 Martin Zack
Martin Zack- 1Cognitive Psychopharmacology Laboratory, Neuroscience Department, Centre for Addiction and Mental Health, Toronto, ON, Canada
- 2Translational Neuroscience Program, Department of Psychiatry, School of Medicine, University of Pennsylvania, Philadelphia, PA, USA
- 3Biopsychology Section, Neuroscience Department, Centre for Addiction and Mental Health, Toronto, ON, Canada
Addiction is considered to be a brain disease caused by chronic exposure to drugs. Sensitization of brain dopamine (DA) systems partly mediates this effect. Pathological gambling (PG) is considered to be a behavioral addiction. Therefore, PG may be caused by chronic exposure to gambling. Identifying a gambling-induced sensitization of DA systems would support this possibility. Gambling rewards evoke DA release. One episode of slot machine play shifts the DA response from reward delivery to onset of cues (spinning reels) for reward, in line with temporal difference learning principles. Thus, conditioned stimuli (CS) play a key role in DA responses to gambling. In primates, DA response to a CS is strongest when reward probability is 50%. Under this schedule the CS elicits an expectancy of reward but provides no information about whether it will occur on a given trial. During gambling, a 50% schedule should elicit maximal DA release. This closely matches reward frequency (46%) on a commercial slot machine. DA release can contribute to sensitization, especially for amphetamine. Chronic exposure to a CS that predicts reward 50% of the time could mimic this effect. We tested this hypothesis in three studies with rats. Animals received 15 × 45-min exposures to a CS that predicted reward with a probability of 0, 25, 50, 75, or 100%. The CS was a light; the reward was a 10% sucrose solution. After training, rats received a sensitizing regimen of five separate doses (1 mg/kg) of d-amphetamine. Lastly they received a 0.5 or 1 mg/kg amphetamine challenge prior to a 90-min locomotor activity test. In all three studies the 50% group displayed greater activity than the other groups in response to both challenge doses. Effect sizes were modest but consistent, as reflected by a significant group × rank association (ϕ = 0.986, p = 0.025). Chronic exposure to a gambling-like schedule of reward predictive stimuli can promote sensitization to amphetamine much like exposure to amphetamine itself.
ifihan
Addiction has been characterized as a brain disease caused by chronic exposure to drugs of abuse (Leshner, 1997). Neuroplasticity is thought to mediate the effects of such exposure (Nestler, 2001). Sensitization of brain dopamine (DA) systems is a form of neuroplasticity implicated in hyper-reactivity to conditioned stimuli (CS) for drugs, and compulsive drug seeking (Robinson ati Berridge, 2001). Sensitization has been operationally defined by increased DA release in response to a CS for reward and by increased locomotor response to pharmacological DA challenge (Robinson ati Berridge, 1993; Pierce ati Kalivas, 1997; Vanderschuren ati Kalivas, 2000). Although sensitization is only one of many brain changes linked with addiction (cf. Robbins ati Everitt, 1999; Koob ati Le Moal, 2008), changes in presynaptic dopamine release have been suggested to represent common neuroadaptations involved in addiction-based drug-seeking (e.g., relapse), in that drugs that induce locomotor sensitization to opiate (e.g., morphine) or stimulant challenge (e.g., amphetamine), also cause reinstatement of extinguished operant responses for heroin or cocaine self-administration—an animal model of relapse (Vanderschuren et al., 1999). Evidence that incentive sensitization (increased value of drug reward) is most pronounced after initial exposure to addictive drugs further suggests that sensitization may be involved in the early stages of addiction as well (Vanderschuren ati Pierce, ọdun 2010).
Pathological gambling (PG) has been described as a behavioral addiction and recently reclassified to the same category as substance dependence disorders in the 5th edition of the Diagnostic and Statistical Manual of Mental Disorders (Frascella et al., 2010; A.P.A., 2013). This implies that PG may be caused by chronic exposure to gambling-like activity, that common mechanisms may mediate the effects of gambling and drug exposure (Zack ati Poulos, 2009; Leeman ati Potenza, 2012); ati pe ifamọ ti ọpọlọ DA awọn ipa ọna le jẹ ọkan pataki eroja ti ilana yii.
Clinical evidence indirectly supports this possibility: Using positron emission tomography (PET) Boileau and colleagues found that male PG subjects exhibit significantly greater striatal DA release in response to amphetamine (0.4 mg/kg) than healthy male controls (Boileau et al., 2013). Overall group differences were significant in the associative and somatosensory striatum. In the limbic striatum, which includes the nucleus accumbens, the groups did not differ. However, in PG subjects, DA release in the limbic striatum correlated directly with the severity of PG symptoms. These findings are consistent with sensitization of brain DA pathways in PG, but also suggest some important differences with human substance dependent individuals and with the classic animal model of amphetamine sensitization. Unlike PG subjects and animals exposed to low doses of amphetamine (cf. Robinson et al., 1982), humans with substance dependence consistently exhibit decreased DA release to a stimulant challenge (Volkow et al., 1997; Martinez et al., 2007), and evidence from animals suggests that this may reflect deficits in DA function during the initial stages of abstinence following binge patterns of substance abuse (Mateo et al., 2005). In studies where stimulant sensitization is demonstrated in animals, enhanced DA release is usually observed in the limbic striatum rather than the dorsal (associative, somatosensory) striatum (Vezina, 2004). However, cue-induced (i.e., conditioned) drug-seeking in animals repeatedly exposed to cocaine has been linked with enhanced DA release in the dorsal striatum, a result thought to indicate a more habitual form of motivated behavior (Ito et al., 2002). Thus, the overall elevation in DA release in dorsal regions in PG subjects may be related to habit-based (inflexible, routinized) reward seeking involving “a progression from ventral to more dorsal domains of the striatum” (Everitt ati Robbins, 2005, p. 1481), lakoko ti itusilẹ DA ti o da lori biba ni limbic striatum ninu awọn koko-ọrọ wọnyi le ṣe deede diẹ sii ni pẹkipẹki si ifamọ ifamọ bi igbagbogbo ṣe apẹrẹ ninu awọn ẹranko. Awọn awari PET ko le ṣafihan boya DA hyper-reactivity jẹ ẹya-tẹlẹ ti awọn koko-ọrọ PG wọnyi, abajade ti ifihan ere, tabi abajade ti ilana miiran patapata. Lati koju ibeere yii, o jẹ dandan lati ṣafihan ifamọ ti ifamọ nipasẹ ifihan ere onibaje ni awọn koko-ọrọ ti o jẹ deede ṣaaju ifihan. Eleyi ji awọn ibeere bi si ohun ti awọn ẹya ara ẹrọ ti ayo ni o seese lati jeki ifamọ.
Skinner noted that the variable schedule of reinforcement was fundamental to gambling’s allure (or at least its persistence) (Skinner, 1953). Betting behavior in a slot machine game conforms well to the basic principles of instrumental conditioning, as reflected by a prospective correlation between monetary payoff and bet size on consecutive spins (Tremblay et al., 2011). Bayi, oniyipada ratio operant fesi han lati pese ohun ita wulo awoṣe ti Iho ẹrọ ayo .
Recent research with animals provides strong initial support for a causal effect of gambling exposure on sensitization. Singer and colleagues examined the effects of 55 1–h daily sessions of fixed (FR20) or variable (VR20) saccharin reinforcement in an operant lever-press paradigm on subsequent locomotor response to low dose (0.5 mg/kg) amphetamine in healthy male (Sprague Dawley) rats (Olorin et al., 2012). Wọn ṣe akiyesi pe, ti ere ba yori si ifamọ, awọn eku ti o farahan si iṣeto oniyipada, eyiti o ṣe adaṣe ere, o yẹ ki o ṣafihan esi nla si amphetamine ju awọn eku ti o farahan si iṣeto ti o wa titi. Gẹgẹbi asọtẹlẹ, ẹgbẹ VR20 ṣe afihan idahun locomotor ti o tobi ju 50% si amphetamine ju ẹgbẹ FR20 lọ. Ni idakeji, awọn ẹgbẹ ṣe afihan ipo deede ni atẹle abẹrẹ iyọ kan. Awọn awari wọnyi jẹrisi pe ifihan onibaje si imuduro oniyipada to lati fa ifasẹyin hyper-si ipenija DA kan ninu awọn ẹranko ti o ni ilera ti a sọtọ si awọn iṣeto oniwun.
A number of questions arise from this result: First, to what extent does the perceived contingency—or lack thereof—between the operant response and its outcome mediate these effects? In learning terms, does this effect involve a “response-outcome expectancy,” or might a similar effect be seen in the absence of an operant response, i.e., “a stimulus-outcome expectancy” in a Pavlovian paradigm (cf. Awọn Bolulu, 1972)? Ẹlẹẹkeji, ṣe iwọn airotẹlẹ laarin iṣẹlẹ iṣaaju (idahun tabi iyanju) ati abajade rẹ ni ipa iwọn ifamọ bi?
Ibeere keji kan ipa ti aidaniloju ninu ifamọ. Fun apẹẹrẹ, ṣe awọn ere ti abajade jẹ laileto nitootọ — airotẹlẹ patapata — ni agbara nla lati fa ifamọ ju awọn ere nibiti awọn aidọgba ti bori ti ṣalaye ni kedere ṣugbọn kii ṣe laileto, paapaa ti oṣuwọn pipe ti ere ba lọ silẹ? Iwadi ti o wa lọwọlọwọ koju awọn ibeere wọnyi.
The experimental design was informed by a seminal study on reward expectancy and DA neuron response in monkeys (Fiorillo et al., 2003). Awọn ẹranko ti o wa ninu iwadi yẹn gba ẹsan oje kan (US) labẹ 0, 25, 50, 75, tabi 100% awọn iṣeto ipin ipin. Awọn iṣeto naa jẹ apẹrẹ nipasẹ 1 ti 4 oriṣiriṣi CS (awọn aami). Eto 0% jiṣẹ ẹsan nigbagbogbo bi iṣeto 100%, ṣugbọn o yọkuro CS. Oṣuwọn ibọn ti awọn neurons DA lakoko aarin laarin ibẹrẹ CS ati ifijiṣẹ AMẸRIKA tabi yiyọ kuro jẹ iwọn igbẹkẹle bọtini. Iwadi na rii pe idahun DA pọ si bi iṣẹ ti aidaniloju ti ifijiṣẹ ere. Nitorinaa, labẹ iṣeto 100% CS ti yọ iṣẹ ṣiṣe kekere kuro, labẹ awọn eto 25 ati 75%, CS yọkuro iwọntunwọnsi ati awọn ipele iṣẹ ṣiṣe ti o jọra, ati labẹ iṣeto 50% CS yọkuro iṣẹ ṣiṣe to pọ julọ. Ninu ọran kọọkan, oṣuwọn ibọn pọ si ni akoko aarin aarin CS-US, ie, bi ireti ti sunmọ eso.
Awọn awari wọnyi fihan pe iṣẹ ṣiṣe DA kii ṣe iyatọ nikan pẹlu boya tabi kii ṣe ẹsan jẹ idaniloju (Ipin Ti o wa titi) tabi aidaniloju (Ipin Ayipada), ṣugbọn tun yatọ ni ipin idakeji si iye alaye nipa ifijiṣẹ ere ti o gbejade nipasẹ CS. Ni ipo 100%, CS nfa ireti ere ati tun sọ asọtẹlẹ ifijiṣẹ rẹ ni pipe. Ni awọn ipo 25 ati 75%, CS ṣe ifojusọna ireti ati asọtẹlẹ ifijiṣẹ ere ni mẹta ninu awọn igba mẹrin. Ni ipo 50% CS n fa ireti ireti ṣugbọn ko pese alaye nipa ifijiṣẹ ere kọja aye nikan. Da lori awọn awari wọn, Fiorillo et al. pari: "Eleyi aidaniloju-induced ilosoke ninu dopamine le tiwon si funlebun-ini ti ayo "(P. 1901).
The effects of 50% variable reward in a single session should not change over the course of multiple sessions because the likelihood of reward remains entirely unpredictable on every trial. Thus, when considering the conditions that would maximize chronic activation of DA neurons over repeated episodes of gambling the 50% schedule should engender the most enduring as well as the most robust effect. This is noteworthy given that the long run rate of reward (payoff > 0) observed over thousands of spins on a commercial slot machine was 45.8% (Tremblay et al., 2011). Nitorinaa, ere oniyipada 50% han lati ṣe afihan deede iṣeto isanwo ti a nṣakoso nipasẹ awọn ẹrọ ayokele gangan.
Iwadi lọwọlọwọ lo awọn iṣeto idabobo kanna bi Fiorillo et al. ni a onibaje ifihan, laarin-ẹgbẹ 'apẹrẹ pẹlu eku. Awọn ẹranko lọ ~ ọsẹ 3 ti awọn akoko idabobo ojoojumọ, nibiti CS (ina) ti so pọ pẹlu US kan (iye kekere ti sucrose). Lẹhin ipele ikẹkọ, awọn ẹranko sinmi ṣaaju iṣayẹwo ti ifamọ ti atọka nipasẹ idahun locomotor si amphetamine. Da lori awọn iwe-iwe, o ti sọtẹlẹ pe awọn eku ti o farahan si awọn iṣeto ere oriṣiriṣi kii yoo yato ninu ihuwasi locomotor ọfẹ ti oogun wọn ṣugbọn yoo ṣe afihan awọn ipele ti o yatọ pupọ ti locomotion ni atẹle amphetamine, pẹlu ẹgbẹ 50% ti n ṣafihan idahun locomotor nla si oogun naa ni ibatan si awọn ẹgbẹ miiran lori ilana awọn abere, apẹrẹ ti yoo nireti ti awọn ẹranko 50% ti farahan tẹlẹ si awọn abere afikun ti amphetamine funrararẹ (ie, ifamọ agbelebu).
Ṣe idanwo 1
Awon nkan ise nkan ati awon ona lati se nkan
Awọn koko
Awọn ẹgbẹ mẹrin (n = 8/ẹgbẹ) ti agbalagba (300-350 g) awọn eku Sprague-Dawley ọkunrin (Charles River, St. Constant, Quebec, Canada) ni a gbe ni ẹyọkan ni awọn apoti polycarbonate ti o han gbangba (20 × 43 × 22 cm) labẹ iyipada 12: 12 ina-dudu ọmọ. Wọn ti gba ipolongo libitum access to food and water, and daily handling by an experimenter for 2 weeks prior to the study. Each group was conditioned under one of four variable reward schedules: 0, 25, 50, or 100%. The 75% group was omitted in this initial study, as Fiorillo et al. (2003) found equivalent post-CS DA release under 25 and 75% reward schedules, such that both conditions led to greater DA release than did the 100% CS-US condition, but less than the 50% condition.
ohun elo
Wiwọle si awọn igbejade sucrose ati si CS ni a pese ni ẹyọkan ninu awọn apoti idamu iṣẹ (33 × 31 × 29 cm). Apotin dopodopo yin awuwlena po linlinnamẹwe huhlọnnọ de po, he tin to adó nukọn. Imọlẹ ti o wa ni oke iwe irohin naa ṣiṣẹ bi CS. Dipper olomi ti o ni idari solenoid le gbe soke si ilẹ ti iwe irohin naa. Awọn iṣẹlẹ ti o wa ninu apoti jẹ iṣakoso nipasẹ ohun elo Med Associates ati sọfitiwia, ni lilo eto inu ile ti a kọ sinu MED-PC. Idanwo Locomotor ni a ṣe ni ẹyọkan ni awọn agọ Plexiglas (27 × 48 × 20 cm). Ẹyẹ kọọkan ni ipese pẹlu eto ibojuwo ti o ni awọn sẹẹli fọto-tan ina mẹfa lati ṣe awari gbigbe petele.
ilana
ikẹkọ. Iwadi naa ni a ṣe ni ibamu pẹlu awọn ilana iṣe ti a ṣeto nipasẹ Igbimọ Kanada lori Itọju Ẹranko. Awọn eku jẹ ounjẹ-ihamọ si 90% ti iwuwo ara wọn fun iye akoko ikẹkọ ati gbele ni ẹyọkan. Eku kọọkan gba awọn ọjọ 15 ti ikẹkọ ere sucrose (ojutu omi 10% ni 0.06 milimita fun ẹsan): Awọn ọjọ itẹlera 5 × 3 ọsẹ, pẹlu awọn ipari ose. A tọju awọn ẹranko lori chow boṣewa ṣaaju ati lẹhin ipele ikẹkọ; ifihan sucrose jẹ ihamọ si awọn igba ikẹkọ ~ meedogun-40-min. Ipejọ ojoojumọ kọọkan ni awọn igbejade iyanju 15 (ina kan; CS), ọkọọkan ti yapa nipasẹ aarin-idanwo aarin ti 120 s. Imọlẹ naa wa ni apa oke ti iwe irohin naa, o wa lori fun awọn iṣẹju 25, pẹlu sucrose ti o wa lakoko awọn iṣẹju 5 to kẹhin. Ninu ọran ti ẹgbẹ 0 sucrose dipper ti dide ni gbogbo 140 s (fun awọn iṣẹju 5) ṣugbọn ina ayun naa ko tan. Eyi dọgba aarin laarin awọn ifarahan ti dipper ni ẹgbẹ 0 ati awọn ẹgbẹ miiran (120 + 25 s). Ilana itọju kọọkan duro ~ 40 min. Ni apapọ, ẹgbẹ 25 gba sucrose lẹẹkan fun gbogbo awọn igbejade CS mẹrin; ẹgbẹ 50 gba sucrose lẹẹkan fun gbogbo meji CS ifarahan, ati ẹgbẹ 100 gba sucrose lẹhin ti gbogbo CS igbejade.
HIV. Ọsẹ meji lẹhin iraye si sucrose ti o kẹhin (tabi “itumọ”) igba, idahun locomotor si d-amphetamine (AMPH; ip) jẹ iṣiro. Awọn eku ni a fun ni awọn akoko 2-h mẹta lati ṣe deede si awọn apoti locomotor, atẹle nipasẹ awọn akoko idanwo AMPH mẹfa. Awọn ọjọ idanwo AMPH waye ni awọn aaye arin 1-wk. Ni awọn ọjọ idanwo, awọn eku ni a fun ni iṣẹju 30 lati ṣe deede si awọn apoti lẹhinna gba iwọn 0.5 mg / kg kan ti AMPH tẹle, ni awọn akoko ọsẹ lọtọ, nipasẹ awọn iwọn 1.0 mg / kg marun (iwọn iwọn kan fun ọjọ kan) ni awọn ọjọ idanwo 1 nipasẹ 5 Locomotion post-AMPH ni a ṣe ayẹwo fun awọn iṣẹju 90 ni igba kọọkan.
Data analytic ona
Awọn itupalẹ iṣiro ni a ṣe pẹlu SPSS (v. 16 ati v. 21; SPSS Inc., Chicago IL). Idahun ihuwasi lẹsẹkẹsẹ si CS ni a ṣe ayẹwo ni awọn ofin ti awọn pokes imu sinu iho nibiti a ti pin sucrose naa. Nọmba apapọ ti awọn pokes imu lakoko aarin yii (5 s fun idanwo) lẹhinna ni akawe si nọmba apapọ ti awọn pokes imu fun iye akoko kanna (5 s) ni aropin ni akoko ti CS ko si. Ẹgbẹ × Ikoni ANOVAs ti imu-pokes pẹlu CS ti o wa ati isansa tọpa imudani ti idahun iyasoto si ifẹnukonu ati awọn idahun aibikita imu poke awọn idahun labẹ awọn iṣeto oriṣiriṣi lori akoko ti awọn akoko ikẹkọ sucrose 15.
Awọn ipa ti itọju lori awọn idahun locomotor ni a ṣe ayẹwo pẹlu Ẹgbẹ × Ikoni ANOVAs fun ipele ibugbe ti ko ni oogun (awọn akoko mẹta), ifamọ tẹlẹ 0.5 mg/kg AMPH ipenija (igba kan), ati lakoko igba marun-1 mg / kg AMPH Ilana ifamọ, nigbati a nireti awọn ẹgbẹ lati yatọ ni idahun si awọn iwọn lilo ti AMPH leralera. Ẹgbẹ × Ikoni ANOVAs tun ṣe ayẹwo awọn idahun locomotor ti ko ni oogun lakoko akoko ibugbe abẹrẹ ṣaaju-30-min lati igba idanwo AMPH kọọkan. Awọn afiwera ti a gbero ṣe ayẹwo iyatọ ninu iṣẹ ṣiṣe fun ẹgbẹ 50 vs. ẹgbẹ 0 (ko si iṣakoso ireti) ati ẹgbẹ 100 (ko si iṣakoso aidaniloju), nipasẹ ọna ti t- awọn idanwo (Howell, 1992), using the MS error and df error terms for the relevant effect (i.e., group or group × session interaction) from the ANOVA (Winer, 1971). Awọn itupalẹ aṣa ilopọpo ṣe idanwo profaili ti awọn ayipada ni akoko awọn akoko.
Lati pinnu boya awọn idahun isunmọ ni wiwa ati isansa ti CS lakoko awọn akoko ikẹkọ sucrose 15 ṣe alabapin si iyatọ ninu idahun locomotor si AMPH, tabi awọn iyatọ ẹgbẹ alaja ni idahun AMPH, awọn itupalẹ atẹle ti covariance (ANCOVAs) ni a ṣe lori AMPH data locomotor, pẹlu lapapọ awọn pokes imu (apao fun awọn akoko 15) nigbati CS ko si bi alajọpọ. Ipa pataki ti covariate yoo tọka pe awọn idahun ọna ti ko ni oogun ni iwọntunwọnsi (ni ipa ti agbara) awọn ipa ti ẹgbẹ tabi igba. Idinku ni pataki ti awọn ipa ti ẹgbẹ tabi igba ni iwaju iṣọpọ pataki kan yoo fihan pe awọn idahun isunmọ ti ni ilaja (ṣiṣiro fun) awọn ipa ti ẹgbẹ tabi igba. Idinku pataki ti ẹgbẹ tabi awọn ipa igba ni isansa ti ipa isomọ pataki kan yoo ṣe afihan ipadanu ti agbara iṣiro lasan nitori ibi-ipo ti df lati ọrọ aṣiṣe si iṣọpọ, ati pe kii yoo ni ipa lori itumọ ti awọn ipa ti ẹgbẹ tabi igba.
awọn esi
Imu pokes nigba sucrose karabosipo igba
CS lọwọlọwọ. Nọmba 1A fihan awọn pokes imu ti o tumọ fun awọn ẹgbẹ 25, 50, ati 100 lakoko ti CS wa lori awọn akoko isọdọtun sucrose 15 (awọn pokes imu ko ni koodu fun ẹgbẹ 0, eyiti ko gba CS). Ẹgbẹ 3 × 15 ANOVA ti ṣe agbejade awọn ipa akọkọ pataki ti Ẹgbẹ, F(2, 21) = 5.63, p = 0.011, ati Igba, F(14, 294) = 14.00, p <0.001, pẹlu ibaraẹnisọrọ Ẹgbẹ pataki × Ibaṣepọ, F(28, 294) = 2.93, p <0.001. Olusin 1A tọkasi pe ipa akọkọ ti Ikoni ṣe afihan ilosoke ninu awọn pokes imu kọja awọn akoko ni gbogbo awọn ẹgbẹ mẹta, ati pe ipa akọkọ ti Ẹgbẹ ṣe afihan awọn ikun gbogbogbo ti o ga julọ ni ẹgbẹ 100 vs. ibaraenisepo fun aṣa onigun, F(2, 21) = 4.42, p = 0.030, tọkasi igbega ni iyara, fibọ, ati ipele ni pipa ni awọn ere imu lori awọn akoko ni ẹgbẹ 100, bii ilodi si ilosoke laini lori awọn akoko ni ẹgbẹ 50, ati ilosoke laini aijinile lori awọn akoko ni ẹgbẹ 25.
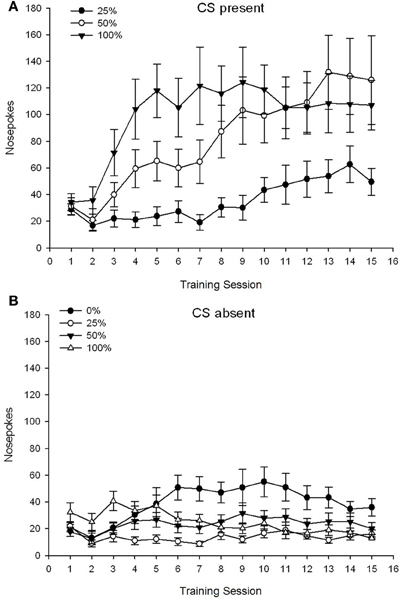
Figure 1. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Sprague Dawley rats (n = 8/ẹgbẹ) ti o farahan si ẹsan sucrose (ojutu 10%) ti a firanṣẹ labẹ 0, 25, 50, tabi 100% awọn iṣeto oniyipada. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (A) Scores when CS was present (5 s × 15 trials). (B) Scores when CS was absent (average for 5 × 15 s while light was off).
CS ko si. Nọmba 1B ṣe afihan awọn pokes imu ti o tumọ fun gbogbo awọn ẹgbẹ mẹrin fun iye akoko deede (awọn idanwo 5 s × 15) ni aropin ni akoko ti CS ko si. Ẹgbẹ 4 × 15 ANOVA ti pese awọn ipa akọkọ pataki ti Ẹgbẹ, F(3, 28) = 7.06, p = 0.001, ati Igba F(14, 392) = 2.84, p <0.001, pẹlu ibaraẹnisọrọ Ẹgbẹ pataki × Ibaṣepọ, F(42, 392) = 3.93, p <0.001. Ibaraṣepọ Ẹgbẹ pataki × Ibaṣepọ fun aṣa kuadiratiki, F(3, 28) = 3.91, p = 0.019, pẹlu ko si ibaraenisepo fun aṣa onigun, F(3, 28) <0.93, p > 0.44, ṣe afihan profaili “inverted-U” ti awọn imu imu lori awọn akoko ni ẹgbẹ 0, ni ilodi si profaili iduroṣinṣin gbogbogbo lori awọn akoko ni awọn ẹgbẹ miiran.
Ibugbe si awọn iyẹwu locomotor
ANOVA Ẹgbẹ 4 × 3 ni ipa akọkọ ti Ikoni, F(2, 56) = 5.67, p = 0.006, ko si si awọn ipa pataki miiran, F(3, 28) <1.60, p > 0.21. Itumọ (SE) awọn fifọ ina fun 2 h ni awọn apoti locomotor jẹ 1681 (123) ni igba 1, 1525 (140) lori igba 2, ati 1269 (96) lori igba 3. Awọn afiwera ti a gbero ko ri awọn iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ. 0 tabi ẹgbẹ 100 ni igba akọkọ tabi ipari ibugbe, t(84) <1.69, p > 0.05. Nitorinaa, ni laisi AMPH, ifihan leralera si awọn apoti idanwo ni nkan ṣe pẹlu idinku deede ni iṣẹ-ṣiṣe locomotor lẹẹkọkan ni awọn ẹgbẹ mẹrin (ie, ipa Ikoni), ati pe ko si idahun iyatọ bi iṣẹ ti iṣeto ikẹkọ sucrose (ko si ibaraenisepo) .
Awọn akoko idanwo
Awọn ipa ti ifamọ tẹlẹ 0.5 mg/kg ipenija AMPH.
Pre-abẹrẹ locomotion. Ẹgbẹ 4 kan ni ọna kan ANOVA ti idahun locomotor lakoko akoko ibugbe abẹrẹ ṣaaju-30-min ko ni awọn ipa pataki, F(3, 28) <1.05, p > 0.38. Awọn afiwera ti a gbero ko rii iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100, t(32) <0.87, p > 0.40. Nitoribẹẹ, awọn iyatọ ipilẹ ti o wa ni ipo iṣaju abẹrẹ ko ṣe akọọlẹ fun awọn iyatọ ẹgbẹ ni idahun locomotor si AMPH. Itumọ (SE) awọn fifọ ina fun apẹẹrẹ jẹ 559 (77).
Locomotion abẹrẹ lẹhin-abẹrẹ la igba ibugbe ti ko ni oogun ti o kẹhin. ANOVA Ẹgbẹ 4 × 2 ṣe afiwe awọn idahun locomotor awọn ẹgbẹ lori igba ibugbe ikẹhin, ati lẹsẹkẹsẹ lẹhin ifamọ ṣaaju 0.5 mg/kg AMPH ipenija. Awọn ikun fun igba ibugbe (120 min) ni iwọn lati ni ibamu pẹlu iye akoko akoko idanwo AMPH (90 min) (Dimegili ibugbe aise × 90/120). Onínọmbà naa ni ipa akọkọ pataki ti Ikoni, F(1, 28) = 34.16, p <0.001, ati pe ko si awọn ipa pataki miiran, F(3, 28) <2.26, p > 0.10. Ipa Ikoni ṣe afihan ilosoke ninu awọn fifọ ina (SE) ni idahun si iwọn lilo, lati 952 (72) si 1859 (151). Awọn afiwera ti a gbero ko rii awọn iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100 ni idahun si iwọn lilo, t(56) <1.72, p > 0.10. Sibẹsibẹ, aṣẹ ipo ti awọn ikun fifọ ina (M; SE) ni ibamu pẹlu arosọ: ẹgbẹ 50 (2205; 264)> ẹgbẹ 0 (2025; 203)> ẹgbẹ 100 (1909; 407)> ẹgbẹ 25 (1296; 299) .
Awọn ipa ti 1 mg / kg AMPH.
Pre-abẹrẹ locomotion. Ẹgbẹ 4 × 5 ANOVA ti idahun locomotor lakoko ipo ibugbe iṣaaju-abẹrẹ 30-min lori awọn akoko idanwo AMPH 1 mg/kg ti mu ipa akọkọ ti Ikoni, F(4, 112) = 43.64, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(3, 28) <0.97, p > 0.42. Awọn afiwera ti a gbero ko rii iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100 ni igba idanwo akọkọ tabi ikẹhin, t(140) <0.84, p > 0.30. Nitorina, awọn iyatọ ipilẹ ti o wa ni agbegbe ko ṣe iṣiro fun awọn iyatọ ẹgbẹ ni idahun locomotor si AMPH. Itumọ (SE) awọn ikun fifọ tan ina fun ipele iṣaju iwọn lilo ni awọn akoko 1–5 jẹ: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
Locomotion lẹhin-abẹrẹ. Nọmba 2 fihan awọn ipa ti awọn abẹrẹ marun ti 1 mg/kg AMPH (ọkan fun ọsẹ kan) lori awọn iṣiro iṣẹ ṣiṣe locomotor ni awọn ẹgbẹ mẹrin. ANOVA Ẹgbẹ 4 × 5 ni ipa akọkọ ti Ikoni, F(4, 112) = 8.21, p <0.001, ipa akọkọ ti ẹgbẹ, F(2, 45) = 3.28, p = 0.085, ko si si ibaraenisepo pataki, F(12, 122) <0.77, p > 0.68.
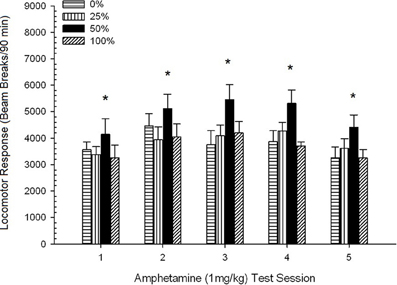
Figure 2. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
Awọn afiwera ti a gbero ṣe afihan pe awọn nọmba 50 ẹgbẹ yatọ si pataki si ẹgbẹ 0, t(14) = 2.19, p = 0.037, ati ẹgbẹ 100, t(14) = 2.36, p = 0.025 [ati pe o yatọ si diẹ si ẹgbẹ 25, t(14) = 2.03, p = 0.051]. Nitorinaa, ni ẹgbẹ 50, idahun locomotor si 1 mg/kg AMPH ni igbẹkẹle ju ti awọn ẹgbẹ mẹta miiran kọja gbogbo awọn akoko idanwo marun. Itupalẹ aṣa ilopọpo ṣe awari aṣa kuadiratiki pataki kan kọja awọn akoko, F(1, 28) = 32.47, p <0.0001, ko si si awọn aṣa pataki miiran, F(1, 28) <1.78, p > 0.19. Olusin 2 fihan pe abajade yii ṣe afihan apẹrẹ “iyipada U” kọja awọn akoko.
Iṣakoso fun iyatọ ninu idahun imu poke lakoko ikẹkọ sucrose
Atẹle ANCOVA ti awọn idahun locomotor si 1 miligiramu / kg AMPH, pẹlu awọn pokes imu (CS ti o wa) bi covariate, ninu awọn ẹgbẹ mẹta ti o gba CS, ti pese ipa akọkọ ti ẹgbẹ, F(2, 20) = 3.07, p = 0.069, ati pe ko si awọn ipa ti o ni ibatan iṣọpọ pataki, F(4, 80) <0.05, p > 0.85. Nitorinaa, ọna ti o dahun lakoko ikẹkọ ko ṣe alaye iyatọ pataki ninu idahun locomotor si 1 mg/kg AMPH ni awọn ẹgbẹ 25, 50, tabi 100.
Atẹle ANCOVA ti awọn idahun locomotor si 1 miligiramu/kg AMPH, pẹlu awọn pokes imu (CS aisi) bi iṣọpọ, ti mu ipa pataki ti iṣọpọ, F(1, 27) = 6.17, p = 0.020, ipa akọkọ pataki ti Ẹgbẹ, F(3, 27) = 4.13, p = 0.016, Ikoni abala kan × ibaraenisepo Ajọpọ, p = 0.080, ko si si awọn ipa pataki miiran, F(4, 108) <1.48, p > 0.21. Nitorinaa, ọna aibikita (aibikita) ti n dahun lakoko ikẹkọ ṣe alaye iyatọ pataki ni idahun locomotor si 1 mg/kg AMPH. Bibẹẹkọ, iyatọ yii kii ṣe agbekọja pẹlu iyatọ ti o ni ibatan ẹgbẹ, nitori ifisi ti covariate ninu itupalẹ pọ kuku ju dinku pataki ti ipa ẹgbẹ.
fanfa
Awọn data imu poke lakoko ti CS wa fihan pe awọn ẹgbẹ ti gba ajọṣepọ laarin CS ati ifijiṣẹ sucrose bi a ti ṣe afihan nipasẹ ilosoke ninu awọn idahun ti o da lori awọn akoko ikẹkọ. Profaili ti idahun lori awọn akoko lakoko ti CS wa ni iyanju pe awọn iṣeto 100 ati 50% CS-US jẹ doko gidi ni imunadoko ọna, lakoko ti iṣeto 25% mu alekun iwọntunwọnsi diẹ sii ni ọna idawọle. Awọn data imu imu nigba ti CS ko si daba pe awọn ẹgbẹ ti o gba eyikeyi ninu awọn eto ikẹkọ CS-sucrose mẹta (ẹgbẹ 25, 50, 100) kọ ẹkọ ni kiakia lati dinku awọn imu imu wọn ni isansa ti CS, lakoko ti awọn ẹranko ni ẹgbẹ 0 , eyiti ko gba CS, nikan kọ ẹkọ lati dinku ihuwasi isunmọ wọn si iwọn ti o lopin lẹhin ikẹkọ lọpọlọpọ.
Awọn data ibugbe fihan pe awọn ẹgbẹ ko yatọ ṣaaju AMPH ati pe ifihan leralera si awọn apoti idanwo ni nkan ṣe pẹlu idinku idahun locomotor laisi oogun. Nitoribẹẹ, awọn iyatọ laarin ẹgbẹ ati idahun ti o pọ si lori awọn iwọn lilo ti AMPH leralera ko le ṣe ikalara si awọn iyatọ iṣaaju-tẹlẹ ninu ihuwasi locomotor.
Awọn abajade ti ipenija ifamọ tẹlẹ pẹlu 0.5 mg/kg AMPH jẹrisi pe oogun naa pọ si iṣẹ ṣiṣe locomotor ni ibatan si ọjọ ibugbe laisi oogun ikẹhin. Ni ila pẹlu idawọle, ẹgbẹ 50 ni ipo ti o ga ju awọn ẹgbẹ 0 tabi 100 (bakannaa ẹgbẹ 25) ni awọn ọna ti idahun ti o tumọ si iwọn lilo, botilẹjẹpe awọn iyatọ iyatọ laarin awọn ẹgbẹ ko ṣe pataki.
Fun awọn akoko ifamọ, awọn afiwera ti awọn ẹgbẹ laarin awọn ẹgbẹ fihan pe ifihan ṣaaju si 50% ere sucrose ti o ni majemu yori si ilosoke pataki ni idahun locomotor si iwọn 1.0 mg/kg ti amphetamine ni ibatan si awọn iṣeto mẹta miiran. Ipa yii han gbangba lati iwọn lilo akọkọ ati pe ko yipada ni riri lori awọn iwọn lilo leralera. Onínọmbà aṣa tọkasi esi biphasic (fun apẹẹrẹ kikun) si awọn iwọn lilo ti AMPH leralera, jijẹ si iwọn lilo kẹta ati dinku lẹhinna. Awọn abajade ti atẹle ANCOVA pẹlu imu-pokes (CS asan) bi covariate ṣe idaniloju pe awọn iyatọ ninu awọn idahun locomotor awọn ẹgbẹ mẹrin si 1 miligiramu/kg AMPH ko ni ilaja nipasẹ ọna aibikita ti n dahun lakoko awọn akoko ikẹkọ sucrose.
Ipa ẹgbẹ lakoko awọn akoko ifamọ wa ni ibamu pẹlu idawọle wa. Ipa igba bi-phasic ko ni ibamu pẹlu ilọsiwaju ti a nireti ni awọn idahun locomotor pẹlu awọn iwọn AMPH ti o leralera. Eyi le jẹ ibatan si aarin iwọn lilo. Lati koju ọrọ yii, ilana kan (awọn iwọn lilo ojoojumọ miiran) ti o han lati fa ilọsiwaju deede ni idahun locomotor si awọn iwọn 1.0 mg/kg ti AMPH (ie, ifamọ ihuwasi) yẹ ki o lo. Ipa ti ilana ifamọ ti AMPH lori esi ti o tẹle si ipenija 0.5 mg/kg keji yoo ṣe atilẹyin siwaju si gbogbogbo ti ipa yii. Ifisi ipenija iyo saju si AMPH yoo pinnu ipa ti ireti tabi abẹrẹ ti o ni ibatan (fun apẹẹrẹ, aapọn) awọn ipa lori idahun locomotor si AMPH. Ifisi ti ẹgbẹ sucrose 75% kan yoo ṣe iranlọwọ lati ṣalaye ipa ti aidaniloju ere vs. awọn idahun isunmọ ti o ni ibatan si ipo agbegbe labẹ AMPH (lilo awọn pokes imu pẹlu CS ti o wa bi covariate), awọn pokes imu tun jẹ koodu fun ẹgbẹ 50 lakoko aarin nigba ti CS wa ni awọn ẹgbẹ mẹrin miiran (ie, ki imu pokes lati gbogbo marun). awọn ẹgbẹ-pẹlu ẹgbẹ 25 ti ko gba CS-le wa ninu igbekale ti iṣọkan pẹlu CS ti o wa bi iṣọkan). Awọn isọdọtun wọnyi ni a dapọ si ninu idanwo 0.
Ṣe idanwo 2
Awon nkan ise nkan ati awon ona lati se nkan
The methodology of experiment 2 was similar to that of experiment 1 but revised to better approximate a regimen found to reliably induce AMPH sensitization (Fletcher et al., 2005). Awọn ayipada jẹ bi atẹle: (a) Ẹgbẹ 75% CS-sucrose (n = 8) wa ninu; (b) Lakoko ikẹkọ sucrose, awọn eku (ayafi fun ẹgbẹ 0) gba awọn igbejade 20 CS (ina) (ni idakeji si 15 ni idanwo 1); (c) Awọn igbejade CS ni ọkọọkan yapa nipasẹ aropin aarin-idanwo ti 90 s; ibiti: 30-180 s (vs. 120 s ni adanwo 1), eyi ti o ṣe aiṣedeede ilosoke ninu awọn idanwo ikẹkọ lati dọgba iye akoko ikẹkọ kọọkan si ti idanwo 1; (d) iye akoko kọọkan ti awọn akoko ibugbe mẹta ti dinku lati 120 si 90 min lati ni ibamu pẹlu iye akoko awọn akoko idanwo; (e) Iyọ (ip, 1 milimita / kg) ipenija (90 min) ni a ṣafikun (ọjọ ikẹkọ lẹhin-sucrose 8), lati ṣe ayẹwo awọn ipa locomotor ti abẹrẹ fun kan (fun apẹẹrẹ, ireti, wahala); (f) Awọn akoko ifamọ 1 mg/kg ni a waye ni awọn ọjọ ọsẹ miiran (awọn ọjọ ikẹkọ lẹhin-lẹhin 12-21) dipo ni awọn aaye arin ọsẹ bi ni idanwo 1; (g) Paapọ pẹlu ifamọ tẹlẹ 0.5 mg / kg ipenija AMPH (ọjọ ikẹkọ lẹhin-ọjọ 9) ifamọ lẹhin keji 0.5 mg/kg ipenija AMPH (ọjọ ikẹkọ lẹhin-sucrose 28), lati ṣe idanwo gbogbogbo ti ipa ifamọ kọja awọn abere; (h) awọn pokes imu nigba ti CS wa ni koodu fun gbogbo awọn ẹgbẹ (pẹlu ẹgbẹ 0); (i) awọn pokes imu nigba ti CS ko si ni a gbasilẹ ni pato lati aarin 5-s lẹsẹkẹsẹ ṣaaju ibẹrẹ ti CS si atọka ọna ti o ti tọjọ.
awọn esi
Imu pokes nigba sucrose karabosipo igba
Ẹgbẹ 5 × 15 Ikoni × 2 Ipele (CS ti o wa, CS ko si) ANOVA ti awọn poki imu ti mu awọn ipa akọkọ pataki ti Ẹgbẹ, F(4, 19) = 2.89, p = 0.050, Igba F(14, 266) = 2.28, p = 0.006, ati Ipele, F(1, 19) = 14.72, p = 0.001, bakanna bi ibaraenisepo ọna mẹta, F(56, 266) = 1.38, p = 0.050. Awọn panẹli (A, B) ti Figure 3 Idite awọn ikun tumọ imu awọn ẹgbẹ fun CS ti o wa ati awọn ipele ti ko si CS, ni atele. Ifiwera ti awọn panẹli meji naa ṣafihan pe ipa akọkọ ti Ipele ṣe afihan awọn idahun imu poke gbogbogbo diẹ sii nigbati CS wa bayi la ko si. Nítorí náà, àwọn ìdáhùn tí a fọwọ́ kàn án ṣẹlẹ̀ lọ́pọ̀ ìgbà ju àwọn ìdáhùn tí a kò tíì tọ́jọ́ lọ. Awọn ipa akọkọ ti Ẹgbẹ ati Apejọ ko ni itumọ ni imurasilẹ nitori ibaraenisepo aṣẹ ti o ga julọ. Abajade igbehin yii ṣe afihan isọdọkan awọn ikun fun awọn ẹgbẹ marun ni ipele iduroṣinṣin kekere kan kọja awọn akoko nigbati CS ko si (Eyaworan 3B), papọ pẹlu iyatọ ti awọn ikun si giga (ẹgbẹ 75, ẹgbẹ 100), agbedemeji (ẹgbẹ 50), ati kekere (ẹgbẹ 0, ẹgbẹ 25) awọn ipele imu ti n dahun lori awọn akoko nigbati CS wa (Figure). 3A). Ninu awọn aṣa ilopọ pupọ ti aṣẹ kekere (laini, quadratic, cubic) nikan ibaraenisepo ọna mẹta fun aṣa laini sunmọ pataki, F(4, 19) = 2.32, p = 0.094, ti n ṣe afihan ilosoke monotonic gbogbogbo ni awọn pokes imu lori awọn akoko ni ẹgbẹ 75 ati isọdọtun iyara diẹ sii ni giga, agbedemeji, ati awọn ipele kekere ti idahun ni awọn ẹgbẹ miiran nigbati CS wa.
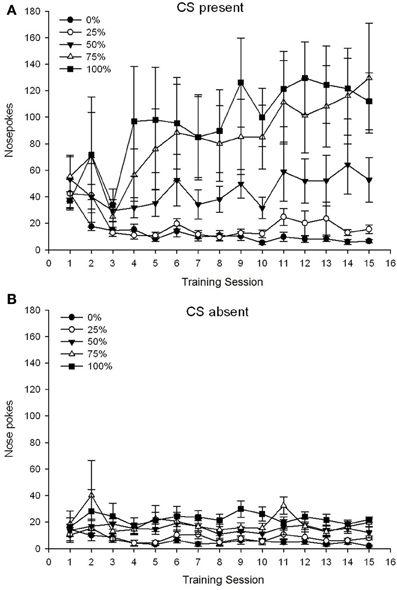
Figure 3. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Sprague Dawley rats (n = 8/ẹgbẹ) ti o farahan si ẹsan sucrose (ojutu 10%) ti a firanṣẹ labẹ 0, 25, 50, 75, tabi 100% awọn iṣeto oniyipada. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (A) Scores when CS was present (5 s × 20 trials). (B) Scores when CS was absent (average for 5 × 20 s while light was off).
Habituation to locomotor apoti
ANOVA Ẹgbẹ 5 × 3 ti awọn idahun locomotor ti ko ni oogun ti mu ipa akọkọ pataki kan ti Ikoni, F(2, 70) = 60.01, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <0.70, p > 0.60. Awọn afiwera ti a gbero ti ẹgbẹ 50 pẹlu ẹgbẹ 0 ati pẹlu ẹgbẹ 100 lori awọn akoko ibugbe akọkọ ati ikẹhin ko pese awọn ipa pataki, tawọn <0.84, p > 0.40. Nitorinaa, tumọ si idahun locomotor ti ko ni oogun ni awọn ẹgbẹ bọtini ko yatọ ṣaaju idanwo. Nọmba apapọ (SE) ti awọn fifọ ina fun iṣẹju 90 jẹ 2162 (118) lori igba 1, 1470 (116) lori igba 2, ati 1250 (98) ni igba 3.
Awọn akoko idanwo
Iyọ. Ẹgbẹ 5 × 2 ANOVA ṣe afiwe idahun locomotor lori igba ibugbe ipari ati igba ipenija iyo. ANOVA naa ni ipa akọkọ ti Ikoni, F(1, 35) = 62.46, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <0.65, p > 0.64. Olusin 4 Awọn igbero ẹgbẹ tumọ si ati fihan pe ipa Ikoni ṣe afihan idinku gbogbogbo ni esi locomotor lati igba ibugbe laisi oogun ti o kẹhin si igba iyọ, eyiti ko yatọ nipasẹ ẹgbẹ. Nitorinaa, idinku ninu idahun locomotor ti a rii lori awọn akoko ibugbe mẹta tẹsiwaju lori ifihan ti ko ni oogun kẹrin si awọn apoti idanwo.
Figure 4. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) on the last of 3 drug-free habituation sessions and on a subsequent session after saline injection (i.p., 1 ml/kg) in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
Awọn ipa ti 0.5 mg / kg AMPH.
Pre-abẹrẹ locomotion. Ẹgbẹ 5 × 2 ANOVA ti ipo iṣaju abẹrẹ (30-min) lori iṣaaju- ati lẹhin-ifamọ 0.5 mg/kg AMPH awọn ọjọ idanwo ni ipa akọkọ pataki ti Ikoni, F(1, 35) = 13.39, p = 0.001, ko si si awọn ipa pataki miiran, F(4, 35) <1.79, p > 0.15. Awọn afiwera ti a gbero ko rii awọn iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100 ni igba akọkọ, t(70) <1.00, p > 0.30. Bibẹẹkọ, lori ẹgbẹ igba keji (ifojusi lẹhin-ifiweranṣẹ) ẹgbẹ 50 (1203; 121) ṣe afihan ni pataki diẹ sii diẹ sii awọn fifọ tan ina abẹrẹ tẹlẹ (M; SE) ju ẹgbẹ 100 (756; 103) lọ, t(70) = 5.11, p <0.001, ṣugbọn ko yato si ẹgbẹ 0 (1126; 211), t(7) <0.88, p > 0.40. Nitoribẹẹ, awọn iyatọ ti ipilẹṣẹ ni agbegbe ko ṣe akọọlẹ fun awọn iyatọ ẹgbẹ ni idahun locomotor si iwọn 0.5 mg / kg akọkọ ti AMPH ṣugbọn o le ti ṣe alabapin si awọn iyatọ laarin ẹgbẹ 50 ati ẹgbẹ 100 ni idahun locomotor si iwọn 0.5 mg / kg keji ti AMPH . Itumọ (SE) awọn fifọ tan ina fun ipele iṣaaju-abẹrẹ lori akọkọ ati keji 0.5 mg/kg awọn akoko idanwo AMPH jẹ 757 (41) ati 974 (59).
Locomotion lẹhin-abẹrẹ. ANOVA Ẹgbẹ 5 × 2 ti idahun locomotor si 0.5 mg/kg AMPH ṣaaju ati lẹhin ilana ilana ifaramọ iwọn lilo 5 ti mu ipa akọkọ ti Ikoni, F(1, 35) = 76.05, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <1.10, p > 0.37. Olusin 5 ṣe afihan awọn ikun tumọ fun ẹgbẹ kọọkan ati igba.
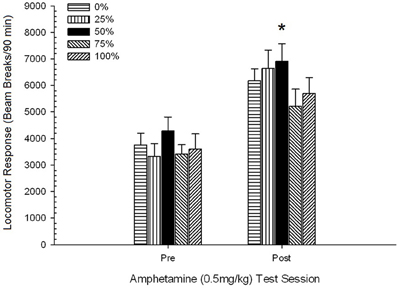
Figure 5. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 0.5 mg/kg d-amphetamine on separate sessions before and after a 5-session sensitizing regimen of d-amphetamine (1.0 mg/kg; i.p. per session) in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
Nọmba naa fihan pe ipa Ikoni kan pẹlu ilosoke pataki ni awọn ifasilẹ tan ina apapọ apapọ (SE) fun iṣẹju 90 lati iwọn 0.5 mg/kg iwọn 1, 3674 (216) si 0.5 mg/kg iwọn lilo 2, 6123 (275). Aini ibaraenisepo tabi ipa ẹgbẹ daba pe ifamọ si AMPH ko yatọ ni igbẹkẹle laarin awọn ẹgbẹ. Laibikita aini awọn ipa ti o ni ibatan ẹgbẹ pataki ni ANOVA, ayewo ti eeya naa ṣafihan pe ẹgbẹ 50 ṣe afihan idahun ti o tobi julọ si mejeeji akọkọ ati keji 0.5 mg/kg abere. Awọn afiwera ti a gbero ti idahun si iwọn lilo 0.5 mg/kg akọkọ ti fihan ko si iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100, t's(35) <0.48, p > 0.50. Bibẹẹkọ, ni idahun si keji (ifamọ lẹhin) 0.5 mg/kg iwọn lilo, ẹgbẹ 50 ṣe afihan ipo ti o tobi pupọ ju ẹgbẹ 0 lọ, t(35) = 2.00, p <0.05, bakanna bi ẹgbẹ 100, t(35) = 3.29, p <0.01.
Ni imọlẹ ti iyatọ ẹgbẹ pataki ni ipo abẹrẹ iṣaaju ni akoko 0.5 mg/kg AMPH keji ti o royin loke, atẹle 5 Group × 2 Session ANCOVA ti idahun locomotor si 0.5 mg/kg AMPH ni a ṣe, iṣakoso fun iṣaaju- locomotion abẹrẹ lori igba keji. Itupalẹ yii ṣe ipa pataki ti covariate, F(1, 34) = 8.65, p = 0.006, ipa akọkọ ti Ikoni F(1, 34) = 10.83, p = 0.002, ko si si awọn ipa pataki miiran, F(4, 34) <0.85, p > 0.50. Ni pataki, awọn afiwera ti a gbero ti o da lori aṣiṣe MS ati aṣiṣe df lati ANCOVA jẹrisi pe tumọ si idahun locomotor si iwọn 0.5 mg/kg keji ti AMPH wa ni pataki pupọ ni ẹgbẹ 50 ju ẹgbẹ 100 lọ, t(34) = 3.09, p <0.01, ati ẹgbẹ 0, t(34) = 1.88, p <0.05 (tailed-ọkan), nigbati iyatọ abẹrẹ iṣaaju lati igba 2 ti ṣakoso. Nitorinaa, ẹgbẹ 50 ṣe afihan idahun locomotor lẹhin ifarabalẹ ti o tobi pupọ si 0.5 mg/kg AMPH ju ẹgbẹ 100 tabi ẹgbẹ 0 lọ, ati pe awọn iyatọ ẹgbẹ wọnyi ko ni ilaja nipasẹ iṣaju iṣaju-abẹrẹ ni awọn ọjọ idanwo.
Awọn ipa ti 1.0 mg / kg AMPH.
Pre-abẹrẹ locomotion. ANOVA Ẹgbẹ 5 × 5 ti awọn ikun abẹrẹ iṣaaju-30-min fun awọn akoko ifamọ AMPH 1 miligiramu/kg ti pese ipa akọkọ ti Ikoni, F(4, 140) = 16.70, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <0.94, p > 0.45. Awọn afiwera ti a gbero ko rii iyatọ pataki ni ipo abẹrẹ iṣaaju laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100 ni igba akọkọ, t(175) <1.66, p > 0.10. Sibẹsibẹ, ni igba ikẹhin, ẹgbẹ 50 (1167; 140) ṣe afihan ni pataki diẹ sii awọn fifọ ina ina (M; SE) ju ẹgbẹ 100 (1000; 99) lọ, t(175) = 2.35, p <0.05, ṣugbọn ko yato si ẹgbẹ 0 (1085, 120), t(175) <1.16, p > 0.20. Nitorinaa, awọn iyatọ ninu ipo abẹrẹ iṣaaju ṣe alabapin si awọn iyatọ laarin awọn ẹgbẹ 50 ati 100 ni idahun locomotor si iwọn 1 mg/kg ipari AMPH. Itumọ (SE) awọn fifọ tan ina lapapọ fun apẹẹrẹ lakoko ipele abẹrẹ ṣaaju fun Awọn akoko 1 si 5 jẹ: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Locomotion lẹhin-abẹrẹ. Ẹgbẹ 5 × 5 Ikoni ANOVA ti awọn idahun si 1 mg/kg AMPH ni ipa akọkọ pataki ti Ikoni, F(4, 140) = 6.72, p <0.001, Ibaṣepọ Ẹgbẹ kan × Ibaṣepọ, F(16, 140) = 1.57, p = 0.085, ko si si ipa akọkọ ti Ẹgbẹ, F(4, 35) <0.44, p > 0.77. Awọn itupalẹ aṣa ilopọpo ṣe afihan aṣa laini pataki kan, F(1, 35) = 9.19, p = 0.005, ati aṣa onigun, F(1, 35) = 21.63, p <0.001, lori awọn akoko 1 nipasẹ 5. Ṣe nọmba 6 ṣe afihan awọn iṣiro locomotor tumọ fun ẹgbẹ kọọkan ati igba.
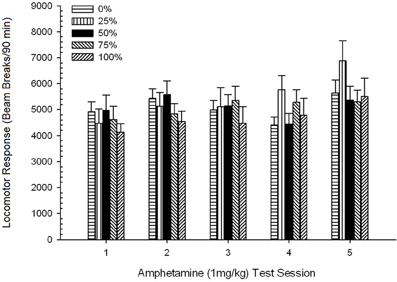
Figure 6. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Sprague Dawley rats (n = 8/ group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
Nọmba naa fihan pe ipa Ikoni ṣe afihan ilosoke pataki ni awọn ifasilẹ tan ina apapọ apapọ (SE) fun apẹẹrẹ ni kikun lati igba 1, 4624 (213) si igba 5, 5736 (272), ifẹsẹmulẹ ifarahan ti ifamọ si AMPH. Aṣa onigun tọkasi maxima ibatan lori awọn akoko 1, 3, ati 5, pẹlu awọn dips lori awọn akoko 2 ati 4, ni pataki fun awọn ẹgbẹ 0 ati 50. Nọmba naa tun ṣafihan pe, laibikita aini ibaraenisepo pataki, ẹgbẹ 25 ṣe afihan ni ilọsiwaju ni ilọsiwaju locomotor esi lori awọn akoko ati pe o yatọ pupọ si awọn ẹgbẹ miiran lori awọn akoko 4 ati 5 (9 ati 22% tobi ni atele, ju ẹgbẹ ti o ga julọ ti atẹle). Awọn afiwera ti a gbero rii pe ẹgbẹ 50 ko yatọ ni pataki lati awọn ẹgbẹ 0 tabi 100, t(175) <0.89, p > 0.40 lori akoko idanwo AMPH akọkọ tabi ipari 1 mg/kg.
Iṣakoso fun iyatọ ninu idahun imu poke lakoko ikẹkọ sucrose
Meji 5 Group × 2 Ikoni ANCOVAs ti idahun locomotor si 0.5 mg/kg AMPH ṣaaju ati lẹhin ilana ilana ifamọ, pẹlu lapapọ awọn pokes imu lakoko ikẹkọ sucrose pẹlu CS ti o wa ati pẹlu CS ti ko si bi awọn alabaṣiṣẹpọ lọtọ, ko rii awọn ipa pataki fun boya covariate, F(1, 18) <1.03, p > 0.31. Nitorina, ọna idahun lakoko ikẹkọ ko ṣe agbedemeji awọn iyatọ ẹgbẹ ni idahun si 0.5 mg / kg AMPH.
Meji 5 Group × 5 Ikoni ANCOVAs ti idahun locomotor si 1 mg/kg lakoko awọn akoko ifamọ pẹlu awọn pokes imu lapapọ (CS ti o wa, CS aisi) bi awọn akojọpọ lọtọ ko pese awọn ipa pataki ti covariate lakoko ti CS wa, F(4, 104) <1.04, p > 0.38, ati ipa akọkọ alapin ti covariate lakoko ti CS ko si, F(1, 18) = 3.32, p = 0.085.
fanfa
Awọn abajade iwadi yii ko ṣe atilẹyin igbagbogbo pe ẹgbẹ 50 yoo ṣe afihan idahun locomotor ti o ga julọ lori awọn akoko ti a fiwe si awọn ẹgbẹ miiran. Awọn data AMPH miligiramu/kg jẹri ifarahan ti ifamọ pẹlu ilana iwọn lilo ọjọ-keji. Ilana ti o wa laarin awọn ẹgbẹ ṣe afihan aṣa kan fun ifarabalẹ ti o pọju lakoko awọn akoko ikẹhin ni ẹgbẹ 1, laisi iru ẹri fun ẹgbẹ 25. Ni idakeji, awọn abajade iwọn lilo 50 mg / kg ṣe afihan aṣa kan fun ifarabalẹ nla ni ẹgbẹ 0.5, lakoko kanna. akoko ifẹsẹmulẹ ilosoke gbogbogbo pataki ni idahun locomotor kọja awọn ẹgbẹ si keji la akọkọ 50 mg/kg iwọn lilo AMPH. Ipa asan ti abẹrẹ iyọ jẹri pe ireti tabi aapọn ti o ni ibatan abẹrẹ ko ṣe alabapin si awọn ipa AMPH.
The nose poke data again revealed an overall increase in approach responding over the course of training sessions when the CS was present, with no corresponding increase when the CS was absent. Therefore, the animals appeared to acquire the association between the CS and the prospect of sucrose reward. Group differences in the frequency of nose pokes when the CS was present conformed roughly to the frequency of reward delivery under the respective schedules, with groups 75 and 100 displaying the most nose pokes, group 50 displaying intermediate numbers of nose pokes, and groups 0 and 25 displaying the fewest nose pokes. These results suggest that the CS came to control approach responding in a manner consistent with the overall probability of reward. Although speculative, one possible explanation for the lower nose poke rates with CS present in group 50 in experiment 2 vs. experiment 1 may be the shortening of the inter-trial interval, as longer inter-trial intervals (experiment 1) appear to encourage impulsive tendencies and this is associated with increased turnover of DA in anterior cingulate, prelimbic and infralimbic cortices (Dalley et al., 2002). Nitorinaa, idinku 30% ni aarin-idanwo laarin idanwo 2 (ati 3) le ti yipada awọn ipele DA ti cortical ati igbega yiyan diẹ sii (ie, itọsọna nipasẹ igbohunsafẹfẹ ibatan ti ẹsan) vs. impulsive (kii ṣe itọsọna nipasẹ igbohunsafẹfẹ ere) ọna fesi ni ẹgbẹ 50 lakoko awọn idanwo ikẹkọ ni idanwo 2 bi akawe pẹlu idanwo 1.
Aini awọn ipa ti o ni ibatan idapọmọra pataki fun awọn pokes imu ni ipo CS ti o wa ninu awọn ANCOVAs tọkasi pe isunmọ idahun lakoko ikẹkọ sucrose ko ṣe agbedemeji awọn ipa ti awọn iṣeto CS-sucrose oriṣiriṣi lori awọn idahun si AMPH. Ipa pataki pataki ti covariate fun ipo isansa CS ni ANCOVA ti awọn idahun locomotor si 1 mg/kg AMPH ni imọran pe ifarahan si idahun ti ko ni oogun ti o ti tọjọ ṣe alaye diẹ ninu awọn iyipada ninu awọn ipa locomotor ti AMPH lakoko awọn akoko ifamọ.
Papọ, ẹri naa ni imọran pe awọn ipa ti itan-itumọ le jẹ akiyesi diẹ sii pẹlu 0.5 AMPH ju pẹlu 1 mg/kg AMPH, ati pe ilana kan ti o ṣe agbejade ifamọ ni laisi ifọwọyi eyikeyi le ṣe ṣoki tabi ṣe laiṣe awọn ipa ti putative ifamọ-igbega ifọwọyi ihuwasi (ie, ere oniyipada onibaje).
Behavioral sensitization to AMPH is a robust effect in the laboratory. However, outside the laboratory, only a minority of individuals who gamble chronically escalate to pathological levels. Although risk for sensitization is related to risk for addiction (or drug seeking), especially for psychostimulants (Vezina, 2004; Flagel et al., 2008), many factors aside from sensitization risk may predispose one to addiction (e.g., Verdejo-Garcia et al., 2008; Conversano et al., 2012; Volkow et al., 2012). Bibẹẹkọ, awọn ifosiwewe abuda ti o funni ni ailagbara si ifamọ le ṣe ajọṣepọ pẹlu itan-itumọ lati tẹnuba awọn ipa ti ẹsan aisọtẹlẹ (ie, 50% iṣeto CS-US) lori imuṣiṣẹda eto DA. Lati ṣe iwadii iṣeeṣe yii, idanwo 3 lo ilana kanna bi idanwo 2 ṣugbọn o lo igara Lewis dipo awọn eku igara Sprague Dawley.
Sprague Dawley rats display intermediate levels of DA transporters, with lower levels than Wistar strain rats (Zamudio et al., 2005), but higher levels than Wistar-Kyoto rats (a “depressive”-like strain) in the nucleus accumbens, amygdala, ventral tegmental area and substantia nigra (Jiao et al., 2003). This profile may render Sprague Dawley rats only moderately sensitive to environmental or pharmacological manipulations of DA function. In contrast, Lewis rats exhibit low levels of DA transporters as well as D2 and D3 DA receptors in the nucleus accumbens and dorsal striatum compared to other strains (e.g., F344) (Flores ati al., Ọdun 1998). These morphological differences may contribute to Lewis rats’ differential response to DA manipulations. Lewis rats also exhibit a range of accentuated responses to experimental drug manipulations compared to other strains (e.g., F344). Most importantly, Lewis rats display greater sensitization to methamphetamine, characterized by low response to initial doses but higher response to later doses (Camp et al., 1994). Lewis rats also exhibit greater locomotor sensitization to a range of doses of cocaine (Kosten et al., 1994; Haile et al., 2001). Da lori ilana ti awọn ipa yii, a ro pe awọn eku Lewis yoo jẹ ki a ṣe iwadii boya ifaragba si ifamọ n pọ si awọn ipa ti iṣeto kondisona lori esi ti o tẹle si AMPH.
Ṣe idanwo 3
Awon nkan ise nkan ati awon ona lati se nkan
Ilana naa jẹ kanna gẹgẹbi ni idanwo 2, yato si lilo awọn eku Lewis (200-225 g lori dide, Charles River, Quebec, Canada).
awọn esi
Imu pokes nigba sucrose karabosipo igba
Ẹgbẹ 5 × 15 Ikoni × 2 Ipele (CS ti o wa, CS ko si) ANOVA ti awọn poki imu ti mu awọn ipa akọkọ pataki ti Ẹgbẹ, F(4, 34) = 6.12, p = 0.001, Igba, F(14, 476) = 3.42, p <0.001, ati Ipele, F(1, 34) = 20.83, p <0.001, bakanna bi ibaraenisepo ọna mẹta, F(56, 476) = 1.56, p = 0.008. Awọn panẹli (A, B) ti Figure 7 Idite awọn ikun tumọ imu awọn ẹgbẹ fun CS ti o wa ati awọn ipele ti ko si CS, ni atele. Ifiwera ti awọn panẹli meji naa ṣafihan pe ipa akọkọ ti Ipele ṣe afihan awọn idahun imu poke gbogbogbo diẹ sii nigbati CS wa bayi la ko si. Nitorinaa, awọn idahun cued waye ni pataki diẹ sii nigbagbogbo ju awọn idahun ti ogbo ṣaaju lọ. Awọn ipa akọkọ ti Ẹgbẹ ati Apejọ ko ni itumọ ni imurasilẹ nitori ibaraenisepo aṣẹ ti o ga julọ. Ibaraṣepọ ọna mẹta ṣe afihan isọdọkan ti awọn ikun fun awọn ẹgbẹ marun ni ipele iduroṣinṣin kekere ti o jo kọja awọn akoko nigbati CS ko si [Igbimọ (B)], papọ pẹlu iyatọ ti awọn ikun nigbati CS wa sinu awọn profaili ti o ni iyasọtọ fun ẹgbẹ kọọkan ti o jọra ilana ipo ipo wọn ti igbohunsafẹfẹ ere: lati ga julọ (ẹgbẹ 100) si ti o kere julọ (ẹgbẹ 25) [Panel] (A)]. Ilana laini nikan fun ibaraenisepo jẹ pataki, F(4, 34) = 4.03, p = 0.009, ti n ṣe afihan ilosoke deede ni gbogbo igba ni awọn pokes imu lori awọn akoko ni ẹgbẹ 100 nigbati CS wa bi o lodi si profaili aisedede ti o jo ti ilosoke ninu awọn pokes imu kọja awọn akoko ni awọn ẹgbẹ miiran lakoko ipele yii.
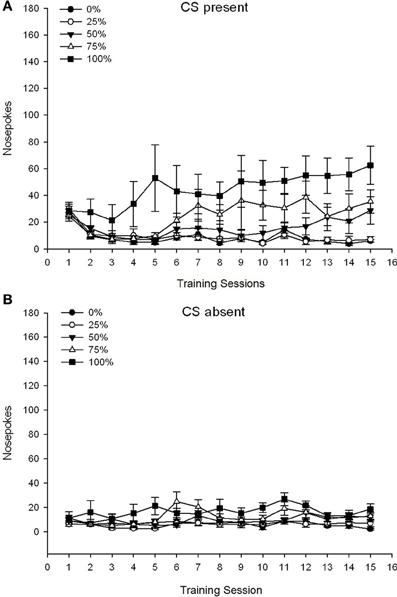
Figure 7. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Lewis rats (n = 8/ẹgbẹ) ti o farahan si ẹsan sucrose (ojutu 10%) ti a firanṣẹ labẹ 0, 25, 50, 75, tabi 100% awọn iṣeto oniyipada. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (A) Scores when CS was present (5 s × 20 trials). (B) Scores when CS was absent (average for 5 × 20 s while light was off).
Habituation to locomotor apoti
ANOVA Ẹgbẹ 5 × 3 ni ipa akọkọ ti Ikoni, F(2, 70) = 23.07, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(8, 70) <1.47, p > 0.18. Apẹrẹ curvilinear ti awọn ikun locomotor (SE) farahan lati igba 1, 1076 (74), nipasẹ igba 2, 644 (48), si igba 3, 762 (59). Awọn afiwera ti a gbero ti ẹgbẹ 50 pẹlu ẹgbẹ 0 ati pẹlu ẹgbẹ 100 lori awọn akoko ibugbe akọkọ ati ikẹhin ṣafihan awọn isinmi ina kekere ti o dinku ni ẹgbẹ 50 (M = 911; SE = 109) la ẹgbẹ 0 (M = 1103; SE = 176) lori igba ibugbe 1, t(105) = 2.02, p <0.05, ṣugbọn ko si iyatọ laarin ẹgbẹ 50 ati ẹgbẹ 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, lori igba yii. Ẹgbẹ 50 ko yatọ ni pataki lati boya ẹgbẹ 0 tabi ẹgbẹ 100 lori igba ibugbe ipari, t(105) <0.93, p > 0.30. Nitorinaa, tumọ si idahun locomotor ti ko ni oogun ni awọn ẹgbẹ bọtini ko yatọ nigbagbogbo ṣaaju idanwo.
Awọn akoko idanwo
Iyọ. Ẹgbẹ 5 × 2 ANOVA ti awọn idahun locomotor lori igba ibugbe ikẹhin ati igba idanwo iyo ni ipa akọkọ pataki ti Ikoni, F(1, 35) = 50.12, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <0.57, p > 0.68. Olusin 8 fihan ẹgbẹ tumọ si awọn ikun fun awọn akoko meji ati tọka pe ipa Ikoni ṣe afihan idinku nla lati ibugbe si idanwo iyọ. Bayi, gbigba ti awọn abẹrẹ fun kan (fun apẹẹrẹ, ireti, aapọn) ko mu idahun locomotor pọ si.
Figure 8. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) on the last of 3 drug-free habituation sessions and on a subsequent session after saline injection (i.p., 1 ml/kg) in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
Awọn ipa ti 0.5 mg / kg AMPH.
Pre-abẹrẹ locomotion. Ẹgbẹ 5 × 2 Ikoni ANOVA ti ipo abẹrẹ iṣaju iṣaju ti pese ipa akọkọ pataki ti Ikoni, F(1, 35) = 15.04, p <0.001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <1.19, p > 0.33. Awọn afiwera ti a gbero ko rii iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100 lori boya igba idanwo, t(70) <0.99, p > 0.30. Nitorina, awọn iyatọ ti ipilẹṣẹ ni ipo iṣaju-abẹrẹ ko ṣe iṣiro fun awọn iyatọ ẹgbẹ ni idahun locomotor si 0.5 mg / kg AMPH. Itumọ (SE) awọn fifọ tan ina fun ipele iṣaaju-abẹrẹ fun akọkọ ati keji (ifojusi lẹhin-ifiweranṣẹ) awọn akoko 0.5 mg/kg jẹ 325 (25) ati 473 (36).
Locomotion lẹhin-abẹrẹ. ANOVA Ẹgbẹ 5 × 2 ti idahun locomotor si awọn abere 0.5 mg/kg ti a firanṣẹ ṣaaju ati lẹhin onibaje 1 mg/kg AMPH ni ipa akọkọ ti Ikoni, F(1, 34) = 87.44, p <0.0001, ati pe ko si awọn ipa pataki miiran, F(4, 34) <0.94, p > 0.45. Olusin 9 n gbero awọn iṣiro locomotor tumọ fun ẹgbẹ kọọkan ati igba ati fihan pe ipa Ikoni ṣe afihan esi gbogbogbo ti o pọ si si iwọn lilo 0.5 mg/kg keji, ni ibamu pẹlu ifamọ. Nọmba naa tun fihan pe awọn ẹgbẹ ṣe bakanna ni igba 1, ṣugbọn ẹgbẹ 50 naa ṣe afihan iṣẹ-ṣiṣe locomotor diẹ sii ju awọn ẹgbẹ miiran lọ ni igba 2. Awọn afiwera ti a gbero ni idahun si akọkọ 0.5 mg / kg iwọn lilo ti fihan ko si iyatọ pataki laarin ẹgbẹ 50 ati ẹgbẹ 0 tabi ẹgbẹ 100, t(35) <1.28, p > 0.20. Sibẹsibẹ, ẹgbẹ 50 ṣe afihan idahun locomotor ti o tobi pupọ si iwọn 0.5 mg/kg keji ju ti ẹgbẹ 0 lọ, t(35) = 4.32, p <0.001, tabi ẹgbẹ 100, t(35) = 2.24, p <0.05.
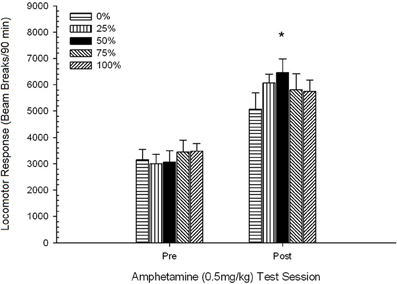
Figure 9. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 0.5 mg/kg d-amphetamine on separate sessions before and after a 5-session sensitizing regimen of d-amphetamine (1.0 mg/kg; i.p. per session) in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
Awọn ipa ti 1 mg / kg AMPH.
Pre-abẹrẹ locomotion. Ẹgbẹ 5 × 5 ANOVA ti awọn ikun abẹrẹ iṣaaju-30-min fun awọn akoko ifamọ ti mu ipa akọkọ ti Ikoni, F(4, 140) = 4.10, p = 0.004, ko si si awọn ipa pataki miiran, F(4, 35) = 1.25, p > 0.31. Awọn afiwera ti a gbero rii pe awọn fifọ tan ina lakoko ipele abẹrẹ iṣaaju (M; SE) kere pupọ ni ẹgbẹ 50 (395; 62) ju ni ẹgbẹ 100 (508; 62), t(175) = 2.58, p <0.01, ṣugbọn kii ṣe ẹgbẹ 0, t(175) <1.83, p > 0.10, lori 1 mg/kg AMPH igba 1. Ni ipari 1 mg/kg AMPH akoko, awọn afiwera ti a ti pinnu tun ri pe iṣaju abẹrẹ ni ẹgbẹ 50 (378; 60) jẹ pataki ti o kere ju ni ẹgbẹ 100 (650; 75) ), t(175) = 6.17, p <0.001, ṣugbọn kii ṣe ni ẹgbẹ 0, t(175) <1.84, p > 0.10. Gẹgẹbi itọsọna ti awọn iyatọ ẹgbẹ wọnyi (ẹgbẹ iṣakoso = ẹgbẹ 50) jẹ idakeji si apẹrẹ ti a ti sọ tẹlẹ, awọn iyatọ ẹgbẹ ni ipo abẹrẹ lẹhin-abẹrẹ ti o ni ibamu pẹlu iṣeduro ko le ṣe iyasọtọ si awọn iyatọ ipilẹ ti iṣaju-abẹrẹ. Itumọ (SE) awọn fifọ tan ina lapapọ lakoko ipele abẹrẹ ṣaaju fun Awọn akoko 1 si 5 jẹ: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Locomotion lẹhin-abẹrẹ. Ẹgbẹ 5 × 5 Awọn akoko ANOVA ti awọn idahun si awọn abere 1 mg/kg ti pese ipa akọkọ pataki ti Ikoni, F(4, 140) = 6.15, p <0.001, ati pe ko si awọn ipa pataki miiran, F(4, 35) <0.57, p > 0.68. Awọn itupalẹ aṣa ilopọpo ṣe afihan aṣa laini pataki kan, F(1, 35) = 9.34, p = 0.004, ati aṣa onigun, F(1, 35) = 5.08, p = 0.031, abajade igbehin ti n tọka si maxima ibatan lori awọn akoko 3 ati 5. Ṣe nọmba 10 Idite awọn ikun wọnyi ati fihan pe, laibikita aini ibaraenisepo pataki ninu ANOVA, ẹgbẹ 50 ṣe afihan ipo nla ti o tobi ju awọn ẹgbẹ mẹrin miiran lọ ni idahun si iwọn lilo 1 mg/kg ikẹhin. Nitorinaa, awọn afiwera ti a gbero ṣe afihan awọn ikun ti o ga julọ ni pataki lori igba 5 ni ẹgbẹ 50 ju gbogbo awọn ẹgbẹ miiran lọ, t(35) > 3.68, p <0.001.
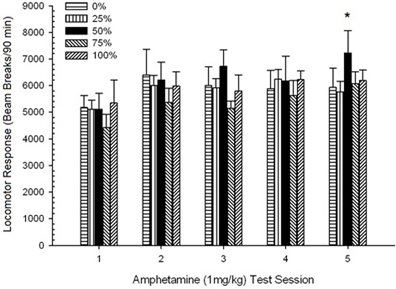
Figure 10. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
Iṣakoso fun iyatọ ninu idahun imu poke lakoko ikẹkọ sucrose
Meji 5 Group × 2 Ikoni ANCOVAs ti idahun locomotor si 0.5 mg/kg AMPH ṣaaju ati lẹhin ilana ilana ifamọ, pẹlu lapapọ awọn pokes imu lakoko ikẹkọ sucrose pẹlu CS ti o wa ati pẹlu CS ti ko si bi awọn alabaṣiṣẹpọ lọtọ, ko rii awọn ipa pataki fun boya covariate, F(1, 32) <0.44 p > 0.51. Meji 5 Group × 5 Ikoni ANCOVAs ti idahun locomotor si 1 mg/kg AMPH lakoko awọn akoko ifamọ pẹlu awọn pokes imu lapapọ (CS lọwọlọwọ, CS aisi) bi awọn akojọpọ lọtọ ko pese awọn ipa pataki ti covariate lakoko ti CS wa tabi ko si, F(1, 33) <0.14, p > 0.71. Nitorinaa, ọna idahun laisi oogun ko ṣe akọọlẹ fun awọn iyatọ ẹgbẹ ninu awọn idahun locomotor si boya iwọn lilo AMPH.
fanfa
Sensitization developed to the effects of repeated 1.0 mg/kg amphetamine. The habituation and saline data confirm that this effect was not due to pre-existing differences, expectancy, or stress-related responses to the injection. The ANCOVAs with nose pokes confirm that these effects were not due to drug-free approach behavior. The nose poke data themselves indicated that the groups acquired the association between the CS and prospect of sucrose reward. The groups’ rank level of nose-poke responding at the end of training matched the overall frequency of reward under the different schedules from highest (group 100) to lowest (group 0), as it did in experiment 2. The relatively lower overall mean nose poke levels in this experiment compared to experiments 1 and 2 may reflect more selective approach responding to cues for reward in Lewis rats (Kosten et al., 2007).
Awọn data iwọn lilo 0.5 mg/kg fihan pe idahun locomotor akọkọ si AMPH ni awọn eku Lewis (Eyaworan 9) ni itumo timole akawe si Sprague Dawley eku (ṣàdánwò 2; Figure 5), ṣugbọn ilosoke laarin ẹgbẹ ni idahun si iwọn lilo keji ni awọn eku Lewis jẹ akude (o fẹrẹ ilọpo meji idahun si iwọn lilo 0.5 mg/kg akọkọ) ni atẹle ilana 5-akoko AMPH Ni pataki julọ, ẹgbẹ 50 ṣe afihan idahun locomotor nla kan. ju gbogbo awọn ẹgbẹ ayafi ẹgbẹ 25 si keji (ie, ifamọ lẹhin-ifiweranṣẹ) 0.5 mg/kg iwọn AMPH ati idahun locomotor ti o tobi ju gbogbo awọn ẹgbẹ miiran lọ, pẹlu ẹgbẹ 25, si iwọn 1 mg/kg ipari AMPH (akoko ifamọ ipari) .
Itupalẹ akopọ ti awọn ipo ẹgbẹ kọja awọn adanwo
To determine the reliability of group differences in sensitization, a non-parametric analysis assessed the contingency between group and rank of mean locomotor response to the second (post-chronic AMPH) 0.5 mg/kg dose and the final 1.0 mg/kg dose of AMPH from the 3 experiments. The analysis yielded a significant effect, φ = 0.986, p = 0.025, ti n ṣe afihan otitọ pe ẹgbẹ 50 ni ipo akọkọ ni gbogbo ṣugbọn ọkan ninu awọn afiwera. Ipo ti o ga julọ ti ẹgbẹ 50 ni akawe si gbogbo awọn ẹgbẹ miiran ni idahun si keji (AMPH lẹhin-chronic) iwọn 0.5 mg/kg ni a fihan ni Nọmba 5 (ayẹwo 2) ati Figure 9 (àdánwò 3). Ipo ti o ga julọ ti ẹgbẹ 50 ni ibatan si awọn ẹgbẹ miiran ni idahun si iwọn lilo 1.0 mg/kg ti o kẹhin jẹ afihan ni Nọmba 2 (ayẹwo 1) ati Figure 10 (àdánwò 3). Iyatọ kan si apẹrẹ yii ni idahun si iwọn lilo 1.0 mg/kg ikẹhin ni awọn eku Sprague-Dawley ni idanwo 2.
Gbogbogbo ijiroro
The present series of experiments tested the hypothesis that chronic exposure to a gambling-like schedule of reward can sensitize brain DA pathways much like chronic exposure to drugs of abuse. Evidence for such an effect would suggest that neuroplasticity, of the same kind thought to contribute to drug addiction, can be induced by chronic exposure to unpredictable reward schedules. In line with the literature on drug addiction, locomotor response to 0.5 and 1.0 mg/kg doses of AMPH indexed DA system reactivity, with greater locomotion in response to later doses operationally defining sensitization (cf. Robinson ati Berridge, 1993; Pierce ati Kalivas, 1997; Vanderschuren ati Kalivas, 2000).
Lapapọ, awọn abajade wa ni ila pẹlu idawọle wa. Bibẹẹkọ, wọn tun tọkasi iyipada akude ninu awọn ipa idanwo nitori awọn ifosiwewe ilana. Awọn ipa ti iṣeto iṣeto jẹ iwọntunwọnsi ṣugbọn ni ibamu, pẹlu ẹgbẹ 50 ti n ṣe afihan esi ti o tobi ju awọn ẹgbẹ mẹrin miiran lọ si awọn abere mejeeji ti o tẹle awọn ilana iwọn lilo marun. Biotilejepe ìwò F-awọn iye fun awọn ipa ti o ni ibatan ẹgbẹ ninu awọn itupale iyatọ nigbagbogbo kii ṣe pataki, awọn iyatọ ẹgbẹ bọtini ni a timo pẹlu awọn afiwera ti a gbero ni ọna meji. Ni ọran yii o yẹ ki o ṣe akiyesi pe, “Ironu lọwọlọwọ, sibẹsibẹ, ni pataki lapapọ [fun F ni ANOVA] ko wulo. Ni akọkọ, awọn idawọle ti idanwo nipasẹ idanwo gbogbogbo ati idanwo afiwera pupọ yatọ pupọ, pẹlu awọn ipele agbara ti o yatọ pupọ. Fun apẹẹrẹ, apapọ F kosi pin awọn iyatọ laarin awọn ẹgbẹ kọja nọmba awọn iwọn ti ominira fun awọn ẹgbẹ. Eyi ni ipa ti diluting apapọ F in the situation where several group means are equal to each other but different from some other mean” (Howell, 1992, p. 338). Eyi ni deede ipo ti o lo ninu awọn adanwo lọwọlọwọ, nibiti a ti nireti ẹgbẹ 50 lati yatọ si ẹgbẹ 0 ati awọn iṣakoso ẹgbẹ 100 ṣugbọn ko si iyatọ laarin awọn ẹgbẹ iṣakoso wọnyi ti asọtẹlẹ fun ẹgbẹ 25 tabi ẹgbẹ 75.
Awọn data poke imu timo pe, ni gbogbo idanwo, awọn ẹranko gba ajọṣepọ laarin CS ati ireti ti ẹsan sucrose. Ifiweranṣẹ laarin igbohunsafẹfẹ poke imu fun awọn ẹgbẹ oriṣiriṣi ati igbohunsafẹfẹ gbogbogbo ti ẹsan labẹ awọn iṣeto ikẹkọ oniwun wọn ni imọran pe iwọn apapọ ti ẹsan sucrose ṣe itọsọna idahun ti ko ni oogun. Sibẹsibẹ, aisi ipa ilaja ti awọn imu imu lori awọn idahun locomotor ti o ni ibatan si AMPH ni awọn ANCOVA fihan pe awọn ilana lọtọ ni o wa labẹ awọn ihuwasi meji.
Ni awọn igba miiran, ipa ti iṣeto iṣeto ni o han ni idahun si iwọn lilo AMPH akọkọ; ni awọn igba miiran o farahan nikan lẹhin awọn iwọn lilo leralera. Awọn iyatọ ẹgbẹ ni idahun locomotor si iwọn lilo AMPH akọkọ daba pe ifihan si awọn eto ere bii ere jẹ to funrararẹ lati fa ifamọ. Awọn iyatọ ti ẹgbẹ ni ipo ti o tẹle awọn iwọn AMPH lọpọlọpọ tọkasi ipa arekereke diẹ sii ti o le ṣe afihan bi “ailagbara,” eyiti o farahan nikan nigbati a ba papọ pẹlu ifihan ti nlọ lọwọ si aṣoju ifarabalẹ akọkọ (ie, amphetamine).
Differences in the pattern of response across experiments suggest that a longer interval between training and initial AMPH challenge may maximize the opportunity to detect the inherent sensitizing effect of the conditioning treatment. This in turn suggests that effects of conditioned reward exposure may incubate over time, a phenomenon also seen with stimulant sensitization (Grimm et al., 2006). Ilana ti idahun si awọn abere meji ti amphetamine ni imọran pe iwọn lilo 0.5 mg/kg le jẹ imunadoko diẹ sii ni fifihan awọn ipa ti itan-itumọ. Eyi ni imọran ni imọran pe awọn ipa imudara labẹ ilana ikẹkọ lọwọlọwọ jẹ abele diẹ ati pe o le jẹ camouflaged nipasẹ awọn ipa aja labẹ awọn iwọn AMPH ati awọn ipo ti o ṣe ipilẹṣẹ. de novo ijẹrisi.
In experiment 3, the biphasic pattern of response to the 0.5 mg/kg doses and progressive emergence of superiority in group 50 is consistent with the expected profile for Lewis rats in response to methamphetamine (Camp et al., 1994). Eyi ṣe atilẹyin atilẹyin si iwulo ti awọn awari lọwọlọwọ ati daba ni lqkan laarin awọn ifosiwewe ti o jẹ ailagbara iwọntunwọnsi si ifamọ psychostimulant ati si awọn iṣeto ere bii ere.
Across experiments, the post-sensitization locomotor response of group 50 generally exceeded that of the other groups under different doses of amphetamine and in different strains of animals. However, the high within-group variability and modest between-group effect sizes indicate a role for other factors in DA system reactivity to amphetamine following exposure to varying schedules of conditioned sucrose reward. Although responses of DA neurons to reward signals may provide a coarse model of gambling (Fiorillo et al., 2003), bii gbogbo awọn awoṣe, isonu ti alaye wa fun idi ti parsimony — ie, lati ṣafihan ilana bọtini kan. Bi abajade, apẹẹrẹ ti awọn ipa kọja awọn ipo CS-US ni atilẹba Fiorillo et al. iwadi ko ni kikun ni kikun si idahun locomotor si amphetamine. Siwaju awọn isọdọtun ti awoṣe ni a pe fun ni kikun awọn aaye ti ayokele ti o ni ipa lori iṣẹ eto DA.
Taken together, the results of this series of experiments provide provisional support for the hypothesis that chronic exposure to gambling-like schedules of reward enhances the reactivity of the brain DA system to psychostimulant challenge. As such, they extend the findings of Singer et al. (2012) who demonstrated that, relative to a fixed schedule, prior exposure to a variable reinforcement schedule in an operant paradigm enhances subsequent locomotor response to amphetamine. More specifically, the present findings point to uncertainty of reward delivery as the critical factor underlying the effects of variable reward. The magnitude of effects in the operant paradigm was substantially greater than the effects found in the present experiments. This may reflect greater chronic exposure to the gambling-like activity (55 vs. 15 days); it may reflect the effects of requiring an operant response to elicit the reward (i.e., a role for agency) rather than passive exposure, as in the present study. Increasing the duration of training in the present paradigm would help to resolve these questions.
The validity of variable reward and reinforcement schedules as models of gambling cannot be gleaned from these experiments. Future research that examines the impact of conditioning history on risk-taking behavior in rodent gambling tasks could address this issue. Similarly, the correspondence between the behavioral sensitization found here and the elevated striatal DA response to amphetamine recently found in pathological gamblers must await further investigation (Boileau et al., 2013). Micro-dialysis le koju ibeere yii, ati pe asọtẹlẹ ti o da lori data eniyan yoo jẹ pe itusilẹ DA ti o tobi julọ ninu ẹgbẹ 50 “phenotype ere” yoo ṣe akiyesi ni gbangba ni dorsal (sensorimotor) striatum kuku ju ventral (limbic) striatum . Ifọwọsi ti 50% oniyipada CS + ifihan ere ni awọn paragile miiran yoo ṣe atilẹyin ohun elo rẹ bi awoṣe esiperimenta ododo ti PG.
Whereas some forms of gambling clearly entail an instrumental response (e.g., slot machines), in other forms of gambling (e.g., lottery) the link between the action (purchasing the ticket, i.e., placing the bet), the cues for reward (i.e., lottery numbers) and the reward itself (the winning number and monetary payoff) is much more diffuse. Nevertheless, activation of DA during the CS-US interval may well occur. This may explain why, when the “winning number” is announced, attention is riveted as each individual lottery ball drops in succession to compose the specific sequence of digits in the winning number. Although the probability of a specific digit occurring is mathematically defined, the outcome for each individual lottery ball is binary—hit (matches the player’s number) or miss (does not match the player’s number)—and the outcome on any given trial is unknown. Such a scenario may better characterize the experience of group 50 in the present experiments, where reward was provided non-contingently but also unpredictably and the CS merely indicated the potential for reward without revealing whether it would occur on a given trial. Slot machines are more strongly linked with PG than are lottery tickets (Cox et al., 2000; Bakken et al., 2009), indicating an important role for instrumental factors (and immediacy) in the rewarding aspects of gambling for this population (Loba et al., 2001). Bibẹẹkọ, ilana Pavlovian ti a ṣe awoṣe ninu awọn adanwo lọwọlọwọ (CS + ere ti ko ni idaniloju) yoo han pe o jẹ pataki ti ko ba to ipin ti iriri ere.
Paapọ pẹlu aini ibeere ohun elo ti o yege, nọmba awọn ẹya apẹrẹ miiran le ti ṣe alabapin si iwọntunwọnsi ati ilana iyipada ti awọn ipa idanwo. Awọn ẹgbẹ naa yatọ ni ifihan sucrose gbogbogbo bi airotẹlẹ laarin CS ati ẹsan sucrose. Botilẹjẹpe eyi le ti ṣe alabapin si iyipada laarin ẹgbẹ, ko le ṣalaye ni imurasilẹ idi ti awọn ẹranko ti o ni ifihan sucrose ti o tobi julọ (ẹgbẹ 100) ṣe afihan ifamọ kere ju ẹgbẹ 50. Ni afikun, ẹgbẹ 0 ko gba iyanju ṣaaju ifihan sucrose lori gbogbo idanwo. Botilẹjẹpe eyi ṣe idiwọ ifojusọna ifẹnukonu ti ẹsan, ko ṣakoso fun wiwa itunu ṣaaju ifijiṣẹ ere, eyiti o wa ni gbogbo awọn ẹgbẹ miiran. Lati koju ọran yii, iwadii ọjọ iwaju yẹ ki o pẹlu ipo kan nibiti awọn ẹranko gba ere lori gbogbo idanwo ni atẹle ifihan laileto si ayun didoju (ie, ti wiwa rẹ ko ṣe ifihan agbara fun ẹsan).
Another design limitation is the potential emergence of adjunctive behavior that could influence the effects of training schedule. In the face of uncertainty, animals may develop superstitious behaviors designed to enhance perceived control and reduce uncertainty-induced DA activation (cf. Harris et al., 2013). Nitorinaa o ṣee ṣe pe awọn apakan ti a ko ṣakoso ti apẹrẹ adanwo jẹ ki awọn ẹranko ṣe aiṣedeede awọn ipa ti iṣeto mimu. Iru ipa bẹẹ le ṣe alabapin si iwọntunwọnsi ati idahun iyipada si amphetamine ni ẹgbẹ 50 ni atẹle ikẹkọ CS + sucrose. Iwadi ojo iwaju yẹ ki o ṣe igbasilẹ ihuwasi lẹẹkọkan, laisi awọn pokes imu, lakoko awọn akoko ikẹkọ lati ṣe idanwo iṣeeṣe yii, ati iṣakoso fun iṣiro ti o yẹ ki o farahan. Nitoripe iru ihuwasi bẹẹ yoo nireti lati koju tabi dẹkun awọn ipa ti aidaniloju iṣeto-iṣeto, idahun locomotor si amphetamine ni ẹgbẹ 50 yẹ ki o ni ilọsiwaju nigbati o ba jẹ iṣakoso (ilana ilana tabi iṣiro). Nitorinaa, apẹrẹ ti o wa lọwọlọwọ (laisi iṣakoso) n pese idanwo Konsafetifu ti awọn ipa ti 50% CS + ere lori ifamọ amphetamine.
In terms of external validity, the use of male rats also limits the generalizability of the results. The lack of a clear “punishment” condition also differs from gambling, where large monetary losses are common and exert important motivational effects (Nieuwenhuis et al., 2005; Singh and Khan, 2012). The ability to accumulate reward is also absent from the present paradigm and cumulative winnings in a slot machine game have been found to interact with DA manipulations in humans (Tremblay et al., 2011; Smart et al., 2013). Bakanna, awọn anfani fun a jackpot jẹ ẹya pataki iyato laarin awọn bayi awoṣe ki o si gangan ayo .
Despite these limitations, the present results suggest that 50% variable CS + reward can engage DA pathways implicated in the reinforcing effects of gambling (Fiorillo et al., 2003; Anselme, 2013). Cross-sensitization of response to AMPH following this gambling-like schedule is consistent with a pivotal role for DA in gambling and psychostimulant drug effects (Zack ati Poulos, 2009), and extends earlier studies on cross-priming of motivation to gamble by AMPH in pathological gamblers (Zack ati Poulos, 2004). The present results also indirectly suggest that modest doses of AMPH, which do not cause supra-physiological DA release, may better model brain activity in response to intermittent reward signals (i.e., during gambling) than exposure to high (i.e., binge-like) doses of stimulant drugs (cf. Vanderschuren ati Pierce, ọdun 2010). Atilẹyin taara fun ifọrọranṣẹ yii le jẹri nipasẹ ṣiṣe iṣiro itusilẹ DA ni idahun si iṣeto CS-US oniyipada 50% ati awọn abere oriṣiriṣi ti AMPH ni lilo microdialysis.
Lati oju iwoye idanwo kan, awoṣe Pavlovian ti o wa lọwọlọwọ ati awoṣe oniṣẹ iṣaaju ti imuduro oniyipada mejeeji han lati ṣe agbekalẹ phenotype kan ti o dabi olutaja eleto eniyan. Bi iru, nwọn pese kan niyelori iranlowo si rodent ayo awọn iṣẹ-ṣiṣe eyi ti awoṣe ayo ihuwasi (bi awọn kan ti o gbẹkẹle odiwon) sugbon ni, titi bayi, nikan oojọ ti ni ilera eranko, awọn deede ti eda eniyan awujo gamblers. Da lori awọn iwe-kikọ, awọn ẹranko ti o farahan si ere oniyipada le yatọ daradara ni awọn iṣẹ ṣiṣe wọnyi, ni pataki ni idahun si awọn oogun DA-ergic. Apapọ awọn eku ayo phenotype pẹlu ayo awọn iṣẹ-ṣiṣe le laye ilana idagbasoke ti oogun fun awọn itọju ti PG, eyi ti o le wa ko le ni kikun aṣepe pẹlu awọn ẹranko ni ilera nikan. Awọn isọdọtun siwaju ninu apẹrẹ esiperimenta ati ilana ikẹkọ, gẹgẹ bi a ti ṣalaye loke, yẹ ki o mu ibaramu pọ si laarin awọn ẹranko ti o ni ikẹkọ ni apẹrẹ yii ati awọn onijagidijagan pathological gangan.
From the clinical-sociological standpoint, the finding that exposure to 50% variable CS + reward, which closely matches the reward schedule on a commercial slot machine (Tremblay et al., 2011), yi awọn ọpọlọ DA eto ni gbẹkẹle ati ki o fífaradà awọn ọna ni imọran wipe, ni awọn igba miiran, ayo aṣayan iṣẹ-ṣiṣe, bi oloro ti abuse, le jẹ a "pathogen" o lagbara ti nfa afẹsodi. Bibẹẹkọ, iwọn ipa iwọntunwọnsi ati iyipada giga ni idahun si ẹsan 50% CS + daba pe, bii awọn oogun ilokulo, ifarahan fun awọn eto ere bii ere lati ṣe igbega afẹsodi yoo dale pupọ lori profaili eewu ti tẹlẹ ti olutayo. Bibẹẹkọ, lati ṣafipamọ eewu eewu ti awọn ẹni-kọọkan si ifihan ti o pọju awọn ipa ti o ni ibatan si ayokele, o dabi ẹni pe awọn eto imulo ti a lo lati ṣe idiwọ lilo ati dinku ipalara lati awọn oogun ilokulo le tun gbooro si ere paapaa.
Gbólóhùn Ìfẹnukò Ìdánilójú
Awọn onkọwe sọ pe iwadi ti ṣe iwadi ni laisi awọn iṣowo ti owo tabi ti owo ti a le sọ bi ipọnju ti o ni anfani.
Acknowledgments
Iwadi yii ni inawo nipasẹ awọn ifunni lati Awọn Imọ-jinlẹ Adayeba ati Igbimọ Iwadi Imọ-ẹrọ ti Ilu Kanada si Paul J. Fletcher. A dupe tọkàntọkàn Ms. Djurdja Djordjevic fun mura awọn isiro.
jo
Anselme, P. (2013). Dopamine, motivation, and the evolutionary significance of gambling-like behaviour. Behav. Ipolowo ọlọ. 256, 1 – 4. doi: 10.1016 / j.bbr.2013.07.039
A.P.A. (2013). Aisan ati iwe afọwọkọ Iṣiro ti Awọn rudurudu Ọpọlọ, 5th Edn. Arlington, VA: Atejade Psychiatric Amerika.
Bakken, I. J., Gotestam, K. G., Grawe, R. W., Wenzel, H. G., and Oren, A. (2009). Gambling behavior and gambling problems in Norway 2007. Scand. J. Psychol. 50, 333 – 339. doi: 10.1111 / j.1467-9450.2009.00713.x
Boileau, I., Payer, D., Chugani, B., Lobo, D. S., Houle, S., Wilson, A. A., et al. (2013). Ni vivo evidence for greater amphetamine-induced dopamine release in pathological gambling: a positron emission tomography study with [C]-(+)-PHNO. Mol. Awoasinwin doi: 10.1038/mp.2013.163. [Epub ahead of print].
Bolles, R. C. (1972). Reinforcement, expectancy, and learning. Psychol. Rev. 79, 394–409. doi: 10.1037/h0033120
Camp, D. M., Browman, K. E., and Robinson, T. E. (1994). The effects of methamphetamine and cocaine on motor behavior and extracellular dopamine in the ventral striatum of Lewis versus Fischer 344 rats. Agbejade ọlọjẹ. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G., and Dell’Osso, L. (2012). Pathological gambling: a systematic review of biochemical, neuroimaging, and neuropsychological findings. Harv. Rev. Psychiatry 20, 130-148. ni: 10.3109 / 10673229.2012.694318
Cox, B. J., Kwong, J., Michaud, V., and Enns, M. W. (2000). Problem and probable pathological gambling: considerations from a community survey. Le. J. Awoasinwin 45, 548-553.
Dalley, J. W., Theobald, D. E., Eagle, D. M., Passetti, F., and Robbins, T. W. (2002). Deficits in impulse control associated with tonically-elevated serotonergic function in rat prefrontal cortex. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Everitt, BJ, ati Robbins, TW (2005). Awọn ọna ṣiṣe ti imudaniloju fun afẹsodi oogun: lati awọn iṣe si awọn iwa si ipa. Nat. Neurosci. 8, 1481–1489. doi: 10.1038 / nn1579
Fiorillo, CD, Tobler, PN, ati Schultz, W. (2003). Ṣiṣe ifaminsi ti oye iṣeeṣe ere ati idaniloju nipasẹ awọn iṣan dopamine. Science 299, 1898 – 1902. doi: 10.1126 / Imọ.1077349
Flagel, S. B., Watson, S. J., Akil, H., and Robinson, T. E. (2008). Individual differences in the attribution of incentive salience to a reward-related cue: influence on cocaine sensitization. Behav. Ipolowo ọlọ. 186, 48 – 56. doi: 10.1016 / j.bbr.2007.07.022
Fletcher, P. J., Tenn, C. C., Rizos, Z., Lovic, V., and Kapur, S. (2005). Sensitization to amphetamine, but not PCP, impairs attentional set shifting: reversal by a D1 receptor agonist injected into the medial prefrontal cortex. Psychopharmacology (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Flores, G., Wood, G. K., Barbeau, D., Quirion, R., and Srivastava, L. K. (1998). Lewis and Fischer rats: a comparison of dopamine transporter and receptors levels. Agbejade ọlọjẹ. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Frascella, J., Potenza, M. N., Brown, L. L., and Childress, A. R. (2010). Shared brain vulnerabilities open the way for nonsubstance addictions: carving addiction at a new joint? Ann. NY Acad. Sci. 1187, 294 – 315. doi: 10.1111 / j.1749-6632.2009.05420.x
Grimm, J. W., Buse, C., Manaois, M., Osincup, D., Fyall, A., and Wells, B. (2006). Time-dependent dissociation of cocaine dose-response effects on sucrose craving and locomotion. Behav. Pharmacol. 17, 143–149. doi: 10.1097/01.fbp.0000190686.23103.f8
Haile, C. N., Hiroi, N., Nestler, E. J., and Kosten, T. A. (2001). Differential behavioral responses to cocaine are associated with dynamics of mesolimbic dopamine proteins in Lewis and Fischer 344 rats. Synapse 41, 179–190. doi: 10.1002 / syn.1073
Harris, J. A., Andrew, B. J., and Kwok, D. W. (2013). Magazine approach during a signal for food depends on Pavlovian, not instrumental, conditioning. J. Exp. Psychol. Animu. Iwa. Ilana. 39, 107–116. doi: 10.1037/a0031315
Howell, D. C. (1992). Statistical Methods for Psychology. Boston, MA: Duxbury.
Ito, R., Dalley, J. W., Robbins, T. W., and Everitt, B. J. (2002). Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J. Neurosci. 22, 6247 – 6253.
Jiao, X., Pare, W. P., and Tejani-Butt, S. (2003). Strain differences in the distribution of dopamine transporter sites in rat brain. Prog. Neuropsychopharmacol. Biol. Aimakadi 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Koob, G. F., and Le Moal, M. (2008). Review. Neurobiological mechanisms for opponent motivational processes in addiction. Philos. Atagba R. Soc. Lọndọnu. B Biol. Sci. 363, 3113–3123. doi: 10.1098 / rstb.2008.0094
Kosten, T. A., Miserendino, M. J., Chi, S., and Nestler, E. J. (1994). Fischer and Lewis rat strains show differential cocaine effects in conditioned place preference and behavioral sensitization but not in locomotor activity or conditioned taste aversion. J. Pharmacol. Exp. Ther. 269, 137 – 144.
Kosten, T. A., Zhang, X. Y., and Haile, C. N. (2007). Strain differences in maintenance of cocaine self-administration and their relationship to novelty activity responses. Behav. Neurosci. 121, 380–388. doi: 10.1037/0735-7044.121.2.380
Leeman, RF, ati Potenza, MN (2012). Awọn afijq ati awọn iyatọ laarin awọn pathological ayo ati nkan na lilo ségesège: kan aifọwọyi lori impulsivity ati compulsivity. Psychopharmacology (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Leshner, A. I. (1997). Addiction is a brain disease, and it matters. Science 278, 45 – 47. doi: 10.1126 / Imọ.278.5335.45
Loba, P., Stewart, S. H., Klein, R. M., and Blackburn, J. R. (2001). Manipulations of the features of standard video lottery terminal (VLT) games: effects in pathological and non-pathological gamblers. J. Gambl. Ikẹkọ. 17, 297–320. doi: 10.1023/A:1013639729908
Martinez, D., Narendran, R., Foltin, R. W., Slifstein, M., Hwang, D. R., Broft, A., et al. (2007). Amphetamine-induced dopamine release: markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am. J. Onimọran 164, 622-629. doi: 10.1176 / appi.ajp.164.4.622
Mateo, Y., Lack, C. M., Morgan, D., Roberts, D. C., and Jones, S. R. (2005). Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology 30, 1455-1463. Ṣe: 10.1038 / sj.npp.1300687
Nestler, EJ (2001). Ipilẹ molikula ti ṣiṣu igba pipẹ ti o wa labẹ afẹsodi. Nat. Rev. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
Nieuwenhuis, S., Heslenfeld, D. J., von Geusau, N. J., Mars, R. B., Holroyd, C. B., and Yeung, N. (2005). Activity in human reward-sensitive brain areas is strongly context dependent. Awọn aworan Neuro 25, 1302-1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pierce, R. C., and Kalivas, P. W. (1997). A circuitry model of the expression of behavioral sensitization to amphetamine-like psychostimulants. Ọpọlọ Res. Ọpọlọ Res. Osọ. 25, 192. doi: 10.1016/S0165-0173(97)00021-0
Robbins, T. W., and Everitt, B. J. (1999). Drug addiction: bad habits add up. Nature 398, 567-570. ni: 10.1038 / 19208
Robinson, T. E., Becker, J. B., and Presty, S. K. (1982). Long-term facilitation of amphetamine-induced rotational behavior and striatal dopamine release produced by a single exposure to amphetamine: sex differences. Agbejade ọlọjẹ. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Robinson, TE, ati Berridge, KC (1993). Ipilẹ nkankikan ti ifẹkufẹ oogun: imọ-imọ-imọ-imọran ti afẹsodi. Ọpọlọ Res. Ọpọlọ Res. Osọ. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, ati Berridge, KC (2001). Atunse-ifamọra ati afẹsodi. afẹsodi 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Singer, B. F., Scott-Railton, J., and Vezina, P. (2012). Unpredictable saccharin reinforcement enhances locomotor responding to amphetamine. Behav. Ipolowo ọlọ. 226, 340 – 344. doi: 10.1016 / j.bbr.2011.09.003
Singh, V., and Khan, A. (2012). Decision making in the reward and punishment variants of the iowa gambling task: evidence of “foresight” or “framing”? Iwaju. Neurosci. 6:107. doi: 10.3389 / fnins.2012.00107
Skinner, B. F. (1953). Imọ ati Ihuwa Eniyan. New York, NY: Free Press.
Smart, K., Desmond, R. C., Poulos, C. X., and Zack, M. (2013). Modafinil increases reward salience in a slot machine game in low and high impulsivity pathological gamblers. Neuropharmacology 73, 66-74. doi: 10.1016/j.neuropharm.2013.05.015
Tremblay, A. M., Desmond, R. C., Poulos, C. X., and Zack, M. (2011). Haloperidol modifies instrumental aspects of slot machine gambling in pathological gamblers and healthy controls. Okudun. Biol. 16, 467 – 484. doi: 10.1111 / j.1369-1600.2010.00208.x
Vanderschuren, L. J., and Kalivas, P. W. (2000). Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology (Berl.) 151, 99 – 120. doi: 10.1007 / s002130000493
Vanderschuren, L. J., and Pierce, R. C. (2010). Sensitization processes in drug addiction. Curr. Top. Behav. Neurosci. 3, 179–195. doi: 10.1007/7854_2009_21
Vanderschuren, L. J., Schoffelmeer, A. N., Mulder, A. H., and De Vries, T. J. (1999). Dopaminergic mechanisms mediating the long-term expression of locomotor sensitization following pre-exposure to morphine or amphetamine. Psychopharmacology (Berl.) 143, 244 – 253. doi: 10.1007 / s002130050943
Verdejo-Garcia, A., Lawrence, A. J., and Clark, L. (2008). Impulsivity as a vulnerability marker for substance-use disorders: review of findings from high-risk research, problem gamblers and genetic association studies. Neurosci. Biobehav. Rev. 32, 777 – 810. doi: 10.1016 / j.neubiorev.2007.11.003
Vezina, P. (2004). Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci. Biobehav. Rev. 27, 827 – 839. doi: 10.1016 / j.neubiorev.2003.11.001
Volkow, N. D., Wang, G. J., Fowler, J. S., Logan, J., Gatley, S. J., Hitzemann, R., et al. (1997). Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. Nature 386, 830 – 833. doi: 10.1038 / 386830a0
Volkow, ND, Wang, GJ, Fowler, JS, ati Tomasi, D. (2012). Afẹsodi circuitry ninu awọn eniyan ọpọlọ. Lododun. Rev. Pharmacol. Egboogi. 52, 321–336. doi: 10.1146/annurev-pharmtox-010611-134625
Winer, B. (ed.). (1971). Statistical Principles in Experimental Design. New York, NY: McGraw-Hill.
Zack, M., and Poulos, C. X. (2004). Amphetamine primes motivation to gamble and gambling-related semantic networks in problem gamblers. Neuropsychopharmacology 29, 195-207. Ṣe: 10.1038 / sj.npp.1300333
Zack, M., and Poulos, C. X. (2009). Parallel roles for dopamine in pathological gambling and psychostimulant addiction. Curr. Drug Abuse Rev. 2, 11 – 25. doi: 10.2174 / 1874473710902010011
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F., and Flores, G. (2005). Strain differences of dopamine receptor levels and dopamine related behaviors in rats. Ọpọlọ Res. Bull. 65, 339 – 347. doi: 10.1016 / j.brainresbull.2005.01.009
Keywords: pathological gambling, sensitization, amphetamine, dopamine, uncertainty
Citation: Zack M, Featherstone RE, Mathewson S and Fletcher PJ (2014) Chronic exposure to a gambling-like schedule of reward predictive stimuli can promote sensitization to amphetamine in rats. Iwaju. Behav. Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Ti gba: 01 Kọkànlá Oṣù 2013; Ti gba: 23 Oṣu Kini 2014;
Published online: 11 February 2014.
Satunkọ nipasẹ:
Ọmọbinrin Bryan F., University of Michigan, Orilẹ Amẹrika
Àyẹwò nipasẹ:
Louk Vanderschuren, University of Utrecht, Netherlands
Ruud Van Den Bos, Radboud University Nijmegen, Netherlands
Patrick Anselme, University of Liège, Bẹljiọmu
Copyright © 2014 Zack, Featherstone, Mathewson and Fletcher. This is an open-access article distributed under the terms of the Aṣẹ Ipese Creative Commons (CC BY). Lilo, pinpin tabi atunse ni awọn apejọ miiran ti jẹ idaniloju, ti a fun ni akọwe tabi onilẹwe ti o ni akọkọ ati pe a ṣe apejuwe atilẹba ti o wa ninu iwe akọọlẹ yii, ni ibamu pẹlu ilana ẹkọ ti a gba. A ko lo lilo, pinpin tabi atunse ti ko ni ibamu pẹlu awọn ofin wọnyi.
*Correspondence: Martin Zack, Cognitive Psychopharmacology Laboratory, Neuroscience Department, Centre for Addiction and Mental Health, 33 Russell Street, Toronto, ON M5S 2S1, Canada e-mail: [imeeli ni idaabobo]