ديف ريف. 2008. 28(1): 62-77. دوى: 10.1016 / j.dr.2007.08.003
ملخص
المراهقة هي فترة تطورية تتميز بقرارات وأفعال دون المستوى الذي يؤدي إلى زيادة حدوث الإصابات غير المتعمدة والعنف ، وإدمان الكحول والمخدرات ، والحمل غير المقصود والأمراض المنقولة جنسياً. فشلت التفسيرات العصبية التقليدية والمعرفية لسلوك المراهقين في تفسير التغيرات غير الخطية في السلوك التي لوحظت خلال فترة المراهقة ، نسبة إلى الطفولة والبلوغ. تقدم هذه المراجعة تصوراً معقولاً بيولوجياً للآليات العصبية الكامنة وراء هذه التغيرات غير الخطية في السلوك ، كإستجابة متصاعدة للحوافز بينما لا يزال التحكم في الاندفاع غير ناضج نسبياً خلال هذه الفترة. توفر الدراسات الحديثة التي أجريت على الإنسان والحيوان أساسًا بيولوجيًا لهذه النظرة ، مما يوحي بالتطور التفاضلي لأنظمة المكافآت الحوفية المتعلقة بأنظمة التحكم من أعلى إلى أسفل خلال فترة المراهقة مقارنة بالطفولة والبلوغ. قد يتفاقم هذا النمط التنموي لدى هؤلاء المراهقين الذين لديهم ميول نحو المخاطرة ، مما يزيد من خطر النتائج السيئة.
وفقا للمركز الوطني للإحصاءات الصحية ، هناك أكثر من وفاة المراهقين 13,000 في الولايات المتحدة كل عام. ينجم ما يقرب من 70٪ من هذه الوفيات عن حوادث السيارات ، والإصابات غير المتعمدة ، والقتل ، والانتحار (إيتون وآخرون ، 2006). تظهر نتائج مسح سلوك الشباب الوطني 2005 (YRBS) أن المراهقين ينخرطون في سلوكيات تزيد من احتمالية الوفاة أو المرض عن طريق قيادة السيارة بعد الشرب أو بدون حزام الأمان وحمل السلاح واستخدام مواد غير مشروعة والانخراط في ممارسة الجنس بدون حماية مما أسفر عن الحمل غير المقصود والأمراض المنقولة جنسيا ، بما في ذلك العدوى بفيروس نقص المناعة البشرية (إيتون وآخرون ، 2006). تؤكد هذه الإحصائيات أهمية فهم الخيارات والإجراءات الخطرة في أوساط المراهقين.
تم افتراض عدد من الفرضيات المعرفية والبيولوجية العصبية لسبب إشراك المراهقين في سلوك الاختيار دون المستوى الأمثل. في مراجعة حديثة للأدبيات المتعلقة بتطور دماغ المراهق البشري ، Yurgelun-Todd (2007) يشير إلى أن التطور المعرفي من خلال سنوات المراهقة يرتبط مع زيادة تدريجية في كفاءة قدرات السيطرة الإدراكية. توصف هذه الكفاءة بأنها تعتمد على نضوج القشرة المخية قبل الجبهية كما يتضح من زيادة النشاط في مناطق ما قبل الجبهي البؤري (Rubia وآخرون ، 2000; تام ، مينون ، وريس ، 2002) ونقص النشاط في مناطق الدماغ غير ذات صلة (براون وآخرون ، 2005; Durston وآخرون ، 2006).
هذا النمط العام ، للتحكم المعرفي المحسّن مع نضوج قشرة الفص الجبهي ، يشير إلى زيادة خطية في النمو من الطفولة إلى البلوغ. ومع ذلك ، فإن الخيارات والإجراءات غير المثلى التي تمت ملاحظتها خلال فترة المراهقة تمثل تغيرًا غير خطي في السلوك يمكن تمييزه عن الطفولة والبلوغ ، كما يتضح من المركز الوطني للإحصاءات الصحية حول سلوك المراهقين ومعدل وفياتهم. إذا كان التحكم المعرفي وقشرة قبل الجبهية غير ناضجة هي أساس سلوك الاختيار دون المستوى الأمثل ، فيجب أن يبدو الأطفال متشابهين بشكل ملحوظ أو حتى أسوأ من المراهقين ، بالنظر إلى قشرة الفص الجبهي الأقل تطورا والقدرات المعرفية. وبالتالي ، فإن وظيفة قبل الجبهية غير ناضجة وحدها ، لا يمكن أن يفسر سلوك المراهقين.
يجب أن يعامل التصور الدقيق للتغييرات المعرفية والعصبية خلال فترة المراهقة ، مرحلة المراهقة على أنها فترة تطورية انتقالية (الرمح ، 2000) ، بدلاً من لقطة واحدة في الوقت (كيسي ، توتنهام ، ليستون ، ودورستون ، 2005). وبعبارة أخرى ، لفهم هذه الفترة التنموية ، تعتبر التحولات من سن المراهقة والخروج منها ضرورية لتمييز الصفات المميزة لهذه المرحلة من التطور. إن إرساء مسارات تنموية للعمليات المعرفية والعصبية أمر ضروري لتوصيف هذه التحولات وتقييد التفسيرات حول التغيرات في السلوك خلال هذه الفترة. على المستوى الإدراكي أو السلوكي ، يصف المراهقون على أنهم مندفعون (أي يفتقرون إلى السيطرة الإدراكية) ويأخذون المخاطر مع هذه التركيبات المستخدمة بشكل مترادف ودون تقدير للمسارات التنموية المتميزة لكل منها. على مستوى البيولوجيا العصبية ، تقترح الدراسات التصويرية والحيوانية البشرية قواعد بيولوجية عصبية متميزة ومسارات تنموية للأنظمة العصبية التي تكمن وراء هذه التركيبات المنفصلة من التحكم في الانفعالات والقرارات الخطرة.
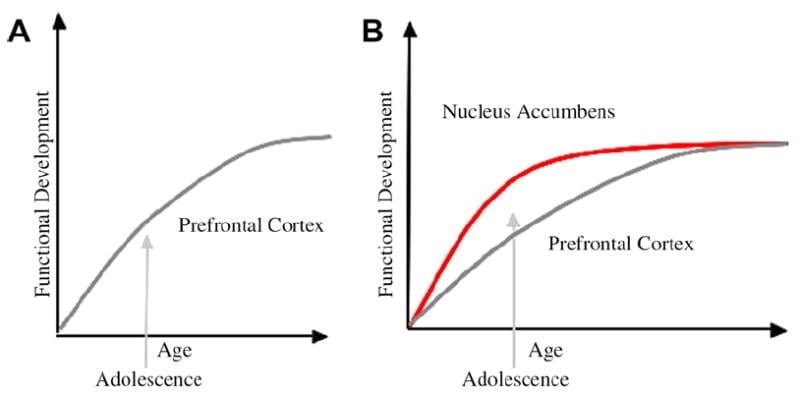
لقد طورنا نموذجًا بيولوجيًا عصبيًا لتطور المراهقين في هذا الإطار الذي يعتمد على نماذج القوارض (Laviola، Adriani، Terranova، & Gerra، 1999; الرمح ، 2000) ودراسات التصوير الحديثة من المراهقة (Ernst et al.، 2005; جالفان ، هير ، فوس ، جلوفر ، وكيسي ، 2007; Galvan et al.، 2006). التين 1 أدناه يصور هذا النموذج. على اليسار هو التوصيف التقليدي للمراهقة كما هو مرتبط بشكل شبه حصري بنضج قشرة الفص الجبهي. على اليمين يوجد نموذجنا العصبي الحيوي المقترح الذي يوضح كيف يجب اعتبار المناطق القاعية تحت القاعية والجبهة القبلية من أعلى لأسفل. يوضح الرسم البياني المسارات التنموية المختلفة لهذه الأنظمة ، مع تطور الأنظمة الحوفية في وقت مبكر عن مناطق السيطرة الأمامية. ووفقًا لهذا النموذج ، يكون الفرد متحيزًا بشكل أكبر من خلال المناطق الحوفية الناضجة وظيفيًا خلال فترة المراهقة (أي اختلال التوازن الحوفي بالنسبة للسيطرة الأمامية) مقارنة بالأطفال ، الذين لا تزال هذه الأنظمة (أي الحوفي والجانب الجبهي) في طور النمو. ومقارنة مع البالغين ، الذين تنضج هذه الأنظمة بالكامل. يوفر هذا المنظور أساسًا للتحولات غير الخطية في السلوك عبر التطوير ، نظرًا للنضج المبكر لهذا الجهاز الحوفي نسبة إلى منطقة التحكم الأمامية الجبهية الأقل نضجا من أعلى لأسفل. مع التطوير والخبرة ، يوفر الاتصال الوظيفي بين هذه المناطق آلية للتحكم من أعلى إلى أسفل في هذه المناطق (هير ، فوس ، جلوفر ، وكيسي ، 2007 أ). علاوة على ذلك ، يوفق النموذج بين تناقض الإحصاءات الصحية للسلوك المحفوف بالمخاطر خلال فترة المراهقة ، مع الملاحظة الذكية رينا وفرلي (2006) أن المراهقين قادرون على فهم وفهم مخاطر السلوكيات التي ينخرطون فيها. وفقا لنموذجنا ، في الحالات البارزة عاطفيا ، فإن النظام الحوفي سيفوز على أنظمة التحكم بالنظر إلى نضجه بالنسبة لنظام التحكم الجبهي. يتم توفير أدلة من الدراسات التصويرية السلوكية والبشرية لدعم هذا النموذج في سياق الإجراءات في سياقات مجزية وعاطفية (Galvan et al.، 2006, 2007; هير ، فوس ، جلوفر ، وكيسي ، 2007 ب; هير وآخرون ، 2007a). بالإضافة إلى ذلك ، فإننا نتوقع لماذا قد يتطور الدماغ بهذه الطريقة ، ولماذا قد يكون بعض المراهقين معرضين لخطر أكبر لاتخاذ قرارات دون المستوى مما يؤدي إلى نتائج طويلة الأجل أفقر (Galvan et al.، 2007; هير وآخرون ، 2007b).
وقد اقترح أن التفسير التقليدي لسلوك المراهقين يرجع إلى التطور المطول لقشرة الفص الجبهي (A). يأخذ نموذجنا في الاعتبار تطوير قشرة الفص الجبهي جنبا إلى جنب مع المناطق القاعية تحت القشرية (على سبيل المثال ، النواة المتكئة) التي تورطت في الخيارات والإجراءات الخطرة (ب).
تطوير السلوك الموجه نحو الهدف
حجر الزاوية في التطور المعرفي هو القدرة على قمع الأفكار والإجراءات غير المناسبة لصالح الأهداف الموجهة ، خاصة في وجود حوافز مقنعة (كيسي ، جالفان ، وهير ، 2005; كيسي وآخرون ، 2000b; كيسي ، توماس ، ديفيد-سون ، كونز ، وفرانزين ، 2002 أ; كيسي ، توتنهام ، وفوسيلا ، 2002 ب). أظهر عدد من الدراسات التنموية الكلاسيكية أن هذه القدرة تتطور خلال مرحلة الطفولة والمراهقة (القضية ، 1972; فلافيل ، فيتش ، وتشينسكي ، 1966; كيتنغ وبوبيت ، 1978; باسكوال ليون ، 1970). جادل العديد من أصحاب النظريات بأن التطور المعرفي يرجع إلى زيادة سرعة المعالجة وكفاءتها وليس بسبب زيادة القدرات العقلية (على سبيل المثال ، Bjorkland ، 1985; Bjorkland ، 1987; القضية ، 1985). شمل المنظرين الأخرين بناء العمليات "المثبطة" في حسابهم للتطور المعرفي (هارنيشفيغر وبيوركلاند ، 1993). وفقا لهذا الحساب ، يتميز الإدراك غير الناضج بالتأثر بالتداخل من المصادر المتنافسة التي يجب قمعها (على سبيل المثال ، برينرد ورينا ، 1993; كيسي ، توماس ، ديفيدسون ، كونز ، وفرانزين ، 2002 أ; Dempster، 1993; الماس ، 1985; موناكاتا ويريس ، 2001). وبالتالي فإن السلوك الموجه نحو الأهداف يتطلب التحكم في الدوافع أو تأخير الإشباع من أجل تحسين النتائج ، ويبدو أن هذه القدرة تنضج عبر الطفولة والمراهقة.
وقد وصف سلوك المراهقين بأنه متهور ومحفوف بالمخاطر ، بشكل شبه مرادف ، إلا أن هذه التركيبات تعتمد على العمليات المعرفية والعصبية المختلفة ، التي توحي ببنيات مميزة مع مسارات تنموية مختلفة. على وجه التحديد ، تشير مراجعة الأدبيات إلى أن الاندفاع يتناقص مع التقدم في العمر عبر مرحلة الطفولة والمراهقة (كيسي وآخرون ، 2002a; Casey، Galvan et al.، 2005; Galvan et al.، 2007) ويرتبط بالتطور المطول لقشرة الفص الجبهي (Casey، Galvan et al.، 2005) ، رغم وجود اختلافات في الدرجة التي يكون فيها فرد معين مندفعًا أم لا ، بغض النظر عن العمر.
وعلى النقيض من ذلك ، فإنه بالنسبة للسيطرة على الحركة / الإدراك ، يبدو أن أخذ المخاطر يزداد خلال فترة المراهقة مقارنة بالطفولة والبلوغ ، ويرتبط بالنظم تحت القشرية المعروفة بأنها تشارك في تقييم المكافآت. تقترح دراسات التصوير البشري التي سيتم مراجعتها ، زيادة في التنشيط تحت القشري (على سبيل المثال ، المتكئة) عند اتخاذ خيارات محفوفة بالمخاطر (Kuhnen & Knutson ، 2005; ماثيوز وآخرون ، 2004; مونتاج و بيرنز ، 2002) مبالغ فيه في المراهقين ، بالنسبة للأطفال والبالغين (Ernst et al.، 2005; Galvan et al.، 2006). تقترح هذه النتائج مسارات مختلفة للسلوك القائم على المكافأة أو الحافز ، مع التطور المبكر لهذه الأنظمة المتعلقة بأنظمة التحكم التي تظهر مسارًا تطويريًا مطولًا وخطيًا ، من حيث تجاوز الخيارات والإجراءات غير الملائمة لصالح الأهداف الموجهة.
أدلة من دراسات التصوير العصبي من تطور الدماغ البشري
وقد استندت الأبحاث الحديثة في تطوير دماغ المراهقين إلى التقدم في منهجيات التصوير العصبي التي يمكن استخدامها بسهولة مع نمو البشر. تعتمد هذه الطرق على طرق التصوير بالرنين المغناطيسي (MRI) التين 2) وتشمل: التصوير بالرنين المغناطيسي هيكلية ، والذي يستخدم لقياس حجم وشكل الهياكل ؛ التصوير بالرنين المغناطيسي الوظيفي الذي يستخدم لقياس أنماط نشاط الدماغ. والتصوير الموسع للانتشار (DTI) والذي يستخدم لفهرسة الربط بين مساحات ألياف المادة البيضاء. ويدعم الدليل على نموذجنا التنموي للمنافسة بين المناطق القشرية وتحت القشرية عن طريق الاتصال الهيكلي والوظيفي غير الناضج كما تم قياسه بواسطة DTI و fMRI ، على التوالي.
دراسات التصوير بالرنين المغناطيسي لتطوير دماغ الإنسان
وقد استخدمت العديد من الدراسات التصوير بالرنين المغناطيسي هيكلية لرسم خريطة المسار التشريحي لتطور الدماغ الطبيعي (انظر مراجعة قبل Durston وآخرون ، 2001). على الرغم من أن حجم الدماغ الكلي يبلغ حوالي 90٪ من حجمه البالغ عند بلوغه السادسة ، فإن المكونات الفرعية للمادة الرمادية والبيضاء في الدماغ تستمر في إجراء تغييرات ديناميكية خلال فترة المراهقة. تشير البيانات المستقاة من دراسات التصوير بالرنين المغناطيسي الطويلة الطولية إلى أن حجم المادة الرمادية له نمط U معكوس ، مع وجود اختلاف إقليمي أكبر من المادة البيضاء (Giedd ، 2004; Gogtay وآخرون ، 2004; سويل وآخرون ، 2003; سويل ، طومسون ، وتوجا ، 2004). وبوجه عام ، فإن المناطق التي تقوم بوظائف أساسية ، مثل الأنظمة الحركية والحركية ، تنضج في أقرب وقت ممكن ؛ مناطق الارتباط الأعلى ، والتي تدمج هذه الوظائف الأساسية ، تنضج لاحقًا (Gogtay وآخرون ، 2004; سويل ، طومسون ، وتوجا ، 2004). على سبيل المثال ، تظهر الدراسات التي تستخدم المقاييس المستندة إلى التصوير بالرنين المغناطيسي أن فقدان المادة الرمادية القشرية يحدث في أقرب وقت في المناطق الحسية الأولية وأحدث في القِسَف الأمامية الجبهية الأمامية الجبهية والصدرية الجانبية (Gogtay وآخرون ، 2004). يتناسب هذا النمط مع دراسات ما بعد الرئيسيات غير البشرية والرئيسية التي تظهر أن القشرة المخية قبل الجبهية هي واحدة من آخر مناطق الدماغ تنضج (بورجوا ، جولدمان-راكيتش ، وراكيتش ، 1994; Huttenlocher ، 1979). على النقيض من المادة الرمادية ، يزداد حجم المادة البيضاء في نمط خطي تقريبًا ، ويتزايد خلال التطور إلى مرحلة البلوغ (Gogtay وآخرون ، 2004). يفترض أن هذه التغييرات تعكس الميالين المستمر للمحاور من قبل الخلايا قليلة التغصن التي تعزز التوصيل العصبي والاتصالات.
على الرغم من الاهتمام الأقل بالمناطق تحت القشرية عند فحص التغيرات الهيكلية ، إلا أن بعض التغيرات الكبرى في الدماغ عبر التنمية تظهر في هذه المناطق ، خاصة في العقد القاعدية (سويل وآخرون ، 1999، انظر التين 3) وخاصة في الذكور (Giedd وآخرون ، 1996). التغيرات التنموية في الحجم الهيكلي في العقد القاعدية والمناطق قبل الجبهية مثيرة للاهتمام في ضوء العمليات التنموية المعروفة (على سبيل المثال ، الشجرة الشجيرية ، موت الخلايا ، التشذيب المتشابك ، الميالين) التي تحدث أثناء الطفولة والمراهقة. تسمح هذه العمليات بضبط وتدعيم الصلات بين المناطق قبل الجبهية وتحت القشرية مع التطوير والتعلم الذي قد يتزامن مع تحكم إدراكي أكبر. كيف ترتبط هذه التغييرات الهيكلية بالتغييرات المعرفية؟ هناك عدد من الدراسات التي تتعلق بالنضج الهيكلي للفص الجبهي والدالة المعرفية باستخدام التدابير النفسية العصبية والإدراكية (على سبيل المثال ، سويل وآخرون ، 2003). على وجه التحديد ، تم الإبلاغ عن الارتباطات بين أحجام القشرة القبلية الأمامية القاعدية القائمة على التصوير بالرنين المغناطيسي والقياسات الأساسية للسيطرة الإدراكية (أي القدرة على تجاوز الاستجابة غير الملائمة لصالح أخرى أو لخفض الانتباه نحو خاصية التحفيز غير ذات الصلة لصالح سمة التحفيز ذات الصلة)كيسي ، ترينور وآخرون ، 1997). تشير هذه النتائج إلى أن التغييرات المعرفية تنعكس في التغيرات الهيكلية في الدماغ وتؤكد أهمية القشرة القشرية تحت القشرية بالإضافة إلى القشرة القشرية (على سبيل المثال ، القشرة المخية قبل الجبهية).

دراسات DTI لتطوير الدماغ البشري
تشير دراسات القثاء المستندة إلى التصوير بالرنين المغنطيسي التي تم استعراضها إلى أن الروابط القشرية يتم ضبطها بشكل جيد مع القضاء على الإفراط في المشابك وتقوية الروابط ذات الصلة بالتنمية والخبرة. تقدم التطورات الحديثة في تكنولوجيا التصوير بالرنين المغناطيسي ، مثل DTI أداة محتملة لدراسة دور مسالك معينة من المادة البيضاء لتطوير الدماغ والسلوك بمزيد من التفصيل. تتعلق هذه الورقة بدراسات التصوير العصبي التي ربطت تطوير مساحات الألياف مع تحسينات في القدرة المعرفية. على وجه التحديد ، وقد أظهرت الجمعيات بين مقاييس DTI المستندة إلى تطوير المادة البيضاء قبل الجبهية والسيطرة الإدراكية لدى الأطفال. في دراسة واحدة ، كان تطور هذه القدرة مرتبطًا بشكل إيجابي بمناطق الألياف قبل الجبهية الجدارية (ناجي ، ويستربرج ، وكلينجبيرج ، 2004) بما يتفق مع الدراسات التصويرية للأعصاب الوظيفية التي تظهر التجميع التفاضلي لهذه المناطق في الأطفال مقارنة بالبالغين.
باستخدام نهج مماثل ، ليستون وآخرون. (2005) أظهرت أن المادة البيضاء تتسرب بين العقد القَبْليَّة القَبَليَّة القَطَنية وكتل الألياف الداعمة تستمر في النمو خلال الطفولة إلى مرحلة البلوغ ، ولكن هذه السلالم فقط بين القشرة المخية قبل الجبهية والعقد القاعدية ترتبط بالتحكم في الانفعالات ، مقاسة بالأداء على go / nogo مهمة. تم تحديد مساحات الألياف قبل الجبهية عن طريق مناطق المصالح المحددة في دراسة الرنين المغناطيسي الوظيفي باستخدام نفس المهمة. في كل من دراسات DTI النمائية ، تم ربط مقاييس الألياف مع التطور ، ولكن تم توضيح خصوصية بعض مسالك الألياف مع الأداء المعرفي عن طريق فصل المسالك المعينة (Liston وآخرون ، 2005) أو القدرة المعرفية (ناجي وآخرون ، 2004). هذه النتائج تؤكد على أهمية دراسة ليس فقط التغييرات الإقليمية ، ولكن الدارات ذات الصلة عند تقديم مطالبات حول التغيرات التي تعتمد على العمر في ركائز العصبية من التطور المعرفي.
دراسات التصوير بالرنين المغناطيسي الوظيفي من التطور السلوكي والدماغ
على الرغم من أن التغيرات الهيكلية التي تم قياسها بواسطة التصوير بالرنين المغناطيسي و DTI قد ارتبطت بالتغييرات السلوكية أثناء التطوير ، فإن المقاربة الأكثر مباشرة لفحص ارتباط البنية - الوظيفة هي قياس التغيرات في الدماغ والسلوك في وقت واحد ، كما هو الحال مع الرنين المغناطيسي الوظيفي. القدرة على قياس التغييرات الوظيفية في الدماغ النامية مع التصوير بالرنين المغناطيسي لديها إمكانات كبيرة في مجال العلوم التنموية. في سياق الورقة الحالية ، يوفر الرنين المغناطيسي الوظيفي وسيلة للحد من تفسيرات سلوك المراهقين. كما ذكرنا سابقًا ، يُعتقد أن تطوير القشرة المخية قبل الجبهية يلعب دورًا مهمًا في نضوج القدرات المعرفية الأعلى مثل اتخاذ القرار والتحكم المعرفي (كيسي وتوتنهام وفوسيلا 2002 ب; كيسي ، ترينور وآخرون ، 1997). تم استخدام العديد من النماذج ، جنبا إلى جنب مع fMRI ، لتقييم الأساس العصبي لهذه القدرات ، بما في ذلك المطراف ، Stroop والمهام go / nogo (Casey، Castellanos et al.، 1997; Casey، Giedd، & Thomas، 2000a; Durston وآخرون ، 2003). وتظهر هذه الدراسات مجتمعة أن الأطفال يجندون مناطق قبل الفصوص متميزة ولكنها غالباً أكبر وأكثر انتشاراً عند أداء هذه المهام من البالغين. يصبح نمط النشاط داخل مناطق الدماغ المركزية لأداء المهام (أي ، الذي يرتبط بالأداء المعرفي) أكثر تركيزًا أو ضبطًا دقيقًا مع التقدم في العمر ، في حين أن المناطق غير المرتبطة بأداء المهام تتضاءل في النشاط مع التقدم في السن. وقد لوحظ هذا النمط عبر كلا المقطع العرضي (براون وآخرون ، 2005) والدراسات الطولية (Durston وآخرون ، 2006) وعبر مجموعة متنوعة من النماذج. على الرغم من أن الدراسات التصويرية للتصوير العصبي لا يمكن أن تميز بشكل محدد آلية مثل هذه التغييرات التنموية (على سبيل المثال ، التشجير الشجيرية ، التقليم المتشابك) ، فإن النتائج تعكس التطور داخل ، وصقل التوقعات من وإلى ، وتنشيط مناطق الدماغ مع النضج ، وتشير إلى أن هذه التغييرات تحدث على فترة طويلة فترة من الزمن (براون وآخرون ، 2005; بونج ، دودوكوفيتش ، توماسون ، فيديا ، وغابرييلي ، 2002; كيسي ، ترينور وآخرون ، 1997; كيسي وآخرون ، 2002a; كرون ، دونوهو ، هونوميشل ، وينديلكين ، وبانج ، 2006; لونا وآخرون ، 2001; موسى وآخرون ، 2002; Schlaggar وآخرون ، 2002; Tamm et al.، 2002; توماس وآخرون، 2004; Turkeltaub، Gareau، Flowers، Zeffiro، & Eden، 2003).
كيف يمكن لهذه المنهجية أن تطلعنا على ما إذا كان المراهقون يفتقرون بالفعل إلى السيطرة الإدراكية الكافية (المندفعة) أم أنهم ينطوي على مخاطر في خياراتهم وأفعالهم؟ يظهر التحكم في الاندفاع الذي تم قياسه بواسطة مهام التحكم الإدراكي مثل مهمة go / nogo نمطًا خطيًا للتطور عبر الطفولة والمراهقة كما هو موضح أعلاه. ومع ذلك ، فقد بدأت دراسات التصوير العصبي الحديثة بدراسة المعالجة ذات الصلة بالمكافأة الخاصة بالمخاطرة عند المراهقين (Bjork et al.، 2004; Ernst et al.، 2005; مايو وآخرون ، 2004). وقد ركزت هذه الدراسات في المقام الأول على منطقة المتكئين ، وهي جزء من العقد القاعدية تشارك في التنبؤ بالمكافأة ، بدلا من توصيف تطور هذه المنطقة بالتزامن مع مناطق التحكم من الأعلى إلى الأسفل (القشرة الأمامية الجبهية). على الرغم من أن تقرير حديث عن نشاط قبل الجبهي أقل فطرية في المراهقين بالنسبة للبالغين خلال مهمة صنع القرار النقدي بشأن سلوك المخاطرة قد تم عرضه (إيشيل ونيلسون وبلير وباين وإرنست 2007).
بشكل عام ، قامت بعض الدراسات بفحص كيفية تطور دارات المكافأة في المناطق دون القشرية (على سبيل المثال ، المتكئة) بالتزامن مع تطور المناطق قبل الجبهية القشرية. علاوة على ذلك ، كيف تتزامن هذه التغييرات العصبية مع سلوكيات المكافأة ، والاندفاعية ، وسلوك المخاطرة لا تزال غير معروفة نسبياً. يقترح نموذجنا العصبي البيولوجي أن الجمع بين الاستجابة المتصاعدة للمكافآت وعدم النضج في مناطق السيطرة السلوكية قد يحرض المراهقين على السعي للحصول على مكاسب فورية ، وليس طويلة الأجل ، وربما يفسرون الزيادة في اتخاذ القرارات الخطرة والسلوكيات الاندفاعية. يوفر تتبع القشرة تحت القشرية (على سبيل المثال ، المتكئفين) والقشرية (على سبيل المثال ، قبل الجبهي) في عملية صنع القرار عبر مرحلة الطفولة إلى مرحلة البلوغ ، قيودًا إضافية على ما إذا كانت التغييرات التي يتم الإبلاغ عنها في مرحلة المراهقة محددة لهذه الفترة من التطور ، أو تعكس النضج الذي يحدث بشكل مطرد في نمط خطي إلى حد ما من الطفولة إلى مرحلة البلوغ.
الأدلة التجريبية من دراسة الرنين المغناطيسي الوظيفي الأخيرة تساعد على دعم نموذجنا العصبي الحيوي وتتخذ نهجًا انتقاليًا لفهم المراهقة من خلال فحص التغييرات السابقة للمراهقة وبعدها. في هذه الدراسة (Galvan et al.، 2006) ، فحصنا الاستجابات السلوكية والعصبية لمكافأة التلاعب عبر التنمية ، مع التركيز على مناطق الدماغ المتورطة في التعلم والسلوك المتعلق بالثقافة في الحيوانات (هيكوساكا وواتانابي ، 2000; Pecina، Cagniard، Berridge، Aldridge، & Zhuang، 2003; شولتز ، 2006) ودراسات التصوير للبالغين (على سبيل المثال ، كنوتسون وآدامز وفونغ وهومر 2001; O ، Doherty ، Kringelbach ، Rolls ، Hornak ، Andrews ، 2001; Zald وآخرون ، 2004) وفي دراسات الإدمان (هايمان ومالينكا ، 2001; فولكو ولي ، 2004). بناء على نماذج القوارضLaviola et al.، 1999; الرمح ، 2000) وأعمال التصوير السابقة (Ernst et al.، 2005) ، افترضنا أنه بالنسبة إلى الأطفال والبالغين ، سيظهر المراهقون نشاطًا مبالغًا فيه للتكثيف ، بالتنسيق مع توظيف أقل نضجًا لمناطق التحكم الأمامية الجبهية من أعلى لأسفل. إن الأعمال الحديثة التي تبين تأخر الاتصال الوظيفي بين هذه المناطق قبل الجبهية والحورية تحت القشرية في مرحلة المراهقة بالنسبة للبالغين ، توفر آلية للافتقار إلى التحكم من أعلى إلى أسفل في هذه المناطق (هير وآخرون ، 2007a).
كانت النتائج التي توصلنا إليها متسقة مع نماذج القوارض (Laviola، Macri، Morley-Fletcher، & Adriani، 2003) ودراسات التصوير السابقة (Ernst et al.، 2005) يقترح تعزيز النشاط المتكئ على المكافآت خلال فترة المراهقة. في الواقع ، بالنسبة إلى الأطفال والبالغين ، أظهر المراهقون استجابة متكافلة مبالغ فيها تحسبًا للمكافأة. ومع ذلك ، أظهر كل من الأطفال والمراهقين استجابة أقل نضجا في مناطق السيطرة قبل الجبهية من البالغين. تشير هذه النتائج إلى أن المسارات التنموية المختلفة لهذه المناطق قد تكمن في تعزيز النشاط المترابط ، بالنسبة إلى الأطفال أو البالغين ، والذي قد يرتبط بدوره بسلوكيات الاندفاعية والمحفوفة بالمخاطر التي لوحظت خلال هذه الفترة من التطور (انظر التين 4).

تم الإبلاغ عن التوظيف التفاضلي للمناطق قبل الجبهية وتحت القشرية عبر عدد من الدراسات التنموية الرنين المغناطيسي الوظيفي (كيسي وآخرون ، 2002b; Monk et al.، 2003; توماس وآخرون، 2004). عادة ما يتم تفسير هذه النتائج من حيث المناطق قبل الجبهية غير ناضجة بدلا من عدم التوازن بين التنمية الإقليمية قبل الجبهية وتحت القشرية. تقديم أدلة على المناطق قبل الجبهية في توجيه الإجراءات المناسبة في سياقات مختلفة (ميلر وكوهين ، 2001) قد يعوق النشاط الجبهي غير الناضج التقدير المناسب للنتائج المستقبلية وتقييم الخيارات الخطرة ، وبالتالي قد يكون أقل تأثيرا على تقييم المكافأة من المتكئين. يتطابق هذا النمط مع الأبحاث السابقة التي تظهر ارتفاع القشرة تحت القشرية ، نسبة إلى النشاط القشري عندما تتحيز القرارات من خلال المكاسب الفورية على المدى الطويل (مكلور ، لايبسون ، لوينشتاين ، وكوهين ، 2004). علاوة على ذلك ، تم إظهار نشاط المتكاتف مع الرنين المغناطيسي الوظيفي للارتباط بشكل إيجابي مع سلوكيات المخاطرة التالية (Kuhnen & Knutson ، 2005). خلال فترة المراهقة ، بالنسبة إلى مرحلة الطفولة أو مرحلة البلوغ ، قد لا توفر القشرة المخية قبل الجبهية غير الناضجة تحكمًا كافيًا من الأعلى إلى الأسفل لمناطق معالجة المكافأة النشطة تنشيطًا (على سبيل المثال ، المتكئة) ، مما يؤدي إلى تأثير أقل على أنظمة الفص الجبهي (القشرة الأمامية الأمامية) نسبة إلى المتكئة في المكافأة تقييم.
لماذا تتم برمجة الدماغ لتطوير هذه الطريقة؟
المراهقة هي الفترة الانتقالية بين الطفولة والبلوغ وغالبا ما تحدث مع سن البلوغ. البلوغ يمثل بدايات النضج الجنسي (Graber & Brooks-Gunn ، 1998) ويمكن تحديدها بواسطة علامات بيولوجية. يمكن وصف المراهقة بأنها انتقال تدريجي إلى مرحلة البلوغ مع دورة زمنية مبكِّرة للجينوم (الرمح ، 2000). ومن الناحية النظرية ، فإن فترة المراهقة هي الفترة التي يتم فيها اكتساب مهارات الاستقلال لزيادة النجاح عند الانفصال عن حماية الأسرة ، على الرغم من زيادة فرص الظروف الضارة (مثل الإصابة والاكتئاب والقلق وتعاطي المخدرات والإدمان)كيلي وشوشيه ولاندري ، 2004). تنتشر السلوكيات التي تسعى إلى الاستقلالية عبر الأنواع ، مثل الزيادات في التفاعلات الاجتماعية الموجهة من الأقران وتكثيف السلوكيات التي تبحث عن الجدة والمخاطرة. العوامل النفسية والاجتماعية تؤثر على نزوع المراهقين للسلوك الخطر. ومع ذلك ، فإن السلوك المحفوف بالمخاطر هو نتاج اختلال توازن بيولوجي بين زيادة الجدة والإحساس بالاقتران مع "الكفاءة التنظيمية الذاتية" غير الناضجة (شتاينبرغ ، 2004). تشير بياناتنا البيولوجيا العصبية إلى حدوث ذلك من خلال التطوير التفاضلي لهذين النظامين (الحوفي والتحكم).
قد تشير التكهنات إلى أن هذا النمط التنموي هو ميزة تطورية. أنت بحاجة إلى الانخراط في سلوك شديد الخطورة لترك عائلتك وقريةك للعثور على رفيقة ومجازفة في نفس الوقت الذي تدفع فيه الهرمونات إلى المراهقين للبحث عن شركاء جنسيين. في مجتمع اليوم ، قد تمتد فترة المراهقة إلى أجل غير مسمى ، مع الأطفال الذين يعيشون مع والديهم ولديهم الاعتماد المالي واختيار الزملاء في وقت لاحق من الحياة ، قد يعتبر هذا التطور غير مناسب.
هناك أدلة عبر الأنواع لزيادة البحث عن الجدة والمخاطرة خلال سنوات المراهقة. البحث عن الأقران من نفس الجنس والقتال مع الأهل ، والتي تساعد جميعًا في جعل المراهق بعيدًا عن أرض الوطن للتزاوج يُرى في الأنواع الأخرى بما في ذلك القوارض ، والرئيسيات غير البشرية وبعض الطيور (الرمح ، 2000). بالنسبة للبالغين ، تظهر الفئران ذات الجين الزائدة سلوكيات بحثية جديدة في نموذج حدا الاختيار الحر (Laviola et al.، 1999). تشير الدلائل الكيميائية العصبية إلى أن التوازن في الدماغ المراهق بين أنظمة الدوبامين القشرية ودون القشرية ، يبدأ في التحول نحو مستويات الدوبامين القشرية الأكبر خلال فترة المراهقة (الرمح ، 2000). وقد ظهر إنزيم دوبرامين طويل الأمد مماثل خلال فترة المراهقة إلى مرحلة البلوغ في قشرة فصائل ما قبل الجبهية البدائية أيضًا.روزنبرغ ولويس ، 1995). وهكذا يبدو أن هذا الارتفاع الواضح في المخاطرة يكون عبر الأنواع ولها أغراض تكيفية هامة.
الاستعدادات البيولوجية ، والتنمية ، والمخاطر
تم التعرف على الفروق الفردية في السيطرة على الدوافع والمخاطر في علم النفس لبعض الوقت (بينثين وسلوفيتش وسيفرسون 1993). ولعل أحد الأمثلة الكلاسيكية للاختلافات الفردية التي ذكرت في هذه القدرات في أدب علم النفس الاجتماعي والمعرفي والنمو هو تأخير الإشباع (ميشيل ، شودا ، ورودريجيز ، 1989). يتم تقييم تأخير الإشباع عادة في الأطفال 3- لأطفال عمرهم 4 عام. يسأل الطفل ما إذا كانوا يفضلون مكافأة صغيرة (ملف تعريف ارتباط واحد) أو مكافأة كبيرة (اثنين من ملفات تعريف الارتباط). ثم يُخبر الطفل أن المجرب سيغادر الغرفة من أجل التحضير للأنشطة القادمة ويشرح للطفل أنه إذا بقيت في مقعدها ولم يأكل الكعكة ، فستحصل على المكافأة الكبيرة. إذا كان الطفل لا أو لا يمكنه الانتظار ، فعليه قرع الجرس لاستدعاء المجرب وبالتالي الحصول على المكافأة الأصغر. بمجرد أن يتضح أن الطفل يفهم المهمة ، فهي جالسة على الطاولة مع المكافتين والجرس. يتم تقليل الانحرافات في الغرفة ، مع عدم وجود ألعاب أو كتب أو صور. يعود المجرب بعد مرور 15 دقيقة أو بعد أن يدق الطفل الجرس ، أو يؤكل المكافآت ، أو يظهر أي علامات الشدة. أظهر Mischel أن الأطفال يتصرفون عادة بإحدى الطريقتين التاليتين: (1) إما أنهم يقرعون الجرس على الفور تقريبًا من أجل الحصول على الكوكي ، مما يعني أنهم يحصلون على واحد فقط. أو (2) ينتظرون ويحسنون مكاسبهم ، ويستقبلون كلا من ملفات تعريف الارتباط. تشير هذه الملاحظة إلى أن بعض الأفراد أفضل من الآخرين في قدرتهم على التحكم في الدوافع في مواجهة الحوافز البارزة للغاية ويمكن ملاحظة هذا التحيز في مرحلة الطفولة المبكرة (Mischel et al.، 1989) ويبدو أنها ستبقى طوال فترة المراهقة وفترة الشباب (Eigsti وآخرون ، 2006).
ما الذي يمكن أن يفسر الاختلافات الفردية في اتخاذ القرار والسلوك الأمثل؟ وقد افترض بعض المنظرين أن دارة الدوبامين الدوبامينية ، المتورطة في معالجة المكافآت ، تكمن وراء السلوك المحفوف بالمخاطر. قد تكون الاختلافات الفردية في هذه الدائرة ، مثل المتغيرات الأليلية في الجينات ذات الصلة بالدوبامين ، والتي ينتج عنها القليل جدا أو الكثير من الدوبامين في المناطق تحت القشرية ، مرتبطة بالميل إلى الانخراط في سلوك محفوف بالمخاطر (O'Doherty ، 2004). أظهرت النواة المتكئة زيادة في النشاط مباشرة قبل اتخاذ خيارات محفوفة بالمخاطر على نماذج المخاطر النقدية (Kuhnen & Knutson ، 2005; ماثيوز وآخرون ، 2004; مونتاج و بيرنز ، 2002) وكما هو موضح سابقًا ، يظهر المراهقون نشاطًا متكاليًا مبالغ فيه لمكافأة النتائج المتعلقة بالأطفال أو البالغين (Ernst et al.، 2005; Galvan et al.، 2006). بشكل جماعي ، تشير هذه البيانات إلى أن المراهقين قد يكونون أكثر عرضة للاختبارات الخطرة كمجموعة (بستاني وشتاينبرغ ، 2005) ، ولكن بعض المراهقين سيكونون أكثر عرضة من غيرهم للانخراط في سلوكيات محفوفة بالمخاطر ، مما يعرضهم لخطر أكبر على النتائج السلبية. لذلك من المهم النظر إلى التباين الفردي عند فحص العلاقات المعقدة بين الدماغ والسلوك المتعلقة بالمجازفة والمكافأة في المجموعات التطورية.
لاستكشاف الفروق الفردية في سلوك المخاطرة ، جالفان وآخرون. (2007) فحصت مؤخرا العلاقة بين النشاط في الدوائر العصبية المتعلّقة بالجوّ تحسبًا لمكافأة مالية كبيرة مع مقاييس السمات الشخصية للمجازفة والاندفاع في مرحلة المراهقة. تم الحصول على التصوير بالرنين المغناطيسي الوظيفي والمقاييس المجهولة لتقرير التقييم الذاتي للسلوك المحفوف بالمخاطر ، وتصور المخاطر والاندفاع لدى الأفراد الذين تتراوح أعمارهم بين 7 و 29 سنوات. كان هناك ارتباط إيجابي بين نشاط المتكئين واحتمال الانخراط في سلوك محفوف بالمخاطر عبر التنمية. تباين هذا النشاط كدالة لتقييم الأفراد للنتائج الإيجابية أو السلبية المتوقعة لمثل هذا السلوك. الأفراد الذين ينظرون إلى السلوكيات المحفوفة بالمخاطر بأنها تؤدي إلى عواقب وخيمة ، نشطوا المتكئفين أقل لمكافأة. كان الدافع وراء هذا الارتباط إلى حد كبير من قبل الأطفال ، مع تصنيف البالغين عواقب مثل هذا السلوك ممكن. لم ترتبط تصنيفات الاندفاع مع نشاط المتكئين ، ولكن مع التقدم في السن. تشير هذه النتائج إلى أنه خلال فترة المراهقة ، قد يكون بعض الأفراد أكثر عرضة للانخراط في سلوكيات محفوفة بالمخاطر بسبب التغيرات التنموية بالتنسيق مع التغير في استعداد فرد معين للانخراط في سلوك محفوف بالمخاطر ، بدلاً من التغييرات البسيطة في الاندفاع (انظر التين 5).

تم وصف سلوك المراهقين بشكل متهور ومحفوف بالمخاطر (شتاينبرغ ، 2004, 2007) ، ولكن هذا الاستعراض لأدبيات التصوير يشير إلى ركائز عصبية بيولوجية مختلفة ومسارات تنموية مختلفة لهذه السلوكيات. على وجه التحديد ، يرتبط الاندفاع بتطور جبهي فطري غير ناضج ويقلل تدريجياً من الطفولة إلى البلوغ (Casey، Galvan et al.، 2005). الارتباط السلبي بين درجات الاندفاع والسن في الدراسة بواسطة جالفان وآخرون. (2007) يدعم كذلك هذه الفكرة. في المقابل ، يرتبط أخذ المخاطر بزيادة في النشاط المتكئ (Kuhnen & Knutson ، 2005; ماثيوز وآخرون ، 2004; مونتاج و بيرنز ، 2002) ، وهذا مبالغ فيه في المراهقين ، بالنسبة للأطفال والبالغين (Ernst et al.، 2005; Galvan et al.، 2006). وهكذا لا يمكن تفسير اختيارات وسلوك المراهقين عن طريق الاندفاع أو التطور المطول لقشرة الفص الجبهي فقط ، حيث يتوقع أن يكون الأطفال أكثر عرضة للمخاطر. توفر هذه النتائج أساسًا عصبيًا لكون بعض المراهقين معرضين لخطر أكبر من غيرهم ، ولكنها توفر أيضًا أساسًا لكيفية اختلاف سلوك المراهقين عن الأطفال والبالغين الذين يواجهون المخاطر.
بشكل جماعي ، توحي هذه البيانات أنه على الرغم من اعتبار المراهقين كمجموعة من المتقدمين للمخاطر (بستاني وشتاينبرغ ، 2005) ، سيكون بعض المراهقين أكثر عرضة من غيرهم للانخراط في سلوكيات محفوفة بالمخاطر ، مما يعرضهم لخطر أكبر على النتائج السلبية. تؤكد هذه النتائج على أهمية النظر في التباين الفردي عند فحص العلاقات المعقدة بين الدماغ والسلوك المتعلقة بالمجازفة والمكافأة في المجموعات التطورية. علاوة على ذلك ، قد تساعد هذه الاختلافات الفردية والتنموية في تفسير ضعف بعض الأفراد في اتخاذ المخاطر المرتبطة بتعاطي المخدرات ، وفي النهاية ، الإدمان.
استنتاجات
تظهر الدراسات التصويرية البشرية التغيرات الهيكلية والوظيفية في المناطق الأمامية ()Giedd وآخرون ، 1996, 1999; Jernigan et al.، 1991; سويل وآخرون ، 1999. للمراجعة، Casey، Galvan et al.، 2005) التي يبدو أنها زيادات موازية في السيطرة الإدراكية والتنظيم الذاتي (كيسي ، ترينور وآخرون ، 1997; لونا وسويني ، 2004; لونا وآخرون ، 2001; Rubia وآخرون ، 2000; شتاينبرغ ، 2004وانظر أيضا: شتاينبرغ ، 2008، هذه المسألة). يبدو أن هذه التغييرات تظهر تحولًا في تنشيط مناطق ما قبل الجبهية من الانتشار إلى مزيد من التعيين المركزي بمرور الوقت (براون وآخرون ، 2005; Bunge et al.، 2002; كيسي ، ترينور وآخرون ، 1997; Durston وآخرون ، 2006; موسى وآخرون ، 2002) والتجنيد المرتفع للمناطق دون القشرية خلال فترة المراهقة (كيسي وآخرون ، 2002a; Durston وآخرون ، 2006; لونا وآخرون ، 2001). على الرغم من أن دراسات التصوير العصبي لا يمكن أن تميز بشكل محدد آلية مثل هذه التغييرات التنموية ، فإن هذه التغيرات في الحجم والبنية قد تعكس التطور داخل ، وصقل التوقعات من وإلى مناطق الدماغ هذه أثناء النضج مما يوحي بضبط النظام مع التطور.
مجتمعة ، تشير النتائج التي تم تجميعها هنا إلى أن السلوك المتزايد للمخاطرة في مرحلة المراهقة يرتبط مع مسارات تنموية مختلفة للمناطق دون القشرية ومناطق السيطرة القشرية. يمكن أن تتفاقم هذه التغييرات التنموية من خلال الفروق الفردية في نشاط أنظمة المكافآت. على الرغم من أن المراهقة قد تميزت كفترة تتميز بسلوكيات المكافأة والتعامل مع المخاطر (بستاني وشتاينبرغ ، 2005; الرمح ، 2000الفروق الفردية في الاستجابات العصبية للمكافأة ، تؤهب بعض المراهقين ليأخذوا مخاطر أكثر من غيرهم ، مما يجعلهم أكثر عرضة للنتائج السلبية. توفر هذه النتائج أساسًا حيويًا من خلال تجميع النتائج المختلفة المتعلقة بسلوك المخاطرة في مرحلة المراهقة وفهم الفروق الفردية والعلامات التنموية من أجل الميل إلى الانخراط في السلوك السلبي.
شكر وتقدير
وأيد هذا العمل في جزء من المنح المقدمة من المعهد الوطني للإدمان على المخدرات R01 DA18879 والمعهد الوطني للصحة العقلية 1P50 MH62196.
مراجع حسابات
- بينتين ألف ، Slovic P ، سيفرسون H. دراسة نفسية من إدراك المخاطر للمراهقين. مجلة المراهقة. 1993.16: 153-168. [مجلات]
- Bjork JM، Knutson B، Fong GW، Caggiano DM، Bennett SM، Hommer DW. تنشيط الدماغ الحافز - التنشيط في المراهقين: أوجه التشابه والاختلاف بين الشباب. مجلة علم الأعصاب. 2004.24: 1793-1802. [مجلات]
- Bjorkland DF. دور المعرفة المفاهيمية في تطوير المنظمة في ذاكرة الأطفال. In: Brainerd CJ، Pressley M، editors. العمليات الأساسية في تطوير الذاكرة: التقدم في بحوث التنمية المعرفية. نيويورك: سبرينغر فيرلاغ 1985. pp. 103 – 142.
- Bjorkland DF. تساهم التغيرات العمرية في قاعدة المعرفة في تطوير ذاكرة الأطفال: مراجعة تفسيرية. مراجعة تنموية. 1987.7: 93-130.
- Bourgeois JP، Goldman-Rakic PS، Rakic P. Synaptogenesis in the prefrontal cortex of rhesus monkeys. القشرة الدماغية. 1994.4: 78-96. [مجلات]
- Brainerd CJ، Reyna VF. استقلال الذاكرة وتداخل الذاكرة في التطور المعرفي. مراجعة نفسية. 1993.100: 42-67. [مجلات]
- Brown TT، Lugar HM، Coalson RS، Miezin FM، Petersen SE، Schlaggar BL. التغييرات التنموية في تنظيم وظيفي دماغي الإنسان لتوليد الكلمات. القشرة الدماغية. 2005.15: 275-290. [مجلات]
- Bunge SA، Dudukovic NM، Thomason ME، Vaidya CJ، Gabrieli JD. مساهمات الفص الجبهي غير الناضجة في السيطرة الإدراكية عند الأطفال: أدلة من الرنين المغناطيسي الوظيفي. الخلايا العصبية. 2002.33: 301-311. [مجلات]
- حالة R. التحقق من بناء القدرات Piagetian الجديد. مجلة علم نفس الطفل التجريبي. 1972.14: 287-302.
- حالة ر. التطور الفكري: الولادة حتى سن البلوغ. New York: Academic Press؛ 1985.
- Casey BJ، Castellanos FX، Giedd JN، Marsh WL، Hamburger SD، Schubert AB. وآخرون. تداعيات الحق في الدوائر الأمامية في التغلب على رد الفعل واضطراب نقص الانتباه / فرط النشاط. مجلة الأكاديمية الأمريكية للطب النفسي للأطفال والمراهقين. 1997.36: 374-383. [مجلات]
- Casey BJ، Galvan A، Hare TA. التغييرات في تنظيم وظيفي دماغي أثناء التطور المعرفي. الرأي الحالي في علم الأعصاب. 2005.15: 239-244. [مجلات]
- Casey BJ، Giedd JN، Thomas KM. تطور الدماغ البنيوي والوظيفي وعلاقته بالتطور المعرفي. علم النفس البيولوجي. 2000a.54: 241-257. [مجلات]
- Casey BJ، Thomas KM، Davidson MC، Kunz K، Franzen PL. فصل الوظيفة المخطط والحصين بطريقة تنموية مع مهمة توافق التحفيز والاستجابة. مجلة علم الأعصاب. 2002a.22: 8647-8652. [مجلات]
- Casey BJ، Thomas KM، Welsh TF، Badgaiyan RD، Eccard CH، Jennings JR، et al. تفكك نزاع الاستجابة والاختيار المقصود والمتوقع مع التصوير بالرنين المغناطيسي الوظيفي. وقائع الأكاديمية الوطنية للعلوم. 2000b.97: 8728-8733.
- Casey BJ، Tottenham N، Fossella J. Clinical، imaging، lesion and genetic approach into a model of cognitive control. علم النفس النفسي التنموي. 2002b.40: 237-254. [مجلات]
- Casey BJ، Tottenham N، Liston C، Durston S. Imaging the develop your brain: ماذا تعلمنا عن التطور المعرفي؟ الاتجاهات في العلوم المعرفية. 2005.9: 104-110.
- Casey BJ، Trainor RJ، Orendi JL، Schubert AB، Nystrom LE، Giedd JN، et al. دراسة التصوير بالرنين المغناطيسي الوظيفي التنموي من التنشيط قبل الجبهي أثناء أداء مهمة go-no-go. مجلة علم الأعصاب الإدراكي. 1997.9: 835-847.
- Crone E، Donohue S، Honomichl R، Wendelken C، Bunge S. Brain regional mediating flexible rule use during development. مجلة علم الأعصاب. 2006.26: 11239-11247. [مجلات]
- Dempster FN. مقاومة التداخل: التغيرات التنموية في آلية المعالجة الأساسية. In: Howe ML، Pasnak R، editors. الموضوعات الناشئة في التنمية المعرفية Volume 1: Foundations. نيويورك: سبرينغر. 1993. pp. 3 – 27.
- الماس ألف تطوير القدرة على استخدام استدعاء لتوجيه العمل ، كما يتضح من أداء الرضع على AB. نمو الطفل. 1985.56: 868-883. [مجلات]
- Durston S، Davidson MC، Thomas KM، Worden MS، Tottenham N، Martinez A، et al. التلاعب حدودي من الصراع ومنافسة الاستجابة باستخدام الرنين المغناطيسي الوظيفي السريعة ذات الصلة الحدث التجريبي. Neuroimage. 2003.20: 2135-2141. [مجلات]
- Durston S، Davidson MC، Tottenham N، Galvan A، Spicer J، Fossella J، et al. التحول من الانتشار إلى النشاط القشري البؤري مع التطور. العلوم التنموية. 2006.1: 18-20. [مجلات]
- Durston S، Hulshoff Pol HE، Casey BJ، Giedd JN، Buitelaar JK، van Engeland H. Anatomical MRI of the human human brain: What have learn؟ مجلة الأكاديمية الأمريكية للطب النفسي للمراهقين. 2001.40: 1012-1020.
- Eaton LK، Kinchen S، Ross J، Hawkins J، Harris WA، Lowry R، et al. مراقبة سلوكيات مخاطر الشباب - الولايات المتحدة ، 2005 ، ملخصات المراقبة. المراضة والوفيات تقرير أسبوعي. 2006.55: 1-108. [مجلات]
- Eigsti IM، Zayas V، Mischel W، Shoda Y، Ayduk O، Dadlani MB، et al. التنبؤ بالسيطرة المعرفية من مرحلة ما قبل المدرسة وحتى المراهقة المتأخرة وصغر سن الرشد. العلوم النفسية. 2006.17: 478-484. [مجلات]
- Ernst M، Nelson EE، Jazbec S، McClure EB، Monk CS، Leibenluft E، et al. اللوزة والنواة المتكئة في الردود على استلام وإغفال المكاسب في البالغين والمراهقين. Neuroimage. 2005.25: 1279-1291. [مجلات]
- Eshel N، Nelson EE، Blair RJ، Pine DS، Ernst M. Neural substrates of choice selection in adults and adults: Development of ventrolateral prefrontal and front fronted cortices. Neuropsychologia. 2007.45: 1270-1279. [بك المادة الحرة] [مجلات]
- Flavell JH، Feach DR، Chinsky JM. بروفة شفوية عفوية في مهمة ذاكرة كدالة للعمر. نمو الطفل. 1966.37: 283-299. [مجلات]
- Galvan A، Hare TA، Parra CE، Penn J، Voss H، Glover G، et al. إن التطور السابق للمتلازمين بالنسبة لقشرة الدماغ الأمامية يمكن أن يكمن وراء سلوك المخاطرة عند المراهقين. مجلة علم الأعصاب. 2006.26: 6885-6892. [مجلات]
- Galvan A، Hare T، Voss H، Glover G، Casey BJ. المخاطرة والمخ المراهق: من هو في خطر؟ العلوم التنموية. 2007.10: F8-F14. [مجلات]
- Gardener M، Steinberg L. Peer effect on risk taking، risk preferference، and risky making making in adolescence and adulthood: An experimental study. علم النفس التنموي. 2005.41: 625-635. [مجلات]
- جيد. التصوير بالرنين المغناطيسي الهيكلي للمخ سن المراهقة. حوليات أكاديمية نيويورك للعلوم. 2004.1021: 77-85. [مجلات]
- Giedd JN، Blumenthal J، Jeffries NO، Castellanos FX، Liu H، Zijdenbos A، et al. تطور الدماغ أثناء الطفولة والمراهقة: دراسة التصوير بالرنين المغناطيسي طوليًا. الطبيعة العصبية. 1999.2: 861-863.
- Giedd JN، Snell JW، Lange N، Rajapakse JC، Casey BJ، Kozuch PL، et al. التصوير بالرنين المغناطيسي الكمي لتطوير دماغ الإنسان: Ages 4 – 18. القشرة الدماغية. 1996.6: 551-560. [مجلات]
- Gogtay N، Giedd JN، Lusk L، Hayashi KM، Greenstein D، Vaituzis AC، et al. رسم الخرائط الديناميكي للتنمية القشرية البشرية خلال الطفولة وحتى سن البلوغ المبكر. وقائع الأكاديمية الوطنية للعلوم في الولايات المتحدة الأمريكية. 2004.101: 8174-8179. [بك المادة الحرة] [مجلات]
- Graber JA، Brooks-Gunn J. Puberty. In: Blechman EA، Brownell KD، editors. الطب السلوكي والمرأة دليل شامل. نيويورك ، نيويورك: مطبعة جويلفورد. 1998. pp. 51 – 58.
- Hare TA، Voss HU، Glover GH، Casey BJ. دماغ المراهقين والمخاطر المحتملة للقلق والاكتئاب. 2007a مقدم للنشر.
- Hare TA، Voss HU، Glover GH، Casey BJ. تبرز المنافسة بين النظم الحبيبية الأمامية الجبهية والحشوية تحت القفوية التفاعلية خلال مرحلة المراهقة. 2007b مقدم للنشر.
- Harnishfeger KK، Bjorkland F. The ontogeny of inhib mechanism: A renewed approach to cognitive development. In: Howe ML، Pasnek R، editors. الموضوعات الناشئة في التنمية المعرفية. المجلد. 1. نيويورك: سبرينغر فيرلاغ 1993. pp. 28 – 49.
- Hikosaka K، Watanabe M. تأخير نشاط العصبونات الفدارية الأمامية والجانبية للقردة مع اختلاف المكافآت. القشرة الدماغية. 2000.10: 263-271. [مجلات]
- Huttenlocher PR. كثافة متشابكة في القشرة الأمامية للإنسان - التغيرات التنموية وآثار الشيخوخة. بحوث الدماغ. 1979.163: 195-205. [مجلات]
- Hyman SE، Malenka RC. الإدمان والدماغ: البيولوجيا العصبية للإكراه واستمراره. مراجعات الطبيعة 2001.2: 695-703.
- Jernigan TL، Zisook S، Heaton RK، Moranville JT، Hesselink JR، Braff DL. تشوهات التصوير بالرنين المغناطيسي في النويات العدسية والقشرة الدماغية في الفصام. المحفوظات للطب النفسي العام. 1991.48: 811-823.
- Keating DP، Bobbitt BL. الفروق الفردية والتنموية في مكونات المعالجة المعرفية للقدرة العقلية. نمو الطفل. 1978.49: 155-167.
- Kelley AE، Schochet T، Landry C. حوليات أكاديمية نيويورك للعلوم. 2004.1021: 27-32. [مجلات]
- Knutson B، Adams CM، Fong GW، Hommer D. Anticipation of increase money reward reward selectively recruits nucleus accumbens. مجلة علم الأعصاب. 2001.21: RC159. [مجلات]
- Kuhnen CM، Knutson B. The neural basis of financial risk taking. الخلايا العصبية. 2005.47: 763-770. [مجلات]
- Laviola G، Adriani W، Terranova ML، Gerra G. عوامل الخطر النفسية للتعرض للمخاطر النفسية للمراهقين من البشر والنماذج الحيوانية. علم الأعصاب والمراجعات Biobehavior. 1999.23: 993-1010. [مجلات]
- Laviola G، Macri S، Morley-Fletcher S، Adriani W. ملخص سلوكيات المخاطرة في الفئران المراهقين: محددات نفسية بيولوجية وتأثير متجانس مبكر. علم الأعصاب والمراجعات Biobehavior. 2003.27: 19-31. [مجلات]
- Liston C، Watts R، Tottenham N، Davidson MC، Niogi S، Ulug AM، et al. إن البنية المجهرية للدماغ الأمامي تعدل التوظيف الفعال للسيطرة الإدراكية. القشرة الدماغية. 2005.16: 553-560. [مجلات]
- لونا B ، سويني JA. ظهور وظيفة الدماغ التعاونية: دراسات FMRI من تطوير تثبيط الاستجابة. حوليات أكاديمية نيويورك للعلوم. 2004.1021: 296-309. [مجلات]
- Luna B، Thulborn KR، Munoz DP، Merriam EP، Garver KE، Minshew NJ، et al. ينضج نضج وظيفة الدماغ الموزعة على نطاق واسع التطور المعرفي. Neuroimage. 2001.13: 786-793. [مجلات]
- Matthews SC، et al. التنشيط الانتقائي للنواة المتكئة أثناء اتخاذ القرار المخاطرة. Neuroreport. 2004.15: 2123-2127. [مجلات]
- May JC، Delgado MR، Dahl RE، Stenger VA، Ryan ND، Fiez JA، et al. التصوير بالرنين المغناطيسي الوظيفي المرتبط بالحدث المتعلق بدائرة الدماغ ذات الصلة بالمكافئات عند الأطفال والمراهقين. الطب النفسي البيولوجي. 2004.55: 359-366. [مجلات]
- McClure SM، Laibson DI، Loewenstein G، Cohen JD. الأنظمة العصبية المنفصلة تقدر المكافآت المالية الفورية. العلم. 2004.306: 503-507. [مجلات]
- Miller EK، Cohen JD. نظرية تكاملية لوظيفة القشرة الجبهية. المراجعة السنوية لعلم الأعصاب. 2001.24: 167-202.
- Mischel W، Shoda Y، Rodriguez MI. تأجيل الإشباع في الأطفال. العلم. 1989.244: 933-938. [مجلات]
- Monk CS، McClure EB، Nelson EE، Zarahn E، Bilder RM، Leibenluft E، et al. عدم النضج لدى المراهقين في ارتباط الدماغ المرتبط بالانتباه بتعبيرات الوجه العاطفية. Neuroimage. 2003.20: 420-428. [مجلات]
- Montague PR، Berns GS. الاقتصاد العصبي والركائز البيولوجية للتقييم. الخلايا العصبية. 2002.36: 265-284. [مجلات]
- Moses P، Roe K، Buxton RB، Wong EC، Frank LR، Stiles J. Functional MRI of global and local processing in children. Neuroimage. 2002.16: 415-424. [مجلات]
- Munakata Y، Yerys BE. جميعا معا الآن: عندما تختفي الفوارق بين المعرفة والعمل. العلوم النفسية. 2001.12: 335-337.
- ناجي Z ، Westerberg H ، Klingberg T. يرتبط نضج المادة البيضاء مع تطور الوظائف المعرفية أثناء الطفولة. مجلة علم الأعصاب الإدراكي. 2004.16: 1227-1233. [مجلات]
- O'Doherty J، Kringelbach ML، Rolls ET، Hornak J، Andrews C. Abstract reward and punishment representations in the human thebitofrontal cortex. الطبيعة العصبية. 2001.4: 95-102. [مجلات]
- O'Doherty JP. تمثيل المكافآت والتعلم المتصل بالمكافأة في الدماغ البشري: رؤى من تصوير الأعصاب. الآراء الحالية في علم الأعصاب. 2004.14: 769-776.
- باسكوال-ليون جيه. نموذج رياضي للانتقال في مراحل تطور بياجيه. اكتا Psychologica. 1970.32: 301-345.
- Pecina S، Cagniard B، Berridge KC، Aldridge JW، Zhuang X. Hyperdopaminergic mutant mice have higher "wanting" but not "liking" for sweet rewards. مجلة علم الأعصاب. 2003.23: 9395-9402. [مجلات]
- Reyna VF، Farley F. المخاطر والعقلانية في صنع القرار للمراهقين: الآثار المترتبة على النظرية والممارسة والسياسة العامة. العلوم النفسية في المصلحة العامة. 2006.7: 1-44.
- روزنبرغ DR ، لويس DA. النضج بعد الولادة من تعصيب الدوباميني من القشرة الأمامية والخلفية القرد الحركي: A tyrosine hydroxylase immunohistochemical analysis. مجلة علم الأعصاب المقارن. 1995.358: 383-400. [مجلات]
- Rubia K، Overmeyer S، Taylor E، Brammer M، Williams SC، Simmons A، et al. جبهات وظيفية مع تقدم العمر: رسم خرائط المسارات النمائية العصبية مع fMRI. علم الأعصاب والمراجعات Biobehavior. 2000.24: 13-19. [مجلات]
- Schlaggar BL، Brown TT، Lugar HM، Visscher KM، Miezin FM، Petersen SE. الاختلافات التشوهية العصبية الوظيفية بين البالغين والأطفال في سن المدرسة في معالجة الكلمات المفردة. العلم. 2002.296: 1476-1479. [مجلات]
- نظريات Schultz W. السلوكية وعلم الأعصاب من المكافأة. الاستعراضات السنوية لعلم النفس. 2006.57: 87-115.
- Sowell ER، Peterson BS، Thompson PM، Welcome SE، Henkenius AL، Toga AW. رسم تغيير القشرية عبر أعمار البشر. الطبيعة العصبية. 2003.6: 309-315.
- Sowell ER، Thompson PM، Holmes CJ، Jernigan TL، Toga AW. في الدليل على نضوج الدماغ في مرحلة ما بعد المراهقة في المناطق الأمامية والموجهة. الطبيعة العصبية. 1999.2: 859-861.
- Sowell ER، Thompson PM، Toga AW. التغييرات في رسم الخرائط في القشرة البشرية طوال فترة الحياة. الأعصاب. 2004.10: 372-392. [مجلات]
- الرمح ليرة لبنانية. الدماغ في سن المراهقة والمظاهر السلوكية المرتبطة بالعمر. علم الأعصاب والمراجعات Biobehavior. 2000.24: 417-463. [مجلات]
- شتاينبرغ لام. أخذ المخاطر في مرحلة المراهقة: ما هي التغييرات ، ولماذا؟ حوليات أكاديمية نيويورك للعلوم. 2004.1021: 51-58. [مجلات]
- شتاينبرغ لام. أخذ المخاطر في مرحلة المراهقة: وجهات نظر جديدة من الدماغ والعلوم السلوكية. الاتجاهات الحالية في العلوم النفسية. 2007.16: 55-59.
- شتاينبرغ ل. منظور علم الأعصاب الاجتماعي حول الأخذ بالمراهقين. مراجعة تنموية. 2008.28: 78-106. [بك المادة الحرة] [مجلات]
- Tamm L، Menon V، Reiss AL. نضج وظيفة الدماغ المرتبطة تثبيط الاستجابة. مجلة الأكاديمية الأمريكية للطب النفسي للأطفال والمراهقين. 2002.41: 1231-1238. [مجلات]
- Thomas KM، Hunt RH، Vizueta N، Sommer T، Durston S، Yang Y، et al. دليل على الاختلافات التطورية في التعلم التسلسلي الضمني: دراسة FMRI للأطفال والكبار. مجلة علم الأعصاب الإدراكي. 2004.16: 1339-1351. [مجلات]
- Turkeltaub PE، Gareau L، Flowers DL، Zeffiro TA، Eden GF. تطوير الآليات العصبية للقراءة. الطبيعة العصبية. 2003.6: 767-773.
- Volkow ND، Li TK. الإدمان على المخدرات: إن العصبية السلوكية ذهبت بعيدا. مراجعات الطبيعة 2004.5: 963-970.
- Yurgelun-Todd D. التغييرات العاطفية والإدراكية خلال فترة المراهقة. الرأي الحالي في علم الأعصاب. 2007.17: 251-257. [مجلات]
- Zald DH، Boileau I، El-Dearedy W، Gunn R، McGlone F، Dichter GS، et al. انتقال الدوبامين في المخطط البشري أثناء مهام المكافأة النقدية. مجلة علم الأعصاب. 2004.24: 4105-4112. [مجلات]