COMENTARIS: Com estudis posteriors revelaran DeltaFosB és l'interruptor molecular comú tant per a addiccions de drogues com per conducta. És un factor de transcripció que significa que afecta quins gens estan activats o desactivats. Com s’ha dit en altres llocs, les drogues addictives només segresten els mecanismes normals. Per això, és ximple suggerir que no poden existir addiccions conductuals.
Proc Natl Acad Sci EUA A. 2001 25; 98 (20): 11042 – 11046.
doi: 10.1073 / pnas.191352698.
Eric J. Nestler *, Michel Barrot i David W. Self
Departament de Psiquiatria i Centre de Neurociència Bàsica, Centre Mèdic Sud-oest de la Universitat de Texas, 5323 Boulevard Harry Hines, Dallas, TX 75390-9070
abstracte
La longevitat d'algunes de les anomalies de comportament que caracteritzen l'addicció a les drogues ha suggerit que la regulació de l'expressió gènica neuronal pot estar implicada en el procés pel qual les drogues d'abús causen un estat d'addicció. JoL'augment de la evidència suggereix que el factor de transcripció BFosB representa un mecanisme pel qual les drogues d'abús produeixen canvis relativament estables en el cervell que contribueixen al fenotip d'addicció. ΔFosB, membre de la família de factors de transcripció de Fos, s'acumula dins d’un subconjunt de neurones del nucli accumbens i dorsal striatum (regions cerebrals importants per a l’addicció) després d’administrar repetidament molts tipus de drogues d’abús. Una acumulació similar de FosB ocorre després d’un funcionament compulsiu, el que suggereix que ΔFosB pot acumular-se en resposta a molts tipus de comportaments compulsius. És important destacar que ΔFosB persisteix a les neurones durant períodes de temps relativament llargs a causa de la seva extraordinària estabilitat. Per tant, ΔFosB representa un mecanisme molecular que podria iniciar i després mantenir canvis en l’expressió gènica que persisteixen molt després de l’extinció de l’exposició als medicaments. Els estudis en ratolins transgènics inducibles que sobreexpressen ΔFosB o un inhibidor negatiu dominant de la proteïna proporcionen evidència directa que ΔFosB provoca un augment de la sensibilitat als efectes conductuals de les drogues d'abús i, possiblement, un augment del comportament de la recerca de drogues. Aquest treball dóna suport a la idea que ΔFosB funciona com un tipus de "interruptor molecular" sostingut que converteix gradualment les respostes agudes en medicaments a adaptacions relativament estables que contribueixen a la plasticitat neuronal i conductual a la llarga de l’addicció.
La investigació sobre l'addicció es centra en entendre les maneres complexes en què les drogues d'abús canvien el cervell per causar anomalies de comportament que caracteritzen l'addicció. Un dels reptes fonamentals en el camp és identificar canvis induïts pel fàrmac en el cervell relativament estables per tenir en compte les anomalies de comportament que són particularment de llarga durada. Per exemple, un addicte humà pot estar en major risc de recaiguda fins i tot després d'anys d'abstinència.
L’estabilitat d’aquestes anomalies de comportament ha donat lloc al suggeriment que poden estar mediats, almenys en part, mitjançant canvis en l’expressió gènica (1 – 3). Segons aquest punt de vista, l'exposició repetida a una droga d'abús pertorba repetidament la transmissió en determinades sinapsis del cervell que són sensibles al fàrmac. Aquestes pertorbacions finalment senyalen a través de les cascades de missatgeria intracel·lular al nucli, on inicialment inicien i després mantenen canvis en l’expressió de gens específics. Un mecanisme primari a través del qual les vies de transducció de senyals influeixen en l'expressió gènica és la regulació dels factors de transcripció, proteïnes que s'uneixen a regions reguladores dels gens i modificen la seva transcripció.
Per tant, un dels objectius de la investigació sobre l’addicció ha estat identificar factors de transcripció que s’alteren a les regions del cervell implicades en l’addicció després de l’administració crònica de fàrmacs d’abús. Diversos factors de transcripció s'han identificat durant la darrera dècada (1 – 6). L'objectiu d'aquesta revisió és un factor de transcripció anomenat BFOSB.
Inducció de ΔFosB per drogues d'abús
BFosB, codificat pel gen fosB, és membre de la família de factors de transcripció Fos, que també inclouen c-Fos, FosB, Fra1 i Fra2 (7). Aquestes proteïnes de la família Fos es heterodimerizan amb proteïnes de la família Jun (c-Jun, JunB o JunD) per formar factors de transcripció actius AP-1 (proteïna activadora-1) que s'uneixen a llocs AP-1 (seqüència consens: TGAC / GTCA) presents a els promotors de certs gens per regular la seva transcripció.
Aquestes proteïnes de la família Fos s’indueixen de manera ràpida i transitòria en regions específiques del cervell després d’administrar de forma aguda molts fàrmacs d’abús (fig. 1) (8 – 11). Les regions destacades són el nucli accumbens i el dorsal striatum, que són mediadors importants de les respostes conductuals als medicaments, en particular els seus efectes gratificants i activadors de locomotors (12, 13). Aquestes proteïnes tornen als nivells basals en hores d’administració de drogues.
figura 1
Esquema que mostra l'acumulació gradual de FosB enfront de la inducció ràpida i transitòria d'altres proteïnes de la família Fos en resposta a drogues d'abús. (A) L'autoradiograma il·lustra la inducció diferencial d'aquestes proteïnes mitjançant l'estimulació aguda (1-2 h després d'una única exposició a un fàrmac) versus estimulació crònica (1 dia després de l'exposició repetida a medicaments). (B) Diverses onades de proteïnes similars a Fos [formades per c-Fos (isoformes 52 a 58-kDa), FosB (isoformes 46 a 50-kDa), ΔFosB (isoforma 33-kDa) i Fra1 o Fra2 ( 40 kDa)] s’indueixen al nucli accumbens i a les neurones estriatals dorsals per administració aguda d’un medicament d’abús. També s'indiquen les isoformes bioquímiques de ΔFosB (35-37 kDa); també són induïdes (encara que a nivells baixos) després de l'administració aguda de fàrmacs, però persisteixen en el cervell durant llargs períodes a causa de la seva estabilitat. (C) Amb l'administració de medicaments repetits (per exemple, dues vegades al dia), cada estímul agut indueix un nivell baix de les isoformes ΔFosB estables, que està indicat pel conjunt inferior de línies superposades que indiquen ΔFosB induït per cada estímul agut. El resultat és un augment gradual dels nivells totals de BFosB amb estímuls repetits durant un curs de tractament crònic, que està indicat per la creixent línia gradual al gràfic.
S'observen respostes molt diferents després d’administrar crònicament medicaments d’abús (fig. 1). Les isoformes bioquímicament modificades de ΔFosB (massa molecular 35-37 kDa) s'acumulen dins de les mateixes regions del cervell després d'una exposició repetida a medicaments, mentre que la resta de membres de la família Fos mostren tolerància (és a dir, reducció de la inducció en comparació amb les exposicions inicials de medicaments). Aquesta acumulació de ΔFosB s'ha observat per a la cocaïna, la morfina, l’anfetamina, l’alcohol, la nicotina i la fenciclidina.e (11, 14 – 18). Hi ha algunes evidències que aquesta inducció és selectiva per al subconjunt que conté la dinorfina / substància P de neurones espinoses mitjanes localitzades en aquestes regions cerebrals (15, 17), encara que es necessita més treball per establir-ho amb certesa. Les isoformes 35 a 37-kDa de ΔFosB es dimeritzen predominantment amb JunD per formar un complex AP-1 actiu i durador dins d'aquestes regions cerebrals (19, 20). Aquestes isoformes de BFosB s'acumulen amb l’exposició crònica a medicaments a causa de la seva vida mitjana extraordinàriament llarga (21) i, per tant, persisteixen a les neurones durant almenys diverses setmanes després del cessament de l’administració del fàrmac. És interessant observar que aquestes isoformes BFosB són productes altament estables d’un gen precoç immediat (fosB). L'estabilitat de les isoformes ΔFosB proporciona un nou mecanisme molecular pel qual els canvis induïts per fàrmacs en l'expressió gènica poden persistir malgrat períodes relativament llargs de retirada de fàrmacs.
Tot i que el nucli accumbens juga un paper crític en els efectes gratificants de les drogues d'abús, es creu que funciona normalment mitjançant la regulació de les respostes als reforçadors naturals, com ara menjar, beguda, sexe i interaccions socials (12, 13). Com a resultat, hi ha un considerable interès en un possible paper d’aquesta regió cerebral en altres comportaments compulsius (per exemple, menjar excessivament patològic, joc, exercici, etc.). Per aquest motiu, vam examinar si ΔFosB està regulat en un model animal de funcionament compulsiu. De fet, les isoformes estables de 35 a 37-kDa de ΔFosB són induïdes selectivament dins del nucli accumbens en rates que mostren un comportament compulsiu. †
Identitat bioquímica de les isoformes estables BFosB
Com es va esmentar anteriorment, les isoformes ΔFosB que s'acumulen després de l’administració crònica d’un fàrmac d’abús o compulsiu mostren una massa molecular de 35 – 37 kDa. Es poden diferenciar de la isoforma de 33-kDa de ΔFosB que s'indueix ràpidament però de manera transitòria després d’una única exposició de fàrmacs (Fig. 1) (14, 19, 22). Les proves actuals suggereixen que la isoforma 33-kDa és la forma nativa de la proteïna, que es modifica per formar els productes 35- a 37-kDa més estables (19, 21). No obstant això, la naturalesa de la modificació bioquímica que converteix la isoforma inestable 33-kDa en les isoformes estables de 35 a 37-kDa ha romàs obscura. S'ha especulat que la fosforilació pot ser responsable (11). Per exemple, la inducció de ΔFosB és atenuada en ratolins que no tenen DARPP-32, una proteïna enriquida amb estriat (23, 24). Com que DARPP-32 regula l'activitat catalítica de la proteïna fosfatasa-1 i la proteïna quinasa A (25, 26), el requisit d'aquesta proteïna per a l'acumulació normal de les isoformes ΔFosB estables suggereix un possible paper per a la fosforilació en la generació d'aquests productes estables.
Paper de BFosB en plasticitat conductual a drogues d'abús
La comprensió de la funció de FosB en la drogodependència ha sorgit en gran part de l’estudi de ratolins transgènics en els quals ΔFosB pot ser induït selectivament dins del nucli accumbens i altres regions estriatals d’animals adults (27, 28). És important destacar que aquests ratolins sobreexpressen ΔFosB selectivament a les neurones espinoses mitjanes que contenen la dinorfina / substància P, on es creu que els fàrmacs indueixen la proteïna. El fenotip del comportament dels ratolins xFosB-sobreexpressius, que de moltes maneres s'assembla a animals després de l'exposició crònica a les drogues, es resumeix a la taula 1. Els ratolins mostren respostes locomotores augmentades a la cocaïna després de l'administració crònica i aguda (28). També mostren una major sensibilitat cap als efectes gratificants de la cocaïna i la morfina en els assajos de condicionament del lloc (11, 28) i s'autocontrolaran dosis més baixes de cocaïna que els que no sobreexpressen BFosB., En canvi, aquests animals mostren un locomotor condicionat normal. la sensibilització a la cocaïna i l’aprenentatge espacial normal en el laberint d’aigua de Morris (28). Taquestes dades indiquen que ΔFosB augmenta la sensibilitat d'un animal a la cocaïna i potser a altres drogues d'abús i pot representar un mecanisme de sensibilització relativament prolongada a les drogues.
 estriatum
estriatum
| Augment de l’activació locomotora en resposta a l’administració aguda i repetida de cocaïna. |
| Augment de les respostes gratificants a la cocaïna i la morfina en assaigs de condicionament del lloc. |
| Major auto-administració de baixes dosis de cocaïna. |
| Augment de la motivació per la cocaïna en assajos de ràtio progressiva. |
| Augment de les respostes ansiolítiques a l’alcohol. |
| Augment del comportament compulsiu. |
Basat en dades de refs. 28 i 29.† ‡ §¶
Plasticitat del comportament mediada per ΔFosB al nucli accumbens-dorsal striatum
IA més, hi ha proves preliminars que els efectes de mayFosB poden transcendir més enllà d’una regulació de la sensibilitat de les drogues per si mateixes a comportaments més complexos relacionats amb el procés d’addicció. Els ratolins que expressen BFosB treballen més per autoadministrar-se la cocaïna en assajos autoadministratius de relació progressiva,ggesting que ΔFosB pot sensibilitzar els animals a les propietats motivacionals d'incentius de la cocaïna i, per tant, conduir a una propensió a la recaiguda després de la retirada de droguesEls ratolins que expressen FOSB també mostren efectes ansiolítics millorats de l'alcohol, un fenotip que s'ha associat amb un augment de la ingesta d'alcohol en humans. Junts, aquests primers descobriments suggereixen que ΔFosB, a més d’augmentar la sensibilitat als fàrmacs d’abús, produeix canvis qualitatius en el comportament que promouen la conducta que busca la droga. Per tant, ΔFosB pot funcionar com un "interruptor molecular" sostingut que ajuda a iniciar i després mantenir aspectes crucials de l'estat addicte. Una qüestió important en la investigació actual és si l’acumulació de BFosB durant l’exposició al fàrmac promou un comportament de recerca de drogues després de períodes prolongats de retirada, fins i tot després que els haveFosB s’han normalitzat (vegeu més avall).
Adult Els ratolins que sobreexpressen ΔFosB selectivament dins del nucli accumbens i el dorsal striatum també presenten un major funcionament compulsiu en comparació amb els controladors de llars. † Aquestes observacions plantegen la interessant possibilitat que l'acumulació de ΔFosB en aquestes neurones tingui un paper més general en la formació i manteniment de records d'hàbits i compulsius. comportaments, potser reforçant l’eficàcia dels circuits neuronals en què funcionen aquestes neurones.
ΔFosB s'acumula a determinades regions del cervell fora del nucli accumbens i del dorsal striatum després de l'exposició crònica a la cocaïna. Destaca entre aquests les regions són l’amígdala i l’escorça prefrontal medial (15). Un dels principals objectius de la investigació actual és entendre les contribucions de la inducció de ΔFosB en aquestes regions al fenotip de l’addicció.
Els treballs anteriors sobre ratolins eliminatoris fosB van revelar que aquests animals no desenvolupen la sensibilització als efectes locomotors de la cocaïna, cosa que és coherent amb les troballes dels ratolins sobreexpressors ΔFosB esmentats anteriorment (22). No obstant això, els mutants fosB van mostrar una sensibilitat millorada als efectes aguts de la cocaïna, cosa que és incompatible amb aquests altres descobriments. Tanmateix, la interpretació de les troballes amb els mutants fosB és complicada pel fet que a aquests animals no només els falta ΔFosB, sinó també FosB complet. A més, els mutants no tenen proteïnes a tot el cervell i des de les primeres etapes del desenvolupament. De fet, treballs més recents donen suport a conclusions dels ratolins sobreexpressors ΔFosB: sobreexpressió induïble d’un mutant truncat de c-Jun, que actua com a antagonista negatiu dominant de osFosB, selectivament en el nucli accumbens i l’estrat dorsal mostra una sensibilitat reduïda als efectes gratificants de la cocaïna Aquestes troballes emfatitzen la precaució que s’ha d’utilitzar en la interpretació de resultats de ratolins amb mutacions constitutives i il·lustren la importància dels ratolins amb mutacions induïbles i específiques del tipus cel·lular en estudis de plasticitat al cervell adult.
Gènes objectiu per a ΔFosB
Atès que BFosB és un factor de transcripció, és probable que la proteïna produeixi plasticitat conductual a través d’alteracions en l’expressió d’altres gens. ΔFosB es genera mitjançant splicing alternatiu del gen fosB i manca d'una part del domini de transactivació C-terminal present en FosB de longitud completa. Com a resultat, es va proposar originalment que ΔFosB funcionés com a repressor transcripcional (29). Tanmateix, el treball en cultius cel·lulars ha demostrat clarament que ΔFosB pot induir o reprimir Transcripció mediada per AP-1 en funció del lloc AP-1 específic utilitzat (21, 29-31). El FosB de longitud completa exerceix els mateixos efectes que BFosB en certs fragments promotors, però efectes diferents sobre els altres. Cal treballar més per entendre els mecanismes que sustenten aquestes variades accions de BFosB i FosB.
El nostre grup ha utilitzat dos enfocaments per identificar gens diana per a osFosB. Un és l’enfocament genètic candidat. Inicialment, vam considerar els receptors de glutamat de l’àcid α-amino-3-hidroxi-5-metil-4-isoxazolepropionic (AMPA) com a objectius suposats, atès el paper important de la transmissió glutamatergica al nucli accumbens. Els treballs realitzats fins ara indiquen que una subunitat en particular del receptor de glutamat AMPA, GluR2, pot ser un objectiu de bona fe per a osFosB (figura 2). L'expressió de GluR2, però no l'expressió d'altres subunitats del receptor AMPA, augmenta en el nucli accumbens (però no en l'estri dorsal) després de la sobreexpressió de ΔFosB (28), i l'expressió d'un mutant negatiu dominant atenua la capacitat de la cocaïna per induir la proteïna. A més, el promotor del gen GluR2 conté un lloc consensuat AP-1 que uneix ΔFosB (28). La sobreexpressió de GluR2 al nucli accumbens, mitjançant l’ús de transferència gènica mediada per virus, augmenta la sensibilitat d’un animal als efectes gratificants de la cocaïna, imitant així part del fenotip vist en ratolins que expressen expressFosB (28). La inducció de GluR2 podria explicar la reducció de la sensibilitat electrofisiològica de les neurones del nucli accumbens als agonistes del receptor AMPA després de l’administració crònica de cocaïna (32), perquè els receptors AMPA que contenen GluR2 mostren una conductança global reduïda i una permeabilitat Ca2 + reduïda. La reducció de la resposta d’aquestes neurones a les aportacions excitadores pot augmentar les respostes a una droga d’abús. Tot i això, es desconeixen les maneres en què els senyals dopaminèrgics i glutamatergics del nucli accumbens regulen el comportament addictiu; això requerirà un nivell de comprensió del circuit neuronal, que encara no està disponible.
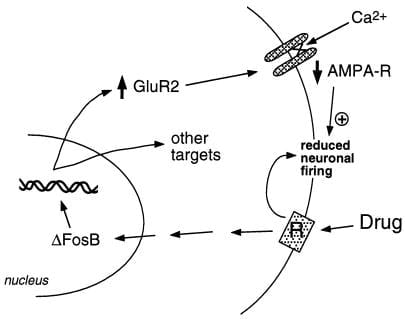
figura 2
La subunitat del receptor de glutamat AMPA, GluR2, és un objectiu suposat per a osFosB. Es mostra com la inducció mediada per ΔFosB de GluR2 pot alterar la capacitat de resposta fisiològica de les neurones del nucli accumbens i provocar respostes sensibilitzades a les drogues d'abús. Segons aquest esquema, les drogues d'abús produeixen els seus efectes aguts de reforç mitjançant la inhibició de les neurones del nucli accumbens. Amb una exposició repetida, els medicaments indueixen osFosB, que regula nombrosos gens diana, inclòs el GluR2. Això augmenta la proporció de receptors AMPA (AMPA-R) a les neurones del nucli accumbens que contenen la subunitat GluR2, cosa que provoca una reducció del corrent general d’AMPA i una reducció del corrent de Ca2 +. Aquesta excitabilitat reduïda pot fer que les neurones siguin més sensibles als efectes inhibidors aguts dels medicaments i, per tant, als efectes de reforç dels medicaments..
Un altre objectiu putatiu de FosB és el gen que codifica la dinorfina. Com es va dir anteriorment, la dinorfina s'expressa en el subconjunt del nucli accumbens de les neurones espinoses mitjanes que mostren la inducció de ΔFosB. Dynorphin sembla funcionar en un bucle de retroalimentació intercel·lular: la seva alliberament inhibeix les neurones dopaminèrgiques que innerven les neurones espinoses mitjanes, a través de receptors opiáceos κ presents en terminals nervioses dopaminèrgiques al nucli accumbens i també en cossos i dendrites de la zona ventral tegmental (Fig. 3) (33 – 35). Aquesta idea és coherent amb la capacitat d’un agonista del receptor κ, després d’administrar-lo a qualsevol d’elles, per disminuir la reoberturad (35).
REl treball ecent ha indicat que ΔFosB disminueix l’expressió de la dinorfina, could que podria contribuir a la millora dels mecanismes de recompensa vistos amb la inducció de BFosB. Curiosament, un altre factor de transcripció regulat per fàrmacs, el CREB (proteïna d’enllaç de l’element de resposta d’AMPc) (2, 3), exerceix l’efecte contrari: indueix l’expressió de la dinorfina al nucli accumbens i redueix les propietats gratificants de la cocaïna i la morfina. (4). **
BL’activació de CREB induïda per fàrmacs es dissipa ràpidament després d’administrar medicaments; aquesta regulació recíproca de la dinorfina per CREB i ΔFosB podria explicar els canvis de comportament recíprocs que es produeixen durant les fases primerenques i tardanes de la retirada, amb símptomes emocionals negatius i reducció de la sensibilitat farmacèutica durant les primeres fases de la retirada i la sensibilització cap als efectes motivacionals d’incentius i gratificants de les drogues predominants en els moments posteriors.
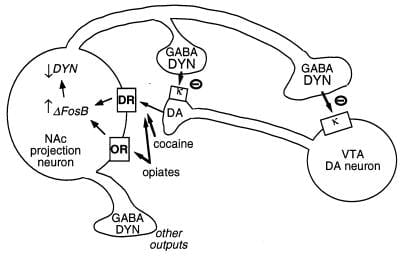
figura 3
La dinorfina és un objectiu putatiu per a ΔFosB. Es mostra una neurona de la zona tegmental ventral (VTA) de la dopamina (DA) inervant una classe de nucli accumbens (NAc) neurona de projecció GABAergic que expressa la dinorfina (DYN). Dynorphin serveix un mecanisme de retroalimentació en aquest circuit: la dynorphin, alliberada de les terminals de les neurones NAc, actua sobre receptors opioides κ situats en terminals nervioses i cossos cel·lulars de les neurones DA per inhibir el seu funcionament. ΔFosB, mitjançant l’inhibició de l’expressió de la dinorfina, pot reduir aquest bucle de retroalimentació i millorar les propietats gratificants de les drogues d’abús. No es mostra l’efecte recíproc del CREB en aquest sistema: el CREB millora l’expressió de la dinorfina i, per tant, atenua les propietats gratificants de les drogues d’abús. (4). Àcid GABA, γ-aminobutíric; DR, receptor de dopamina; OR, receptor opioide.
El segon enfocament utilitzat per identificar gens diana per a osFosB implica l’anàlisi de microarrays d’ADN. La sobreexpressió induïble de osFosB augmenta o disminueix l’expressió de nombrosos gens al nucli accumbens (36). Tot i que ara es necessita un treball considerable per validar cadascun d’aquests gens com a dianes fisiològiques de osFosB i per entendre la seva contribució al fenotip d’addicció, sembla que un objectiu important és Cdk5 (cinasa-5 dependent de la ciclina). Per tant, Cdk5 es va identificar inicialment com a ΔFosB regulat per l’ús de microarrays i, posteriorment, es va demostrar que estava induït al nucli accumbens i a l’estrat dorsal després de l’administració crònica de cocaïna (37). OsFosB activa el gen cdk5 a través d’un lloc AP-1 present al promotor del gen (36). Juntes, aquestes dades donen suport a un esquema en què la cocaïna indueix l’expressió de Cdk5 en aquestes regions cerebrals mitjançant ΔFosB. La inducció de Cdk5 sembla alterar la senyalització dopaminèrgica almenys en part a través de l’augment de la fosforilació de DARPP-32 (37), que es converteix d’un inhibidor de la proteïna fosfatasa-1 a un inhibidor de la proteïna quinasa A després de la seva fosforilació per Cdk5 (26).
Paper de BFosB en la mediació de la plasticitat "permanent" davant les drogues d'abús
Tot i que el senyal ΔFosB té una vida relativament llarga, no és permanent. BFosB es degrada gradualment i ja no es pot detectar en el cervell després dels mesos de retirada de fàrmacs 1 – 2, tot i que certes anomalies de comportament persisteixen durant períodes de temps molt més llargs. Per tant, ΔFosB per se no sembla que sigui capaç de mediar aquestes anomalies de comportament semipermanents. La dificultat per trobar les adaptacions moleculars que sustenten els canvis de comportament extremadament estables associats a l'addicció són anàlogues als reptes que afronten el camp de la memòria i l'aprenentatge. Tot i que hi ha models mòbils i moleculars elegants d’aprenentatge i memòria, no s’ha pogut identificar fins ara les adaptacions moleculars i cel·lulars prou llargues per tenir en compte records de comportament altament estables. De fet, ΔFosB és l’adaptació més longeva que es coneix al cervell adult, no només en resposta a drogues d’abús, sinó també a qualsevol altra pertorbació (que no impliqui lesions). Han evolucionat dues propostes, tant en els camps de l’addicció com de l’aprenentatge i la memòria, per explicar aquesta discrepància.
Una possibilitat és que canvis més transitoris en l’expressió gènica, com ara els mediats a través de BFosB o altres factors de transcripció (per exemple, CREB), pot mediar canvis més de llarga durada en la morfologia neuronal i en l’estructura sinàptica. Per exemple, Un augment de la densitat de les espines dendrítiques (especialment un augment de les espines dobles) acompanya a la major eficàcia de les sinapsis glutamatergiques en neurones piramidals de l'hipocamp durant la potenciació a llarg termini (38-40), i paral·lel a la millora de la sensibilitat conductual a la cocaïna mediada a nivell de neurones espinoses mitjanes del nucli accumbens (41). No se sap si aquests canvis estructurals són prou llargs per tenir en compte els canvis de comportament altament estables, tot i que aquests últims persisteixen almenys durant el mes de retirada de medicaments. Les evidències recents plantegen la possibilitat que ΔFosB, i la seva inducció de Cdk1, sigui un mediador de canvis induïts per fàrmacs en l’estructura sinàptica del nucli accumbens (Fig. 5) ‡‡ Així, la infusió d’un inhibidor de Cdk4 al nucli accumbens evita que capacitat d’exposició repetida de cocaïna per augmentar la densitat de la columna dendrítica d’aquesta regió. Això és coherent amb l'opinió que Cdk5, que està enriquit en cervell, regula l'estructura neural i el creixement (vegeu refs. 5 i 36). És possible, encara que de cap manera demostrat, que tals canvis en la morfologia neuronal puguin sobreviure a la mateixa senyal ΔFosB.
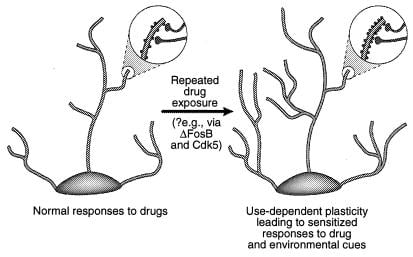
figura 4
Regulació de l'estructura dendrítica per drogues d'abús. Es mostra l'expansió de l'arbre dendrític d'una neurona després de l'exposició crònica a una droga d'abús, com s'ha observat amb la cocaïna al nucli accumbens i a l'escorça prefrontal (41). Les àrees d'ampliació mostren un augment de les espines dendrítiques, que es postula que es produeix juntament amb els terminals nerviosos activats. Aquest augment de la densitat de la columna dendrítica es pot mediar a través de ΔFosB i la consegüent inducció de Cdk5 (vegeu el text). Aquestes alteracions en l'estructura dendrítica, que són similars a les observades en alguns models d'aprenentatge (per exemple, potenciació a llarg termini), podrien mediar respostes sensibilitzades de llarga vida a drogues d'abús o indicis ambientals. [Reproduït amb permís de la ref. 3 (Copyright 2001, Macmillian Magazines Ltd.)].
Una altra possibilitat és que l’inducció transitòria d’un factor de transcripció (per exemple, ΔFosB, CREB) condueix a canvis més permanents en l’expressió gènica mitjançant la modificació de les cromàtiquesn. Es creu que aquests i molts altres factors de transcripció activen o reprimeixen la transcripció d’un gen objectiu mitjançant la promoció de l’acetilació o desacetilació, respectivament, de les histones properes al gen (42). Tot i que tal acetilació i desacetilació de les histones pot aparèixer molt ràpidament, és possible que BFosB o CREB puguin produir adaptacions més duradores en la maquinària enzimàtica que controla la acetilació de les histones. BFosB o CREB també poden promoure canvis de llarga durada en l’expressió gènica mitjançant la regulació d’altres modificacions de la cromatina (per exemple, DNA o histil metilació) que s’han implicat en els canvis permanents en la transcripció de gens que es produeixen durant el desenvolupament (vegeu refs. 42 i 43) . Tot i que aquestes possibilitats continuen sent especulatives, podrien proporcionar un mecanisme pel qual les adaptacions transitòries a un fàrmac d’abús (o alguna altra pertorbació) condueixen a conseqüències conductuals essencials durant tota la vida.
referències
- ↵
- Nestler EJ,
- Hope BT,
- Widnell KL
(1993) Neuron 11: 995 – 1006.
CrossRefMedlineWeb de la ciència
- ↵
- Berke JD,
- Hyman SE
(2000) Neuron 25: 515 – 532.
CrossRefMedlineWeb de la ciència
- ↵
- Nestler EJ
(2001) Nat Rev Neurosci 2: 119 – 128.
CrossRefMedlineWeb de la ciència
- ↵
- Carlezon WA Jr,
- Thome J,
- Olson VG,
- Lane-Ladd SB,
- Brodkin ES,
- Hiroi N,
- Duman RS,
- Neve RL,
- Nestler EJ
(1998) Ciència 282: 2272 – 2275.
Text complet abstracte / GRATUÏT
- O'Donovan KJ,
- GT de Tourtellotte,
- Millbrandt J,
- Baraban JM
(1999) Tendències Neurosci 22: 167 – 173.
CrossRefMedlineWeb de la ciència
- ↵
- Mackler SA,
- Korutla L,
- Cha XY,
- Koebbe MJ,
- Fournier KM,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210 – 6217.
Text complet abstracte / GRATUÏT
- ↵
- Morgan JI,
- Curran T
(1995) Tendències Neurosci 18: 66 – 67.
CrossRefMedlineWeb de la ciència
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci EUA 88: 1291 – 1295.
Text complet abstracte / GRATUÏT
- Graybiel AM,
- Moratalla R,
- Robertson HA
(1990) Proc Natl Acad Sci EUA 87: 6912 – 6916.
Text complet abstracte / GRATUÏT
- Hope B,
- Kosofsky B,
- Hyman SE,
- Nestler EJ
(1992) Proc Natl Acad Sci EUA 89: 5764 – 5768.
Text complet abstracte / GRATUÏT
- ↵
- Kelz MB,
- Nestler EJ
(2000) Curr Opin Neurol 13: 715 – 720.
CrossRefMedlineWeb de la ciència
- ↵
- Koob GF,
- Sanna PP,
- Bloom FE
(1998) Neuron 21: 467 – 476.
CrossRefMedlineWeb de la ciència
- ↵
- RA sàvia
(1998) Dependència de l’alcohol de drogues.
CrossRefMedlineWeb de la ciència
- ↵
- Hope BT,
- Nye HE,
- Kelz MB,
- Self DW,
- Iadarola MJ,
- Nakabeppu I,
- Duman RS,
- Nestler EJ
(1994) Neuron 13: 1235 – 1244.
CrossRefMedlineWeb de la ciència
- ↵
- Nye H,
- Hope BT,
- Kelz M,
- Iadarola M,
- Nestler EJ
(1995) J Pharmacol Exp Ther 275: 1671 – 1680.
Text complet abstracte / GRATUÏT
- Nye HE,
- Nestler EJ
(1996) Mol Pharmacol 49: 636 – 645.
- ↵
- Moratalla R,
- Elibol B,
- Vallejo M,
- Graybiel AM
(1996) Neuron 17: 147 – 156.
CrossRefMedlineWeb de la ciència
- ↵
- Pich EM,
- Pagliusi SR,
- Tessari M,
- Talabot-Ayer D,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) Ciència 275: 83 – 86.
Text complet abstracte / GRATUÏT
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Hiroi N,
- Nakabeppu I,
- Hope BT,
- Nestler EJ
(1995) Mol Pharmacol 48: 880 – 889.
- ↵
- Hiroi N,
- Brown J,
- Ye H,
- Saudou F,
- Vaidya VA,
- Duman RS,
- Greenberg ME,
- Nestler EJ
(1998) J Neurosci 18: 6952 – 6962.
Text complet abstracte / GRATUÏT
- ↵
- Chen J,
- Kelz MB,
- Hope BT,
- Nakabeppu I,
- Nestler EJ
(1997) J Neurosci 17: 4933 – 4941.
Text complet abstracte / GRATUÏT
- ↵
- Hiroi N,
- Brown J,
- Haile C,
- Ye H,
- Greenberg ME,
- Nestler EJ
(1997) Proc Natl Acad Sci EUA 94: 10397 – 10402.
Text complet abstracte / GRATUÏT
- ↵
- Fienberg AA,
- Hiroi N,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Nishi A,
- Cheramy A,
- O'Callaghan JP,
- Miller D,
- Cole DG,
- et al.
(1998) Ciència 281: 838 – 842.
Text complet abstracte / GRATUÏT
- ↵
- Hiroi N,
- Feinberg A,
- Haile C,
- Greengard P,
- Nestler EJ
(1999) Eur J Neurosci 11: 1114 – 1118.
CrossRefMedlineWeb de la ciència
- ↵
- Greengard P,
- Allen PB,
- Nairn AC
(1999) Neuron 23: 435 – 447.
CrossRefMedlineWeb de la ciència
- ↵
- JA Bibb,
- Snyder GL,
- Nishi A,
- Yan Z,
- Meijer L,
- Fienberg AA,
- Tsai LH,
- Kwon YT,
- JA Girault,
- Czernik AJ,
- et al.
(1999) Nature (Londres) 402: 669 – 671.
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Sakai N,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Duman RS,
- Nestler EJ
(1998) Mol Pharmacol 54: 495 – 503.
Text complet abstracte / GRATUÏT
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Whisler K,
- Gilden L,
- Beckmann AM,
- Steffen C,
- Zhang YJ,
- Marotti L,
- Self SW,
- et al.
(1999) Nature (Londres) 401: 272 – 276.
- ↵
- Dobrazanski P,
- Noguchi T,
- Kovary K,
- Rizzo CA,
- Lazo PS,
- Bravo R
(1991) Mol Cell Biol 11: 5470 – 5478.
Text complet abstracte / GRATUÏT
- Nakabeppu I,
- Nathans D
(1991) Cel·la 64: 751 – 759.
CrossRefMedlineWeb de la ciència
- ↵
- Yen J,
- Saviesa RM,
- Tratner I,
- Verma IM
(1991) Proc Natl Acad Sci EUA 88: 5077 – 5081.
Text complet abstracte / GRATUÏT
- ↵
- White FJ,
- Hu XT,
- Zhang XF,
- Wolf ME
(1995) J Pharmacol Exp Ther 273: 445 – 454.
Text complet abstracte / GRATUÏT
- ↵
- Hyman SE
(1996) Neuron 16: 901 – 904.
CrossRefMedlineWeb de la ciència
- Kreek MJ
(1997) Pharmacol Biochem Behav 57: 551 – 569.
CrossRefMedlineWeb de la ciència
- ↵
- Shippenberg TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449 – 455.
CrossRefMedlineWeb de la ciència
- ↵
- Chen JS,
- Zhang YJ,
- Kelz MB,
- Steffen C,
- Ang ES,
- Zeng L,
- Nestler EJ
(2000) J Neurosci 20: 8965 – 8971.
Text complet abstracte / GRATUÏT
- ↵
- JA Bibb,
- Chen JS,
- Taylor JR,
- Svenningsson P,
- Nishi A,
- Snyder GL,
- Yan Z,
- Sagawa ZK,
- Nairn AC,
- Nestler EJ,
- et al.
(2001) Nature (Londres) 410: 376 – 380.
- ↵
- Luscher C,
- Nicoll RA,
- Malenka RC,
- Muller D
(2000) Nat Neurosci 3: 545 – 550.
CrossRefMedlineWeb de la ciència
- Malinow R,
- Mainen ZF,
- Hayashi Y
(2000) Curr Opin Neurobiol 10: 352 – 357.
CrossRefMedlineWeb de la ciència
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133 – 141.
CrossRefMedlineWeb de la ciència
Robinson, TE i Kolb, B. (1999) (1997) Eur. J. Neurosci.11, 1598-1604.
- ↵
- Carey M,
- Smale ST
(2000) Regulació transcripcional en eucariotes (Cold Spring Harbor Lab. Press, Plainview, NY).
- ↵
- Spencer VA,
- Davie JR
(1999) Gen 240: 1 – 12.
CrossRefMedlineWeb de la ciència
 Facebook
Facebook- Twitter
- Google+
- CiteULike
- deliciós
 Digg
Digg- Mendeley
Articles publicats per HighWire Press que citen aquest article
- Llei de premis naturals i medicaments sobre mecanismes comuns de plasticitat neuronal amb {Delta} FosB com a mediador clau J. Neurosci. 2013 33 (8) 3434-3442
- Drogues, delictes i l'epigenètica de l'alostasis hedònica Journal of Contemporary Criminal Justice 2012 28 (3) 314-328
- abstracte
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- abstracte
- Text complet (HTML)
- Text complet (PDF)
- La morfina activa el factor E de vint-i-sis factors de transcripció: el factor 1 / factor de resposta de sèrum a través de quinases regulades per senyal extracel·lular 1 / 2 a les cèl·lules F11 derivades de les neurones dels ganglis de l'arrel dorsal J. Pharmacol. Exp. Hola. 2012 342 (1) 41-52
- Mecanisme molecular per a una droga de passarel·la: canvis epigenètics iniciats per l'expressió gènica primària de la nicotina per cocaïna Sci Transl Med 2011 3 (107) 107ra109
- Auto-administració de sacarosa i cocaïna millorada i medicament induït per induir després de la pèrdua de VGLUT2 a les neurones de dopamina del cervell mig en ratolins J. Neurosci. 2011 31 (35) 12593-12603
- La hipòxia crònica intermitent augmenta la pressió arterial i l'expressió de FosB / {Delta} FosB a les regions autònomes centrals Am. J. Physiol. Regul. Integració. Comp. Physiol. 2011 301 (1) R131-R139
- L’absència del receptor GPR37 / PAEL perjudica la fosforilació estratal Akt i ERK2, l’expressió {Delta} FosB i la preferència del lloc condicionada a l’anfetamina i la cocaïna FASEB J. 2011 25 (6) 2071-2081
- La relació entre la durada de l'exposició inicial a l'alcohol i la persistència de la tolerància molecular és marcadament no lineal J. Neurosci. 2011 31 (7) 2436-2446
- La producció d’imatges bioluminiscents In Vivo revela l’activació de l’activador de proteïna-1 amb regulació redox en un nucli paraventricular de ratolins amb hipertensió renovascular Hipertensió 2011 57 (2) 289-297
- La sobreexpressió estriatal de {Delta} FosB reprodueix moviments involuntaris crònics inducits per levodopa J. Neurosci. 2010 30 (21) 7335-7343
- Mediació epigenètica de les influències ambientals en els trastorns psicòtics majors Schizophr Bull 2009 35 (6) 1045-1056
- Sondes MRI basades en ADN per a la detecció específica de l'exposició crònica a l'anfetamina en cervells vius J. Neurosci. 2009 29 (34) 10663-10670
- Plasticitat de la columna vertebral dendrítica alterada en rates de retirada de cocaïna J. Neurosci. 2009 29 (9) 2876-2884
- La pantalla de sobreexpressió a Drosophila identifica les funcions neuronals de GSK-3 {beta} / shaggy com a regulador de la plasticitat de desenvolupament dependent de AP-1 Genètica 2008 180 (4) 2057-2071
- RM de la transcripció: una nova visió del cervell viu Neurocientífic 2008 14 (5) 503-520
- {Delta} Inducció de FosB a la Còrtex Orbitofrontal, que media la tolerància a la disfunció cognitiva induïda per la cocaïna J. Neurosci. 2007 27 (39) 10497-10507
- Vulnerabilitat persistent davant la reincorporació del comportament que busca la metamfetamina en ratolins mutants del factor neurotròfic derivat de la línia cel·lular glial. FASEB J. 2007 21 (9) 1994-2004
- {Delta} FosB al nucli Accumbens regula el comportament i la motivació instrumentals reforçats pels aliments J. Neurosci. 2006 26 (36) 9196-9204
- Regulació de l'estabilitat {Delta} per FosB per fosforilació. J. Neurosci. 2006 26 (19) 5131-5142
- Expressió de receptors mutants de NMDA en dopamina Les cèl·lules que contenen receptors D1 impedeixen la sensibilització per la cocaïna i disminueix la preferència de cocaïna J. Neurosci. 2005 25 (28) 6651-6657
- Els receptors de dopamina D1 modifiquen {Delta} la inducció de FosB en estriat de rata després de l'administració intermitent de morfina J. Pharmacol. Exp. Hola. 2005 314 (1) 148-154
- Neurobiologia de ratolins seleccionats per a una activitat voluntària d'alta roda Integració. Comp. Biol. 2005 45 (3) 438-455
- Efectes de la privació d’aigua i la seva rehidratació en la tinció de c-Fos i FosB al nucli supraàctic de la rata i la regió de la lamina Am. J. Physiol. Regul. Integració. Comp. Physiol. 2005 288 (1) R311-R321
- Inducció transcripcional del gen FosB / {Delta} FosB per estrès mecànic en osteoblasts J Biol Chem 2004 279 (48) 49795-49803
- Inducció de {Delta} FosB en Estructures cerebrals relacionades amb la recompensa després de l'estrès crònic J. Neurosci. 2004 24 (47) 10594-10602
- La dosificació genètica Sim1 modula la resposta alimentària homeostàtica davant un augment del greix dietètic en ratolins Am. J. Physiol. Endocrinol. Metab. 2004 287 (1) E105-E113
- Anàlisi de microarrays d’ADN d’expressió gènica en astròcits de cap del nervi òptic humà en resposta a la pressió hidrostàtica Physiol. Genòmica 2004 17 (2) 157-169
- El superóxido està involucrat en l’activació del sistema nerviós central i la simpatoexcitació del fracàs cardíac induït per infarto miocardi Circ. Res. 2004 94 (3) 402-409
- Receptors d’adenosina A2A en la neuroadaptació a l’estimulació dopaminèrgica repetida: implicacions per al tractament de les discinesies en la malaltia de Parkinson Neurologia 2003 61 (90116) S74-81
- Localització citoplasmàtica contra localització nuclear de proteïnes relacionades amb Fos a la granota, rana esculenta, testicles: efecte in vivo i directe en viu d’un agonista hormonal alliberador de gonadotropina Biol. Reprod. 2003 68 (3) 954-960
- Els ratolins periadolescents mostren una major regulació del FosB Delta en resposta a la cocaïna i l'anfetamina J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB regula el funcionament de la roda J. Neurosci. 2002 22 (18) 8133-8138
- L’activitat del CREB al nucli accumbens controla l’enllaç de les respostes conductuals als estímuls emocionals Proc. Natl. Acad. Sci. Estats Units 2002 99 (17) 11435-11440
- Psicogenòmica: oportunitats per entendre les addiccions J. Neurosci. 2001 21 (21) 8324-8327
