Front Behav Neurosci. 2015 Mei 19;9:128. doi: 10.3389/fnbeh.2015.00128. e-versameling 2015.
Andreatta M1, Pauli P1.
Abstract
In klassieke kondisionering word 'n aanvanklik neutrale stimulus (gekondisioneerde stimulus, CS) geassosieer met 'n biologies opvallende gebeurtenis (ongekondisioneerde stimulus, US), wat pyn (aversiewe kondisionering) of voedsel (aptytlike kondisionering) kan wees. Na 'n paar assosiasies, is die CS in staat om onderskeidelik óf verdedigende óf volkome reaksies te inisieer. In teenstelling met aversiewe kondisionering, word aptytkondisionering selde by mense ondersoek, hoewel die belangrikheid daarvan vir normale en patologiese gedrag (bv. vetsug, verslawing) onmiskenbaar is. Die huidige studie het ten doel om dierebevindinge oor eetluskondisionering te vertaal na mense wat voedsel as 'n VS gebruik. Drie-en-dertig deelnemers is tussen 8:10 en XNUMX:XNUMX sonder ontbyt ondersoek om te verseker dat hulle honger voel. Tydens twee verkrygingsfases het een geometriese vorm (avCS+) 'n aversiewe VS (pynlike elektriese skok) voorspel, 'n ander vorm (appCS+) het 'n aptytlike VS voorspel (sjokolade of sout pretzel volgens die deelnemers se voorkeur), en 'n derde vorm (CS– ) het nie een van die VSA voorspel nie. In 'n uitwissingsfase is hierdie drie vorms plus 'n nuwe vorm (NUUT) weer aangebied sonder Amerikaanse aflewering. Valensie- en opwekkingsgraderings sowel as skrik- en velgeleiding (SCR)-reaksies is as leerindekse ingesamel. Ons het suksesvolle aversiewe en aptytwekkende kondisionering gevind. Aan die een kant is die avCS+ gegradeer as meer negatief en meer opwindend as die CS– en het skrikversterking en verhoogde SCR geïnduseer. Aan die ander kant is die appCS+ meer positief gegradeer as die CS– en het skrik-demping en groter SCR veroorsaak. Samevattend het ons dierebevindings by (honger) mense suksesvol bevestig deur aptytlike leer en normale afkeerleer te demonstreer.
Inleiding
Die voorspelling van bedreiging en voedsel is van kardinale belang vir enige organisme se voortbestaan. In klassieke kondisionering (Pavlov, 1927), gaan 'n leidraad 'n aversiewe gebeurtenis vooraf, soos 'n ligte pynlike elektriese skok (aversiewe ongekondisioneerde stimulus, VS), of 'n aptytgebeurtenis, soos 'n voedselkorrel (aptyttige US), verskeie kere. Daarna is hierdie stimulus alleen (nou gemerk CS, CS+) in staat om onderskeidelik óf defensiewe óf volkome response te ontlok. Eersgenoemde soort assosiatiewe leer word aversiewe kondisionering genoem, terwyl laasgenoemde appetitiewe kondisionering genoem word.
Alhoewel die voorspelling van eetlusgebeure net so belangrik is vir oorlewing as die voorspelling van afkerende gebeurtenisse, word aptytkondisionering merkwaardig minder by diere ondersoek (Bouton en Peck, 1989; Koch et al., 1996; McDannald et al., 2011, vir 'n resensie sien Martin-Soelch et al., 2007) sowel as by mense (Klucken et al., 2009, 2013; Austin en Duka, 2010; Delgado et al., 2011; Levy en Glimcher, 2011). Hierdie gebrek aan navorsing kan moontlik te wyte wees aan die kompleksiteit van die aptyt-paradigma in vergelyking met die aversiewe een. Byvoorbeeld, kos as 'n primêre versterker moet gelewer word wanneer die organisme honger is om lonend te wees (vir 'n onlangse resensie sien Dickinson en Balleine, 1994; Clark et al., 2012). In menslike navorsing is hierdie moeilikheid oorkom deur geld te gebruik (Austin en Duka, 2010; Delgado et al., 2011; Levy en Glimcher, 2011) of erotiese prente (Klucken et al., 2009, 2013). Neurobeeldingstudies het egter daarop gewys dat primêre (dws versnaperinge of drankies) en sekondêre (dws geld) versterkers sommige algemene breinstreke (bv. striatum) aktiveer, maar ook duidelike patrone van aktivering behels (Delgado et al., 2011; Levy en Glimcher, 2011).
Na ons kennis het slegs 'n paar menslike kondisioneringstudies die effekte van primêre aptytversterkers soos reuk (Gottfried et al., 2002), water (Kumar et al., 2008), of kos (Preost et al., 2012). In die eerste studie, Gottfried et al. (2002) geassosieerde neutrale gesigte (CS'e) met óf 'n onaangename, 'n aangename of 'n neutrale reuk. Interessant genoeg het hulle groter aktivering gevind in die orbitofrontale korteks (OFC) en in die ventrale striatum in reaksie op die aptytwekkende CS+ teenoor die aversiewe CS+. Hulle het tot die gevolgtrekking gekom dat die OFC die waarde van die reuk verwerk en betrokke is by die oordrag van die affektiewe waarde van die olfaktoriese (VS) na die visuele (CS) sisteem. Die (ventro) striatale aktivering is geïnterpreteer as weerspieëling van die aptytwekkende CR wat deur die aptytlike CS+ ontlok word. In die tweede studie, Kumar et al. (2008) hulle deelnemers (gesonde kontrolepersoneel en pasiënte met ernstige depressie) vroegoggend na die laboratorium genooi en hulle gevra om nie gedurende die nag te drink om te verseker dat hulle dors was voor die skandering nie. Fraktale prente was CSs en 0.1 ml water die VSA. Interessant genoeg, onder ander aktiverings, het gesonde deelnemers (maar nie depressiepasiënte nie) groter aktivering in die ventrale striatum tot die aptytwekkende CS+ getoon, wat daarop dui dat hierdie stimulus as lonend verwerk is. In die derde studie, Prévost et al. (2012) het ook fraktale prente as CS'e en óf soet óf sout versnaperinge as US aangebied volgens die deelnemers se voorkeur. Die CS is vir 6 s aangebied en gedurende die laaste sekonde is 'n kosprentjie addisioneel aangebied. Elke keer as die kosprentjie verskyn het, het die eksperimenteerder 'n stukkie kos in die deelnemers se hande geplaas, wat toegelaat is om dadelik die peuselhappie te eet. Ongelukkig het die skrywers nie die breinaktivering aan die aptytwekkende CS+ tydens die klassieke kondisioneringsfase gerapporteer nie; hulle het egter minder hartvertraging na die beloonde CS+ waargeneem in vergelyking met die nie-beloonde CS–, wat differensiële kondisioneringseffekte aandui.
Die gebrek aan studies wat die skrikreaksie as 'n indeks vir eetlus kondisionering gebruik, is verbasend, veral in die lig van die breë gebruik daarvan in aversiewe kondisionering. Skrikreaksie is 'n voorvaderlike en outomatiese verdedigingsreaksie teen skielike, onverwagte en sterk afkeerlike gebeurtenisse (Koch, 1999). Hierdie verdedigingsreaksie word bemiddel deur 'n relatief eenvoudige neuronale pad wat die kogleêre wortelneurone, die caudale pontine kern van die retikulêre vorming (PnC) en spinale motoneurone (Fendt en Fanselow, 1999; Koch, 1999). Dierestudies het aan die lig gebring dat skrikpotensiëring afhang van die projeksies van die amygdala na die PnC (Fendt en Fanselow, 1999; Koch, 1999), terwyl skrik-demping afhang van 'n ongeskonde nucleus accumbens (NAcc, Koch, 1999). Sodanige modulasie van die skrikreaksie is 'n nuttige implisiete maatstaf vir die valensie van die voorgestelde voorgrondstimuli. Potensiasie dui dus op negatiewe valensie, terwyl verswakking positiewe valensie aandui, beide sonder om grootliks deur kognitiewe prosesse beïnvloed te word (Hamm en Weike, 2005; Andreatta et al., 2010). Na ons kennis het slegs een dierstudie aptytkondisionering ondersoek deur skrikreaksies as 'n afhanklike maatstaf te meet. Interessant genoeg het die ondersoekde rotte skrik-demping getoon vir 'n CS+ (dws lig) wat verband hou met die lewering van 'n sukrose-oplossing. Sodanige verswakking is veral benadeel in daardie diere met NAcc-letsels, maar nie in dié met amygdala-letsels nie, wat daarop dui dat die NAcc 'n spesifieke rol speel om aptytwekkende CR's te ontlok en om skrikreaksies te versag (Koch et al., 1996).
Die huidige studie is na ons kennis die eerste om hierdie aptytkondisioneringsparadigma na mense te vertaal deur primêre versterkers te gebruik as US, maw soet (sjokolade Smarties)®) of sout (klein sout krakeling) kos, en skrik modulasie as 'n maatstaf van CR'e. Ons het verwag dat die appCS + sterk aptytwekkende CR's sou aktiveer, soos weerspieël in skrikverswakking, verbeterde SCR en positiewe valensiegradering in vergelyking met die ander stimuli, dws die avCS + en die CS–.
Materiaal en metodes
Deelnemers
Twee-en-veertig vrywilligers het aanvaar om aan die studie deel te neem en het kursuskrediete ontvang. Nege deelnemers is van die analise uitgesluit, twee weens tegniese probleme, drie omdat hulle as nie-reagerende deelnemers gekodeer is (gemiddelde skrikamplitude < 5 μV), en vier omdat hulle nie genoeg skrikreaksies per toestand getoon het nie (minimum = 2; vir besonderhede, sien Materiale en Metode). Uiteindelik is 33 deelnemers in die analise oorweeg (16 mans; gemiddelde ouderdom: 22.09 jaar, SD: 2.84; reeks: 18–29 jaar). Vier deelnemers was nie inheemse Duitsers nie, en ses was linkshandig. Een deelnemer het regdeur die eksperiment onbewus gebly van die CSs-USs-assosiasies (sien Prosedure), maar ons het besluit om nie hierdie deelnemer uit te sluit nie omdat sy antwoorde normaal was en nie resultate beïnvloed het nie.
Materiale
Ongekondisioneerde stimuli (VSA)
Twee soorte US is gebruik. As averse US het ons 'n effens pynlike elektriese skok op die nie-dominante voorarm van deelnemers toegepas. Die elektriese skok is gelewer deur middel van twee elektrodes met 9 mm deursnee en spasiëring 30 cm. Die elektriese stimulus het bestaan uit 'n polsstimulus met 'n frekwensie van 50 Hz en duur van 200 ms, gegenereer deur 'n stroomstimulator (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, VK, 400 V, maksimum van 9.99 mA). Die intensiteit van die elektriese skok is individueel bepaal deur 'n drempelprosedure wat voorheen beskryf is (Andreatta et al., 2010). Kortliks, deelnemers het twee reekse stygende en dalende intensiteite in intervalle van 0.5 mA ondergaan. Hulle moes elke stimulus beoordeel op 'n visuele skaal wat wissel van 0 (voel glad niks) tot 10 (regtig intense pyn) met 4 as 'n anker vir die drumpel (net merkbare pyn). Die gemiddelde intensiteit van die elektriese stimulus was 2.12 mA (SD = 0.56) en dit is as pynlik gegradeer (M = 6.45, SD = 1.73). Die aptytlike VSA het bestaan uit óf 'n sjokolade (Smarties®) of klein sout pretzel. Die keuse van die aptytlike US het afgehang van die deelnemer se individuele voorkeur soos gerapporteer tydens die voorlopige onderhoud. Deelnemers moes naamlik rapporteer of hulle gewoonlik sout of soet kos tydens ontbyt eet. Hulle kon ook vrylik kies of hulle die sjokolade of die sout pretzel verkies tydens die eksperiment self. Op die ou end het 22 deelnemers die sjokolade en 11 die klein sout pretzel gekies.
Gekondisioneerde stimuli (CS)
Geometriese vorms (blou vierkant, geel sirkel, groen driehoek, rooi seshoek) met 'n diagonaal van 8 cm is as CS'e aangebied. Die vorms is vir 8 s in die middel van 'n swart rekenaarskerm aangebied. Een vorm (avCS+) was altyd geassosieer met die aversiewe US (pynlike elektriese skok), een vorm (appCS+) was altyd geassosieer met die aptytlike US (sjokolade of sout krakeling), een vorm (CS–) was nooit geassosieer met óf die aversiewe of die aptytlike US's, en die vierde vorm (NUUT) is in die uitsterwingsfase aangebied, maar nie tydens die verkrygingsfase nie om die neutraliteit daarvan te verseker.
Skrik sonde
Wit geraas van 103 dB met 'n duur van 50 ms is as 'n skrik-sonde gebruik. Die akoestiese stimuli is binauraal oor oorfone aangebied en het ewekansig 4–6 s na die vorm se aanvang plaasgevind.
vraelyste
Voor en na die eksperiment moes deelnemers die Duitse weergawes van die State-Trait Anxiety Inventory (STAI, Laux et al., 1981) en die Positiewe Negatiewe Affekte-skedule (PANAS, Krohne et al., 1996). Die STAI is 'n inventaris om deelnemers se eienskap en/of staatsangs te assesseer en bestaan uit 20 items vir beide die eienskap en die staat weergawes. Deelnemers se angsvlak voor (M = 37.06, SD = 7.80) en daarna (M = 39.33, SD = 9.16) die eksperiment het nie betekenisvol verander nie [t(32) = 1.61, p = 0.117]. Eienskap-angstellings in die huidige steekproef het tussen 20 en 58 gewissel (M = 36.6, SD = 8.98), wat vergelykbaar is met die gepubliseerde normale reeks volwassenes (Laux et al., 1981). Die PANAS (Krohne et al., 1996) is 'n indeks vir positiewe en die negatiewe bui. Individue met hoë tellings op die positiewe-affekte-skaal (PAS) is geneig tot emosies soos entoesiasme, terwyl individue met hoë tellings op die negatiewe-affekte-skaal (NAS) geneig is tot emosies soos nood. Elke item bestaan uit 'n byvoeglike naamwoord, en deelnemers dui op 'n skaal wat wissel van 1 (baie effens) tot 5 (uiters) aan tot watter mate die byvoeglike naamwoord hul gevoelens op daardie spesifieke oomblik weerspieël. Geen beduidende verskille is gevind in deelnemers se negatiewe invloed as gevolg van die eksperiment nie [begin: M = 11.67, SD = 2.29; einde: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Op een of ander manier het deelnemers van die begin af hul positiewe bui aansienlik verander (M = 26.72, SD = 4.70) tot die einde (M = 23.66, SD = 7.02) van die eksperiment [t(31) = 3.11, p = 0.004]. Hierdie afname in die deelnemers se positiewe bui kon moontlik verband hou met die onaangenaamheid van die paradigma (pynlike elektriese skokke sowel as aversiewe wit geraas is aangebied).
Prosedure
By aankoms in die laboratorium het deelnemers 'n ingeligte toestemmingsvorm gelees en onderteken wat deur die etiese komitee van die Departement Sielkunde van die Universiteit van Würzburg goedgekeur is. Hulle is nie ingelig oor die gebeurlikheid tussen CS'e en US'e nie. Nadat die vraelyste ingevul is, is die elektrodes geheg en die pyndrempelprosedure is uitgevoer soos hierbo beskryf.
Gedurende die gewenningsfase, is die vier meetkundige vorms twee keer aangebied met 'n inter-proef-interval (ITI) wat wissel tussen 18 en 25 s (gemiddeld: 21.5 s). Geen VS- of skriktoetse is gedurende hierdie fase afgelewer nie.
Voor die verkrygingsfase is sewe aanvalle van wit geraas elke 7–15 s gelewer om die aanvanklike skrikreaktiwiteit te verminder.
Die volgende twee verkrygingsfases was identies (Figuur 1). Elke verkrygingsfase het uit 24 proewe bestaan: 8 CS– proewe, 8 avCS+ proewe en 8 appCS+ proewe. Die CS-volgorde was pseudorandom met die enigste beperking dat dieselfde stimulus nie meer as twee keer in 'n ry aangebied kon word nie. Die avCS+ is veral saam met 'n weerligstraal aangebied as 'n simbool vir die elektriese skok en die pynlike US is teen die afslag afgelewer. Die appCS+ is in samestelling aangebied met 'n beeld van Smarties of 'n sout pretzel en die deelnemer kon 'n Smarties of 'n pretzel uit 'n pot kies. Die CS– is saam met 'n verbodsimbool aangebied en geen US is afgelewer nie. Tydens drie van die 8 CS-aanbiedings van elke tipe, is 'n skrik-sonde tussen 4 en 6 s na stimulusaanvang afgelewer. Drie bykomende skrik-probes is tydens die ITI's aangebied om hul onvoorspelbaarheid te verseker en om skrik-habituasie te verminder. Die ITI, bestaande uit 'n swart skerm, het tussen 18 en 25 s gewissel met 'n gemiddelde van 21.5 s.
FIGUUR 1
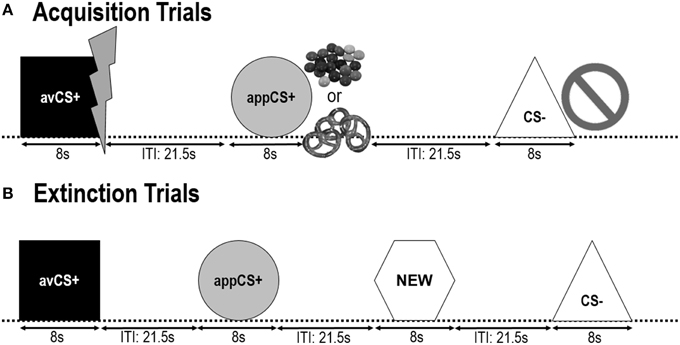
Figuur 1. Proewe tydens die twee verkrygingsfases (A) en die uitwissingsfase (B). Deelnemers het geleer dat een vorm (avCS+) 'n ligte pynlike elektriese skok voorspel, een vorm (appCS+) het óf 'n stukkie sjokolade óf 'n klein sout krakeling voorspel (volgens hul voorkeur), en 'n derde vorm (CS–) het geen voorspel biologies opvallende gebeurtenis. Elke vorm is saam met 'n prentjie aangebied wat 'n elektriese skok, smarties/sout pretzel, of niks uitbeeld, afhangende van die assosiasie met die VSA. Tydens die uitwissingsfase is die drie geometriese vorms weereens aangebied, maar geen US'e is afgelewer nie. Daarbenewens is 'n vierde geometriese vorm (NUUT) as 'n neutrale kontrole aangebied.
Gedurende die uitsterwingsfase, het deelnemers die drie meetkundige vorms (dws die avCS+, die appCS+ en die CS–) weer gesien plus 'n nuwe neutrale vorm (NUUT). Geen US is afgelewer nie en die vorms is nie saam met foto's van flitse, sjokolade/sout pretzels of bans aangebied nie. Elke stimulus is agt keer in 'n pseudorandom volgorde aangebied (dws dieselfde stimulus is nie meer as twee keer in 'n ry aangebied nie), wat 32 proewe geskep het. Skriksonde-stimuli is tydens 4 uit 8 stimulus-aanbiedings van elke CS-tipe aangebied. Soos in die verkryging en die gewenningsfase, het die ITI's tussen 18 en 25 s gewissel, en 4 bykomende skriktoetse is onvoorspelbaar tydens die ITI's afgelewer.
Na elke fase het deelnemers die valensie (aangenaamheid) en die opwekking (opwindend) van die CS'e beoordeel met visuele analoogskale (VAS) wat wissel van 1 tot 9. Die valansskaal het gewissel van "1" wat "baie onaangenaam" tot "9" aandui. wat "baie aangenaam" aandui; die opwekkingskaal het gewissel van "1" wat "kalm" aandui tot "9" wat "opwindend" aandui. Daarbenewens is gebeurlikheidsgraderings na die twee verkrygingsfases en die uitwissingsfase beoordeel. Deelnemers het 'n geometriese vorm vir 1 s gesien en dan moes hulle aandui of hierdie vorm met die elektriese skok geassosieer word, met die sjokolade/sout krakeling, met niks, en of hulle nie in staat was om enige assosiasie te maak nie. Veral, alle deelnemers (behalwe een) was bewus van gebeurlikheid na Verkryging 2.
Data vermindering
Fisiologiese response is aangeteken met 'n V-Amp 16 versterker en Vision Recorder V-Amp Edition sagteware (weergawe 1.03.0004, BrainProducts Inc., München, Duitsland). 'n Monsternemingstempo van 1000 Hz en 'n 50 Hz-kerffilter is toegepas. Die vanlyn ontledings is uitgevoer met Brain Vision Analyzer (weergawe 2.0; BrainProducts Inc., München, Duitsland).
Skrikreaksie
Skrikreaksie is deur middel van elektromiografie (EMG) aan die linkerkant gemeet orbicularis oculi spier met twee 5 mm Ag/AgCl elektrodes. In ooreenstemming met riglyne (Blumenthal et al., 2005), is een elektrode onder die pupil geplaas en die tweede een 1 cm lateraal. Die grond- en verwysingselektrodes is onderskeidelik op die regter- en linkermastoïede geplaas. Voordat die elektrodes geheg is, is die vel liggies afgeskuur en met alkohol skoongemaak om impedansie onder 10 kΩ te hou. Die elektromiografiese sein is vanlyn gefiltreer met 'n 28 Hz lae afsnyfilter en 'n 500 Hz hoë afsnyfilter. Toe is die EMG-sein reggestel en 'n bewegende gemiddelde van 50 ms is toegepas. Ons het die 50 ms voor die begin van die skrik-sonde as 'n basislyn gebruik (Grillon et al., 2006). Reaksies op skriktoetse is met die hand aangeteken, en proewe met oormatige basislynverskuiwings (±5 μV) of bewegingsartefakte is van verdere analise uitgesluit. Skrikreaksies laer as 5 μV is as nul gekodeer en oorweeg vir die berekening van skrikgrootte (Blumenthal et al., 2005). Altesaam 10.4% van proewe is verwerp, en 'n minimum van 2 uit 3 skrikreaksies in die verkrygingsfases en 4 uit 8 skrikreaksies in die uitsterwingsfase vir elke toestand was nodig om die deelnemer in die analisepoel te hou. Om hierdie rede is vier deelnemers uitgesluit. Die piek amplitude is gedefinieer as die maksimum piek relatief tot basislyn gedurende die 20-120 ms tydvenster na die aanvang van skrik sonde. Die rou data is dan genormaliseer binne-vakke met behulp van z-tellings en dan T-tellings om die invloed van individuele veranderlikheid te verminder en sielkundige prosesse beter op te spoor. Die T-tellings is gemiddeld vir elke toestand (avCS+, appCS+, CS–, NEW en ITI). Ten einde skrikpotensiëring of skrikverswakking te ondersoek, is die tellings vir die ITI skrikreaksies afgetrek van die skrikreaksies vir elke toestand.
Velgeleidingsreaksie (SCR)
Velgeleidingsreaksie (SCR) is aangeteken deur gebruik te maak van twee 5 mm Ag/AgCl elektrodes wat op die palm van die nie-dominante hand geplaas is. Die galvaniese reaksie is vanlyn gefiltreer met 'n 1 Hz hoë afsnyfilter. Die SCR is gedefinieer as die verskil (in μS) tussen die responsaanvang (1–3 s na stimulusaanvang) en die responspiek (Tranel en Damasio, 1994; Delgado et al., 2011). Proewe wat skriktoetse bevat, is nie in die ontleding van die SCR oorweeg nie. Response onder 0.02 μS is as nul gekodeer. Vyf verdere deelnemers is uitgesluit van die SCR-analise omdat hulle 'n gemiddelde SCR laer as 0.02 μS gehad het. Die rou velgeleidingsdata is vierkantswortel getransformeer om die verspreiding te normaliseer en tellings is vir elke toestand afsonderlik gemiddeld vir die twee verkrygingsfases (avCS+, appCS+, CS–) en die uitsterwingsfase (avCS+, appCS+, CS– en NEW ).
Statistiese analise
Alle data is ontleed met SPSS vir Windows (weergawe 20.0, SPSS Inc.). Vir die fisiologiese response is geskeide meerveranderlike variansieanalises (ANOVA) vir die twee verkrygingsfases en die uitsterwingsfase bereken. Die ANOVA vir die verkrygingsfases het stimulus (avCS+, appCS+, CS–) en fase (Verkryging 1, Verkryging 2) as binne-vakke faktore gehad. Die ANOVA vir die uitsterwingsfase het slegs stimulus (avCS+, appCS+, CS–, NEW) as 'n binne-subjekte-faktor gehad. Valensie-, opwekking- en gebeurlikheidsgraderings is ontleed met aparte ANOVA's wat die binne-vakke faktore stimulus (avCS+, appCS+, CS– en NEW) en fase bevat. Hierdie faktor het vier vlakke gehad vir die valensie- en opwekkingsgraderings (T1: na gewenningsfase, T2: na eerste verkrygingsfase, T3: na tweede verkrygingsfase, T4: na uitsterwingsfase), maar drie vlakke vir die gebeurlikheidsgraderings (T1: na eerste verkrygingsfase, T2: na tweede verkrygingsfase, T3: na uitsterwingsfase).
Die alfa (α) vlak is gestel op 0.05 vir alle ontledings. Die effekgrootte word as gedeeltelike η gerapporteer2.
Results
Die valensie- en opwekkingsgraderings vir elke fase word in Figuur uitgebeeld 2; die skrikreaksies en SCR's word in Figuur uitgebeeld 3.
FIGUUR 2
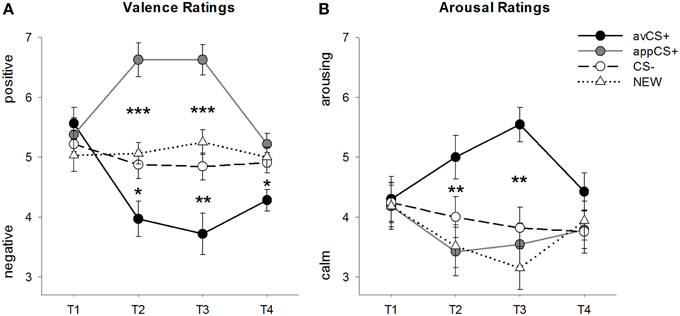
Figuur 2. Graderings vir valensie (A) en opwekking (B). Lyne (met standaardfoute) beeld die graderings na die gewenningsfase (T1), Verkryging 1 (T2), Verkryging 2 (T3) en die uitsterwingsfase (T4) uit. Die aversiewe CS+ (swart soliede lyn) het negatiewe valensie en hoë opwekking verkry na die twee verkrygingsfases in vergelyking met die CS– (swart stippellyn) en die NUWE (swart stippellyn). Dit is belangrik dat die aptytwekkende CS+ (grys soliede lyn) positiewe valensie verkry het in vergelyking met die CS– en die NUWE. *p <0.05, **p > 0.01, ***p <0.001.
FIGUUR 3
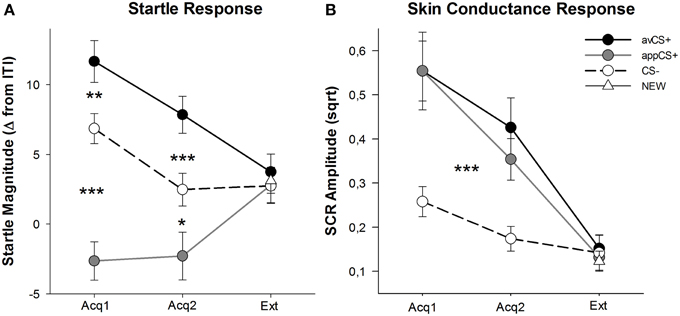
Figuur 3. Skrikreaksies (A) en velgeleiding (B) (met standaardfoute) tydens die eerste verkrygingsfase (Acq1), die tweede verkrygingsfase (Acq2) en die uitwissingsfase (Ext). Skrikreaksies is aansienlik gepotensiëer tot die aversiewe CS+ (swart soliede lyn) en aansienlik verswak tot die aptytwekkende CS+ (grys soliede lyn) in vergelyking met die CS– (swart stippellyn) tydens die verkrygingsfases. SCR was aansienlik groter vir die avCS+ en die appCS+ in vergelyking met die CS–. Geen verskille is tydens die uitsterwingsfase aan die lig gebring nie. *p <0.05, **p > 0.01, ***p <0.001.
Ratings
Die ANOVA vir die valensie graderings tydens verkryging het beduidende hoofeffekte van stimulus geopenbaar [F(3, 93) = 17.26, GG-ε = 0.801, p < 0.001, gedeeltelike η2 = 0.358] en fase [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, gedeeltelike η2 = 0.096] sowel as 'n beduidende interaksie tussen stimulus en fase [F(9, 279) = 9.54, GG-ε = 0.463, p < 0.001, gedeeltelike η2 = 0.235]. Volg op t-toetse dui aan dat die valensies van die vier meetkundige vorms aan die begin van die eksperiment identies was (ps > 0.19), terwyl die avCS+ na beide Verkryging 1 en Verkryging 2 as veral negatief gegradeer is en die appCS+ as veral positief. Spesifiek, die avCS+ is as meer negatief gevalueer in vergelyking met die CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], die NUWE [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p < 0.001], en die appCS+ [Acq1: t(31) = 5.41, p < 0.001; Acq2: t(31) = 6.11, p < 0.001]. Die appCS+ is as aansienlik meer positief as die CS- [Acq1: t(31) = 4.99, p < 0.001; Acq2: t(31) = 5.31, p < 0.001] en die NUWE [Acq1: t(31) = 4.92, p < 0.001; Acq2: t(31) = 4.14, p < 0.001]. Verskille tussen die CS– en die NUWE was nooit betekenisvol nie (ps > 0.18).
Wat belangrik is, is dat geen beduidende verskille gevind is wanneer die valensiegraderings vir die appCS+ wat met sjokolade vs. sout pretzel geassosieer word, vergelyk word nie [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Na die uitwissingsfase is die avCS+ steeds as meer negatief gegradeer in vergelyking met die CS– [t(31) = 2.40, p = 0.023], die NUWE [t(31) = 3.40, p = 0.002], en die appCS+ [t(31) = 3.35, p = 0.002]. Inteendeel, die valensie van die appCS+ het nie meer verskil van die CS– [t(31) = 1.77, p = 0.086] of die NUWE [t(31) = 1.07, p = 0.293].
Die ANOVA vir die opwekking graderings tydens verkryging het 'n beduidende hoofeffek van stimulus geopenbaar [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, gedeeltelike η2 = 0.181], maar nie van fase nie [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, gedeeltelike η2 = 0.038], en 'n beduidende interaksie tussen stimulus en fase [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, gedeeltelike η2 = 0.124]. Volg op t-toetse het geen betekenisvolle verskille tussen die stimuli met betrekking tot hul aanvanklike opwekking aangedui nie (ps > 0.74). Na die twee verkrygingsfases is die avCS+ egter meer opwindend gegradeer as die NUWE [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p < 0.001] en die appCS+ [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p < 0.001], en effens meer opwindend as die CS– na die eerste verkrygingsfase [t(32) = 1.96, p = 0.058], maar aansienlik meer opwindend na die tweede verkrygingsfase [t(32) = 3.65, p = 0.001]. In teenstelling met die valensiegraderings, het opwekking van die appCS+ nie verskil van die opwekking van die CS– en die NUWE (ps > 0.13) na die verkrygingsfases.
Net soos met die valensiegraderings, het CS– en NEW nie verskil in hul opwekking nie (ps > 0.07).
Soos die geval was vir valensie, is geen verskille geopenbaar vir die appCS+ wat met die sjokolade en die appCS+ geassosieer word met die sout pretzel [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Na die uitwissingsfase is geen beduidende verskille in die opwekkingsgraderings geopenbaar nie (ps > 0.08).
Vir bykomende post-hoc t-toetse wat die graderings na die gewenningsfase, die eerste en tweede verkrygingsfase en die uitsterwingsfase vergelyk, sien Aanvullende Materiale.
Skrikreaksie
Die ANOVA vir die verkrygingsfases het 'n hoofeffek van stimulus [F(2, 64) = 49.92, GG-ε = 0.964, p < 0.001, gedeeltelike η2 = 0.609], maar nie fase [F(1, 32) = 3.16, p = 0.085, gedeeltelike η2 = 0.090], en 'n beduidende interaksie Stimulus × Fase [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, gedeeltelike η2 = 0.095]. Volg op t-toetse het beduidende skrikpotensiasie aan die avCS+ getoon in vergelyking met die CS– tydens beide die eerste [t(32) = 3.27, p = 0.003] en die tweede [t(32) = 4.00, p < 0.001] verkrygingsfases. Skrikreaksies op die avCS+ is ook aansienlik versterk in vergelyking met dié op die appCS+, weer na beide Verkryging 1 [t(32) = 8.20, p < 0.001] en Verkryging 2 [t(32) = 5.74, p < 0.001]. Dit is belangrik dat die skrikwekkende omvang van die appCS+ aansienlik verswak is in vergelyking met die CS- tydens beide verkryging 1 [t(32) = 6.34, p < 0.001] en Verkryging 2 [t(32) = 2.91, p = 0.007]. Weereens en in ooreenstemming met die graderings, is geen differensiële skrikreaksies vir die appCS+ geopenbaar wanneer dit met die sjokolade of met die sout pretzel geassosieer word nie [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Tydens die uitsterwingsfase is geen betekenisvolle effek gevind nie [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, gedeeltelike η2 = 0.008].
Velgeleidingsreaksie (SCR)
Van die ANOVA's vir die SCR tydens die twee verkrygingsfases, die hoof effekte stimulus [F(2, 54) = 18.04, GG-ε = 0.908, p < 0.001, gedeeltelike η2 = 0.401] en fase [F(1, 27) = 20.91, p < 0.001, gedeeltelike η2 = 0.436] blyk betekenisvol te wees, maar nie hul interaksie nie [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, gedeeltelike η2 = 0.024]. Post-hoc t-toetse het aansienlik groter SCR aan die avCS+ [t(27) = 6.46, p < 0.001] en na die appCS+ [t(27) = 4.84, p < 0.001] in vergelyking met die CS–, terwyl deelnemers vergelykbare SCR getoon het met die avCS+ en die appCS+ [t(27) = 0.64, p = 0.527]. Daar is veral geen verskille in SCR's vir die sjokolade-appCS+ en die sout pretzel-appCS+ gevind nie [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Soos waar was vir die graderings en die skrikreaksie, is geen beduidende effekte vir die uitsterwingsfase gevind nie [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, gedeeltelike η2 = 0.010].
Bespreking
Die doel van hierdie studie was om dierebevindinge na mense te vertaal deur 'n klassieke aptytkondisioneringsparadigma te gebruik met 'n primêre versterker as ongekondisioneerde stimulus (VS). Om dit te doen, het deelnemers vroegoggend sonder ontbyt na die laboratorium gekom om te verseker dat hulle honger is, en volgens hul voorkeur is stukkies sjokolade of sout pretzels as eetlus gebruik. Tydens die verkrygingsfase het een geometriese vorm (avCS+) geassosieer geraak met 'n ligte pynlike elektriese skok (aversiewe VS), 'n ander vorm (appCS+) met die aptytlike US, en 'n derde vorm (CS–) met nóg die aptytlike US nóg met die afwykende VSA. Resultate dui op suksesvolle aversiewe en aptytwekkende kondisionering op die eksplisiete verbale vlak (dws graderings), op die implisiete gedragsvlak (bv. skrikreaksie) en op die fisiologiese vlak (bv. SCR). Spesifiek, die avCS+ in vergelyking met die CS– het meer negatiewe valensiegraderings, hoër opwekkingsgraderings, skrikpotensiëring en groter SCR ontlok. Die belangrikste is dat die appCS+ in vergelyking met die CS– meer positiewe valensiegraderings, skrik-demping en groter SCR veroorsaak het. Ons bevindinge oor aversiewe kondisionering was in ooreenstemming met verwagtinge, aangesien vorige studies bevind het dat 'n stimulus voorspellende bedreiging (avCS+) as aversief beoordeel word, groter vreesreaksies ontlok en fisiologiese opwekking verhoog (Fendt en Fanselow, 1999; Hamm en Weike, 2005; Andreatta et al., 2010, 2013). Ons resultate oor eetluskondisionering was ook in lyn met vorige menslike en dierstudies wat getoon het dat 'n stimulusvoorspellende beloning (appCS+) as positief beoordeel word, vreesreaksies inhibeer en fisiologiese opwekking verhoog (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin en Duka, 2010; Preost et al., 2012). Na ons kennis, is dit die eerste studie wat gekondisioneerde skrik-demping by mense aantoon na 'n stimulus wat 'n primêre beloning voorspel. Dit is belangrik dat ons die resultate van 'n dierestudie kon oordra en bevestig (Koch et al., 1996). Hierdie dierstudie het getoon dat skrikverswakking by rotte afhang van projeksies vanaf die NAcc (deel van die ventrale striatum) na die PnC. Daarom kan die skrik-demping in ons studie NAcc-aktiwiteit impliseer, wat ook in lyn is met fMRI-bevindinge (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy en Glimcher, 2011). Gevolglik kom ons tot die gevolgtrekking dat ons aptytkondisioneringsparadigma suksesvol was, soos beide die eksplisiete (graderings) en die implisiete (skrikverswakking) positiewe valensie aangedui het.
Benewens hierdie nuwe maar redelik voorspelbare bevinding, verdien nog twee interessante resultate vermelding. Eerstens het verbale en fisiologiese opwekkingsreaksies op die appCS+ gedissosieer. Tweedens het ons vinniger uitwissing van die aptytwekkende CR's gevind in vergelyking met die aversiewe CR's.
Die SCR dui simpatiese aktivering aan, wat verhoog word tot beide die beloning-geassosieerde stimulus (appCS +) en die bedreiging-geassosieerde stimulus (avCS +). Hierdie resultaat is in ooreenstemming met 'n vorige kondisioneringstudie, waarin erotiese prente as aptytwekkende US (Klucken et al., 2013), en 'n studie wat vergelykbare SCR onthul met 'n aanduiding wat geld voorspel en met 'n aanduiding wat 'n afkerende geraas voorspel (Austin en Duka, 2010). SCR is veral 'n oriënterende reaksie wat verband hou met die aktivering van die simpatiese sisteem. Hierdie reaksie is voorgestel om die voorbereiding van 'n gedragsreaksie op motiverend opvallende gebeure te weerspieël (Bradley, 2009). Op grond hiervan dink ons dat beide die bedreiging-geassosieerde en die beloning-geassosieerde stimuli 'n voorbereidende reaksie vir opeenvolgende gedragsreaksies ontlok het. Met ander woorde, voedsel- en pynseine het onderskeidelik voorbereiding vir benadering en vermydingsgedrag geïnisieer. In teenstelling met die hoë fisiologiese opwekking, het verbale reaksies lae opwekking vir die beloning-geassosieerde stimulus aangedui. Dit is moontlik dat verbale opwekking meer beïnvloed kan word deur die opwindende aard van die VSA eerder as die fisiologiese aktivering per se. Trouens, ons dink dat die appCS+ vs. die avCS+ as laag teenoor hoogs opwindend gegradeer is omdat hulle onderskeidelik aan lae en hoogs opwindende USs gekoppel is. Ongelukkig het ons nie opwekkingsgraderings vir die VSA's ingesamel nie, en daarom sal dit aan toekomstige studies wees om hierdie hipotese eksplisiet te toets.
Tydens die uitwissingsfase is geen VSA afgelewer nie. Dit het moontlik 'n nuwe inhiberende leer genaamd uitwissing veroorsaak (vir 'n oorsig sien Milad and Quirk, 2012), met beide die aversiewe en die aptytlike CR's wat as gevolg daarvan afneem. Op die gedrags- (dws skrikreaksie) en op die fisiologiese (dws SCR) vlakke, was geen differensiële reaksies op die avCS+, die appCS+ en die CS– meer waarneembaar nie. Net so het die eksplisiete opwekking van die gekondisioneerde stimuli wat na die uitwissingsfase geassesseer is op 'n lae vlak gelykgestel, wat suksesvolle uitsterwingsleer voorstel. Die avCS+ is egter steeds aansienlik meer negatief as die CS– gegradeer, terwyl die appCS+ valensie nie meer van die CS– valensie verskil het nie. Die stadiger uitwissing van die aversiewe eksplisiete reaksie kan wees as gevolg van 'n evolusionêre konserwatisme, wat beteken dat bedreigingseine veral moeilik is om te vergeet omdat 'n nie-reaksie op 'n bedreigingsein lewensgevaarlik kan wees.
Een vraag bly steeds: hoekom het die skrikreaksie (dws die implisiete valensie) dan heeltemal uitgedoof tydens die uitwissingsfase? Eerstens is hierdie response oor die hele verloop van die fase bereken. Daarom is dit denkbaar dat diskriminerende CR's steeds tydens die eerste proewe van die uitwissingsfase waarneembaar sal wees. Op 'n verkennende wyse het ons hierdie hipotese gevolg en gekyk na beide die skrikreaksies en SCR dwarsdeur die uitwissingsfase (sien aanvullende materiaal). Alhoewel ons nie beduidende verskille gevind het nie, het ons 'n effens hoër skrik-omvang vir die avCS + waargeneem in vergelyking met die CS- en die nuwe beheerstimulus. Ons het ook aan die begin van die uitwissingsfase effens meer skrik-demping vir die appCS+ waargeneem in vergelyking met die CS– en die nuwe beheerstimulus, wat egter binne 'n paar proewe verdwyn het. SCR na die appCS+ het reeds na 2de uitwissingsproef gedaal, terwyl die SCR na die avCS+ hoër gebly het vir byna alle uitwissing in vergelyking met die SCR na die NUWE. Alhoewel ons hierdie resultate met groot omsigtigheid moet interpreteer, het die skrikreaksies gelyk met die valensiegraderings. Boonop ondersteun beide skrikreaksies en SCR's die idee van 'n evolusionêre konserwatisme teenoor bedreigingstimuli.
Ten slotte moet ons 'n paar beperkings van hierdie studie erken. Eerstens, as gevolg van 'n tegniese probleem kon ons nie graderings van aangenaamheid (en opwinding) vir die sjokolade en die sout pretzel rapporteer nie. Die aptytwekkende CR's dui egter daarop dat deelnemers die twee US's inderdaad as aptytlik ervaar het. Tweedens het die duur van die aversiewe en die aptytlike US baie verskil. Die pynlike elektriese skok is dus gelewer teen die verstelling van die avCS+ en het presies 200 ms geduur, terwyl die sjokolade en die sout krakeling ongeveer 2 s na die aanvang van die appCS+ aan deelnemers aangebied is, en die duur was ondefinieerbaar omdat dit afhang van hoe vinnig die individu het hulle geëet. Die keuse om die aptytlike US's op hierdie manier te lewer, was gebaseer op 'n vorige studie in mense (Preost et al., 2012). Dit sou egter metodologies meer elegant wees om 'n aptytwekkende VSA te lewer wat meer vergelykbaar is met die aversiewe VSA, soos 'n slukkie sap of gemmerbier teen appCS+-afset. Om vir hierdie groot perseptuele verskil te vergoed, het ons meetkundige vorms aangebied in samewerking met 'n visuele stimulus wat die VSA simboliseer. Derdens kan ons nie beslis die moontlikheid uitsluit dat die vinnig uitgedoofde fisiologiese response in die uitsterwingsfase aan 'n metodologiese aspek te wyte is nie. Trouens, die visuele stimuli in hierdie fase is nie saam met die Amerikaanse simbool aangebied soos wat die geval was in die verkrygingsfases nie. Dit is moontlik dat die aanbieding van die CS'e alleen die CR'e kan beïnvloed het en om daardie rede was geen beduidende verskille sigbaar tydens die eerste uitsterwingsproewe nie. Ons vinnige uitgedoofde antwoorde stem egter ooreen met die uitgedoofde antwoorde in 'n vorige studie waarin die CS+ (gesig) in samestelling met die VSA (geskreeu) tydens verkryging aangebied is, maar nie tydens uitwissing nie (Lissek et al., 2008).
Ten slotte, ons het suksesvolle afkeer en aptytwekkende gekondisioneerde reaksies op onderskeidelik 'n stimulus wat met bedreiging geassosieer word en op 'n stimulus wat met beloning geassosieer word, gevind. Interessant genoeg het die eksplisiete (graderings) en die implisiete (skrikrefleks) vlak van reaksies op 'n sinergiese wyse gewerk, deurdat die avCS+ as negatief en geïnduseerde skrikpotensiasie gerapporteer is en die appCS+ as positiewe en geïnduseerde skrik-demping gerapporteer is. Verder het eksplisiete (graderings) en fisiologiese (SCR) opwekking van die appCS+ gedissosieer, wat twee afsonderlike prosesse weerspieël.
Konflik van belangstelling
Die skrywers verklaar dat die navorsing gedoen is in die afwesigheid van enige kommersiële of finansiële verhoudings wat as 'n potensiële botsing van belange beskou kan word.
Erkennings
Hierdie werk is ondersteun deur die Samewerkende Navorsingsentrum "Vrees, angs en angsversteurings," SFB-TRR 58, projek B1.
Aanvullende materiaal
Die aanvullende materiaal vir hierdie artikel kan aanlyn gevind word by: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Verwysings
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E., en Pauli, P. (2013). Pyn voorspelbaarheid keer valensie graderings van 'n verligting-geassosieerde stimulus. Voorkant. Syst. Neurosci. 7:53. doi: 10.3389/fnsys.2013.00053
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B., en Pauli, P. (2010). 'n Kloof tussen implisiete en eksplisiete gekondisioneerde valensie na pynverligting by mense. Proc. Biol. Wetenskap. 277, 2411–2416. doi: 10.1098/rspb.2010.0103
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Austin, AJ en Duka, T. (2010). Meganismes van aandag vir aptytwekkende en aversiewe uitkomste in Pavloviaanse kondisionering. Behav. Brein Res. 213, 19-26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV, en van Boxtel, A. (2005). Komiteeverslag: riglyne vir elektromiografiese studies van menslike skrik-oogknippering. Psychophysiology 42, 1-15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Bouton, ME en Peck, CA (1989). Kontekseffekte op kondisionering, uitwissing en herinstelling in 'n aptytsame kondisioneringsvoorbereiding. Anim. Leer. Gedrag. 17, 188-198. doi: 10.3758 / BF03207634
CrossRef Volledige teks | Google Scholar
Bradley, MM (2009). Natuurlike selektiewe aandag: oriëntering en emosie. Psychophysiology 46, 1-11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Clark, JJ, Hollon, NG, en Phillips, PEM (2012). Pavloviaanse waardasiestelsels in leer en besluitneming. Kur. Opin. Neurobiol. 22, 1054-1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Delgado, MR, Jou, RL, en Phelps, EA (2011). Neurale sisteme onderliggend aan aversiewe kondisionering by mense met primêre en sekondêre versterkers. Front. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Dickinson, A. en Balleine, B. (1994). Motiveringsbeheer van doelgerigte aksie. Anim. Leer. Gedrag. 22, 1-18. doi: 10.3758 / BF03199951
CrossRef Volledige teks | Google Scholar
Fendt, M. en Fanselow, MS (1999). Die neuroanatomiese en neurochemiese basis van gekondisioneerde vrees. Neurosci. Biobehav. Ds. 23, 743–760. doi: 10.1016/j.ijpsycho.2012.09.006
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Gottfried, JA, O'Doherty, J., en Dolan, RJ (2002). Appetitiewe en aversiewe reukleer by mense bestudeer met behulp van gebeurtenisverwante funksionele magnetiese resonansbeelding. J. Neurosci. 22, 10829–10837. Aanlyn beskikbaar by: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B., en Johnson, L. (2006). Kontekskondisionering en gedragsvermyding in 'n virtuele werklikheidsomgewing: effek van voorspelbaarheid. Biol. Psigiatrie 60, 752-759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Hamm, AO, en Weike, AI (2005). Die neuropsigologie van vreesleer en vreesregulering. Int. J. Psigofisiol. 57, 5–14. doi: 10.1016/j.ijpsycho.2005.01.006
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Klucken, T., Schwekendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Neurale aktiverings van die verkryging van gekondisioneerde seksuele opwekking: effekte van gebeurlikheidsbewustheid en seks. J. Seks. Med. 6, 3071-3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). Die 5-HTTLPR polimorfisme word geassosieer met veranderde hemodinamiese response gedurende appetitiewe kondisionering. Neurie. Brein Mapp. 34, 2549-2560. doi: 10.1002 / hbm.22085
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Koch, M. (1999). Die neurobiologie van skrik. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Koch, M., Schmid, A., en Schnitzler, H.-U. (1996). Plesier-demping van skrik word ontwrig deur letsels van die nucleus accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W., en Tausch, A. (1996). Untersuchungen mit 'n Duitse weergawe van "Positive and Negative Affect Schedule" (PANAS). Diagnostica 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I., en Steele, JD (2008). Abnormale temporele verskil beloning-leer seine in ernstige depressie. Brein 131, 2084-2093. doi: 10.1093 / brein / awn136
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P., en Spielberger, CD (1981). Das staatseienskap Angstinventar. Weinheim: Beltz-toets.
Levy, DJ, en Glimcher, PW (2011). Vergelyk appels en lemoene: gebruik beloningspesifieke en beloning-algemene subjektiewe waardevoorstelling in die brein. J. Neurosci. 31, 14693-14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Verhoogde vrees kondisionering tot sosiaal relevante ongekondisioneerde stimuli in sosiale angsversteuring. Am. J. Psigiatrie 165, 124-132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Martin-Soelch, C., Linthicum, J., en Ernst, M. (2007). Appetitiewe kondisionering: neurale basisse en implikasies vir psigopatologie. Neurosci. Biobehav. Ds. 31, 426-440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Abstract | CrossRef Volledige teks | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y., en Schoenbaum, G. (2011). Ventrale striatum en orbitofrontale korteks word albei benodig vir modelgebaseerde, maar nie modelvrye, versterkingsleer nie. J. Neurosci. 31, 2700-2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Milad, MR, en Quirk, G. (2012). Vrees uitsterwing as 'n model vir translasie neurowetenskap: tien jaar van vooruitgang. Annu. Eerw. Psychol. 63, 129-111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Pavlov, IP (1927). Gekondisioneerde reflekse: 'n ondersoek na die fisiologiese aktiwiteit van die serebrale korteks. Londen: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM, en O'Doherty, JP (2012). Neurale korrelate van spesifieke en algemene Pavlovian-tot-instrumentele oordrag binne menslike amygdalêre substreke: 'n hoë-resolusie fMRI-studie. J. Neurosci. 32, 8383–8390. doi: 10.1523/jneurosci.6237-11.2012
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Tranel, D., en Damasio, H. (1994). Neuroanatomiese korrelate van elektrodermale velgeleidingsreaksies. Psychophysiology 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Abstract | CrossRef Volledige teks | Google Scholar
Sleutelwoorde: klassieke kondisionering, beloning, straf, skrikrefleks, velgeleidingsreaksie
Aanhaling: Andreatta M en Pauli P (2015) Appetitiewe vs. Aversiewe kondisionering by mense. Front. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Ontvang: 11 Februarie 2015; Aanvaar: 05 Mei 2015;
Gepubliseer: 19 Mei 2015.
Geredigeer deur:
Niels Birbaumer, Universiteit van Tuebingen, Duitsland
Nagesien deur:
Tim Klucken, Justus Liebig Universiteit Giessen, Duitsland
Alessandro Angrilli, Universiteit van Padova, Italië
Kopiereg © 2015 Andreatta en Pauli. Dit is 'n oop-toegang artikel versprei onder die bepalings van die Creative Commons Erkenning Lisensie (CC BY). Die gebruik, verspreiding of reproduksie in ander forums word toegelaat, mits die oorspronklike skrywer (s) of lisensiegewer gekrediteer word en dat die oorspronklike publikasie in hierdie joernaal aangehaal word, in ooreenstemming met die aanvaarde akademiese praktyk. Geen gebruik, verspreiding of voortplanting word toegelaat wat nie aan hierdie bepalings voldoen nie.
*Korrespondensie: Marta Andreatta, Departement Sielkunde (Biologiese Sielkunde, Kliniese Sielkunde en Psigoterapie), Universiteit van Würzburg, Marcusstraße 9-11, D-97070 Würzburg, Duitsland, [e-pos beskerm]
