أمامي. الطب النفسي ، 25 فبراير 2016 | http://dx.doi.org/10.3389/fpsyt.2016.00024
جاريد غودمان و مارك جي باكارد*
- قسم علم النفس ، معهد تكساس إيه آند إم لعلم الأعصاب ، جامعة تكساس إيه آند إم ، كوليج ستيشن ، تكساس ، الولايات المتحدة الأمريكية
وقد حظي الرأي القائل بأن أنظمة الذاكرة المستقلة تشريحيا بشكل تفاضلي تسهم في تطوير الإدمان على المخدرات والانتكاس دعم واسع النطاق. هذه المراجعة الوجيزة تعيد النظر في هذه الفرضية كما اقترح في الأصل 20 منذ سنوات (1) ويسلط الضوء على العديد من التطورات الأخيرة. تشير الأبحاث المكثفة التي تستخدم مجموعة متنوعة من نماذج التعلم الحيواني إلى أن الأنظمة العصبية القابلة للفصل تتوسط في أنواع متميزة من التعلم والذاكرة. من المحتمل أن يساهم كل نظام ذاكرة بمكونات فريدة للسلوك المكتسب الداعم لإدمان المخدرات والانتكاس. على وجه الخصوص ، قد يعكس التحول من استخدام العقاقير الترويحية إلى تعاطي المخدرات القهري تحولًا عصبيًا من السيطرة الإدراكية على السلوك الذي يتوسطه مخطط الحصين / الظهارة النصفية نحو التحكم المعتاد في السلوك الذي يتوسطه المخطط الظهاري المجهري (DLS). بالإضافة إلى ذلك ، قد يشكل الإجهاد / القلق عامل مساعد ييسر الذاكرة المعتمِدة على DLS ، وقد يستخدم هذا كآلية السلوكيات العصبية الكامنة وراء زيادة تعاطي المخدرات والانتكاس في البشر بعد أحداث الحياة المجهدة. إن الأدلة التي تدعم النظرة المتعددة للأنظمة على إدمان المخدرات تأتي في الغالب من دراسات التعلم والذاكرة التي استخدمت كمعززات للمواد الإدمانية التي غالبًا ما تُعتبر ضمن سياق أبحاث إدمان المخدرات ، بما في ذلك الكوكايين والكحول والأمفيتامينات. بالإضافة إلى ذلك ، تشير الأدلة الحديثة إلى أن نهج أنظمة الذاكرة قد يكون مفيدًا أيضًا في فهم المصادر الموضعية للإدمان والتي تعكس المخاوف الصحية الناشئة ، بما في ذلك استخدام الماريجوانا ، والنظام الغذائي عالي الدهون ، واللعب بألعاب الفيديو.
المُقدّمة
غالباً ما ينظر الباحثون إلى آليات التعلم والسلوك لشرح كيفية اكتساب وعيوب علم النفس البشري. تم تقديم مثال على هذا التطبيق من قبل نورمان م. وايت الذي استخدم مبادئ نظرية التعلم الكلاسيكية والأدلة التجريبية التي تدعم وجود أنظمة ذاكرة متعددة في الدماغ لتوفير نهج جديد ومؤثر لإدمان المخدرات (1). وعلى وجه التحديد ، أشار وايت إلى أن الأدوية يمكن أن تلعب دور "المعززات" التي ، مثل الغذاء أو الماء في مهمة تعلم ، تقوي الروابط بين المنبهات والسياقات والسلوك المتعلق بالمخدرات لتشجيع تعاطي المخدرات ، وعلى مدار الوقت ، الإدمان. كما أدرج وايت الفرضية الناشئة بأن هناك أنواعًا مختلفة من الذاكرة تتوسطها أنظمة عصبية مفصولة. ووفقًا لهذا الرأي الجديد ، يمكن للأدوية أن تعدل بشكل مباشر الأنظمة العصبية المتعددة ، وهذه الأنظمة العصبية تعمل على تشفير مكونات متميزة للذاكرة المتعلقة بالمخدرات والتي ، عند التعبير عنها ، تشجع على المزيد من تعاطي المخدرات.
يصادف العام 2016 الذكرى السنوية 20th لنظم أنظمة الذاكرة المتعددة لإدمان المخدرات كما وصفها White. يقوم هذا الاستعراض بإعادة النظر في هذه الفرضية المؤثرة ، مع تسليط الضوء على بعض التطورات الحديثة المهمة التي لم تثبت فقط الفرضية الأصلية فحسب ، بل أسفرت أيضًا عن رؤى إضافية حول كيفية دعم أنظمة الذاكرة المتعددة لإدمان المخدرات.
عرض أنظمة الذاكرة المتعددة للإدمان
تشير الأدلة المتقاربة من الدراسات التي تستخدم البشر والحيوانات الدنيا إلى أن ذاكرة الثدييات تتوسطها أنظمة عصبية مستقلة نسبيا [للمراجعات ، انظر المرجع. (2-4)]. تم إجراء التجارب المبكرة لفصل أنظمة الذاكرة المتعددة بشكل أساسي في المتاهة الشعاعية وأوضحت وظائف فريدة للذاكرة في قرن آمون ، المخطط الظهري ، اللوزة المخية (5, 6). يتوسط الحصين شكلًا معرفيًا / مكانيًا للذاكرة ، في حين يتوسط المخطط الظهري ذاكرة العادة التحفيز والاستجابة (S – R). تتوسط اللوزة بوساطة العلاقات بين بافلوفان والعلاقات التحفيزية والترابطية (6, 7) ، في حين أيضا تحتل الدور التنظيمي من الإثارة العاطفية على أنواع أخرى من الذاكرة (8-12).
ضمن سياق عرض الأنظمة المتعددة للذاكرة ، الأبيض (1) اقترح أن قرن آمون ، المخطط الظهري ، واللوزة تشفر مكونات فريدة من الذكريات المتعلقة بالمخدرات (انظر الشكل 1). يشفر قرن آمون المعرفة الصريحة المتعلقة بالعلاقة بين الإشارات والأحداث (أي رابطات التحفيز - المنبه) في سياق العقار. والأهم من ذلك ، أن الحصين لا يشفر الاستجابات السلوكية ، بل بالأحرى يمكن استخدام المعلومات التي يتم الحصول عليها من قبل قرن آمون لتوليد الاستجابات السلوكية المناسبة لتلقي تعزيز الأدوية. من ناحية أخرى ، يشكّل المخطط الظهري الارتباطات بين المنبهات المرتبطة بالمخدرات والاستجابات السلوكية. قد يسمح هذا بعرض تقديم جديلة مرتبطة بالمخدرات لتفعيل استجابة سلوكية تلقائية تؤدي إلى تعاطي المخدرات (على سبيل المثال ، طريقة التشغيل أو الضغط على ذراع آلية). ترمز اللوزة إلى علاقات بافلوفين-الارتباطية ، مما يسمح بأن ترتبط الإشارات الحيادية في سياق الدواء بمكافأة الدواء. تتفاعل الحيوانات في وقت لاحق مع هذه الإشارات الشرعية بشكل مشابه لكيفية تفاعلها في الأصل مع الدواء. على وجه التحديد ، تنشط الإشارات الشرطية الاستجابات العاطفية المكيفة ، بما في ذلك الحالات العاطفية الداخلية والنهج المشروط تجاه (أو في بعض الحالات تجنب من) الإشارات المشروطة. هناك عنصر آخر حاسم في فرضية وايت هو أن الأدوية يمكن أن تعدل وظيفة الذاكرة لكل منطقة من مناطق الدماغ هذه. وهكذا ، يمكن للأدوية تعزيز قدرتها على الإدارة الذاتية عن طريق زيادة توحيد الذكريات المتعلقة بالمخدرات المشفرة من قبل قرن آمون ، اللوزة ، والمخطط الظهري (انظر الشكل 1).
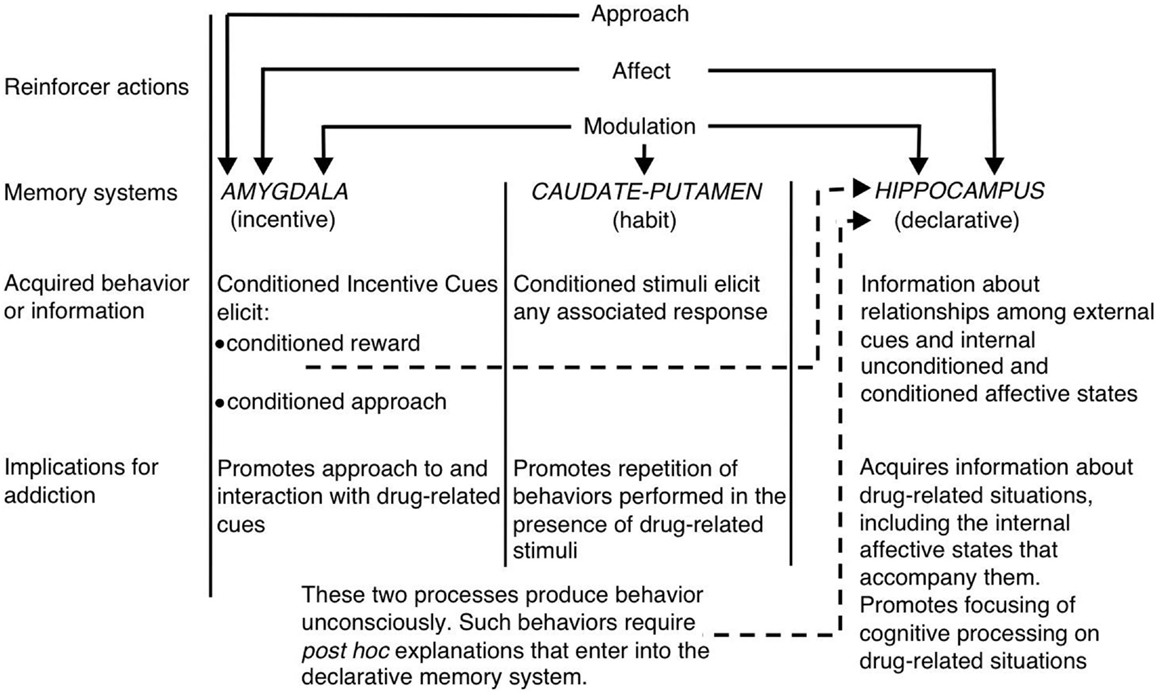
الشكل 1. الأبيض (1) عرض أنظمة ذاكرة متعددة لإدمان المخدرات. مثل المعززات الطبيعية ، تمتلك العقاقير المسببة للإدمان العديد من "الإجراءات المعززة" ، بما في ذلك القدرة على استدعاء التأثير الإيجابي / السلبي ، والنهج ، وتعديل أنظمة الذاكرة. تتوسط كل من اللوزة المخية والبوتامين المذنبة (أي المخطط الظهري) والحصين أنظمة الذاكرة القابلة للفصل ، ويفترض أن كل نظام ذاكرة يشفر مكونات فريدة من الذكريات المتعلقة بالعقاقير. نظرًا لخصائصها المعدلة للذاكرة ، يمكن للعقاقير المسببة للإدمان أن تعزز إدارتها الذاتية من خلال تعزيز وظيفة هذه الأنظمة. (أعيد طبعه من White بإذن من John Wiley & Sons.)
تماشيا مع وجهة نظر أنظمة الذاكرة المتعددة لإدمان المخدرات ، تشير الدلائل الواسعة إلى أدوار حاسمة في قرن آمون ، المخطط الظهري ، واللوزة في إدمان المخدرات والانتكاس لمجموعة متنوعة من المواد الضارة [للمراجعة ، انظر المرجع. (13)]. يبدو أن الحصين الظهري له دور في التحكم السياقي في البحث عن عقار للكوكايين (14-16). تتوسط المنطقة الجانبية من المخطط الظهري (DLS) ذراع S-R المعتاد للضغط على الكوكايين والكحول (17, 18) ، و amygdala basolateral (BLA) يتوسط المخدرات مشروطة البحث عن الكوكايين والكحول والهيروين (19-22). كما تتفق مع فرضية وايت ، يمكن للمواد من سوء المعاملة تعديل وظائف ذاكري من قرن آمون ، المخطط الظهري ، واللوزة (23-31).
ساهمت الدراسات الحديثة في إدخال تعديلات جديدة على نهج أنظمة الذاكرة المتعددة لإدمان المخدرات. تتضمن الميزات الأساسية لهذا العرض المعاصر (1) تحولًا عصبيًا مع مرور الوقت إلى ذاكرة العادة المعتمدة على DLS ، (2) التفاعلات التنافسية بين أنظمة الذاكرة ، (3) دور الإجهاد والقلق في تعزيز البحث المعتاد عن العقاقير ، و (4) تطبيق هذه الفرضية على مصادر جديدة ناشئة من الإدمان.
التحول العصبي من الإدراك إلى العادة
في حالات التعلم التجريبية ، تستخدم الموضوعات عادة السلوك الهادف عند حل إحدى المهام في البداية. ومع ذلك ، بعد التدريب المكثف ، يصبح السلوك مستقلاً ويمكن إجراؤه بقليل من الاهتمام أو النية أو المجهود الإدراكي ، مما يشكل "عادة" [للمراجعة ، راجع المرجع. (32)]. في مظاهرات مبكرة لهذا التحول من السيطرة الإدراكية على السلوك إلى العادة ، تم تدريب القوارض باستخدام مكافأة الطعام في مهمة متاهة مزدوجة الحل (33-35). في هذه المهمة ، تم إطلاق الفئران من نفس وضعية البدء (على سبيل المثال ذراع الجنوب) واضطرت إلى إحداث تحكُّم متناسق في تقاطع المتاهة لتلقي مكافأة الطعام دائمًا في ذراع الهدف نفسه (على سبيل المثال ، اجعل اليسار دائمًا اتجه إلى العثور على الطعام في الذراع الغربية). يمكن للفئران أن تحل هذه المهمة من خلال تعلم استجابة ثابتة للجسم أو عن طريق اتخاذ أي استجابة ضرورية للذهاب إلى نفس الموقع المكاني. ولتحديد الإستراتيجية التي تتبعها الجرذان ، قام الباحثون بتنفيذ اختبار اختبار حيث تم إطلاق الحيوانات من ذراع البداية المعاكس (على سبيل المثال ، الذراع الشمالي). إذا كانت الحيوانات تتخذ المنعطف المعاكس للانتقال إلى موقع الهدف الأصلي ، فقد تم تحديدهم كمتعلمين مكان. إذا صنعت الحيوانات نفس منعطف الجسم أثناء التدريب (أي الذهاب إلى الذراع المقابلة لموقع الهدف الأصلي) ، فقد تم تحديد الحيوانات على أنها متعلمة استجابة. تشير الأدلة إلى أنه بعد بعض التدريب ، تعرض معظم الحيوانات التعلم في المكان ، بينما بعد تدريب مكثف ، تتحول الحيوانات إلى التعلم بالاستجابة المعتادة (34-36). ومن المثير للاهتمام أن هذا التحول من التعلم بالمكان إلى التعلم استجابة قد يعكس التحول العصبي. يتم توسط الاستخدام الأولي للتعلم في مكان في هذه المهمة من قبل الحصين والظهراني المخطط [DMS (36, 37)] ، في حين أن استخدام التعلم استجابة بعد تدريب الموسعة بوساطة DLS (36).
بالإضافة إلى المظاهرات المبكرة باستخدام متاهة زائد (34, 35) ، وقد تبين في وقت لاحق التحول السلوكي إلى الذاكرة الاعتيادية باستخدام نماذج الضغط رافعة رافعة فعالة (38-42). في هذه المهام التعليمية المفيدة ، تقوم الحيوانات في البداية بالضغط بشكل هادف من أجل الحصول على النتيجة وسوف تتوقف عن الضغط على الرافعة بمجرد تخفيض قيمة الغذاء. ومع ذلك ، بعد الحيوانات تدريب واسعة النطاق سوف تتحول إلى الاستجابة المعتادة وسوف تستمر في الضغط على رافعة حتى بعد أن تم تخفيض قيمة الغذاء (40). كما هو موضح في الأصل في متاهة زائد (36) ، يمكن أيضًا أن يعزى الانتقال من الإدراك إلى العادة في مهام التعلم الآلية إلى التحول العصبي التشريحي. يتم التحكم في السيطرة الإدراكية الأولية للسلوك في هذه المهام التعليمية الأساسية من قبل الحصين و DMS (43, 44) ، في حين أن الاستجابة المعتادة في وقت لاحق يتم بوساطة DLS (18, 45, 46).
اقترح العديد من الباحثين أن التحول العصبي إلى الذاكرة الاعتيادية الظاهرة في المتاهة ومهام التعلم الأساسية قد يكمن أيضا في التحول من استخدام المخدرات الترويحي إلى تعاطي المخدرات القهري (13, 47-50). بالتوافق مع هذه الفرضية ، أظهر المحققون مجموعة متنوعة من المواد المتعرضة للإساءة التي يتوسط بها نظام إدارة الوجهات السياحية استجابة موجهة نحو الهدف من أجل تعزيز العقاقير وتوسيط DLS في الاستجابة المعتادة لتعاطي المخدرات (18, 31, 51-53).
بالنظر إلى الإمكانات العالية لإساءة استخدام بعض الأدوية ، اقترح الباحثون أن العقاقير المسببة للإدمان قد تعزز وظيفة الذاكرة المعتمدة على DLS وبالتالي تسرع التحول من السيطرة السلوكية على الإدراك إلى السلوك المعتاد. بما يتفق مع هذه الفرضية ، فإن التعرض المتكرر للأمفيتامين أو الكوكايين يسهل التحول من الهدف الموجه إلى الاستجابة المعتادة لتعزيز الغذاء في مهام ضغط الرافعة الآلية (31, 54-59). بالإضافة إلى ذلك ، فإن الضغط بالضغط على المواد المسببة للإدمان (مثل الكحول أو الكوكايين) مقابل المكافأة الغذائية قد ارتبط بزيادة الاستجابة المعتادة مقابل الاستجابة الموجهة نحو الأهداف (24, 60, 61). في البشر ، يظهر الأفراد الذين يعتمدون على الكحول استجابة أكثر اعتدالًا في مهمة تعلم مفيدة ، مقارنةً بالأفراد غير المعتمدين على السيطرة (62). وقد لوحظ هذا تعزيز الذاكرة العادة تعتمد على DLS من المخدرات الإدمانية في مهام التعلم المتاهة القوارض. ارتبط الكوكايين والأمفيتامين والتعرض للكحول بالتعليم المعزز في مهام متاهة تعتمد على DLS أو استخدام أكبر لاستراتيجيات الاستجابة المعتمدة على DLS في إصدارات الحل المزدوج للمتاهة (25, 63, 64). في البشر ، ارتبط استخدام المواد المعتادة ، بما في ذلك الكحول والتبغ ، بزيادة استخدام الاستراتيجيات الملاحية الظهارية المعتمدة على المخططات في متاهة افتراضية (65). وهكذا ، فإن بعض تعاطي المخدرات قد يعزز ذاكرة العادة المعتمدة على DLS ، وهذا الانخراط المتزايد لنظام الذاكرة DLS قد يعجل الانتقال من استخدام المخدرات الترويحي إلى تعاطي المخدرات المعتاد. هذه الآلية المقترحة تتفق مع الأبيض (1) اﻻدﻋﺎء اﻷﺻﻠﻲ ﺑﺄن ﻋﻘﺎﻗﻴﺮ اﻹﺳﺎءة ﻗﺪ ﺗﺴﻬﻞ ﻓﻲ ﺑﻌﺾ اﻷﺣﻴﺎن إدارﺗﻬﺎ اﻟﺬاﺗﻴﺔ ﻣﻦ ﺧﻼل ﺗﻌﺰﻳﺰ وﻇﺎﺋﻒ أﻧﻈﻤﺔ اﻟﺬاآﺮة
التنافس بين أنظمة الذاكرة
على الرغم من أنه من الممكن أن المخدرات الإدمانية تعزيز الذاكرة العادة مباشرة عن طريق تعزيز وظيفة DLS [على سبيل المثال ، المرجع. (29)] ، وهناك احتمال آخر هو أن تعاطي المخدرات يعزز عادة الذاكرة بشكل غير مباشر عن طريق تعديل أنظمة الذاكرة الأخرى. تستدعي هذه الآلية البديلة الفرضية القائلة بأن أنظمة الذاكرة تتنافس في بعض حالات التعلم من أجل التحكم في التعلم ، ومن خلال إضعاف وظيفة نظام ذاكرة واحد ، يمكن تحسين وظيفة نظام سليم آخر (11, 66). بشكل ملحوظ ، قد يتنافس قرن آمون و DLS في بعض الأحيان من أجل السيطرة على التعلم ، حيث تعمل آفة الحصين على تعزيز وظيفة الذاكرة المعتمدة على DLS (5, 6, 67, 68). يمكن أيضًا إظهار التفاعلات التنافسية في مهام الحل المزدوج ، عندما يؤدي ضعف نظام ذاكرة واحد إلى استخدام استراتيجية بوساطة نظام سليم آخر. على سبيل المثال ، تعرض الحيوانات التي تعطى آفات DMS الاستجابة المعتادة على DLS للاستجابة لمكافأة الطعام في مهام التعلم الأساسية (44).
بالنظر إلى التفاعلات التنافسية التي تنشأ في بعض الأحيان بين أنظمة الذاكرة ، هناك احتمال واحد هو أن بعض تعاطي المخدرات قد يعزز ذاكرة العادة المعتمدة على DLS بشكل غير مباشر من خلال إضعاف آليات الذاكرة المعرفية التي توسط بها DMS و hippocampus. كما ذكرنا سابقًا ، يرتبط الكحول بزيادة استخدام ذاكرة العادة المعتمدة على DLS في المتاهة ونماذج ضغط الذراع النشطة (24, 61, 62, 64, 65). تشير الأدلة أيضا إلى أن الكحول يضعف التعلم في مهام الذاكرة المكانية المعتمدة على قرن آمون [(64, 69-72)؛ للمراجعة ، راجع المرجع. (73)] ، وكذلك في مهام تعلم الانعكاس التي تعتمد على DMS (74-77). بالتوافق مع التفاعل التنافسي بين أنظمة الذاكرة ، فقد تم الافتراض بأن الكحول قد يسهل الذاكرة المعتمِدة على DLS بشكل غير مباشر عبر إضعاف آليات الذاكرة المعرفية (78).
تجدر الإشارة إلى أنه بالإضافة إلى الكحول ، فقد ارتبط العديد من العقاقير بنقص الذاكرة المعرفية. التعرض للمورفين والهيروين والميثامفيتامين ، MDMA (النشوة) ، أو الكوكايين المزمن بالمثل تنتج ضعف الذاكرة المكانية المعتمدة على الحصين عبر مجموعة متنوعة من المهام (79-89). من المغري التكهن بأن ، كما هو مقترح بالنسبة للكحول ، فإن ضعف الذاكرة الإدراكي الناتج عن العقاقير المسببة للإدمان قد يعزز بشكل غير مباشر الذاكرة المعتمَدة على DLS ، وأن هذا قد يكون آلية تسمح للإدارة الذاتية للمخدرات أن تصبح معتادة في تعاطي المخدرات. ومن ناحية أخرى ، من الممكن أيضًا أن تحدث عجوزات التعلم المكاني الناتجة عن الأدوية المسببة للإدمان بشكل غير مباشر عن طريق تعزيز عمليات الذاكرة المعتمدة على DLS. بما يتفق مع هذه الفرضية ، فإن تنشيط نشاط CREB في DLS يضعف الذاكرة المكانية المعتمدة على الحصين (90) ، في حين أن تثبيط نشاط CREB في DLS يعكس ضعف الذاكرة المكانية الناتجة عن المورفين (91).
دور الاجهاد والقلق
وهناك اعتبار إضافي فيما يتعلق بنهج أنظمة الذاكرة المتعددة لإدمان المخدرات هو دور الإجهاد. تشير الأدلة المتقاربة إلى أن الاستثارة العاطفية القوية تسهل ذكرى الذاكرة المعتمدة على DLS في القوارض والبشر [للمراجعات ، راجع المرجع. (9-12)]. إدارة الأدوية المسببة للتشنج تعزز تعلم الاستجابة المعتمدة على DLS في الماء زائد المتاهة (92-97). هذا التحسن في ذاكرة العادة المعتمدة على DLS يلاحظ أيضًا بعد التعرض لحدوث ضغوط سلوكية غير مشروطة [على سبيل المثال ، تقييد مزمن ، صدمة ذيل ، رائحة مفترسة ، إلخ.)98-101)] والتعرض لمثيرات الخوف المرهقة [نغمة مقترنة بالصدمة سابقًا (102, 103)]. على الرغم من أن ثبت في الأصل في القوارض (92) ، وهذا تعزيز الذاكرة العادة الناجمة عن الإثارة العاطفية القوية كما تم أظهر على نطاق واسع في البشر (99, 104-110).
لا تزال الآليات التي تسمح بالضغط / القلق لتسهيل ذاكرة العادة غير معروفة إلى حد كبير. ومع ذلك ، تشير الأدلة إلى الدور التحوري الحرج لل BLA (93-95, 100). بالتوافق مع التفاعل التنافسي بين أنظمة الذاكرة ، تشير بعض الأدلة أيضًا إلى أن الإجهاد / القلق قد يؤدي إلى تعزيز ذاكرة العادة المعتمدة على DLS بشكل غير مباشر من خلال إضعاف وظيفة الحصين (94, 95).
قد يكون تعزيز ذاكرة العادة بعد الإجهاد أو القلق مفيدًا لفهم بعض العوامل البارزة التي تؤدي إلى تعاطي المخدرات. أي ، أحداث الحياة المجهدة أو الفترات المزمنة لفترات طويلة من التوتر / القلق ترتبط بزيادة التعرض للإدمان على المخدرات والانتكاس في البشر (111-117) ، وقد تم إجراء ملاحظات مشابهة في النماذج الحيوانية للإدارة الذاتية للمخدرات [للمراجعة ، انظر المرجع. (118)]. وقد اقترح الباحثون أن تتفق مع تأثير الإثارة العاطفية على أنظمة ذاكرة متعددة (10) ، قد يؤدي الإجهاد الحاد أو المزمن إلى تعزيز إدمان المخدرات والانتكاس في البشر عن طريق إشراك عمليات الذاكرة التي تعتمد على DLS (9, 49, 119). بالتوافق مع هذا الاقتراح ، يرتبط الإجهاد لدى الأفراد المعتمدين على الكوكايين بانخفاض النشاط المعتمد على مستوى الأكسجين في الدم (BOLD) في قرن آمون وزيادة النشاط في المخطط الظهري ، وترتبط هذه التغييرات في نشاط BOLD مع الرغبة الشديدة في تعاطي الكوكايين الناتج عن الإجهاد (120).
مصادر الادمان الناشئة
وبصرف النظر عن تعاطي المخدرات ، فقد تم استخدام فرضية أنظمة الذاكرة المتعددة في الآونة الأخيرة لفهم مصادر الإدمان الناشئة الأخرى. على سبيل المثال ، أدى الارتفاع في السمنة على مدى العقود القليلة الماضية إلى ارتفاع مماثل في الاهتمام التجريبي ، مع العديد من الباحثين رسم أوجه الشبه بين إدمان المخدرات والإفراط في تناول الطعام [للمراجعة ، انظر المرجع. (121-123)]. وقد أشارت بعض الأدلة الحديثة إلى أنه مثل إدمان المخدرات ، يمكن أن يعزى إدمان الغذاء جزئياً إلى زيادة إشراك ذاكرة العادة المعتمدة على DLS. في الفئران ، يسهّل استهلاك الطعام الشبيه بنهم الإزاحة الانتقال من السيطرة الإدراكية إلى السلوك المعتاد (124, 125). علاوة على ذلك ، يرتبط السلوك المعتاد في حيوانات الحبل مع زيادة النشاط DLS ويمكن الوقاية من خلال منع مستقبلات AMPA أو الدوبامين D1 في DLS (125). كما أن السمنة التي يسببها النظام الغذائي ارتبطت في الآونة الأخيرة باستخدام ذاكرة العادة في مهمة Y-maze (126).
اضطراب سلوكي ناشئ آخر يوازي بعض ميزات إدمان المخدرات هو لعبة فيديو أو لعبة إدمان ألعاب الفيديو [للمراجعة ، انظر المرجع. (127)]. مثل إدمان المخدرات ، فقد ارتبط اللعب المفرط في لعبة الفيديو على المدى الطويل بخفض مستقبلات الدوبامين D2 في المخطط الظهري (dorsal striatum)128). ويرتبط اللعب العاب الفيديو أيضا لزيادة تفعيل المخطط الظهري (129, 130) ، وتتوقع أحجام مخططة ظهرية أكبر مستويات أعلى من مهارة ألعاب الفيديو (131). من المرجح أن يستخدم الأشخاص الذين يمارسون ألعاب فيديو تعمل بشكل منتظم ذاكرة العادة الثانوية المرتبطة بالمخططات الظاهرية في متاهة افتراضية (132) ، وتؤدي ممارسة لعبة الفيديو التمهيدي سابقًا إلى الرد المعتاد على الاستجابة الموجهة نحو الهدف في مهمة صنع القرار على مرحلتين (133). وبالتالي ، فإن لعب ألعاب الفيديو ، كما هو مقترح لتعاطي المخدرات ، قد يحسن إدمان ألعاب الفيديو من خلال إشراك نظام الذاكرة المعتمد على DLS.
وأخيرًا ، قد يكون نهج أنظمة الذاكرة المتعددة مفيدًا أيضًا في فهم إدمان الماريجوانا. على الرغم من أن الماريجوانا قد تكون أقل عرضة للإساءة من غيرها من المواد غير المشروعة التي تعتبر معتادة في سياق أبحاث إدمان المخدرات (مثل الكوكايين ، والمورفين ، والهيروين ، وما إلى ذلك) ، إلا أن استخدام القنب الثقيل يمكن مع ذلك أن يشجع على الاعتماد على المخدرات وأعراض الانسحاب كما هو ملاحظ مع أدوية أخرى. إساءة (134-137). وقد اقترح مؤخرا أن إدمان الماريجوانا يمكن أن يعزى جزئيا إلى زيادة مشاركة الذاكرة المعتمد على DLS (138). في حين أن التعرض الحاد للقنب يضعف وظيفة الذاكرة المعتمدة على DLS (139, 140) ، يؤدي التعرض المتكرر للقنب إلى زيادة الاستجابة المعتادة على DLS في مهمة تعلم مفيدة (141). بالإضافة إلى ذلك ، يعرض مستخدمو القنب الثقيل عملية تنشيط أكبر للمخطط الظهري ، بالنسبة لغير المستخدمين ، عند إجراء نسخة من الماريجوانا لمهمة الارتباط الضمني (142) ، والمشاركين الذين لديهم تاريخ من استخدام الحشيش هم أكثر عرضة لاستخدام الذاكرة الاعتيادية الظهارية المعتمدة على المخطط في المتاهة الافتراضية (65).
بالنظر إلى التطبيق الناجح لنهج نظام الذاكرة لمصادر الإدمان الناشئة ، من المعقول الافتراض بأن أنظمة الذاكرة المتعددة قد تكون متورطة أيضًا في الأمراض السلوكية الأخرى المرتبطة بالإدمان ، مثل التسوق القهري وإدمان الإنترنت والإدمان على الجنس. في الواقع ، ما إذا كان نهج نظام الذاكرة قد يكون مفيدًا لفهم المقامرة المرضية قد حظي أيضًا ببعض الاهتمام (143, 144).
وفي الختام
لقد أثبتت عشرين عامًا من الأدلة التجريبية صحة1) نهج ذاكرة متعددة نهج لإدمان المخدرات. تشير الأدلة إلى أن الحصين يتوسط التحكم السياقي للإدارة الذاتية للمخدر ، تتوسط الـ DLS الاستجابة المعتادة لـ S-R من أجل تعزيز العقاقير ، وتتوسط اللوزة اللولبية عقارًا مشروطًا. بالإضافة إلى ذلك ، أدى البحث اللاحق إلى رؤى إضافية فيما يتعلق بنظريات أنظمة الذاكرة المتعددة لإدمان المخدرات بما في ذلك التحول إلى ذاكرة العادة ، والتنافس بين أنظمة الذاكرة ، ودور التوتر والقلق.
يجب أن يحاول البحث المستقبلي دمج مقاربة نظم الذاكرة مع نظريات الإدمان الأخرى ، مثل العمليات التحفيزية للخصم (145). سيكون من المفيد أيضا أن تدرج في أنظمة الذاكرة عرض ميزات إضافية للإدمان ، مثل الاعتماد على المخدرات ، والتسامح ، والانسحاب. على الرغم من أن المراجعة الحالية تركزت في الغالب على مناطق الدماغ التي كان البيض يعتبرها في الأصل (أي ، الحصين ، المخطط الظهري ، واللوزة) ، يجب ملاحظة أن مناطق الدماغ الإضافية المتعلقة بالتعلم والذاكرة كانت متورطة بشكل حاسم في إدمان المخدرات والانتكاس. ، بما في ذلك قشرة الفص الجبهي الإنسي والنقط المتكئة [للمراجعة ، راجع المرجع. (13)]. وأخيرًا ، على الرغم من أنه خارج نطاق المراجعة الحالية ، إلا أنه يجب الاعتراف بأن هناك أدلة مستفيضة تشير إلى أن التغيرات الخلوية والجزيئية في نظام دوبامين الدماغ المتوسط تساهم أيضًا في الإدمان (146).
على الرغم من أن ذكريات العادة قد يكون من الصعب السيطرة عليها بشكل خاص ، إلا أن بعض الأدلة تشير إلى أن الذاكرة التي تعتمد على DLS ، بمجرد الحصول عليها ، يمكن في بعض الحالات قمعها (147) أو حتى عكسه (148, 149). وبالتالي ، فمن المحتمل أن تكون التلاعبات الدوائية والإجراءات السلوكية التي تؤدي إلى عكس أو إخماد ذاكرة العادة في نماذج التعلم الحيوانية من الممكن تكييفها لعلاج الإدمان على المخدرات والانتكاس في البشر.
الكاتب الاشتراكات
ساهم كل من JG و MP في أفكار وكتابة للمراجعة المصغّرة الحالية.
بيان تضارب المصالح
يعلن المؤلفون أن البحث أجري في غياب أي علاقات تجارية أو مالية يمكن تفسيرها على أنها تضارب مصالح محتمل.
مراجع حسابات
1. NM الأبيض. العقاقير التي تسبب الادمان كمعززات: إجراءات جزئية متعددة على أنظمة الذاكرة. الإدمان (1996) 91(7):921–50. doi: 10.1111/j.1360-0443.1996.tb03586.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
2. NM الأبيض ، ماكدونالد RJ. أنظمة ذاكرة متوازية متعددة في دماغ الفئران. Neurobiol تعلم Mem (2002) 77(2):125–84. doi:10.1006/nlme.2001.4008
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
3. سكوير LR. نظم الذاكرة في الدماغ: تاريخ موجز ومنظور الحالي. Neurobiol تعلم Mem (2004) 82(3):171–7. doi:10.1016/j.nlm.2004.06.005
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
4. NM الأبيض ، باكارد MG ، ماكدونالد RJ. انفصال أنظمة الذاكرة: القصة تتكشف. Behav Neurosci (2013) 127(6):813–34. doi:10.1037/a0034859
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
5. Packard MG، Hirsh R، White NM. التأثيرات التفاضلية للإصابات النواة والفوقية على اثنين من مهام متاهة شعاعية: دليل على أنظمة ذاكرة متعددة. J Neurosci (1989) 9(5): 1465-72.
6. ماكدونالد RJ ، الأبيض NM. تفكك ثلاثي لأنظمة الذاكرة: الحصين ، اللوزة ، والمخطط الظهري. Behav Neurosci (1993) 107(1):3–22. doi:10.1037/0735-7044.107.1.3
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
7. مارين S. علم الأعصاب من تكييف خوف بافلوفان. انو ريف نيوروسكي (2001) 24(1):897–931. doi:10.1146/annurev.neuro.24.1.897
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
8. McGaugh JL. ينقسم اللوزة لتوحيد الذكريات للتجارب المثيرة للعاطفة. انو ريف نيوروسكي (2004) 27: 1-28. دوى: 10.1146 / annurev.neuro.27.070203.144157
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
9. باكارد MG. القلق والإدراك والعادة: منظور أنظمة ذاكرة متعددة. الدماغ الدقة (2009) 1293: 121-8. دوى: 10.1016 / j.brainres.2009.03.029
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
10. Packard MG، Goodman J. الإثارة العاطفية وأنظمة الذاكرة المتعددة في الدماغ في الثدييات. الجبهة Behav Neurosci (2012) 6: 14. دوى: 10.3389 / fnbeh.2012.00014
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
11. Packard MG، Goodman J. العوامل التي تؤثر على الاستخدام النسبي لأنظمة الذاكرة المتعددة. الحصين (2013) 23(11):1044–52. doi:10.1002/hipo.22178
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
12. Schwabe L. الإجهاد ومشاركة أنظمة الذاكرة المتعددة: دمج الدراسات الحيوانية والإنسانية. الحصين (2013) 23(11):1035–43. doi:10.1002/hipo.22175
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
13. Everitt BJ، Robbins TW. الأنظمة العصبية من التعزيز لإدمان المخدرات: من الإجراءات إلى العادات للإكراه. نات نيوروسكي (2005) 8(11):1481–9. doi:10.1038/nn1579
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
14. Fuchs RA، Evans KA، Ledford CC، Parker MP، Case JM، Mehta RH، et al. دور قشرة الفص الجبهي الظهري الظهري ، اللوزة الوحشية basolateral ، و hippocampus الظهرية في السياق عودة الكوكايين في الفئران. Neuropsychopharmacology (2005) 30(2):296–309. doi:10.1038/sj.npp.1300579
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
15. Fuchs RA، Eaddy JL، Su ZI، Bell GH. تفاعل ال amygdala basolateral مع hippocampus الظهرية وقشرة الفص الجبهي الظهارة dorsomedial ينظم السياق المستحث بالسياسة من إعادة البحث عن الكوكايين في الفئران. يورو J Neurosci (2007) 26(2):487–98. doi:10.1111/j.1460-9568.2007.05674.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
16. Kramar CP، Barbano MF، Medina JH. مطلوبة مستقبلات الدوبامين D1 / D5 في قرن آمون الظهرية لاكتساب والتعبير عن ذاكرة واحدة مرتبطة الكوكايين المرتبطة. Neurobiol تعلم Mem (2014) 116: 172-80. دوى: 10.1016 / j.nlm.2014.10.004
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
17. Zapata A، Minney VL، Shippenberg TS. التحول من الهدف الموجه إلى تعاطي الكوكايين المعتاد بعد تجربة طويلة في الفئران. J Neurosci (2010) 30(46):15457–63. doi:10.1523/JNEUROSCI.4072-10.2010
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
18. Corbit LH، Nie H، Janak PH. البحث عن الكحول المعتاد: دورة زمنية ومساهمة المناطق دون الإقليمية للمخطط الظهري. بيول الطب النفسي (2012) 72(5):389–95. doi:10.1016/j.biopsych.2012.02.024
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
19. Whitelaw RB، Markou A، Robbins TW، Everitt BJ. إن آفات اللوزة الملقحة للأعصاب في اللوزة القاعدية الجانبية تضعف من اكتساب سلوك يسعى للكوكايين في إطار جدول زمني من الدرجة الثانية من التعزيز. علم الادوية النفسية (1996) 127(1–2):213–24. doi:10.1007/BF02805996
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
20. Alderson HL، Robbins TW، Everitt BJ. آثار آفات excitotoxic من اللوزة الوحشية basolateral على اكتساب السلوك تسعى الهيروين في الفئران. علم الادوية النفسية (2000) 153(1):111–9. doi:10.1007/s002130000527
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
21. Gabriele A، راجع RE. تعطيل عكوس اللوزة الجانبية الوحشية ، ولكن ليس البويدات الوحشية الظهارية الوحشية ، يخفف من توطيد التوعية الكوكايين-جديلة في نموذج إعادة البحث عن المخدرات. يورو J Neurosci (2010) 32(6):1024–9. doi:10.1111/j.1460-9568.2010.07394.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
22. Sciascia JM، Reese RM، Janak PH، Chaudhri N. يتم تنشيط البحث عن الكحول من قبل العظة Pavlovian المنفصلة عن طريق سياقات الكحول والتوسط عن طريق الإشارة الغلوتامات في اللوزة الوحشية basolateral. Neuropsychopharmacology (2015) 40: 2801-12. دوى: 10.1038 / npp.2015.130
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
23. Packard MG، Teather LA. تعديل اللوغز من أنظمة الذاكرة المتعددة: الحصين و caudate-putamen. Neurobiol تعلم Mem (1998) 69(2):163–203. doi:10.1006/nlme.1997.3815
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
24. Dickinson A، Wood N، Smith JW. البحث عن طريق الكحول من قبل الفئران: العمل أو العادة؟ QJ Exp Psychol B (2002) 55(4):331–48. doi:10.1080/0272499024400016
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
25. Udo T، Ugalde F، DiPietro N، Eichenbaum HB، Kantak KM. آثار استمرار الإدارة الذاتية للكوكايين على التعلم المعتمد على المخططات المعتمدة على اللوزة المخية في الفئران. علم الادوية النفسية (2004) 174(2):237–45. doi:10.1007/s00213-003-1734-1
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
26. Wood SC، Fay J، Sage JR، Anagnostaras SG. الكوكايين و بافلوفيان تكييف الخوف: تحليل تأثير الجرعة. Behav Brain Res (2007) 176(2):244–50. doi:10.1016/j.bbr.2006.10.008
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
27. Wood SC، Anagnostaras SG. الذاكرة ومضادات نفسية: تعديل تكييف بافلوفان بواسطة الأمفيتامين في فئران C57BL / 6. علم الادوية النفسية (2009) 202(1–3):197–206. doi:10.1007/s00213-008-1185-9
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
28. Iñiguez SD، Charntikov S، Baella SA، Herbert MS، Bolaños-Guzmán CA، Crawford CA. يسهّل التعرض للكوكايين بعد التدريب تدريب الذاكرة المكانية في فئران C57BL / 6. الحصين (2012) 22(4):802–13. doi:10.1002/hipo.20941
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
29. DePoy L، Daut R، Brigman JL، MacPherson K، Crowley N، Gunduz-Cinar O، et al. ينتج الكحول المزمن عمليات تعديل عصبي لتعليم الرأسية المخطط. بروك Natl أكاد العلوم الولايات المتحدة الأمريكية (2013) 110(36):14783–8. doi:10.1073/pnas.1308198110
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
30. Leri F، Nahas E، Henderson K، Limebeer CL، Parker LA، White NM. آثار ما بعد التدريب على الهيروين و d-amphetamine على توطيد التعلُّم من أجل البقاء ، والتكيف مع الخوف. ي Psychopharmacol (2013) 27(3):292–301. doi:10.1177/0269881112472566
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
31. Schmitzer-Torbert N، Apostolidis S، Amoa R، O'Rear C، Kaster M، Stowers J، et al. إدارة ما بعد التدريب الكوكايين تسهل عملية تعلم العادات وتحتاج إلى القشرة اللعابية وطبقية الظهرية الجانبية. Neurobiol تعلم Mem (2015) 118: 105-12. دوى: 10.1016 / j.nlm.2014.11.007
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
32. نولتون ب. العقد القاعدية: تشكيل العادة. In: Jaeger D، Jung R، editors. موسوعة علم الأعصاب الحاسوبية. نيويورك: سبرينغر (2014). ص. 1-17.
33. Tolman EC، Ritchie BF، Kalish D. Studies in spatial learning. IV. نقل التعلم مكان لمسارات بدء أخرى. J Exp Psychol (1947) 37(1):39–47. doi:10.1037/h0062061
34. Ritchie BF، Aeschliman B، Pierce P. Studies in spatial learning. VIII. مكان الأداء واقتناء ترتيبات المكان. J Comp Physiol Psychol (1950) 43(2):73–85. doi:10.1037/h0055224
35. هيكس LH. آثار overtraining على اكتساب وعكس تعلم المكان والاستجابة. النائب Psychol (1964) 15(2):459–62. doi:10.2466/pr0.1964.15.2.459
36. Packard MG، McGaugh JL. إن تثبيط نواة الحصين أو النواة المذنبة مع ليدوكائين يؤثر بشكل تفاضلي على التعبير عن المكان وتعلم الاستجابة. Neurobiol تعلم Mem (1996) 65(1):65–72. doi:10.1006/nlme.1996.0007
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
37. يين HH ، نولتون BJ. مساهمات المناطق دون الإقليمية في الموضع لوضعها والاستجابة لها. تعلم ميم (2004) 11(4):459–63. doi:10.1101/lm.81004
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
38. Adams CD، Dickinson A. Instrumental response after following reinforcer defaluation. QJ Exp Psychol (1981) 33B: 109-12. دوى: 10.1080 / 14640748108400816
39. Adams CD، Dickinson A. Actions and habits: variations in associative representations during instrumental learning. In: Spear NE، Miller RR، editors. معالجة المعلومات في الحيوانات: آليات الذاكرة. Hillsdale، NJ: Erlbaum (1981). ص. 143-65.
40. آدمز CD. الاختلافات في حساسية الرد الفعال على تخفيض قيمة المعزز. QJ Exp Psychol (1982) 34B: 77-98. دوى: 10.1080 / 14640748208400878
41. ديكنسون أ ، نيكولاس دي جي. تعلم حافز غير ذي صلة أثناء التكييف الفعال: دور معزز محرك الأقراص وعوامل تعزيز الاستجابة. QJ Exp Psychol (1983) 35B: 249-63. دوى: 10.1080 / 14640748308400909
42. Dickinson A، Nicholas DJ، Adams CD. آثار الطوارئ مفيدة على القابلية لخفض قيمة التخفيض. QJ Exp Psychol (1983) 35B: 35-51. دوى: 10.1080 / 14640748308400912
43. Corbit LH، Balleine BW. دور hippocampus في تكييف فعال. J Neurosci (2000) 20(11): 4233-9.
44. Yin HH، Ostlund SB، Knowlton BJ، Balleine BW. دور المخطط الظهاري الظهري في تكييف فعال. يورو J Neurosci (2005) 22:513–23. doi:10.1111/j.1460-9568.2005.04218.x
45. Yin HH، Knowlton BJ، Balleine BW. الآفات من striatum ظهرية جانبية الحفاظ على النتائج المتوقعة ولكن تعطيل تشكيل العادة في التعلم مفيدة. يورو J Neurosci (2004) 19:181–9. doi:10.1111/j.1460-9568.2004.03095.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
46. Quinn JJ، Pittenger C، Lee AS، Pierson JL، Taylor JR. لا تعتمد العادات المعتمِدة على الأحجار على الزيادات والنقصان في القيمة المعززة لدى الفئران. يورو J Neurosci (2013) 37: 1012-21. دوى: 10.1111 / ejn.12106
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
47. يين سمو. من الإجراءات إلى العادات: عمليات تجديد عصبي تؤدي إلى الاعتماد. الكحول الصحة الصحة (2008) 31(4): 340-4.
48. Belin D، Jonkman S، Dickinson A، Robbins TW، Everitt BJ. عمليات التعلم الموازية والتفاعلية داخل العقد القاعدية: الصلة لفهم الإدمان. Behav Brain Res (2009) 199(1):89–102. doi:10.1016/j.bbr.2008.09.027
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
49. Schwabe L، Dickinson A، Wolf OT. الإجهاد والعادات والإدمان على المخدرات: منظور نفسي عصبي ثقافي. إكسب كلينيك Psychopharmacol (2011) 19(1):53–63. doi:10.1037/a0022212
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
50. Hogarth L ، Balleine BW ، Corbit LH ، Killcross S. آليات التعلم التعاقبي التي تدعم الانتقال من تعاطي المخدرات الترويحي إلى الإدمان. Ann NY Acad Sci (2013) 1282(1):12–24. doi:10.1111/j.1749-6632.2012.06768.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
51. Murray JE، Belin D، Everitt BJ. تفكك مزدوج للسيطرة الظهارية الظهارية والظهرية على اكتساب وأداء البحث عن الكوكايين. Neuropsychopharmacology (2012) 37(11):2456–66. doi:10.1038/npp.2012.104
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
52. Clemens KJ، Castino MR، Cornish JL، Goodchild AK، Holmes NM. الركائز السلوكية والعصبية من تشكيل العادات في الجرذان عن طريق الوريد النيكوتين بالإدارة الذاتية. Neuropsychopharmacology (2014) 39: 2584-93. دوى: 10.1038 / npp.2014.111
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
53. Corbit LH، Nie H، Janak PH. يعتمد الاستجابة المعتادة للكحول على كل من مستقبلات AMPA و D2 في المخطط الجانبي الظهاري. الجبهة Behav Neurosci (2014) 8: 301. دوى: 10.3389 / fnbeh.2014.00301
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
54. Schoenbaum G، Setlow B. الكوكايين يجعل الأعمال غير حساسة للنتائج ولكن ليس الانقراض: الآثار المترتبة على وظيفة amygdalar تغيير المداري. Cereb اللحاء (2005) 15(8):1162–9. doi:10.1093/cercor/bhh216
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
55. Nelson A، Killcross S. Amphetamine التعرض يعزز تشكيل العادة. J Neurosci (2006) 26(14):3805–12. doi:10.1523/JNEUROSCI.4305-05.2006
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
56. Nordquist RE، Voorn P، De Mooij-van Malsen JG، Joosten RNJMA، Pennartz CMA، Vanderschuren LJMJ. القيمة المعززة المعززة والتشكيل المتسارع للعادة بعد العلاج المتكرر للأمفيتامين. Eur Neuropsychopharmacol (2007) 17(8):532–40. doi:10.1016/j.euroneuro.2006.12.005
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
57. LeBlanc KH، Maidment NT، Ostlund SB. ويسهِّل التعرض المتكرر للكوكايين التعبير عن دوافع التحفيز والحث على التحكم المعتاد في الجرذان. بلوس واحد (2013) 8: e61355. دوى: 10.1371 / journal.pone.0061355
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
58. Nelson AJ ، Killcross S. تم عكس تشكيل العجلة المتسارع بعد التعرض للأمفيتامين بواسطة D1 ، ولكن تم تعزيزه بواسطة D2 ، مضادات المستقبلات. جبهة عصبيه (2013) 7: 76. دوى: 10.3389 / fnins.2013.00076
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
59. Corbit LH، Chieng BC، Balleine BW. آثار التعرض المتكرر للكوكايين على تعلم العادات وانعكاسها بواسطة N-acetylcysteine. Neuropsychopharmacology (2014) 39(8):1893–901. doi:10.1038/npp.2014.37
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
60. مايلز إف جي ، إيفريت بي جي ، ديكينسون أ. الكوكايين عن طريق الفم تسعى من الفئران: العمل أو العادة؟ Behav Neurosci (2003) 117(5):927–38. doi:10.1037/0735-7044.117.5.927
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
61. Mangieri RA، Cofresí RU، Gonzales RA. الإيثانول الذي تبحث عنه الجرذان لفيف إيفانز ليس دائما سلوك موجه نحو الهدف. بلوس واحد (2012) 7: e42886. دوى: 10.1371 / journal.pone.0042886
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
62. Sjoerds Z، De Wit S، Van Den Brink W، Robbins TW، Beekman ATF، Penninx BWJH، et al. أدلة السلوك والتصوير العصبي لاعتماد أكثر من اللازم على تعلم العادة في المرضى الذين يعتمدون على الكحول. ترجمة علم النفس (2013) 3(12): e337. دوى: 10.1038 / tp.2013.107
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
63. Packard MG، McGaugh JL. يحسن Quinpirole و d-amphetamine إدارة posttraining الذاكرة على تمييز مكاني و cued في متاهة مائية. علم النفس (1994) 22(1): 54-60.
64. Matthews DB، Ilgen M، White AM، Best PJ. إدارة الإيثانول الحاد تعوق الأداء المكاني بينما تسهل الأداء غير الرياضي في الجرذان. Neurobiol تعلم Mem (1999) 72(3):169–79. doi:10.1006/nlme.1998.3900
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
65. ويرتبط Bohbot VD ، Balso D ، كونراد K ، كونيشي K ، ليتون م. Caudate الاستراتيجيات الملاحية تعتمد النواة مع زيادة استخدام العقاقير الادمانية. الحصين (2013) 23(11):973–84. doi:10.1002/hipo.22187
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
66. Poldrack RA، Packard MG. التنافس بين أنظمة الذاكرة المتعددة: الأدلة المتقاربة من دراسات الدماغ البشري والحيواني. Neuropsychologia (2003) 41(3):245–51. doi:10.1016/S0028-3932(02)00157-4
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
67. ماثيوز ديسيبل ، وأفضل PJ. Fimbria / fornix الآفات تسهيل تعلم مهمة استجابة غير طبية. Psychon Bull Rev (1995) 2(1):113–6. doi:10.3758/BF03214415
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
68. Schroeder JP، Wingard JC، Packard MG. يكشف التثبيط العكسي بعد التدريب للحصيات عن التداخل بين أنظمة الذاكرة. الحصين (2002) 12(2):280–4. doi:10.1002/hipo.10024
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
69. ماثيوز دي بي ، سيمون PE ، أفضل PJ. الإيثانول الحاد يضعف الذاكرة المكانية ولكن ليس الذاكرة المحفزة / استجابة في الفئران. كحول الكحول إكسب Res (1995) 19(4):902–9. doi:10.1111/j.1530-0277.1995.tb00965.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
70. White AM، Elek TM، Beltz TL، Best PJ. الأداء المكاني أكثر حساسية للإيثانول من الأداء غير الرياضي بغض النظر عن القربان. كحول الكحول إكسب Res (1998) 22(9):2102–7. doi:10.1111/j.1530-0277.1998.tb05922.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
71. Matthews DB، Morrow AL، Tokunaga S، McDaniel JR. إدارة الإيثانول الحاد وإدارة allopregnanolone الحاد تضعف الذاكرة المكانية في مهمة موريس للمياه. كحول الكحول إكسب Res (2002) 26(11):1747–51. doi:10.1111/j.1530-0277.2002.tb02479.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
72. بيري آر بي ، ماثيوز ديسيبل. إدارة الإيثانول الحاد يضعف بشكل انتقائي الذاكرة المكانية في الفئران C57BL / 6J. كحول (2004) 32(1):9–18. doi:10.1016/j.alcohol.2003.09.005
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
73. Silvers JM، Tokunaga S، Berry RB، White AM، Matthews DB. ضعف في التعلم المكاني والذاكرة: الإيثانول ، اللوبيريجنولونات ، و hippocampus. دماغ ريف القس (2003) 43(3):275–84. doi:10.1016/j.brainresrev.2003.09.002
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
74. Badanich KA، Becker HC، Woodward JJ. تأثير التعرض المزمنة للإيثانول المتقطع على السلوكيات التي تعتمد على القشرة الجبهية الأمامية والمتوسطة في الفئران. Behav Neurosci (2011) 125(6):879–91. doi:10.1037/a0025922
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
75. Coleman LG Jr، He J، Lee J، Styner M، Crews FT. المراهق الشراهة يغير الكبار ناقلات التعبير الجيني في الدماغ ، والسلوك ، وحجم الدماغ الإقليمي ، والكيمياء العصبية في الفئران. كحول الكحول إكسب Res (2011) 35(4):671–88. doi:10.1111/j.1530-0277.2010.01385.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
76. Kuzmin A ، Liljequist S ، Meis J ، Chefer V ، Shippenberg T ، Bakalkin G. نوبات معتدلة من الإيثانول جرعة معتدلة يضعف وظيفة المعرفية في الفئران Wistar. المدمن Biol (2012) 17(1):132–40. doi:10.1111/j.1369-1600.2010.00224.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
77. Coleman LG، Liu W، Oguz I، Styner M، Crews FT. يزيل علاج الإيثانول للمراهقين الأحجام الدماغية الإقليمية للبالغين ، وبروتين المصفوفة خارج الخلية والمرونة السلوكية. Pharmacol Biochem Behav (2014) 116: 142-51. دوى: 10.1016 / j.pbb.2013.11.021
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
78. Matthews DB، Silvers JR. استخدام إدارة الإيثانول الحاد كأداة للتحقيق في أنظمة ذاكرة متعددة. Neurobiol تعلم Mem (2004) 82(3):299–308. doi:10.1016/j.nlm.2004.06.007
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
79. Broiding HW، Morford LL، Inman-Wood SL، Fukumura M، Vorhees CV. يعتمد 3 و 4-methylenedioxymethamphetamine (الإكستاسي) على التعلم والإعاقات في الذاكرة على عمر التعرض خلال التطور المبكر. J Neurosci (2001) 21(9): 3228-35.
80. Williams MT، Morford LL، Wood SL، Wallace TL، Fukumura M، Broening HW، et al. يقوم العلاج التطوري d-methamphetamine بتحفيز انتقائي للملاحة المكانية في الذاكرة المرجعية في متاهة موريس المائية مع الحفاظ على الذاكرة العاملة. المشبك (2003) 48(3):138–48. doi:10.1002/syn.10159
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
81. Vorhees CV، Reed TM، Skelton MR، Williams MT. التعرض ل 3 و 4-methylenedioxymethamphetamine (MDMA) في أيام ما بعد الولادة 11-20 يقوم بتحويل الإشارة إلى عدم وجود عجز في الذاكرة في متاهة موريس المائية في الجرذان: آثار التعلم السابق. Int J Dev Neurosci (2004) 22(5):247–59. doi:10.1016/j.ijdevneu.2004.06.003
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
82. Cohen MA، Skelton MR، Schafer TL، Gudelsky GA، Vorhees CV، Williams MT. التعلم والذاكرة بعد التعرض الوليدي ل 3 و 4-methylenedioxymethamphetamine (الإكستاسي) في الفئران: التفاعل مع التعرض في مرحلة البلوغ. المشبك (2005) 57(3):148–59. doi:10.1002/syn.20166
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
83. Skelton MR، Williams MT، Vorhees CV. العلاج مع MDMA من P11-20 يعطل التعلم المكاني وتعلم تكامل المسار في الفئران المراهقين ولكن فقط التعلم المكاني في الفئران الأكبر سنا. علم الادوية النفسية (2006) 189(3):307–18. doi:10.1007/s00213-006-0563-4
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
84. Ma MX، Chen YM، He J، Zeng T، Wang JH. آثار المورفين وانسحابها على الذاكرة المتاهة المكانية Y-maze في الفئران. علم الأعصاب (2007) 147(4):1059–65. doi:10.1016/j.neuroscience.2007.05.020
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
85. Belcher AM، Feinstein EM، O'Dell SJ، Marshall JF. الميثامفيتامين يؤثر على ذاكرة الاعتراف: مقارنة نظم الجرعات المتصاعدة واليوم الواحد. Neuropsychopharmacology (2008) 33(6):1453–63. doi:10.1038/sj.npp.1301510
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
86. Tramullas M، Martínez-Cué C، Hurlé MA. إن الإدارة المزمنة للهيروين إلى الفئران تنتج تنظيمًا للبروتينات المرتبطة بالبرمجة الدماغية وتضعف التعلم المكاني والذاكرة. الجهاز العصبي (2008) 54(4):640–52. doi:10.1016/j.neuropharm.2007.11.018
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
87. North A، Swant J، Salvatore MF، Gamble-George J، Prins P، Butler B، et al. يؤدي التعرض للميثامفيتامين المزمن إلى حدوث عجز في الذاكرة متأخّر يدوم طويلاً. المشبك (2013) 67(5):245–57. doi:10.1002/syn.21635
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
88. Fole A، Martin M، Morales L، Del Olmo N. Effects of chronic cocaine treatment during adolescence in Lewis and Fischer-344 rats: novel location recognition recognition and changes in synaptic plasticity in adulthood. Neurobiol تعلم Mem (2015) 123: 179-86. دوى: 10.1016 / j.nlm.2015.06.001
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
89. Zhou M، Luo P، Lu Y، Li CJ، Wang DS، Lu Q، et al. يؤدي اختلال التوازن بين تعبير HCN1 و HCN2 في منطقة CA1 الحصينية إلى إعاقة التعلم المكاني والذاكرة في الجرذان التي تتعرض للمورفين المزمنة. بروغ Neuropsychopharmacol بيول الطب النفسي (2015) 56: 207-14. دوى: 10.1016 / j.pnpbp.2014.09.010
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
90. Kathirvelu B، Colombo PJ. آثار تعبير CREB بوساطة الفيروسات في المخطط الظهاري الجانبي: تعزيز الذاكرة والأدلة على التفاعلات التنافسية والتعاونية مع قرن آمون. الحصين (2013) 23(11):1066–74. doi:10.1002/hipo.22188
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
91. Baudonnat M، Guillou JL، Husson M، Vandesquille M، Corio M، Decorte L، et al. تعطيل تأثير المكافأة الناتجة عن الأدوية على التعلم المكاني ولكن غير الموجه: تأثير بروتين كيناز بروتيني A / cAMP مسار بروتين ربط عنصر الارتباط. J Neurosci (2011) 31:16517–28. doi:10.1523/JNEUROSCI.1787-11.2011
92. Packard MG، Wingard JC. اللوزة وتعديل "العاطفي" من الاستخدام النسبي لأنظمة ذاكرة متعددة. Neurobiol تعلم Mem (2004) 82(3):243–52. doi:10.1016/j.nlm.2004.06.008
93. Elliott AE، Packard MG. intra-amygdala anxiogenic infusion قبل التحفيز استرجاع الفئران نحو استخدام الذاكرة العادة. Neurobiol تعلم Mem (2008) 90(4):616–23. doi:10.1016/j.nlm.2008.06.012
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
94. Wingard JC، Packard MG. اللوزة وتشكيل عاطفي للمنافسة بين الذاكرة المعرفية والعادة. Behav Brain Res (2008) 193(1):126–31. doi:10.1016/j.bbr.2008.05.002
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
95. Packard MG ، غابرييل A. الحقن المخدرات anxiogenic الطرفية يؤثر بشكل تفاضلي على الذاكرة المعرفية والعادة: دور اللوزة الوحشية basolateral. علم الأعصاب (2009) 164(2):457–62. doi:10.1016/j.neuroscience.2009.07.054
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
96. Leong KC، Goodman J، Packard MG. Buspirone يمنع التأثير المعزز للأدوية المضادة للتشنج RS 79948-197 على دمج ذاكرة العادة. Behav Brain Res (2012) 234(2):299–302. doi:10.1016/j.bbr.2012.07.009
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
97. Goodman J، Leong KC، Packard MG. تعزيز جلايكورتيكود للذاكرة الظهرية تعتمد على المخطط الافتراضي يتطلب نشاط noradrenergic المتزامنة. علم الأعصاب (2015) 311: 1-8. دوى: 10.1016 / j.neuroscience.2015.10.014
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
98. كيم جيه جي ، لي هج ، هان جي إس ، باكارد MG. اللوزة أمر بالغ الأهمية لتشكيل الإجهاد المستحث من تقوية قرن آمون على المدى الطويل والتعلم. J Neurosci (2001) 21(14): 5222-8.
99. Schwabe L، Dalm S، Schächinger H، Oitzl MS. ينظم الإجهاد المزمن استخدام استراتيجيات التعلم المكانية والاستجابة التحفيزية في الفئران والإنسان. Neurobiol تعلم Mem (2008) 90(3):495–503. doi:10.1016/j.nlm.2008.07.015
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
100. Leong KC، Packard MG. يؤثر التعرض للرائحة المفترسة على الاستخدام النسبي لأنظمة الذاكرة المتعددة: دور اللوزة الجانبية الأساسية. Neurobiol تعلم Mem (2014) 109: 56-61. دوى: 10.1016 / j.nlm.2013.11.015
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
101. Taylor SB، Anglin JM، Paode PR، Riggert AG، Olive MF، Conrad CD. قد يساعد الإجهاد المزمن في تعيين الدوائر العصبية المتعلقة بالإدمان والإدمان من خلال إعادة هيكلة الخلايا العصبية. علم الأعصاب (2014) 280: 231-42. دوى: 10.1016 / j.neuroscience.2014.09.029
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
102. Leong KC، Goodman J، Packard MG. يعزز إعادة التعرّض لمرحلة ما بعد التدريب للمخاوف المكيّفة من تعزيز الذاكرة وتحيّز الفئران نحو استخدام التعلم بالاستجابة المعزّزة للطبقات الجانبية الظاهرة. Behav Brain Res (2015) 291: 195-200. دوى: 10.1016 / j.bbr.2015.05.022
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
103. Goode TE، Leong KC، Goodman J، Maren S، Packard MG. يتم بوساطة تحسين الذاكرة التي تعتمد على المخطط باستخدام خوف مشروط بواسطة مستقبلات بيتا الأدرينالية في اللوزة الدماغية الجانبية. علم الأعصاب من الإجهاد (في الصحافة). دوى: 10.1016 / j.ynstr.2016.02.004
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
104. Schwabe L، Oitzl MS، Philippsen C، Richter S، Bohringer A، Wippich W، et al. ينظم الإجهاد استخدام استراتيجيات التعلم المكانية مقابل التحفيز والاستجابة عند البشر. تعلم ميم (2007) 14(1–2):109–16. doi:10.1101/lm.435807
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
105. Schwabe L، Schächinger H، de Kloet ER، Oitzl MS. يعمل الكورتيزون كمحول بين أنظمة الذاكرة. J Cogn Neurosci (2010) 22(7):1362–72. doi:10.1162/jocn.2009.21278
106. Schwabe L، Tegenthoff M، Höffken O، Wolf OT. النشاط الجلوكوكورتيكويدي والنورادريني المتزامن يغير سلوك الأدوات من الهدف الموجه إلى السيطرة المعتادة. J Neurosci (2010) 30(24):8190–6. doi:10.1523/JNEUROSCI.0734-10.2010
107. Schwabe L، Tegenthoff M، Höffken O، Wolf OT. يمنع حاجز مستقبلات مينيرالوكورتيكويد التشكيل الناتج عن الإجهاد لأنظمة الذاكرة المتعددة في الدماغ البشري. بيول الطب النفسي (2013) 74(11):801–8. doi:10.1016/j.biopsych.2013.06.001
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
108. Schwabe L، Wolf OT. الإجهاد يطالب سلوك العادة في البشر. J Neurosci (2009) 29(22):7191–8. doi:10.1523/JNEUROSCI.0979-09.2009
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
109. Schwabe L، Wolf OT. إجهاد الضغط البارد الذي تم تقييمه اجتماعياً بعد أن يساعد التعلم الفعال العادات على العمل الموجه نحو الهدف. Psychoneuroendocrinology (2010) 35(7):977–86. doi:10.1016/j.psyneuen.2009.12.010
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
110. Guenzel FM، Wolf OT، Schwabe L. Glucocorticids تعزز تكوين الذاكرة المحفزة الاستجابة في البشر. Psychoneuroendocrinology (2014) 45: 21-30. دوى: 10.1016 / j.psyneuen.2014.02.015
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
111. هيغنز RL ، مارلات GA. الخوف من التقييم الشخصي كمحدد لاستهلاك الكحول لدى من يشربون الاجتماعيين الذكور. J Abnorm Psychol (1975) 84(6):644–51. doi:10.1037/0021-843X.84.6.644
112. Marlatt GA، Gordon JR. محددات الانتكاس: الآثار المترتبة على الحفاظ على تغيير السلوك. In: Davidson PO، Davidson SM، editors. الطب السلوكي: تغيير نمط الحياة. نيويورك: برون / مازل (1980). ص. 410-52.
113. نيوكومب إم دي ، بنتلير PM. تأثير تعاطي المخدرات للمراهقين والدعم الاجتماعي لمشاكل الشباب: دراسة طولية. J Abnorm Psychol (1988) 97:64–75. doi:10.1037/0021-843X.97.1.64
114. والاس ق. المحددات النفسية والبيئية للانتكاس في مدخني الكوكايين الكراك. J علاج سوء المعاملة الثانوية (1989) 6(2):95–106. doi:10.1016/0740-5472(89)90047-0
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
115. Kaplan HB، Johnson RJ. العلاقات بين الظروف المحيطة بالاستخدام الأولي للمخدرات غير المشروعة وتصعيد تعاطي المخدرات: تخفيف الآثار المترتبة على الجندر وتجارب المراهقين المبكرة. In: Glantz M، Pickens R، editors. الضعف تجاه تعاطي المخدرات. واشنطن العاصمة: الجمعية الأمريكية لعلم النفس (1992). ص. 200-358.
116. Harrison PA، Fulkerson JA، Beebe TJ. استخدام متعدد المواد بين ضحايا الاعتداء البدني والجنسي للمراهقين. إساءة معاملة الطفل (1997) 21:529–39. doi:10.1016/S0145-2134(97)00013-6
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
117. Chilcoat HD، Breslau N. Posttraumumatic disorder and drug disorders: testing tests pathways. قوس الجنرال الطب النفسي (1998) 55(10):913–7. doi:10.1001/archpsyc.55.10.913
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
118. Piazza PV، Le Moal M. The Role of stress in drug self-administration. اتجاهات Pharmacol العلوم (1998) 19(2):67–74. doi:10.1016/S0165-6147(97)01115-2
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
119. Goodman J، Leong KC، Packard MG. التعديل العاطفي لأنظمة الذاكرة المتعددة: الآثار المترتبة على البيولوجيا العصبية لاضطراب ما بعد الصدمة. القس Neurosci (2012) 23(5–6):627–43. doi:10.1515/revneuro-2012-0049
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
120. Sinha R، Lacadie C، Skudlarski P، Fulbright RK، Rounsaville BJ، Kosten TR، et al. النشاط العصبي المرتبط بإجهاد الكوكايين الناجم عن الإجهاد: دراسة التصوير بالرنين المغناطيسي الوظيفي. علم الادوية النفسية (2005) 183(2):171–80. doi:10.1007/s00213-005-0147-8
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
121. Avena NM، Rada P، Hoebel BG. أدلة لإدمان السكر: التأثيرات السلوكية والعصبية الكيميائية الناجمة عن تناول السكر المفرط والمتقطع. Neurosci Biobehav Rev (2008) 32(1):20–39. doi:10.1016/j.neubiorev.2007.04.019
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
122. Gearhardt AN، Corbin WR، Brownell KD. الإدمان على الغذاء: فحص للمعايير التشخيصية للاعتماد. J Addict Med (2009) 3(1):1–7. doi:10.1097/ADM.0b013e318193c993
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
123. Smith DG، Robbins TW. الأسس العصبية الحيوية من السمنة والشراهة عند تناول الطعام: الأساس المنطقي لاعتماد نموذج الإدمان على الغذاء. بيول الطب النفسي (2013) 73(9):804–10. doi:10.1016/j.biopsych.2012.08.026
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
124. de Jong JW، Meijboom KE، Vanderschuren LJ، Adan RA. ويرتبط انخفاض السيطرة على تناول الطعام مستساغ في الفئران مع السلوك المعتاد والضعف الانتكاس: الفروق الفردية. بلوس واحد (2013) 8(9): e74645. دوى: 10.1371 / journal.pone.0074645
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
125. Furlong TM، Jayaweera HK، Balleine BW، Corbit LH. إن الاستهلاك الشبيه بشراهة من طعام مستساغ يسرع من السيطرة المعتادة على السلوك ويعتمد على تفعيل المخطط الظهاري الوحشي. J Neurosci (2014) 34(14):5012–22. doi:10.1523/JNEUROSCI.3707-13.2014
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
126. Hargrave SL، Davidson TL، Zheng W، Kinzig KP. الحمية الغربية تحفز تسرب الدم في الحاجز الدماغي وتغير الاستراتيجيات المكانية في الجرذان. Behav Neurosci (2016) 130(1):123–35. doi:10.1037/bne0000110
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
127. Smith KL، Hummer TA، Hulvershorn LA. ألعاب الفيديو المرضية وعلاقتها باضطرابات تعاطي المخدرات. Curr Addict Rep (2015) 2(4):302–9. doi:10.1007/s40429-015-0075-6
128. وينشتاين صباحا. إدمان ألعاب الكمبيوتر وألعاب الفيديو - مقارنة بين مستخدمي الألعاب وغير المستخدمين. صباحا J المخدرات تعاطي الكحول (2010) 36(5):268–76. doi:10.3109/00952990.2010.491879
129. Kätsyri J، Hari R، Ravaja N، Nummenmaa L. The importantent important: higher fMRI reward responses to winning against a human against a computer feeent during interactive video game playing. Cereb اللحاء (2013) 23(12):2829–39. doi:10.1093/cercor/bhs259
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
130. Kätsyri J، Hari R، Ravaja N، Nummenmaa L. مجرد مشاهدة اللعبة ليست كافية: مكافأة fMRI مخططي الاستجابة للنجاحات والفشل في لعبة فيديو أثناء اللعب النشط والمنفذ. الجبهة هوم نيوروسكي (2013) 7: 278. دوى: 10.3389 / fnhum.2013.00278
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
131. Erickson KI، Boot WR، Basak C، Neider MB، Prakash RS، Voss MW، et al. يتنبأ حجم القدرات بمستوى اكتساب مهارات ألعاب الفيديو. Cereb اللحاء (2010) 20: 2522-30. دوى: 10.1093 / cercor / bhp293
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
132. West GL، Drisdelle BL، Konishi K، Jackson J، Jolicoeur P، Bohbot VD. ترتبط لعبة ألعاب الفيديو الاعتيادية بالاستراتيجيات الملاحية المعتمدة على النواة. Proc R Soc B (2015) 282(1808). دوى: 10.1098 / rspb.2014.2952
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
133. Liu S، Schad DJ، Kuschpel MS، Rapp MA، Heinz A. Music and video gaming during breaks: impact of abitual versus goal-making decision making. ورقة مقدمة في الاجتماع السنوي 45 لجمعية علم الأعصاب. Chicago، IL: Society for Neuroscience (2015).
134. de Fonseca FR، Carrera MRA، Navarro M، Koob GF، Weiss F. Activation of corticotropin-releasing factor in the limbic system during cannabinoid withdraw. علوم (1997) 276(5321):2050–4. doi:10.1126/science.276.5321.2050
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
135. Cornelius JR، Chung T، Martin C، Wood DS، Clark DB. انسحاب القنب أمر شائع بين المراهقين الباحثين عن العلاج مع اعتماد القنب والاكتئاب الشديد ، ويترافق مع الانتكاس السريع للاعتماد. المدمن Behav (2008) 33(11):1500–5. doi:10.1016/j.addbeh.2008.02.001
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
136. Greene MC، Kelly JF. انتشار انسحاب القنب وتأثيره على استجابة ونتائج علاج المراهقين: تحقيقات مستقبلية في 12-month. J Addict Med (2014) 8: 359-67. دوى: 10.1097 / ADM.0000000000000064
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
137. واغنر فو ، أنتوني جي سي. من أول تعاطي المخدرات إلى الاعتماد على المخدرات ؛ فترات النمو من خطر الاعتماد على الماريجوانا والكوكايين والكحول. Neuropsychopharmacology (2002) 26:479–88. doi:10.1016/S0893-133X(01)00367-0
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
138. غودمان جي ، باكارد MG. تأثير القنب على عمليات التعلم والذاكرة من المخطط الظهري. Neurobiol تعلم Mem (2015) 125: 1-14. دوى: 10.1016 / j.nlm.2015.06.008
139. Rueda-Orozco PE، Soria-Gomez E، Montes-Rodriguez CJ، Martínez-Vargas M، Galicia O، Navarro L، et al. وظيفة محتملة من endocannabinoids في اختيار استراتيجية الملاحة عن طريق الفئران. علم الادوية النفسية (2008) 198(4):565–76. doi:10.1007/s00213-007-0911-z
140. غودمان جي ، باكارد MG. الحقن المخططة الطرفية وداخل الظهرية من ناهض مستقبلات القنب WIN 55,212-2 تضعف توحيد الذاكرة استجابة التحفيز. علم الأعصاب (2014) 274: 128-37. دوى: 10.1016 / j.neuroscience.2014.05.007
141. Nazzaro C، Greco B، Cerovic M، Baxter P، Rubino T، Trusel M، et al. يعمل تشكيل قناة SK على إنقاذ اللدونة والقدرة على التحمل على العادة في مجال تحمل القنب. نات نيوروسكي (2012) 15: 284-93. دوى: 10.1038 / nn.3022
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
142. Ames SL، Grenard JL، Stacy AW، Xiao L، He Q، Wong SW، et al. التصوير الوظيفي لارتباط الماريجوانا الضمني أثناء الأداء في اختبار ارتباط ضمني (IAT). Behav Brain Res (2013) 256: 494-502. دوى: 10.1016 / j.bbr.2013.09.013
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
143. Redish AD، Jensen S، Johnson A. A unified framework for addiction: vulnerabilities in the decision process. Behav Brain Sci (2008) 31(04):415–37. doi:10.1017/S0140525X0800472X
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
144. Brevers D، Bechara A، Cleeremans A، Noël X. Iowa Gambling Task (IGT): twenty years after - gambling disorder and IGT. Psychol الجبهة (2013) 4: 665. دوى: 10.3389 / fpsyg.2013.00665
145. Koob GF، Le Moal M. Neurobiological mechanisms for demonstrative motivational processes in addiction. Philos Trans R Soc B Biol Sci (2008) 363(1507):3113–23. doi:10.1098/rstb.2008.0094
146. Hyman SE، Malenka RC، Nestler EJ. الآليات العصبية للإدمان: دور التعلم والذاكرة المتعلقة بالجوائز. انو ريف نيوروسكي (2006) 29: 565-98. دوى: 10.1146 / annurev.neuro.29.051605.113009
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
147. Goodman J، Packard M. يحدد نظام الذاكرة أثناء عملية الاستحواذ فعالية بروتوكولات الانقراض المختلفة. الجبهة Behav Neurosci (2015) 9: 314. دوى: 10.3389 / fnbeh.2015.00314
148. Palencia CA، Ragozzino ME. مساهمة مستقبلات NMDA في المخطط الظهاري الجانبي لتعلم استجابة الأنانية. Behav Neurosci (2005) 119(4):953–60. doi:10.1037/0735-7044.119.4.953
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
149. Rueda-Orozco PE، Montes-Rodriguez CJ، Soria-Gomez E، Méndez-Díaz M، Prospéro-García O. Imprement of endocannabinoids activity in the dorsolateral striatum يؤخر انقراض السلوك في مهمة الذاكرة الإجرائية في الجرذان. الجهاز العصبي (2008) 55(1):55–62. doi:10.1016/j.neuropharm.2008.04.013
الكلمات الرئيسية: الذاكرة ، إدمان المخدرات ، الحصين ، المخطط ، اللوزة ، الإجهاد ، القلق
الاقتباس: غودمان جا و Packard MG (2016) أنظمة الذاكرة والمدمن الدماغ. أمامي. الطب النفسي 7: 24. doi: 10.3389 / fpsyt.2016.00024
تم الاستلام: 01 December 2015؛ تم القبول: 11 February 2016؛
نشرت: شنومكس فبراير شنومكس
حرره:
ديفيد فنسنت، National National de la Recherche Scientifique (CNRS)، France
تمت مراجعته من قبل:
جاك ميتشوجامعة بوردو 1 ، فرنسا
روبرتو Ciccocioppoجامعة كاميرينو ، إيطاليا
حقوق النشر: © 2016 Goodman و Packard. هذا هو مقال مفتوح الوصول موزعة وفقا لشروط ترخيص Creative Commons Attribution (CC BY). يُسمح بالاستخدام أو التوزيع أو الاستنساخ في المنتديات الأخرى بشرط أن يُنسب إلى المؤلف الأصلي (المؤلفين) أو المرخِّص الأصلي وأن المنشور الأصلي في هذه المجلة يتم الاستشهاد به ، وفقًا للممارسات الأكاديمية المقبولة. لا يسمح باستخدام أو توزيع أو إعادة إنتاج لا يتوافق مع هذه الشروط.
* المراسلات: مارك جي باكارد ، [البريد الإلكتروني محمي]