Front. Behav. Neurosci., 23 Maart 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Translasionele Navorsingsinstituut en Instituut vir Gesondheid en Biomediese Innovasie, Queensland Universiteit van Tegnologie, Brisbane, QLD, Australië
- 2Skool vir Biomediese Wetenskappe, die Universiteit van Queensland, Brisbane, QLD, Australië
Die moderne dieet het hoogs versoet geword, wat lei tot ongekende vlakke van suikerverbruik, veral onder adolessente. Terwyl chroniese langtermyn suiker inname bekend is om by te dra tot die ontwikkeling van metaboliese afwykings, insluitend obesiteit en tipe II diabetes, is min bekend met betrekking tot die direkte gevolge van langtermyn, binge-like suikerverbruik op die brein. BOmdat suiker die vrystelling van dopamien in die nucleus accumbens (NAc) kan veroorsaak, is dit soortgelyk aan misbruikmiddels. Ons ondersoek veranderings in die morfologie van neurone in hierdie brein streek na kort- (4 weke) en langtermyn (12 weke) soos sukroseverbruik met behulp van 'n intermitterende twee-bottel keuseparadigma. Ons het Golgi-Cox-kleuring gebruik om medium-spinyneurone (MSNs) van die NAc-kern en dop van kort- en langtermyn-sukroseverbruikende rotte te bevrug en vergelyk dit met die ouderdom-pasgemaakte waterkontroles. Ons toon dat langdurige binge-agtige sukroseverbruik die totale dendritiese lengte van NAc-skulp MSN's aansienlik verlaag het in vergelyking met die ouderdomstemperaat-beheerratte. Ons het ook bevind dat die herstrukturering van hierdie neurone hoofsaaklik veroorsaak is deur verminderde distale dendritiese kompleksiteit. Daarenteen het ons verhoogde ruggraatdigtheid by die distale takbestellings van NAc-skulp MSNs van langtermyn-sukroseverbruikende rotte waargeneem. Gekombineer, hierdie resultate beklemtoon die neuronale effekte van langdurige binge-like inname van sukrose op NAc shell MSN morfologie.
Inleiding
Oor die afgelope 40 jaar is daar 'n gedokumenteerde styging in die verbruik van suikerverzoete drank en kosse wat bygevoegde suikers bevat (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), met verslae wat vermoed dat tot en met 75% van alle kosse en drankse hoë hoeveelhede toegevoegde suikers bevat (Ford en Dietz, 2013; Bray en Popkin, 2014). Gedurende hierdie tydperk was daar ook 'n gelyktydige toename in die voorkoms van vetsug en tipe II diabetes, veral onder adolessente (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Onlangse studies het getoon dat oorgewig en vetsugtige kinders dikwels hoë hoeveelhede toegevoegde suiker verbruik, maar die bydrae van hoë suiker wat dieet bevat aan die verhoogde voorkoms van oorgewig en vetsugtige kinders bly omstrede (Hu, 2013; Bray en Popkin, 2014; Bucher Della Torre et al., 2015).
Terwyl 'n groeiende bewyse dui daarop dat die verbruik van hoë suikerdieet gedeeltelik kan bydra tot gewigstoename by kinders en adolessente (Malik et al., 2010; Te Morenga et al., 2013; Bray en Popkin, 2014), is minder aandag gegee aan nadelige nie-metaboliese gevolge wat voortspruit uit oormatige suiker inname. Interessant genoeg, kom 'n paar algemene gedrags- en sielkundige patrone voor onder 'n deel van diegene wat diëte wat suiker bevat en in stand hou. Die mees opvallende is die ontwikkeling van eetversteurings, insluitende binge-eet, gekombineer met die gelyktydige aanvang van sielkundige simptome, insluitende gebrek aan motivering en depressie (hersien in Sheehan en Herman, 2015). Daarbenewens omdat binge-eetende individue dikwels 'n verlies aan beheer toon en 'n onvermoë om hul suiker inname te beperk, is dit waarskynlik dat hierdie gedrag ontstaan as gevolg van neurologiese aanpassings in breinstreke wat die hedoniese waarde van hoogs smaaklike kos evalueer (Saper et al., 2002; Lutter en Nestler, 2009; Kenny, 2011). Hierdie rasionaal word ook ondersteun deur bewyse in die mens wat aantoon dat suiker en soetheid dringend kan wees wat soortgelyk is aan dié wat deur verslawende middels soos alkohol en nikotien veroorsaak word (Volkow et al., 2012).
Alhoewel die verslawende eienskappe van suiker steeds spekulatief is, word hierdie waarnemings gekombineer met studies wat die bydrae toon van oormatige suiker inname tot veranderinge in beloningskringe en die ontwikkeling van verslawende gedrag en emosionele toestande in diermodelle (Avena et al., 2008; Benton, 2010; Ventura et al., 2014), waarborg die behoefte aan verdere ondersoek. Vorige studies in knaagdiere het getoon dat intermitterende toegang tot sukrose die aktiwiteit van verskeie neurotransmitters binne die mesolimbiese stelsel insluitend dopamien, opioïede en asetielcholien verander. Avena et al., 2008). Binge-like verbruik van sukrose het getoon dat dopamien vrylating in die nucleus accumbens (NAc), soortgelyk aan dwelms van misbruik, fasiliteer (Avena et al., 2008). Verder het ons getoon dat langtermynverbruik van sukrose met behulp van 'n 24 h intermitterende toegang twee-bottel keuseparadigma (Simms et al., 2008) modulateer nikotiniese asetielcholien reseptor (nAChR) uitdrukking in die NAc (Shariff et al., In pers). Interessant genoeg het ons ook opgemerk dat nAChR-verbindings bekend is om dopamien- en asetielcholienaktiwiteit in die NAc te moduleer, het verskillende effekte op sukroseverbruik na kort- en langtermyn inname (Shariff et al., In pers).
Alhoewel hierdie studies ooreenkomste getoon het in die gedrags- en neurochemiese veranderinge wat veroorsaak word deur intermitterende toegang tot suiker en dwelmmiddels, is dit nie bekend of hierdie effekte die veranderinge in neuronale morfologie in die NAc fasiliteer nie. THy is in teenstelling met misbruikstowwe, insluitende kokaïen, amfetamien en nikotien, wat goed gekenmerk word deur veranderinge in die morfologie van medium-spierneurone (MSN's) in die NAc, insluitende verhoogde ruggraatdigtheid en veranderde dendritiese kompleksiteit (Robinson en Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Aangesien ons voorheen getoon het dat die langtermynblootstelling (12week) na alkohol en sukrose met die intermitterende twee-bottelkeuse-paradigma is, lewer dit 'n differensiële respons op farmakoterapeutiese intervensies in vergelyking met korttermyn inname (4 weke; Steensland et al., 2007; Shariff et al., In pers), het ons die gevolge van kort- en langtermyn-sukroseverbruik op MSN-morfologie in die NAc beoordeel. Ons het toegelaat dat adolessente rotte sukrose op 'n binge-manier gebruik vir 4 (korttermyn) of 12 (langtermyn) weke en dan die morfologie van NAc MSNs van kort- en langtermyn-sukroseverbruikende rotte ontleed en vergelyk dit met ouderdom-pas beheermaatreëls wat net toegang tot water gegee word. Ons resultate toon dat MSN's van die NAc-dop verander word na lang- maar nie korttermyn-sukroseverbruik nie, met dendritiese lengte, maar verhoogde distale dendritiese ruggraatdigtheid. Verder het ons gevind dat die morfologie van MSNs van die NAc-kern relatief ongeskonde gebly het na kort- en langtermyn-sukroseverbruik. Hierdie resultate beklemtoon 'n direkte neurologiese gevolg van langtermyn-sukroseverbruik op 'n binge-agtige wyse. Daarbenewens demonstreer hierdie data die behoefte aan verdere studies wat daarop gemik is om die molekulêre en neurochemiese veranderinge wat die morfologiese herstrukturering van NAc-skulp MSN's vergesel, deur langdurige bingeagtige sukrose-inname te vergemaklik.
Materiaal en metodes
Etiekverklaring
Alle eksperimentele prosedures is uitgevoer in ooreenstemming met die Australiese Kode vir die Sorg en Gebruik van Diere vir Wetenskaplike Doeleindes, 8th Uitgawe (Nasionale Gesondheids- en Mediese Navorsingsraad, 2013). Die protokolle is goedgekeur deur die Animal Ethics Committee van die Queensland Universiteit van Tegnologie en die Animal Ethics Committee van die Universiteit van Queensland.
Diere en Behuising
Vyf weke oue (adolessente) manlike Wistar rotte (Beheer: 176.4 ± 4.8 g; Sukrose: 178.3 ± 5.0 g) (LNR, WA, Australië), is individueel gehuisves in geventileerde dubbelvlakpleksiglas® hokke. Die rotte is geklimatiseer met die individuele behuisingsomstandighede, hantering en omkering-ligsiklus 5 dae voor die aanvang van die eksperimente. Alle rotte is gehuisves in 'n klimaat-beheerde 12-uur omkering lig / donker siklus (ligte af by 9 am) kamer met standaard rat chow en water beskikbaar ad libitum.
Intermitterende Toegang Twee-Bottle Choice Drink Paradigma
Die intermitterende toegang 5% sukrose twee-bottel keuse drink paradigma (Simms et al., 2008) is aangepas vanaf Wys (1973). Alle vloeistowwe is aangebied in 300 ml-gegradueerde plastiekbottels met vlekvrye staal-drinkstowwe wat deur die aanvang van die donker ligsiklus deur twee grommets in die voorkant van die hok aangebring is. Gewigte van elke bottel is aangeteken voor bottelaanbieding. Twee bottels is gelyktydig aangebied: een bottel wat water bevat; die tweede bottel met 5% (w / v) sukrose. Die plasing van die 5% (w / v) sukrose-bottel is afgewissel met elke blootstelling om beheer vir syvoorkeure te beheer. Flesse is 24 h geweeg nadat die vloeistowwe aangebied is, en metings is na die naaste 0.1 g geneem. Die gewig van elke rot is ook gemeet om die gram sukrose inname per kilogram liggaamsgewig te bereken. Op dag 1 van die drinkperiode, het rotte (n = 6-9) is toegang gegee tot een bottel 5% (w / v) sukrose en een bottel water. Na 24 h is die sukrose bottel vervang met 'n tweede waterbottel wat beskikbaar was vir die volgende 24 h. Hierdie patroon is op Woensdae en Vrydae herhaal. Die rotte het op alle ander dae onbeperkte toegang tot water gehad. Binge-like verbruik van sukrose het gelei tot 'n toename in totale sukrose inname (ml) oor tyd (Aanvullende Figuur 1) en is gepaard gegaan met stabiele baseline drinkvlakke gebaseer op liggaamsgewig [20 ± 5 g / kg van die 5% (w / v)] gedurende die korttermyn [4 weke (13 drink sessies)] en langtermyn [ ~ 12 weke (37 drink sessies)] drinkperiodes. 'N Aparte groep beheerratte (n = 6-9) was onder dieselfde toestande soos hierbo beskryf, toegang tot water in beide bottels (dws geen sukrose). Die gemiddelde liggaamsgewig van beheer en sukrose wat rotte verbruik het aan die einde van korttermynblootstelling was onderskeidelik 405.7 ± 40.8 g en 426.4 ± 31.2 g. Aan die einde van langtermynblootstelling was die gemiddelde liggaamsgewig vir beheer- en sukrosegroepe 578.8 ± 53.4 g en 600.2 ± 45.2 g.
Golgi-Cox Staining
Na die laaste drink sessie is rotte vanaf die dierefasiliteit oorgedra om die breinmonsters by die histologiefasiliteit van die Skool vir Biomediese Wetenskappe, Universiteit van Queensland (St Lucia, Australië), toe te pas. Alle goedgekeurde maatreëls is geneem om stres tydens vervoer te verminder, waarna rotte toegelaat word om oornag te herstel. Die volgende dag is rotte geoffer deur natriumpentobarbital oordosis (60-80 mg / kg, ip Vetcare, Brisbane, Australië) en intracardiaal perfuse met ~ 300 ml kunsmatige serebro-spinale vloeistof wat daarin vervat is, (in mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, en 10 D-glukose. Elke dier is dan gedepapiteer en die brein verwyder en in die donker ingesluit in Golgi-Cox-oplossing wat 5% kaliumdichromaat, 5% kaliumchromaat en 5% mercuric chloride bevat (alle chemikalieë van Sigma-Aldrich) wat vars 3 dae gemaak is. voor opoffering soos hierbo beskryf (Rutledge et al., 1969). Golgi-Cox vlek inkubasie en na-verwerking metodes is verander van Ranjan en Mallick (2010). Hersiene van korttermyn-sukroseverbruikende diere is vir 6-dae by 37 ° C geïnkubineer, terwyl die brein van langtermyn-sukroseverbruikende diere vir 10-dae geïnkubeer is, met een verandering na vars Golgi-Cox-oplossing na 4-dae van inkubasie.
Na inkubasie is 300 μm koronale snitte gesny met 'n vibrerende Zeiss Hyrax V50 mikrotoom (Carl Zeiss, Duitsland). Snye is dan opeenvolgend in 24-putplate geplaas met 30% (w / v) sukrose in 0.1 M-fosfaatgebufferde sout en geprosesseer soos uiteengesit in (Ranjan en Mallick, 2010). Kortliks is gedeeltes gedehidreer in 50% etanol vir 5 min, dan in 0.1 M NH geplaas.4OH oplossing vir 30 min, twee keer met gedistilleerde water vir 5 min gesuiwer en in Fujihunt film fixer (Fujifilm, Singapoer) vir 30 min in die donker geplaas. Die snye is dan twee keer in gedistilleerde water vir 2 min gesuiwer en elk twee keer in 70, 90, 95 en 100% etanol vir 5 min elk. Die afdelings is dan in CXA-oplossing (1: 1: 1 chloroform: xyleen: alkohol) vir 10 min skoongemaak en in DPX (Sigma-Aldrich) op Superfrost Plus-skyfies (Menzel-Glaser, Lomb Scientific, Australië) gemonteer (Menzel-Glaser, Duitsland). Die skyfies is in die donker gelaat om oornag by kamertemperatuur te droog.
Neuronale Keuring en Opsporing binne die Nucleus Accumbens
Koronale snye tussen bregma + 2.8 en + 1.7 is ondersoek vir MSN's binne die kern en dop van die NAc, met behulp van die laterale ventrikel en die anterior kommissie as landmerke met behulp van 'n rotbreinatlas (Paxinos en Watson, 2007) (Figuur 1). Die kontoerfunksie in Neurolucida 7 (MBF Bioscience, VT, VSA) is gebruik om die NAc-kern en NAc-dop in elke sny af te sny (Figuur 2). Tussen 2 en 9 neurone per streek per dier is opgespoor vir dendritiese lengte parameters met behulp van 'n 63x doel of vir ruggraatdigtheid (aangegee as stekels per 100 μm) met 'n 100x doel op 'n Zeiss Axioskop II (Carl Zeiss, Duitsland) met behulp van 'n outomatiese xyz verhoog gedryf deur Neurolucida® 7 sagteware (MBF Biosciences, VT, VSA). Alle opsporing is blindelings uitgevoer ten opsigte van behandeling. Morfologiese parameters van Golgi-Cox geïmpregneerde neurone is geanaliseer op 'n wyse soortgelyk aan vorige verslae (Klenowski et al., 2015).
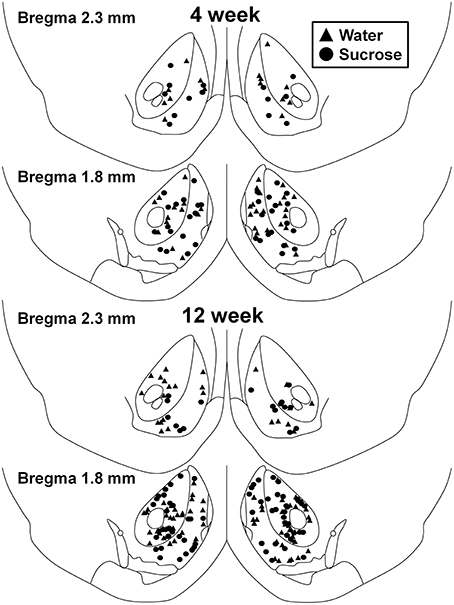
Figuur 1. Kaart wat liggings toon van middelpuntige spierneurone wat uit die kernklem en die dop van 4- en 12-week sukrose wat rotte verbruik, en ouderdom-gekontroleerde kontrole. Top twee panele wys plekke van neurone wat uit die kernklem en die dop van 4 weekbeheer (driehoeke) en sukrose (sirkels) diere getoets word. Onderste twee panele toon posisies van neurone wat uit 12 weekbeheer (driehoeke) en sukrose (sirkels) diere getoets word.
Statistiese analise
Gemiddelde en standaard fout van die gemiddelde (SEM) is bereken vir elke datastel met die dier as n, met behulp van die gemiddelde morfometrie data van al die kern of dop NAc MSNs (n = 7 vir NAc dop en n = 6 vir NAc kern 4-week, n = 9 vir groepe van 12 weke). Waar aangedui, ongepaarde twee stert studente ttoetse of tweerigting-ANOVA's met Bonferroni-na-toetse is uitgevoer vir alle ontledings wat die vergelyking van groep beteken, met behulp van GraphPad Prism-weergawe 6.02 (GraphPad Software, San Diego, CA). Statistiese betekenisvolheid is aanvaar by P <0.05. Alle gegewens in die resultate word aangebied as gemiddeld ± SEM. Persentasieveranderings word bereken as relatief tot die kontrolewaarde.
Results
Medium Spiny Neurons van die Nucleus Accumbens Shell het afgeneem Dendritiese Lengte, verminderde Dendritiese Kompleksiteit, maar verhoogde gemiddelde ruggraatdigtheid by Distale Takorders Volg Lang- maar nie Korttermyn Sukroseverbruik nie.
Na korttermyn (4 weke) sukroseverbruik was daar geen beduidende verskille in NAc-skulp MSN morfometriese parameters (Table 1). Daar was ook geen beduidende verskille tussen korttermyn-sukroseverbruik en waterbeheer nie. NAc-skulp MSN's in ontledings met betrekking tot sentrifugale takorder. Namens dendritiese segmente per takorde (P = 0.4111), gemiddelde dendritiese lengte per takorde (P = 0.5581) en gemiddelde ruggraatdigtheid per takorde (P = 0.2977, tweerigting-ANOVA's) was nie beduidend verskillend tussen groepe nie. 'N Plekkaart wat die benaderde posisies van die gesamploneerde neurone toon, word in Figuur getoon 1.
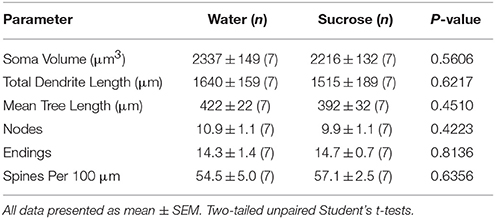
Tabel 1. Algemene morfologiese parameters van medium-stekelneurone uit die nukleusakkapsel-dop van korttermyn-sukroseverbruikende rotte en ouderdom-gekontroleerde waterkontroles.
Na aanleiding van langtermyn (12 weke) sukroseverbruik, is die totale dendritiese booglengte van NAc-skulp MSN's met 21% verlaag in vergelyking met waterverbruikende beheermaatreëls (Water: 1827 ± 148 μm, n = 9; Sukrose 1449 ± 78 μm, n = 9, *P = 0.0384, tweestert-ongepaarde studente t-toets, Figuur 2, Tabel 2). Vergelyking van die gemiddelde aantal dendritiese bifurkasies (nodusse) en dendritiese eindpunte tussen die water- en sukrosegroepe het 'n verminderde (hoewel nie beduidende) vlak van dendritiese kompleksiteit in NAc-skulp MSN's (nodusse: Water 12.9 ± 1.4 n = 9, Sukrose 10.1 ± 0.8 n = 9, P = 0.0879; eindpunte: Water 17.9 ± 1.4 n = 9, Sukrose 14.8 ± 0.7 n = 9, P = 0.0657, tweestert-ongepaarde studente ttoets, tabel 2). Daar was geen verandering in soma volume nie (P = 0.9400), gemiddelde dendritiese boomlengte (P = 0.1646) of totale ruggraatdigtheid (P = 0.3662) in NAc-skulp MSN's van langtermyn-sukroseverbruikende rotte in vergelyking met waterbeheer. Hierdie morfometriese parameters word in Tabel uiteengesit 2.
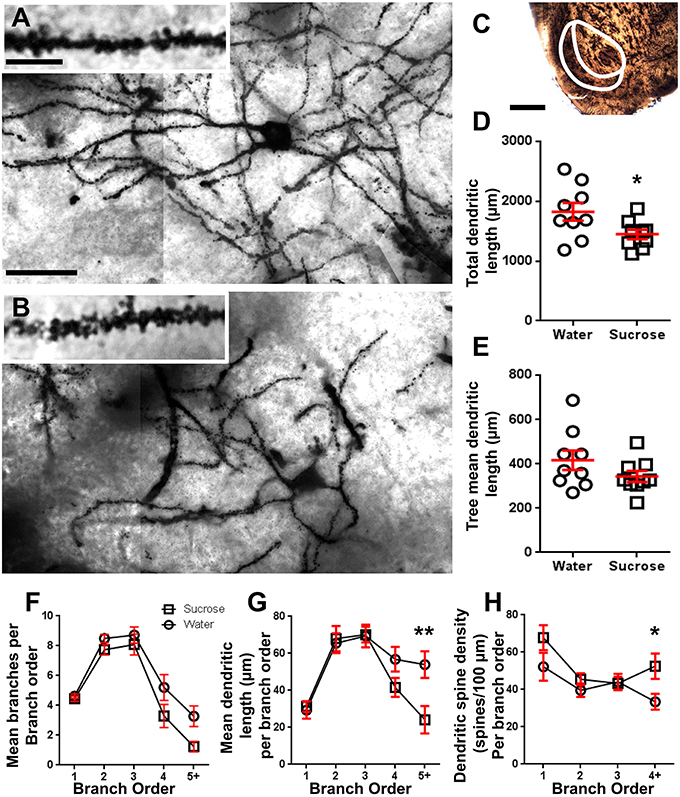 Figuur 2. Verminderde dendritiese booglengte en verhoogde distale dendritiese ruggraatdigtheid van medium-stekelneurone (MSN's) van die Nucleus Accumbens (NAc) -dop van langtermyn-Suksrose-behandelde rotte in vergelyking met beheermaatjies. (A, B) toon voorstellings van beheer (boonste) en langtermyn (12 week) sukrose (onderste) behandelde helderveld z-stapmosaïek van Golgi-geïmpregneerde MSN's van die NAc-dop (63x-vergroting). Inset van (A, B) toon beheer en langtermyn-sukrose-behandelde helderfield beelde van Golgi-geïmpregneerde MSN dendriete en dendritiese stekels uit die NAc-dop (100x vergroting). (C) toon die anatomiese streke waaruit MSN's in hierdie studie geneem is. (D) toon 'n verspreidingsgrafiek van verlaagde totale MSN dendritiese arbor (gemiddelde ± SEM) van die NAc-dop in langtermyn-sukrose-diere (blokkies) in vergelyking met kontroles (sirkels), ongepaarde studente t-toets, *P <0.05, n = 9; beheer en n = 9; 12 week sukrose. (E) toon 'n verspreidingsgrafiek van onveranderde gemiddelde MSN dendritiese boomlengte (gemiddelde ± SEM) van die NAc-dop in langtermyn-sukrose-diere (blokkies) in vergelyking met kontroles (sirkels), ongepaarde studente tToets, P > 0.05, n = 9; beheer en n = 9; 12 week sukrose. Takorde-analise (gemiddelde ± SEM) van dendritiese segmentnommer per takorder (F), gemiddelde dendritiese lengte per takorder (G) en dendritiese ruggraatdigtheid per takorder (H). Langtermyn-sukroseverbruik het dendritiese lengte by distale takordes (5 +) en verhoogde dendritiese ruggraatdigtheid by distale takorders (4 +) vergeleke met kontroles (G, H), tweerigting-ANOVA's met Bonferroni-na-toetse, *P <0.05, **P <0.01, n = 9; beheer en n = 9; langtermyn sukrose. Skaalbalke: (A, B) = 20 μm; inset van (A, B) = 10 μm; (C) = 1 mm.
Figuur 2. Verminderde dendritiese booglengte en verhoogde distale dendritiese ruggraatdigtheid van medium-stekelneurone (MSN's) van die Nucleus Accumbens (NAc) -dop van langtermyn-Suksrose-behandelde rotte in vergelyking met beheermaatjies. (A, B) toon voorstellings van beheer (boonste) en langtermyn (12 week) sukrose (onderste) behandelde helderveld z-stapmosaïek van Golgi-geïmpregneerde MSN's van die NAc-dop (63x-vergroting). Inset van (A, B) toon beheer en langtermyn-sukrose-behandelde helderfield beelde van Golgi-geïmpregneerde MSN dendriete en dendritiese stekels uit die NAc-dop (100x vergroting). (C) toon die anatomiese streke waaruit MSN's in hierdie studie geneem is. (D) toon 'n verspreidingsgrafiek van verlaagde totale MSN dendritiese arbor (gemiddelde ± SEM) van die NAc-dop in langtermyn-sukrose-diere (blokkies) in vergelyking met kontroles (sirkels), ongepaarde studente t-toets, *P <0.05, n = 9; beheer en n = 9; 12 week sukrose. (E) toon 'n verspreidingsgrafiek van onveranderde gemiddelde MSN dendritiese boomlengte (gemiddelde ± SEM) van die NAc-dop in langtermyn-sukrose-diere (blokkies) in vergelyking met kontroles (sirkels), ongepaarde studente tToets, P > 0.05, n = 9; beheer en n = 9; 12 week sukrose. Takorde-analise (gemiddelde ± SEM) van dendritiese segmentnommer per takorder (F), gemiddelde dendritiese lengte per takorder (G) en dendritiese ruggraatdigtheid per takorder (H). Langtermyn-sukroseverbruik het dendritiese lengte by distale takordes (5 +) en verhoogde dendritiese ruggraatdigtheid by distale takorders (4 +) vergeleke met kontroles (G, H), tweerigting-ANOVA's met Bonferroni-na-toetse, *P <0.05, **P <0.01, n = 9; beheer en n = 9; langtermyn sukrose. Skaalbalke: (A, B) = 20 μm; inset van (A, B) = 10 μm; (C) = 1 mm.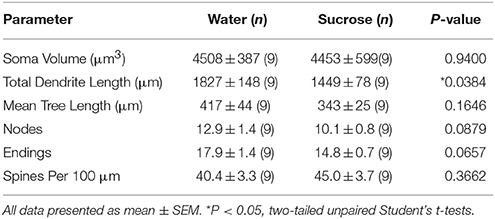
Tabel 2. Algemene morfologiese parameters van medium-stekelneurone uit die nukleusakkapsel-dop van langtermyn-sukroseverbruikende rotte en ouderdom-gekontroleerde waterkontroles.
Na aanleiding van die karakterisering van die algemene dendritiese morfologie van langtermyn-sukrose wat NAc-skulp MSN's verbruik, het ons dendritiese arborisasies en ruggraatdigtheid geanaliseer met betrekking tot hul takorder eienskappe. Ons omvattende beoordeling van die dendritiese bome het die aantal dendritiese segmente per takorde gekwantifiseer, die gemiddelde lengte van dendritiese segmente per takorder en gemiddelde ruggraatdigtheid per takorde van NAc-skulp MSNs van waterbeheer en langtermyn-sukroseverbruikende rotte. 'N Opsomming van die takorderdata en analise word in Tabel uiteengesit 3.
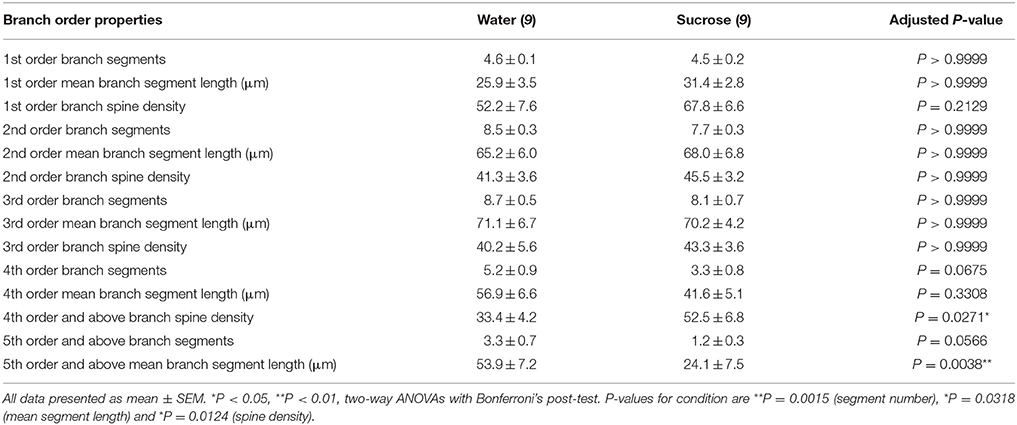 Tabel 3. Takordragseienskappe van medium-spinyneurone van langtermyn-sukrose en waterdrottende rotte.
Tabel 3. Takordragseienskappe van medium-spinyneurone van langtermyn-sukrose en waterdrottende rotte.Die gemiddelde dendritiese taksegmentnommer per takorde van NAc-skulp MSN's is aansienlik verminder in langtermyn-sukroseverbruikende rotte in vergelyking met waterkontroles (**P = 0.0015, tweerigting-ANOVA). Bonferroni-na-toetse het 'n neiging getoon na 'n verminderde aantal taksegmente by 4th (Water: 5.2 ± 0.9, n = 9; Sukrose 3.3 ± 0.8, n = 9, P = 0.0675, Figuur 2F, Tabel 3), en 5th orde en bo takopdragte (Water: 3.3 ± 0.7, n = 9; Sukrose 1.2 ± 0.3, n = 9, P = 0.0566, Figuur 2F, Tabel 3). Die gemiddelde dendritiese segmentlengte per takorde van NAc-skulp MSN's is ook aansienlik verminder in langtermyn-sukroseverbruikende rotte in vergelyking met waterkontroles (*P = 0.0444, tweerigting-ANOVA). Bonferroni-na-toetse het 'n vermindering van 55% by 5th-orde takke en daarbuite gewys (Water: 53.9 ± 7.2 μm, n = 9; Sukrose 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figuur 2G, Tabel 3).
Takorde analise het 'n beduidende toename in die dendritiese ruggraatdigtheid van NAc-skulp MSNs van langtermyn-sukroseverbruikende rotte vergeleke met beheermaatreëls (*P = 0.0124, tweerigting-ANOVA). Bonferroni-na-toetse het 'n stygingsdigtheidsverhoging van 57% by distale 4th-orde takke en daarbuite getoon. (Water: 33.4 ± 4.2, n = 9; Sukrose 52.5 ± 6.8, n = 9, P = 0.0271 *, inset van figure 2A, B, H, Tabel 3). Verteenwoordigende beelde van algehele MSN-argitektuur en distale ruggraatdigtheid (inset) word in figure weergegee 2A, B.
Saam met hierdie resultate word aangedui dat korttermyn-sukroseverbruik min invloed het op morfologiese parameters van MSN's binne die NAc-dop. Na langdurige verbruik, is daar egter 'n beduidende afname in die neuronale boonlengte en kompleksiteit, veral in distale dendritiese takke. Gelyktydige toename in distale ruggraatdigtheid is ook duidelik in NAc-skulp MSN's van langtermyn-sukroseverbruikende rotte.
Medium Spiny Neurons van die Nucleus Accumbens Core het die vertakkingskompleksiteit verlaag na lang, maar nie korttermyn-sukroseverbruik nie.
Na korttermyn sukroseverbruik was daar geen beduidende verskille in NAc kern MSN morfometriese parameters nie (Tabel 4). Daar was ook geen beduidende verskille tussen 4-week sukrose verbruik en waterbeheer kern MSNs in ontledings wat verband hou met sentrifugale tak orde. Namens dendritiese segmente per takorde (P = 0.7717), gemiddelde dendritiese lengte per takorde (P = 0.2096), en gemiddelde ruggraatdigtheid per takorde (P = 0.3521, tweerigting-ANOVA's) verskil nie tussen groepe nie.
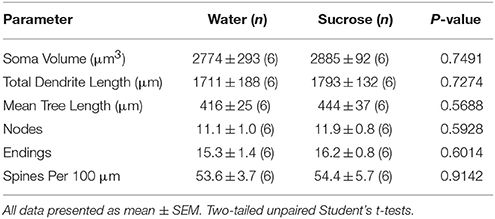 Tabel 4. Algemene morfologiese parameters van medium-stekelneurone uit die kern-kern van korttermyn-sukrose-verterende rotte en ouderdom-gekontroleerde waterbeheer.
Tabel 4. Algemene morfologiese parameters van medium-stekelneurone uit die kern-kern van korttermyn-sukrose-verterende rotte en ouderdom-gekontroleerde waterbeheer.Langdurige sukroseverbruik het ook geen betekenis gehad op NAc kern MSN morfometriese parameters nie (Tabel 5). Die gemiddelde dendritiese taksegmentnommer per takorde van NAc-kern MSNs is aansienlik verminder in langtermyn-sukroseverbruikende rotte in vergelyking met waterkontroles (*P = 0.0416, tweerigting-ANOVA), maar daar was geen beduidende verskille in gemiddelde dendritiese lengte per takorder nie (P = 0.0995) en gemiddelde ruggraatdigtheid per takorde (P = 0.4888, tweerigting-ANOVA's) tussen MSN's in die NAc-kern van langtermyn-sukroseverbruikende rotte in vergelyking met waterbeheer. Saam gesien, ons data toon dat die NAc-kern nie so reageer op langtermyn-sukroseverbruik in vergelyking met MSN's van die NAc-skulpgebied nie.
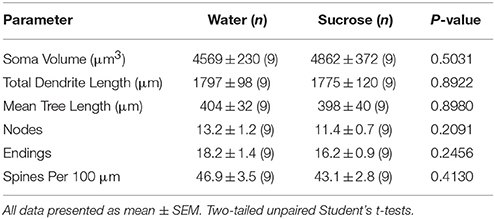 Tabel 5. Algemene morfologiese parameters van medium-stekelneurone uit die kern-kern van langtermyn-sukroseverbruikende rotte en ouderdom-gekontroleerde waterbeheer.
Tabel 5. Algemene morfologiese parameters van medium-stekelneurone uit die kern-kern van langtermyn-sukroseverbruikende rotte en ouderdom-gekontroleerde waterbeheer.Bespreking
Die verhoogde beskikbaarheid van hoogs versoete voedsel in die Westerse dieet het nie net bygedra tot die verhoogde voorkoms en ekonomiese las van vetsug en tipe II diabetes nie, maar dit het ook gelei tot 'n aanvang van eetversteurings soos binge-eet (Swanson et al., 2011; Kessler et al., 2013; Davis, 2015). Alhoewel die verslawende eienskappe van suikers, insluitende fruktose en sukrose, spekulatief bly, is daar 'n opvallende ooreenkoms in die gedrags- en neurale korrelate wat manifesteer as gevolg van oor-eet en langdurige dwelmgebruik (Avena et al., 2008, 2011). Boonop aktiveer suiker die brein se beloningskringe op 'n manier wat soortgelyk is aan dwelmmiddels (Volkow et al., 2012), en die resultate van menslike studies dui daarop dat suiker en soetigheid dwing kan veroorsaak wat vergelykbaar is met diegene wat deur verslawende middels veroorsaak word, soos alkohol en nikotien (Volkow et al., 2012). Ons het dus 'n model van binge-sukroseverbruik in rotte gebruik om die effekte van kort- (4 weke) en langtermyn (12 weke) sukroseverbruik op neuronale morfologie van MSNs in die NAc te bepaal, 'n belangrike komponent van die oorvleuelende beloningskring Dit word deur suiker en verslawende middels gemoduleer. Ons wys dat MSN's van die NAc-dop van chroniese langtermyn-sukroseverbruikende rotte aansienlik verminderde dendritiese lengte en kompleksiteit het, maar verhoogde distale dendritiese ruggraatdigtheid. Langtermyn sukroseverbruik het geen invloed gehad op die morfologie van MSNs van die NAc-kern nie, terwyl korttermyn-sukroseverbruik ook geen beduidende effek op MSN-morfologie van die NAc-kern of -skulp gehad het nie. Hierdie resultate toon nie net 'n direkte effek van langdurige binge-like sukrose-inname op neuronale morfologie van NAc-skulp MSNs nie, maar ook die potensiële skadelike gevolge van die langdurige verbruik van hoë suikerbevattende diëte.
Die NAc, wat deel vorm van die ventrale striatum, bestaan hoofsaaklik uit MSNs, wat morfologies gekenmerk word as mediumgrootte neurone met uitgebreide dendritiese arborisasies en hoë ruggraatdigtheid (Kemp en Powell, 1971; Graveland en DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). Glutamatergiese en dopaminerge neurone is die twee primêre afferente insette aan die NAc, hoofsaaklik met die dendritiese skagte en ruggraat van MSNs (Groves, 1980; Kaiya en Namba, 1981; Groves et al., 1994). Spesifiek, die NAc-dop en -kern ontvang glutamatergiese insette uit funksioneel afsonderlike kortikale gebiede (Brog et al., 1993). Die NAc-dop word ook geïnterpreteer deur opwekkende afferente uit subkortiese streke soos die hippokampus, thalamus en basolaterale amygdala (Brog et al., 1993; Wright en Groenewegen, 1995). Vorige studies het getoon dat hierdie glutamatergiese insette 'n sleutelrol speel in motivering en doelgerigte gedrag soos kos en beloning soekende (Maldonado-Irizarry et al., 1995; Kelley en Swanson, 1997; Reynolds en Berridge, 2003; Richard en Berridge, 2011). Die ander oorheersende inset op NAc MSNs is afkomstig van dopaminerge afferente wat uit die ventrale tegmentale area uitsteek (Lindvall en Björklund, 1978; Veening et al., 1980; Kalivas en Miller, 1984). Interessant genoeg, het vorige studies met soortgelyke modelle van intermitterende suikertoegang getoon dat die gevolglike binge-like verbruik 'n toename in ekstrasellulêre dopamien in die NAc tot gevolg het (ook al in mindere mate) tot dwelmmiddels (Rada et al., 2005; Avena et al., 2006), en kan dopamien reseptor uitdrukking moduleer (Colantuoni et al., 2001, 2002) in die NAc kern en dop. Interessant genoeg veroorsaak binge-like verbruik van sukrose 'n toename in inname oor tyd, soortgelyk aan selfadministrasie van dwelmmiddels soos kokaïen en heroiN (Ahmed en Koob, 1998; Ahmed et al., 2000, 2003) wat verband hou met die ontwikkeling van 'n "verslawende soortgelyke" staat.
Ons ontleding van takordemorfometrie toon dat die algehele vermindering in dendritiese lengte van NAc-skulp-MSN's wat veroorsaak word deur langtermyn-sukrose-inname, hoofsaaklik uit reduksies in die kompleksiteit van distale takordes lei. Ons het verminderde distale vertakking (4th en 5th orde en bo takorders) waargeneem en die gemiddelde lengte by 5th-orde en hoër dendriete het aansienlik verminder, gekombineer met verhoogde ruggraatdigtheid by hierdie takorders. 'N Algemene faktor wat hierdie tipe dendritiese herstrukturering moontlik beïnvloed, sluit in veranderinge in sinaptiese konnektiwiteit en / of funksie (Russo et al., 2010). Vorige studies het getoon dat glutamatergiese sinapse op MSN's hoofsaaklik gevorm word op stekels, veral by distale dendriete (Groenewegen et al., 1999). Daarbenewens, mede-lokalisering van dopamien- en glutamatergiese insette van die prefrontale korteks (Sesack en Pickel, 1992), hippokampus (Totterdell en Smith, 1989; Sesack en Pickel, 1990), en amygdala (Johnson et al., 1994) is op dendritiese stekels van MSNs waargeneem. Hierdie waarnemings, gekombineer met die verhoogde ruggraatdigtheid na langtermyn-sukroseverbruik wat in ons studie voorkom, ondersteun die vorming van verhoogde opwekkende insette. Daarom ontstaan die moontlikheid waar aanhoudende effekte wat veroorsaak word deur langdurige binge-like sukrose inname, verhoogde opwindende sinaptiese aktiwiteit by die distale dendriete van MSN's in die NAc-dop kan fasiliteer. Gevolglik kan vermindering en / of terugtrekking van distale dendriete tot gevolg hê deur middel van 'n sinaptiese homeostatiese meganisme (Reissner en Kalivas, 2010), maar dit moet nog bepaal word.
Dit is interessant om daarop te let dat Crombag en kollegas het getoon dat daar geen ruggraatdigtheidstoename in die NAc-dop na 4-week sukroseverbruik deur die neuspoot selfadministrasieparadigma was nie, ten spyte van 'n meer robuuste verkryging en 'n hoër reaksietempo vir sukrose in vergelyking met met amfetamien (Crombag et al., 2005). Hul waarneming van 'n afwesigheid van verandering in ruggraatdigtheid by 4 weke weerspieël ons bevindinge. In teenstelling hiermee toon ons studie egter dat die volgende langtermyn (12 week) blootstelling aan chroniese sukroseverbruik, 'n beduidende toename in distale ruggraatdigtheid op die MSN's van die sukrose-ervaringsrotte ervaar. Verder het ons laboratorium voorheen getoon dat die langtermyn (12 week) sukroseverbruik 'n differensiële farmakologiese reaksie op farmakoterapeutiese middels fasiliteer wat getoon is om dopamien- en asetielcholienreaksies op die vlak van die NAc te modelleer (Shariff et al., In pers). Saamgestel, dui dit daarop dat langtermyn (12 weke en verder) sukrose blootstelling, wat meer akkuraat weerspieël word van die werklike scenario's, lei tot morfologie aanpassings op die vlak van die NAc.
In terme van dwelmmiddels, veroorsaak herhaalde blootstelling aan verskillende middels langdurige veranderinge in die struktuur van dendriete en dendritiese stekels. Byvoorbeeld, amfetamiene en kokaïen verhoog beide ruggraatdigtheid in die NAc in beide dop en kern (Robinson en Kolb, 2004). Nikotienblootstelling het ook getoon om ruggraatdigtheid in die NAc-dop te verhoog. Omgekeerd lei morfienblootstelling tot 'n afname in ruggraatdigtheid en dendritiese takkompleksiteit (Robinson en Kolb, 2004). In terme van langtermyn-sukroseverbruik het ons 'n toename in ruggraatdigtheid, soortgelyk aan amfetamien, kokaïen en nikotien, waargeneem en teenoor die effek van morfien waargeneem. In teenstelling met amfetamien en kokaïen, maar soortgelyk aan nikotien, is die toename van ruggraatdigtheid op langtermyn blootstelling aan sukrose beperk tot die NAc-dop. Dit is ook interessant dat veranderinge in beide dendritiese vertakkings (Robinson en Kolb, 1999) en ruggraatdigtheid (Li et al., 2003) wat deur amfetamien of kokaïen geproduseer word, is beperk tot distale dendriete van MSNs in die NAc, wat die bevindinge in ons studie weerspieël. Verder, en bevestigend aan die veranderinge wat hierbo beskryf is, is ook sukroseverbruik voorheen getoon om opwindende sinaptiese sterkte op akkumulale dopamienneurone te verbeter (Stuber et al., 2008b) sowel as ander komponente van die mesolimbic beloning pad (Stuber et al., 2008a; Chen et al., 2010). Saam met mekaar, dit plaas sucrose as 'n kragtige modulator van neuron morfologie na langdurige swaar gebruik, wat verwant is aan die effekte waargeneem word van dwelms van misbruik.
Alhoewel verdere ondersoeke nodig is om die sellulêre en sinaptiese meganismes te identifiseer wat bydra tot die morfologiese veranderinge wat in hierdie studie gesien word, toon ons resultate betekenisvolle neuronale effekte wat voortspruit uit langtermyn-sukroseverbruik. In die besonder, 'n oorweging wat nie in ons studie ondersoek is nie, is of die waargenome morfologiese effekte van sukrose ook verkry kan word met nie-kalorie-versoeters soos sakkara. In hierdie verband is dit belangrik om daarop te let dat Lenoir en kollegas het getoon dat intense soetigheid kokaïenbeloning oortref, of dit deur sakkara of sukrose gegenereer word (Lenoir et al., 2007). Verder, 'n onlangse studie gepubliseer deur ons laboratorium (Shariff et al., In pers) toon dat varenikline, 'n nikotiniese asetielcholien-reseptor-partiële agonist, beide sukrose- en sarcharininname in knaagdiere verminder het, volgens dieselfde langtermyn-intermitterende toegangsregime wat in die huidige studie gebruik is. Interessant genoeg, het vorige studies ooreenkomste getoon in die akute effekte van nie-kalorie-versoeters soos sarkarien en sukrose op die vlak van die NAc (Scheggi et al., 2013; Tukey et al., 2013; Carelli en Wes, 2014). Nog verdere studies moet bepaal of nie-kalorie-versoeters langtermyn-effekte kan oplewer wat soortgelyk is aan veranderinge in die morfologie van die NAc-skulp-MSNs wat veroorsaak word deur die langtermyn-sukroseverbruik wat hier genoem word.
Die gebrek aan effek op NAc MSN morfologie na korttermyn sukroseverbruik, beklemtoon die belangrikheid van langtermynstudies om die impak van langdurige misbruik van dwelms of natuurlike belonings soos sukrose te bepaal. Wat afhanklikheid betref, is nie net herhaalde siklusse van binge-inname en onthouding-sleutelkomponente van die verslawingskringloop nie, maar 'n groeiende bewyse het getoon dat die oorgang na afhanklikheid 'n progressiewe proses is wat dikwels oor 'n lang tydperk voorkom. Alhoewel die verslawende eienskappe van suikers onseker bly, word die plausibiliteit van verslawing aan ander nie-dwelmbelonings soos seks, dobbel en kos toenemend ondersoek. Die resultate van hierdie studie voeg meriete by die hipotese dat suikers soos sukrose potensieel verslawende eienskappe het as gevolg van langtermyn, binge-like verbruik. Ons resultate het ook implikasies vir die groeiende aantal kinders en adolessente wat ongehoorlike eetgewoontes (hoë suikerverbruik en binge-eet) in volwassenheid handhaaf. In ooreenstemming met die verhoogde risiko om metaboliese effekte te ontwikkel, is dit ook moontlik dat neurologiese en psigiatriese gevolge wat gemoedsrus en motivering beïnvloed, ook uit hierdie gedrag kan voortspruit.
Skrywer Bydraes
Deelname aan navorsingsontwerp: PK, SB. Gevoerde eksperimente: PK, MS, AB, MF, EM. Data analise: PK, MF, MS. Interpreteer die data en dra by tot die skryf van die manuskrip: PK, MS, MF, EM, MB, SB. Alle outeurs lees en goedgekeur die finale manuskrip vir indiening.
Konflik van belangstelling
Die skrywers verklaar dat die navorsing gedoen is in die afwesigheid van enige kommersiële of finansiële verhoudings wat as 'n potensiële botsing van belange beskou kan word.
Die beoordelaars SC, SA en die hanteringsredakteur het hul gedeelde affiliasie verklaar, en die hanteringsredakteur verklaar dat die proses steeds voldoen aan die standaarde van 'n billike en objektiewe hersiening.
Erkennings
Hierdie werk is ondersteun deur befondsing van toekennings van die Australiese Navorsingsraad (FT1110884) na SB en die Nasionale Gesondheids- en Mediese Navorsingsraad (1061979) na SB en MB.
Aanvullende materiaal
Die aanvullende materiaal vir hierdie artikel kan aanlyn gevind word by: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Aanvullende figuur 1. Sucrose inname en voorkeur van 4 en 12 week sukrose wat rotte verteer. (A, B) toon toename in totale sukrose inname (ml) oor 4 en 12 weke blootstelling. (C, D) toon hoë voorkeur vir sukrose oor water gedurende periodes van sukrose aanbieding.
Verwysings
Ahmed, SH en Koob, GF (1998). Oorskakeling van matige tot oormatige dwelminname: verandering in hedoniese setpunt. Wetenskap 282, 298-300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF, en Parsons, LH (2003). Escalatie van kokaïen-selfadministrasie is nie afhanklik van veranderde kokaïen-geïnduceerde kerne wat dopamienvlakke behels nie. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR, en Koob, GF (2000). Aanhoudende toename in die motivering om heroïen in rotte te neem met 'n geskiedenis van dwelm eskalasie. Neuropsigofarmakologie 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Tik 2-diabetes by kinders: kliniese aspekte en risikofaktore. Horm Res 57 (Suppl. 1), 19-28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG, en Gold, MS (2011). Oorvleueling in die nosologie van middelmisbruik en ooreet: die translasionele implikasies van "voedselverslawing". Kur. Dwelmmisbruik Eerw. 4, 133-139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P., en Hoebel, BG (2008). Bewyse vir suikerverslawing: gedrags- en neurochemiese effekte van intermitterende, oormatige suiker inname. Neurosci. Biobehav. Op 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N. en Hoebel, BG (2006). Sucrose sham voer op 'n binge skedule vrylating dopamine herhaaldelik en elimineer die asetielcholien versadiging reaksie. Neurowetenskap 139, 813-820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Die aanneemlikheid van suikerverslawing en sy rol in vetsug en eetversteurings. Clin. Nutr. 29, 288-303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA, en Popkin, BM (2014). Dieet suiker en liggaamsgewig: het ons 'n krisis bereik in die epidemie van vetsug en diabetes ?: gesondheid word veroordeel! Gooi die suiker op. Diabetes Care 37, 950-956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY, en Zahm, DS (1993). Die patrone van afferente innervering van die kern en dop in die "accumbens" deel van die rat ventrale striatum: immunohistochemiese opsporing van retrogradelie-vervoerde fluoro-goud. J. Comp. Neurol. 338, 255-278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., en Kruseman, M. (2015). Suiker-versoete drank- en vetsugrisiko by kinders en adolessente: 'n sistematiese analise oor hoe metodologiese kwaliteit gevolgtrekkings kan beïnvloed. J. Acad. Nutr. Dieet. [Epub voor druk]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM, en Wes, EA (2014). Wanneer 'n goeie smaak sleg word: neurale meganismes onderliggend aan die opkoms van negatiewe invloed en gepaardgaande natuurlike beloning-devaluasie deur kokaïen. Neuro Farmacologie 76 (Pt B), 360-369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW, en Bonci, A. (2010). Sinaptiese plastisiteit in die mesolimbiese stelsel: terapeutiese implikasies vir middelmisbruik. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Bewyse dat tussenposes, oormatige suiker inname veroorsaak endogene opioïde afhanklikheid. Obes. Res. 10, 478-488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Oormatige suiker inname verander bindend vir dopamien en mu-opioïede reseptore in die brein. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. en Robinson, TE (2005). Opposisie-effekte van amfetamien self-administrasie ervaring op dendritiese stekels in die mediale en orbitale prefrontale korteks. Cereb. korteks 15, 341-348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Voorkoms van tipe 1 en tipe 2 diabetes onder kinders en adolessente van 2001 tot 2009. JAMA 311, 1778-1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Die epidemiologie en genetika van binge eating disorder (BED). CNS Spectr. 20, 522-529. doi: 10.1017 / s1092852915000462
Ford, ES, en Dietz, WH (2013). Neigings in energie-inname onder volwassenes in die Verenigde State: bevindinge van NHANES. Am. J. Clin. Nutr. 97, 848-853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD en Ogden, CL (2014). Voorkoms van oorgewig, vetsug en uiterste vetsug onder volwassenes: Verenigde State, 1960-1962 deur 2011-2012. Atlanta, GA: CDC.
Graveland, GA, en DiFiglia, M. (1985). Die frekwensie en verspreiding van mediumgrootte neurone met ingewande kerne in die primaat en knaagdier neostriatum. Brein Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV, en Voorn, P. (1999). Konvergensie en segregasie van ventrale striatale insette en uitsette. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Sinaptiese eindes en hul postsynaptiese teikens in neostriatum: sinaptiese spesialisasies wat uit die analise van seriële afdelings geopenbaar word. Proc. Natl. ACAD. Sci. VSA 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC, en Young, SJ (1994). 5-hidroksiedopamien-gemerkte dopaminerge aksone: driedimensionele rekonstruksies van aksone, sinapse en postsynaptiese teikens in rat neostriatum. Neurowetenskap 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Besluit: daar is voldoende wetenskaplike bewyse dat dalende suikerverzoete drankverbruik die voorkoms van vetsug- en vetsugverwante siektes sal verminder. Obes. Op 14, 606-619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z., en Totterdell, S. (1994). Invoer van die amygdala na die ratkern-accumbens: sy verhouding met tyrosienhidroksilase-immunoreaktiwiteit en geïdentifiseerde neurone. Neurowetenskap 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. en Namba, M. (1981). Twee tipes dopaminerge senuweesterminale in die ratneostriatum. 'N Ultrastruktuurstudie. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW, en Miller, JS (1984). Neurotensienneurone in die ventrale tegmentale area projekteer na die mediale nucleus accumbens. Brein Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ, en Emson, PC (1990). Projeksie subtipes van rat neostriatale matriks selle geopenbaar deur intracellulêre inspuiting van biocytine. J. Neurosci. 10, 3421-3438.
Kelley, AE en Swanson, CJ (1997). Voeding geïnduseer deur blokkade van AMPA en kainate reseptore binne die ventrale striatum: 'n mikroinfusie kartering studie. Behav. Brein Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM, en Powell, TP (1971). Die sinaptiese organisasie van die caudate-kern. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Beloningsmeganismes in vetsug: nuwe insigte en toekomstige rigtings. Neuron 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Die voorkoms en korrelate van binge eating disorder in die Wêreldgesondheidsorganisasie Wêreld Geestesgesondheidsopnames. Biol. Psigiatrie 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, en Bartlett, SE (2015). Strukturele en funksionele karakterisering van dendritiese arbors en GABAergiese sinaptiese insette op interneurone en hoofselle in die rat basolaterale amygdala. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., en Ahmed, SH (2007). Intense soetheid oortref kokaïenbeloning. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. en Robinson, TE (2003). Die ligging van volgehoue amfetamien-geïnduceerde veranderinge in die digtheid van dendritiese stekels op medium stekelneurone in die nukleus accumbens en caudate-putamen. Neuropsigofarmakologie 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O., en Björklund, A. (1978). Anatomie van die dopaminerge neuron stelsels in die rotbrein. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M., en Nestler, EJ (2009). Homeostatiese en hedoniese seine wisselwerking in die regulering van voedselinname. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ, en Kelley, AE (1995). Glutamaat-reseptore in die nukleusbodemskyf beheer voergedrag via die laterale hipotalamus. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Despres, JP en Hu, FB (2010). Suiker-versoete drank, vetsug, tipe 2 diabetes mellitus, en kardiovaskulêre siekte risiko. Sirkulasie 121, 1356-1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM, en Popkin, BM (2012). Gebruik van kalorie- en niekaloriese versoeters in Amerikaanse verbruikersverpakte voedsel, 2005-2009. J. Acad. Nutr. Dieet. 112, 1828-1834 e1821-e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM, en Popkin, BM (2002). Neigings in energie-inname in die VSA tussen 1977 en 1996: soortgelyke verskuiwings wat oor ouderdomsgroepe gesien word. Obes. Res. 10, 370-378. doi: 10.1038 / oby.2002.51
Paxinos, G., en Watson, C. (2007). Die Rat Brein in Stereotaksiese Koördinate. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Wat is verkeerd met die Amerikaanse benadering tot vetsug? Virtuele Mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM, en Hoebel, BG (2005). Daaglikse bingeing op suiker stel herhaaldelik dopamien in die accumbens dop. Neurowetenskap 134, 737-744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW, en McNeill, TH (1989). Golgi studie van die muis striatum: ouderdomsverwante dendritiese veranderinge in verskillende neuronale populasies. J. Comp. Neurol. 279, 212-227. doi: 10.1002 / cne.902790205
Ranjan, A. en Mallick, BN (2010). 'N Gewysigde metode vir konsekwente en betroubare Golgi-cox kleuring in aansienlik verminderde tyd. Front. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Tik 2 diabetes mellitus by kinders en adolessente. Wêreld J. Diabetes 4, 270-281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ, en Kalivas, PW (2010). Die gebruik van glutamaat homeostase as 'n teiken vir die behandeling van verslawende siektes. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM, en Berridge, KC (2003). Glutamaat-motiverende ensembles in kernklemme: Rostrocaudale dopgradiente van vrees en voeding. EUR. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM, en Berridge, KC (2011). Nucleus accumbens dopamien / glutamaat interaksie skakelaars modi genereer begeerte versus dread: D (1) alleen vir eetlustige eet, maar D (1) en D (2) saam vir vrees. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE, en Kolb, B. (1999). Veranderinge in die morfologie van dendriete en dendritiese stekels in die kernklem en prefrontale korteks na herhaalde behandeling met amfetamien of kokaïen. EUR. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE, en Kolb, B. (2004). Strukturele plastisiteit wat verband hou met blootstelling aan dwelmmiddels. Neuro Farmacologie 47 (Suppl. 1), 33-46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC, en Nestler, EJ (2010). Die verslaafde sinaps: meganismes van sinaptiese en strukturele plastisiteit in kernklemme. Neigings Neurosci. 33, 267-276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J., en Beatty, N. (1969). 'N Studie van piramidale sel-akson-collaterale in intakte en gedeeltelik geïsoleerde volwasse serebrale korteks. Brein Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC, en Elmquist, JK (2002). Die behoefte om te voed: homeostatiese en hedoniese beheer van eet. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, en Gambarana, C. (2013). Invloed van smaaklikheid op motivering om te werk vir kalorie- en nie-kalorie-voedsel in nie-voedsel-beroofde en voedsel-beroofde rotte. Neurowetenskap 236, 320-331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR, en Pickel, VM (1990). In die rat mediale nucleus accumbens, hippocampale en katekolaminerge terminale kom saam op stekelneurone en is in teenstelling met mekaar. Brein Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR, en Pickel, VM (1992). Prefrontale kortikale efferente in die rotssynaps op ongemerkte neuronale teikens van katecholamien terminale in die nucleus accumbens septi en op dopamienneurone in die ventrale tegmentale area. J. Comp. Neurol. 320, 145-160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (in pers). Neuronale nikotiniese asetielcholien reseptor modulators verminder suiker inname. PLoS ONE.
Sheehan, DV en Herman, BK (2015). Die sielkundige en mediese faktore wat verband hou met onbehandelde eetgewoontes. Prim. Care Companion CNS Disord. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Onderbroke toegang tot 20% etanol veroorsaak hoë etanolverbruik by Long-Evans- en Wistar-rotte. Alkohol. Clin. Exp. Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, en Bartlett, SE (2007). Varenicline, 'n alfa4beta2 nikotiniese asetielcholienreseptor gedeeltelike agonis, verlaag selektief etanolverbruik en soek. Proc. Natl. ACAD. Sci. VSA 104, 12518-12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. en Bonci, A. (2008a). Vrywillige inname van etanol verhoog die opwindende sinaptiese sterkte in die ventrale tegmentale gebied. Alkohol. Clin. Exp. Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Beloningvoorspellende leidrade verhoog opwindende sinaptiese sterkte op dopamienneurone in die middelbrein. Wetenskap 321, 1690-1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J., en Merikangas, KR (2011). Voorkoms en korrelasies van eetversteurings by adolessente. Resultate van die nasionale aanvulling op adolessente vir replikasie van die comorbiditeitsopname. Boog. Gen. Psychiatrie 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., en Mann, J. (2013). Dieet suikers en liggaamsgewig: sistematiese oorsig en metaanalises van gerandomiseerde gekontroleerde studies en kohortstudies. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S., en Smith, AD (1989). Konvergensie van hippokampale en dopaminerge toevoer na geïdentifiseerde neurone in die kern van die rat. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., et al. (2013). Inname van sukrose veroorsaak vinnige handel in AMPA-reseptore. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM, en Lieven, PA (1980). Die aktuele organisasie van die afferente tot die caudatoputamen van die rot. 'N Peperwortel-peroxidase-studie. Neurowetenskap 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R., en Contreras, AM (2014). Neurobiologiese basis van hunkering na koolhidrate. Voeding 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., en Baler, R. (2012). Beloning vir voedsel en dwelmmiddels: oorvleuelende stroombane in vetsug en verslawing by mense. Kur. Top. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Wyse, RA (1973). Vrywillige inname van etanol by rotte na blootstelling aan etanol op verskillende skedules. Psychopharmacologia 29, 203-210. doi: 10.1007 / BF00414034
Wright, CI, en Groenewegen, HJ (1995). Patrone van konvergensie en segregasie in die mediale nucleus accumbens van die rot: verwantskappe van prefrontale kortikale, middellyntalamiese en basale amigdaloïde afferente. J. Comp. Neurol. 361, 383-403. doi: 10.1002 / cne.903610304
Sleutelwoorde: binge-agtige verbruik, langtermyn, mediumstekige neuron, nucleus accumbens, sukrose
Aanhaling: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC en Bartlett SE (2016) Langdurige verbruik van sukrose in 'n binge-agtige manier, verander die morfologie van mediumstekelige neurone in die Nucleus Accumbens-skulp. Front. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Ontvang: 03 Desember 2015; Aanvaar: 07 Maart 2016;
Gepubliseer: 23 Maart 2016.
Geredigeer deur:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Frankryk
Nagesien deur:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Frankryk
Stéphanie Caille, Centre National de la Recherche Scientifique, Frankryk
Kopiereg © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham en Bartlett. Dit is 'n oop-toegang artikel versprei onder die voorwaardes van die Creative Commons Erkenning Lisensie (CC BY). Die gebruik, verspreiding of reproduksie in ander forums word toegelaat, mits die oorspronklike skrywer (s) of lisensiegewer gekrediteer word en dat die oorspronklike publikasie in hierdie joernaal aangehaal word, in ooreenstemming met die aanvaarde akademiese praktyk. Geen gebruik, verspreiding of voortplanting word toegelaat wat nie aan hierdie bepalings voldoen nie.
* Korrespondensie: Selena E. Bartlett, [e-pos beskerm]
