Is jy daar God Dit is my dopamienneuron
September 30, 2013 · deur Talia Lerner
Dopamienneurone is van die mees gestudeerde, mees sensasionele neurone daar buite. Die afgelope tyd het hulle egter 'n bietjie van 'n identiteitskrisis gegaan. Wat is 'n dopamienneuron? Enkele interessante onlangse wendings in dopamiennavorsing het die mite definitief ontbloot dat dopamienneurone almal van aard is - en jy moet enige studie wat hulle as sodanig behandel, bevraagteken.
Daar is baie maniere waarop 'n dopamienneuron (gedefinieer as 'n neuron wat die neurotransmitterdopamien vrystel) nie net 'n dopamienneuron is nie. Ek sal hier op drie baie cool maniere fokus:
- Nie alle neurone wat dopamien vrystel, laat dit terselfdertyd op dieselfde plek toe nie. Dopamienneurone het verskillende rolle in breinfunksie en gedrag, afhangende van hoe hulle in neurale bane aangeraak word.
- Nie alle neurone wat dopamien vrylating vrylaat, is slegs dopamien. Sommige kan ook ander neurotransmitters vrystel, wat diepgaande effekte kan hê op hoe hulle neurale kringfunksie beïnvloed.
- Nie alle neurone wat dopamien vrylaat, laat altyd dopamien vry. Sommige neurone kan hul dopamien sintese masjinerie aan of af skakel. As gevolg van hierdie vermoë, kan hulle nie eers in vorige navorsing as dopamienneurone erken word nie.
Voordat ek hierdie opwindende nuwe bevindinge beskryf, laat ek jou die standaard Neuroscience 101 bekendstelling gee aan dopamienneurone. Hierdie invloedryke teorie van dopamienneuronfunksie kom by ons uit Wolfram Schultz en kollegas se 1997-wetenskapskrif, "'N Neurale Substraat van Voorspelling en Beloning. " Dit het getoon dat dopamienneurone, wat teen 'n mate van agtergrond vuur brand, meer brand as gevolg van onvoorspelbare, maar nie voorspelbare voordele nie. Verder, as jy 'n beloning verwag en dit nie kry nie, brand die dopamienneurons minder. Hierdie bevinding het Schultz et al. Gelei. om voor te stel dat dopamienneurone kodeer "voorspellingsfout beloon." Dit beteken dat hulle jou vertel of dinge so goed, beter of erger is as wat jy verwag het. Schultz et al. gaan voort om te sê: "Die antwoorde van hierdie neurone is relatief homogeen. Verskillende neurone reageer op dieselfde wyse en verskillende aptitiewe stimuli ontlok soortgelyke neuronale reaksies. Alle antwoorde kom voor in die meerderheid dopamienneurone (55 tot 80%). "
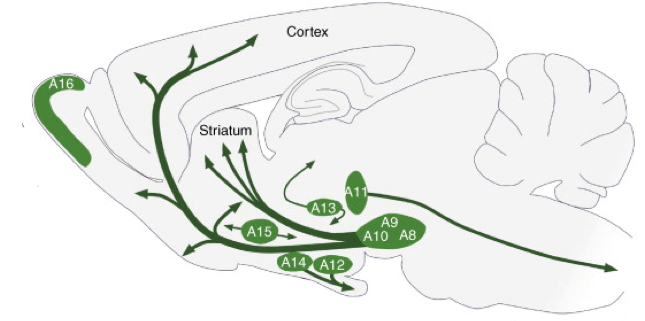
Die rol van dopamienneurone as rekenaars van voorspellingsfoute bly 'n fassinerende en waardige navorsingslyn, maar as beloningvoorspellingsfout ALLE is wat dopamienneurone doen, wat het ons dan nodig vir 400,000-600,000? * Hier is 'n kaart van waar die brein se dopamienneurone is geleë (in 'n dwarssnit van 'n knaagdierbrein):
Verspreiding van dopamienneuronselgroepe A8-A16 in die volwasse knaagdierbrein. Aangepas uit Björklund, A. & Dunnett, SB Dopamine neuronstelsels in die brein: 'n opdatering. Tendense in neurowetenskappe 30, 194–202 (2007).
* By mense Daar is 160,000-320,000 in ape en net 20,000-45,000 in knaagdiere.
As ons na hierdie diagram kyk, lyk dit asof daar 'n paar bruto anatomiese onderskeidings tussen groepe dopamienneurone is, en daarom word hulle A8-A16 gemerk. Daar is ook fyner anatomiese onderskeidings, wat nie-so-subtiele funksionele implikasies het. In die eerste studiegids sal ek hier fokus, Lammel et al. stel oor die onderskeid tussen dopamienneurone in die ventrale tegmentale area (VTA, of A10 in die bostaande foto) deur hul konneksie met ander breinareas. Lammel et al. het opgemerk dat daar ten minste twee skeibare bevolkings van dopamienneurone in die VTA is. Een populasie kry insetseine vanaf 'n breinarea genaamd die laterodorsale tegmentum en stuur uitsetseine na 'n breinarea genaamd die nucleus accumbens (noem hierdie LDT-dopamien-NAc neurone). Die ander bevolking kry insette van die laterale habenula en stuur uitsette na die prefrontale korteks (noem hierdie LHb-dopamien-PFC neurone). So wat? Word die feit dat hierdie dopamienneurone in verskillende breinkringe bedraad is, glad vir gedrag? Lammel et al. het getoon dat dit saak maak. Wanneer (gebruik optogenetics!) Hulle het die insette aan LDT-dopamien-NAc neurone in muise geaktiveer. Hulle het gevind dat die diere positiewe assosiasies gevorm het met die konteks waarin hulle gestimuleer is. Hulle het verkies om meer tyd in die deel van 'n boks te spandeer, waar hulle die breinstimulasie gekry het. In teenstelling hiermee, wanneer Lammel et al. het die insette aan LHb-dopamien-PFC neurone geaktiveer, die presiese teenoorgestelde is waargeneem. Diere vermy 'n deel van 'n boks waar hulle die stimulasie gekry het. In 'n ander studie deur dieselfde groep, toe die muise van nature iets goed of iets sleg beleef het, is die sterkte van hierdie afsonderlike stroombane differensieel gemoduleer. Muise wat kokaïen gegee het, het 'n verhoogde sterkte van die LDT-dopamien-NAc-pad getoon, maar geen verandering in die LHb-dopamien-PFC-pad nie. Muise wat 'n irritasie op hul poot gegee het, het geen verandering in die LDT-dopamien-NAc-pad gehad nie, maar 'n verhoogde sterkte van die LHb-dopamien-PFC-pad.
Ondermyn Schultz et al. Se aanvanklike bewering dat dopamienneurone homogeen is, Lammel et al. ontdek dat hulle nie is nie. Hierdie hersiening het waarskynlik plaasgevind as gevolg van die toenemende sensitiwiteit van die beskikbare gereedskap, wat redelik verander het van die 1990s na die 2010s. Nuwer en beter gereedskap, in kombinasie met 'n bietjie kreatiwiteit, het Lammel et al toegelaat. om subtiliteite te onderskei wat nie vir Schultz et al toeganklik was nie. In die onthulling van hierdie subtiliteite, Lammel et al. het gehelp om die hubris te demonstreer om te glo dat jy 'n hele klas neurone uitgevind het omdat jy antwoorde in 55-80% van 'n bevolking sien, veral as jy nie heeltemal seker is of nie Ek het daardie populasie gedefinieer. (Die vraag van die definisie van dopamienneurone tydens in vivo neurale opnames is 'n HELE ander probleem). Al die krediet in die wêreld aan Schultz et al. vir die verligting van die vuur van dopamiennavorsing, maar dit was meer van 'n beginpunt as 'n eindpunt.
Groepering van neurone deur die breinbane waarin hulle deelneem, maak 'n klomp sin as jy probeer uitvind hoe breinbane werk. Maar wat as jy die dopamien-gedeelte van dopamienneurone probeer uitvind? Die meeste dopamienneuronnavorsing het aanvaar dat wanneer 'n dopamienneuron brande, dit die neurotransmitter dopamien vrystel, 'n klein molekule wat so lyk:
Trouens, dit is hoe ons "dopamienneuron" omskryf het. Maar soos die wetenskap dikwels voorkom, lyk dit nie so maklik nie. In die tweede lyn van onlangse navorsing sal ek hier bespreek, wetenskaplikes het bewyse getoon dat dopamienneurone ander neurotransmittermolekules, glutamaat en GABA, saam met vrystelling van dopamien kan mede vrylating.
Eintlik sal verskillende subgroepe van dopamienneurone waarskynlik oorwegend glutamaat of GABA vrystel. Studies by Hnasko et al. en Stuber et al. het gedemonstreer dat dopamienneurone in die VTA mede-vrystelling glutamaat. Eerstens het hulle opgemerk dat baie VTA-dopamienneurone 'n glutamaatvervoerder uitbeeld, naamlik VGLUT2, 'n proteïen wat glutamaat verpak vir vrystelling van neurone. Het die teenwoordigheid van VGLUT2 beteken dat dopamienneurone in vergelyking met dopamien glutamaat verpak? Om na hierdie vraag te kyk, het die wetenskaplikes gekyk na die response van neurone in die nukleusakkapels (een plek waar dopamiene neurone uitsette na, die bespreking van Lammel et al. Hierbo) na dopamienneuronstimulasie sien. Inderdaad, hulle waargeneem vinnige, opwindende antwoorde van die kern pasiënte neurone stimuleer van VTA dopamienneurone van 'n tipe wat in ooreenstemming sou wees met 'n glutamatergiese eerder as 'n dopaminerge reaksie. Hierdie response is geblokkeer deur antagoniste van glutamaatreseptore, maar nie deur antagoniste van dopamienreseptore nie. Verder, in muise wat geneties gemanipuleer is om VGLUT2 in dopamienneurone te ontbreek, is geen sulke antwoorde gesien nie.
Die mede-vrystelling van glutamaat mag egter nie in alle dopamienneurone voorkom nie. Soos in Lammel et al. Se studies raak verbindings. Stuber et al. het opgemerk dat dopamienneurone in 'n naburige area genaamd die substantia nigra (A9), wat uitsette na die dorsale striatum stuur, nie bewys van glutamaat vrystelling toon nie. Die negatiewe uitslag is nog kontroversieel. Nog 'n groep, Tritsch et al., het bewyse van glutamaat vrystelling deur substantia nigra dopamienneurone waargeneem. Daarbenewens het hulle getoon dat hierdie substantia nigra dopamienneurone ook nog 'n neurotransmitter mede vrylaat: GABA. Vreemd genoeg, substantia nigra dopamienneurone druk egter nie VGAT, die normale GABA-vervoerder uit nie. In plaas daarvan, Tritsch et al. het bevind dat VMAT, die dopamien-vervoerder, ook GABA kan vervoer, dit verpak vir sinaptiese vrystelling saam met dopamien. Tritsch et al. Se bevinding kan veralgemeen as substantia nigra dopamienneurone. Solank as wat daar 'n paar GABA is, kan enige iets wat VMAT uitdruk, moontlik ook die GABA pak en loslaat. Een sleutelvraag wat uit Tritsch et al se studie voorkom, is presies waar en wanneer die GABA in die substantia nigra gesintetiseer word. Tog is dit daar.
Die implikasies van glutamaat en GABA mede-vrystelling van dopamienneurone bly vir die grootste deel nog te sien. Die enigste gerapporteerde gedragseffek kom uit die Hnasko et al. papier. Hulle wys dat muise wat VGLUT2 in dopamienneurone ontbreek, minder rondloop in reaksies op kokaïen as normale muise. Dis nou al. As niks anders nie, toon dit hoeveel meer ons moet leer oor die verskynsel van sender-mede-vrylating.
Tot dusver het ons gesien dat dopamienneurone verskillende dinge kan aandui as hulle in verskillende breinbane verbind word, en dat hulle hul toegewysde rol in 'n breinbaan ten minste gedeeltelik kan gebruik deur ander chemikalieë as net dopamien. In die derde lyn van navorsing sal ek hier ondersoek, ons sal nog 'n laag baie komplekse kompleksiteit by die prentjie voeg: dopamienneurone kan die manier verander waarop hulle aan 'n breinbaan deelneem deur te verander of hulle maak of nie vrystelling van dopamien. In hierdie geval, Dulcis et al. kyk na 'n effens verskillende groep dopamienneurone van dié waaroor ek nou gepraat het, in die hipotalamus. Hulle het opgemerk dat die aantal dopamienneurone by rotte skynbaar fluktueer met die lengte van die "daglig" wat die rotte ervaar. Ek sit daglicht in aanhalingstekens omdat dit nie werklike daglig is nie - net of die ligte in 'n baie beheerde laboratorium-omgewing is of nie. Die meeste laboratoriumdiere sien 12 uur se lig per dag, maar Dulcis et al. het ook net 5 uur per dag of tot 19 probeer. Rotte wat lang dae ervaar het, het minder dopamienneurone in hul hipotalamus gehad, terwyl rotte wat kort dae ervaar het, meer gehad het. By verdere ondersoek het hulle vasgestel dat die veranderinge in die aantal dopamienneurone in die verskillende ligtoestande nie te wyte was aan neurone wat doodgaan en gebore word nie. Dieselfde neurone was eintlik daar in alle omstandighede, maar hulle het hul dopamien-nes aan of af geskakel. Dit is nog nie duidelik waarom lig blootstelling hierdie veranderings veroorsaak of wat die presiese gedragsgevolge is nie. Rotte wat lang dae gehad het, en minder dopamienneurone het gevolglik depressiewe en angstige gedrag getoon (hou in gedagte dat rotte nag is en verkies die donker). So het rotte wie se hipotalamiese dopamienneurone met 'n gifstowwe doodgemaak is. As die dopamienneurone egter met 'n toksien doodgemaak word terwyl die rotte 12 uur lig per dag kry en dan is die rotte slegs 5 uur lig per dag gegee, is voorheen nie-dopaminerge neurone gewerf om dopamien vry te laat en minder depressief en angstig gedrag is waargeneem. Pretty cool! En belangriker, hierdie werk demonstreer dat neurone ons nie eers voorheen geïdentifiseer het nie, aangesien dopamienneurone onder die regte omstandighede kan transformeer. Sommige aspekte van ons brein is gebou om stabiel te wees, maar baie verander al die tyd, wat ons toelaat om te internaliseer en aan te pas by ons ervaring.
Na al hierdie studies, wat het ons geleer? Vir my is die groot prentjie wegneem dat die begrip van die brein beteken om kompleksiteit te waardeer. Om effens meer spesifiek te wees, beteken dit om molekules en selle te verbind met stroombane en gedrag om definisies van biologiese entiteite te verskaf wat die modaliteite van studie dek. Nie meer groepeer neurone uitsluitlik deur een neurotransmitter wat hulle kan vrystel nie. Hierdie groepering kan soms nog tersaaklik wees, maar soos ons in die bogenoemde studies gesien het, nie altyd nie. Om te dink aan die herdefiniëring van die groep wat formeel bekend staan as dopamienneurone, moet ons ook terugkyk na die dekades van vorige literatuur met die perspektief wat na vore kom. Dit is nie dat die data in ouer dopamienneuronstudies verkeerd is nie, maar gevolgtrekkings is dalk nie heeltemal wat ons gedink het hulle was nie. Dit kan 'n goeie ding wees. Baie arcane argumente oor presies wat dopamienneurone enkodeer, kan eintlik uiteindelik opgelos word deur te verstaan dat hulle baie verskillende dinge in verskillende kontekste doen. Moenie bekommerd wees nie: dit mag dalk verwarrend lyk, maar dit is die baie normale proses van wetenskapsveroudering. Nie net is die proses normaal nie, dit is absoluut noodsaaklik. Wetenskaplikes moet voortdurend ons definisies bevraagteken en hersien om beduidende konseptuele vooruitgang te weerspieël.
Definisies kan verwarrend wees. Hulle kan ook taamlik vervelig wees, en ek is bekommerd dat hulle te dikwels mense van die wetenskap af wegjaag. Toe ek 'n beginbiologie student was, het ek ure spandeer om flitskaarte te maak om my te help memoriseer wat soos eindelose definisies gelyk het. Ek het dit beskou as 'n vervelige maar noodsaaklike inisiatief vir die bioloogsklub. Eintlik het ek self gesê dat ek die woordeskat moes leer om hoër orde kwessies met werkende wetenskaplikes te kan bespreek. Wat ek waardeer het soos wat ek verder gegaan het in my loopbaan, is hoe genuanseerde diegene wat skynbaar swart-en-wit, reg of verkeerde definisies is - hoeveel subtiliteit en geskiedenis hulle verpak het. Wetenskaplike definisies, soos die definisie van 'n dopamienneuron, gee nie net 'n gemeenskaplike taal nie; hulle struktureer die aard van ons ondersoeke. Ons benodig hierdie struktuur om voort te gaan met ons eksperimente, maar ons moet ook bewus wees van die maniere waarop hierdie definisies ons kan beperk. Ons vergelyk gedefinieerde groepe met mekaar. Ons praat oor groepgemiddeldes. So presies watter dinge in ons groepe ingesluit word, kan dramaties beïnvloed hoe ons data lyk en wat ons besluit, beteken dit. So, ons moet altyd bewus wees van die vooroordele inherent in ons kategorisering. Miskien is definisies nie so saai nie! Besprekings van hierdie voorbehoudsbepalings kan 'n bietjie meer intro kursusmateriaal oplewer terwyl studente leer hoe om te dink soos ware wetenskaplikes.
Die besondere vraag om neuronale seltipes te definieer, blyk redelik tydig te wees. Net 'n paar weke gelede, die eerste tussentydse verslag van die BRAIN-inisiatief werkgroep het uitgekom (sien ook Astra Bryant's pos oor die onderwerp). Daarin word nege hoë prioriteitsnavorsingsareas vir FY 2014 uiteengesit. Die eerste hiervan is 'n sensus van seltipes. 'Die verslag erken die probleme wat ek hier bespreek het:
Daar is nog nie 'n konsensus oor wat 'n neuronale tipe is nie, aangesien 'n verskeidenheid faktore insluitende ondervinding, konnektiwiteit en neuromodulators die molekulêre, elektriese en strukturele eienskappe van aanvanklik soortgelyke neurone kan diversifiseer. In sommige gevalle mag daar nie eens skerp grense wees wat subtipes van mekaar skei nie. Nietemin is daar algemene ooreenstemming dat tipes voorlopig bepaal kan word deur invariante en algemeen intrinsieke eienskappe, en dat hierdie klassifikasie 'n goeie beginpunt vir 'n sensus kan bied. So, die sensus moet begin met goed beskryf groot klasse neurone (bv. Opwindende piramidale neurone van die korteks) en gaan dan voort na fyner kategorieë binne hierdie klassifikasies. Hierdie sensus sal geneem word met die wete dat dit aanvanklik onvolledig sal wees en oor iterasies sal verbeter.
Die antwoord op die vraag "Wat is 'n dopamienneuron?" Kom nie heeltemal voor nie, maar 'n hoë profiel-herkenning van die vraag en die befondsing wat daarop volg, is 'n belangrike eerste stap. Jammer daaroor.
