Front. Behav. Neurosci., 02 April 2014 |
Carla Kalkhoven1, Cor Sennef1, Ard Peeters1 en Ruud van die Bos2*
- 1Chardon Pharma, Herpen, Nederland
- 2Departement Organiese Dierfisiologie, Fakulteit Natuurwetenskappe, Radboud Universiteit Nijmegen, Nijmegen, Nederland
Huntington se siekte (HD) is 'n genetiese, neurodegeneratiewe siekte, wat spesifiek stralatale neurone van die indirekte weg beïnvloed, wat lei tot 'n progressiewe afname in spierkoördinasie en verlies van emosionele en kognitiewe beheer. Interessant genoeg, bevat vooroordeel vir patologiese dobbelary en ander verslawings stoornisse in dieselfde cortico-striatale stroombane wat in HD geraak word, en vertoon soortgelyke disinhibisieverwante simptome, insluitende veranderende sensitiwiteit vir strawwe en belonings, impulsiwiteit en onvermoë om langtermyn voordele te oorweeg. oor korttermynbelonings. Beide HD-pasiënte en patologiese spelers toon ook soortgelyke prestasie-tekorte op riskante besluitnemingstake, soos die Iowa Dobbeltaak (IGT). Hierdie ooreenkomste dui daarop dat HD-pasiënte 'n waarskynlike risikogroep vir dobbelprobleme is. Sulke probleme is egter net bykans waargeneem in HD-pasiënte. In hierdie oorsig beoog ons om die risiko van patologiese dobbelary in HD, sowel as die onderliggende neurobiologiese meganismes, te karakteriseer. Veral met die huidige opkoms van maklik toeganklike internetgokgeleenthede, is dit belangrik om hierdie risiko's te verstaan en dienooreenkomstig toepaslike pasiëntondersteuning te verskaf. Op grond van neuropatologiese en gedragsbevindinge, stel ons voor dat HD-pasiënte nie 'n groter neiging het om risiko's te soek en dobbel te begin nie, maar dat hulle 'n groter kans het om 'n verslawing te ontwikkel sodra hulle aan dobbelaktiwiteite deelneem. Daarom moet huidige en toekomstige ontwikkelinge van internetgokmoontlikhede en verwante verslawings met omsigtigheid beskou word, veral vir kwesbare groepe soos HD-pasiënte.
Inleiding
Huntington se siekte (HD) is 'n genetiese neurodegeneratiewe siekte, geërf op 'n outosomale dominante wyse. Die siekte word gekenmerk deur progressiewe motoriese, kognitiewe en gedragsimptome, wat gewoonlik tussen 30 en 50 jaar oud word, en lei tot voortydige dood in 10-20 jaar na siekte begin. HD word veroorsaak deur 'n mutasie in die Huntingtin geen (HTT), wat lei tot proteïen aggregasie, deregulering van verskeie sellulêre prosesse, en uiteindelik sel dood. Neuronale degenerasie kom aanvanklik selektief voor in die striatum (caudate-kern en putamen), waar dit cortico-striatale weë beïnvloed wat dien om motoriese en kognitiewe funksies te beheer (Reiner et al., 2011; Vonsattel et al., 2011). Op die motoriese vlak word hierdie degeneratiewe proses as ongeorganiseerde bewegings (chorea) uitgedruk, terwyl pasiënte op 'n kognitiewe / gedragsvlak 'n "uitvoerende disfunksiesindroom" toon. Dit sluit onder meer impulsiwiteit, swak risikobepaling en die onvermoë om 'n swak verloop van aksie (Hamilton et al., 2003; Duff et al., 2010b). Soortgelyke gedrags- en kognitiewe simptome word gesien in verslawende gedrag wat verband hou met stowwe of aktiwiteite (Newman, 1987; Rosenblatt, 2007; Iacono et al., 2008). Daarom kan verwag word dat HD-pasiënte die risiko loop om verslawings te ontwikkel. Besluitnemingsparadigmas in laboratoriuminstellings het inderdaad voorgestel tekorte in riskante besluitneming in gevorderde HD pasiënte (bv. Stout et al., 2001), en patologiese dobbelary is toevallig in hierdie pasiëntgroep waargeneem (De Marchi et al., 1998). Hierdie bevindings is egter skaars, en verbasend min studies het direk simptome en gevolge van byvoorbeeld gedrags disinhibisie in HD ondersoek.
In hierdie resensie sal ons argumenteer dat HD-pasiënte 'n risikogroep kan wees vir die ontwikkeling van problematiese dobbelary. Eerstens word problematiese dobbelary gekenmerk deur vakke se onvermoë om dobbelary te stop ten spyte van finansiële, persoonlike of professionele probleme. Gebaseer op neurobiologiese steurnisse en gedragsimptome, blyk dit dat die kapasiteit om dobbelgedrag op te hou, in HD-pasiënte verminder of afwesig is. Tweedens, as gevolg van die meer liberale houding teenoor dobbelary en toenemende moontlikhede van wettige en onwettige internet dobbelary (sien bv. Griffiths, 2003), kan ons verwag dat die voorkoms van dobbelprobleme in die komende jare sal toeneem. Toenemende toeganklikheid kan spesifiek 'n risiko vir kwesbare groepe, soos HD pasiënte, wat nie voorheen aan sulke risiko's blootgestel is nie.
Oor die algemeen kan veranderende eksterne toestande en behandelingsmetodes onverwagte en ongewenste effekte hê op pasiëntgedrag, veral in komplekse neurologiese siektes. Sulke effekte word maklik gemis wanneer gedragsimptome nie gereeld herwaardeer word nie. Dit kan die beste geïllustreer word deur die geval van Parkinson se siekte, waar die bekendstelling van geneesmiddelbehandeling met dopamien-agoniste tot impulsbeheerstoornisse gelei het, soos kompulsiewe dobbelary, inkopies, eet en hipersexualiteit, wat veroorsaak is deur die overstimulasie van die mesolimbiese dopaminerge sisteem (Dodd et al., 2005; Witjas et al., 2012; Weintraub et al., 2013). Hierdie newe-effekte is egter nie erken tot jare na die bekendstelling van dopamien-agonistiese terapieë in kombinasie met maatskaplike veranderinge wat verband hou met (die beskikbaarheid van) inkopies, voedselverbruik, seksualiteit, internet en dobbelary. Hierdie voorbeeld illustreer dat herassessering van risikofaktore belangrik is om effektiewe behandeling en leiding te gee aan pasiënte in die gesig van 'n veranderende omgewing.
Hier sal ons die siekteprofiel van HD ondersoek in verband met verslawing, dobbelprobleme en besluitnemingstekorte. In Afdeling HD: Neuropatologie, Simptome en Progressie, progressie van HD simptome sal bespreek word in verhouding tot stoornisse in cortico-striatale stroombane wat betrokke is by taakleer, sensitiwiteit vir straf, en kognitiewe / impulskontrole. In Afdeling Risikobeweging en Patologiese Dobbel Gedrag in HD, sal die neurobiologiese profiel van HD-pasiënte bespreek word in die konteks van dobbel en gevestigde risiko-neem en besluitnemingstoetse, soos die Iowa Dobbeltaak (IGT) en die Cambridge Dobbeltaak (CGT). In Afdeling Bespreking, sal ons bespreek hoe 'n karakterisering van dobbel risiko's kan lei tot aanbevelings vir HD pasiënte en hul versorgers oor hoe om hierdie probleem te hanteer en watter situasies die beste vermy word. Ons beoog ook om nog onbeantwoorde vrae te identifiseer, wat as uitgangspunt kan dien vir toekomstige navorsing oor die voorkoms en risiko's van dobbelprobleme by HD-pasiënte.
HD: Neuropatologie, Simptome en Progressie
Neurobiologiese siekte meganismes
HD word veroorsaak deur 'n onstabiele CAG (trinucleotide, cytosine-adenine-guanine) herhaling in die koderingstreek van die HTT-geen, wat lei tot die produksie van mutant huntingtin-proteïen (Htt) met 'n uitgebreide poliglutamien (polyQ) -strek (MacDonald et al., 1993). Die aantal trinucleotied herhalings is omgekeerd gekorreleer met die ouderdom van siekte begin (Snell et al., 1993; Stine et al., 1993). Die meerderheid van HD-pasiënte het 40-55 herhalings wat tipiese volwasse aanvalle veroorsaak, terwyl uitbreidings van meer as 70 herhalings lei tot jeugdigheidsversteuring. Individue met minder as 35 CAG herhalings in die HTT geen sal nie HD ontwikkel nie. Alhoewel die presiese meganismes van HD-patogenese onbekend bly en nie hier in besonderhede bespreek kan word nie, behels hulle die vorming van proteïenaggregate deur polyQ uitgebrei Htt, asook die interaksie van mutant Htt met talle proteïene wat betrokke is by energiemetabolisme, proteïen en blaassel. vervoer en regulering van geen transkripsie (Li en Li, 2004; Jones en Hughes, 2011). Die gevolglike deregulering van hierdie sellulêre prosesse lei uiteindelik tot neuronale degenerasie deur meganismes wat eksitotoksisiteit en apoptose insluit.
Neuronale degenerasie word aanvanklik beperk tot die basale ganglia, waar die medium stekelneurone in die striatum (caudate-kern en putamen) spesifiek geraak word (Vonsattel en DiFiglia, 1998; Kassubek et al., 2004). Die striatum ontvang sy hoof-eksitatoriese (glutamatergiese) inset van kortikale gebiede, terwyl dit sy dopaminerge insette van die substantia nigra ontvang. Die striatum het twee hoof inhibitiewe (GABA-ergiese) uitsette: 'n direkte en 'n indirekte pad (Figuur 1A). Striatale neurone van die direkte padprojek na die interne globus pallidus (GPi), wat op sy beurt inhibitoriese projeksies aan die thalamus het. Die thalamus gee aanleiding tot die vernaamste opwekkende insette van die korteks. So, in effek, aktiveer die aktivering van die direkte striatale weg GPi-aktiwiteit, wat weer thalamokortiese aktiwiteit versteur, en sodoende beweging- en kognitiewe funksies vergemaklik. Die indirekte striatale pad, aan die ander kant, projekte aan die eksterne GP (GPe), wat op sy beurt inhibitiewe projeksies na die subthalamiese kern (STN) stuur. Die STN stuur opwindende projeksies na die GPi. Gevolglik aktiveer die aktivering van die indirekte striatale roete die STN, wat dit toelaat om die GPi te aktiveer, wat weer die thalamokortiese aktiwiteit inhibeer, die beweging en kognitiewe funksies onderdruk. Adaptiewe gedrag lei tot 'n (delikate) balans van aktiwiteit in die direkte en indirekte pad. Patologie in die indirekte weg is die sleutel tot HD en ontwrig die balans in striatale beheer wat lei tot 'n verlies van inhibitiewe beheer oor motoriese funksionering en gedrag (Figuur 1B; Albin et al., 1989; Alexander en Crutcher, 1990).
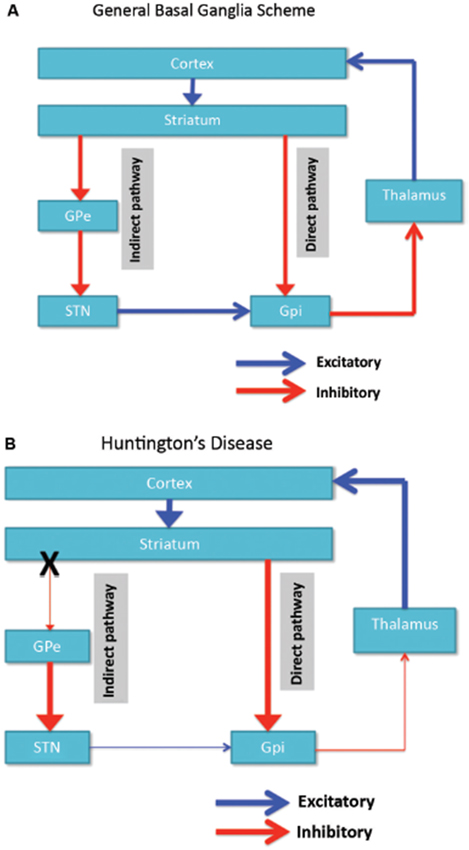
Figuur 1. (A) Vereenvoudigde skema van die organisasie van kortikobasale ganglia-netwerke (kortikale, striatale, pallidale en thalamiese gebiede) wat die direkte en indirekte weë in normale brein toon. (B) Spesifieke degenerasie van die indirekte weg (X) in HD lei tot 'n afname in inhibitiewe beheer oor kortikale funksies. GPe: eksterne globus pallidus; GPi: interne globus pallidus; STN: subthalamiese kern. rooi: inhibitiewe (GABA) bane, Blou: opwekkende (glutamaat) paaie.
Cortico-basale ganglia-kringe, wat verbindings tussen kortikale gebiede, streitale gebiede, pallidale gebiede en thalamiese gebiede insluit, word op 'n parallelle wyse georganiseer wat verskillende funksies in die organisasie van gedrag onderworpe. Soveel uitstekende resensies bestaan oor die anatomie en funksie van hierdie stroombane (bv. Alexander et al., 1986, 1990; Alexander en Crutcher, 1990; Yin en Knowlton, 2006; Verny et al., 2007; Yin et al., 2008; Haber en Knutson, 2010; Sesack en Grace, 2010), ons beklemtoon net 'n paar kwessies wat bevorderlik is vir ons oorsig. Eerstens bestaan daar 'n dorsale tot ventrale topografiese organisasie in beide kortikale en striatale gebiede. Dus, die dorsale prefrontale areas word geassosieer met dorsale striatale areas, terwyl die meer ventrale prefrontale areas geassosieer word met meer ventrale striatale areas (insluitend die nucleus accumbens). Tweedens kan breedweg drie funksioneel verskillende stroombane beskryf word. Die sensorimotoriese kring sluit die sensorimotoriese striatum (putamen) en sensorimotoriese kortikasies in wat verband hou met die uitvoering van motoriese gedrag. Die assosiatiewe / kognitiewe beheerbaan behels die dorsolaterale prefrontale korteks, anterior cingulêre korteks en assosiatiewe striatum (caudate-kern). Hierdie kring is veral relevant vir uitvoerende funksionering, dit wil sê, dit is betrokke by kognitiewe beheer, beplanning en werkgeheue. Daarbenewens is dit betrokke by die bevordering van langtermyn-adaptiewe gedrag deur die versterking of staking van instrumenteel gedrag, dit wil sê sekwense gedragsgedrag, geleer in interaksie met die omgewing (Kravitz et al., 2012; Paton en Louie, 2012). Die limbiese kring sluit die orbitofrontale korteks, ventromediale prefrontale korteks, amygdala en limbiese striatum in (nucleus accumbens). Hierdie kring is veral relevant vir die evaluering van die affektiewe waarde van stimuli, wat die verwagte beloning of straf van 'n opkomende stimulus, keuse of gebeurtenis, emosionele beheer en aanpasbare (emosionele) leer aandui (O'Doherty et al., 2001; Rushworth et al., 2007; Van den Bos et al., 2013b, 2014).
Patologie in HD word waargeneem in beide die putamen en caudate-kern (Vonsattel en DiFiglia, 1998; Kassubek et al., 2004; Vonsattel, 2008; Vonsattel et al., 2011; Hadzi et al., 2012). Daarbenewens volg in beide strukture atrofie 'n kenmerkende patroon wat begin in die dorsale en caudale streke en beweeg na die ventrale en rostrale streke as die siekte vorder (Vonsattel en DiFiglia, 1998; Kassubek et al., 2004; Vonsattel, 2008). Maar vroeë atrofie is ook in sommige kernstudies waargeneem in die nucleus accumbens en globus pallidus (Van die Bogaard et al., 2011; Sánchez-Castañeda et al., 2013). Alhoewel stoornisse in die sensorimotoriese kring (putamen) verband hou met die motoriese simptome, kan stoornisse in die assosiatiewe / kognitiewe beheerbaan (caudate-kern) verband hou met uitvoerende disfunksie, en veroorsaak tekorte in bv. Werkgeheue in vroeë HD-pasiënte (Lawrence et al., 1996; Bonelli en Cummings, 2007; Wolf et al., 2007). Versteurings in die limbiese kring, soos as gevolg van vroeë atrofie in die nukleus, kan verband hou met apatie en depressie (Bonelli en Cummings, 2007; Unschuld et al., 2012). Progressiewe atrofie in die striatum kan lei tot 'n opeenvolgende disfunksie van cortico-striatale bane. Byvoorbeeld, die ventrale caudate-kern is ook deel van die orbitofrontale kring, wat geraak word as die siekte vorder. Disfunksie van hierdie kring is verwant aan gedragsversteuring (Bonelli en Cummings, 2007). Uiteindelik kan degenerasie na ander breinareas versprei, insluitend ander dele van die basale ganglia (pallidale gebiede en thalamus), hippokampus, amygdala en kortikale gebiede in die laat stadiums van die siekte.
Kortom, HD word gekenmerk deur 'n spesifieke degenerasie van striatale neurone wat aan die indirekte roete behoort. Soos die siekte vorder, versprei atrofie van die striatum langs 'n caudale-rostral- en dorsale ventrale gradiënt wat 'n opeenvolgende versteuring van cortico-striatale bane veroorsaak. Die gevolglike verlies aan inhibitiewe beheer in hierdie kringe is direk verwant aan die progressie van motoriese, kognitiewe en gedragsimptome in HD, soos hieronder bespreek.
Simptome van HD
HD word gekenmerk deur 'n verskeidenheid progressiewe motoriese, kognitiewe en gedragsimptome. Die eerste simptome ontstaan gewoonlik op middeljarige ouderdom, met 'n gemiddelde aanvangs ouderdom van 40, alhoewel 'n klein persentasie pasiënte ly aan jeugdige HD, wat voor die ouderdom van 20 begin. Aangesien die simptome en progressie van jeugdige inskrywings HD ietwat van volwassenheidsstoornis is, sal ons op hierdie laasgenoemde groep fokus. Een van die eerste simptome wat in HD voorkom, is chorea (onwillekeurige bewegingsversteuring), en 'n kliniese diagnose word gewoonlik na aanvang van bewegingsafwykings gemaak (Shannon, 2011). Sommige studies meld egter subtiele kognitiewe en emosionele veranderinge voor aanvang van motoriese simptome, en die presiese volgorde van voorkoms en progressie van HD simptome bly 'n onderwerp van debat. Tog is verskeie omvattende resensies van die kliniese manifestasies van HD beskikbaar (Roos, 2010; Anderson, 2011; Shannon, 2011).
Motoriese simptome
Motoriese simptome begin duidelik in die vroeë stadiums van HD, en is gewoonlik die eerste simptome wat opgemerk word in laboratoriuminstellings en eerstegraadse familielede van HD-pasiënte (De Boo et al., 1997; Kirkwood et al., 1999, 2001). Motorstoornisse blyk te begin as 'n disfunksie in fout terugvoerbeheer (Smith et al., 2000), in ooreenstemming met die rol van die cortico-striatale motoriese kring in sensorimotoriese leer en beheer (Graybiel et al., 1994). Die eerste tekens van motoriese abnormaliteite is dikwels subtiele onwillekeurige bewegings (chorea) van bv. Gesigsspiere, vingers en tone ("twitching"), hiperrefleksie en oordrewe vrywillige bewegings (Young et al., 1986; Shannon, 2011), wat lei tot 'n algemene voorkoms van rusteloosheid en lompheid in vroeë HD pasiënte. Hierdie abnormale bewegings is subtiel en begin eers onopgemerk, maar word geleidelik versleg en versprei na alle ander spiere oor tyd. Ander vroeë motoriese simptome sluit in stadige of vertraagde saccadiese oogbewegings (Peltsch et al., 2008) en disartrië (Ramig, 1986; Young et al., 1986). Dysartria, 'n motoriese spraakversteuring, lei tot probleme met artikulasie en verswakking van woorde, wat spraak progressief moeiliker maak om te verstaan. Dysfagie (slukprobleme) word waargeneem in die meeste pasiënte met 'n aanvang in die middel siektetoestande, en word geleidelik vererger totdat pasiënte nie meer onaangeraak kan eet nie en dikwels 'n voedingsbuis in laat stadium HD benodigHeemskerk en Roos, 2011). Ander, nie-choreiese motoriese simptome wat gewoonlik in die middelstadium voorkom, sluit in komplekse gangversteuring, posturale onstabiliteit en distolonie (onwillekeurige spierkontraksies wat stadige herhalende bewegings en abnormale posture veroorsaak), wat dikwels gepaard gaan met gereelde val (Koller en Trimble, 1985; Tian et al., 1992; Louis et al., 1999; Grimbergen et al., 2008). Styfheid en bradykinesie (traagheid van beweging en reflekse) word soms waargeneem, maar word meestal beperk tot gevalle van jong begin HDBittenbender en Quadfasel, 1962; Hansotia et al., 1968). Hierdie motoriese simptome is in ooreenstemming met disfunksie van die sensorimotoriese (en assosiatiewe / kognitiewe beheer) cortico-striatale stroombane wat algemeen in HD geraak word.
Gedrags- en psigiatriese simptome
Gedragsversteurings in HD kan kompleks en moeilik wees om te klassifiseer, en hul voorkoms en begin is hoogs veranderlik tussen individue. Verder kan dit soms moeilik wees om gedragsversteurings te onderskei van normale hantering van 'n ontstellende siekte (Caine en Shoulson, 1983). Die aantal studies wat gedragsimptome in HD gekenmerk het, is beperk, en gevolglik is daar relatief min insig in hul voorkoms in die siekte (Van Duijn et al., 2007). Die mees algemene en konsekwente gedrags- en emosionele simptome in HD is prikkelbaarheid, apatie en depressie, wat voorkom met 'n voorkoms van ongeveer 50% (Caine en Shoulson, 1983; Folstein en Folstein, 1983; Craufurd et al., 2001; Kirkwood et al., 2001; Van Duijn et al., 2007, 2014; Tabrizi et al., 2009). Beide prikkelbaarheid en apatie word soms waargeneem in pre-manifeste HD-pasiënte (Tabrizi et al., 2009; Van Duijn et al., 2014), en ook depressie is aangemeld by vroeë kliniese stadiums (Shiwach, 1994; Julien et al., 2007; Epping et al., 2013). Hierdie affektiewe simptome is een van die eerste nie-motoriese simptome wat deur eerstegraadse familielede opgemerk word (Kirkwood et al., 2001). Tipiese apatie-verwante simptome, wat geleidelik erger word gedurende die siekte, sluit in gebrek aan energie, motivering en inisiatief, verminderde deursettingsvermoë en kwaliteit van werk, gebrekkige oordeel, swak selfversorging en emosionele klap (Craufurd et al., 2001; Kirkwood et al., 2001). Depressiewe simptome is verwant aan verhoogde aktiwiteit in die ventromediale prefrontale korteks (Unschuld et al., 2012). Irritabiliteit word geassosieer met orbitofrontale kring disfunksie, wat lei tot verminderde beheer oor emosionele response in die amygdala (Klöppel et al., 2010).
Ander, minder algemeen waargeneemde psigiatriese simptome en afwykings in HD is angs, obsessiewe-kompulsiewe versteuring, manie, skisofrenie-agtige psigotiese simptome, soos paranoia, hallusinasies en wanorde (Caine en Shoulson, 1983; Folstein en Folstein, 1983; Craufurd et al., 2001; Kirkwood et al., 2001; Van Duijn et al., 2007). Hierdie simptome kom gewoonlik nie voor tot mid- of laat stadiums van die siekte nie, hoewel dit per ongeluk in prekliniese HD-pasiënte gerapporteer is (Duff et al., 2007). Obsessiewe-kompulsiewe versteuring is geassosieer met skade aan die orbitofrontale korteks en anterior cingulêre korteks, terwyl skisofrenie, 'n afwyking wat tekorte in organisering, beplanning en aandag behels, verband hou met dorsolaterale prefrontale korteks disfunksie (Tekin en Cummings, 2002).
Daar word voorgestel dat die meeste psigiatriese simptome in HD eintlik deel uitmaak van 'n breë, ondefinieerbare "frontale lobe sindroom" of "uitvoerende disfunksie sindroom", wat simptome soos apatie, prikkelbaarheid, disinhibisie, impulsiwiteit, obsessiwiteit en volharding insluit (Lyketsos et al., 2004; Rosenblatt, 2007), wat almal algemeen in HD-pasiënte waargeneem word (Hamilton et al., 2003; Duff et al., 2010b). Die literatuur dui daarop dat die aanvang en progressie van gedragsimptome in HD heterogeen is, met affektiewe afwykings wat die meeste voorkom en met vroeë aanvang, terwyl angs, obsessief-kompulsiewe versteuring en psigotiese simptome minder algemeen voorkom en gewoonlik later in die siekte voorkom . Hierdie psigiatriese simptome word geassosieer met disfunksie van limbiese en assosiatiewe / kognitiewe beheer cortico-striatale stroombane wat algemeen in HD geraak word.
Kognitiewe simptome
Kognitiewe afname is nog 'n belangrike aspek van HD-patologie. Baie studies het spesifiek gefokus op die voorkoms van kognitiewe simptome in prekliniese en vroeë kliniese stadiums van HD, in die hoop om vroeë kliniese biomarkers van die siekte te ondersoek (hersien in Papp et al., 2011; Dumas et al., 2013). Algehele resultate dui daarop dat subtiele kognitiewe veranderinge waargeneem kan word tot 5-10 jaar voor aanvang van motoriese simptome met voldoende sensitiewe metodes. Een studie het selfs bevind dat ongeveer 40% van pasiënte by prekliniese en vroeë kliniese stadiums van HD reeds voldoen aan die kriteria vir ligte kognitiewe inkorting ('n versteuring wat verband hou met beperkte geheueverlies, wat nie aan die kriteria vir diagnose van demensie voldoen nie; Duff et al., 2010a). Egter nie alle studies ondersteun hierdie bevindinge nie (Blackmore et al., 1995; Giordani et al., 1995; De Boo et al., 1997; Kirkwood et al., 2001). Oor die algemeen stem die literatuur saam dat inligtingverwerking en psigomotoriese spoed in hierdie vroeë stadium veral geraak word (Rothlind et al., 1993; Kirkwood et al., 1999; Verny et al., 2007; Paulsen et al., 2008). Ander algemeen waargeneemde vroeë kognitiewe inkortings sluit in probleme met aandag, (werk) geheue en visuospatiale prestasie (Jason et al., 1988; Rothlind et al., 1993; Foroud et al., 1995; Lawrence et al., 1996; Hahn-Barma et al., 1998; Verny et al., 2007; Paulsen et al., 2008; Tabrizi et al., 2009; Papp et al., 2011; Stout et al., 2011). Kognitiewe onbuigsaamheid is waargeneem in vroeë siektepasiënte (Jason et al., 1988), in watter stadium ekstra-dimensionele verskuiwings spesifiek benadeel word, terwyl omkeerleer steeds ongeskonde is (Lawrence et al., 1996). Sodoende kan pasiënte nog steeds stimuluswaarde herwaardeer en nuwe stimulus-beloningsvoorwaardes binne dieselfde dimensie (bv. Vorm of kleur) leer, maar probleme ondervind om hul aandag te skuif na 'n ander dimensie (bv. Van kleur na vorm) soos vereis deur die nuwe taak reël om beloning te verkry. In latere stadiums van die siekte, veroorsaak kognitiewe onbuigsaamheid en volharding ook verswakte herleerleer in HD-pasiënte (Josiassen et al., 1983; Lange et al., 1995). Hierdie vordering van simptome is in ooreenstemming met die spesifieke disfunksie van die dorsolaterale prefrontale baan vroeg in die siekte, aangesien ekstdimensionele stelverskuiwing bemiddel word deur die dorsolaterale prefrontale korteks, terwyl omkeerleer gemedieer word deur die orbitofrontale korteks (Dias et al., 1996; McAlonan en Brown, 2003). Ander vroeë gestremdhede sluit in ongeorganiseerde gedrag, gebrekkige beplanning, swak oordeel, en verminderde gedrags- en emosionele beheer (Watkins et al., 2000; Paradiso et al., 2008; Duff et al., 2010b). Disinhibisie is waargeneem in vroeë HD-pasiënte, wie se prestasie verswak is aan take wat inhibering van voorkragtige, maar onvanpaste reaksies benodig (Holl et al., 2013). Ten slotte het verskeie studies bevind dat voorkliniese HD-pasiënte in die erkenning van negatiewe emosies soos woede, afgryse, vrees en hartseer benadeel word. Emosionele erkenning neem progressief af, en kan versprei na probleme met neutrale emosies in vroeë kliniese stadiums van die siekte (Johnson et al., 2007; Tabrizi et al., 2009; Labuschagne et al., 2013). Hierdie fenotipe hou verband met disfunksie van die orbitofrontale korteks, wat betrokke is by die verwerking van emosionele en beloningsinligting (Henley et al., 2008; Ille et al., 2011).
Studies met diermodelle van HD toon soortgelyke kognitiewe gestremdhede as dié waargeneem word by menslike pasiënte. Alhoewel nie alle studies stewige kognitiewe tekorte vind nie (Fielding et al., 2012), bevindings in rot- en muismodelle van HD sluit in angs, verhoogde responsiwiteit teenoor negatiewe emosionele stimuli, en gestremdhede in omkering leer en strategie verskuiwing (Faure et al., 2011; Abada et al., 2013). Een studie het spesifieke vroeë tekorte in omkering geleer voor aanvang van motoriese simptome in 'n rotmodel van HD (Fink et al., 2012). Interessant genoeg lyk dit of HD-diere 'n verhoogde reaksie op negatiewe emosionele stimuli het, terwyl menslike pasiënte minder erkenning van negatiewe emosies toon. Op die oomblik is dit onduidelik of dit weerspieël verskille in taak geadministreer (erkenning van emosies teenoor gedragsreaksies op dreigende stimuli), spesieverwante verskille in die uitkoms van patologie of 'n fundamentele verskil tussen die ratmodel en die menslike toestand. Oor die algemeen toon studies in beide menslike pasiënte en dieremodelle van HD dat 'n wye verskeidenheid kognitiewe funksies reeds in die vroeë HD aangetas kan word. Vroeë abnormaliteite sluit hoofsaaklik tekorte in aandag, geheue, kognitiewe buigsaamheid en emosionele erkenning in. In hierdie vroeë stadium het pasiënte dikwels 'n gebrek aan bewustheid van hul eie (afname in) kognitiewe vermoëns (Hoth et al., 2007). Met verloop van tyd word kognitiewe simptome steeds erger, wat uiteindelik lei tot ernstige subkortikale dementie in die laat stadiums van die siekte. Alhoewel die voorkoms van simptome oor die algemeen in ooreenstemming is met opeenvolgende verswakking van assosiatiewe / kognitiewe beheer en limbiese cortico-striatale stroombane, onderskeidelik, kan spesifieke funksies wat verband hou met die limbiese kring ook reeds in die vroeë stadium HD geraak word.
Gevolgtrekking
Motoriese, gedrags- en kognitiewe simptome in HD is in die verlede omvattend bestudeer en bly 'n onderwerp van belang as gevolg van die wye verskeidenheid en veranderlikheid in die voorkoms en aanvang van hierdie simptome oor pasiënte. Oor die algemeen is gedrags- en kognitiewe simptome verwant aan drie frontale gedragskategorieë: apatie, uitvoerende disfunksie en disinhibition. Die kombinasie van hierdie simptome word soms na verwys as "uitvoerende disfunksie sindroom". Al hierdie simptome hou verband met tekorte in die cortico-striatale bane wat die orbitofrontale korteks, dorsolaterale prefrontale korteks en anterior cingulêre korteks insluit. Soos hierbo bespreek, het neuropatologiese studies 'n geleidelike degenerasie van die striatum waargeneem in 'n dorsale tot ventrale rigting in HD-pasiënte. Alhoewel die gedrags- en kognitiewe waarnemings gedeeltelik saamstem met 'n progressiewe inkorting van kortikostrale bane, lyk die simptomatiese bevindings meer diffus as wat verwag word op grond van patologiese waarnemings. Aanvang en progressie van gedrags- en kognitiewe simptome in HD is hoogs heterogeen, wat aandui dat die skade aan striatale streke meer veranderlik en wydverspreid kan wees in vroeë stadiums van HD as voorheen gedink. Hierdie siening word ondersteun deur bewyse uit verskeie strukturele beeldvormingstudies (Thieben et al., 2002; Rosas et al., 2005; Van die Bogaard et al., 2011).
Risikobeweging en Patologiese Dobbel Gedrag in HD
Patologiese dobbelary
Alhoewel baie mense ontspanningsgeleentheid kan speel, kan dit vir sommige van hulle 'n oop probleem word, aangesien hulle patologiese vorme van hierdie gedrag ontwikkel. Patologiese dobbelary word gekenmerk deur 'n oormatige drang om te dobbel, ondanks duidelike negatiewe finansiële, persoonlike en professionele gevolge. Dit is onlangs as 'n verslawing in DSM-V geklassifiseer, aangesien dit baie ooreenstem met middelmisbruikstoornisse in beide diagnostiese kriteria en neuropatologie (Van Holst et al., 2010; Clark en Goudriaan, 2012). Patologiese dobbelary sal die eerste en enigste "gedragsverslawing" wees wat binne die kategorie "Addiction and Related Disorders ". Nietemin moet daarop gelet word dat verskille bestaan tussen verslawing aan psigoaktiewe stowwe en verslawing aan dobbelary. Eerstens, bevredigende drang na psigoaktiewe stowwe lê in die gebruik van die stof waarvan die effek bekend is, terwyl die bevrediging van die drang na dobbelary 'n onsekere uitkoms kan hê, aangesien geld gewen kan word of nie, tensy dit die daad van dobbel self is, byvoorbeeld as 'n opwindende aktiwiteit. Sodoende kan patologiese dobbelary meer heterogeen wees in hierdie opsig en ook 'n meer onsekere uitkoms as dwelmmisbruik. Daar moet op gelet word dat uitkomsveranderlikheid, insluitende beide oorwinnings en verliese, van kritieke belang is vir die ontwikkeling van dobbelverslawing, aangesien dit 'n wisselende intermitterende versterkingspatroon bied, wat die sterkste vorm van instrumentale / klassieke kondisionering is (Sharpe, 2002; Fiorillo et al., 2003). Tweedens kan psigo-stowwe sterker aktiwiteit in die brein en perifere senuweestelsel verander as dobbelary, as gevolg van hul direkte farmakologiese aktiwiteit by verskeie neurotransmitterstelsels, wat sodoende verslawende prosesse versnel, wat dwelmmisbruik 'n sterker vorm van verslawing maak.
Die onderliggende neurobiologiese meganismes van dobbel is kompleks en behels baie verskillende breinstreke en neurotransmitterstelsels (hersien in Raylu en Oei, 2002; Goudriaan et al., 2004; Potenza, 2013). Vooroordeel tot verslawing is verwant aan 'n verminderde dopamien D2-reseptore in die striatum, wat in 'n terugvoerlus funksioneer om verdere dopamien vrystelling te inhibeer. Die gevolglike hiperaktiwiteit van dopaminerge bane verhoog sensitiwiteit vir beloning, motivering en positiewe versterking van die verslawende gedrag (Volkow et al., 2002; Di Chiara en Bassareo, 2007). Spesifieke motiveringsveranderings wat voorkom wanneer patologiese dobbelary ontwikkel, sluit in verhoogde motivering om te dobbel (Van Holst et al., 2012) en groter aandag aan dobbelverwante stimuli (Brevers et al., 2011a,b). Daarbenewens het patologiese spelers die kognitiewe beheer oor gedrag in die algemeen verminder, soos aangedui deur verlaagde prestasie op reaksie inhibisie take, verhoogde impulsiwiteit, en 'n voorkeur vir onmiddellike oorvertraagde belonings in neurokognitiewe take (Goudriaan et al., 2004; Brevers et al., 2012a; Van den Bos et al., 2013a).
Patologiese dobbelaars verrig swak in vergelyking met kontroles op formele beloningsverwante riskante besluitnemingstake (bv. Cavedini et al., 2002; Brand et al., 2005; Brevers et al., 2012b; resensie: Brevers et al., 2013). Hierdie swak prestasie is onafhanklik van of take eksplisiete en stabiele reëls bevat vir oorwinnings en verliese soos die spel van dobbelstaak (Brand et al., 2005) of of vakke deur proef-en-fout moet leer, watter keuses is op die langtermyn voordelig, soos die IGT (Cavedini et al., 2002; Brevers et al., 2012b; sien Afdeling Risikovolle besluitneming deur HD-pasiënte op laboratoriumtaak vir besonderhede van hierdie taak). Die dobbelstoornis was egter eerder gekorreleer met prestasie op besluitnemingstake waarin die waarskynlikheid van uitkoms onbekend is (IGT) as met take met eksplisiete reëls (Brevers et al., 2012b). Hierdie waarneming is interessant, aangesien die tweede helfte van die IGT by vakke geleer is in die normale vakke, wat verband hou met take met eksplisiete reëls. Gesamentlik, dui hierdie data dus aan dat in patologiese dobbelstremminge in besluitneming kan voortspruit uit beide verminderde uitvoerende beheer, wat verband hou met meer eksplisiete reëls en versteurde beloningstraf (emosionele) prosessering, wat meer verband hou met proef-en- fout leer om langtermynwaarde van opsies te assesseer (Van den Bos et al., 2013a, 2014). Daarbenewens stel dit voor dat steurnisse in laasgenoemde 'n predisponerende faktor kan wees vir die toename van dobbelgedrag.
Uit hierdie studies is dit duidelik dat neurobiologiese predisposisie vir die ontwikkeling van patologiese dobbelgedrag versteurings in beide die assosiatiewe / kognitiewe beheerkring en die limbiese kring behels (Van den Bos et al., 2013a). As gevolg hiervan, patologiese spelers vertoon verminderde kognitiewe beheer, verhoogde impulsiwiteit, en verhoogde sensitiwiteit vir beloning, wat almal aspekte van gedragsdesinhibisie is (Iacono et al., 2008). Die kans dat 'n individu 'n verslawing in sy lewe ontwikkel, hang egter ook af van baie ander aspekte, soos vroeëre ervarings en omgewingsrisiko's.
Patologiese dobbelary in HD: Epidemiologiese bewyse
Met die toenemende aantal moontlikhede wat die internet bied, is daar ook in die afgelope jaar 'n styging in beide wettige en onwettige aanlyngokgeleenthede. Hierdie maklik toeganklike en dikwels onbeheerde dobbelaktiwiteite kan 'n risiko vir enigeen met 'n verhoogde vatbaarheid vir dobbelverslawing inhou, maar mag andersins nie by sulke aktiwiteite betrokke raak nie (Griffiths, 2003). HD-pasiënte is een van die groepe waarvoor internet dobbel so 'n risiko kan uitmaak, omdat gedragsverstopping - 'n algemene kenmerk in die siekte - 'n belangrike faktor is in die ontwikkeling van verslawings (Iacono et al., 2008). Inderdaad, soos hierbo genoem, toon HD-pasiënte verskeie tekens van disinhibisie, soos geïrriteerdheid, verminderde reaksie-inhibisie en verminderde emosionele erkenning, vroeg in die siekte. Ander simptome wat in HD waargeneem word, en die invloed van pasiënte op rasionele besluite kan beïnvloed, is kognitiewe onbuigsaamheid, volharding, swak oordeel en verminderde selfbewustheid. Behalwe hierdie simptomatiese ooreenkomste tussen HD-pasiënte en patologiese spelers, toon beide groepe strukturele en funksionele abnormaliteite in soortgelyke cortico-striatale bane.
In die lig van hierdie ooreenkomste tussen patologiese spelers en HD-pasiënte, kan ons verwag dat die voorkoms van dobbelprobleme onder HD-pasiënte verhoog sal word in vergelyking met die normale bevolking. Tog het slegs een studie tot dusver gevalle van patologiese dobbelary in 'n Italiaanse familie met HD gerapporteer (De Marchi et al., 1998). In hierdie familie is twee individue gediagnoseer met patologiese dobbelary rondom die ouderdom van 18, voor die aanvang van kliniese tekens van HD. Ander epidemiologiese studies het nie hieroor gerapporteer nie, alhoewel benadeelde besluitneming, risiko-opname en swak oordeel is getoon dat dit 'n risiko vir HD-pasiënte inhou om belangrike lewensbesluite en finansiële sake te hanteer (Klitzman et al., 2007; Shannon, 2011). Net so word verslae oor verwante kwessies soos middelmisbruik en verslawing aan internetgebruik ontbreek in die huidige literatuur oor HD-patologie. Op die oomblik is dit onduidelik of die afwesigheid van verslae van dobbelprobleme in die HD-literatuur veroorsaak word deur 'n gebrek aan aandag vir hierdie verskynsel, of of daar werklik geen toename in patologiese dobbelary onder HD-pasiënte is nie. Verskeie redes kan verduidelik waarom sulke probleme nie gereeld aangemeld is nie. Eerstens, selfs al is die voorkoms van patologiese dobbelary in HD verhoog, sal dit waarskynlik nog net 'n klein persentasie pasiënte beïnvloed. In kombinasie met die feit dat die HD-geaffekteerde bevolking self in aantal beperk is, kan dit veroorsaak dat dobbelprobleme onopgemerk word as 'n spesifieke probleem in hierdie pasiëntgroep. Tweedens kan die gebrek aan dobbelprobleme in HD verband hou met die onvermoë of onwilligheid van pasiënte om die huis te verlaat weens motoriese afwykings en dikwels waargeneem tekens van apatie en depressie. Voor die aanvang van internet dobbel, het dit moontlik HD-pasiënte van openbare dobbelplekke soos die casino besoek. Ten slotte blyk adolessensie 'n sensitiewe tydperk vir die ontwikkeling van dobbelprobleme te wees (Van den Bos et al., 2013a), terwyl die meeste HD-pasiënte nie later in die lewe disinhibisieverwante simptome toon nie. Met die opkoms van internetverwante aktiwiteite van adolessente kan hulle egter vorme van ontspanningsgedrag soos aanlyn-dobbel aanleer, wat ontwikkel tot 'n probleem wanneer HD-simptome later in die lewe manifesteer. Alhoewel die omgewing waarin dobbelbewuste HD-pasiënte hulself bevind, moontlik nie so 'n gedrag in die verlede bevorder het nie, is dit duidelik dat 'n groter toeganklikheid en beskikbaarheid van dobbelgeleenthede van die huis die voorkoms van verwante probleme in die HD-bevolking kan verander .
Risikovolle besluitneming deur HD-pasiënte op laboratoriumtaak
Laboratoriumtake word algemeen gebruik om kognitiewe en gedragsafwykings in neurologiese afwykings te assesseer. Om insig te verkry in die prosesse en gestremdhede wat betrokke is by besluitneming en risiko-gedrag, is verskeie take ontwikkel, insluitende die IGT (Bechara et al., 1994) en die CGT (Rogers et al., 1999). Op die IGT word deelnemers aangebied met vier kaarte. Hulle word aangeraai om kaarte uit hierdie dekke te kies, waarmee hulle geld kan wen of verloor; Die doel van die taak is om soveel geld as moontlik te wen. Die dekke verskil van mekaar in die frekwensie en hoeveelheid wins en verliese. Twee hiervan is "slegte" dekke, wat lei tot 'n algehele verlies op die lang termyn, en twee is "goeie" dekke wat tot 'n algehele wins lei. Die deelnemers word egter nie hierdie inligting gegee nie en moet ontdek watter dekke die voordele tydens die eksperimente is. Normale, gesonde, deelnemers leer die reëls van die taak suksesvol na 'n sekere aantal steekproewe, en begin uiteindelik die twee "goeie" dekke verkies. Nietemin is daar beduidende individuele verskille in prestasie selfs onder gesonde deelnemers, insluitend byvoorbeeld duidelike geslagsverskille (Van den Bos et al., 2013b). Op die CGT word deelnemers aangebied met 'n ry 10 bokse van twee verskillende kleure, en moet 'n probabilistiese besluit maak in watter kleurkas 'n teken is verborge. Hulle moet dan kredietpunte op hul vertroue in hierdie besluit speel. In hierdie taak word alle relevante inligting aan die deelnemer tydens die eksperiment voorgelê, en toetse is onafhanklik, wat die werkgeheue en leer eis minimaliseer. Albei dobbelstake is goed gevestig, en die IGT word aanvaar as 'n geldige simulasie van werklike besluitneming (Buelow en Suhr, 2009), terwyl die KWB besonder nuttig is om besluitneming buite 'n leerkonteks te studeer.
HD pasiënte is getoets op beide die Iowa en Cambridge Dobbeltaak. In 'n studie met tussenstadium pasiënte, Stout et al. (2001) bevind dat prestasie op die IGT verminder is in vergelyking met normale vakke. Die verskil in prestasie het duidelik geword in die tweede deel van die taak; waar vakke normaalweg begin om 'n voorkeur vir die goeie dekke te toon, het HD-pasiënte gereeld keuses uit die slegte dekke gemaak. Dit dui daarop dat HD-pasiënte ook nie geleer het watter dekke voordelig was nie, of om kaarte van die slegte dekke te kies ten spyte van hierdie kennis. Die skrywers het opgemerk dat verskeie HD-deelnemers aangedui het dat sommige dekke nadelig was, maar steeds steeds kaarte van daardie dekke gekies het, wat daarop dui dat HD-pasiënte die reëls van die taak kan leer, maar nie 'n voordelige keuringspatroon kan afdwing nie en weerstaan reageer op individuele strawwe en belonings. Nietemin is verminderde prestasie geassosieer met verswakte herinnering en konseptualisering, wat aanleiding gee dat die skrywers bespiegel dat HD-pasiënte probleme ondervind om die langtermyn gevolge van die keuse van kaarte van 'n bepaalde dek te leer of te onthou. HD-pasiënte het ook hoër op disinhibisie as gesonde beheermaatreëls getoon, maar hierdie maatreël is nie met taakverrigting gekorreleer nie. In 'n opvolg van dieselfde data, Stout en kollegas, het drie kognitiewe besluitnemingsmodelle vergelyk om die prestasie tekort van HD pasiënte te verduidelik, en gevind dat dit die beste verklaar is deur tekorte in werkgeheue en deur toenames in roekeloosheid en impulsiwiteit (Busemeyer en Stout, 2002). Verergerde prestasie van HD-pasiënte op die IGT kan ook verband hou met 'n verminderde impak van verliese op hierdie pasiënte, wat gevind is deur die meet van die velgeleidingsreaksies tydens die IGT (Campbell et al., 2004). Hierdie bevinding is in ooreenstemming met verswakte erkenning van negatiewe emosies in HD-pasiënte (Johnson et al., 2007; Ille et al., 2011), en stel voor dat hulle minder sensitief vir groot strawwe is, en daarom minder geneig om weg te draai van die slegte kaartdek. Veral die tweede deel van die IGT vereis die vermoë om benadeelde aksies te onderdruk in reaksie op strawwe, terwyl die bevordering van winsgewende aksies (die Visser et al., 2011; Van den Bos et al., 2013b, 2014).
'N Beperkte aantal ander studies het riskante besluitneming in vroeë stadiums van HD getoets, maar het nie prestasieprobleme by hierdie pasiënte op die IGT of die CGT gevind nie (Watkins et al., 2000; Holl et al., 2013). Dit blyk dus dat waardedaling in besluitneming en die risiko van dobbelprobleme nie ontwikkel tot tussen stadiums van die siekte nie. Hierdie studies het egter gestremdhede in take gevind wat beplanning en remming van voorkragtige reaksies in vroeë HD-pasiënte vereis. Dit blyk dus dat HD-pasiënte eers subtiele probleme met inhibisie, beplanning, emosionele herkenning en werkgeheue ontwikkel. By sommige pasiënte kan dit reeds lei tot probleme met oordeel en besluitneming in vroeë stadiums van die siekte, maar die meeste HD-pasiënte het nie probleme met riskante besluitnemingstake totdat hulle 'n intermediêre stadium van die siekte bereik nie.
Neurobiologiese Meganismes van Besluitneming in HD
Neurobiologiese weë onderliggend aan normale besluitnemingsprosesse in die IGT
Die neurobiologiese meganismes onderliggend aan besluitnemingsprosesse in die IGT is goed bestudeer en beskryf (sien bv. Bechara et al., 2000; Doya, 2008; die Visser et al., 2011; Van den Bos et al., 2013b, 2014). Normale uitvoering van hierdie taak vereis 'n interaksie tussen die limbiese en assosiatiewe / kognitiewe beheer cortico-striatale stroombane. Aktiwiteit in die limbiese kring word vermoedelik dominante tydens die eerste fase van die IGT, waartydens dit betrokke is by verkenningsgedrag, reageer op belonings en straf en die affektiewe waardes van kort- en langtermynuitkomste van besluite in die taak (Manes et al., 2002; Clark en Manes, 2004; Fellows en Farah, 2005; Gleichgerrcht et al., 2010; die Visser et al., 2011; Van den Bos et al., 2014). Die assosiatiewe / kognitiewe beheerbaan is egter meer belangrik tydens die tweede deel van die IGT wanneer impulsiewe reaksies op belonings en strawwe vir langtermynvoordeel onderdruk moet word, voordelige gedragspatrone versterk en negatiewe patrone onderdruk (Manes et al., 2002; Clark en Manes, 2004; Fellows en Farah, 2005; Gleichgerrcht et al., 2010; die Visser et al., 2011; Van den Bos et al., 2014).
Neurobiologiese afwykings in IGT-besluitnemingsprosesse in HD
Aangesien besluitnemingsprosesse in die IGT 'n interaksie tussen limbiese en assosiatiewe / kognitiewe beheer cortico-striatale stroombane behels, is dit nie verbasend dat HD-pasiënte in die uitvoering van hierdie taak benadeel word nie. Een van die waarnemings deur Stout en kollegas is dat die impak van verlies op besluitneming in HD-pasiënte verminder word (Campbell et al., 2004). Dit is in ooreenstemming met bevindings dat hierdie pasiënte benadeel word in die erkenning van negatiewe emosies, en kan verklaar word deur versteurings in die orbitofrontale korteks (Ille et al., 2011). Die orbitofrontale korteks is belangrik vir emosionele prosessering, en word in normale vakke geaktiveer in reaksie op strawwe en belonings in 'n besluitnemingstaak (O'Doherty et al., 2001). Nog 'n bevinding deur Stout et al. (2001) is dat die vertoning van HD-pasiënte op die IGT gekorreleer is met verminderde konseptualisering en langtermyngeheue-maatreëls op die Mattis Dementia Rating Scale. 'N Versuim om te leer of te onthou watter dekke op die langtermyn voordelig is, kan geassosieer word met verminderde aktiwiteit van die assosiatiewe / kognitiewe beheerkring, wat benodig word vir langtermynbeplanning en impulsbeheer (Manes et al., 2002; Clark en Manes, 2004; Fellows en Farah, 2005; Gleichgerrcht et al., 2010). Dit is ook in ooreenstemming met spesifieke tekorte van die indirekte pad in HD, aangesien 'n onlangse studie toon dat die indirekte pad belangrik is vir strafgevoeligheid in 'n versterkings-leertaak (Kravitz et al., 2012; Paton en Louie, 2012). Insensitiwiteit vir die toekomstige gevolge van 'n besluit kan ook veroorsaak word deur ventromediale prefrontale korteks disfunksie, aangesien soortgelyke onsensitiwiteit waargeneem word by pasiënte met skade aan hierdie prefrontale area (Bechara et al., 1994). Dus, verminderde prestasie van HD-pasiënte op die IGT kan veroorsaak word deur 'n kombinasie van disfunksies in cortico-striatale bane wat die orbitofrontale korteks, ventromediale prefrontale korteks en dorsolaterale prefrontale korteks insluit. Dit lei tot verminderde responsiwiteit vir straf in die eerste fase van die taak en die versuim om te leer watter dekke op lang termyn voordelig is, beplan dienooreenkomstig en onderdruk impulsiewe response in die tweede fase van die IGT.
Bespreking
HD en Patologiese Dobbelary: Wat is die risiko's?
Die tipiese skikking van motoriese, emosionele en kognitiewe simptome van HD word veroorsaak deur progressiewe striatale atrofie wat die verskillende cortico-striatale bane beïnvloed. Alhoewel die aanvang en progressie van gedrags- en kognitiewe simptome hoogs heterogeen blyk, word motor- en kognitiewe stroombane tipies vroeg in die siekte getref, terwyl die limbiese kring in 'n later stadium geraak word. Interessant genoeg behels neurobiologiese toereikendheid tot patologiese dobbelary en ander verslawings stoornisse in dieselfde cortico-striatale stroombane wat in HD geraak word. Ten spyte van hierdie opvallende ooreenkomste, is egter in die mediese literatuur HD nie geassosieer met patologiese dobbelary of ander verslawende gedrag nie. Slegs een studie tot dusver het 'n gesin beskryf waarin dobbelprobleme in verskeie HD-geraakde familielede plaasgevind het (De Marchi et al., 1998). Ons bespiegel dat pasiënte se motoriese simptome, sowel as hul ouderdom en sosiale omgewing, tot dusver hulle verhinder het om patologiese dobbelary te ontwikkel, ten spyte van hul verhoogde vatbaarheid vir sulke probleme. Aan die ander kant kan die dikwels gediagnoseerde depressie verwag word om impulsiwiteit en die risiko van dobbelprobleme te verhoog, gebaseer op korrelasiestudies (Clarke, 2006). Nog 'n verduideliking vir die gebrek aan waarnemings van dobbelprobleme in HD kan verband hou met verskille in die onderliggende neuropatologie. Alhoewel die kognitiewe versteurings hoogs gelyk is tussen patologiese spelers en HD-pasiënte, is die emosionele veranderinge van 'n ander aard. Patologiese spelers toon hoofsaaklik 'n verhoogde sensitiwiteit vir belonings, en dring daarop aan om te begin en gaan voort om te dobbel. HD, aan die ander kant, is geassosieer met 'n verminderde sensitiwiteit vir strawwe en negatiewe emosies. Hierdie verskil kan 'n belangrike rede wees waarom HD pasiënte nie 'n groter neiging het om dobbelary te begin of betrokke te raak by ander lonende, verslawende gedrag nie.
Nietemin kan stoornisse in die limbiese cortico-striatale kring van HD-pasiënte steeds riskante besluitneming in situasies met onseker uitkoms bevorder, soos getoon in die IGT (Doya, 2008). Daarbenewens maak die kombinasie van verminderde sensitiwiteit vir straf, versuim om impulsiewe reaksies op onmiddellike belonings te inhibeer, en onvermoë om langtermyn vertraagde belonings te oorweeg en voordelige gedragspatrone dienooreenkomstig te handhaaf, maak dit vir HD-pasiënte moontlik om dobbelprobleme te ontwikkel wanneer hulle 'n situasie wat sulke gedrag bevorder. Kenmerkende probleme van HD-pasiënte met strategie verskuiwing en simptome van kognitiewe onbuigsaamheid en volharding kan bydra tot die progressie van patologiese gedrag in hierdie situasies. Ons stel dus voor dat HD-pasiënte nie 'n groter neiging het om dobbelary of ander verslawende gedrage wat inherent is aan hul neuropatologie, te begin nie, maar dat hulle 'n groter risiko het om 'n verslawing te ontwikkel wanneer hulle dobbelary speel. In ooreenstemming met hierdie idee is daar waargeneem dat pasiënte met 'n frontale letsel impulsief word en dikwels swak besluite neem, maar dat hulle nie 'n verhoogde risikogedrag het nie (Miller, 1992; Bechara et al., 2000). Dit dui daarop dat gestremde besluitneming en risiko-neem of -gedraggedrag nie noodwendig saam voorkom nie, en dat verskillende kombinasies van limbiese en assosiatiewe / kognitiewe beheerskakelstoornisse verskillende effekte op riskante besluitneming en dobbelgedrag kan hê. Ons hipotese sal ook verduidelik waarom HD pasiënte nie waargeneem word om slegter op die CGT te doen nie. Aangesien alle inligting oor kanse en waardes van oorwinnings en verliese voorop staan in hierdie taak, mag HD-pasiënte nie nadelige strategieë ontwikkel nie, omdat hulle nie aktief risiko's soek nie. Dit sal egter in meer gevorderde siektepasiënte getoets moet word.
As HD-pasiënte inderdaad 'n groter risiko het om patologiese dobbelgedrag te ontwikkel wanneer hulle met die toepaslike situasie aangebied word, kan die opkoms van maklik toeganklike internetgokgeleenthede 'n spesifieke risiko vir hierdie pasiëntgroep inhou. Selfs as hulle nie hierdie situasies aktief opspoor nie, is HD-pasiënte nou baie meer geneig om dobbelgeleenthede te bespeur as wat hulle in die verlede was. Dit geld veral vir pasiënte wat die meeste van hul tyd by die huis spandeer as gevolg van hul simptome, waar die internet 'n belangrike manier kan wees om hulle te beset. 'N Hoër waarskynlikheid om aan dobbelgedrag deel te neem, kan dus 'n buitensporige toename in verwante probleme in die HD-bevolking veroorsaak. Ons stel voor dat opsigters bewus moet wees van hierdie moontlike risiko's en verkieslik probeer om te verhoed dat HD-pasiënte van (aanlyn) dobbelaktiwiteite betrokke raak. Daarbenewens beweer ons dat klinici gereeld die risiko en voorkoms van dobbelverwante probleme in die HD-bevolking moet evalueer om gepaste behandeling en leiding aan pasiënte en versorgers te kan gee.
toekomstige rigtings
Benewens epidemiologiese studies om die voorkoms van patologiese dobbelary en ander verslawings in HD te assesseer, kan verskeie rade navorsing voorgestel word om ons begrip van die kwessies wat in hierdie vraestel bespreek word, te verhoog. Eerstens, dit sal interessant wees om prestasie-tekorte op die IGT direk aan stoornisse in cortico-striatale aktiwiteit by HD-pasiënte te koppel. Hiervoor kan HD-pasiënte se breinaktiwiteitspatrone bestudeer word met funksionele magnetiese resonansiebeeldvorming tydens die uitvoer van die IGT, en vergelyk met aktiwiteit in normale vakke. Aktiwiteit in die striatum, dorsolaterale prefrontale korteks en orbitofrontale korteks sal na verwagting in HD-pasiënte verminder word tydens die besluitneming op die IGT.
Om die gedrags- en neurobiologiese aspekte van dobbelgedrag in HD in meer besonderhede te bestudeer, kan tans beskikbare knaagdier siekte modelle gebruik word. Op 'n gedragsvlak kan hierdie diere verwag word om verlaagde prestasie op die IGT te toon, soortgelyk aan menslike pasiënte. Knaagdier weergawes van die IGT is beskikbaar (hersiening: die Visser et al., 2011) en die betrokkenheid van verskillende neuronale strukture in hierdie modelle is goed gekenmerk (die Visser et al., 2011; Van den Bos et al., 2013a, 2014). Dus, sulke eksperimente is haalbaar en kan gekombineer word met 'n grondige analise van onderliggende neuronale veranderinge in knaagdiermodelle van HD met behulp van 'n verskeidenheid tegnieke. Verder, met die aanvang van meer ekologiese geldige navorsingsmetodes en -instrumente om die ontwikkeling van patologiese gedrag te bepaal, kan die risiko vir die ontwikkeling van patologiese dobbelary onder (semi) natuurlike toestande in beide mense en diere bestudeer word (Van den Bos et al., 2013a). Saam sal hierdie studies van dobbelverwante simptome en onderliggende neuropatologie in beide menslike pasiënte en diermodelle van HD ons 'n beter begrip gee van die risiko's wat verband hou met dobbel- en moontlik ander verslawende gedrag in HD en verbeter ons vermoë om te voorsien gepaste behandeling en leiding.
Konflik van belangstelling
Die skrywers verklaar dat die navorsing gedoen is in die afwesigheid van enige kommersiële of finansiële verhoudings wat as 'n potensiële botsing van belange beskou kan word.
Verwysings
Abada, YK, Schreiber, R., en Ellenbroek, B. (2013). Motoriese, emosionele en kognitiewe tekorte in volwasse BACHD-muise: 'n model vir Huntington se siekte. Behav. Brein Res. 238, 243-251. doi: 10.1016 / j.bbr.2012.10.039
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Albin, RL, Young, AB en Penney, JB (1989). Die funksionele anatomie van basale ganglia versteurings. Neigings Neurosci. 12, 366–375. doi: 10.1016/0166-2236(89)90074-x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Alexander, GE, en Crutcher, MD (1990). Funksionele argitektuur van basale ganglia stroombane: neurale substrate van parallelle verwerking. Neigings Neurosci. 13, 266–271. doi: 10.1016/0166-2236(90)90107-l
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Alexander, GE, Crutcher, MD, en DeLong, MR (1990). Basale ganglia-talamokortiese bane: parallelle substrate vir motor-, oculomotoriese, "prefrontale" en "limbiese" funksies. Prog. Brein Res. 85, 119–146. doi: 10.1016/s0079-6123(08)62678-3
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Alexander, GE, DeLong, MR, en Strick, PL (1986). Parallelle organisasie van funksioneel gesegregeerde stroombane wat basale ganglia en korteks verbind. Annu. Ds. Neurosci. 9, 357-381. doi: 10.1146 / annurev.neuro.9.1.357
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Anderson, KE (2011). “Hoofstuk 2 - die siekte van Huntington,” in Handboek van Kliniese Neurologie, eds WJ Weiner en E. Tolosa, hiperkinetiese bewegingsversteurings (Londen: Elsevier), 15-24.
Bechara, A., Damasio, AR, Damasio, H. en Anderson, SW (1994). Onverskilligheid vir toekomstige gevolge as gevolg van skade aan menslike prefrontale korteks. Kognisie 50, 7–15. doi: 10.1016/0010-0277(94)90018-3
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Bechara, A., Damasio, H., en Damasio, AR (2000). Emosie, besluitneming en die orbitofrontale korteks. Cereb. korteks 10, 295-307. doi: 10.1093 / cercor / 10.3.295
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Bittenbender, JB, en Quadfasel, FA (1962). Vaste en aketiese vorme van Huntington's chorea. Boog. Neurol. 7, 275-288. doi: 10.1001 / archneur.1962.04210040027003
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Blackmore, L., Simpson, SA en Crawford, JR (1995). Kognitiewe prestasie in die VK-steekproef van presimptomatiese mense wat die gene vir Huntington se siekte dra. J. Med. Genet. 32, 358-362. doi: 10.1136 / jmg.32.5.358
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Bonelli, RM, en Cummings, JL (2007). Frontale-subkortiese kringe en gedrag. Dialogues Clin. Neurosci. 9, 141-151.
Brand, M., Kalbe, E., Labudda, K., Fujiwara, E., Kessler, J. en Markowitsch, HJ (2005). Besluitneming gestremdhede by pasiënte met patologiese dobbelary. Psigiatrie Res. 133, 91-99. doi: 10.1016 / j.psychres.2004.10.003
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Brevers, D., Bechara, A., Cleeremans, A., and Noël, X. (2013). Iowa-dobbeltaak (IGT): twintig jaar daarna - dobbelstoornis en IGT. Front. Psychol. 4: 665. doi: 10.3389 / fpsyg.2013.00665
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Brevers, D., Cleer Mans, A., Bechara, A., Laloyaux, C., Kornreich, C., Verbanck, P., et al. (2011a). Tydsduur van aandagtig vooroordeel vir dobbel inligting in probleem dobbel. Psychol. Verslaafde. Behav. 25, 675-682. doi: 10.1037 / a0024201
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Brevers, D., Cleer Mans, A., Goudriaan, AE, Bechara, A., Kornreich, C., Verbanck, P., et al. (2012b). Besluitneming onder dubbelsinnigheid, maar nie onder risiko nie, hou verband met die probleem van dobbelary. Psigiatrie Res. 200, 568-574. doi: 10.1016 / j.psychres.2012.03.053
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Brevers, D., Cleer Mans, A., Verbruggen, F., Bechara, A., Kornreich, C., Verbanck, P., et al. (2012a). Impulsiewe aksie, maar impulsiewe keuse bepaal die probleem van dobbelstoornis. PLoS One 7: e50647. doi: 10.1371 / journal.pone.0050647
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Brevers, D., Cleer Mans, A., Tibboel, H., Bechara, A., Kornreich, C., Verbanck, P., et al. (2011b). Verlaagde aanduiding vir dobbel-verwante stimuli in probleemgokkers. J. Behav. En daar. Exp. Psigiatrie 42, 265-269. doi: 10.1016 / j.jbtep.2011.01.005
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Buelow, MT, en Suhr, JA (2009). Konstrueer die geldigheid van die iowa dobbeltaak. Neuropsychol. Op 19, 102–114. doi: 10.1007/s11065-009-9083-4
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Busemeyer, JR, en Stout, JC (2002). 'N Bydrae van kognitiewe besluitmodelle vir kliniese assessering: ontbinding van prestasie op die Bechara-dobbeltaak. Psychol. Evalueer. 14, 253-262. doi: 10.1037 / 1040-3590.14.3.253
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Caine, ED, en Shoulson, I. (1983). Psigiatriese sindrome in Huntington se siekte. Am. J. Psigiatrie 140, 728-733.
Campbell, MC, Stout, JC, en Finn, PR (2004). Verlaagde outonome responsiwiteit vir dobbeltaakverliese in Huntington se siekte. J. Int. Neuropsychol. Soc. 10, 239-245. doi: 10.1017 / s1355617704102105
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Cavedini, P., Riboldi, G., Keller, R., D'Annucci, A., en Bellodi, L. (2002). Voorste lob disfunksie in patologiese dobbelpasiënte. Biol. Psigiatrie 51, 334–341. doi: 10.1016/s0006-3223(01)01227-6
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Clark, L., en Goudriaan, AE (2012). "Neuroimaging in probleem dobbel," in Ensiklopedie van Verslawende Gedrag, Ed PM Miller (Londen: Elsevier).
Clark, L., en Manes, F. (2004). Sosiale en emosionele besluitneming as gevolg van frontale lobbesering. Neurocase 10, 398-403. doi: 10.1080 / 13554790490882799
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Clarke, D. (2006). Impulsiwiteit as 'n bemiddelaar in die verhouding tussen depressie en probleem dobbel. Pers. Individ. Verskil. 40, 5-15. doi: 10.1016 / j.paid.2005.05.008
Craufurd, D., Thompson, JC, en Snowden, JS (2001). Gedragsveranderinge in Huntington siekte. Neuropsieliatrie Neuropsigol. Behav. Neurol. 14, 219-226.
De Boo, GM, Tibben, A., Lanser, JB, Jennekens-Schinkel, A., Hermans, J., Maat-Kievit, A., et al. (1997). Vroeë kognitiewe en motoriese simptome in geïdentifiseerde draers van die geen vir Huntington siekte. Boog. Neurol. 54, 1353-1357. doi: 10.1001 / archneur.1997.00550230030012
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
De Marchi, N., Morris, M., Mennella, R., La Pia, S. en Nestadt, G. (1998). Vereniging van obsessiewe-kompulsiewe versteuring en patologiese dobbelary met Huntington's siekte in 'n Italiaanse stamboom: moontlike assosiasie met die mutasie van Huntington se siekte. Acta Psychiatr. Scand. 97, 62–65. doi: 10.1111/j.1600-0447.1998.tb09964.x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
de Visser, L., Homberg, JR, Mitsogiannis, M., Zeeb, FD, Rivalan, M., Fitoussi, A., et al. (2011). Knaagdier weergawes van die iowa dobbel taak: geleenthede en uitdagings vir die verstaan van besluitneming. Front. Neurosci. 5: 109. doi: 10.3389 / fnins.2011.00109
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Di Chiara, G., en Bassareo, V. (2007). Beloningstelsel en verslawing: wat dopamien doen en nie doen nie. Kur. Opin. Pharmacol. 7, 69-76. doi: 10.1016 / j.coph.2007.02.001
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Dias, R., Robbins, TW, en Roberts, AC (1996). Dissociation in prefrontal cortex of affective and attentional shift. Aard 380, 69-72. doi: 10.1038 / 380069a0
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Dodd, M., Klos, K., Bower, J., Geda, Y., Josephs, K., en Ahlskog, J. (2005). Patologiese dobbelary wat veroorsaak word deur dwelms wat gebruik word om parkinson siekte te behandel. Boog. Neurol. 62, 1377-1381. doi: 10.1001 / archneur.62.9.noc50009
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Doya, K. (2008). Modulators van besluitneming. Nat. Neurosci. 11, 410-416. doi: 10.1038 / nn2077
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Duff, K., Paulsen, JS, Beglinger, LJ, Langbehn, DR en Stout, JC (2007). Psigiatriese simptome in Huntington se siekte voor diagnose: die voorspelling-HD studie. Biol. Psigiatrie 62, 1341-1346. doi: 10.1016 / j.biopsych.2006.11.034
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Duff, K., Paulsen, JS, Beglinger, LJ, Langbehn, DR, Wang, C., Stout, JC, et al. (2010b). "Frontale" gedrag voor die diagnose van Huntington se siekte en sy verhouding tot merkers van siekteprogressie: bewys van vroeë gebrek aan bewustheid. J. Neuropsychiatry Clin. Neurosci. 22, 196-207. doi: 10.1176 / appi.neuropsych.22.2.196
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Duff, K., Paulsen, J., Mills, J., Beglinger, LJ, Moser, DJ, Smith, MM, et al. (2010a). Ligte kognitiewe inkorting in voorgeskrewe Huntington siekte. Neurologie 75, 500–507. doi: 10.1212/wnl.0b013e3181eccfa2
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Dumas, EM, van die Bogaard, SJ, Middelkoop, HA en Roos, RA (2013). 'N Oorsig van kognisie in Huntington se siekte. Front. Biosci. (Schol. Ed.) 5, 1-18. doi: 10.2741 / s355
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Epping, EA, Mills, JA, Beglinger, LJ, Fiedorowicz, JG, Craufurd, D., Smith, MM, et al. (2013). Karakterisering van depressie in prodromale Huntington siekte in die neurobiologiese voorspellers van HD (PREDICT-HD) studie. J. Psychiatr. Res. 47, 1423-1431. doi: 10.1016 / j.jpsychires.2013.05.026
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Faure, A., Höhn, S., Von Hörsten, S., Delatour, B., Raber, K., Le Blanc, P., et al. (2011). Veranderde emosionele en motiverende verwerking in die transgeniese ratmodel vir Huntington se siekte. Neurobiol. Leer. Mem. 95, 92-101. doi: 10.1016 / j.nlm.2010.11.010
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Fellows, LK, en Farah, MJ (2005). Verskillende onderliggende gestremdhede in die besluitneming na aanleiding van ventromediale en dorsolaterale frontale lobbeskadiging by mense. Cereb. korteks 15, 58-63. doi: 10.1093 / cercor / bhh108
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Fielding, SA, Brooks, SP, Klein, A., Bayram-Weston, Z., Jones, L., en Dunnett, SB (2012). Profiele van motoriese en kognitiewe inkorting in die transgeniese ratmodel van Huntington se siekte. Brein Res. Bul. 88, 223-236. doi: 10.1016 / j.brainresbull.2011.09.011
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Fink, KD, Rossignol, J., Crane, AT, Davis, KK, Bavar, AM, Dekorver, NW, et al. (2012). Vroeë kognitiewe disfunksie in die HD 51 CAG transgeniese ratmodel van Huntington se siekte. Behav. Neurosci. 126, 479-487. doi: 10.1037 / a0028028
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Fiorillo, CD, Tobler, PN, en Schultz, W. (2003). Diskrete kodering van beloningswaarskynlikheid en onsekerheid deur dopamienneurone. Wetenskap 299, 1898-1902. doi: 10.1126 / science.1077349
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Folstein, SE, en Folstein, MF (1983). Psigiatriese kenmerke van Huntington se siekte: onlangse benaderings en bevindinge. Psychiatr. Dev. 1, 193-205.
Foroud, T., Siemers, E., Kleindorfer, D., Bill, DJ, Hodes, ME, Norton, JA, et al. (1995). Kognitiewe tellings in draers van Huntington se siekte-gen in vergelyking met nie-draers. Ann. Neurol. 37, 657-664. doi: 10.1002 / ana.410370516
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Giordani, B., Berent, S., Boivin, MJ, Penney, JB, Lehtinen, S., Markel, DS, et al. (1995). Longitudinale neuropsigologiese en genetiese koppelingsanalise van persone met 'n risiko vir Huntington se siekte. Boog. Neurol. 52, 59-64. doi: 10.1001 / archneur.1995.00540250063014
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Gleichgerrcht, E., Ibanez, A., Roca, M., Torralva, T., en Manes, F. (2010). Besluitnemingskognisie in neurodegeneratiewe siektes. Nat. Eerw. Neurol. 6, 611-623. doi: 10.1038 / nrneurol.2010.148
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Goudriaan, AE, Oosterlaan, J., die Beurs, E., en Van den Brink, W. (2004). Patologiese dobbelary: 'n omvattende oorsig van biobehaviorale bevindinge. Neurosci. Biobehav. Op 28, 123-141. doi: 10.1016 / j.neubiorev.2004.03.001
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Graybiel, AM, Aosaki, T., Flaherty, AW, en Kimura, M. (1994). Die basale ganglia en adaptiewe motoriese beheer. Wetenskap 265, 1826-1831. doi: 10.1126 / science.8091209
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Griffiths, M. (2003). Internet dobbel: kwessies, bekommernisse en aanbevelings. Cyberpsychol. Behav. 6, 557-568. doi: 10.1089 / 109493103322725333
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Grimbergen, YAM, Knol, MJ, Bloem, BR, Kremer, BPH, Roos, RAC, en Munneke, M. (2008). Herfs en gangversteurings in Huntington se siekte. Mov. Disord. 23, 970-976. doi: 10.1002 / mds.22003
Haber, SN, en Knutson, B. (2010). Die beloningskring: koppeling van primaatanatomie en menslike beeldvorming. Neuropsigofarmakologie 35, 4-26. doi: 10.1038 / npp.2009.129
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Hadzi, TC, Hendricks, AE, Latourelle, JC, Lunetta, KL, Cupples, LA, Gillis, T., et al. (2012). Assessering van kortikale en striatale betrokkenheid by 523 Huntington-siekte-brein. Neurologie 79, 1708–1715. doi: 10.1212/wnl.0b013e31826e9a5d
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Hahn-Barma, V., Deweer, B., Dürr, A., Dode, C., Feingold, J., Pillon, B., et al. (1998). Is kognitiewe veranderinge die eerste simptome van Huntington se siekte? 'N Studie van geendraers. J. Neurol. Neurochir. Psigiatrie 64, 172-177. doi: 10.1136 / jnnp.64.2.172
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Hamilton, JM, Salmon, DP, Corey-Bloom, J., Gamst, A., Paulsen, JS, Jerkins, S., et al. (2003). Gedragsafwykings dra by tot die funksionele afname in Huntington se siekte. J. Neurol. Neurochir. Psigiatrie 74, 120-122. doi: 10.1136 / jnnp.74.1.120
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Hansotia, P., Cleeland, CS, en Chun, RW (1968). Juvenile Huntington's chorea. Neurologie 18, 217-224.
Heemskerk, A.-W., en Roos, RAC (2011). Dysfagie in Huntington se siekte: 'n oorsig. disfagie 26, 62–66. doi: 10.1007/s00455-010-9302-4
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Henley, SMD, Wild, EJ, Hobbs, NZ, Warren, JD, Frost, C., Scahill, RI, et al. (2008). Defecte emosie herkenning in vroeë HD is neuropsigologies en anatomies generies. Neuropsychologia 46, 2152-2160. doi: 10.1016 / j.neuropsychologia.2008.02.025
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Holl, AK, Wilkinson, L., Tabrizi, SJ, Painold, A., en Jahanshahi, M. (2013). Selektiewe uitvoerende wanfunksie, maar onaktiewe risikobesluitende besluitneming in die vroeë Huntington-siekte. Mov. Disord. 28, 1104-1109. doi: 10.1002 / mds.25388
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Hoth, KF, Paulsen, JS, Moser, DJ, Tranel, D., Clark, LA, en Bechara, A. (2007). Pasiënte met Huntington se siekte het verswakte bewustheid van kognitiewe, emosionele en funksionele vermoëns. J. Clin. Exp. Neuropsychol. 29, 365-376. doi: 10.1080 / 13803390600718958
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Iacono, WG, Malone, SM, en McGue, M. (2008). Gedragsversteuring en die ontwikkeling van vroeëre verslawing: algemene en spesifieke invloede. Annu. Eerw. Clin. Psychol. 4, 325-348. doi: 10.1146 / annurev.clinpsy.4.022007.141157
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Ille, R., Schäfer, A., Scharmüller, W., Enzinger, C., Schöggl, H., Kapfhammer, HP, et al. (2011). Emosie-herkenning en ervaring in Huntington-siekte: 'n voxel-gebaseerde morfometrie studie. J. Psigiatrie Neurosci. 36, 383-390. doi: 10.1503 / jpn.100143
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Jason, GW, Pajurkova, EM, Suchowersky, O., Hewitt, J., Hilbert, C., Reed, J., et al. (1988). Presimptomatiese neuropsigologiese inkorting in Huntington se siekte. Boog. Neurol. 45, 769-773. doi: 10.1001 / archneur.1988.00520310079021
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Johnson, SA, Stout, JC, Solomon, AC, Langbehn, DR, Aylward, EH, Cruce, CB, et al., En Predict-HD Ondersoekers van die Huntington Studiegroep (2007). Beyond disgust: verswakte erkenning van negatiewe emosies voor diagnose in Huntington se siekte. Brein 130, 1732-1744. doi: 10.1093 / brein / awm107
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Jones, L., en Hughes, A. (2011). "Patogene meganismes in Huntington se siekte," in Internasionale oorsig van Neurobiologie, eds J. Brotchie, E. Bezard en P. Jenner, Patofisiologie, Farmakologie en Biochemie van Dyskinesia (Londen: Akademiese Pers), 373-418.
Josiassen, RC, Curry, LM, en Mancall, EL (1983). Ontwikkeling van neuropsigologiese tekorte in Huntington se siekte. Boog. Neurol. 40, 791-796. doi: 10.1001 / archneur.1983.04050120041005
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Julien, CL, Thompson, JC, Wild, S., Yardumian, P., Snowden, JS, Turner, G., et al. (2007). Psigiatriese versteurings in prekliniese Huntington se siekte. J. Neurol. Neurochir. Psigiatrie 78, 939-943. doi: 10.1136 / jnnp.2006.103309
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Kassubek, J., Juengling, FD, Kioschies, T., Henkel, K., Karitzky, J., Kramer, B., et al. (2004). Topografie van serebrale atrofie in vroeë Huntington se siekte: 'n voxel-gebaseerde morfometriese MRI-studie. J. Neurol. Neurochir. Psigiatrie 75, 213-220. doi: 10.1136 / jnnp.2002.009019
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Kirkwood, SC, Siemers, E., Stout, JC, Hodes, ME, Conneally, PM, Christian, JC, et al. (1999). Longitudinale kognitiewe en motoriese veranderinge onder presimptomatiese Huntington siekte geen draers. Boog. Neurol. 56, 563-568. doi: 10.1001 / archneur.56.5.563
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Kirkwood, SC, Su, JL, Conneally, P., en Foroud, T. (2001). Progressie van simptome in die vroeë en middelstadia van Huntington siekte. Boog. Neurol. 58, 273-278. doi: 10.1001 / archneur.58.2.273
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Klitzman, R., Thorne, D., Williamson, J., Chung, W., en Marder, K. (2007). Besluitneming oor reproduktiewe keuses onder individue wat die risiko van Huntington se siekte het. J. Genet. Couns. 16, 347–362. doi: 10.1007/s10897-006-9080-1
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Klöppel, S., Stonnington, CM, Petrovic, P., Mobbs, D., Tüscher, O., Craufurd, D., et al. (2010). Irritasie in prekliniese Huntington se siekte. Neuropsychologia 48, 549-557. doi: 10.1016 / j.neuropsychologia.2009.10.016
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Koller, WC, en Trimble, J. (1985). Die gang-abnormaliteit van Huntington se siekte. Neurologie 35, 1450-1454. doi: 10.1212 / wnl.35.10.1450
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Kravitz, AV, Tye, LD, en Kreitzer, AC (2012). Spesifieke rolle vir direkte en indirekte pasiëntstreekse neurone in versterking. Nat. Neurosci. 15, 816-818. doi: 10.1038 / nn.3100
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Labuschagne, I., Jones, R., Callaghan, J., Whitehead, D., Dumas, EM, Say, MJ, et al. (2013). Emosionele gesigherkenningstekorte en medikasie-effekte in pre-manifestasie deur stadium II Huntington se siekte. Psigiatrie Res. 207, 118-126. doi: 10.1016 / j.psychres.2012.09.022
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Lange, KW, Sahakian, BJ, Quinn, NP, Marsden, CD, en Robbins, TW (1995). Vergelyking van die uitvoerende en visuospatiale geheuefunksie in Huntington se siekte en demensie van Alzheimer-tipe wat ooreenstem met die graad van demensie. J. Neurol. Neurochir. Psigiatrie 58, 598-606. doi: 10.1136 / jnnp.58.5.598
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Lawrence, AD, Sahakian, BJ, Hodges, JR, Rosser, AE, Lange, KW, en Robbins, TW (1996). Uitvoerende en meganiese funksies in die vroeë Huntington-siekte. Brein 119, 1633-1645. doi: 10.1093 / brein / 119.5.1633
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Li, SH en Li, XJ (2004). Huntingtin-proteïen interaksies en die patogenese van Huntington se siekte. Trends Genet. 20, 146-154. doi: 10.1016 / j.tig.2004.01.008
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Louis, ED, Lee, P., Quinn, L., en Marder, K. (1999). Dystonie in Huntington se siekte: voorkoms en kliniese eienskappe. Mov. Disord. 14, 95–101. doi: 10.1002/1531-8257(199901)14:1<95::aid-mds1016>3.0.co;2-8
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Lyketsos, CG, Rosenblatt, A., en Rabins, P. (2004). Verlore frontale sindroom of "Uitvoerende disfunksie sindroom". Psychosomatics 45, 247-255. doi: 10.1176 / appi.psy.45.3.247
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
MacDonald, ME, et al., En Huntington's Disease Collaborative Research Group (1993). 'N Nuwe gen wat 'n trinucleotied herhaling bevat wat op Huntington se siekte-chromosome uitgebrei en onstabiel is. Cell 72, 971–983. doi: 10.1016/0092-8674(93)90585-e
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Manes, F., Sahakian, B., Clark, L., Rogers, R., Antoun, N., Aitken, M., et al. (2002). Besluitnemingsprosesse as gevolg van skade aan die voorfrontale korteks. Brein 125, 624-639. doi: 10.1093 / brein / awf049
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
McAlonan, K., en Brown, VJ (2003). Orbitale prefrontale korteks bemiddel omkeer leer en nie aanstootlike stel verskuiwing in die rot. Behav. Brein Res. 146, 97-103. doi: 10.1016 / j.bbr.2003.09.019
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Miller, LA (1992). Impulsiwiteit, risiko-opname en die vermoë om gefragmenteerde inligting te sintetiseer na frontale lobektomie. Neuropsychologia 30, 69–79. doi: 10.1016/0028-3932(92)90015-e
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Newman, JP (1987). Reaksie op straf in ekstras en psigopate: implikasies vir die impulsiewe gedrag van ontsmettende individue. J. Res. Pers. 21, 464–480. doi: 10.1016/0092-6566(87)90033-x
O'Doherty, J., Kringelbach, ML, Rolls, ET, Hornak, J., en Andrews, C. (2001). Abstrakte beloning en strafvoorstellings in die menslike orbitofrontale korteks. Nat. Neurosci. 4, 95-102. doi: 10.1038 / 82959
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Papp, KV, Kaplan, RF en Snyder, PJ (2011). Biologiese merkers van kognisie in prodromale Huntington se siekte: 'n oorsig. Brein Cogn. 77, 280-291. doi: 10.1016 / j.bandc.2011.07.009
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Paradiso, S., Turner, BM, Paulsen, JS, Jorge, R., Ponto, LLB, en Robinson, RG (2008). Neurale basisse van disfunksie in die vroeë Huntington-siekte. Psigiatrie Res. 162, 73-87. doi: 10.1016 / j.pscychresns.2007.04.001
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Paton, JJ, en Louie, K. (2012). Beloning en straf verlig. Nat. Neurosci. 15, 807-809. doi: 10.1038 / nn.3122
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Paulsen, JS, Langbehn, DR, Stout, JC, Aylward, E., Ross, CA, Nance, M., et al. (2008). Deteksie van Huntington se siekte dekades voor diagnose: die voorspelling-HD studie. J. Neurol. Neurochir. Psigiatrie 79, 874-880. doi: 10.1136 / jnnp.2007.128728
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Peltsch, A., Hoffman, A., Armstrong, I., Pari, G., en Munoz, DP (2008). Saakadiese gestremdhede in Huntington se siekte. Exp. Brein Res. 186, 457–469. doi: 10.1007/s00221-007-1248-x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Potenza, MN (2013). Neurobiologie van dobbelgedrag. Kur. Opin. Neurobiol. 23, 660-667. doi: 10.1016 / j.conb.2013.03.004
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Ramig, LA (1986). Akoestiese ontledings van fonasie by pasiënte met Huntington se siekte. Voorlopige verslag. Ann. Otol. Rhinol. Laryngol. 95, 288-293.
Raylu, N. en Oei, TPS (2002). Patologiese dobbelary. 'N Omvattende hersiening. Clin. Psychol. Op 22, 1009–1061. doi: 10.1016/S0272-7358(02)00101-0
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Reiner, A., Dragatsis, I., en Dietrich, P. (2011). "Genetika en neuropatologie van Huntington se siekte," in Internasionale oorsig van Neurobiologie, eds J. Brotchie, E. Bezard en P. Jenner, Patofisiologie, Farmakologie en Biochemie van Dyskinesia (Londen: Akademiese Pers), 325-372.
Rogers, RD, Everitt, BJ, Baldacchino, A., Black Shaw, AJ, Swainson, R., Wynne, K., et al. (1999). Dissociable deficits in die besluitneming kognisie van chroniese amfetamin misbruikers, opiate misbruik, pasiënte met fokale skade aan prefrontale korteks en tryptofaan-uitgeput normale vrywilligers: bewyse vir mono-aminergiese meganismes. Neuropsigofarmakologie 20, 322–339. doi: 10.1016/s0893-133x(98)00091-8
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Roos, RA (2010). Huntington se siekte: 'n kliniese oorsig. Orphanet J. Rare Dis. 5:40. doi: 10.1186/1750-1172-5-40
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Rosas, HD, Hevelone, ND, Zaleta, AK, Greve, DN, Salat, DH en Fischl, B. (2005). Streeks kortikale verdunning in die prekliniese Huntington-siekte en sy verhouding tot kognisie. Neurologie 65, 745-747. doi: 10.1212 / 01.wnl.0000174432.87383.87
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Rosenblatt, A. (2007). Neuropsigiatrie van Huntington se siekte. Dialogues Clin. Neurosci. 9, 191-197.
Rothlind, JC, Bylsma, FW, Peyser, C., Folstein, SE en Brandt, J. (1993). Kognitiewe en motoriese korrelate van alledaagse funksionering in die vroeë Huntington-siekte. J. Nerv. Ment. Dis. 181, 194–199. doi: 10.1097/00005053-199303000-00008
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Rushworth, MFS, Behrens, TEJ, Rudebeck, PH en Walton, ME (2007). Kontrasterende rolle vir cingulate en orbitofrontale korteks in besluite en sosiale gedrag. Neigings Cogn. Sci. 11, 168-176. doi: 10.1016 / j.tics.2007.01.004
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Sánchez-Castañeda, C., Cherubini, A., Elifani, F., Peran, P., Orobello, S., Capelli, G., et al. (2013). Op soek na huntington siekte biomarkers deur multimodale, dwarsdeursnee basale ganglia beelding. Neurie. Brein Mapp. 34, 1625-1635. doi: 10.1002 / hbm.22019
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Sesack, SR, en Grace, AA (2010). Cortico-basale ganglia beloning netwerk: microcircuitry. Neuropsigofarmakologie 35, 27-47. doi: 10.1038 / npp.2009.93
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Shannon, KM (2011). “Hoofstuk 1 - Huntington-siekte - kliniese tekens, simptome, presimptomatiese diagnose en diagnose,” in Handboek van Kliniese Neurologie, eds WJ Weiner en E. Tolosa, hiperkinetiese bewegingsversteurings (Londen: Elsevier), 3-13.
Sharpe, L. (2002). 'N Gereformuleerde kognitiewe gedragsmodel van probleem dobbel. 'N Biopsigososiale perspektief. Clin. Psychol. Op 22, 1–25. doi: 10.1016/s0272-7358(00)00087-8
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Shiwach, R. (1994). Psigopatologie in Huntington se siekte pasiënte. Acta Psychiatr. Scand. 90, 241–246. doi: 10.1111/j.1600-0447.1994.tb01587.x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Smith, MA, Brandt, J., en Shadmehr, R. (2000). Motoriese afwyking in Huntington se siekte begin as 'n disfunksie met foutiewe terugvoerbeheer. Aard 403, 544-549. doi: 10.1038 / 35000576
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Snell, RG, MacMillan, JC, Cheadle, JP, Fenton, I., Lazarou, LP, Davies, P., et al. (1993). Verhouding tussen trinucleotied herhaling uitbreiding en fenotipiese variasie in Huntington se siekte. Nat. Genet. 4, 393-397. doi: 10.1038 / ng0893-393
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Stine, OC, Pleasant, N., Franz, ML, Abbott, MH, Folstein, SE, en Ross, CA (1993). Korrelasie tussen die aanvangsleeftijd van Huntington se siekte en die lengte van die trinucleotied herhaling in IT-15. Neurie. Mol. Genet. 2, 1547-1549. doi: 10.1093 / hmg / 2.10.1547
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Stout, JC, Paulsen, JS, Queller, S., Solomon, AC, Whitlock, KB, Campbell, JC, et al. (2011). Neurokognitiewe tekens in prodromale Huntington siekte. neuropsigologie 25, 1-14. doi: 10.1037 / a0020937
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Stout, JC, Rodawalt, WC, en Siemers, ER (2001). Risikovolle besluitneming in Huntington se siekte. J. Int. Neuropsychol. Soc. 7, 92-101. doi: 10.1017 / S1355617701711095
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Tabrizi, SJ, Langbehn, DR, Leavitt, BR, Roos, RA, Durr, A., Craufurd, D., et al. (2009). Biologiese en kliniese manifestasies van Huntington se siekte in die longitudinale TRACK-HD studie: deursnee-analise van basislyn data. Lancet Neurol. 8, 791–801. doi: 10.1016/S1474-4422(09)70170-X
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Tekin, S. en Cummings, JL (2002). Frontale-subkortiese neuronale bane en kliniese neuropsigiatrie: 'n opdatering. J. Psychosom. Res. 53, 647–654. doi: 10.1016/s0022-3999(02)00428-2
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Thieben, MJ, Duggins, AJ, Good, CD, Gomes, L., Mahant, N., Richards, F., et al. (2002). Die verspreiding van strukturele neuropatologie in prekliniese Huntington se siekte. Brein 125, 1815-1828. doi: 10.1093 / brein / awf179
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Tian, J., Herdman, SJ, Zee, DS, en Folstein, SE (1992). Posturale stabiliteit by pasiënte met Huntington se siekte. Neurologie 42, 1232-1238. doi: 10.1212 / wnl.42.6.1232
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Unschuld, PG, Joel, SE, Pekar, JJ, Reading, SA, Oishi, K., McEntee, J., et al. (2012). Depressiewe simptome in prodromale Huntington se siekte korreleer met stroop-interferensie verwante funksionele konnektiwiteit in die ventromediale prefrontale korteks. Psigiatrie Res. 203, 166-174. doi: 10.1016 / j.pscychresns.2012.01.002
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van den Bogaard, SJ, Dumas, EM, Acharya, TP, Johnson, H., Langbehn, DR, Scahill, RI, et al. (2011). Vroeë atrofie van pallidum en accumbens kern in Huntington se siekte. J. Neurol. 258, 412–420. doi: 10.1007/s00415-010-5768-0
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van den Bos, R., Davies, W., Dellu-Hagedorn, F., Goudriaan, AE, Granon, S., Homberg, J., et al. (2013a). Kruis-spesies benaderings tot patologiese dobbelary: 'n oorsig wat seksverskille, adolessente kwesbaarheid en ekologiese geldigheid van navorsingshulpmiddels rig. Neurosci. Biobehav. Op 37, 2454-2471. doi: 10.1016 / j.neubiorev.2013.07.005
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van den Bos, R., Homberg, J., en de Visser, L. (2013b). 'N Kritiese oorsig van geslagsverskille in besluitnemingstake: fokus op die iowa-dobbeltaak. Behav. Brein Res. 238, 95-108. doi: 10.1016 / j.bbr.2012.10.002
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van die Bos, R., Koot, S., en de Visser, L. (2014). 'N knaagdier weergawe van die iowa dobbel taak: 7 jaar van vordering. Front. Psychol. 5: 203.doi: 10.3389 / fpsyg.2014.00203
Van Duijn, E., Kingma, EM, en Van der Mast, RC (2007). Psigopatologie in geverifieerde Huntington se siekte geen draers. J. Neuropsychiatry Clin. Neurosci. 19, 441-448. doi: 10.1176 / appi.neuropsych.19.4.441
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van Duijn, E., Reedeker, N., Giltay, EJ, Eindhoven, D., Roos, RAC, en van der Mast, RC (2014). Kursus van geïrriteerdheid, depressie en apatie in Huntington se siekte met betrekking tot motoriese simptome gedurende 'n twee jaar opvolgperiode. Neurodegener. Dis. 13, 9-16. doi: 10.1159 / 000343210
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van Holst, RJ, Van den Brink, W., Veltman, DJ, en Goudriaan, AE (2010). Waarom dobbelaars nie wen nie: 'n oorsig van kognitiewe en neuroimaging-bevindinge in patologiese dobbelary. Neurosci. Biobehav. Op 34, 87-107. doi: 10.1016 / j.neubiorev.2009.07.007
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Van Holst, RJ, Veltman, DJ, Büchel, C., Van den Brink, W., en Goudriaan, AE (2012). Verwronge verwagte kodering in probleem dobbelary: is die verslawende in afwagting? Biol. Psigiatrie 71, 741-748. doi: 10.1016 / j.biopsych.2011.12.030
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Verny, C., Allain, P., Prudean, A., Malinge, MC, Gohier, B., Scherer, C., et al. (2007). Kognitiewe veranderinge in asimptomatiese draers van die Huntington-siekte mutasie geen. EUR. J. Neurol. 14, 1344-1350. doi: 10.1111 / j.1468-1331.2007.01975.x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Volkow, ND, Fowler, JS, en Wang, GJ (2002). Rol van dopamien in dwelmversterking en verslawing by die mens: resultate van beeldstudie. Behav. Pharmacol. 13, 355–366. doi: 10.1097/00008877-200209000-00008
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Vonsattel, JP, en DiFiglia, M. (1998). Huntington siekte. J. Neuropathol. Exp. Neurol. 57, 369–384. doi: 10.1097/00005072-199805000-00001
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Vonsattel, JPG, Keller, C. en Cortes Ramirez, EP (2011). Hoofstuk 4 - Huntington se siekte - neuropatologie ”in Handboek van Kliniese Neurologie, eds W. Weiner en E. Tolosa, hiperkinetiese bewegingsversteurings (Londen: Elsevier), 83-100.
Vonsattel, JPG (2008). Huntington siekte modelle en menslike neuropatologie: ooreenkomste en verskille. Acta Neuropathol. 115, 55–69. doi: 10.1007/s00401-007-0306-6
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Watkins, LHA, Rogers, RD, Lawrence, AD, Sahakian, BJ, Rosser, AE en Robbins, TW (2000). Verswakte beplanning, maar ongeskonde besluitneming in die vroeë Huntington-siekte: implikasies vir spesifieke fronto-stratale patologie. Neuropsychologia 38, 1112–1125. doi: 10.1016/s0028-3932(00)00028-2
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Weintraub, D., Papay, K., Siderowf, A., en Parkinson se Progression Markers Initiative. (2013). Sifting vir impulsbeheersymptome by pasiënte met de novo Parkinson-siekte: 'n gevallestudie-studie. Neurologie 80, 176–180. doi: 10.1212/wnl.0b013e31827b915c
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Witjas, T., Eusebio, A., Fluchère, F., en Azulay, JP (2012). Verslawende gedrag en Parkinson se siekte. Eerw. Neurol. (Parys) 168, 624-633. doi: 10.1016 / j.neurol.2012.06.014
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Wolf, RC, Vasic, N., Schönfeldt-Lecuona, C., Landwehrmeyer, GB, en Ecker, D. (2007). Dorsolaterale prefrontale korteks disfunksie in presimptomatiese Huntington se siekte: bewyse uit gebeurtenisverwante fMRI. Brein 130, 2845-2857. doi: 10.1093 / brein / awm210
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Yin, HH, en Knowlton, BJ (2006). Die rol van die basale ganglia in gewoontevorming. Nat. Ds. Neurosci. 7, 464-476. doi: 10.1038 / nrn1919
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Yin, HH, Ostlund, SB, en Balleine, BW (2008). Beloningsgeleide leer buite dopamien in die kernklem: die integrerende funksies van kortikobasale ganglia-netwerke. EUR. J. Neurosci. 28, 1437-1448. doi: 10.1111 / j.1460-9568.2008.06422.x
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Young, AB, Shoulson, I., Penney, JB, Starosta-Rubinstein, S., Gomez, F., Travers, H., et al. (1986). Huntington se siekte in Venezuela Neurologiese kenmerke en funksionele afname. Neurologie 36, 244-249. doi: 10.1212 / WNL.36.2.244
Pubmed Abstract | Pubmed Volledige teks | CrossRef Volledige teks
Sleutelwoorde: Huntington se siekte, risiko-neem, dobbel, prefrontale korteks, basale ganglia, disinhibtie
Aanhaling: Kalkhoven C, Sennef C, Peeters A en van den Bos R (2014) Risiko-opname en patologiese dobbelgedrag in Huntington se siekte. Front. Behav. Neurosci. 8: 103. doi: 10.3389 / fnbeh.2014.00103
Ontvang: 30 November 2013; Papier hangende gepubliseer: 18 Januarie 2014;
Aanvaar: 12 Maart 2014; Gepubliseer aanlyn: 02 April 2014.
Geredigeer deur:
Patrick Anselme, Universiteit van Luik, België
Nagesien deur:
Damien Brevers, Université Libre de Bruxelles, België
Bryan F. Sanger, Universiteit van Michigan, VSA
