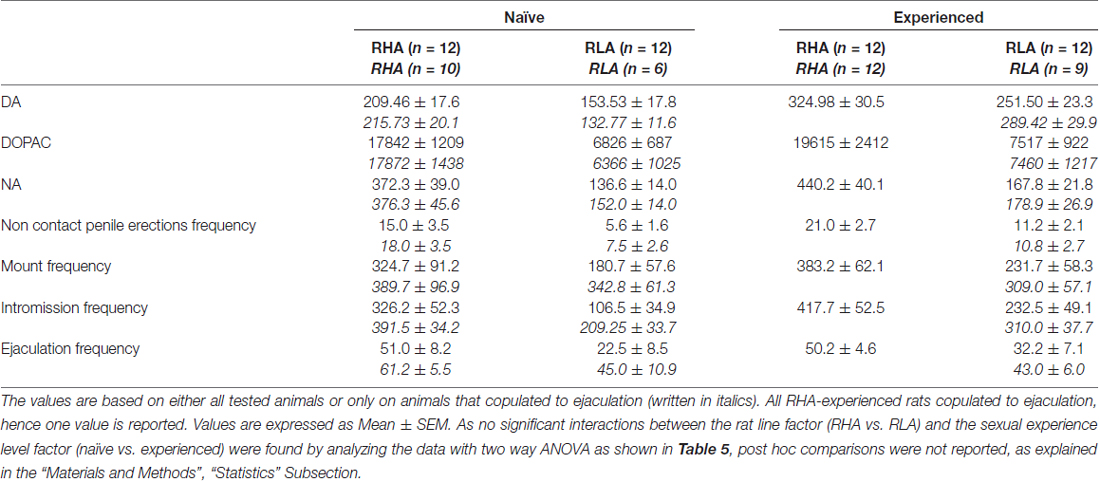 Tablo 4. Şekillerde gösterilen sonuçlardan elde edilen ortalama AUC'lerin analizi ile DA, DOPAC ve NA konsantrasyonlarındaki ve cinsel açıdan deneyimsiz ve deneyimli RHA ve RLA sıçanları arasındaki cinsel davranıştaki farklılıkların genel değerlendirmesi 2 ve 3.
Tablo 4. Şekillerde gösterilen sonuçlardan elde edilen ortalama AUC'lerin analizi ile DA, DOPAC ve NA konsantrasyonlarındaki ve cinsel açıdan deneyimsiz ve deneyimli RHA ve RLA sıçanları arasındaki cinsel davranıştaki farklılıkların genel değerlendirmesi 2 ve 3.
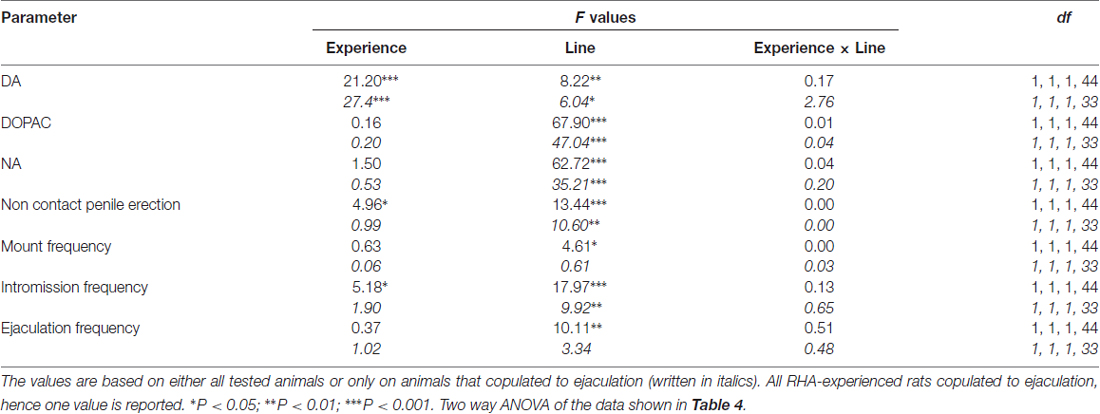 Tablo 5. F Tabloda rapor edilen veriler üzerinde gerçekleştirilen iki yönlü ANOVA'ların değerleri ve anlamlılık düzeyleri 4 denek faktörleri arasında sıçan çizgisi (RHA vs. RLA) ve cinsel deneyim düzeyi (naif vs. deneyimli) kullanılarak.
Tablo 5. F Tabloda rapor edilen veriler üzerinde gerçekleştirilen iki yönlü ANOVA'ların değerleri ve anlamlılık düzeyleri 4 denek faktörleri arasında sıçan çizgisi (RHA vs. RLA) ve cinsel deneyim düzeyi (naif vs. deneyimli) kullanılarak.
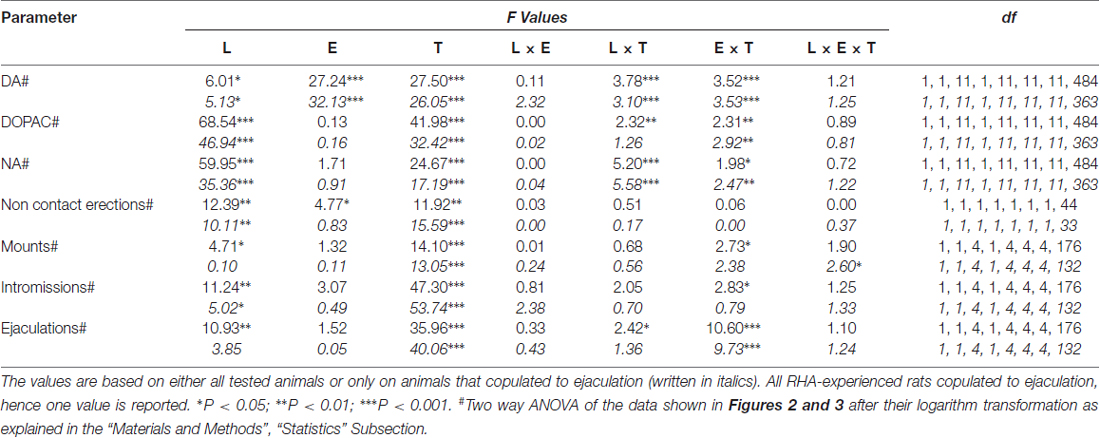 Tablo 6. F Şekillerde gösterilen sonuçlar üzerinde gerçekleştirilen tekrarlanan ölçümler için genel faktöriyel ANOVA'ların değerleri ve anlamlılık seviyeleri 2 ve 3 denek faktörleri arasında sıçan çizgisi (L; RHA vs. RLA) ve cinsel deneyim düzeyi (E) (naif vs. deneyimli) ve denek faktörü dahilinde zaman (T) (diyalizat fraksiyonları) kullanılarak.
Tablo 6. F Şekillerde gösterilen sonuçlar üzerinde gerçekleştirilen tekrarlanan ölçümler için genel faktöriyel ANOVA'ların değerleri ve anlamlılık seviyeleri 2 ve 3 denek faktörleri arasında sıçan çizgisi (L; RHA vs. RLA) ve cinsel deneyim düzeyi (E) (naif vs. deneyimli) ve denek faktörü dahilinde zaman (T) (diyalizat fraksiyonları) kullanılarak.
 Şekil 2. Cinsel açıdan saf olanlardan mPFC diyalizatlarındaki Dopamin (DA), DOPAC ve noradrenalin (NA) konsantrasyonları (A,C,E) ve deneyimli RHA ve RLA erkek sıçanları (B,D,F) alıcı bir kadınla cinsel aktivite sırasında. mPFC'ye stereotaksik olarak implante edilen bir mikrodiyaliz probu ile her iki soyun cinsel açıdan naif (yani daha önce cinsel açıdan alıcı bir dişiyle hiç karşılaşmamış) ve cinsel açıdan deneyimli (beş çiftleşme testinden geçmiş) çiftleşme kafesine ayrı ayrı yerleştirildi. Mikrodiyaliz probu, "Malzemeler ve Yöntemler" Bölümünde açıklandığı gibi diyaliz tamponu ile perfüze edildi. Bazal değerlerin belirlenmesi için dört diyalizat alikuotunun toplanmasından sonra, erişilemeyen alıcı bir dişi, çiftleştirme aparatının küçük kafesinin içine yerleştirildi (zaman = 0, erişim yok.fe). 30 dakika sonra küçük kafes çıkarıldı ve 75 dakika boyunca çiftleşmeye izin verildi (çiftleşme), ardından dişi çiftleşme kafesinden (fe.remov) çıkarıldı. Deney sırasında temassız ereksiyonlar ve çiftleşme parametreleri ölçüldü ve her 15 dakikada bir diyalizat alikotları toplandı ve "Malzemeler ve Yöntemler" Bölümünde açıklandığı gibi dopamin, DOPAC ve NA açısından analiz edildi. Değerler, grup başına 12 sıçanın tümü tarafından elde edilen değerlerin (RHA_all = düz yeşil çizgiler, RLA_all = düz kahverengi çizgiler) veya yalnızca boşalma için çiftleşenlerin (RHA_cop = kesikli kırmızı çizgiler; RLA_cop = kesikli mavi çizgiler) değerlerinin ± SEM'si anlamına gelir. Deneyimli RHA sıçanlarında tüm hayvanlar çiftleşmeye ulaştı ve değerler aynıdır. *P Grubun bazal değerlerine göre (dişi yok) < 0.05 (RHA_all için yeşil; RHA_cop için kırmızı, RLA_all için kahverengi, RLA_cop için mavi); #P RLA grubunun karşılık gelen değerlerine göre < 0.05 (kırmızı, RHA_all vs. RLA_all; yeşil, RHA_cop vs. RLA_cop); §P Cinsel açıdan naif sıçanların zaman uyumlu değerlerine göre < 0.05 (yeşil, deneyimli RHA_all vs. saf RHA_all; kırmızı, deneyimli RHA_cop vs. saf RHA_polis; kahverengi, deneyimli RLA_all vs. saf RLA_all; mavi, deneyimli RLA_cop vs. saf RLA_cop) ("Malzemeler ve Yöntemler", "İstatistikler" Alt Bölümünde açıklandığı gibi, logaritmik dönüşümlerinden sonra gösterilen veriler üzerinde yapılan iki yönlü ANOVA'lar ve ardından Tukey'nin HSD testleri).
Şekil 2. Cinsel açıdan saf olanlardan mPFC diyalizatlarındaki Dopamin (DA), DOPAC ve noradrenalin (NA) konsantrasyonları (A,C,E) ve deneyimli RHA ve RLA erkek sıçanları (B,D,F) alıcı bir kadınla cinsel aktivite sırasında. mPFC'ye stereotaksik olarak implante edilen bir mikrodiyaliz probu ile her iki soyun cinsel açıdan naif (yani daha önce cinsel açıdan alıcı bir dişiyle hiç karşılaşmamış) ve cinsel açıdan deneyimli (beş çiftleşme testinden geçmiş) çiftleşme kafesine ayrı ayrı yerleştirildi. Mikrodiyaliz probu, "Malzemeler ve Yöntemler" Bölümünde açıklandığı gibi diyaliz tamponu ile perfüze edildi. Bazal değerlerin belirlenmesi için dört diyalizat alikuotunun toplanmasından sonra, erişilemeyen alıcı bir dişi, çiftleştirme aparatının küçük kafesinin içine yerleştirildi (zaman = 0, erişim yok.fe). 30 dakika sonra küçük kafes çıkarıldı ve 75 dakika boyunca çiftleşmeye izin verildi (çiftleşme), ardından dişi çiftleşme kafesinden (fe.remov) çıkarıldı. Deney sırasında temassız ereksiyonlar ve çiftleşme parametreleri ölçüldü ve her 15 dakikada bir diyalizat alikotları toplandı ve "Malzemeler ve Yöntemler" Bölümünde açıklandığı gibi dopamin, DOPAC ve NA açısından analiz edildi. Değerler, grup başına 12 sıçanın tümü tarafından elde edilen değerlerin (RHA_all = düz yeşil çizgiler, RLA_all = düz kahverengi çizgiler) veya yalnızca boşalma için çiftleşenlerin (RHA_cop = kesikli kırmızı çizgiler; RLA_cop = kesikli mavi çizgiler) değerlerinin ± SEM'si anlamına gelir. Deneyimli RHA sıçanlarında tüm hayvanlar çiftleşmeye ulaştı ve değerler aynıdır. *P Grubun bazal değerlerine göre (dişi yok) < 0.05 (RHA_all için yeşil; RHA_cop için kırmızı, RLA_all için kahverengi, RLA_cop için mavi); #P RLA grubunun karşılık gelen değerlerine göre < 0.05 (kırmızı, RHA_all vs. RLA_all; yeşil, RHA_cop vs. RLA_cop); §P Cinsel açıdan naif sıçanların zaman uyumlu değerlerine göre < 0.05 (yeşil, deneyimli RHA_all vs. saf RHA_all; kırmızı, deneyimli RHA_cop vs. saf RHA_polis; kahverengi, deneyimli RLA_all vs. saf RLA_all; mavi, deneyimli RLA_cop vs. saf RLA_cop) ("Malzemeler ve Yöntemler", "İstatistikler" Alt Bölümünde açıklandığı gibi, logaritmik dönüşümlerinden sonra gösterilen veriler üzerinde yapılan iki yönlü ANOVA'lar ve ardından Tukey'nin HSD testleri).
ÇIKTILAR
RHA ve RLA Sıçanları Farklı Sayıda Temassız Ereksiyon ve Farklı Çiftleşme Davranışı Modelleri Gösteriyor
Daha önceki çalışmalarla uyumlu olarak (Sanna ve diğerleri, 2014a,b, 2015), farklı sayıda cinsel açıdan saf erkek RHA ve RLA sıçanları, intraserebral mikrodiyaliz yoluyla mPFC'den diyalizat alikotlarının toplanması sırasında alıcı bir dişi ile bir araya getirildiklerinde cinsel aktiviteye dahil oldu. Kısaca, bu çalışmada cinsel açıdan saf 10 erkek RHA sıçanından 12'u (%83) ilk çiftleşme testinde boşalmayla çiftleşirken, mikrodiyaliz deneyi sırasında 6 saf RLA sıçanından yalnızca 12'sı (%50) boşaldı. Bu fark aynı zamanda cinsel açıdan deneyimli erkek RHA ve RLA sıçanlarında da bulundu (beş çiftleşme testinden sonra), zayıflatılmış olmasına rağmen, mikrodiyaliz sırasında 12 RHA sıçanının tamamı 9 RLA sıçanından 12'una karşı boşalmaya ulaştı. Her zaman önceki çalışmalarla uyumlu olarak, bu çalışmada da iki Romalı sıçan çizgisi, erişilemeyen alıcı bir dişiye maruz kaldıklarında farklı gecikme süreleri ve temassız penis ereksiyonlarının yanı sıra cinsel etkileşim sırasında farklı çiftleşme davranışı modelleri sergiledi. Bu farklılıklar, belirli bir gruptaki tüm deney hayvanlarının verileri dikkate alındığında, boşalmayla çiftleşip eşleşmediklerine bakılmaksızın (örneğin, bu davranışı göstermeyen hayvanlara, yani boşalmayla çiftleşmeyen sıçanlara tam puanlar atanarak) bulunmuştur. boşalma) veya yalnızca belirli bir grubun davranışını gösteren hayvanların verileri dikkate alındığında (örneğin, boşalmayla çiftleşen hayvanlar; Tablo) 1). Buna göre, temassız ereksiyon (NCPE) değerlerinin ve çiftleşme aktivitesinin ilk serisinde iki yönlü ANOVA'larla ölçülen çiftleşme parametrelerinin istatistiksel analizleri, ya tüm deney fareleri göz önüne alındığında ya da yalnızca çiftleşen fareler göz önüne alındığında iki sıçan çizgisi arasında önemli farklılıklar ortaya çıkardı. dört deney grubunda (bkz. Tablo 2). Hatta tüm deney hayvanlarından elde edilen veriler dikkate alındığında, RHA sıçanlarında RLA sıçanlarına göre temassız ereksiyon sayısının daha yüksek olduğu ve NCPEL, ML, IL ve PEI değerlerinin anlamlı derecede kısa olduğu görüldü. Ayrıca RHA sıçanlarında RLA sıçanlarına kıyasla EF ve CE daha yüksekken III önemli ölçüde daha kısaydı. İki Roma sıçan soyu arasındaki yukarıdaki farklardan bazıları, tekrarlanan çiftleşme testleriyle azalma veya yok olma eğilimindeydi. Bununla birlikte, cinsel davranışın tekrarlanan cinsel deneyimlerle sabitlenmesinden sonra bu farklılıkların bazıları hala mevcuttu. EF hariç, yalnızca dört deney grubundaki ejakülasyonla çiftleşen hayvanların değerleri dikkate alındığında benzer sonuçlar elde edildi (bkz. Tablolar) 1, 2).
Cinsel Açıdan Tecrübesiz ve Deneyimli RHA ve RLA Sıçanlarından Alınan mPFC Diyalizatlarında Hücre Dışı Dopamin, DOPAC ve Noradrenalinin Bazal Konsantrasyonları
Mevcut deneysel koşullar altında, cinsel açıdan saf RHA ve RLA sıçanlarının tümünün (örneğin, boşalmayla çiftleşip eşleşmediklerine bakılmaksızın) mPFC'den elde edilen diyalizatlardaki dopamin, DOPAC ve NA miktarları dopamin için 2.32 pg ve 2.11 pg idi. 322.56 ul diyalizatta DOPAC için sırasıyla 124.42 pg ve 6.05 pg ve NA için sırasıyla 2.24 pg ve 20 pg. Cinsel deneyime sahip RHA ve RLA sıçanlarının mPFC'sinden elde edilen diyalizatlarda da benzer miktarlar ölçüldü (dopamin: 3.73 pg ve 3.33 pg, DOPAC: 353.62 pg ve 133.32 pg, NA: RHA ve RLA sıçanlarında sırasıyla 7.17 pg ve 2.78 pg) . Bu değerler, RHA sıçanlarının mPFC'sinde hücre dışı dopamin ve DOPAC için sırasıyla ≅0.8–1.2 nM ve ≅95–100 nM konsantrasyonunu ve hücre dışı NA için ≅1.8–2.2 nM konsantrasyonunu ve ≅0.8–1.2 nM konsantrasyonunu gösterir. RLA sıçanlarının mPFC'sinde hücre dışı dopamin ve DOPAC için sırasıyla ≅35–40 nM ve hücre dışı NA için ≅0.6–0.9 nM (Tablo) 3). Yukarıdaki değerler, diyaliz tamponunun mPFC hücre dışı sıvı ile 2 saatlik dengelenme süresinden sonra elde edildi. Orijinal dopaminin geri kazanımı, diyaliz problarının DOPAC ve NA'sının %20'ye yakın olduğu tahmin edildiğinden, hücre dışı dopamin, DOPAC ve NA konsantrasyonlarının dopamin için her iki satırda da ≅4–5 nM'ye yakın olduğu, ≅500 olduğu tahmin edilebilir. ve RHA ve RLA sıçanlarının mPFC'sinde sırasıyla DOPAC için ≅ 200 nM ve NA için ≅10 ve ≅4 nM. Logaritma dönüşümünden sonra yukarıdaki değerler üzerinde yapılan tekrarlanan ölçümler için faktöriyel ANOVA'lar, cinsel açıdan deneyimsiz ve cinsel açıdan deneyimli sıçanlar arasındaki bazal dopamin seviyelerinde önemli farklılıklar ortaya çıkardı (F(1,44,132) = 26.05, P < 0.001) ve DOPAC'ın bazal seviyelerinde (F(1,44,132) = 63.36, P < 0.001) ve NA (F(1,44,132) = 42.14, P < 0.001) RHA ve RLA sıçanları arasında (dişinin küçük kafese yerleştirilmesinden önce toplanan son dört örnek). Mikrodiyaliz deneyi sırasında yalnızca boşalmayla çiftleşen sıçanlar dikkate alındığında benzer değerler bulundu (Tablo 3). Ayrıca bu durumda, logaritma dönüşümünden sonra değerler üzerinde yapılan tekrarlanan ölçümler için faktöriyel ANOVA'lar, cinsel açıdan deneyimsiz ve cinsel açıdan deneyimli sıçanlar arasındaki bazal dopamin seviyelerinde önemli farklılıklar ortaya çıkardı (F(1,33,99) = 26.97, P < 0.001) ve DOPAC'ın bazal seviyelerinde (F(1,33,99) = 42.95, P < 0.001) ve NA (F(1,33,99) = 27.63, P < 0.001) RHA ve RLA sıçanları arasında.
Cinsel Deneyimli ve Deneyimli RHA ve RLA Sıçanlarından Elde Edilen mPFC Diyalizatlarındaki Hücre Dışı Dopamin, DOPAC ve Noradrenalin Konsantrasyonları Cinsel Aktivite Sırasında Farklı Şekilde Değişir
İçteki küçük kafeste erişilemeyen alıcı dişinin varlığı ve ardından gelen doğrudan cinsel etkileşim, hem cinsel açıdan saf hem de deneyimli erkek RHA ve RLA sıçanlarından elde edilen mPFC diyalizatlarındaki hücre dışı dopamin, DOPAC ve NA konsantrasyonlarını artırdı; ancak bunlar arasında önemli farklılıklar vardı. ya tüm deney hayvanları ya da yalnızca boşalmayla çiftleşenler dikkate alındığında iki sıçan çizgisi ve deneyim düzeyi koşulları (Şekil 2). Aslında, dopamin, DOPAC ve NA konsantrasyonlarının AUC'lerinin iki yönlü ANOVA'ları, tüm testin genel dopamin, DOPAC ve NA içeriklerinin, RLA sıçanlarına kıyasla RHA sıçanlarında ve cinsel açıdan önemli ölçüde daha yüksek olduğunu gösterdi. saf sıçanlarla karşılaştırıldığında daha deneyimlidir (bkz. Tablo 4 AUC değerleri ve Tablo için 5 için F değerler ve anlamlılık düzeyi). Bu farklılıklar, deney boyunca iki cinsel açıdan saf ve deneyimli sıçan soyunun dopamin, DOPAC ve NA miktarlarının tekrarlanan ölçümleri için noktadan noktaya faktöriyel ANOVA'larla daha da doğrulandı; bu, Çizgi, Deneyim, Zaman ve anlamlı Çizgi × Zaman'ın önemli ana etkilerini ortaya çıkardı. ve Deneyim × Zaman birinci dereceden etkileşimler (bkz. Tablo 6 için F değerler ve anlamlılık düzeyi).
Cinsel Deneyimsiz RHA ve RLA Sıçanlarından Elde Edilen mPFC Diyalizatlarındaki Hücre Dışı Dopamin, DOPAC ve Noradrenalin Konsantrasyonları Cinsel Aktivite Sırasında Farklı Şekilde Değişir
Cinsel açıdan naif RHA ve RLA sıçanlarında hücre dışı dopamin, erişilemez dişinin huzuruna konulduğunda ilk önce artarken DOPAC artmadı, NA ise yalnızca RHA sıçanlarında arttı. Saf RHA sıçanlarında hücre dışı dopamin, DOPAC ve NA ile olduğu gibi cinsel etkileşimin ilk 15 dakikasında daha da arttı. Hücre dışı dopamin, DOPAC ve NA'daki artışlar tüm çiftleşme süresi boyunca devam etti; dopamin, DOPAC ve NA sırasıyla 60 dakika, 75 dakika ve 75 dakikada zirve değerine ulaştı, ardından hücre dışı dopamin ve daha az ölçüde DOPAC ve NA , dişi çıkarıldıktan sonra bazal değerlere benzer değerlere düştü.
Öte yandan, hücre dışı dopamin, DOPAC ve NA'daki ilk anlamlı artış ve en yüksek değer, saf RLA sıçanlarında çiftleşmenin ilk 15 dakikasında meydana geldi. Bununla birlikte, RHA sıçanlarından farklı olarak, RLA sıçanlarında hücre dışı dopamin, DOPAC ve NA, dişiyle doğrudan etkileşimin ilk 30 dakikasında bazal değerlere benzer değerlere dönme eğilimi gösterdi (bkz. Şekiller) 2A,C,E).
Son olarak, çiftleşme sırasında hücre dışı dopamindeki artışlar, cinsel açıdan naif RHA'da RLA sıçanlarına kıyasla daha büyük ve daha kalıcıydı (bkz. 2A) oysa dişinin yokluğunda konsantrasyonları iki sıçan hattında aynı olmasa da benzerdi (yukarıya ve Tabloya bakın) 3). İlginçtir ki, her ne kadar iki Roma soyu arasında DOPAC artışı yüzdesi ve daha az ölçüde alıcı dişinin varlığı sırasındaki NA konsantrasyonları açısından bir fark bulunmasa da, DOPAC ve NA konsantrasyonlarının mutlak değerlerinde iki kattan fazla bir fark bulunmaktadır. iki sıçan çizgisi arasında hem bazal koşullarda gözlendi (bkz. Tablo 3) ve alıcı dişiyle test boyunca (bkz. Şekiller) 2C,E).
Cinsel Deneyimli RHA ve RLA Sıçanlarından Elde Edilen mPFC Diyalizatlarındaki Hücre Dışı Dopamin, DOPAC ve Noradrenalin Konsantrasyonları Cinsel Aktivite Sırasında Farklı Şekilde Değişir
Cinsel açıdan naif RHA ve RLA sıçanlarında bulunduğu gibi, erişilemeyen dişinin varlığı ve ardından gelen doğrudan cinsel etkileşim, cinsel açıdan deneyimli erkek RHA ve RLA sıçanlarından elde edilen mPFC diyalizatında hücre dışı dopamin ve DOPAC artışına yol açtı. Bununla birlikte, bu durumda da, ya tüm deney hayvanları göz önüne alındığında ya da daha az bir ölçüde, yalnızca boşalma ile çiftleşenler göz önüne alındığında, sıçan soyuna bağlı farklılıklar bulunmuştur (Şekil 1). 2). Cinsel açıdan deneyimli RHA ve RLA sıçanlarında, hücre dışı dopamindeki ilk artış, erişilemeyen dişi sıçanlarda meydana geldi. Daha sonra her iki sıçan soyunda da çiftleşme sırasında dopamin konsantrasyonları arttı. Dopamin konsantrasyonları çiftleşmeden 45-60 dakika sonra zirve değerlerine ulaştı ve çiftleşme testinin sonunda bazal değerlere doğru yavaş yavaş azaldı (bkz. 2B). Cinsel açıdan naif sıçanlara benzer şekilde, hücre dışı dopamin konsantrasyonları, cinsel açıdan deneyimli RHA sıçanlarında, RLA sıçanlarına kıyasla, esas olarak çiftleşme fazının orta kısmında (30-45 dakikalık çiftleşmeden sonra) toplanan diyalizat kısımlarında daha yüksekti. DOPAC konsantrasyonları, yalnızca RHA sıçanlarında erişilemeyen dişinin varlığı sırasında artarken, çiftleşme sırasında her iki sıçan soyunda da benzer ölçüde arttı (75-90 dakikada tepe değerleri) ve sürenin sonunda bazal değerlere dönme eğilimi gösterdi. çiftleşme testi (bkz. Şekil 2D). NA konsantrasyonları ayrıca, RHA sıçanlarında 90 dakikada ve RLA sıçanlarında 45 dakikada zirve değerlerine ulaşan dişinin tanıtılmasından sonra her iki cinsel deneyimi olan sıçan soylarında da arttı (bkz. Şekil 2F). Dopaminden farklı olarak (yukarıya bakınız), tüm hayvanlar ve yalnızca boşalmaya ulaşmış hayvanlar dikkate alındığında, her iki hattan cinsel açıdan deneyimli sıçanlarda DOPAC ve NA konsantrasyonlarındaki artışlarda benzer fark bulunmuştur. Bununla birlikte, cinsel açıdan deneyimsiz sıçanlarda görüldüğü gibi, DOPAC ve NA konsantrasyonlarındaki artış yüzdeleri benzer olmasına rağmen, test boyunca (yani öncesinde, sırasında ve sonrasında) cinsel açıdan deneyimli RHA ve RLA sıçanları arasında DOPAC ve NA'nın mutlak değerlerinde önemli farklılıklar bulunmuştur. alıcı dişinin varlığı), RHA sıçanları RLA sıçanlarından daha yüksek değerler gösterir (bkz. Tablo 3 bazal değerler ve Rakamlar için 2D,K testin tamamındaki değerler için).
Cinsel Deneyim, Cinsel Aktivite Sırasında RHA ve RLA Sıçanlarından Elde Edilen mPFC Diyalizatlarındaki Dopamin ve Noradrenalin Konsantrasyonlarındaki Değişiklikleri Etkiliyor
Cinsel açıdan naif ve deneyimli RHA ve RLA sıçanlarında hücre dışı dopamin ve NA konsantrasyonlarının karşılaştırılması, tüm deney hayvanları dikkate alındığında, cinsel deneyimin her iki sıçan soyunda hücre dışı dopamin konsantrasyonunu ve daha az ölçüde NA konsantrasyonunu değiştirdiğini gösterdi. veya yalnızca boşalmak için çiftleşenler (Şekiller) 2A,B,E,F). Buna göre, her iki soydaki cinsel açıdan deneyimsiz ve deneyimli sıçanlar arasında dopaminin bazal değerlerinde önemli farklılıklar meydana geldi (bkz. 3) ve çiftleşme aktivitesi sırasındaki dopamin değerleri, cinsel açıdan deneyimli RHA ve RLA sıçanlarında, saf muadillerine kıyasla genel olarak daha yüksekti (bkz. 2B). Üstelik, cinsel açıdan deneyimli RHA sıçanlarında, erişilemeyen dişinin varlığı sırasında bulunan dopamin artışı, cinsel açıdan naif sıçanlarda görülen aynı zamansal modeli izlese de, çiftleşme sırasında zamansal modelde farklılıklar bulundu. Aslında bu durumda, cinsel açıdan saf RHA sıçanlarında gözlemlenenden farklı olarak (60 dakikada bir ana tepe noktası), iki ana tepe değeri bulundu; birincisi çiftleşmeden 15 dakika sonra ve ikincisi çiftleşmeden 45 dakika sonra. Bu son artıştan sonra dopamin değerleri bazal değerlere dönme eğiliminde oldu (bkz. 2B).
Benzer bir tablo cinsel açıdan deneyimsiz ve deneyimli RLA farelerinde de bulundu. Aslında, RHA sıçanlarında bulunana benzer şekilde, deneyimli RLA sıçanlarında da hücre dışı dopamindeki üç ana artışla karakterize edilen zamansal bir model bulunmuştur: ilki erişilemeyen dişide ve diğer ikisi çiftleşme sırasında, yani 15 dakika ve 60 dakika sonra. en az çiftleşme (bkz. Şekiller) 2A, B). Genel olarak cinsel deneyim, cinsel açıdan naif durumdan deneyimli duruma geçerken her iki sıçan soyunda dopamin salınımının zamansal modelinde daha uzun süreli değişiklikler üretiyor gibi görünüyor.
Şekillerde gösterildiği gibi 2C-FHer iki hattaki saf ve deneyimli sıçanlar arasında DOPAC ve NA konsantrasyonunda yalnızca küçük farklılıklar tespit edildi. Bununla birlikte, bazalde, deneyimsiz sıçanlara kıyasla, hücre dışı NA'da daha yüksek değerlere doğru anlamlı olmayan bir eğilim gözlendi (alıcı dişinin tanıtılmasından önce, bkz. Tablo). 3) ve AUC değerlerinin ortaya çıkardığı genel NA miktarlarında (bkz. Tablo 4) tüm testten elde edilen hücre dışı konsantrasyonlar üzerinden hesaplanır (alıcı dişinin varlığı öncesinde ve sırasında). Son olarak, her iki sıçan soyundan deneyimli sıçanlar saf benzerleriyle karşılaştırıldığında NA konsantrasyonlarının zamansal düzeni çok benzerdi; bu, cinsel deneyimin, artış düzenindeki değişikliklerden ziyade hücre dışı NA'da genel bir artışa yol açabileceğini gösterir (bkz. Şekiller) 2E, K).
Cinsel Açıdan Tecrübesiz ve Deneyimli RHA ve RLA Sıçanlarından Elde Edilen mPFC Diyalizatlarındaki Hücre Dışı Dopamin ve Noradrenalin Konsantrasyonlarındaki Değişiklikler, Cinsel Davranıştaki Değişikliklerle Eşzamanlı Olarak Ortaya Çıkıyor
Test sırasında hem cinsel açıdan saf hem de deneyimli RHA ve RLA sıçanlarında bulunan mPFC'den elde edilen diyalizatlardaki hücre dışı dopamin, DOPAC ve NA konsantrasyonlarındaki farklılıklar (Şekil 2), diğerlerinin yanı sıra temassız penil ereksiyonları da içeren, deneylerin iki ana aşaması sırasında ölçülen farklı cinsel parametrelerdeki değişikliklerle birlikte meydana geldi (yani, sırasıyla kadınlara erişilemez ve erkekler için erişilebilir) (Şekiller 3A, B), bağlar (Şekiller 3C D), girişler (Şekiller 3E, K) ve boşalma (Şekiller 3G, H). Bu farklılıklar ya tüm deney hayvanları dikkate alındığında ya da yalnızca boşalmak için çiftleşen hayvanlar dikkate alındığında bulundu. Aslında, bu parametrelerin AUC değerlerinin iki yönlü ANOVA'larla bir ön analizi (bkz. 4 AUC değerleri ve Tablo için 5 için F değerler ve anlamlılık düzeyi) ve ardından tekrarlanan ölçümler için faktöriyel ANOVA ile deney boyunca bu parametrelerin değerlerinin noktadan noktaya analizi, Çizgi, Deneyim, Zaman, anlamlı birinci dereceden Çizgi × Zaman ve Deneyim × Zaman etkileşimlerinin önemli ana etkilerini ortaya çıkardı, ve montajlar için önemli bir ikinci dereceden etkileşim (bkz. Tablo 6 için F değerler ve anlamlılık düzeyi). Buna göre daha önceki çalışmalarla uyumlu olarak (Sanna ve diğerleri, 2015): (i) cinsel açıdan naif RHA sıçanları, RLA muadillerine göre daha fazla temassız penil ereksiyon, montaj, intromisyon ve boşalma gösterdi; (ii) cinsel açıdan deneyimli RHA sıçanları, RLA muadillerine göre daha yüksek sayıda temassız penil ereksiyon, giriş ve boşalma gösterdi; ve (iii) her iki soydan cinsel açıdan deneyimli sıçanlar ve özellikle RLA sıçanları, cinsel açıdan saf muadillerine göre daha fazla temassız penil ereksiyon, giriş ve boşalma gösterdi. Aslında, RHA sıçanlarında cinsel deneyimin neden olduğu değişikliklerin çoğu, davranışsal parametrelerin sıklığından ziyade zaman içerisinde meydana geldi (örneğin, deneyimli sıçanlar esas olarak testin ilk yarısında çiftleşirken, naif sıçanlar çiftleşti). Test boyunca benzer bir çiftleşme eğilimi), RLA farelerinde cinsel davranışta, özellikle de cinsel performansta daha belirgin bir genel iyileşme, deneyimli RLA farelerindeki çiftleşme parametrelerinin değerlerindeki değişikliklerle karşılaştırıldığında ortaya çıktığı gibi, cinsel deneyim tarafından üretildi. naif meslektaşları.
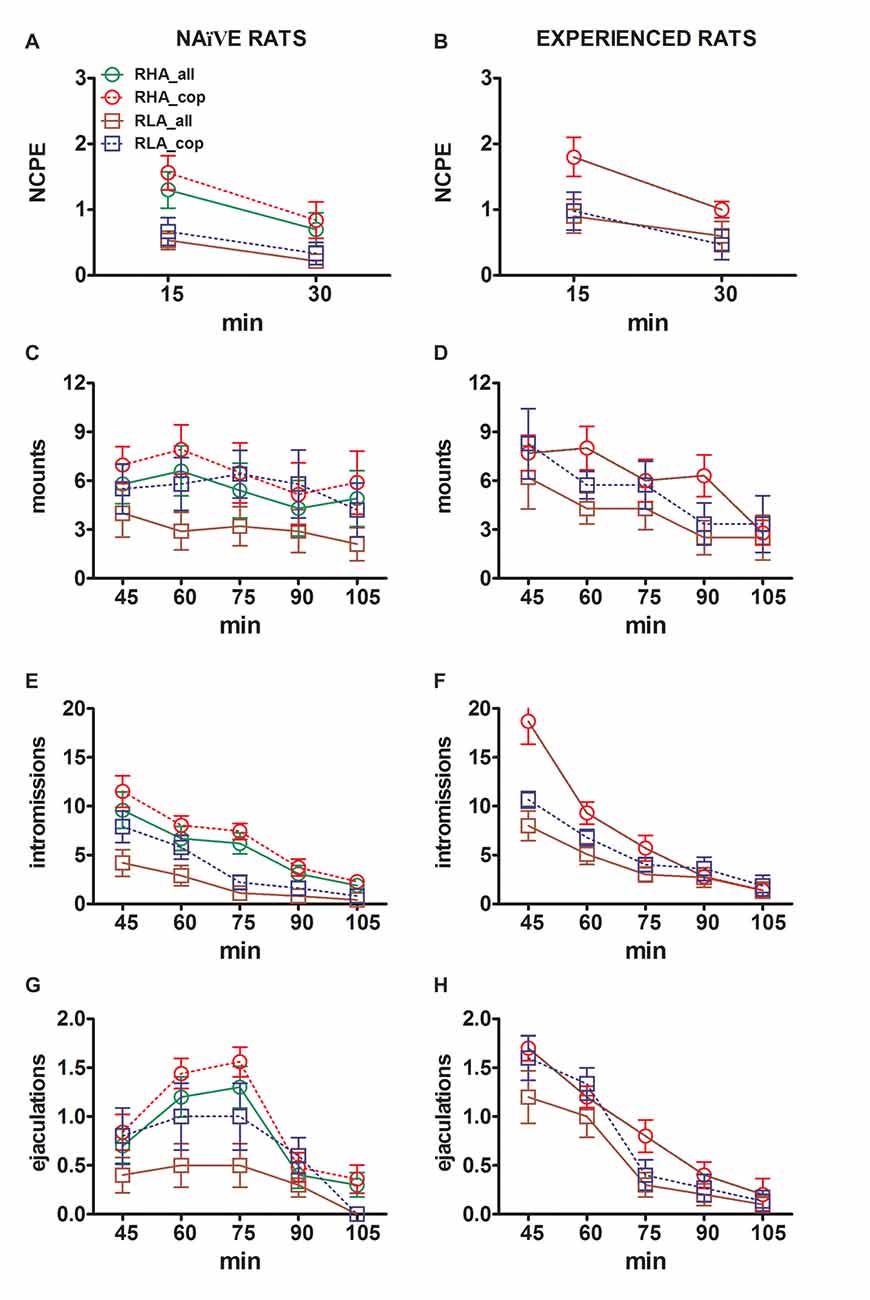 Şekil 3. Cinsel açıdan saf olanlardan kaydedilen temassız penis ereksiyonları (NCPE), montajlar, girişler ve boşalmaların sayısındaki değişiklikler (A,C,E,G) ve deneyimli RHA ve RLA fareleri (B,D,F,H) Şekilde bildirilen mikrodiyaliz deneyleri sırasında 2. Tüm deney koşulları, Şekil açıklamasında açıklananlarla aynıydı. 2. Değerler, grup başına 12 sıçanın tümü tarafından elde edilen değerlerin (tam zamanlı puanlar atananlar dahil; RHA = düz yeşil çizgiler, RLA = düz kahverengi çizgiler) veya yalnızca boşalmayla çiftleşenlerin (RHA = kesikli kırmızı çizgiler) elde edilen değerlerinin ± SEM anlamına gelir. ; RLA = kesikli mavi çizgiler). Deneyimli RHA sıçanlarında tüm hayvanlar çiftleşmeye ulaştı ve değerler aynıdır. Sıçan çizgisi faktörü (RHA'ya karşı RLA) arasında anlamlı bir etkileşim bulunmadığından, cinsel deneyim düzeyi faktörü (naif vs. deneyimli) ve zaman, Tablo'da gösterildiği gibi iki yönlü ANOVA'larla veriler analiz edilerek bulunmuştur. 6, Post hoc “Gereçler ve Yöntemler”, “İstatistikler” Alt Bölümünde açıklandığı gibi karşılaştırmalar raporlanmamıştır.
Şekil 3. Cinsel açıdan saf olanlardan kaydedilen temassız penis ereksiyonları (NCPE), montajlar, girişler ve boşalmaların sayısındaki değişiklikler (A,C,E,G) ve deneyimli RHA ve RLA fareleri (B,D,F,H) Şekilde bildirilen mikrodiyaliz deneyleri sırasında 2. Tüm deney koşulları, Şekil açıklamasında açıklananlarla aynıydı. 2. Değerler, grup başına 12 sıçanın tümü tarafından elde edilen değerlerin (tam zamanlı puanlar atananlar dahil; RHA = düz yeşil çizgiler, RLA = düz kahverengi çizgiler) veya yalnızca boşalmayla çiftleşenlerin (RHA = kesikli kırmızı çizgiler) elde edilen değerlerinin ± SEM anlamına gelir. ; RLA = kesikli mavi çizgiler). Deneyimli RHA sıçanlarında tüm hayvanlar çiftleşmeye ulaştı ve değerler aynıdır. Sıçan çizgisi faktörü (RHA'ya karşı RLA) arasında anlamlı bir etkileşim bulunmadığından, cinsel deneyim düzeyi faktörü (naif vs. deneyimli) ve zaman, Tablo'da gösterildiği gibi iki yönlü ANOVA'larla veriler analiz edilerek bulunmuştur. 6, Post hoc “Gereçler ve Yöntemler”, “İstatistikler” Alt Bölümünde açıklandığı gibi karşılaştırmalar raporlanmamıştır.
Tartışma
Bu çalışma, rahatsız edici koşullara yanıt olarak belirgin şekilde farklı başa çıkma stilleri sergileyen RHA ve RLA sıçanlarının olduğunu gösteren önceki çalışmaların bulgularını doğruluyor ve genişletiyor (Driscoll ve Bättig, 1982; Giorgi ve diğerleri, 2003a; Steimer ve Driscoll, 2003), erişilemeyen alıcı bir dişinin varlığında ve klasik çiftleşme testleri sırasında farklı davranış kalıpları gösterir (Sanna ve diğerleri, 2014a,b, 2015). Buna göre, bu çalışma aynı zamanda cinsel açıdan saf RHA sıçanlarının (daha önce hiç alıcı bir dişiyle karşılaşmamış) cinsel açıdan saf RLA sıçanlarına kıyasla daha yüksek cinsel motivasyon ve daha iyi cinsel performans sergilediğini, örneğin RHA sıçanlarının daha yüksek sayıda temassız penil ereksiyon (feromon- uyarılmış penis ereksiyonları cinsel uyarılmanın bir göstergesi olarak kabul edilir; Sachs ve diğerleri, 1994; Sachs, 2000) ve önceki çalışmalarda da görüldüğü gibi çeşitli çiftleşme parametrelerinde farklı cinsellik yanlısı değişiklikler (Sanna ve diğerleri, 2014a,b, 2015). Bu farklılıklar, cinsel deneyimin kazanılmasından sonra, yani cinsel davranışın alıcı bir dişiye beş maruz kalma ve çiftleşme testleriyle sabitlenmesinden sonra her iki Roma fare soyunda da hala devam etti. Bu çalışma aynı zamanda ilk kez hem RHA hem de RLA sıçan soylarında, naif veya deneyimli durumda, temassız penil ereksiyonların ve cinsel etkileşimin, hücre dışı dopamin (ve onun ana metaboliti DOPAC) konsantrasyonlarında bir artışla eş zamanlı olarak meydana geldiğini göstermektedir. ) ve NA, her iki Roma sıçan soyunun mPFC'sinden intraserebral mikrodiyaliz yoluyla elde edilen diyalizatlarda. İlginç bir şekilde, mPFC'deki hücre dışı dopamin ve NA'daki bu artışlar, sıçan fenotipindeki veya cinsel deneyim düzeyindeki farklılıklarla ilişkili görünmektedir. Buna göre, Sonuçlar bölümünde kapsamlı bir şekilde gösterildiği gibi, hem cinsel açıdan naif hem de deneyimli RHA sıçanları, cinsel davranışın hem iştah açıcı hem de tüketim aşamasında RLA muadilleriyle karşılaştırıldığında hücre dışı dopamin ve NA'da daha yüksek ve uzun süreli artışlar sergiledi. Üstelik, her iki Roma soyundan cinsel açıdan deneyimli sıçanlar, saf muadillerine kıyasla cinsel davranışın her iki aşamasında mPFC'de hücre dışı dopamin ve NA'da daha yüksek ve uzun süreli artışlar sergiledi. Son olarak, mPFC'deki hücre dışı dopamin ve NA'daki yukarıdaki tüm farklılıklar, açıkça cinsel davranışın davranışsal parametrelerindeki farklılıklarla ilişkiliydi; örneğin, her iki Roma soyundan cinsel açıdan deneyimli sıçanlar, saf muadilleriyle karşılaştırıldığında daha yüksek cinsel motivasyon ve daha iyi çiftleşme performansları sergiledi. ve cinsel açıdan naif ve deneyimli RHA sıçanları, cinsel açıdan naif ve deneyimli RLA muadilleriyle karşılaştırıldığında daha yüksek cinsel motivasyon ve daha iyi çiftleşme performansları sergiledi.
Cinsel aktivite sırasında mPFC diyalizatında bulunan hücre dışı dopamin artışlarına gelince, ilk artış, cinsel açıdan alıcı dişinin çiftleşme aparatına dahil edilmesinden sonraki ilk 15 dakikada, dişinin erkek için erişilemez olduğu zaman meydana geldi; bu genellikle şunu gösterir: bu durumda temassız penis ereksiyonları. Hücre dışı dopaminde ikinci ve daha büyük bir artış, dişinin çiftleşme için erkeğin erişimine açılmasından sonraki ilk 15 dakika içinde meydana geldi. İlginç bir şekilde, saf durumda hücre dışı dopamin, ikinci bölümde bazal değerlere dönme eğilimi gösteren erişilebilir dişi ile testin ilk bölümünde esas olarak artarken, cinsel deneyimin yaşandığı durumda dopamin test boyunca yüksek değerlere ulaşarak yüksek kaldı her iki Roma sıçan hattında 45-60 dakikalık çiftleşmeden sonra.
Cinsel açıdan naif ve deneyimli durumdaki RHA ve RLA sıçanları arasındaki cinsel aktivitenin farklı evreleri sırasında hücre dışı dopaminde yukarıda belirtilen farklılıkların yanı sıra, bu çalışma aynı zamanda daha önce iki Roma sıçan soyu arasında hücre dışı DOPAC konsantrasyonunda beklenmedik önemli ve son derece anlamlı farklılıklar da göstermektedir (örn. , bazal değerler) ve alıcı dişinin çiftleşme aparatına dahil edilmesinden sonra. Buna göre, mPFC diyalizatındaki hücre dışı DOPAC'ın bazal değerleri, RHA sıçanlarında, RLA sıçanlarında bulunanlardan yaklaşık 2.6 kat daha yüksek bulundu. RHA ve RLA sıçanları arasındaki bu fark, bu deney koşullarında iki Roma sıçan soyundan elde edilen mPFC diyalizatlarındaki hücre dışı dopaminin bazal değerlerindeki çok mütevazı farklılıklara rağmen, cinsel açıdan deneyimsiz ve deneyimli koşullarda ortaya çıktı ve devam etti. Birlikte ele alındığında bu farklılıklar, iki Romalı sıçan soyu arasında mPFC düzeyinde farklı bir dopamin dönüşümünü gösterebilir. Buna göre, DOPAC konsantrasyonları genellikle salınan ve daha sonra dopaminerjik sinir terminalleri tarafından yeniden yakalanan ve monoaminooksidaz (MAO;) tarafından DOPAC'a dönüştürülen dopamin miktarını yansıtır. Carlsson, 1975). Bu nedenle, RHA sıçanlarında bulunan daha yüksek bazal DOPAC konsantrasyonları, RHA sıçanlarının mPFC'sindeki dopaminerjik sinir uçları tarafından daha yüksek miktarlarda dopamin salındığını ve yeniden yakalandığını, örneğin RHA sıçanlarının mezokortikal dopaminerjik sisteminin RLA ile karşılaştırıldığında daha yüksek bir bazal aktivitesini düşündürmektedir. sıçanlar. RHA sıçanlarında RLA sıçanlarına göre daha yüksek bir mezokortikal dopaminerjik ton, hem cinsel açıdan saf hem de deneyimli RHA sıçanlarında mPFC diyalizatında cinsel aktivite sırasında RLA muadillerine göre bulunan daha yüksek hücre dışı dopamin konsantrasyonları tarafından da desteklenir. Bu bağlamda, daha yüksek bir dopaminerjik tonun, RHA ve RLA sıçanlarında mevcut olan farklı ve hatta zıt davranış özelliklerinin çoğundan en azından kısmen sorumlu olduğunun kabul edildiğini hatırlamak yerinde olacaktır. Buna göre, RHA sıçanları aktif başa çıkanlar, son derece dürtüsel, yenilik ve heyecan arayanlardır ve çeşitli bağımlılık ilaçlarını almaya ve kötüye kullanmaya eğilimlidirler; RLA sıçanları ise tepkisel başa çıkanlar, aşırı duygusaldır ve depresif benzeri semptomlar geliştirmeye eğilimlidir.Zeier ve diğerleri, 1978; Giorgi ve diğerleri, 1994, 2003b, 2007; Corda ve diğerleri, 1997, 2014; Escorihuela ve diğerleri, 1999; Steimer ve Driscoll, 2003; Lecca ve diğerleri, 2004; Giménez-Llort ve diğerleri, 2005; Carrasco ve diğerleri, 2008; Fattore ve diğerleri, 2009; Moreno ve diğerleri, 2010; Coppens ve diğerleri, 2012; Díaz-Moran ve diğerleri, 2012; Sabariego ve diğerleri, 2013; Manzo ve diğerleri, 2014a,b; Oliveras ve diğerleri, 2015), RHA sıçanlarına kıyasla cinsel açıdan saf ve deneyimli RHA sıçanlarının daha yüksek cinsel motivasyonu ve daha iyi çiftleşme performansı dahil (Sanna ve diğerleri, 2015). Cinsel açıdan naif ve deneyimli RHA sıçanlarının mPFC'sinden diyalizatta bulunan hücre dışı dopamin konsantrasyonlarındaki daha yüksek artış, RLA muadillerine göre cinsel açıdan naif ve deneyimli RHA sıçanlarının çekirdek accumbens'inden diyalizatta bulunan daha yüksek hücre dışı dopamin konsantrasyonlarına benzer. (Sanna ve diğerleri, 2015). Bununla birlikte, mPFC'den farklı olarak, hem cinsel açıdan saf hem de deneyimli RHA ve RLA sıçanlarının çekirdek akümülatörlerinden alınan diyalizattaki bazal hücre dışı dopamin ve DOPAC seviyeleri önemli ölçüde farklılık göstermedi (Sanna ve diğerleri, 2015). Bu nedenle, cinsel açıdan naif ve deneyimli RHA sıçanlarının nükleus accumbens'indeki daha yüksek mezolimbik dopaminerjik ton, alıcı farelerin varlığında RLA muadilleriyle karşılaştırıldığında yalnızca RHA sıçanlarının nükleus accumbens'inden diyalizatlarda bulunan daha yüksek hücre dışı dopamin konsantrasyonları ile çıkarılabilir. dişi ve bu çalışmada mPFC'de meydana geldiği tespit edilen sırasıyla RHA sıçanlarında ve RLA sıçanlarında DOPAC konsantrasyonunun daha yüksek ve daha düşük bazal seviyelerinin ortaya çıkardığı farklı bir bazal dopaminerjik aktivite ile değil. Bu aynı zamanda RHA sıçanlarının mPFC diyalizatındaki hücre dışı dopamin seviyelerinin RLA sıçanlarınınkinden daha yüksek bulunduğunu, buna karşılık akümbens çekirdeğinin kabuğundaki diyalizattaki benzer dopamin seviyelerinin bulunduğunu gösteren daha önceki raporların sonuçlarıyla da uyumludur. Farklı deney koşulları altında hem RHA hem de RLA sıçanlarının (D'Angio ve diğerleri, 1988; Scatton ve diğerleri, 1988; Willig ve diğerleri, 1991; Giorgi ve diğerleri, 2003a, 2007). mPFC ve akümbens çekirdeği arasındaki yukarıdaki farklılıklara rağmen, cinsel deneyim, her iki Roma sıçan soyunda da cinsel açıdan naif durumdan deneyimli duruma geçerken mPFC'de dopamin salınımının zamansal modelinde daha uzun süreli artışlara neden oluyor gibi görünüyor, ancak bu tür artışlar genellikle RHA sıçanlarında RLA sıçanlarına göre daha yüksekti. Bu, RHA sıçanlarında cinsel deneyimin hücre dışı dopamin artışlarını çiftleşme testinin ilk kısmına kaydırma eğiliminde olduğu, RLA sıçanlarında ise tüm çiftleşme testi boyunca arttırdığı çekirdek accumbens'te bulunandan biraz farklıdır.Sanna ve diğerleri, 2015).
Bildiğimiz kadarıyla, bu çalışma aynı zamanda ilk kez sadece hücre dışı dopaminin değil aynı zamanda hücre dışı NA'nın da cinsel aktivite sırasında hem RHA hem de RLA sıçanlarının mPFC'sindeki diyalizatta dopaminde olduğu gibi arttığını göstermektedir. Roma fare soyuna ve cinsel deneyime bağlı farklılıklar. Özellikle, hem RHA hem de RLA sıçanlarının mPFC'sinden elde edilen diyalizatta çok benzer bulunan bazal dopamin düzeylerinden farklı olarak, RHA sıçanlarının mPFC'sinden elde edilen diyalizattaki hücre dışı NA'nın bazal seviyeleri önemli ölçüde daha yüksekti (yaklaşık 2,5 -kat) RLA sıçanlarına göre daha fazladır. Bu farklılıkların nedenleri bilinmemektedir, ancak yukarıda dopamin ve DOPAC için tartışıldığı gibi, bu sonuçlar, RLA sıçanlarıyla karşılaştırıldığında RHA sıçanlarının en azından mPFC korteksinde daha yüksek bir bazal noradrenerjik tonun mevcut olduğunu gösterebilir. Bu kadar yüksek bir noradrenerjik ton, yukarıda dopamin için tartışıldığı gibi, RHA ve RLA sıçanlarında mevcut olan farklı ve çoğu zaman zıt davranış özelliklerine de dahil olabilir. mPFC'de RHA sıçanlarında RLA sıçanlarına göre daha yüksek bir noradrenerjik tona yönelik daha fazla destek, bazal hücre dışı NA seviyelerindeki farklılıkların hem cinsel açıdan deneyimsiz hem de deneyimli koşullarda cinsel aktivite sırasında da devam ettiğini, örneğin hücre dışı NA düzeylerinin arttığını gösteren sonuçlardan gelir. Erişilemeyen dişiye maruz bırakıldığında RHA sıçanlarında mPFC diyalizatı ve tüm çiftleşme periyodu boyunca RLA sıçanlarına göre çok daha fazla; burada NA seviyeleri çiftleşme periyodunun yalnızca ilk 15 dakikasında arttı, ardından NA seviyeleri bazal değerlere geri döndü . Hücre dışı NA seviyelerindeki bu değişiklikler hem cinsel açıdan deneyimsiz hem de deneyimli durumda görüldü, ancak NA seviyelerindeki artışlar cinsel açıdan deneyimli RHA ve RLA sıçanlarında cinsel açıdan deneyimsiz muadillerine kıyasla daha yüksekti. Dolayısıyla cinsel deneyim, dopamin için daha önce tartışıldığı gibi, RHA sıçanlarının ve daha az ölçüde RLA sıçanlarının mPFC'sindeki noradrenerjik aktivitenin daha da güçlenmesine neden olur. RHA sıçanlarının mPFC'sinde, RLA sıçanlarına kıyasla daha yüksek noradrenerjik ve dopaminerjik tonlar, bazı yorumları hak ediyor. Aslında mPFC'deki noradrenerjik aktivitenin farklı mekanizmalar yoluyla dopamin salınımını etkileyebildiği bildirildi (örneğin bkz. Carboni ve diğerleri, 1990, 2006; Gresch ve diğerleri, 1995; Westernik ve diğerleri, 1998 ve buradaki referanslar). Bunlardan biri, PFC'nin homojenatlarından elde edilen sinaptozomlarda dopamini NA'ya olandan daha yüksek bir afiniteyle bağladığı gösterilen NET'in aktivitesi ile ilgilidir (Boynuz, 1973). Eğer bu mPFC'de meydana gelseydi in vivomPFC'de salınan dopamin, yalnızca dopaminerjik tarafından değil aynı zamanda noradrenerjik sinir terminalleri tarafından NA'nınkinden bile daha yüksek bir hızda yeniden yakalanabilir. Bu da sinaptik yarıktan uzaklaştırılan dopamin miktarını telafi etmek için dopaminerjik terminallerden dopamin salınımında bir artışa (örn. dopaminerjik tonda artış) neden olabilir; dolayısıyla RHA sıçanlarında bulunan daha yüksek noradrenerjik ton, RLA sıçanlarıyla karşılaştırıldığında RHA sıçanlarının mPFC'sinde bulunan daha yüksek dopaminerjik tona en azından kısmen katkıda bulunabilir. Bu olasılığı doğrulamak için daha ileri çalışmalara ihtiyaç vardır. Özellikle, bu çalışmada RHA ve RLA sıçanları arasındaki mPFC'de tanımlanan noradrenerjik aktivite farkı, antidepresan tedavisine farklı yanıt vermelerinde de rol oynayabilir. Aslında, RHA fareleri bir dayanıklılık modeli olarak kabul edilirken, RLA fareleri de depresyona karşı bir kırılganlık modeli olarak kabul ediliyor. Buna göre, desipramin, fluoksetin ve klorimipramin gibi antidepresanlarla yapılan akut veya kronik tedavi, RHA sıçanlarının tepkilerini etkilemeden, RLA sıçanlarında zorunlu yüzme testindeki davranışsal tepkileri önemli ölçüde iyileştirdi.Piras ve diğerleri, 2010, 2014).
Son olarak bu çalışma, mPFC'nin cinsel davranışın kontrolünde rol oynadığını gösteren önceki bulguları doğruluyor ve genişletiyor, ancak bu beyin bölgesinin cinsel davranıştaki kesin rolü açık olmaktan uzaktır (Fernández-Guasti ve diğerleri, 1994; Agmo ve Villalpando, 1995; Agmo ve diğerleri, 1995; Hernández-Gonzáles ve diğerleri, 1998, 2007; Kakeyama ve diğerleri, 2003; Balfour ve arkadaşları, 2006; Afonso ve diğerleri, 2007; Davis ve diğerleri, 2010; Şubat, 2011). Son zamanlarda, erkek bir sıçanın, cinsel açıdan kabul edilemez bir dişiye yaklaşma davranışları sırasında mPFC'de seçici hücre ateşlemesi, tek hücreli ateşleme kayıt paradigması kullanılarak gösterilmiştir.Şubat, 2011). İlginç bir şekilde, erkeğin erişilemez alıcı dişiye ilk maruz kalması sırasında tepki vermeyen nöronlar, sanki önceki deneyim davranışsal tepkilerde varyasyona yol açan plastik değişiklikleri tetikleyebilmiş gibi, ikinci maruz kalma sırasında aktif hale gelir. Bununla birlikte, mPFC'nin ibotenik asidine sahip lezyonların, erkek sıçanların cinsel açıdan alıcı bir dişiyle cinsel davranışlarını veya cinsel ödül için koşullu yer tercihinin ifadesini değiştiremediği bulundu. Bununla birlikte, bu lezyonlu hayvanlarda, caydırıcı uyaranlarla eşleştirildiğinde cinsel aktiviteye karşı koşullu tiksinti oluşturma yeteneğinin tamamen ortadan kalktığı bulunmuştur.Davis ve diğerleri, 2010). Bu, cinsel davranış sırasında mPFC aktivasyonunun, doğal pekiştiricilere verilen doğuştan gelen tepkilerin ifadesinden ziyade, hedefe yönelik davranışların yürütülmesi ve kontrolü için dış ve iç bilgilerin entegrasyonunda bir rol oynadığını göstermektedir (bkz. Goto ve Grace, 2005). Bu çalışma için daha da önemlisi, çeşitli çalışmalar, çekirdek accumbens, ventral tegmental alan, medial preoptik alan, stria terminalisin yatak çekirdeği, bazolateral amigdala ve parvoselüler subparafasiküler talamik çekirdek gibi cinsel motivasyon ve cinsel davranışla ilgili beyin alanlarının girdi aldığını göstermiştir. cinsel aktivite sırasında mPFC'den (bkz. Balfour ve arkadaşları, 2006). Bu beyin alanlarının çoğu, cinsel motivasyon ve ödülden cinsel performansa kadar cinsel davranışın kontrolünde yer alan karmaşık bir sinir devresine katılır. Bu devre, hipotalamusun PVN'sinden (PVN) kaynaklanan ve ventral tegmental alana, nukleus accumbens'e, hipokampusa, amigdalaya, PFC'ye, stria terminalisin yatak çekirdeğine, medulla oblongata'ya ve omuriliğe, mezolimbik/mezokortikal dopaminerjike projekte olan oksitoserjik nöronları içerir. Ventral tegmental alandan nükleus accumbens'e ve PFC'ye uzanan nöronlar, devrenin çeşitli alanlarında lokal ve sistem düzeyinde katılan insertohipotalamik dopaminerjik nöronlar ve glutamaterjik nöronlar (bkz. Melis ve Argiolas, 1995, 2011; Melis ve diğerleri, 2003, 2007, 2009, 2010; Succu ve diğerleri, 2007, 2008, 2011). Bu nedenle, cinsel aktivite sırasında görülen mPFC'den diyalizatlarda hücre dışı dopaminin (aynı zamanda NA'nın da) artması, mPFC'nin bu devrenin aktif bir parçası olduğunu, yani mezokortikal dopaminerjik nöronların cinsel aktivitenin kontrolüne birlikte katıldığını gösterebilir. noradrenerjik, mezolimbik dopaminerjik, oksitosinerjik ve glutamaterjik nöronlarla ilişkilidir, ancak cinsel davranışın ifadesi için kesinlikle gerekli görünmemektedir (Davis ve diğerleri, 2010). Bu hipoteze uygun olarak bu çalışma, cinsel aktivite sırasında mPFC'de dopamin ve NA salınımındaki farklılıkların, temassız penis ereksiyonlarının sayısındaki farklılıklar ve çiftleşme parametrelerindeki değişikliklerle eş zamanlı olarak meydana geldiğini ve cinsel aktivite sırasında daha yüksek dopamin ve NA salınımıyla birlikte ortaya çıktığını göstermektedir. hem Romalı sıçan soylarında hem de hem cinsel açıdan saf hem de deneyimli koşullarda daha yüksek cinsel motivasyon ve daha iyi çiftleşme performanslarıyla ilişkili iştah açıcı ve tüketici aşamalar.
Yukarıda verilen tartışma esas olarak, cinsel aktivite sırasında erkek sıçanların akümbens çekirdeğinde meydana gelen hücre dışı dopaminin iyi bilinen artışına dayanmaktadır (Pfaus ve diğerleri, 1990; Pleim ve diğerleri, 1990; Pfaus ve Phillips, 1991; Damsma ve diğerleri, 1992; Wenkstern ve arkadaşları, 1993). Bununla birlikte, stres veya stresle başa çıkma gibi diğer deneysel koşullarda mPFC'de (ve nukleus accumbens'te) dopamin ve diğer nörotransmiterler salınabilir.Thierry ve diğerleri, 1976; Fadda ve diğerleri, 1978), bunlar kesinlikle cinsel davranışla ilgili değildir. Bu nedenle, mPFC'de (bu çalışma) veya çekirdek accumbens'te hücre dışı dopaminde bulunan dopamin artışının olduğu iddia edilebilir (Sanna ve diğerleri, 2015), cinsel aktiviteden ziyade, cinsel olarak alıcı kadının varlığı ve/veya onunla etkileşime bağlı olarak ortaya çıkan ikincil stresle veya bununla başa çıkmayla ilgilidir. Bunun, rahatsız edici koşullara yanıt olarak önemli ölçüde farklı başa çıkma tarzları ve farklı hipotalamik-hipofiz-adrenal eksen aktivasyonu gösteren RHA ve RLA sıçanlarıyla bir ilgisi olabilir (RLA sıçanları, bu eksenin aktivasyonunu RHA sıçanlarından çok daha yüksek gösterir; bkz. Carrasco ve diğerleri, 2008; Díaz-Moran ve diğerleri, 2012). Bu aynı zamanda stres hormonlarının (yani kortikosteroidlerin) mPFC ve diğer beyin alanlarındaki dopaminerjik aktiviteyi etkileyebileceği olasılığını da arttırmaktadır. Gerçekte, RLA sıçanları tepkisel başa çıkanlardır ve hipomotilite ve donma ile karakterize edilen aşırı duygusal davranışlar sergilerlerken, RHA sıçanları stres etkeni üzerinde kontrol kazanmayı amaçlayan proaktif bir başa çıkma davranışı sergilerler.Driscoll ve Bättig, 1982; Willig ve diğerleri, 1991; Escorihuela ve diğerleri, 1999; Steimer ve Driscoll, 2003; Giorgi ve diğerleri, 2007). Bunun cinsel açıdan deneyimsiz farelerde (daha önce alıcı bir dişiyle hiç etkileşime girmemiş) ve özellikle cinsel açıdan deneyimsiz RLA farelerinde (yukarıya bakın) meydana gelebileceğini tamamen göz ardı etmek imkansız olsa da, bu pek olası değildir. Deney koşullarımızda, her türlü stresten kaçınmak için mümkün olan her şey yapılıyor ("Malzemeler ve Yöntemler" Bölümüne bakın) ve deneyler, erkeklerin dişiyle hemen etkileşime girmeyeceği, ancak dişiye maruz kaldıktan 30 dakika sonra etkileşime gireceği şekilde organize ediliyor. , bu sırada dopaminde ölçülen artış, cinsel etkileşim sırasında meydana gelenden çok daha düşüktür. Roma sıçanlarının mPFC'sinde (ve nükleus accumbens'te) bulunan dopamin artışı (cinsel açıdan saf RHA ve RLA fareleri, mPFC ve nükleus accumbens'ten gelen diyalizattaki bazal hücre dışı dopamin seviyelerine sahiptir, çok benzer) gerçekten dişinin varlığı/etkileşiminden kaynaklanan stres veya onunla başa çıkma: (i) dopamin artışı, erkeklerin dişinin huzuruna çıktığı ilk dönemde çok daha yüksek bulunacaktı, daha sonra değil, çiftleşme sırasında bulunduğu gibi; ve (ii) istikrarlı bir cinsel aktivite elde edildikten sonra, yani öğrenme süreçleri (yenilikle başa çıkma dahil) tamamlandığında ve cinsel aktivitenin gerçekleştirilmesinde yalnızca küçük bir rol oynadığında dopamin artışının ortadan kalkması veya büyük ölçüde azalması beklenebilir. aktivite, aksine cinsel aktivite tekrarlandığında oluşmaya devam eder. Başka bir deyişle, mPFC'deki (ve nukleus accumbens'teki) hücre dışı dopamin, cinsel davranış öğrenildikten sonra bile çiftleşme sırasında her zaman artar. mPFC'deki (ve nukleus accumbens'teki) dopamin aktivitesindeki bu artışın fonksiyonel rolünü açıklığa kavuşturmak için daha ileri çalışmalara ihtiyaç vardır; bkz. buradaki "Giriş ve Referanslar" Bölümü).
Sonuç olarak, bu çalışma ilk kez, cinsel açıdan alıcı bir kadınla etkileşimin varlığının ve hatta daha fazlasının, cinsel açıdan saf (hiçbir zaman cinsel uyaranlara maruz kalmayan) mPFC'den diyalizattaki hücre dışı dopamin ve NA artışına yol açtığını göstermektedir. ) ve cinsel açıdan deneyimli (beş ön çiftleşme testinden geçmiş ve istikrarlı cinsel performans sergileyen) RHA ve RLA fareleri, bu fareler, rahatsız edici koşullara yanıt olarak belirgin şekilde farklı başa çıkma stilleri ve cinsel açıdan alıcı bir dişiyle de farklı çiftleşme davranışı modelleri sergiliyor. Buna göre, cinsel aktivitenin hem öngörücü hem de tüketim aşamaları sırasında, cinsel açıdan saf ve deneyimli RHA sıçanlarının diyalizatındaki dopamin ve NA konsantrasyonları, RLA muadilleriyle karşılaştırıldığında daha yüksek bulundu. Bu sonuçlar kısmen, RHA sıçanlarının mPFC'sindeki dopaminerjik ve noradrenerjik tonun, akümbens çekirdeğindeki dopamin için bildirildiği gibi, RLA sıçanlarınınkinden daha yüksek olmasından kaynaklanıyor olabilir (Sanna ve diğerleri, 2015). Üstelik, hücre dışı mPFC dopamin ve NA, cinsel açıdan deneyimli RHA'da ve daha az ölçüde RLA sıçanlarında, bazal koşullarda ve cinsel davranışın iştah açıcı ve tüketici aşamaları sırasında cinsel açıdan saf muadillerine kıyasla daha yüksek bulundu. Bu, cinsel deneyimin, her iki Roma sıçan soyunun akümbens çekirdeğindeki dopamin için bulunduğu gibi, mPFC'de dopamin ve NA nörotransmisyonunu daha da güçlendiren plastik süreçleri tetiklediğini göstermektedir.Sanna ve diğerleri, 2015). İki Roma soyunun mPFC'sindeki dopaminerjik ve noradrenerjik nörotransmisyondaki yukarıdaki farklılıklar, yalnızca farklı cinsel davranış kalıplarında değil, aynı zamanda bu iki sıçan soyunu karakterize eden farklı ve çoğu zaman zıt davranış özelliklerinde de rol oynayabilir.
Yazar Katkıları
Projeyi FS, AA, MRM, OG ve MGC tasarladı. FS ve JB, cinsel davranış ve mikrodiyaliz deneylerinden elde edilen verileri tasarladı, gerçekleştirdi ve analiz etti. MAP, OG ve MGC, Romalı fareleri seçip yetiştirdi. Çalışmayı FS, AA, MRM, OG ve MGC denetledi. Taslağı FS, AA, MRM, OG ve MGC yazdı. Tüm yazarlar sonuçları tartıştı ve makaleye yorum yaptı.
Harçlar
Bu çalışma, İtalya Üniversite ve Araştırma Bakanlığı'nın (MIUR) (Genç Araştırmacıların Bilimsel Bağımsızlığı, SIR 2014, Kod no. RBSI14IUX7) Sardunya Özerk Bölgesi'nden (ARS) (LR 7/2007) FS'ye verdiği hibelerle kısmen desteklenmiştir. , “Sardunya'da Bilimsel Araştırma ve teknolojik yeniliğin teşviki”, Avrupa Sosyal Fonu, 2007–2013, Proje Kodu No. CRP-59842) OG'ye ve Cagliari Üniversitesi'nden AA, MRM, FS, OG ve MGC'ye.
Çıkar Çatışması Beyanı
Yazarlar, araştırmanın potansiyel bir çıkar çatışması olarak yorumlanabilecek ticari veya finansal ilişkilerin olmadığı durumlarda yapıldığını beyan eder.
Teşekkürler
Hayvan barınağı ve bakımı için Cagliari Üniversitesi'nden Dr. Barbara Tuveri.
Referanslar
Afonso, VM, Sison, M., Lovic, V. ve Fleming, AS (2007). Dişi sıçandaki medial prefrontal korteks lezyonları cinsel ve annelik davranışlarını ve bunların sıralı organizasyonunu etkiler. Behav. Neurosci. 121, 515 – 526. doi: 10.1037 / 0735-7044.121.3.515
Agmo, A. ve Villalpando, A. (1995). Merkezi sinir uyarıcıları, medial prefrontal korteks lezyonları olan erkek sıçanlarda cinsel davranışı kolaylaştırır. Beyin Res. 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A., Villalpando, A., Picker, Z. ve Fernández, H. (1995). Erkek sıçanda medial prefrontal korteks lezyonları ve cinsel davranış. Beyin Res. 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A. ve Melis, MR (1995). Penis ereksiyonunun nöromodülasyonu: nörotransmiterlerin ve nöropeptitlerin rolüne genel bakış. Prog. Neurobiol. 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A. ve Melis, MR (2005). Penis ereksiyonunun merkezi kontrolü: hipotalamusun paraventriküler çekirdeğinin rolü. Prog. Neurobiol. 76, 1 – 21. doi: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A. ve Melis, MR (2013). Geçmişten günümüze nöropeptitler ve cinsel davranışın merkezi kontrolü: bir inceleme. Prog. Neurobiol. 108, 80 – 107. doi: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L. ve Coolen, LM (2006). Erkek sıçanda cinsel davranışı takiben efferentlerin medial prefrontal korteksten sinir aktivasyonuna potansiyel katkıları. Neuroscience 137, 1259 – 1276. doi: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L. ve Coolen, LM (2004). Cinsel Davranış ve cinsiyetle ilişkili çevresel ipuçları, erkek sıçanlarda mezolimbik sistemi harekete geçirir. Nöropsikofarmakoloji 29, 718 – 730. doi: 10.1038 / sj.npp.1300350
Beloate, LN, Omrani, A., Adan, RA, Webb, IC ve Coolen, LM (2016). Erkek sıçan cinsel davranışı sırasında ventral tegmental alan dopamin hücre aktivasyonu, cinsiyetten uzak durmanın ardından nöroplastisiteyi ve d-amfetamin çapraz duyarlılığını düzenler. J. Neurosci. 36, 9949 – 9961. doi: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K. ve Robinson, T. (1998). Dopaminin ödüldeki rolü nedir: hedonik etki, ödül öğrenimi veya teşvik edici belirginlik? Brain Res. Rev. 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965). Sıçanlarda yüksek oranlarda ve düşük kaçınma koşullandırma oranlarına yönelik seçim. Anim. Behav. 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL ve Bignami, G. (1965). Psikogenetik seçilimin bağıntılı etkileri: Roma'nın yüksek ve düşük kaçınmalı sıçan türleri üzerine bir çalışma. Behav. Res. Ther. 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E., Silvagni, A., Vacca, C. ve Di Chiara, G. (2006). Norepinefrin ve dopamin taşıyıcı blokajının, nükleus accumbens kabuğu, stria terminalisin yatak çekirdeği ve prefrontal korteksteki hücre dışı dopamin artışı üzerindeki kümülatif etkisi. J. Neurochem. 96, 473 – 481. doi: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E., Tanda, GL, Frau, R. ve Di Chiara, G. (1990). Noradrenalin taşıyıcısının blokajı, prefrontal kortekste hücre dışı dopamin konsantrasyonlarını artırır: dopaminin alındığına dair kanıt in vivo noradrenerjik terminaller tarafından J. Neurochem. 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975). "Dopamin metabolizmasının reseptör aracılı kontrolü" Sinaptik Öncesi ve Sonrası Reseptörler, editörler E. Usdin ve WE Bunney (New York, NY: Marcel Deker), 49–65.
Carrasco, J., Márquez, C., Nadal, R., Tobeña, A., Fernández-Teruel, A. ve Armario, A. (2008). Kendilenmiş Roma sıçan suşlarında hipotalamus-hipofiz-adrenal eksenin merkezi ve periferik bileşenlerinin karakterizasyonu. Psikonöroendokrinoloji 33, 437 – 445. doi: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T. ve Koolhaas, JM (2012). Romalı yüksek ve düşük kaçınma farelerinde dürtüsellik ve saldırgan davranış: temel farklılıklar ve ergen sosyal stresinin neden olduğu değişiklikler. Physiol. Behav. 105, 1156 – 1160. doi: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D., Piras, G., Di Chiara, G. ve Giorgi, O. (1997). Roma yüksek kaçınma ve Roma düşük kaçınma sıçanlarının CNS'sindeki dopaminerjik ve GABAerjik nörotransmisyonun biyokimyasal parametreleri. Davranış. Genet. 27, 527 – 536. doi: 10.1023 / A: 1021452814574
Corda, MG, Piras, G., Piludu, MA ve Giorgi, O. (2014). Gönüllü etanol tüketiminin çekirdekteki dopamin çıkışı üzerindeki farklı etkileri, roma yüksek ve düşük kaçınma farelerinin kabuğunu accumbens: davranışsal ve beyin mikrodiyaliz çalışması. Dünya J. Neurosci. 4, 279–292. doi: 10.4236/wjns.2014.43031
Dahlstrom, A. ve Fuxe, K. (1964). Merkezi sinir sisteminde monoamin içeren nöronların varlığına dair kanıt. I. Beyin sapı nöronlarının hücre gövdelerinde monoaminlerin gösterilmesi. Açta Physiol. Scand. Ek. 62, 1-54.
Damsma, G., Pfaus, J., Wenkstern, D., Phillips, A. ve Fibiger, H. (1992). Cinsel davranış, erkek sıçanların çekirdeğindeki accumbens ve striatumdaki dopamin iletimini arttırır: yenilik ve hareket ile karşılaştırma. Behav. Neurosci. 106, 181 – 191. doi: 10.1037 / 0735-7044.106.1.181
D'Angio, M., Serrano, A., Driscoll, P. ve Scatton, B. (1988). Stresli çevresel uyaranlar, hipo-duygusal (Roma yüksek kaçınma) sıçanların prefrontal korteksindeki hücre dışı DOPAC seviyelerini arttırır, ancak hiper-duygusal (Roma düşük kaçınma) sıçanları artırmaz. BİR in vivo voltammetrik çalışma. Beyin Res. 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M., Di Sebastiano, AR, Brown, JL, Lehman, MN ve Coolen, LM (2010). Medial prefrontal korteks lezyonları erkek sıçanlarda uyumsuz cinsel davranışlara neden olur. Biol. Psikiyatri 67, 1199 – 1204. doi: 10.1016 / j.biopsych.2009.12.029
Díaz-Moran, S., Palència, M., Mont-Cardona, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., ve diğerleri. (2012). Genetik olarak heterojen sıçanlarda başa çıkma tarzı ve stres hormonu tepkileri: Roma sıçan türleriyle karşılaştırma. Behav. Beyin Res. 228, 203 – 210. doi: 10.1016 / j.bbr.2011.12.002
Driscoll, P. ve Bättig, K. (1982). "Aktif, iki yönlü kaçınma performansındaki aşırı farklılıklar nedeniyle seçilen sıçanların davranışsal, duygusal ve nörokimyasal profilleri" Beynin Genetiği, ed. I. Lieblich (Amsterdam: Elsevier), 95–123.
Dunlop, BW ve Nemeroff, CB (2007). Depresyon patofizyolojisinde dopaminin rolü. Arch. Gen. Psikiyatri 64, 327 – 337. doi: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A., Gil, L., Aguilar, R., Tobeña, A. ve Driscoll, P. (1999). Doğuştan yüksek ve düşük kaçınmalı Roma fareleri: kaygı, yenilik arayışı ve mekik kutusu davranışlarındaki farklılıklar. Physiol. Behav. 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990). Cinsel motivasyon: erkek sıçanların iştah açıcı ve çiftleşme tepkilerinin altında yatan mekanizmaların sinirsel ve davranışsal bir analizi. Neurosci. Biobehav. Rev. 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ ve Robbins, TW (2005). Uyuşturucu bağımlılığı için sinirsel güçlendirme sistemleri: eylemlerden alışkanlıklara ve zorlamalara kadar. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fadda, F., Argiolas, A., Melis, MR, Tissari, AH, Onali, PL ve Gessa, GL (1978). Serebral korteks ve n.accumbens'te 3,4-dihidroksifenilasetik asit (DOPAC) seviyelerinde strese bağlı artış: diazepam ile tersine çevirme. Hayat Sci. 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L., Piras, G., Corda, MG ve Giorgi, O. (2009). Roma'nın yüksek ve düşük kaçınmalı sıçan soyları, intravenöz kokainin kendi kendine uygulanmasının edinilmesi, sürdürülmesi, yok edilmesi ve eski haline getirilmesi açısından farklılık gösterir. Nöropsikofarmakoloji 34, 1091 – 1101. doi: 10.1038 / npp.2008.43
Febo, M. (2011). Cinsel açıdan alıcı dişilere yaklaşırken erkek sıçanlarda prefrontal hücre ateşlemesi: kokainle etkileşimler. Synapse 65, 271 – 277. doi: 10.1002 / syn.20843
Fernández-Guasti, A., Omaña-Zapata, I., Luján, M. ve Condés-Lara, M. (1994). Siyatik sinir ligatürünün cinsel açıdan deneyimli ve deneyimsiz erkek sıçanların cinsel davranışları üzerindeki etkileri: ön kutup dekortikasyonunun etkileri. Physiol. Behav. 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobeña, A. ve Escorihuela, RM (2002). Teşvik arama davranışında farklılık gösteren iki sıçan hattında (RHA/Verh ve RLA/Verh) çevresel zenginleştirmenin yenilik arayışı, sakarin ve etanol alımı üzerindeki kalıcı etkileri. Pharmacol. Biochem. Behav. 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L., Cañete, T., Guitart-Masip, M., Fernández-Teruel, A. ve Tobeña, A. (2005). Yüksek ve düşük kaçınmalı Roma farelerinde apomorfinin neden olduğu iki farklı fenotip. Physiol. Behav. 86, 458 – 466. doi: 10.1016 / j.physbeh.2005.07.021
Giorgi, O., Lecca, D., Piras, G. ve Corda, MG (2005). Çekirdekteki dopamin salınımının farklı aktivasyonu, akut veya tekrarlanan amfetamin enjeksiyonlarından sonra çekirdek ve kabuğun oluşmasına neden olur: Roma yüksek ve düşük kaçınmalı sıçan soylarında karşılaştırmalı bir çalışma. Neuroscience 135, 987 – 998. doi: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O., Lecca, D., Piras, G., Driscoll, P. ve Corda, MG (2003a). Başa çıkma stratejilerinde caydırıcı koşullarla farklılık gösteren, psikogenetik olarak seçilmiş iki fare hattında mezokortikal dopamin salınımı ile korkuya bağlı davranışlar arasındaki ayrışma. Avro. J. Neurosci. 17, 2716 – 2726. doi: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O., Piras, G., Lecca, D., Hansson, S., Driscoll, P. ve Corda, MG (2003b). Roma yüksek ve düşük kaçınmalı sıçanlarda merkezi serotonerjik iletimin farklı nörokimyasal özellikleri. J. Neurochem. 86, 422 – 431. doi: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O., Orlandi, M., Escorihuela, RM, Driscoll, P., Lecca, D. ve Corda, MG (1994). Roma yüksek kaçınma ve Roma düşük kaçınma farelerinin beyninde GABAerjik ve dopaminerjik iletim. Beyin Res. 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O., Piras, G. ve Corda, MG (2007). Psikogenetik olarak seçilmiş Roma yüksek ve düşük kaçınmalı sıçan soyları: uyuşturucu bağımlılığına karşı bireysel hassasiyeti incelemek için bir model. Neurosci. Biobehav. Rev. 31, 148 – 163. doi: 10.1016 / j.neubiorev.2006.07.008
Goto, Y. ve Grace, AA (2005). Hedefe yönelik davranışın çekirdeğinin limbik ve kortikal dürtüsünün dopaminerjik modülasyonu. Nat. Neurosci. 8, 805 – 812. doi: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ ve Finlay, JM (1995). Sıçan medial prefrontal korteksinde endojen norepinefrinin hücre dışı dopamin üzerindeki lokal etkisi. J. Neurochem. 65, 111 – 116. doi: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, M., Guevara, MA, Cervantes, M., Morali, G. ve Corsi-Cabrera, M. (1998). Faktöriyel analiz sonucu erkek sıçanın cinsel etkileşimi sırasında kortiko-frontal EEG'nin karakteristik frekans bantları. J. Fizyoloji 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M., Prieto-Beracoechea, CA, Arteaga-Silva, M. ve Guevara, MA (2007). Sıçanlarda cinsel motivasyonlu bir görev sırasında medial ve orbital prefrontal korteksin farklı işlevselliği. Physiol. Behav. 90, 450 – 458. doi: 10.1016 / j.physbeh.2006.10.006
Boynuz, AS (1973). Sıçan beyni homojenatlarında noradrenerjik ve dopaminerjik nöronlardan sinaptozomlara katekolamin alımının inhibisyonu için yapı-aktivite ilişkileri. Br. J. Pharmacol. 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Hull, EM, Du, J., Lorrain, DS ve Matuszewich, L. (1995). Medial preoptik alanda hücre dışı dopamin: cinsel motivasyon ve çiftleşmenin hormonal kontrolü için çıkarımlar. J. Neurosci. 15, 7465-7471.
Hull, EM, Lorrain, DS, Du, J., Matuszewich, L., Lumley, LA, Putnam, SK, ve diğerleri. (1999). Cinsel davranışın kontrolünde hormon-nörotransmitter etkileşimleri. Behav. Beyin Res. 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Hull, EM, Weber, MS, Eaton, RC, Dua, R., Markowski, VP, Lumley, L., ve diğerleri. (1991). Ventral tegmental bölgedeki dopamin reseptörleri, erkek sıçanlarda çiftleşmenin motor bileşenlerini etkiler, ancak motivasyonel veya refleksif bileşenleri etkilemez. Beyin Res. 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S. ve Panksepp, J. (1999). Çekirdeğin rolü, dopamini motive edilmiş davranışta görevlendirir: ödül arayışına özel referansla birleştirici bir yorum. Brain Res. Rev. 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, M., Sone, H., Miyabara, Y. ve Tohyama, C. (2003). 2,3,7,8-tetraklorodibenzo-p-dioksine perinatal maruz kalma, neokortekste BDNF mRNA'nın aktiviteye bağlı ekspresyonunu ve yetişkinlikte erkek sıçan cinsel davranışını değiştirir. Neurotoxicology 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D., Piras, G., Driscoll, P., Giorgi, O. ve Corda, MG (2004). Çekirdek accumbens'in kabuğundaki ve çekirdeğindeki dopamin çıkışının diferansiyel aktivasyonu, bağımlılık yaratan ilaçlara verilen motor tepkilerle ilişkilidir: Roma yüksek ve düşük kaçınmalı sıçanlarda bir beyin diyalizi çalışması. nörofarmakolojisi 46, 688 – 699. doi: 10.1016 / j.neuropharm.2003.11.011
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Donaire, R., Sabariego, M., Fernández-Teruel, A., ve diğerleri. (2014a). Etanol tercihi ile heyecan/yenilik arayışı arasındaki ilişki. Physiol. Behav. 133, 53 – 60. doi: 10.1016 / j.physbeh.2014.05.003
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A., Papini, MR ve Torres, C. (2014b). Sıçanlarda teşvik kaybının neden olduğu anti-anksiyete kendi kendine ilaç tedavisi. Physiol. Behav. 123, 86 – 92. doi: 10.1016 / j.physbeh.2013.10.002
Masana, M., Bortolozzi, A. ve Artigas, F. (2011). Noradrenerjik ilaçlarla mezokortikal dopaminerjik iletimin seçici olarak arttırılması: Şizofrenide terapötik fırsatlar. Int. J. Neuropsychopharmacol. 14, 53 – 68. doi: 10.1017 / s1461145710000908
Meisel, RL ve Sachs, BD (1994). "Erkek cinsel davranışının fizyolojisi" Üreme Fizyolojisi, Cilt. 2, 2. Baskı, editörler E. Knobil ve J. Neil (New York, NY: Raven Press), 3–96.
Melis, MR ve Argiolas, A. (1995). Dopamin ve cinsel davranış. Neurosci. Biobehav. Rev. 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR ve Argiolas, A. (2011). Penis ereksiyonunun merkezi kontrolü: erkek sıçanlarda oksitosinin rolünün ve bunun dopamin ve glutamik asit ile etkileşiminin yeniden gözden geçirilmesi. Neurosci. Biobehav. Rev. 35, 939 – 955. doi: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., et al. (2007). Ventral tegmental alana enjekte edilen oksitosin, penis ereksiyonunu indükler ve erkek sıçanların hipotalamusunun nükleus accumbens'inde ve paraventriküler çekirdeğinde hücre dışı dopamini arttırır. Avro. J. Neurosci. 26, 1026 – 1035. doi: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., ve diğerleri. (2010). Oksitosin, ventral subikuluma enjekte edildiğinde penis ereksiyonunu indükler: nitrik oksit ve glutamik asidin rolü. nörofarmakolojisi 58, 1153 – 1160. doi: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S., Mascia, MS, Cortis, L. ve Argiolas, A. (2003). Cinsel aktivite sırasında erkek sıçanların paraventriküler çekirdeğinde hücre dışı dopamin artar. Avro. J. Neurosci. 17, 1266 – 1272. doi: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S., Sanna, F., Boi, A. ve Argiolas, A. (2009). Amigdalanın ventral subikulumuna veya posteromedial kortikal çekirdeğine enjekte edilen oksitosin, penis ereksiyonunu indükler ve erkek sıçanların nükleus accumbens'indeki hücre dışı dopamin seviyelerini arttırır. Avro. J. Neurosci. 30, 1349 – 1357. doi: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M., Cardona, D., Gómez, MJ, Sánchez-Santed, F., Tobeña, A., Fernández-Teruel, A., ve diğerleri. (2010). Roma yüksek ve düşük kaçınmalı sıçan türlerinde dürtüsellik karakterizasyonu: davranışsal ve nörokimyasal farklılıklar. Nöropsikofarmakoloji 35, 1198 – 1208. doi: 10.1038 / npp.2009.224
Oliveras, I., Río-Álamos, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., Giorgi, O., ve diğerleri. (2015). Ön uyarı inhibisyonu, doğuştan Romalı yüksek ve düşük kaçınmalı sıçanlarda ve genetik olarak heterojen NIH-HS sıçanlarında uzamsal çalışma belleği performansını öngörür: şizofrenide dikkat öncesi ve bilişsel anomalilerin incelenmesi için uygunluk. Ön. Behav. Neurosci. 9: 213. doi: 10.3389 / fnbeh.2015.00213
Paxinos, G. ve Watson, C. (2004). Stereotaksik Koordinatlarda Sıçan Beyin, 4. Baskı, New York: Academic Press.
Pfaus, JG (2010). Dopamin: erkeklerin en az 200 milyon yıl boyunca çiftleşmesine yardımcı olmak: Kleitz-Nelson ve diğerleri üzerine teorik yorum. (2010). Behav. Neurosci. 124, 877 – 880. doi: 10.1037 / a0021823
Pfaus, JG, Damsma, G., Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG, ve diğerleri. (1990). Cinsel davranış, erkek sıçanda merkezi dopamin iletimini arttırır. Beyin Res. 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG ve Everitt, BJ (1995). “Cinsel davranışın psikofarmakolojisi”, Psikofarmakoloji: Dördüncü Nesil İlerleme, editörler FE Bloom ve DJ Kupfer (New York, NY: Raven Press), 742–758.
Pfaus, JG ve Phillips, AG (1991). Erkek sıçanda cinsel davranışın öngörücü ve tamamlayıcı yönlerinde dopaminin rolü. Behav. Neurosci. 105, 727 – 747. doi: 10.1037 / 0735-7044.105.5.727
Piras, G., Giorgi, O. ve Corda, MG (2010). Antidepresanların, başa çıkma stratejilerinde caydırıcı koşullarla farklılık gösteren, psikogenetik olarak seçilmiş iki sıçan hattının zorla yüzme testindeki performansı üzerindeki etkileri. Psikofarmakoloji 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G., Piludu, MA, Giorgi, O. ve Corda, MG (2014). Kronik antidepresan tedavilerinin, varsayılan bir genetik kırılganlık modelinde (Roma düşük kaçınmalı fareler) ve stresin neden olduğu depresyona karşı direnç (Roma yüksek kaçınmalı fareler) üzerindeki etkileri. Psikofarmakoloji 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V., Mouzon, K., Nestler, EJ, Lehman, MN, ve diğerleri. (2010). Çekirdek accumbens'teki ΔFosB, cinsel ödülün etkilerini güçlendirmek için kritik öneme sahiptir. Genlerin Beyin Davranışı. 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J., Van, S., Frohmader, KS, ve diğerleri. (2014). Ventral tegmental bölgedeki dopaminerjik nöronların endojen opioid kaynaklı nöroplastisitesi, doğal ve opiat ödülünü etkiler. J. Neurosci. 34, 8825 – 8836. doi: 10.1523 / JNEUROSCI.0133-14.2014
Pitchers, KK, Vialou, V., Nestler, EJ, Laviolette, SR, Lehman, MN ve Coolen, LM (2013). Doğal ve ilaç ödülleri, ΔFosB'nin temel aracı olduğu ortak sinirsel esneklik mekanizmaları üzerinde hareket eder. J. Neurosci. 33, 3434 – 3442. doi: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E., Matochik, J., Barfield, R. ve Auerbach, S. (1990). Farelerde çekirdekteki dopamin salınımının erkeksi cinsel davranışla ilişkilendirilmesi. Beyin Res. 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW ve Arnsten, AFT (2009). Fronto-yürütücü fonksiyonun nöropsikofarmakolojisi: monoaminerjik modülasyon. Ann. Rev. Neurosci. 32, 267 – 287. doi: 10.1146 / annurev.neuro.051508.135535
Sabariego, M., Morón, I., Gómez, MJ, Donaire, R., Tobeña, A., Fernández-Teruel, A., ve diğerleri. (2013). Kendi içinde melezlenmiş roman yüksek (RHA-I) ve roman düşük (RLA-I) kaçınma sıçanlarında teşvik kaybı ve hipokampal gen ekspresyonu. Behav. Beyin Res. 257, 62 – 70. doi: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000). Erektil fonksiyonun, erektil disfonksiyonun ve cinsel uyarılmanın fizyolojisi ve sınıflandırılmasına bağlamsal yaklaşımlar. Neurosci. Biobehav. Rev. 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K., Citro, JK, Daniels, SB ve Natoli, JH (1994). Kızgın dişilerden gelen temassız uyarı, sıçanlarda peniste ereksiyona neden olur. Physiol. Behav. 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD ve Barfield, RJ (1976). Sıçanlarda erkeksi çiftleşme davranışının fonksiyonel analizi. Av. Damızlık. Davranış. 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M., Yohn, SE, López-Cruz, L., San Miguel, N. ve Correa, M. (2016). Mezolimbik dopamin ve motive olmuş davranışın düzenlenmesi. Curr. Üst. Behav. Neurosci. 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F., Corda, MG, Melis, MR, Piludu, MA, Giorgi, O. ve Argiolas, A. (2014a). Erkek Romalı yüksek ve düşük kaçınmalı fareler, farklı çiftleşme davranışı modelleri gösterir: Sprague dawley fareleriyle karşılaştırma. Physiol. Behav. 127, 27 – 36. doi: 10.1016 / j.physbeh.2014.01.002
Sanna, F., Piludu, MA, Corda, MG, Argiolas, A., Giorgi, O. ve Melis, MR (2014b). Dopamin, Roma'daki yüksek ve düşük kaçınmalı sıçanların farklı çiftleşme davranışı modellerinde rol oynar: apomorfin ve haloperidol ile yapılan çalışmalar. Pharmacol. Biochem. Behav. 124, 211 – 219. doi: 10.1016 / j.pbb.2014.06.012
Sanna, F., Piludu, MA, Corda, MG, Melis, MR, Giorgi, O. ve Argiolas, A. (2015). Roma yüksek ve düşük kaçınma sıçanları arasındaki cinsel davranış farklılıklarına dopaminin katılımı: intraserebral mikrodiyaliz çalışması. Behav. Beyin Res. 281, 177 – 186. doi: 10.1016 / j.bbr.2014.12.009
Scatton, B., D'Angio, M., Driscoll, P. ve Serrano, A. (1988). Bir in vivo Farklı duygusallık düzeylerine sahip sıçan türlerinde mezokortikal ve mezoakcumbens dopaminerjik nöronların çevresel uyaranlara tepkisinin voltametrik çalışması. Ann. NY Acad. Sci. 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T. ve Driscoll, P. (2003). Psikogenetik olarak seçilmiş Roma yüksek (RHA) ve düşük (RLA) kaçınma farelerinde farklı stres tepkileri ve başa çıkma stilleri: davranışsal, nöroendokrin ve gelişimsel yönler. stres 6, 87 – 100. doi: 10.1080 / 1025389031000111320
Succu, S., Sanna, F., Argiolas, A. ve Melis, MR (2011). Hipokampal ventral subikuluma enjekte edilen oksitosin, ventral tegmental alanda glutamaterjik nörotransmisyonu artırarak erkek sıçanlarda penis ereksiyonunu indükler. nörofarmakolojisi 61, 181 – 188. doi: 10.1016 / j.neuropharm.2011.03.026
Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, ve diğerleri. (2008). Oksitosin, erkek sıçanların ventral tegmental alanına enjekte edildiğinde penis ereksiyonunu indükler: nitrik oksit ve siklik GMP'nin rolü. Avro. J. Neurosci. 28, 813 – 821. doi: 10.1111 / j.1460-9568.2008.06385.x
Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A. ve Melis, MR (2007). Erkek sıçanların hipotalamusunun paraventriküler çekirdeğindeki dopamin reseptörlerinin uyarılması, penis ereksiyonunu indükler ve çekirdek akümülatörlerdeki hücre dışı dopamini arttırır: merkezi oksitosinin katılımı. nörofarmakolojisi 52, 1034 – 1043. doi: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G. ve Glowinski, J. (1976). Mezokortikal DA sisteminin stresle seçici aktivasyonu. Tabiat 263, 242 – 244. doi: 10.1038 / 263242a0
Wenkstern, D., Pfaus, J. ve Fibiger, H. (1993). Erkek sıçanların cinsel açıdan alıcı dişi sıçanlara ilk maruz kalmaları sırasında nükleus accumbens'inde dopamin iletimi artar. Beyin Res. 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik, BHC, De Boer, P., De Vries, JB ve Long, S. (1998). Antipsikotik ilaçlar, sıçan beyninin medial prefrontal korteksinde dopamin ve noradrenalin salınımı üzerinde benzer etkilere neden olur. Avro. J. Pharmacol. 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F., M'Harzi, M., Bardelay, C., Viet, D. ve Delacour, J. (1991). Çalışma belleğinin incelenmesi için psikogenetik bir model olarak Roma suşları: davranışsal ve biyokimyasal veriler. Pharmacol. Biochem. Behav. 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
Anahtar Kelimeler: cinsel davranış, dopamin, noradrenalin, mPFC, mikrodiyaliz, RHA ve RLA sıçanları
Alıntı: Sanna F, Bratzu J, Piludu MA, Corda MG, Melis MR, Giorgi O ve Argiolas A (2017) Dopamin, Noradrenalin ve Romalı Yüksek ve Düşük Kaçınma Oranlı Erkek Sıçanlar Arasındaki Cinsel Davranıştaki Farklılıklar: Medial Prefrontal Kortekste Mikrodiyaliz Çalışması . Ön. Davranış. Nörobilim. 11:108. doi: 10.3389/fnbeh.2017.00108
Alındığı tarih: 22 Aralık 2016; Kabul Tarihi: 19 Mayıs 2017;
Yayın tarihi: 07 Haziran 2017.