أمامي. Behav. Neurosci.، 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 مسرور ر. شريف1, ارنولد بيلمر1, ماثيو جي فوغارتي2, إيريكا WH مو2,
مسرور ر. شريف1, ارنولد بيلمر1, ماثيو جي فوغارتي2, إيريكا WH مو2, 

- 1معهد بحوث الترجمة ومعهد الصحة والابتكار الطبي الحيوي ، جامعة كوينزلاند للتكنولوجيا ، بريسبان ، كوينزلاند ، أستراليا
- 2كلية العلوم الطبية الحيوية ، جامعة كوينزلاند ، بريسبان ، كوينزلاند ، أستراليا
أصبح النظام الغذائي الحديث محليا للغاية ، مما أدى إلى مستويات غير مسبوقة من استهلاك السكر ، ولا سيما بين المراهقين. في حين أن المدخول المزمن طويل الأجل من السكر معروف أنه يسهم في تطوير الاضطرابات الأيضية بما في ذلك السمنة والنوع الثاني من داء السكري ، إلا أنه لا يُعرف إلا القليل عن العواقب المباشرة لاستهلاك السكر على المدى الطويل مثل الشراهة على الدماغ. بيمكن أن يسبب السكر المسبب للإفراز الدوبامين في النواة المتكئة (NAc) بشكل مشابه لعقاقير الإدمان ، حيث قمنا بفحص التغيرات في مورفولوجية الخلايا العصبية في هذه المنطقة الدماغية بعد فترة قصيرة (أسابيع 4) وطويلة الأجل (أسابيع 12) مثل استهلاك السكروز باستخدام نموذج اختيار زجاجة متقطعة اثنين. استخدمنا تلطيخ Golgi-Cox لتلقيح الخلايا العصبية الشوكية المتوسطة (MSNs) من نواة NAc وقذيفة من الفئران المستهلكة على المدى القصير والطويل السكروز وقارنت هذه إلى ضوابط المياه المتطابقة مع العمر. نظهر أن استهلاك السكروز لفترات طويلة يشبه إلى حد كبير انخفاض إجمالي طول شجيري من MSN NAc قذيفة مقارنة مع الفئران السيطرة العمرية المتطابقة. وجدنا أيضا أن إعادة هيكلة هذه الخلايا العصبية نتجت في المقام الأول من تعقيد شجيري البعيدة. بالمقابل ، لاحظنا زيادة كثافة العمود الفقري في أوامر فرع البعيدة من NAc shell MSN من الفئران المستهلكة على السكروز على المدى الطويل. مجتمعة ، هذه النتائج تسليط الضوء على الآثار العصبية من تناول الشراهة لفترات طويلة مثل السكروز على مورفولوجيا NAc قذيفة MSN.
المُقدّمة
خلال السنوات الماضية 40 ، كان هناك زيادة موثقة في استهلاك المشروبات المحلاة بالسكر والأطعمة التي تحتوي على السكريات المضافة (Nielsen et al.، 2002; Popkin ، 2010; Ng et al.، 2012) ، مع تقارير تشير إلى أن ما يصل إلى 75 ٪ من جميع الأطعمة والمشروبات تحتوي على كميات كبيرة من السكريات المضافة (فورد و Dietz ، 2013; براي وبوبكين ، 2014). خلال هذه الفترة ، كان هناك أيضا زيادة متزامنة في انتشار السمنة والنوع الثاني من داء السكري ، لا سيما بين المراهقين (أرسلانيان ، 2002; Reinehr ، 2013; Dabelea et al.، 2014; Fryar وآخرون ، 2014). أظهرت دراسات حديثة أن الأطفال الذين يعانون من زيادة الوزن والسمنة غالبا ما يستهلكون كميات كبيرة من السكر المضاف ، إلا أن مساهمة الوجبات التي تحتوي على نسبة عالية من السكر في زيادة حالات زيادة الوزن والبدانة لا تزال مثيرة للجدل (هو ، 2013; براي وبوبكين ، 2014; Bucher Della Torre et al.، 2015).
في حين أن مجموعة متزايدة من الأدلة تشير إلى أن استهلاك الوجبات الغذائية عالية السكر قد يسهم ، جزئيا ، في زيادة الوزن بين الأطفال والمراهقين (مالك وآخرون ، 2010; Te Morenga et al.، 2013; براي وبوبكين ، 2014) ، وقد تم إيلاء اهتمام أقل إلى الآثار السلبية غير الأيضية الناجمة عن تناول السكر المفرط. ومن المثير للاهتمام أن بعض الأنماط السلوكية والنفسية الشائعة تظهر في الغالب بين مجموعة فرعية من أولئك الذين يأكلون ويحافظون على الوجبات الغذائية التي تحتوي على نسبة عالية من السكر. أبرزها تطور اضطرابات الأكل بما في ذلك الأكل بنهم ، جنبا إلى جنب مع الظهور المتزامن للأعراض النفسية بما في ذلك عدم وجود الدافع والاكتئاب (مراجعة في شيهان وهيرمان ، 2015). بالإضافة إلى ذلك ، نظرًا لأن الأفراد الذين يتناولون الطعام غالباً ما يظهرون فقدان السيطرة وعدم القدرة على الحد من تناولهم للسكر ، فمن المرجح أن هذه السلوكيات تنشأ نتيجة للتكيف العصبي في مناطق الدماغ التي تقيِّم القيمة الطيبة للأطعمة شديدة الاستساغة. (Saper et al.، 2002; Lutter و Nestler ، 2009; كيني ، 2011). هذا الأساس المنطقي مدعوم أيضًا بالدلائل في البشر التي تدل على أن السكر والحلاوة يمكن أن تسبب الرغبة الشديدة التي تتشابه مع تلك الناجمة عن المخدرات الإدمانية مثل الكحول والنيكوتين (Volkow وآخرون ، 2012).
على الرغم من أن خصائص الإدمان للسكر لا تزال مضاربة ، إلا أن هذه الملاحظات تضافرت مع الدراسات التي تدل على مساهمة تناول السكر المفرط للتغيرات في دائرة المكافأة وتطوير السلوكيات المشابهة للادمان والحالات العاطفية في النماذج الحيوانية (Avena et al.، 2008; بنتون ، 2010; Ventura et al.، 2014), يضمن الحاجة لمزيد من التحقيق. وقد أظهرت الدراسات السابقة في القوارض أن الوصول المتقطع إلى السكروز يغير نشاط العديد من الناقلات العصبية ضمن نظام ميزوليببيك بما في ذلك الدوبامين والأفيونات والأسيتيل كولين (تمت مراجعته في Avena et al.، 2008). وقد أظهر الاستهلاك الشبيه بالسكر من السكروز لتسهيل إطلاق الدوبامين في النواة المتكئة (NAc) ، وبالمثل للأدوية من تعاطي (Avena et al.، 2008). وعلاوة على ذلك ، فقد أظهرنا أن استهلاك السكروز على المدى الطويل باستخدام نموذج 24 h متقطعة اختيار اثنين من زجاجة الاختيار (سيمز وآخرون ، 2008ينظم تعبير مستقبلات أستيل النيكوتين (nAChR) في NAc (Shariff وآخرون ، في الصحافة). ومن المثير للاهتمام أننا لاحظنا أيضًا أن مركبات nAChR المعروفة بتعديل نشاط الدوبامين والأسيتيل كولين في NAc ، لها تأثيرات مختلفة على استهلاك السكروز بعد تناول قصير وطويل الأجل (Shariff وآخرون ، في الصحافة).
في حين أظهرت هذه الدراسات أوجه تشابه في التغيرات السلوكية والكيميائية العصبية الناجمة عن الوصول المتقطع للسكر ومخدرات سوء الاستخدام ، فإنه من غير المعروف ما إذا كانت هذه التأثيرات تسهل التغيرات في التشكل العصبوني في NAc. Tله على النقيض من المواد من تعاطي بما في ذلك الكوكايين والأمفيتامين والنيكوتين التي تنتج تغييرات متقنة في مورفولوجيا الخلايا العصبية الشوكية المتوسطة (MSNs) في NAc ، بما في ذلك زيادة كثافة العمود الفقري وتعديل التغصنات المتغيرة (روبنسون و Kolb ، 1999, 2004; لي وآخرون ، 2003; Crombag et al.، 2005). لأننا أظهرنا من قبل أن التعرض الطويل الأمد (أسبوع 12) للكحول والسكروز باستخدام نموذج الإختيار المتقطع للزجاجتين ينتج استجابة تفاضلية للتدخلات العلاجية الدوائية مقارنة بالمدخول قصير المدى (أسابيع 4؛ Steensland et al.، 2007; Shariff وآخرون ، في الصحافة) ، قمنا بتقييم آثار استهلاك السكروز القصير والطويل الأجل على مورفولوجيا MSN في NAc. سمحنا للفئران المراهقين باستهلاك السكروز بطريقة تشبه الشرايين لـ 4 (قصير المدى) أو 12 (على المدى الطويل) ثم قاموا بتحليل مورفولوجية NAc MSNs من الفئران التي تستهلك السكروز على المدى القصير والطويل وقارنت هذا مع ضوابط العمر المتطابقة الذين تم منحهم حق الوصول إلى المياه فقط. تظهر نتائجنا أن MSNs من غلاف NAc تغيرت بعد استهلاك السكروز الطويل الأمد وليس القصير المدى ، بعد أن قللت من طول التغصنات ، ولكن زيادة كثافة العمود الفقري الشجيري البعيدة. علاوة على ذلك ، وجدنا أن مورفولوجيا MSN من نواة NAc ظلت سليمة نسبيا بعد استهلاك السكروز القصير والطويل الأجل. هذه النتائج تسلط الضوء على نتيجة عصبية مباشرة لاستهلاك السكروز على المدى الطويل بطريقة شبيهة بنهم. وعلاوة على ذلك ، توضح هذه البيانات الحاجة إلى مزيد من الدراسات التي تهدف إلى توضيح التغيرات الجزيئية والكيميائية العصبية التي تصاحب إعادة الهيكلة المورفولوجية ل NAc shell MSN التي يسببها تناول السكروز لفترات طويلة مثل الشراهة.
مواد وطرق
بيان الأخلاق
أجريت جميع الإجراءات التجريبية وفقًا للقانون الأسترالي لرعاية واستخدام الحيوانات للأغراض العلمية ، الإصدار 8th (المجلس الوطني للصحة والبحوث الطبية ، 2013). تمت الموافقة على البروتوكولات من قبل لجنة أخلاقيات الحيوان بجامعة كوينزلاند للتكنولوجيا ولجنة أخلاقيات الحيوان بجامعة كوينزلاند.
الحيوانات والاسكان
خمسة عشر أسبوعًا (المراهق) ذكور فئران Wistar (التحكم: 176.4 ± 4.8 g ؛ Sucrose: 178.3 ± 5.0 g) (ARC ، WA ، أستراليا) ، تم إيواءها بشكل فردي في زجاج شبكي مزدوج التهوية® أقفاص. تأقلم الفئران مع ظروف السكن الفردية ، والمناولة ، ودورة الضوء العكسي 5 قبل بدء التجارب بيومين. تم إيواء جميع الجرذان في دورة 12-hr التي يتم التحكم فيها من خلال المناخ (دورة الإضاءة في غرفة 9 am) التي تحتوي على مناخ قياسي libitum الإعلانية.
اختيار اثنين من زجاجة اختيار المتقطعة الشرب النموذج
الوصول المتقطع 5 ٪ السكروز اختيار اثنين من زجاجة الشرب النموذج (سيمز وآخرون ، 2008) تم اقتباسها من الحكمة (1973). تم تقديم جميع السوائل في عبوات بلاستيكية بلاستيكية 300 ml مع أنابيب الشرب الفولاذ المقاوم للصدأ التي تم إدخالها من خلال حلقتين في الجزء الأمامي من القفص بعد بدء دورة الضوء المظلمة. تم تسجيل وزن كل زجاجة قبل تقديم الزجاجة. تم تقديم زجاجتين في وقت واحد: زجاجة تحتوي على ماء ؛ الزجاجة الثانية التي تحتوي على 5٪ (w / v) سكروز. تم التناوب على موضع زجاجة 5٪ (وزن / وزن) مع كل تعريض للتحكم في التفضيلات الجانبية. تم وزن القنينات 24 h بعد عرض السوائل ، وتم أخذ القياسات إلى أقرب 0.1 g. كما تم قياس وزن كل جرم لحساب جرعات مدخول السكروز لكل كيلوغرام من وزن الجسم. في يوم 1 من فترة الشرب ، الفئران (n = 6 – 9) تم منحهم حق الوصول إلى زجاجة واحدة من 5٪ (w / v) سكروز وزجاجة ماء واحدة. بعد 24 h ، تم استبدال زجاجة السكروز بزجاجة ماء ثانية كانت متوفرة لـ 24 h التالي. تم تكرار هذا النمط في أيام الأربعاء والجمعة. كان لدى الفئران وصول غير محدود للمياه في جميع الأيام الأخرى. أدى الاستهلاك الشبيه بالسكر من السكروز إلى تصاعد في إجمالي كمية السكروز (مل) مع مرور الوقت (الشكل التكميلي 1) وكان مصحوبًا بمستويات ثابتة لمستوى الشرب استنادًا إلى وزن الجسم [20 ± 5 g / kg من 5٪ (w / v)] خلال فترة قصيرة [~ 4 أسابيع (جلسات شرب 13)] وطويلة الأجل [ ~ أسابيع 12 (جلسات شرب 37)] فترات الشرب. مجموعة منفصلة من الفئران السيطرة (n = 6 – 9) تم منحهم حق الوصول إلى الماء في كل من القنينتين (أي بدون سكروز) تحت نفس الظروف المذكورة أعلاه. كان متوسط وزن الجسم للسيطرة والفئران التي تستهلك السكروز في نهاية التعرض قصير الأجل هو 405.7 ± 40.8 g و 426.4 ± 31.2 g على التوالي. في نهاية التعرض الطويل الأجل ، كان متوسط وزن الجسم لمجموعات التحكم والسكر هو 578.8 ± 53.4 g و 600.2 ± 45.2 g.
Golgi-Cox Staining
بعد آخر جلسة شرب تم نقل الفئران من المنشأة الحيوانية للسماح بمعالجة عينات الدماغ في منشأة الأنسجة في كلية العلوم الطبية الحيوية ، جامعة كوينزلاند (سانت لوسيا ، أستراليا). وقد اتخذت جميع التدابير المعتمدة للحد من التوتر أثناء النقل ، وبعد ذلك ، سمح للفئران لاسترداد بين عشية وضحاها. في اليوم التالي ، تمت التضحية بالجرذان من خلال جرعة زائدة من بنتوباربيتال الصوديوم (60 - 80 mg / kg ، ip Vetcare ، بريسبان ، أستراليا) و perfused داخل القلب مع ~ 300 ml السائل الدماغي الشوكي الاصطناعي الذي يحتوي ، (بالسيكل): 130 NaCl ، 3 KCl، 26 NaHCO3، 1.25 نا2PO4، 5 MgCl2، 1 CaCl2و 10 D الجلوكوز. ثم تم قطع رأس كل حيوان وإزالة الدماغ واحتضانه في الظلام في محلول Golgi-Cox الذي يحتوي على 5٪ ثنائي كرومات البوتاسيوم ، 5٪ كرومات البوتاسيوم ، وكلوريد الزئبق 5٪ (جميع المواد الكيميائية من Sigma-Aldrich) التي تم تصنيعها أيام 3 طازجة قبل التضحية كما هو موضح سابقا (Rutledge وآخرون ، 1969). تم تعديل حضانة البقع Golgi-Cox وطرق ما بعد المعالجة من رانجان ومالليك (2010). تم تحضين الأدمغة من الحيوانات المستهلكة على المدى القصير السكروز للأيام 6 في 37 درجة مئوية ، في حين تم تحضين العقول من الحيوانات المستهلكة على المدى الطويل السكروز للأيام 10 ، مع تغيير واحد إلى حل Golgi-Cox الطازجة بعد أيام 4 من الحضانة.
بعد الحضانة ، تم قطع 300 corm أقسام الاكليلية باستخدام مشعاع زايس Hyrax V50 تهتز (كارل زايس ، ألمانيا). ثم تم وضع الشرائح بالتتابع في لوحات 24-well المملوءة بسكروز 30 (w / v) في 0.1 M فوسفات مخزني ملحي ومعالجتها كما هو موضح في (رانجان ومالليك ، 2010). باختصار ، تم تجفيف المقاطع في 50٪ من الإيثانول لمدة دقيقة 5 ، ثم تم وضعها في 0.1 M NH4حل OH لـ 30 min ، وشطفه مرتين بالماء المقطر لـ 5 min ووضعه في Fujie Fujifunt film fixer (Fujifilm ، سنغافورة) لـ 30 min في الظلام. تم شطف الشرائح ثم مرتين في الماء المقطر ل 2 دقيقة لكل والمجففة في 70 ، 90 ، 95 ، و 100 ٪ الايثانول مرتين لكل 5 دقيقة لكل منهما. تم بعد ذلك مسح الأجزاء في محلول CXA (1: 1: 1 chloroform: xylene: alcohol) لـ 10 min والمثبتة في DPX (Sigma-Aldrich) على شرائح Superfrost Plus (Menzel-Glaser، Lomb Scientific، Australia) وتراجع الغلاف (Menzel-Glaser ، ألمانيا). تركت الشرائح في الظلام لتجف في درجة حرارة الغرفة طوال الليل.
اختيار الخلايا العصبية وتتبع داخل النواة المتكئة
تم مسح الشرائح التاجية بين bregma + 2.8 و + 1.7 ل MSNs داخل النواة وقشرة من NAc ، وذلك باستخدام البطين الجانبي والامتداد الأمامي كمعالم بمساعدة أطلس دماغ الفئران (Paxinos و Watson ، 2007) (الشكل 1). تم استخدام وظيفة الكنتور في Neurolucida 7 (MBF Bioscience، VT، USA) لترسيم نواة NAc و NAc shell في كل شريحة (الشكل 2). بين الخلايا العصبية 2 و 9 لكل منطقة لكل حيوان تم تتبعها لمعلمات طول شجيري باستخدام هدف 63x أو لكثافات العمود الفقري (تم الإبلاغ عنها كعمود فقري لكل 100 μm) باستخدام هدف 100x في Zeiss Axioskop II (Carl Zeiss، Germany) باستخدام آلية XYZ المرحلة التي يقودها Neurolucida® برنامج 7 (MBF Biosciences، VT، USA). تم تنفيذ جميع تتبع بطريقة أعمى فيما يتعلق بالعلاج. تم تحليل المعلمات المورفولوجية من الخلايا العصبية المشبعة Golgi-Cox بطريقة مشابهة للتقارير السابقة (Klenowski وآخرون ، 2015).
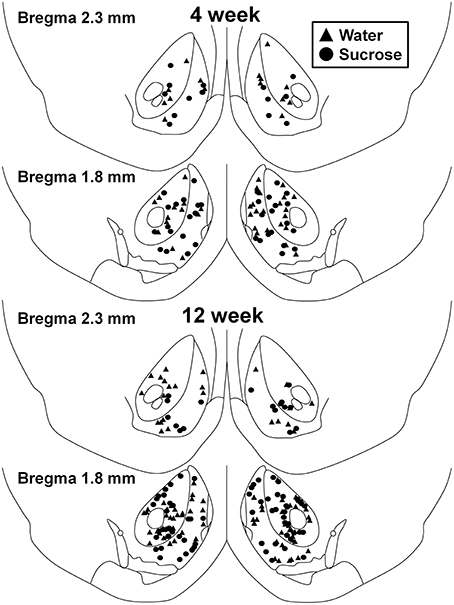
الشكل 1. خريطة توضح مواقع الخلايا العصبية الشوكية المتوسطة التي تم أخذ عينات منها من النواة المتكئة النواة وقذيفة لجرذان 4 و 12 الأسبوعي واستجابة الفئران. تظهر أعلى مجموعتين من مواقع الخلايا العصبية التي تم أخذ عينات منها من النواة المتكئة النواة وقشرة من التحكم في أسبوع 4 (مثلثات) وحيوانات (السكروز). توضح اللوحتان السفليتان مواقع الخلايا العصبية التي تم أخذ عينات منها من التحكم في أسبوع 12 (المثلثات) وحيوانات السكروز (الدوائر).
التحليل الإحصائي
تم حساب متوسط الخطأ المعياري للمتوسط (SEM) لكل مجموعة بيانات مع الحيوان كـ nباستخدام بيانات قياس المورثومتري من كل نواة أو قشرة NAc MSNs (n = 7 لقشرة NAc و n = 6 لـ NNc core 4-week ، n = 9 لمجموعات 12 أسبوعًا). حيث هو مبين ، غير مزاوج اثنين من الطالب الذيل tأجريت اختبارات أو ANOVAs ثنائية الاتجاه مع اختبارات ما بعد Bonferroni لجميع التحليلات التي تنطوي على مقارنة وسائل المجموعة ، وذلك باستخدام الإصدار GraphPad Prism 6.02 (GraphPad Software، San Diego، CA). تم قبول دلالة إحصائية في P <0.05. يتم تقديم جميع البيانات في قسم النتائج كوسائل ± SEM. يتم حساب التغييرات في النسبة المئوية بالنسبة لقيمة التحكم.
النتائج
الخلايا العصبية الشوكية المتوسطة من نواة متكئة Nucleus انخفضت طولها شجيري ، نقص في تعقيد الشجيرات ولكن زيادة متوسط كثافة العمود الفقري عند أوامر فرعية بعيدة بعد استهلاك السكروز طويل الأمد وليس قصير المدى
بعد الاستهلاك قصير الأجل (أسابيع 4) السكروز ، لم تكن هناك فروق ذات دلالة في المعلمات morphometric قذيفة NAc MSN (الجدول 1). لم تكن هناك أيضا فروق ذات دلالة إحصائية بين الاستهلاك على المدى القصير السكروز والتحكم في المياه NAc قذيفة MSN في التحليلات المتعلقة بترتيب الطرد المركزي. وهي شرائح شجيري لكل طلب فرعي (P = 0.4111) ، يعني طول شجيري لكل طلب فرعي (P = 0.5581) وتعني كثافة العمود الفقري لكل طلب فرعي (P = 0.2977 ، ANOVAs ذات الاتجاهين) لا تختلف بشكل كبير بين المجموعات. يظهر في الرسم البياني خريطة الموقع تبين المواقف التقريبية للعصبونات العينة 1.
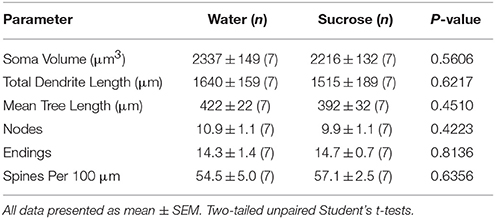
جدول 1. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة من قوقعة الجرذان قصيرة المدى التي تستهلك السكروز والضوابط المائية المتطابقة مع العمر.
بعد فترة طويلة (أسابيع 12) من استهلاك السكروز ، انخفض إجمالي طول الشجرة المتغصنة من NAc shell MSN بواسطة 21٪ مقارنة بالضوابط التي تستهلك المياه (الماء: 1827 ± 148 μm ، n = 9 Sucrose 1449 ± 78 μm، n = 9 ، *P = 0.0384 ، الطالب ثنائي الذيل غير المزاوج tالاختبار ، الشكل 2، الطاولة 2). أظهرت مقارنة متوسط عدد التشعبات (العقد) ونهايات التغصنات بين الماء ومجموعات السكروز مستوى منخفض (وإن لم يكن معنويا) من التعقيد التغصني في NAc shell MSN (العقد: Water 12.9 ± 1.4 n = 9 ، Sucrose 10.1 ± 0.8 n = 9، P = 0.0879 النهايات: المياه 17.9 ± 1.4 n = 9 ، Sucrose 14.8 ± 0.7 n = 9، P = 0.0657 ، الطالب ثنائي الذيل غير المزاوج tالاختبار ، الجدول 2). لم يكن هناك تغيير في حجم سوما (P = 0.9400) ، يعني طول الشجرة المتغصنة (P = 0.1646) أو كثافة العمود الفقري الكلية (P = 0.3662) في NAc shell MSNs من الفئران المستهلكة على المدى الطويل السكروز مقارنة بمراقبة المياه. هذه المعلمات مورفومترية مفصلة في الجدول 2.
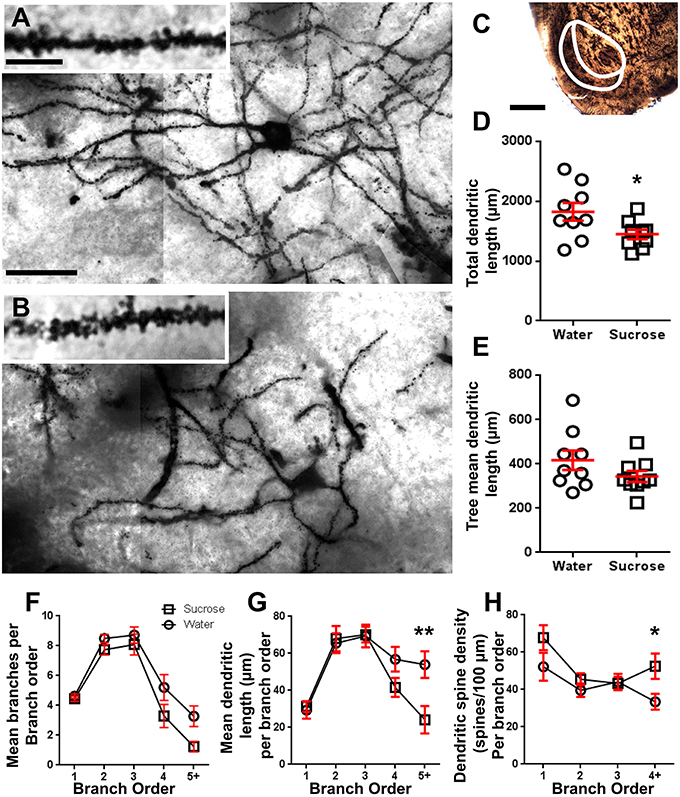 الشكل 2. انخفض طول الشجرة الشجيري وزيادة كثافة العمود الفقري الشجيري البعيدة من الخلايا العصبية الشوكية المتوسطة (ام اس اس) من قذيفة النواة المتكئة (NAc) من الفئران المعالجة على المدى الطويل السكروز مقارنة مع الجرذان السيطرة. (A، B) عرض تمثيلات السيطرة (أعلى) وطويل الأجل (12 الأسبوع) السكروز (القاع) تعامل brightfield zالفسيفساء من الفئران من مشربة Golgi من NAc قذيفة (تكبير 63x). أقحم من (A، B) يظهر السيطرة والسكر على المدى الطويل تعامل الصور brightfield من التشبع الشفاف غولجي MSN وشريط العمود الفقري من قذيفة NAc (التكبير 100x). (C) يبين المناطق التشريحية التي تم أخذ عينات من MSN في هذه الدراسة. (د) يظهر مؤامرة مبعثرة من مجموع الشجرة المتشابكة الكلي MSN (يعني ± SEM) من قذيفة NAc في حيوانات السكروز على المدى الطويل (المربعات) مقارنة مع الضوابط (الدوائر) ، الطلاب غير المتزوجين t-اختبار، *P <0.05 ، n = 9 السيطرة و n = 9 12 أسبوع السكروز. (E) يظهر مؤشرا مبعثرًا لمعدل الشجرة المتشابكة في MSN غير المتغير (متوسط ± SEM) من غلاف NAc في حيوانات السكروز على المدى الطويل (المربعات) مقارنة بالضوابط (الدوائر) ، الطلاب غير الملتزمين t-اختبار، P > 0.05 ، n = 9 السيطرة و n = 9 12 أسبوع السكروز. تحليل ترتيب الفروع (يعني ± SEM) من رقم جزء شجيري لكل طلب فرعي (F)يعني طول شجيري لكل طلب فرعي (G) وكثافة العمود الفقري الشجيري لكل طلب فرعي (H). انخفاض استهلاك السكروز على المدى الطويل طول شجيري في أوامر فرع البعيدة (5 +) وزيادة كثافة العمود الفقري شجيري في أوامر فرع البعيدة (4 +) مقارنة مع الضوابط (G، H)، و ANOVAs ثنائية الاتجاه مع اختبارات ما بعد Bonferroni ، *P <0.05 ، **P <0.01 ، n = 9 السيطرة و n = 9 السكروز على المدى الطويل. قضبان مقياس: (أ ، ب) = 20 μm؛ أقحم من (أ ، ب) = 10 μm؛ (C) = 1 ملم.
الشكل 2. انخفض طول الشجرة الشجيري وزيادة كثافة العمود الفقري الشجيري البعيدة من الخلايا العصبية الشوكية المتوسطة (ام اس اس) من قذيفة النواة المتكئة (NAc) من الفئران المعالجة على المدى الطويل السكروز مقارنة مع الجرذان السيطرة. (A، B) عرض تمثيلات السيطرة (أعلى) وطويل الأجل (12 الأسبوع) السكروز (القاع) تعامل brightfield zالفسيفساء من الفئران من مشربة Golgi من NAc قذيفة (تكبير 63x). أقحم من (A، B) يظهر السيطرة والسكر على المدى الطويل تعامل الصور brightfield من التشبع الشفاف غولجي MSN وشريط العمود الفقري من قذيفة NAc (التكبير 100x). (C) يبين المناطق التشريحية التي تم أخذ عينات من MSN في هذه الدراسة. (د) يظهر مؤامرة مبعثرة من مجموع الشجرة المتشابكة الكلي MSN (يعني ± SEM) من قذيفة NAc في حيوانات السكروز على المدى الطويل (المربعات) مقارنة مع الضوابط (الدوائر) ، الطلاب غير المتزوجين t-اختبار، *P <0.05 ، n = 9 السيطرة و n = 9 12 أسبوع السكروز. (E) يظهر مؤشرا مبعثرًا لمعدل الشجرة المتشابكة في MSN غير المتغير (متوسط ± SEM) من غلاف NAc في حيوانات السكروز على المدى الطويل (المربعات) مقارنة بالضوابط (الدوائر) ، الطلاب غير الملتزمين t-اختبار، P > 0.05 ، n = 9 السيطرة و n = 9 12 أسبوع السكروز. تحليل ترتيب الفروع (يعني ± SEM) من رقم جزء شجيري لكل طلب فرعي (F)يعني طول شجيري لكل طلب فرعي (G) وكثافة العمود الفقري الشجيري لكل طلب فرعي (H). انخفاض استهلاك السكروز على المدى الطويل طول شجيري في أوامر فرع البعيدة (5 +) وزيادة كثافة العمود الفقري شجيري في أوامر فرع البعيدة (4 +) مقارنة مع الضوابط (G، H)، و ANOVAs ثنائية الاتجاه مع اختبارات ما بعد Bonferroni ، *P <0.05 ، **P <0.01 ، n = 9 السيطرة و n = 9 السكروز على المدى الطويل. قضبان مقياس: (أ ، ب) = 20 μm؛ أقحم من (أ ، ب) = 10 μm؛ (C) = 1 ملم.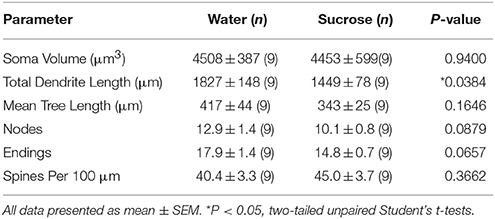
جدول 2. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة القشرة من الفئران المستهلكة لسكر السكر على المدى الطويل والضوابط المائية المطابقة للعمر.
بعد توصيف مورفولوجيا شجيرية عامة من استهلاك السكروز على المدى الطويل NAc shell MSN ، قمنا بتحليل التشعبات الشجرية وكثافات العمود الفقري فيما يتعلق بخصائص ترتيب الفروع. تقييمنا الشامل للأشجار التغصنية كميا عدد الشرائح التغصنية لكل طلب فرعي ، متوسط طول المقاطع التغصنية لكل طلب فرعي و متوسط كثافة العمود الفقري لكل طلب فرعي من NAc shell MSN من التحكم في المياه و الفئران التي تستهلك السكروز على المدى الطويل. يتم تقديم ملخص لبيانات وأوامر الفروع في الجدول 3.
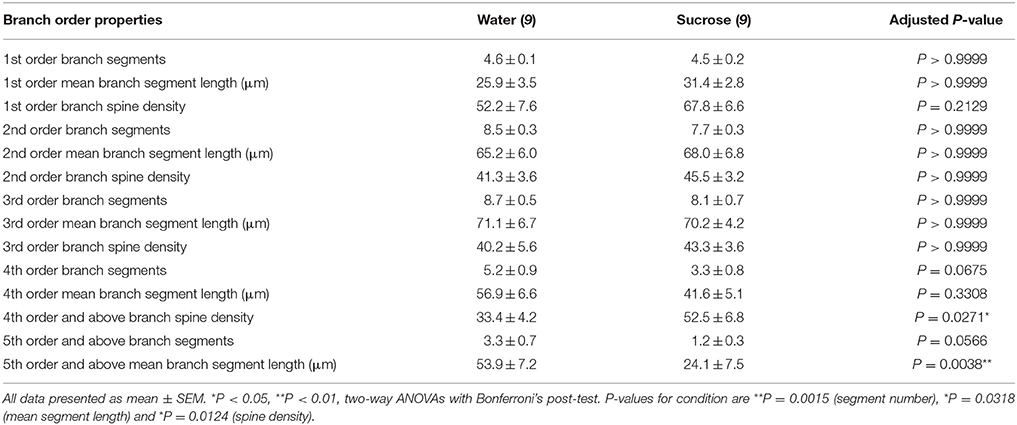 جدول 3. خصائص رتبة فرع من الخلايا العصبية الشوكية المتوسطة من السكروز على المدى الطويل وفئران مياه الشرب.
جدول 3. خصائص رتبة فرع من الخلايا العصبية الشوكية المتوسطة من السكروز على المدى الطويل وفئران مياه الشرب.تم تخفيض عدد أجزاء الفرع المتفرعة المتوسطة لكل طلب فرعي من NAc shell MSNs بشكل كبير في الجرذان المستهلكة لسكر السكر على المدى الطويل مقارنةً بالضوابط المائية (**P = 0.0015 ، ANOVA ذات الاتجاهين). كشفت اختبارات ما بعد Bonferroni عن وجود اتجاه نحو انخفاض عدد قطع الفروع في 4th (الماء: 5.2 ± 0.9 ، n = 9 Sucrose 3.3 ± 0.8 ، n = 9، P = 0.0675 ، الشكل 2F، الطاولة 3) وطلبات 5th وأعلى الفروع (الماء: 3.3 ± 0.7 ، n = 9 Sucrose 1.2 ± 0.3 ، n = 9، P = 0.0566 ، الشكل 2F، الطاولة 3). تم تقليل طول المقطع الشجيري المتوسط لكل طلب فرعي من NAc shell MSN أيضًا بشكل كبير في الجرذان المستهلكة لسكر السكر على المدى الطويل مقارنةً بالضوابط المائية (*P = 0.0444 ، ANOVA ذات الاتجاهين). أظهرت اختبارات ما بعد بونفيروني انخفاضًا في نسبة 55٪ في فروع ترتيب 5th وما بعدها (الماء: 53.9 ± 7.2 μm ، n = 9 Sucrose 24.1 ± 7.5 μm، n = 9 ، **P = 0.0038 ، الشكل 2G، الطاولة 3).
أظهر تحليل ترتيب الفروع زيادة كبيرة في كثافة العمود الفقري الشجيري ل NAc shell MSN من الفئران المستهلكة على السكروز على المدى الطويل مقارنة بالضوابط (*P = 0.0124 ، ANOVA ذات الاتجاهين). أظهرت اختبارات ما بعد Bonferroni زيادة كثافة العمود الفقري لـ 57٪ في فروع النظام 4th البعيدة وما بعدها (Water: 33.4 ± 4.2 ، n = 9 Sucrose 52.5 ± 6.8 ، n = 9، P = 0.0271 *، inset of Figures 2A، B، H، الطاولة 3). صور تمثيلية لهندسة MSN العامة وكثافة العمود الفقري البعيدة (الشكل الداخلي) موضحة في شكل أرقام 2A، B.
مجتمعة ، تشير هذه النتائج إلى أن استهلاك السكروز قصير المدى له تأثير ضئيل على المعلمات المورفولوجية لـ MSNs داخل غلاف NAc. ومع ذلك ، بعد الاستهلاك لفترة طويلة ، هناك انخفاض كبير في طول الشجرة العصبية والتعقيد ، ولا سيما في فروع شجيري البعيدة. زيادة كثافة العمود الفقري البعيدة المتزامنة تظهر أيضًا في NAc shell MSN من الفئران التي تستهلك السكروز على المدى الطويل.
الخلايا العصبية المتوسطة الشوكية من النواة الأساسية Accumbens Core لها تقلل في تعقيد الفروع بعد استهلاك السكروز الطويل الأمد وليس القصير المدة
بعد فترة قصيرة من استهلاك السكروز ، لم تكن هناك فروق ذات دلالة إحصائية في المعلمات morphometric جوهر NAc (الجدول 4). لم يكن هناك أيضا فروق ذات دلالة إحصائية بين استهلاك السكروز في الأسبوع 4 والنواة MSS التحكم في المياه في التحليلات المتعلقة بترتيب الطرد المركزي. وهي شرائح شجيري لكل طلب فرعي (P = 0.7717) ، يعني طول شجيري لكل طلب فرعي (P = 0.2096) ، ويعني كثافة العمود الفقري لكل طلب فرعي (P = 0.3521 ، ANOVAs ذات الاتجاهين) لم تكن مختلفة بين المجموعات.
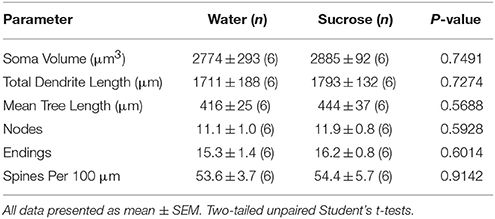 جدول 4. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة الأساسية للجرذان قصيرة الأجل التي تستهلك السكروز وعمر التحكم في الماء.
جدول 4. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة الأساسية للجرذان قصيرة الأجل التي تستهلك السكروز وعمر التحكم في الماء.كما أن استهلاك السكروز لفترات طويلة لم يكن له دلالة على المعلمات المورفومترية الأساسية في NAc MSN (الجدول 5). تم تخفيض عدد أجزاء الفرع المتشابه المتوسط لكل طلب فرعي من NAc core MSs بشكل كبير في الجرذان المستهلكة لسكر السكر على المدى الطويل مقارنةً بالضوابط المائية (*P = 0.0416 ، ANOVA ذات الاتجاهين) ، ولكن لم تكن هناك فروق ذات دلالة إحصائية في متوسط طول الشجيري لكل طلب فرعي (P = 0.0995) وتعني كثافة العمود الفقري لكل طلب فرعي (P = 0.4888 ، ANOVAs ذات الاتجاهين) بين MSN في قلب NAc من الفئران المستهلكة على المدى الطويل السكروز مقارنة مع ضوابط المياه. وإذا أخذنا معاً ، فإن بياناتنا تظهر أن النواة NAc ليست مستجيبة لاستهلاك السكروز على المدى الطويل مقارنةً بـ MSNs من منطقة NAc shell.
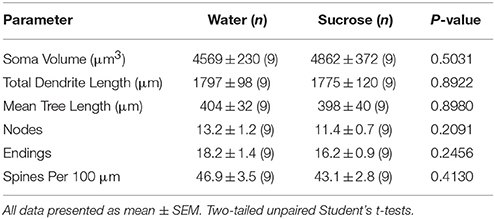 جدول 5. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة الأساسية للجرذان المستهلكة لسكر السكر على المدى الطويل وعمر التحكم في الماء المطابق.
جدول 5. المعلمات المورفولوجية العامة للخلايا العصبية الشوكية المتوسطة من النواة المتكئة الأساسية للجرذان المستهلكة لسكر السكر على المدى الطويل وعمر التحكم في الماء المطابق.مناقشة
زيادة توافر الغذاء المحلى للغاية في النظام الغذائي الغربي لم يساهم فقط في زيادة انتشار العبء الاقتصادي للسمنة والنوع الثاني من مرض السكري ، فقد أدى أيضًا إلى ظهور اضطرابات في الأكل مثل الإفراط في الأكل (سوانسون وآخرون ، 2011; كيسلر وآخرون ، 2013; ديفيس ، 2015). على الرغم من أن الخواص الإدمانية للسكريات بما في ذلك الفركتوز والسكروز تبقى مضاربة ، إلا أن هناك تشابهًا لافتًا في الارتباطات السلوكية والعصبية التي تظهر نتيجة لتناول الطعام على المدى الطويل واستخدامه لفترات طويلة. (Avena et al.، 2008, 2011). بالإضافة إلى ذلك ، يعمل السكر على تنشيط دائرة المكافأة في الدماغ بطريقة تشبه تعاطي المخدرات (Volkow وآخرون ، 2012وتشير النتائج من الدراسات البشرية إلى أن السكر والحلاوة يمكن أن تحفز الرغبة الشديدة التي يمكن مقارنتها بالحجم إلى تلك الناجمة عن المخدرات الإدمانية مثل الكحول والنيكوتين (Volkow وآخرون ، 2012). لذلك ، استخدمنا نموذجًا لاستهلاك الشراهة عند الجرذان لتحديد آثار قصير المدى (أسابيع 4) واستهلاك السكروز طويل الأمد (أسابيع 12) على مورفولوجيا الخلايا العصبية (MSN) في NAc ، وهو مكون أساسي في دوائر المكافآت المتداخلة التي يتم تعديلها بواسطة السكر والأدوية. نظهر أن MSNs من NAc shell من الفئران المستهلكة على السكر على المدى الطويل المزمن قد انخفضت بشكل كبير طول التشعبية والتعقيد ، ولكن زيادة كثافة العمود الفقري الشجيري البعيدة. لم يؤثر استهلاك السكروز على المدى الطويل على مورفولوجيا MSN من نواة NAc ، في حين أن استهلاك السكروز قصير المدى لم يكن له تأثير كبير على مورفولوجيا MSN من نواة NAc أو shell. هذه النتائج لا تظهر فقط تأثير مباشر من تناول السكروز لفترات طويلة مثل الشوك على مورفولوجيا الخلايا العصبية من NAc قذيفة MSN ، لكنها أيضا تسليط الضوء على الآثار الضارة المحتملة للاستهلاك لفترات طويلة من الوجبات الغذائية التي تحتوي على السكر العالي.
يتكون الـ NAc ، الذي يشكل جزءًا من المخطط البطني ، بشكل أساسي من الـ MSN ، والتي تتميز بشكل ملموس بأنها خلايا عصبية متوسطة الحجم ذات شجيرات شجيرية مكثفة وكثافة عالية في العمود الفقري (كيمب وباول ، 1971; Graveland and DiFiglia، 1985; Rafols وآخرون ، 1989; كاواجوتشي وآخرون ، 1990). الخلايا العصبية الغلوتاماتية والدوبامينية هي المدخلين الأساسيين للواقي إلى NAc ، وتتصل في المقام الأول بالأعمدة الشجيرية وأعمدة العمود الفقري لـ MSN. (غروفز ، 1980; Kaiya و Namba ، 1981; Groves et al.، 1994). على وجه التحديد ، تتلقى قشرة NAc ولب القلب المدخلات الجلوتامتية من مناطق قشرية مميزة وظيفيا (Brog et al.، 1993). قذيفة NAc معصوبة أيضاً بواجبات استثارية من المناطق تحت القشرية مثل الحصين ، المهاد والدماغ basolateral (Brog et al.، 1993; Wright و Groenewegen ، 1995). وقد أثبتت الدراسات السابقة أن هذه المدخلات glutamatergic تلعب دورا محوريا في التحفيز والسلوكيات الموجهة نحو الهدف مثل الغذاء والمثابرة مكافأة (مالدونادو-ايريزاري وآخرون ، 1995; كيلي وسوانسون ، 1997; رينولدز وبيريدج ، 2003; ريتشارد وبيريدج ، 2011). المدخلات الأخرى المهيمنة على NAc MSNs هي من مستخلصات الدوبامين التي من المشروع tegmental المنطقة البطنية (Lindvall و Björklund ، 1978; Veening وآخرون ، 1980; كاليفاس وميلر ، 1984). ومن المثير للاهتمام أن الدراسات السابقة التي استخدمت نماذج مماثلة من الوصول المتقطع للسكر أظهرت أن الاستهلاك الشبيه بالناتج الناتج ينتج عنه زيادة في الدوبامين خارج الخلية في NAc بالمثل (وإن كان بدرجة أقل) إلى تعاطي المخدرات (رادا وآخرون ، 2005; Avena et al.، 2006) ، ويمكن تعديل تعبير مستقبلات الدوبامين (Colantuoni وآخرون ، 2001, 2002) في جوهر NAc وقذيفة. ومن المثير للاهتمام ، أن استهلاك السكروز الشبيه بالإفرازات يؤدي إلى تصاعد في المدخول مع مرور الوقت على نحو مماثل للإدارة الذاتية لعقاقير الإساءة مثل الكوكايين والبطلينن (أحمد وكوب ، 1998; أحمد وآخرون ، 2000, 2003) التي ترتبط بتطور حالة "مثل الإدمان".
يوضح تحليلنا لقياسات قياس فرعي ترتيب الفروع أن الانخفاض العام في طول شجيري من NAc shell MSN الذي يسببه تناول السكروز على المدى الطويل ، ينتج في المقام الأول عن تخفيضات في تعقيد أوامر التوصيل البعيدة. لاحظنا انخفاض التفرع البعيدة (4th و 5th ترتيب وأوامر فرع أعلى) وخفض متوسط الطول بشكل كبير في ترتيب 5th وفوق التشعبات ، جنبا إلى جنب مع زيادة كثافة العمود الفقري في هذه الأوامر الفرعية. هناك عامل مشترك من المرجح أن يؤثر على هذا النوع من إعادة الهيكلة التغصنية يتضمن تغييرات في الاتصال و / أو وظيفة متشابك (روسو وآخرون ، 2010). وقد أظهرت الدراسات السابقة أن التشابهات الجلوتاماتية على MSN تتشكل في المقام الأول على العمود الفقري ، ولا سيما في التشعبات البعيدة (Groenewegen وآخرون ، 1999). بالإضافة إلى ذلك ، شارك في توطين الدوبامين ومدخلات الجلوتامين من القشرة الفص الجبهي (Sesack و Pickel ، 1992), الحصين (Totterdell و Smith ، 1989; Sesack و Pickel ، 1990), واللوزة (Johnson et al.، 1994) وقد لوحظت في العمود الفقري شجيري من MSN. هذه الملاحظات جنبا إلى جنب مع زيادة كثافة العمود الفقري بعد الاستهلاك على المدى الطويل السكروز المشاهدة في دراستنا ، ودعم تشكيل مدخلات مثيرات مثيرة. ولذلك ، فإن الاحتمال ينشأ حيث يمكن أن تسهم التأثيرات المستمرة الناجمة عن تناول السكروز لفترات طويلة مثل زيادة نشاط المشابك الاستثارة في التشعبات البعيدة من MSN في غلاف NAc. وبالتالي ، قد ينتج عن تخفيض و / أو سحب التشعبات البعيدة عن طريق آلية استتباب متزامنة (Reissner و Kalivas ، 2010) ، ولكن هذا ما زال يتعين تحديده.
من المثير للاهتمام أن نلاحظ أن Crombag وزملاؤه أظهروا أنه لم تكن هناك زيادة في كثافة العمود الفقري في قشرة NAc بعد استهلاك السكروز في الأسبوع 4 عبر نموذج الإدارة الذاتية للكتف nose-poke على الرغم من اكتساب أكثر قوة ومعدل استجابة أعلى للسكروز عند المقارنة مع الأمفيتامين (Crombag et al.، 2005). تعكس ملاحظةهم لغياب التغيير في كثافة العمود الفقري في أسابيع 4 نتائجنا. على النقيض من ذلك ، توضح دراستنا أنه بعد التعرض الطويل الأمد (الأسبوع 12) لاستهلاك السكروز المزمن ، هناك زيادة كبيرة في كثافة العمود الفقري القاصي على الـ MSNs لجرذان تجارب السكروز. علاوة على ذلك ، أظهر مختبرنا في السابق أن استهلاك السكروز على المدى الطويل (12 week) يسهل الاستجابة الدوائية التفاضلية للعلاجات الدوائية التي ثبت أنها تعدل استجابات الدوبامين والأسيتيل كولين عند مستوى NAc (Shariff وآخرون ، في الصحافة). يؤخذ هذا معا ، وهذا يشير إلى أن السكروز (12 أسابيع وما بعدها) على المدى الطويل التعرض ، وهو أكثر دقة تعكس سيناريوهات العالم الحقيقي ، وينتج في التكيف المورفولوجية على مستوى NAC.
من حيث تعاطي المخدرات ، والتعرض المتكرر لأدوية مختلفة ينتج تغييرات طويلة الأمد في بنية التشعبات والأشواك التغصنية. على سبيل المثال ، تزيد الأمفيتامينات والكوكايين من كثافة العمود الفقري في NAc في كل من القشرة والجوهر (روبنسون و Kolb ، 2004). كما تبين أن تعرض النيكوتين يزيد من كثافة العمود الفقري في غلاف NAc. على العكس ، يؤدي تعرض المورفين إلى انخفاض في كثافة العمود الفقري وتعقيد فرع التغصنات (روبنسون و Kolb ، 2004). من حيث استهلاك السكروز على المدى الطويل ، لاحظنا زيادة في كثافة العمود الفقري تشبه الأمفيتامين والكوكايين والنيكوتين وعكس تأثير المورفين. ومع ذلك ، على عكس الأمفيتامين والكوكايين ، ولكن مماثلة للنيكوتين ، فإن زيادة كثافة العمود الفقري على المدى الطويل التعرض للسكروز يقتصر على قذيفة NAc. ومن المثير للاهتمام أيضا أن التغييرات في كل من المتفرعة dendritic (روبنسون و Kolb ، 1999) وكثافة العمود الفقري (لي وآخرون ، 2003) التي تنتجها الأمفيتامين أو الكوكايين تقتصر على dendrites البعيدة من MSN في NAc ، مما يعكس النتائج في دراستنا. علاوة على ذلك ، وللتأكيد على التغييرات الموضحة أعلاه ، فإن استهلاك السكروز أظهر في السابق أنه يعزز قوة التشابك الاستثارية على الخلايا العصبية الدوبامين التراكمية (Stuber et al.، 2008b) فضلا عن المكونات الأخرى لمسار المكافأة المتوسطة (Stuber et al.، 2008a; تشن وآخرون ، 2010). وإذا ما أخذنا معا في الاعتبار ، فإن هذا يفترض أن السكروز هو أداة قوية لتشكيل الخلايا العصبية بعد الاستخدام الكثيف لفترات طويلة ، وهو ما يماثل التأثيرات التي لوحظت من تعاطي المخدرات.
على الرغم من أن هناك حاجة إلى مزيد من التحقيقات للكشف عن الآليات الخلوية والمتشابكة التي تسهم في التغيرات المورفولوجية التي تظهر في هذه الدراسة ، فإن نتائجنا تظهر آثارًا عصبية كبيرة ناتجة عن استهلاك السكروز على المدى الطويل. على وجه الخصوص ، هناك دراسة لم يتم دراستها في دراستنا هي ما إذا كانت الآثار المورفولوجية المرصودة للسكروز يمكن أيضًا الحصول عليها من المحليات غير السعرية مثل السكرين. في هذا الصدد ، من المهم أن نلاحظ أن لينوار وزملائه قد أظهروا أن الحلاوة الشديدة تفوق المكافأة الكوكايينية ، سواء أكانت من السكرين أو السكروز (Lenoir et al.، 2007). علاوة على ذلك ، دراسة حديثة نشرها مختبرنا (Shariff وآخرون ، في الصحافة) يوضح أن الفارينكلين ، وهو ناهض أستيل النيكوتين الجزئي مستقبلات خفضت كل من تناول السكروز والسكرين في القوارض اتباع نفس نظام الوصول المتقطع على المدى الطويل المستخدمة في هذه الدراسة. ومن المثير للاهتمام ، أظهرت الدراسات السابقة أوجه تشابه في الآثار الحادة للمُحليات غير الحرارية مثل السكرين والسكروز على مستوى NAc (Scheggi وآخرون ، 2013; Tukey وآخرون ، 2013; Carelli and West، 2014). ومع ذلك ، هناك حاجة إلى مزيد من الدراسات لتحديد ما إذا كانت المحليات غير السعرية يمكن أن تحفز تأثيرات طويلة المدى مشابهة للتغيرات في مورفولوجيا NAc shell MSN الناتجة عن استهلاك السكروز على المدى الطويل المذكور هنا.
إن عدم التأثير على مورفولوجيا NAc MSN بعد استهلاك السكروز قصير الأجل ، يسلط الضوء على أهمية تنفيذ دراسات طويلة الأمد لتقييم تأثير تعاطي المخدرات لفترة طويلة أو المكافآت الطبيعية مثل السكروز. من حيث الاعتماد ، ليس فقط دورات متكررة من تناول الشراهة والامتناع عن المكونات الرئيسية لدورة الإدمان ، كشفت مجموعة متزايدة من الأدلة أن الانتقال إلى الاعتماد هو عملية تقدمية غالباً ما تحدث على مدى فترة طويلة من الزمن. على الرغم من أن الخصائص الإدمانية للسكريات لا تزال غير مؤكدة ، إلا أن قابلية الإدمان على المكافآت الأخرى غير الدوائية مثل الجنس ، والمقامرة ، والغذاء يتم التحقيق فيها بشكل متزايد. النتائج من هذه الدراسة تضيف ميزة إلى فرضية أن السكريات مثل السكروز قد يكون لها خصائص إدمانية بعد الاستهلاك الطويل الأجل الشبيه بالأنفجار. نتائجنا لها أيضا آثار بالنسبة للعدد المتزايد من الأطفال والمراهقين الذين يحافظون على عادات الأكل غير الصحية (ارتفاع استهلاك السكر والشراهة في تناول الطعام) إلى مرحلة البلوغ. تماشيا مع زيادة خطر حدوث تأثيرات التمثيل الغذائي فمن الممكن أيضا أن النتائج العصبية والنفسية التي تؤثر على المزاج والتحفيز قد تنتج أيضا عن هذه السلوكيات.
الكاتب الاشتراكات
شارك في تصميم البحوث: PK ، SB. تجارب أجريت: PK، MS، AB، MF، EM. تحليل البيانات: PK ، MF ، MS. تفسر البيانات وساهمت في كتابة المخطوطة: PK ، MS ، MF ، EM ، MB ، SB. قرأ جميع المؤلفين واعتمدوا المخطوطة النهائية لتقديمها.
بيان تضارب المصالح
يعلن المؤلفون أن البحث أجري في غياب أي علاقات تجارية أو مالية يمكن تفسيرها على أنها تضارب مصالح محتمل.
أعلن المراجعون SC و SA ومعالجات التعامل معهم انتمائهم المشترك ، ويذكر محرر المعالجات أن العملية قد استوفت معايير المراجعة العادلة والموضوعية.
شكر وتقدير
تم دعم هذا العمل بتمويل من المنح المقدمة من مجلس البحوث الأسترالي (FT1110884) إلى SB والمجلس الوطني للصحة والبحوث الطبية (1061979) إلى SB و MB.
المواد التكميلية
يمكن العثور على المواد التكميلية لهذه المادة على الإنترنت على الموقع: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
الشكل التكميلي 1. تناول السكروز والأفضلية من 4 و 12 أسبوع السكروز يستهلكان الفئران. (A، B) إظهار التصعيد في إجمالي كمية السكروز (مل) خلال الأسابيع 4 و 12 من التعرض. (C، D) إظهار تفضيل عالي للسكروز فوق الماء أثناء فترات عرض السكروز.
مراجع حسابات
Ahmed، SH، and Koob، GF (1998). الانتقال من المعتدلة إلى الإفراط في تناول الدواء: التغيير في نقطة مجموعة hedonic. علوم 282 و 298 – 300. doi: 10.1126 / science.282.5387.298
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ahmed، SH، Lin، D.، Koob، GF، and Parsons، LH (2003). لا يعتمد تصعيد الكوكائين في تعاطي الكوكايين على النواة المتغيرة للكوكايين المتغيرة والمستويات الدوبامين. J. Neurochem. 86 و 102 – 113. doi: 10.1046 / j.1471-4159.2003.01833.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ahmed، SH، Walker، JR، and Koob، GF (2000). زيادة مستمرة في الدافع لاتخاذ الهيروين في الفئران مع تاريخ من تصعيد المخدرات. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Arslanian، S. (2002). اكتب مرض السكري 2 في الأطفال: الجوانب السريرية وعوامل الخطر. Horm Res 57 (ملحق 1) ، 19 – 28. دوى: 10.1159 / 000053308
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Avena، NM، Bocarsly، ME، Hoebel، BG، and Gold، MS (2011). التداخلات في علم تصنيف الأدوية من تعاطي المخدرات والإفراط في تناول الطعام: الآثار المترجمة "لإدمان الغذاء". داء. إساءة استخدام المخدرات 4 و 133 – 139. دوى: 10.2174 / 1874473711104030133
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Avena، NM، Rada، P.، and Hoebel، BG (2008). أدلة لإدمان السكر: التأثيرات السلوكية والعصبية الكيميائية الناجمة عن تناول السكر المفرط والمتقطع. Neurosci. Biobehav. القس 32 و 20 – 39. doi: 10.1016 / j.neubiorev.2007.04.019
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Avena، NM، Rada، P.، Moise، N.، and Hoebel، BG (2006). السكروز تغذية الشام على جدول الإفراج عن النشرات المتكثفة الدوبامين مرارا وتخلص من استجابة الشبع أستيل. علم الأعصاب 139 و 813 – 820. doi: 10.1016 / j.neuroscience.2005.12.037
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
بينتون ، د. (2010). معقولية إدمان السكر ودوره في السمنة واضطرابات الأكل. كلين. نوتر. 29 و 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Bray و GA و Popkin، BM (2014). السكر الغذائي ووزن الجسم: هل وصلنا إلى أزمة في وباء السمنة ومرض السكري؟ صب على السكر. رعاية مرضى السكري 37 و 950 – 956. doi: 10.2337 / dc13-2085
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Brog، JS، Salyapongse، A.، Deutch، AY، and Zahm، DS (1993). أنماط من تعصيب وارد من جوهر وقذيفة في جزء "المتكئين" من المخطط البطني الفئران: كشف المناعى الكيميائي المنقولة عن طريق نقل فلورو الذهب. J. شركات. Neurol. 338 و 255 – 278. doi: 10.1002 / cne.903380209
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Bucher Della Torre، S.، Keller، A.، Laure Depeyre، J.، and Kruseman، M. (2015). المشروبات المحلاة بالسكر ومخاطر السمنة لدى الأطفال والمراهقين: تحليل منهجي حول كيفية تأثير الجودة المنهجية على النتائج. ج. أكاد. نوتر. حمية. [النشر الإلكتروني قبل الطباعة]. doi: 10.1016 / j.jand.2015.05.020
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Carelli، RM، and West، EA (2014). عندما يتحول الذوق السليم إلى سيء: الآليات العصبية الكامنة وراء ظهور التأثير السلبي وما يرتبط به من تخفيض قيمة المكافأة الطبيعية بالكوكايين. الجهاز العصبي 76 (Pt B) و 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen، BT، Hopf، FW، and Bonci، A. (2010). اللدونة متشابك في نظام mesolimbic: الآثار العلاجية لتعاطي المخدرات. آن. نيويورك أكاد. الخيال العلمي. 1187 و 129 – 139. doi: 10.1111 / j.1749-6632.2009.05154.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Colantuoni، C.، Rada، P.، McCarthy، J.، Patten، C.، Avena، NM، Chadeayne، A.، et al. (2002). دليل على أن تناول السكر المفرط والمتقطع يسبب الاعتماد على المواد الأفيونية الذاتية. الإجسام سمنة. احتياط 10 و 478 – 488. doi: 10.1038 / oby.2002.66
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Colantuoni، C.، Schwenker، J.، McCarthy، J.، Rada، P.، Ladenheim، B.، Cadet، JL، et al. (2001). مدخول السكر المفرط يغير الارتباط بمستقبلات الدوبامين ومو-أفيويد في الدماغ. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Crombag و HS و Gorny و G. و Li و Y. و Kolb و B. و Robinson و TE (2005). عكس الآثار المترتبة على تجربة الأمفيتامين الإدارة الذاتية على العمود الفقري شجيري في القشرة الفص الجبهي وسطي المداري. Cereb. قشرة 15 و 341 – 348. doi: 10.1093 / cercor / bhh136
Dabelea، D.، Mayer-Davis، EJ، Saydah، S.، Imperatore، G.، Linder، B.، Divers، J.، et al. (2014). انتشار نوع 1 ونوع مرض السكري 2 بين الأطفال والمراهقين من 2001 إلى 2009. JAMA 311 و 1778 – 1786. doi: 10.1001 / jama.2014.3201
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Davis، C. (2015). علم الأوبئة وعلم الوراثة من اضطراب الأكل بنهم (BED). الجهاز العصبي المركزي 20 و 522 – 529. دوى: 10.1017 / s1092852915000462
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ford، ES، and Dietz، WH (2013). الاتجاهات في مدخول الطاقة بين البالغين في الولايات المتحدة: نتائج من NHANES. صباحا. جى كلين نوتر. 97 و 848 – 853. doi: 10.3945 / ajcn.112.052662
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Fryar، CD، Carroll، MD، and Ogden، CL (2014). انتشار فرط الوزن والسمنة والسمنة الشديدة بين البالغين: الولايات المتحدة ، 1960 - 1962 من خلال 2011 - 2012. أتلانتا ، جورجيا: CDC.
Graveland، GA، and DiFiglia، M. (1985). تواتر وتوزيع الخلايا العصبية متوسطة الحجم مع نوى مسننة في neostriatum الرئيسيات والقوارض. الدماغ الدقة. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Groenewegen، HJ، Wright، CI، Beijer، AV، and Voorn، P. (1999). التقارب والفصل بين المدخلات والمخرجات الجسدية البطنية. آن. نيويورك أكاد. الخيال العلمي. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
غروف ، PM (1980). النهايات متشابك وأهدافها بعد المشيمة في neostriatum: تم الكشف عن التخصصات متشابك من تحليل المقاطع التسلسلية. بروك. NATL. أكاد. الخيال العلمي. الولايات المتحدة الأمريكية 77 و 6926 – 6929. doi: 10.1073 / pnas.77.11.6926
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Groves، PM، Linder، JC and and Young، SJ (1994). 5 - hydroxydopamine - المسمى المحاور الدوباميني: إعادة بناء ثلاثية الأبعاد للمحاور ، المشابك والأهداف بعد المشبكي في neostriatum الفئران. علم الأعصاب 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
هو ، FB (2013). القرار: هناك أدلة علمية كافية على أن تقليل استهلاك المشروبات المحلاة بالسكر سيقلل من انتشار السمنة والأمراض المرتبطة بالسمنة. الإجسام سمنة. القس 14 و 606 – 619. doi: 10.1111 / obr.12040
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Johnson، LR، Aylward، RL، Hussain، Z.، and Totterdell، S. (1994). مدخلات من اللوزة المخية إلى نواة الفئران المتكئة: علاقتها مع التيروزين hydroxylase immunoreactivity و الخلايا العصبية المحددة. علم الأعصاب 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Kaiya، H.، and Namba، M. (1981). نوعان من محطات الأعصاب الدوبامينية في neostriatum الفئران. دراسة البنية التحتية. علماء الأعصاب. بادئة رسالة. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Kalivas، PW، and Miller، JS (1984). الخلايا العصبية Neurotensin في مشروع المنطقة tegmental بطني إلى النواة الإنسية المتكئة. الدماغ الدقة. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
كاواغوتشي ، واي. ، ويلسون ، سي جيه ، وإيمسون ، كمبيوتر شخصي (1990). أنواع فرعية الإسقاط من الخلايا المصفوفة الفئران حديثي الولادة كشفت عن طريق الحقن داخل الخلايا من البيوسيتين. J. نيوروسكي. شنومكس، شنومكس-شنومكس.
Kelley، AE، and Swanson، CJ (1997). التغذية الناجمة عن الحصار من AMPA ومستقبلات kainate داخل المخطط البطني: دراسة رسم الخرائط microinfusion. Behav. الدماغ الدقة. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Kemp، JM، and Powell، TP (1971). تنظيم متشابك للنواة caudate. Philos. عبر. ر. وند. ب بيول. الخيال العلمي. 262 و 403 – 412. doi: 10.1098 / rstb.1971.0103
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
كيني ، PJ (2011). آليات المكافأة في السمنة: رؤى جديدة واتجاهات مستقبلية. الخلايا العصبية 69 و 664 – 679. doi: 10.1016 / j.neuron.2011.02.016
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Kessler، RC، Berglund، PA، Chiu، WT، Deitz، AC، Hudson، JI، Shahly، V.، et al. (2013). انتشار وارتباط اضطراب الإفراط في الأكل في المسوح العالمية للصحة العقلية لمنظمة الصحة العالمية. بيول. الطب النفسي 73 و 904 – 914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski، PM، Fogarty، MJ، Belmer، A.، Noakes، PG، Bellingham، MC، Bartlett، SE (2015). التوصيف الهيكلي والوظيفي للأشواك التغصنية والمدخلات المشبكية GABAergic على interneurons والخلايا الرئيسية في اللوزة القاعدية الفئران الفرعية. J. نيوروفيزيول. 114 و 942 – 957. doi: 10.1152 / jn.00824.2014
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Lenoir، M.، Serre، F.، Cantin، L.، and Ahmed، SH (2007). حلاوة مكثفة تفوق مكافأة الكوكايين. بلوس ONE 2: e698. doi: 10.1371 / journal.pone.0000698
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Li، Y.، Kolb، B.، and Robinson، TE (2003). موقع التغييرات المستمرة التي يسببها الأمفيتامين في كثافة العمود الفقري شجيري على الخلايا العصبية الشوكية المتوسطة في النواة المتكئة و putamen المذنبة. Neuropsychopharmacology 28 و 1082 – 1085. doi: 10.1038 / sj.npp.1300115
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Lindvall، O.، and Björklund، A. (1978). تشريح نظم الخلايا العصبية الدوبامينية في الدماغ الفئران. حال. الكيميائية الحيوية. Psychopharmacol. شنومكس، شنومكس-شنومكس.
Lutter، M.، and Nestler، EJ (2009). تتفاعل الإشارات التماثلية والاستدعاسية في تنظيم استهلاك الغذاء. ج. 139 و 629 – 632. doi: 10.3945 / jn.108.097618
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
مالدونادو - إريزاري ، سي إس ، سوانسون ، سي جيه ، وكيلي ، AE (1995). مستقبلات الغلوتامات في النواة المتكئة شل التحكم في تغذية السلوك عن طريق المهاد الجانبي. J. نيوروسكي. شنومكس، شنومكس-شنومكس.
Malik و VS و Popkin و BM و Bray و GA و Després و JP و Hu و FB (2010). المشروبات المحلاة بالسكر والسمنة ونوع داء السكري 2 ومخاطر أمراض القلب والأوعية الدموية. تداول 121 و 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ng، SW، Slining، MM، and Popkin، BM (2012). استخدام المحليات السعرات الحرارية وغير القلوية في الأغذية المعلبة المستهلكة في الولايات المتحدة ، 2005-2009. ج. أكاد. نوتر. حمية. 112 و 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Nielsen، SJ، Siega-Riz، AM، and Popkin، BM (2002). الاتجاهات في مدخول الطاقة في الولايات المتحدة بين 1977 و 1996: تظهر تحولات مماثلة عبر الفئات العمرية. الإجسام سمنة. احتياط 10 و 370 – 378. doi: 10.1038 / oby.2002.51
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Paxinos، G.، and Watson، C. (2007). الدماغ الفئران في الإحداثيات التجسيمي. أمستردام. بوسطن ، ماساتشوستس: Academic Press / Elsevier.
بوبكين ، بي إم (2010). ما الخطأ في نهج الولايات المتحدة للسمنة؟ المرشد الظاهري 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Rada، P.، Avena، NM، and Hoebel، BG (2005). الإفراط في تناول السكر يوميا الإفراج عن الدوبامين بشكل متكرر في قذيفة المتكئين. علم الأعصاب 134 و 737 – 744. doi: 10.1016 / j.neuroscience.2005.04.043
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Rafols، JA، Cheng، HW، and McNeill، TH (1989). دراسة جولجي لمخطط الفأر: التغيرات التغصنية المرتبطة بالعمر في التجمعات العصبية المختلفة. J. شركات. Neurol. 279 و 212 – 227. doi: 10.1002 / cne.902790205
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ranjan، A.، and Mallick، BN (2010). طريقة معدلة لتلطيخ Golgi-cox متسقة وموثوق بها في وقت مخفض بشكل ملحوظ. أمامي. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Reinehr، T. (2013). اكتب مرض السكري 2 في الأطفال والمراهقين. العالم J. مرض السكري 4 و 270 – 281. doi: 10.4239 / wjd.v4.i6.270
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Reissner، KJ، and Kalivas، PW (2010). استخدام توازن الغلوتامات كهدف لعلاج الاضطرابات الإدمانية. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Reynolds، SM، and Berridge، KC (2003). مجموعات غلوتامات تحفيزية في النواة المتكئة: تدرجات قذيفة روستوكودال من الخوف والتغذية. يورو. J. نيوروسكي. 17 و 2187 – 2200. doi: 10.1046 / j.1460-9568.2003.02642.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Richard، JM، and Berridge، KC (2011). تتفاعل Nucleus مع أنماط تفاعلات الدوبامين / الغلوتامات لتوليد الرغبة مقابل الرهبة: D (1) وحدها لتناول الطعام الشهية ولكن D (1) و D (2) معاً للخوف. J. نيوروسكي. 31 و 12866 – 12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Robinson، TE، and Kolb، B. (1999). تعديلات في مورفولوجيا التشعبات والأشواك الشجيري في النواة المتكئة والقشرة الجبهية الأمامية بعد العلاج المتكرر مع الأمفيتامين أو الكوكايين. يورو. J. نيوروسكي. 11 و 1598 – 1604. doi: 10.1046 / j.1460-9568.1999.00576.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Robinson، TE، and Kolb، B. (2004). اللدونة الهيكلية المرتبطة التعرض لأدوية سوء المعاملة. الجهاز العصبي 47 (ملحق 1) ، 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Russo، SJ، Dietz، DM، Dumitriu، D.، Morrison، JH، Malenka، RC، and Nestler، EJ (2010). المشبك المدمن: آليات اللدونة المشبكية والهيكلية في النواة المتكئة. اتجاهات neurosci. 33 و 267 – 276. doi: 10.1016 / j.tins.2010.02.002
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Rutledge، LT، Duncan، J.، and Beatty، N. (1969). دراسة لهرمونات محور عصبي الخلية الهرمية في القشرة المخية سليمة وغير معزولة جزئيا. الدماغ الدقة. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper، CB، Chou، TC، Elmquist، JK (2002). الحاجة إلى إطعام: السيطرة على التوحيد والتحكم في الأكل. الخلايا العصبية 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Scheggi، S.، Secci، ME، Marchese، G.، De Montis، MG، and Gambarana، C. (2013). تأثير الاستساغة على التحفيز على العمل من أجل الغذاء من السعرات الحرارية وغير السعرات الحرارية في الجرذان المحرومة من الغذاء والمحرومة من الغذاء. علم الأعصاب 236 و 320 – 331. doi: 10.1016 / j.neuroscience.2013.01.027
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Sesack، SR، and Pickel، VM (1990). في نواة الفئران الإنسية المتكئة ، تتلاقى أطراف الحصين و الكاتيكولامينية على الخلايا العصبية الشوكية و هي في موضعها لبعضها البعض. الدماغ الدقة. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Sesack، SR، and Pickel، VM (1992). افرازات القشرية قبل الجبهية في المشبك الفئران على الأهداف العصبية غير الخاضعة للكتابة من المحطات الكاتيكولامينية في النواة المتكئة septi وعلى الخلايا العصبية الدوبامين في المنطقة tegmental بطني. J. شركات. Neurol. 320 و 145 – 160. doi: 10.1002 / cne.903200202
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Shariff، M.، Quik، M.، Holgate، JY، Morgan، M.، Patkar، OL، Tam، V.، et al. (في الصحافة). نيوترون nicotinic مستقبلات أستيل كولين تقلل من تناول السكر. بلوس ONE.
Sheehan، DV، and Herman، BK (2015). العوامل النفسية والطبية المرتبطة باضطراب الإفراط في الأكل غير المعالج. متزمت. الرعاية رفيق CNS Disord. 17. doi: 10.4088 / PCC.14r01732
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Simms، JA، Steensland، P.، Medina، B.، Abernathy، KE، Chandler، LJ، Wise، R.، et al. (2008). يؤدي الوصول المتقطع إلى 20٪ من الإيثانول إلى استهلاك عالي للإيثانول في فئران Long-Evans و Wistar. الكحول. كلين. إكسب. احتياط 32 و 1816 – 1823. doi: 10.1111 / j.1530-0277.2008.00753.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Steensland، P.، Simms، JA، Holgate، J.، Richards، JK، and Bartlett، SE (2007). Varenicline ، وهو مستقلب جزئية لمستقبلات الأستيل كولين النسيلة ، يقلل بشكل انتقائي من استهلاك الإيثانول والسعي. بروك. NATL. أكاد. الخيال العلمي. الولايات المتحدة الأمريكية 104 و 12518 – 12523. doi: 10.1073 / pnas.0705368104
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Stuber، GD، Hopf، FW، Hahn، J.، Cho، SL، Guillory، A.، and Bonci، A. (2008a). تناول الإيثانول الطوعي يعزز قوة متشابك الاستثارة في المنطقة tegmental بطني. الكحول. كلين. إكسب. احتياط 32 و 1714 – 1720. doi: 10.1111 / j.1530-0277.2008.00749.x
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Stuber، GD، Klanker، M.، de Ridder، B.، Bowers، MS، Joosten، RN، Feenstra، MG، et al. (2008b). تعزز الإشارات التنبؤية المكافئة قوة التشابك الاستثارية على الخلايا العصبية الدوبامين الدماغ المتوسط. علوم 321 و 1690 – 1692. doi: 10.1126 / science.1160873
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Swanson، SA، Crow، SJ، Le Grange، D.، Swendsen، J.، and Merikangas، KR (2011). انتشار وارتباطات اضطرابات الأكل في المراهقين. النتائج من الملحق مراهقة تكرار الاعتلال المشترك الوطنية. قوس. الطب النفسي العام 68 و 714 – 723. doi: 10.1001 / archgenpsychiatry.2011.22
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Te Morenga، L.، Mallard، S.، and Mann، J. (2013). السكريات الغذائية ووزن الجسم: المراجعة المنهجية والتحليلات التلوية للتجارب العشوائية ذات الشواهد ودراسات الأتراب. BMJ 346: e7492. دوى: 10.1136 / bmj.e7492
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Totterdell، S.، and Smith، AD (1989). التقارب من مدخل الحصين والدوبامين على الخلايا العصبية المحددة في النواة المتكئة من الفئران. جي كيم. Neuroanat. شنومكس، شنومكس-شنومكس.
Tukey ، DS ، Ferreira ، JM ، Antoine ، SO ، D'Amour ، JA ، Ninan ، I. ، Cabeza de Vaca ، S. ، et al. (2013). يؤدي تناول السكروز إلى تهريب سريع لمستقبلات AMPA. J. نيوروسكي. 33 و 6123 – 6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Veening، JG، Cornelissen، FM، and Lieven، PA (1980). التنظيم الموضعي ل afferents إلى caudatoputamen من الفئران. دراسة البيروكسيديز الفجل. علم الأعصاب 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Ventura، T.، Santander، J.، Torres، R.، Contreras، AM (2014). أساس العصبية الحيوية من حنين للكربوهيدرات. ركن المعلومات الغذائية 30 و 252 – 256. doi: 10.1016 / j.nut.2013.06.010
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Volkow، ND، Wang، GJ، Fowler، JS، Tomasi، D.، and Baler، R. (2012). مكافأة الغذاء والدواء: تراكب الدوائر في السمنة البشرية والإدمان. داء. أعلى. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
الحكيم ، RA (1973). تناول الإيثانول الطوعي في الجرذان بعد التعرض للإيثانول في جداول مختلفة. Psychopharmacologia 29 و 203 – 210. دوى: 10.1007 / BF00414034
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
Wright، CI، and Groenewegen، HJ (1995). أنماط التقارب والفصل في النواة الأنسية المتكئة للفئران: علاقات القشرة الأمامية القبلية ، المهادية المتوسطة ، القاعدية اللولبية القاعدية. J. شركات. Neurol. 361 و 383 – 403. doi: 10.1002 / cne.903610304
مجلات PubMed | CrossRef النص الكامل | الباحث العلمي من Google
الكلمات الرئيسية: الاستهلاك الشبيه بنهم ، طويل الأجل ، شوكي عصبي متوسط ، نواة متكئة ، سكروز
الاقتباس: Klenowski PM، Shariff MR، Belmer A، Fogarty MJ، Mu EWH، Bellingham MC and Bartlett SE (2016) Prolonged Consumption of Sucrose in a Binge-Like Manner، Alters the Morphology of Medium Spiny Neurons in the Nucleus Accumbens Shell. أمامي. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
تم الاستلام: 03 December 2015؛ تم القبول: 07 March 2016؛
تاريخ النشر: 23 مارس 2016.
حرره:
جوهر نورة أبروس، Institut des Neurosciences de Bordeaux، France
تمت مراجعته من قبل:
سيرج ح. أحمد، National National de la Recherche Scientifique، France
ستيفاني كايلي، National National de la Recherche Scientifique، France
حقوق الطبع والنشر © 2016 Klenowski و Shariff و Belmer و Fogarty و Mu و Bellingham و Bartlett. هذا هو مقال مفتوح الوصول موزعة وفقا لشروط ترخيص Creative Commons Attribution (CC BY). يُسمح بالاستخدام أو التوزيع أو الاستنساخ في المنتديات الأخرى بشرط أن يُنسب إلى المؤلف الأصلي (المؤلفين) أو المرخِّص الأصلي وأن المنشور الأصلي في هذه المجلة يتم الاستشهاد به ، وفقًا للممارسات الأكاديمية المقبولة. لا يسمح باستخدام أو توزيع أو إعادة إنتاج لا يتوافق مع هذه الشروط.
* المراسلات: سيلينا إ. بارتليت ، [البريد الإلكتروني محمي]