YBOP KOMENTARI: Učinak Coolidge stoji iza snage internetskog pornića. Coolidge efekt je pojava koja se primjećuje kod vrsta sisavaca kod kojih mužjaci (a u manjoj mjeri i ženke) pokazuju obnovljeni seksualni interes ako su predstavljeni novim prijemčivim seksualnim partnerima, čak i nakon što su odbili seks od ranije, ali još uvijek dostupnih seksualnih partnera. Seksualna novost nadjačava ovu naviku s ponovnim uzbuđenjem uzrokovanim većim dopaminom. Stalni niz novosti ono je što internetske porniće čini toliko drugačijim od porno prošlosti.
- Dennis F. Fiorino,
- Ariane Coury i
- Anthony G. Phillips
- Journal of Neuroscience, 15. lipnja 1997., 17 (12): 4849-4855;
Sažetak
Coolidge efekt opisuje ponovno pokretanje seksualnog ponašanja u “seksualno nasitenoj” životinji kao odgovor na novi receptivni partner. S obzirom na ulogu mezolimbičkog dopaminskog (DA) sustava u pokretanju i održavanju motiviranog ponašanja, mikrodijalizom je korišten nadzor prijenosa nucleus accumbens (NAC) DA tijekom kopulacije, seksualne sitosti i ponovnog pokretanja seksualnog ponašanja. U skladu s ranijim izvješćima, prezentacija estrusne žene iza ekrana i kopulacija bili su povezani sa značajnim povećanjem izlučivanja NAC DA. Povratak koncentracija NAC DA na početne vrijednosti podudara se s razdobljem seksualne sitosti, iako su koncentracije DA metabolita, dihidroksifeniloctene kiseline i homovanillic kiseline ostale visoke. Prikazivanje nove receptivne ženke iza ekrana rezultiralo je blagim povećanjem NAC DA, što je značajno povećano tijekom ponovnog kopuliranja s novom ženkom. Ovi podaci ukazuju na to da svojstva stimulusa nove receptivne ženke mogu služiti za povećanje prijenosa NAC DA kod seksualno zasićenih mužjaka štakora, a to može biti povezano s ponovnim pokretanjem seksualnog ponašanja.
Uvod
Mužjski štakor koji se kopulirao do sitosti može se potaknuti na ponovno parenje ako se početna ženka zamijeni novom receptivnom ženkom. To je postalo poznato kao Coolidge efekt i uočeno je u brojnim vrstama sisavaca (Wilson i sur., 1963). Opći čimbenici kao što su umor ili motorička depresija nisu dovoljni da objasne prividno stanje seksualnog zasićenja, jer podražaji iz nove ženke još uvijek mogu inducirati kopulaciju. Seksualna sitosti također se mogu farmakološki "preokrenuti", u značajnoj mjeri, primjenom različitih lijekova koji mogu djelovati na različite neurotransmiterske sustave. Ovi lijekovi uključuju yohimbine, 8-OH-DPAT (Rodriguez-Manzo i Fernandez-Guasti, 1994, 1995a), nalakson (Pfaus i Gorzalka, 1987; Rodriguez-Manzo i Fernandez-Guasti, 1995a,b) i apomorfin (Mas i sur., 1995c). Iako se periferno djelovanje tih lijekova ne može isključiti (npr. Adrenergički učinci na erektilnu funkciju), učinci na središnje mehanizme koji podupiru seksualnu sitosti su predloženi na temelju selektivnih eksperimenata središnje noradrenergičke lezije (Rodriguez-Manzo i Fernandez-Guasti, 1995ai eksperimenti mikrodijalize koji su pratili dopaminergički metabolizam u medijalnom predoptičkom području (Mas i sur., 1995a,b).
Budući da središnji mehanizmi mogu posredovati u ponovnom pokretanju seksualnog ponašanja karakterističnog za Coolidge efekt, vjerojatni kandidat je mezolimbički sustav dopamina (DA), projektirajući se iz ventralnog tegmentalnog područja u NAC. Čini se da mezolimbski DA djeluje kao primarni modulator u složenim integrativnim procesima koji uključuju procjenu utjecaja na okoliš, kao što su znakovi seksualno receptivne žene, i organizacija ciljanog ponašanja, uključujući kopulaciju (Fibiger i Phillips, 1986; Blackburn i sur., 1992; Phillips i sur., 1992; LeMoal, 1995; Salamone, 1996).
Iako neuronski DA neuroni reagiraju na primarne nagrade i znakove koji predviđaju nagrađivanje, novi ili nepredvidivi stimulansi okoline potiču neuronsku aktivaciju najudobljivije tijekom ponovljenih treninga (Fabre i sur., 1983; Schultz, 1992; Mirenowicz i Schultz, 1994). Postoji mnogo dokaza koji podupiru važnu olakšavajuću ulogu mezolimbičkog DA u pokretanju i održavanju seksualnog ponašanja štakora (Pfaus i Everitt, 1995), a brojna istraživanja mikrodijalize izvješćuju o povećanju izlučivanja NAC DA tijekom privlačnih i konzumacijskih faza muškog seksualnog ponašanja (Pfaus i sur., 1990; Pleim i sur., 1990; Damsma i sur., 1992; Wenkstern i sur., 1993; Fumero i sur., 1994; Mas i sur., 1995b). Međutim, postoji relativno malo podataka o neurokemijskim korelacijama seksualnog zasićenja i ponovnog pokretanja seksualnog ponašanja. Primjena in vivo mikrodijaliza za praćenje neurotransmisije mezolimbičkog DA tijekom Coolidge efekta pruža jedinstvenu priliku za ispitivanje uloge NAC DA u kopulaciji, seksualnoj sitosti i ponovnom pokretanju kopulacije.
Proveden je pokus mikrodijalize kako bi se utvrdilo sljedeće: (1) da li je početak seksualne sitosti popraćen povratkom izvanstaničnih koncentracija DA u NAC na vrijednosti predpulacije ili niže, i (2) je li ponovno uspostavljanje ponašanja kopulatora u muškog štakora s novom receptivnom ženkom korelira s povećanjem izlučivanja NAC DA.
MATERIJALI I METODE
Subjekti. Mužjaci štakora Sprague Dawley, dobiveni od Centra za skrb o životinjama (University of British Columbia), i ženki štakora Long-Evans, dobiveni od Charles River Canada (St. Constant, Quebec, Kanada), bili su smješteni u kaveze od žičane mreže (18) × 25 × 65 cm, pet po kavezu) u odvojenim sobama za kolonije. Sobe za kolonije održavane su na temperaturi od N20 ° C na obrnutom ciklusu svjetla / mraka 12 hr. Štakori su imali neograničen pristup hrani (Purina Rat Chow) i vodi.
Kirurgija i testiranje ponašanja prije mikrodijalize u mozgu.Ženke štakora bile su ovariektomirane bilateralno pod halotanskom plinskom anestezijom (Fluothane, Ayerst Laboratories) najmanje 4 tjedana prije testiranja. Seksualna receptivnost u ženki podražaja inducirana je potkožnim injekcijama estradiol benzoata (10 μg) i progesterona (500 μg), odnosno 48 i 4 hr, prije svakog ispitivanja. Mužjaci štakora su ispitivani na seksualno ponašanje u dva navrata, 4 d apart, u komorama od pleksiglasa (35 × 35 × 40 cm) s podovima od žičane mreže. Samo muški štakori koji su dostigli kriterij izvedbe, koji je uključivao intromisiju unutar 5 min prezentacije ženke i ejakulaciju unutar 15 min prve intromicije, tijekom dva testa probira su implantirani s mikrodijalizirajućom sondom vodičima za ispitivanje.
Muški štakori (n = 5) anestezirani su ketamin hidrokloridom (100 mg / kg, ip) i ksilazinom (10 mg / kg, ip) prije stereotaksijske kirurgije. Voditeljska kanila s mikrodijalizirajućom sondom (mjerač 19) implantirana je bilateralno preko NAC-a (koordinate iz bregme: anterior, + 1.7 mm; medijska, ± 1.1 mm; ventralna, -1.0 mm; ravna lubanja) i pričvršćena na lubanju zubnim akrilima i zlatarske vijke. Bilateralni vodiči kanile korišteni su kako bi se povećala mogućnost za uspješan mikrodijalizni eksperiment. Srećom, u ovom eksperimentu, potrebna je samo jedna kanila za svakog štakora. Mužjaci štakora su smješteni pojedinačno u velike plastične kaveze s podlogom od kukuruza za ostatak pokusa. Tjedan dana nakon operacije, štakori su testirani na seksualno ponašanje. Tijekom ovog dijela treninga, komora za testiranje bila je opremljena kliznim ekranom od pleksiglasa koji je komoru podijelio na velike i male odjeljke. Mužjaci štakora su uvedeni u veliki odjeljak, a 15 min kasnije, ženka je stavljena iza ekrana. Nakon pripremnog perioda od 15 min, sito je uklonjeno, a štakorima je dopušteno da se kopuliraju za 30 min. Provedena su tri treninga, jedan svaki 4 d. Svi štakori postigli su kriterij uspješnosti tijekom svakog tretmana.
Eksperiment Coolidge efekta. Štakori su unilateralno implantirani pomoću mikrodijaliznih sondi 12-18 h prije eksperimenta Coolidge efekta i stavljeni u veliki odjeljak ispitne komore uz slobodan pristup hrani i vodi. Ujutro eksperimenta, uzorci mikrodijalize su sakupljeni svaki 15 min. Eksperiment se sastojao od sljedećih sedam uzastopnih faza: (1) bazne linije (najmanje 60 min); (2) ženski 1 iza zaslona (15 min); (3) kopulacija sa ženskim 1-om sve do min. 30 min. (4) ponovno uvođenje ženskog 1-a iza zaslona (15 min); (5) pristup ženskom 1-u tijekom 15 min perioda pod uvjetom da nije bilo montiranja (ako je došlo do montaže, ova faza je tretirana kao faza 3); (6) uvođenje ženskog 2-a iza zaslona (15 min); 7) kopulacija sa ženskim 2 za 60 min.
Ponašanje je snimljeno pod slabim osvjetljenjem pomoću JVC video sustava i promatrano na video monitoru koji se nalazio izvan sobe za testiranje. Standardne mjere seksualnog ponašanja zabilježene su pomoću računala i odgovarajućeg softvera (Holmes i dr., 1987).
Nakon eksperimenta mikrodijalize, životinjama je dano predoziranje kloral hidrata i perfundirano intracardialno s fiziološkom otopinom i formalinom (4%). Mozgovi su narezani i zamrznuti, a zatim su koronarni rezovi obojeni krezil ljubičastom bojom kako bi se odredilo postavljanje mikrodijaliznih sondi. Samo su štakori s položajem sondi unutar NAC-a korišteni za bihevioralne i neurokemijske analize.
Mikrodijaliza i HPLC-elektrokemijska detekcija. Sonde za mikrodijalizu bile su koncentrične u izvedbi s polupropusnom membranom od šupljih vlakana (2 mm membrana izložena, 340 μm vanjski promjer, 65000 granična molekulska masa, Filtral 12, Hospal) na distalnom kraju. Sonde su perfundirane na 1.0 μl / min s modificiranom Ringerovom otopinom (0.01 m natrijev fosfatni pufer, pH 7.4, 1.3 mmCaCl23.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) upotrebom štrcaljke za nepropusnost (Hamilton, Reno, NV) i pumpe za injekcije (model 22, Harvard Apparatus, South Natick, MA). Za pričvršćivanje sonde za mikrodijalizu unutar vodične kanile upotrijebljen je vodilica sondi za mikrodijalizu. Za zaštitu cijevi sonde upotrijebljen je čelični svitak, pričvršćen na obrtnu tekućinu (Instech 375) koja je montirana na vrhu ispitne komore.Fiorino et al., 1993).
Mikrodializatni analiti, koji su uključivali DA i njegove metabolite dihiroksifeniloctene kiseline (DOPAC) i homovaniličnu kiselinu (HVA), odvojeni su kromatografijom obrnute faze (kolona Ultrasphere; Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, unutarnji promjer) ) upotrebom 0.083m natrij-acetatnog pufera, pH 3.5 (5% metanol). Koncentracije analita kvantificirane su elektrokemijskom (EC) detekcijom. Uređaj se sastojao od pumpe Bio-Rad (Richmond, CA), instrumenta Valco Instruments (Houston, TX) EC10W dvo-položajnog injektora, ESA (Bedford, MA) Coulochem II EC detektora i dvo-kanalnog snimača dijagrama (Kipp). i Zonen, Bohemia, NY). Parametri elektrokemijskog detektora su sljedeći: elektroda 1, + 450 mV; elektroda 2, -300 mV; i zaštitna stanica, -450 mV. Provedena tipična oporavka sonde vitro i na sobnoj temperaturi, bili su 22% za DA, 18% za DOPAC, i 18% za HVA.
REZULTATI
Ponašanje
Mjere ponašanja iz eksperimenta Coolidge efekta prikazane su u tablici 1, Latencije za montiranje, intromit i ejakulaciju, kao i postejaculacijski interval nakon prve ejakulacije bile su slične onima na prethodnom treningu (podaci nisu prikazani). To sugerira da postupak mikrodijalize nije promijenio normalno seksualno ponašanje. Razvoj seksualnog zasićenja, što pokazuje srednji broj ejakulacija prije nego je zadovoljen kriterij (7.8 ± 0.5), progresivno smanjenje broja intromisija prije svake ejakulacije i progresivno povećanje postejakulacijskog intervala (podaci nisu prikazani) , bio je sličan onom iz prethodnih studija (Plaža i Jordan, 1956; Fowler i Whalen, 1961; Fisher, 1962; Bermant i sur., 1966; Rodriguez-Manzo i Fernandez-Guasti, 1994; Mas i sur., 1995d). Uočena je individualna varijabilnost s obzirom na broj ejakulacija postignutih ženskim 1-om, vrijeme provedeno u kopulaciji sa ženskim 1-om i broj prezentacija ženskog 1-a potrebnih za postizanje kriterija zasićenja (tablica). 1, dolje). Neki su štakori zahtijevali brojne reintrodukcije ženskog 1-a do završetka faze 5 (n = 3). Postupci stavljanja ženskog 1-a iza zaslona i uklanjanje particije mogli su poslužiti kao primarni apetitivni znakovi koji dovode do kopulacije. Također treba napomenuti da je kriterij zasićenosti 30 min bez grebena, iako je prethodno korišten (Beach i Jordan, 1965; Mas i sur., 1995b), je proizvoljan i ne jamči da štakor ne bi imao više vremena. Ipak, kašnjenja ili postupci uklanjanja i zamjene nisu pouzdano rezultirali ponovnom kopulacijom sa ženskim 1-om (npr. Faze 4 i 5).

Ponašanje tijekom eksperimenta Coolidge efekta
Svi štakori su pokazali Coolidge efekt. Aktivnost povezana sa stavljanjem ženskog 2-a iza ekrana i, posebno, uklanjanje particije može pridonijeti tom rezultatu, ali opet, ti događaji sami po sebi nisu bili dovoljni za obnovu kopulacije ranije u eksperimentu. Koriste se usporedbe mjera seksualnog ponašanja sa ženskim 1 i ženskim 2 t testovi s korekcijom Bonferronija. Premda latencije za montažu i intromiciju u odgovoru na ženski 2 nisu se značajno razlikovale od onih u prvom kopulacijskom napadu sa ženskim 1-om, općenito, seksualno ponašanje sa ženskim 2-om bilo je manje robusno, kao što pokazuje značajno manje ejakulacija (srednja vrijednost 0.6 vs 4.2; F = 49.86;p <0.01) i intromisije (prosjek = 11.2 vs 37.0;F = 20.17; p <0.05) tijekom prvog sata. Broj nosača u prvom satu kod ženki 1 i 2 nisu se značajno razlikovali.
Važno je napomenuti da su ženke koje se koriste tijekom dijela eksperimenta zasićenja (tj. Ženskog 1-a) i dalje pokazivale snažno proceptivno (tj. Skakanje i strelovito) i receptivno (tj. Lordozno) ponašanje tijekom cijelog trajanja njihovog kontakta s muškarcem ,
Neurochemistry
Bazalne nanomolarne koncentracije DA i njegovih metabolita u mikrodializatima, predstavljene kao prosjek ± SEM, od prva tri uzorka osnovne linije bile su: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; i HVA, 234.2 ± 49.0 (nekorigirano za oporavak sonde;n = 5). Ove vrijednosti predstavljaju 100% temeljnih rezultata.
Bihevioralno definirane točke podataka, koje odgovaraju svakoj fazi eksperimenta i zajedničke svakom štakoru, korištene su za neurokemijske analize. One su uključivale sljedeće: (1) sedam uzoraka nakon prvog uvođenja ženskog 1-a, (2) četiri uzorka koji su pratili odsutnost ponašanja s ženkama 1, i (3) pet uzoraka nakon predstavljanja ženskog 2-a. Lik 1 ilustrira promjene koncentracija DA (linijski graf, srednji) i DA metaboliti (linijski grafikon, vrh) paralelno oponašanje (bar-grafikon, dno) tijekom testa za Coolidge efekt.
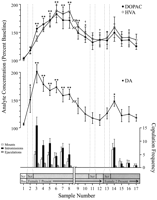
Prikaži veću verziju:
Slika. 1.
Nucleus accumbens neurokemijske korelate seksualnog ponašanja tijekom Coolidge efekta. Prvih osam uzoraka predstavljaju kronološki kontinuirane podatkovne točke od faza 1 do 3. Uzorak 1 je četvrti i posljednji uzorak preliminarne preliminarne analize (Bas). Uzorak 2 predstavlja predstavljanje ženskog 1-a iza zaslona (scr). Nakon 15 min, zaslon je uklonjen, a štakorima je dopušteno da se kopuliraju (uzorci 3-8). razbiti na x-aksija odgovara isključivanju podataka iz tri štakora koji su se kopulirali dulje vrijeme s početnom ženkom. Posljednjih devet uzoraka također je bilo kontinuirano kronološki. Uzorci 9 i 10 odgovaraju razdoblju zasićenja faze 3 (tj. 30 min bez nosača). Ženka 1 je zatim ponovno umetnuta iza zaslona (uzorak 11), a 15 min kasnije, ekran je uklonjen (uzorak 12). Nakon 15 min bez kopulacije, ženski 2 stavljen je iza zaslona (uzorak 13). Uzorci 14-17 odgovaraju kopulaciji sa ženskim 2-om. Broj nosača, intromisija ili ejakulacija povezanih sa svakim 15 min mikrodijalizom uzorak prikazan je u donji stupčasti grafikon, Neurokemijski podaci izraženi su u postocima baznih koncentracija. Promjene u NAC DA (zatvoreni kvadrati), DOPAC (zatvoreni krugovi) i HVA (otvoreni krugovi) izljev su prikazani kao linijski grafikoni, Napravljene su sljedeće usporedbe: osnovni uzorak 1 nasuprot uzoraka 2 – 10; novi osnovni uzorak 10 u odnosu na uzorke 11 i 12; novi osnovni uzorak 12 nasuprot uzoraka 13 – 17 (*p <0.05; ** p <0.01). Neovisno t provedena su ispitivanja između osnovnih vrijednosti (uzorci 1, 10 i 12). Za značajne razlike u odnosu na prvu osnovicu (uzorak 1), †p <0.05.
Odvojene jednosmjerne ANOVA-e s ponovljenim mjerama provedene su na neurokemijskim podacima vezanim uz ženski 1 (uzorci 1-12) i ženski 2 (uzorci 12-17). Apriorno usporedbe su izvršene pomoću Dunnova višestrukog usporedbenog testa (Bonferroni t). Izvršena su sljedeća tri osnovna usporedbe: (1) početna osnovica (uzorak 1) nasuprot uzoraka 2 – 10 (prva izloženost ženskom 1), (2) druga osnovica (uzorak 10) u odnosu na uzorke 11 i 12 (ponovna izloženost ženskom 1) i (3) treću polaznu vrijednost (uzorak 12) u odnosu na uzorke 13 – 17 (izloženost ženskom 2).
Došlo je do značajne ukupne promjene izlučivanja DA kao odgovor na ženski 1F (11,44) = 8.48; p <0.001] i žena 2 [F (5,20) = 2.83;p <0.05]. Značajan porast izljeva DA pronađen je kada je žena 1 bila prisutna iza zaslona (+ 44%,p <0.05; uzorak 2). Tijekom kopulacije koncentracije DA povećale su se i dosegle maksimalnu vrijednost (+ 95%;p <0.01) tijekom prvog kopulatornog napada (uzorak 3). DA je ostao povišen tijekom kopulacije i vratio se na početne koncentracije tek u razdoblju od 30 minuta u kojem nije došlo do povećanja (uzorci 9 i 10). Niti ponovno uvođenje ženke 1 iza zaslona (uzorak 11) niti mogućnost fizičke interakcije, ali bez montiranja (uzorak 12), povišene koncentracije DA u odnosu na drugu osnovnu vrijednost (uzorak 10). Prisutnost ženke 2 iza zaslona (uzorak 13) rezultirala je malim porastom izljeva DA (12%) u odnosu na treću osnovnu vrijednost (uzorak 12) koji nije dostigao statističku značajnost. Obnovljena kopulacija sa ženom 2 rezultirala je značajnim (34%) porastom (p <0.05) u izljevu DA tijekom prvog uzorka kopulacije (uzorak 14). Iako se slabo kopulatorno ponašanje nastavilo tijekom sljedeća tri uzorka, koncentracije DA smanjile su se na početne vrijednosti (uzorci 15-17). Neovisno t testovi provedeni među "osnovnim" uzorcima (tj. 1, 10 i 12) pokazali su da se te vrijednosti nisu značajno razlikovale.
Kod tri štakora koji su nastavili s kopulacijom kada je ženka 1 ponovno uvedena, koncentracije NAC DA su se povećale kada je ženski 1 bio prisutan iza zaslona (raspon, 25-47%) i tijekom kopulacije (raspon, 13 – 37%), u odnosu na uzorak prije ponovnog uvođenja ženke. Međutim, to se povećanje dogodilo samo kada je seksualno ponašanje bilo snažno i dovelo do ejakulacije.
Značajne ukupne promjene u DOPAC-uF (11,44) = 9.57; p <0.001] i HVA [F (11,44) = 12.47; p Pronađene su koncentracije <0. 001] kao odgovor na ženku 1, ali ne i na ženu 2. Koncentracije metabolita blago su porasle (+ 15% u oba slučaja) tijekom prezentacije ženke 1 iza zaslona (uzorak 2), ali to nije bilo značajno statistički. Međutim, došlo je do značajnog povećanja koncentracija DOPAC-a i HVA-e tijekom kopulacije (uzorci 3–8), dostigavši maksimalne vrijednosti (+80, odnosno + 86%; p <0.01) nakon 60 minuta (uzorak 6 u oba slučaja). Iako su se koncentracije metabolita smanjivale tijekom razdoblja spolne neaktivnosti na kraju kontakta sa ženkom 1 (uzorci 9 i 10), koncentracije su i dalje ostale povišene u odnosu na početnu vrijednost (p <0.05 u oba slučaja). Ponovno uvođenje ženke 1 iza paravana (uzorak 11), pristup ženki 1 nakon uklanjanja paravana (uzorak 12) i uvođenje ženke 2 (uzorak 13) nisu rezultirali nikakvim promjenama u koncentraciji metabolita. Lagani, ali statistički beznačajni, porast koncentracija DOPAC-a i HVA (+ 23% u oba slučaja) u odnosu na početnu liniju (uzorak 12) odgovarao je prvom napadu kopulacije sa ženkom 2 (uzorak 14). Taj je porast, međutim, bio kratkotrajan i smanjio se na početne vrijednosti za preostala tri uzorka (15-17). Neovisno ttestovi provedeni među "osnovnim" uzorcima (tj. 1, 10 i 12) pokazali su da su druga i treća osnovna vrijednost (uzorci 10 i 12, respektivno), iako se ne razlikuju međusobno, ostali značajno povišeni u usporedbi s prvim osnovnim uzorkom za DOPAC i HVA (p <0.05 u oba slučaja).
Histologija
Mikrodijalizne sonde bile su smještene u NAC-u (sl.2) u rasponu od + 1.20 do + 1.70 mm od bregme (ravna lubanja). Bilo je i varijabilnosti u mediolateralnoj ravnini; podaci odražavaju uzorkovanje iz ljuske i temeljnih podregija NAC-a.
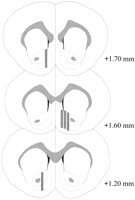
Prikaži veću verziju:
Slika. 2.
Lokacija mikrodijaliznih sondi unutar NAC mužjaka štakora korištenih u eksperimentu Coolidge efekta. Osjenčani pravokutnici odgovaraju izloženoj membranskoj površini sondi za mikrodijalizu. Serijski dijelovi koronalnog mozga su izvučeni izPaxinos i Watson (1986).
RASPRAVA
U skladu s ranijim izvješćima, sadašnji rezultati pokazuju pojačanu transmisiju mezolimbičkog DA povezanu s apetitivnim i konzumatornim komponentama spolnog ponašanja mužjaka štakora kako je procijenjenoin vivo mikrodijaliza (Mas i sur., 1990; Pfaus i sur., 1990;Pleim i sur., 1990; Damsma i sur., 1992; Wenkstern i sur., 1993; Fumero i sur., 1994; Mas i sur., 1995a,b,d). Osim toga, ovi rezultati daju neurokemijsku korelatu za seksualno zasićenje i naknadnu ponovnu inicijaciju kopulacije kao odgovor na novu receptivnu ženku (Coolidge efekt). Ovi podaci ukazuju na to da svojstva stimulusa nove receptivne ženke mogu služiti za povećanje prijenosa NAC DA kod spolno nasitenog mužjaka štakora, što može biti povezano s ponovnim pokretanjem seksualnog ponašanja. To se prvo očituje u blagom povećanju NAC DA tijekom predstavljanja nove ženke iza ekrana i najučinkovitije se javlja kao izraženiji porast tijekom ponovnog kopuliranja sa ženskim 2-om (sl.1).
Prisutnost prve receptivne ženke iza zaslona rezultirala je snažnim povećanjem apetita u NAC DA (44% u odnosu na početnu vrijednost) sličnom po veličini u odnosu na ono što je prijavljeno u prethodnim eksperimentima koristeći sličan dizajn (30%, Pfaus i sur., 1990; 35%Damsma i sur., 1992). Također u skladu s ovim studijama bilo je zapažanje da je izljev NAC DA dodatno pojačan tijekom kopulacije (na> 95% iznad početne vrijednosti u ovom eksperimentu). Iako možemo promatrati konzumatorno ponašanje kao povezano s poboljšanim oslobađanjem NAC DA (Wenkstern i sur., 1993; Wilson i sur., 1995), važno je ispitati pojmove “apetit” i “konzumator” u kontekstu seksualnog ponašanja. Dok je faza u kojoj je ženka prisutna iza ekrana isključivo privlačna ili pripremna, ponašanje tijekom faze kopulacije ne može se smatrati čisto konzumirajućim. Budući da se "appetitive" može upotrijebiti za opisivanje svih ponašanja koja dovode do konzumacije motiviranog ponašanja (kopulacija), primarno ponašanje muških eksponata dok je aktivno u fazi "konzumacije" najbolje se opisuje kao apetit; muškarac provodi većinu svog vremena i truda u potrazi za ženskom parom. U tom smislu, možemo povezati maksimalni NAC DA prijenos s konzumatorom kao i intenzivne apetitivne komponente spolnog ponašanja mužjaka štakora.
Pristup drugoj, novoj ženki rezultirao je obnovljenom kopulacijom u svakom subjektu. Prethodne studije su pokazale da je većina štakora dopuštena za snošenje, koristeći sličan protokol ponašanja u odnosu na onaj korišten u ovom eksperimentu, nije nastavio parenje kada je testiran 24 h kasnije (Plaža i Jordan, 1956). Vrlo je vjerojatno da je prisutnost novih stimulativnih svojstava ženskog 2-a, koje su možda uključivale mirisne kao i vizualne i auditivne znakove, rezultiralo ponovnom kopulacijom. Zanimljivo pitanje, na koje tek treba odgovoriti, je kojim mehanizmom mužjak štakora razlikuje novu ženku od ženke s kojom je nedavno pario. Mjesto za taj mehanizam može biti u glavnom mirisnom sustavu. Prijavljeno je da je integritet ovog sustava bitan za Coolidge efekt u hrčaka (Johnston i Rasmussen, 1984). Međutim, vomeronazalni-pomoćni mirisni sustav, u kojem je nedavno opisan proces feromonalne memorije kod miševa (Kaba i sur., 1994), također je glavni kandidat. U tom smislu, vrijedno je spomenuti da su korištena povećanja NAC DA prijenosa in vivo voltametrija kod mužjaka štakora izloženih posteljini koja je bila izložena ženkama štakora u estrusu (Louillot i sur., 1991; Mitchell i Gratton, 1992). Nadalje, primjena K+ izravno na sloj vomeronazalnog živca dodatne dodatne mirisne žarulje, kao i na samu dodatnu mirisnu žarulju, bio je dovoljan za povećanje prijenosa NAC DA (Mitchell i Gratton, 1992).
Prvi ciklus kopulacije 15 min s ženskim 2-om bio je povezan sa značajnim povećanjem NAC DA. Za razliku od ženskog 1-a, interakcija sa ženskim 2-om nije uzrokovala povećanje NAC DA iste veličine tijekom faze apetita (12%) ili konzumacije (34%). Međutim, ova mala povećanja u NAC DA dobro koreliraju sa smanjenom razinom seksualnog ponašanja prikazanog sa ženskim 2-om u usporedbi sa ženskim 1-om. Koncentracije metabolita ostale su povišene tijekom faze sitosti, što je rezultiralo novim osnovnim koncentracijama (uzorci 10 i 12) koje su bile značajno povišene od početne osnovne vrijednosti (uzorak 1).
Vremensko zaostajanje u povećanju koncentracija DOPAC i HVA tijekom kopulacije u skladu je s njihovim stvaranjem kao metabolita matičnog spoja, DA. Predloženo je da koncentracije metabolita mikrodijalize, barem za vrijeme prirodnog ponašanja koje nije farmakološki potaknuto, daju koristan indeks neuralne aktivnosti (Damsma i sur., 1992; Fumero i sur., 1994). Činjenica da su koncentracije metabolita ostale povišene čak i tijekom razdoblja seksualne neaktivnosti u ovom eksperimentu, kada su se koncentracije DA vratile na pretest početne vrijednosti, baca sumnju na ovaj prijedlog.
Trajno povišenje koncentracije metabolita DA, koje se vidi u ovom eksperimentu, odražava profil preoptičkog područja (mPOA) metabolita DA koji je primijećen u štakora prvog dana nakon što su se kopulirali do zasićenja (Mas i sur., 1995a,b). Neprekidno povišenje DOPAC i HVA koncentracija u NAC ili mPOA nije uvijek opaženo kada je razdoblje parenja trajno, mnogo kraće od vremena potrebnog za postizanje zasićenja. Na primjer, mnoge studije su pokazale da su koncentracije DOPAC-a povećane i ostale povišene tijekom kopulacije, ali su se smanjile na početne vrijednosti ubrzo nakon što je žena uklonjena (Pfaus i sur., 1990; Pleim i sur., 1990; Damsma i sur., 1992;Hull i sur., 1993; Wenkstern i sur., 1993; Hull i sur., 1995). U studiji Mas i sur. (1995b)bazalne izvanstanične koncentracije DOPAC i HVA u mPOA ostale su povišene tijekom 4 uzastopnih dana, što odgovara razdoblju seksualne neaktivnosti. Do četvrtog dana, neposredno prije nego su životinje nastavile s kopulacijom, bazalne koncentracije metabolita bile su blizu vrijednostima presatacije. Autori su usporedili uzorak neurokemijskih promjena s onima koje su viđene nakon primjene blokatora DA receptora (Zetterström i sur., 1984; Imperato i DiChiara, 1985) i sugerirali su da stanje seksualne neaktivnosti može biti posredovano oslobađanjem prolaktina, koji može djelovati kao "endogeni neuroleptik" (Mas i sur., 1995a,b,d). Jasno je da je primjena neuroleptika popraćena povećanjem koncentracije izvanstaničnog metabolita i izlučivanja DA (Zetterström i sur., 1984; Imperato i DiChiara, 1985). Nažalost, Mas i sur. (1995a,b) nisu bili u mogućnosti detektirati mPOA DA koncentracije. U ovoj studiji, koncentracije DA u NAC-u vratile su se na vrijednosti predkupulacije, dok su koncentracije DOPAC i HVA ostale visoke. Ovaj uzorak nije u skladu s ulogom endogenog neuroleptika koji djeluje u NAC-u da izazove seksualnu sitosti.
S obzirom na uključenost mezolimbičkih DA neurona u motivirano ponašanje (Fibiger i Phillips, 1986; Blackburn i sur., 1992; Kalivas i sur., 1993; LeMoal, 1995) i njihovu osjetljivost na nove ekološke podražaje (Fabre i sur., 1983; Schultz, 1992; Mirenowicz i Schultz, 1994), uočena povećanja izvanstaničnih koncentracija NAC DA kao odgovor na novu ženku u skladu su s hipotezom da je aktivnost u ovom sustavu DA važna za ponovno pokretanje seksualnog ponašanja. Osim toga, izvješća o apetitivnom i konzumacijskom povećanju prijenosa DA (Hull i sur., 1993, 1995;Mas i sur., 1995b; Sato i sur., 1995) i neuronsku aktivnost (Shimura i sur., 1994) u mPOA mužjaka štakora tijekom seksualnog ponašanja sugerira da ova struktura može također doprinijeti obnovljenoj kopulaciji koja je karakteristična za Coolidge efekt.
U skladu s općom ulogom za mezolimbički sustav DA u motiviranom ponašanju, dobro je utvrđeno da su izvanstanične koncentracije DA također povišene prije, tijekom i odmah nakon konzumacije obroka, s povratkom na početne vrijednosti X30 min kasnije (Wilson i sur., 1995). Dobro je poznato da na zasićenje izazvano hranom utječu njegova osjetilna svojstva. Ljudi i životinje odbacuju hranu na koju su bili hranjeni do sitosti i unose druge namirnice koje nisu bile pojedene (Rolls, 1986). Time se postavlja pitanje bi li se izlučivanje izvanstaničnog DA u NAC selektivno povećalo prikazom nove vrste hrane, ali ne i od hrane koja je nedavno konzumirana do sitosti na način analogan onome koji je opisan u ovoj studiji u kontekstu seksualna motivacija. Ako se potvrdi, ovaj opći odnos između senzorskih svojstava prirodnih nagrada, sitosti i mezolimbičkog prijenosa DA bi značio kritičnu ulogu ovog neuralnog sustava u regulaciji motivacijskih procesa, čiji poremećaj može dovesti do ozbiljnih poremećaja prehrane i spolne funkcije. ,
REFERENCE
- ↵
- Plaža FA,
- Jordan L
(1956) Seksualna iscrpljenost i oporavak kod mužjaka štakora. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Vremenske karakteristike Coolidgeovog učinka u suparničkom ponašanju mužjaka štakora. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Dopamin djeluje u apetitivnom i obrambenom ponašanju. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Seksualno ponašanje povećava prijenos dopamina u nucleus accumbens i striatum muških štakora: usporedba s novostima i lokomocijom. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Aktivnost neurona u ventralnoj tegmentalnoj regiji majmuna koji se ponaša. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Nagrada, motivacija, spoznaja: psihobiologija mezotelencefalnih dopaminskih sustava. u Priručniku za fiziologiju: živčani sustav IV, ed. Bloom FE, Geiger SD (Američko društvo za fiziologiju, Bethesda, MD), str. 647 – 675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Električna stimulacija mjesta nagrađivanja u ventralnom tegmentalnom području povećava prijenos dopamina u nucleus accumbens štakora. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Učinci varijacije podražaja na spolno zasićenje kod mužjaka štakora. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Varijacija poticajnog stimulusa i seksualnog ponašanja kod mužjaka štakora. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Promjene u prometu monoamina u područjima prednjeg mozga koja su povezana s seksualnim ponašanjem muškaraca: istraživanje mikrodijalize. Brain Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) IBM-PC sustav za prikupljanje podataka za snimanje seksualnog ponašanja glodavaca i za snimanje općih događaja. Physiol Behav 44: 825-828.
- ↵
- Trup EM,
- Eaton RC,
- Mojsije J,
- Lorrain DS
(1993) Kopulacija povećava aktivnost dopamina u medijalnom predoptičkom području mužjaka štakora. Život Sci 52: 935-940.
- ↵
- Trup EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Ekstracelularni dopamin u medijalnom predoptičkom području: implikacije za seksualnu motivaciju i hormonsku kontrolu kopulacije. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Oslobađanje i metabolizam dopamina u štakorima nakon buđenja nakon sistemskih neuroleptika koji su proučavani trans-striatalnom dijalizom. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Individualno prepoznavanje ženki hrčaka od strane mužjaka: uloga kemijskih znakova i mirisnih i vomeronasalnih sustava. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Indukcija olfaktorne memorije aktivacijom metabotropnog receptora glutamata. Znanost 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Kuke MS
(1993) Farmakologija i neuronska kola osjetljivosti na psihostimulanse. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Mezokortikolimbički dopaminergički neuroni. Funkcionalne i regulatorne uloge. u Psihofarmakologiji: četvrta generacija napretka, eds Bloom FE, Kupfer DJ (Raven, New York), str. 283 – 294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Spolni olfaktorni stimulusi izazivaju selektivno povećanje oslobađanja dopamina u nukleusu accumbens muških štakora. Brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Jedini C,
- Guadalupe T
(1990) Povećano oslobađanje dopamina u nukleusu accumbens kopulirajućih mužjaka štakora, što je dokazano voltametrijom in vivo. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Neurokemijske korelacije spolne iscrpljenosti i oporavka procjenjene su mikrodijalizom in vivo. Brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Voltametrijsko i mikrodijalizno praćenje otpuštanja monoaminskih neurotransmitera u mozgu tijekom socioseksualnih interakcija. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Indukcija ponašanja parjenja apomorfinom u spolno nasutih štakora. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
Neurokemija seksualne sitosti. Eksperimentalni model inhibirane želje. u Farmakologija seksualne funkcije i disfunkcije, ed Bancroft J (Raven, New York), str. 1995 – 115.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Važnost nepredvidljivosti odgovora na nadoknadu kod primatnih dopaminergičkih neurona. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Mezolimbičko oslobađanje dopamina izazvano je aktivacijom pomoćnog mirisnog sustava: kronoamperometrijska studija velike brzine. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) Mozak štakora u stereotaksičnim koordinatama (2nd ed). (Akademski, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Seksualno ponašanje povećava središnji prijenos dopamina kod mužjaka štakora. Brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Psihofarmakologija seksualnog ponašanja. u Psihofarmakologiji: četvrta generacija napretka, eds Bloom FE, Kupfer DJ (Raven, New York), str. 743 – 758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioidi i seksualno ponašanje. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Neurobiološki korelati pozitivnih emocionalnih stanja: dopamin, anticipacija i nagrada. u Međunarodnom pregledu studija o emocijama, ed Strongman (Wiley, New York), str. 31 – 50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Korelacija otpuštanja dopamina u nucleus accumbens s muškim seksualnim ponašanjem u štakora. Brain Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Ukidanje spolne iscrpljenosti serotonergičkim i noradrenergičkim sredstvima. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Sudjelovanje centralnog noradrenergičkog sustava u ponovnom uspostavljanju kopulacijskog ponašanja seksualno iscrpljenih štakora od strane yohimbina, naloksona i 8-OH-DPAT. Brain Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Opioidni antagonisti i fenomen seksualne sitosti. Psycho 122: 131-136.
- ↵
- Rolls BJ
(1986) Osjetilna specifičnost. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) Neurokemija ponašanja motivacije: metodološka i konceptualna pitanja u istraživanju dinamičke aktivnosti nucleus accumbens dopamina. J Neurosci metode 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Oslobađanje dopamina u medijalnom predoptičkom području tijekom muškog oponašanja u štakora. Brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Aktivnost dopaminskih neurona u primata koji se ponaša. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) Medijalno preoptičko područje uključeno je u spolno uzbuđenje i učinak kod mužjaka štakora: ponovna procjena neuronske aktivnosti u slobodno pokretnim životinjama. Brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Prijenos dopamina povećava se u nukleusu accumbens mužjaka štakora tijekom njihove prve izloženosti seksualno prijemljivim ženskim štakorima. Brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Dopaminergički korelati motiviranog ponašanja: važnost pogona. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Plaža FA
(1963) Modifikacija seksualnog ponašanja mužjaka štakora proizvedena promjenom stimulusa ženke. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Učinak neuroleptičkih lijekova na otpuštanje i metabolizam dopijuma u striatima kod budnih štakora proučenih intracerebralnom dijalizom. Eur J Pharmacol 106: 27-37.
Članci navode ovaj članak
- Endogena opioidno inducirana neuroplastičnost dopaminergičkih neurona u području ventralnog tegmentala utječe na prirodnu i nagradu za opijate Časopis za neuroznanost, 25 lipanj 2014, 34 (26): 8825-8836
- Neravnoteža u osjetljivosti na različite vrste nagrada u patološkom kockanju Mozak, 1 kolovoz 2013, 136 (8): 2527-2538
- Zakon o nagradama za prirodne i lijekove o zajedničkim mehanizmima neuralne plastičnosti s {Delta} FosB kao ključnim posrednikom Časopis za neuroznanost, 20 veljača 2013, 33 (8): 3434-3442
- Neuronski odgovori u ljusci nuklearnih naslaga tijekom seksualnog ponašanja u mužjaka štakora Časopis za neuroznanost, 1 veljača 2012, 32 (5): 1672-1686
- Romantična ljubav: moždani sustav sisavaca za izbor partnera Filozofske transakcije Kraljevskog društva B: Biološke znanosti, 29 prosinac 2006, 361 (1476): 2173-2186
- Hedonske vruće točke u mozgu Neuroznanstvenik, 1 prosinac 2006, 12 (6): 500-511
- Modulacija središnjim i bazolateralnim amigdalarnim jezgrama dopaminergičkih korelata hranjenja do zasićenosti u jezgrama štakora i medijalnom prefrontalnom korteksom Časopis za neuroznanost, 15 prosinac 2002, 22 (24): 10958-10965
- Učestalost prijelaznih koncentracija dopamina Povećanje dorzalnog i ventralnog strijuma muških štakora tijekom uvođenja konzekultura Časopis za neuroznanost, 1 prosinac 2002, 22 (23): 10477-10486
- Pregled knjige: Dinamika izvanstaničnog dopamina u akutnim i kroničnim djelovanjima kokaina Neuroznanstvenik, 1 kolovoz 2002, 8 (4): 315-322
- Diferencijalna uključenost NMDA, AMPA / kainatnih i dopaminskih receptora u jezgri jezgre nukleusa u stjecanju i izvedbi ponašanja Pavlovskog pristupa Časopis za neuroznanost, 1 prosinac 2001, 21 (23): 9471-9477
- Kontrola ponašanja koje traži kokain pomoću podražaja povezanog s lijekovima kod štakora: Učinci na oporavak ugroženih operant-reagirajućih i izvanstaničnih razina dopamina u amigdali i nucleus accumbens PNAS, 11, 2000, 97 (8): 4321-4326
- Olakšavanje spolnog ponašanja i poboljšanog dopaminskog izlijevanja u nuklearnim udarcima muških štakora nakon senzitivizacije ponašanja izazvane D-amfetaminom Časopis za neuroznanost, 1 siječanj 1999, 19 (1): 456-463
- NEUROZNANOST: Privlačenje pažnje mozga Znanost, 3 listopad 1997, 278 (5335): 35-37
- Dopaminergički korelati osjetilne specifičnosti u medijalnom prefrontalnom korteksu i nuklearnim naslagama štakora Časopis za neuroznanost, 1 listopad 1999, 19 (19): RC29