Neurosci Biobehav Rev. 2011 Jan; 35 (3) 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Nov 2.
- 1Bernard B. Brodie Odjel za neuroznanost, Sveučilište u Cagliariju, Cittadella Universitaria, 09042 Monserrato, CA, Italija. [e-pošta zaštićena]
SAŽETAK
Oksitocin je snažan induktor erekcije penisa kada se ubrizgava u središnji živčani sustav. Kod mužjaka štakora, najosjetljivije područje mozga za proerektilni učinak oksitocina je paraventricularna jezgra hipotalamusa. Ta jezgra i okolna područja sadrže stanična tijela svih oksitocinergičkih neurona koji se šire u područja izvan hipotalamusa i leđnu moždinu. Ovaj pregled pokazuje da oksitocin inducira erekciju penisa i kada se ubrizgava u nekim od tih područja (npr. Ventralno tegmentalno područje, ventralni subikulum hipokampusa, posteromedijalna kortikalna jezgra amigdale i torakalno-lumbalni kičmeni možd). Studije mikroinjekcije u kombinaciji s intracerebralnom mikrodijalizom i studijama dvostruke imunofluorescencije upućuju na to da oksitocin u tim područjima aktivira izravno ili neizravno (uglavnom preko glutaminske kiseline) mezolimbičke dopaminergičke neurone. Dopamin oslobođen u nucleus accumbens zauzvrat aktivira neuronske putove koji dovode do aktivacije in vitro-hipotalamičkih dopaminergičkih neurona u paraventricularnoj jezgri. To aktivira ne samo oksitocinergičke neurone koji se šire u leđnu moždinu i posreduju erekciju penisa, nego i one koji se projektiraju na gore navedena područja izvan hipotalamusa, izravno ili neizravno moduliraju (preko glutaminske kiseline) aktivnost mezolimbičkih dopaminergičkih neurona koji kontroliraju motivaciju i nagradu. Zajedno, ovi neuronski putovi mogu činiti složen hipotetički krug, koji igra ulogu ne samo u fazi konzumacije spolne aktivnosti (erektilna funkcija i kopulacija), već iu motivacijskim i nagrađivanim aspektima anticipativne faze seksualnog ponašanja.
1.Introduction
Erekcija penisa je muški seksualni odgovor koji igra ključnu ulogu u reprodukciji sisavaca uključujući i čovjeka, a to se također može promatrati u kontekstima koji se razlikuju od onih strogo povezanih s reprodukcijom. Ovisno o kontekstu u kojem se javlja erekcija penisa, različiti središnji i periferni živčani i / ili humoralni mehanizmi sudjeluju u njegovoj regulaciji (vidi Meisel i Sachs, 1994; Argiolas i Melis, 1995, 2004, 2005; Sachs, 2000, 2007; McKenna, 2000, Giuliano i Rampin, 2000, 2004, Andersson, 2001, Melis i Argiolas, 1995a, 2003, Hull et al., 2002). Među središnjim neurotransmiterima i neuropeptidima koji kontroliraju erekciju penisa, najpoznatiji su dopamin, serotonin, ekscitatorne aminokiseline, dušikov oksid, adrenokortikotropin, oksitocin i opioidni peptidi, Mogu olakšati ili spriječiti erekciju penisa djelujući u nekoliko područja mozga, srednja preoptička zona, paraventricularna jezgra hipotalamusa, ventralno tegmentalno područje, hipokampus, amigdala, sloj jezgre stria terminalis, nucleus accumbens, medulla oblongata i kičmena moždina (Tablica 1) (vidi Meisel i Sachs, 1994; Witt i Insel, 1994; Stancampiano et al., 1994; Argiolas i Melis, 1995, 2005; Argiolas, 1999; Bancila i sur., 2002; Giuliano i Rampin, 2000; Anderson, 2000, Hull et al., 2001, Coolen et al., 2002).
Oksitocin, neurohipofizni peptid poznat po svojoj hormonskoj ulozi u laktaciji i porođaju, prisutan je kod žena i muškaraca, ne samo u neuronima s staničnim tijelima koji se nalaze u paraventricularnim i supraoptičkim jezgrama hipotalamusa koji se projektiraju na neurohipofizu, ali iu neuronima od paraventricularne jezgre i okolnih struktura do ekstrahipotalamičkih područja mozga (tj. septuma, ventralnog tegmentalnog područja, hipokampusa, amigdale, izduljene medule i kičmene moždine). Smatra se da su ti neuroni uključeni u brojne središnje funkcije, kao što su pamćenje, učenje, pripadnost i socijalno-seksualno ponašanje, uključujući erekciju penisa i ponašanje u kopulaciji (vidi Buijs, 1978; Sofroniew, 1983; Argiolas i Gessa, 1991; Pedersen i dr.) Wagner i Clemens, 1992, Ivell i Russel, 1992, Carter et al., 1993, Tang et al., 1995, Veronneau-Longueville i sur., 1997). Doista, oksitocin olakšava erektilnu funkciju i spolno ponašanje muškaraca kod miševa, štakora, zečeva i majmuna (vidi Argiolas i Gessa, 1998; Carter, 1999; Pedersen et al., 1991; Argiolas i Melis, 1992, 1992; Argiolas, 1995, 2004; Argiolas i 1999). To se može pojaviti i kod ljudi, jer se oksitocin u plazmi povećava seksualnim stimulusima, posebno kod ejakulacije (Carmichael et al., 1987; Murphy i sur., 1987) i manipulacijom dojki i genitalija, koje se obično javljaju tijekom seksualne aktivnosti. odnos (Tindall, 1974).
Olakšavajući učinak oksitocina na seksualno ponašanje muškaraca prvi je pokazao sposobnost intravenskog oksitocina da smanji latenciju do prve ejakulacije i da uspori seksualnu iscrpljenost muških zečeva uparenih s receptivnim ženkama (Melin i Kihlstrom, 1963). Međutim, seksualne učinke oksitocina definitivno su prepoznali tek osamdesetih godina. Zatim je nađeno da oksitocin centralno u nanogramskim količinama može inducirati erekciju penisa (Argiolas et al., 1985, 1986) i poboljšati kopulacijsko ponašanje (Arletti i sur., 1985) kod mužjaka štakora, te povećati lordozu kod ženki štakora (Arletti). i Bertolini, 1985, Caldwell et al., 1986), očigledno djelovanjem na uterine-tipa oksitocinergičke receptore (vidi Argiolas i Melis, 1995, 2004; Argiolas, 1999; Melis i Argiolas, 2003; i reference u njima). Oksitocin poboljšava seksualno ponašanje ne samo kod seksualno snažnih mužjaka štakora (Arletti et al., 1985), nego i kod starijih mužjaka štakora (Arletti et al., 1990), te u dominantnim, ali ne u podređenim, muškim vjevericama (Winslow i Insel). , 1991).
Proerektilni učinak oksitocina ovisan je o testosteronu, budući da je ukinut hipofizektomijom i kastracijom, te obnovljen dodatkom testosterona ili njegovih metabolita, estradiola i 5_-dihidro-testosterona danih zajedno (Melis et al., 1994a). Najosjetljivije područje mozga za indukciju erekcije penisa pomoću oksitocina je paraventricularna jezgra hipotalamusa (Melis et al., 1986), iz koje potječu sve ekstra-hipotalamičke oksitocinergičke projekcije (vidi gore). Ovdje je nađeno da oksitocin može inducirati erekciju penisa (i zijevanje) kada se ubrizgava na tako niskoj dozi kao 3 pmol (vidi Odjeljak 2.1 u nastavku). Oksitocin je izazvao erekciju penisa također kada se ubrizgava bilateralno u CA1 polje hipokampusa, ali ne u dorzalni subikulum (vidi 2.3 dolje), lateralni septum, kaudatnu jezgru, medijalnu preoptičku površinu, ventromedijalnu jezgru hipotalamusa i supraoptična jezgra (Melis et al., 1986). Što se tiče mehanizma kojim oksitocin djeluje u paraventricular nukleusu kako bi izazvao ovaj seksualni odgovor, brojne studije ukazuju da oksitocin aktivira vlastite neurone. U skladu s ovom hipotezom, spolna interakcija povećava FOS, genski proizvod neposrednog ranog gena c-fos u paraventricularnim oksitocinergičkim neuronima koji se šire u leđnu moždinu, a koji su uključeni u kontrolu erekcije penisa (vidi Witt i Insel, 1994 i reference u njemu), i seksualna nemoć (npr. nemogućnost odraslog mužjaka štakora da se sjedinjuje s ovariektomiranom estrogen-progesteronceptiranom receptom) je povezana s mužjacima štakora s niskim razinama oksitocinske mRNA u paraventricularnom jezgru hipotalamusa (Arletti et al., 1997).
Je li oksitocin utječe na fazu predviđanja ili na fazu konzumacije seksualnog ponašanja trenutno nije jasno, Budući da oksitocin inducira erekciju penisa, a glavni učinak oksitocina na ponašanje kopulatora je smanjenje post-ejakulacijskog intervala kod mužjaka štakora (Arletti i sur., 1985), razumno je pretpostaviti da peptid poboljšava spolnu učinkovitost. Međutim, kako oksitocin također povećava socio-seksualnu interakciju (vidi Pedersen et al., 1992; Carter et al., 1997; Ivell i Russel, 1995), i antagonisti oksitocin receptora sprječavaju nekontaktne erekcije (Melis et al., 1999a), smatraju se indeksom seksualnog uzbuđenja (vidi Sachs, 1997, 2000, 2007; Melis et al., 1998, 1999b i reference u njemu), moguća uloga oksitocina u seksualnom uzbuđenju i seksualnoj motivaciji ne može se isključiti.
Ovaj pregled sažima objavljene i neobjavljene rezultate nedavnih studija koje to pokazuju oksitocin inducira erekciju penisa ne samo kada se ubrizgava u paraventricularnu jezgru hipotalamusa, nego iu druga područja izvan mozga, kao što je ventralno tegmentalno područje. (Melis et al., 2007, 2009a; Succu et al., 2008), t.ventralni subikulum hipokampusa i stražnje jezgre amigdale (Melis et al., 2009b, 2010), koji su važni sastojci limbičkog sustava i za koje se smatra da igraju ključnu ulogu u motivaciji i nagrađivanju. Ove studije otkrivaju da oksitocin sudjeluje u neuronskim krugovima, koji uključuju druge neurotransmitore, kao što su dopamin i glutaminska kiselina, i druga područja mozga koja nisu paraventricularna jezgra, npr. Ventralno tegmentalno područje, nucleus accumbens, hipokampus i područja koja tek treba identificirani. Ti su sklopovi vjerojatno posrednici u interakciji između mezolimbičkog i in vitro-hipotalamičkog dopaminergičkog sustava, te igraju ulogu ne samo u fazi konzumacije muškog seksualnog ponašanja (npr. Erekcije penisa i kopulacije), nego iu seksualnoj motivaciji i seksualnoj motivaciji. uzbuđenje, dakle pružanje neuralnog supstrata za objašnjavanje motivacijskih i nagrađivanih svojstava seksualne aktivnosti.
2. Oksitocin utječe na erekciju penisa djelujući u različitim područjima mozga
2.1. Paraventricularna jezgra hipotalamusa
Kao što se spominje iznad, paraventricularna jezgra hipotalamusa uskoro je prepoznata kao područje mozga koje je najosjetljivije na proerektilni učinak oksitocina. Kada se jednostrano ubrizgava u ovu jezgru, nađeno je da je oksitocin aktivan u niskim dozama kao 3 ng (3 pmol) (Melis et al., 1986). Ispitivanja odnosa strukture i aktivnosti pokazala su da je erekcija penisa izazvana oksitocinom posredovana uterinotipskim oksitocinskim receptorima, povezanim s Ca2 + ulaskom u stanična tijela oksitocinergičkih neurona koji se projektiraju na područja izvan hipotalamusa i na aktivaciju sintaze dušikovog oksida. Dušikov oksid zauzvrat djelujući kao unutarstanični glasnik s još nepoznatim mehanizmom (koji ne uključuje gvanilat ciklazu) dovodi do aktivacije oksitocinergičkih neurona koji se šire u moždana područja kičmene moždine i ekstra-hipotalamusa, inducirajući erekciju penisa (slika 1) (vidi i Argiolas i Melis, 1995, 2004, 2005 i reference u njima). Sposobnost oksitocina da aktivira vlastite neurone
Slika 1. (MISSING) Shematski prikaz oksitocinergičkih neurona, koji potječu iz paraventricularne jezgre hipotalamusa i projiciraju se u područja izvan hipotalamusa, kao što su kičmena moždina, VTA, hipokampus, amigdala, itd. dopamin, ekscitatorne aminokiseline, sam oksitocin, peptidi heksarelinskog analoga i VGF-izvedeni peptidi dovode do erekcije penisa, što se može smanjiti i / ili ukinuti stimulacijom GABAergičnih, opioidnih i kanabinoidnih CB1 receptora. Aktivacija oksitocinergičkih neurona je sekundarna u odnosu na aktivaciju sintetaze dušikovog oksida prisutne u tim neuronima. Doista, endogeni dušikov oksid nastao stimulacijom dopamina, ekscitatornih aminokiselina ili oksitocinskih receptora ili egzogenog dušikovog oksida, kao što je dobiven iz donora dušikovog oksida, izravno u paraventricularnu jezgru, aktivira oksitocinergičke neurone još neidentificiranim mehanizmom, koji očito nije povezan s stimulacija gvanilat ciklaze. To uzrokuje otpuštanje oksitocina u leđnoj moždini i izvan-hipotalamičkim područjima mozga. Neke pojedinosti o mehanizmima pomoću kojih oksitocin inducira erekciju penisa kada se oslobode u tim područjima, npr. VTA, ventralni subikulum i amigdala opisani su u odgovarajućim dijelovima područja mozga. Ovdje oksitocin djeluje na vlastite receptore i povećava produkciju NO, što dovodi do erekcije penisa kako je pronađeno u PVN. Međutim, u suprotnosti s PVN, u kaudalnom VTA NO aktivira gvanilat ciklazu. To uzrokuje povećanje koncentracije cGMP-a što dovodi do aktivacije mezolimbičkih dopaminergičkih neurona i erekcije penisa. U VS NO aktivira glutamatergične neurone koji se projektiraju na izvanhipokampalna područja, uključujući VTA. Glutaminska kiselina u VTA aktivira mezolimbičke dopaminergičke neurone, kao što je pronađeno kod oksitocina. Mehanizmi slični onima koji su gore opisani vjerojatno djeluju i kada se erekcija penisa događa u fiziološkom kontekstu, naime kada se muški štakori smjeste u prisustvu nepristupačne receptivne ženke (npr. Nekontaktne erekcije) ili tijekom kopulacije.
u paraventricular jezgre je podržan od strane studija pokazuje da: \ t (1) oksitocinski receptori su prisutni u ovoj jezgri hipotalamusa (Freund-Mercier et al., 1987; Freund-Mercier i Stoeckel, 1995); (2) oksitocin olakšava vlastito oslobađanje in vitro i in vivo (Freund-Mercier i Richard, 1981, 1984; Moos et al., 1984); i (3) Oksitocin uzbuđuje vlastite neurone djelujući u paraventricularnoj jezgri (Yamashita et al., 1987). Štoviše, oksitocinergički sinapsi koji utječu na stanična tijela magnocelularnih oksitocinergičkih neurona također su identificirani u paraventricularnom i supraoptičkom jezgru hipotalamusa. (Theodosis, 1985). Konačno, uništavanje središnjih oksitocerergičkih neurona elektrolitičkim ili kemijskim ekscitotoksičnim lezijama paraventricularne jezgre, koja u potpunosti uništava sadržaj oksitocina u središnjem živčanom sustavu i kralježničnoj moždini, ukida ne samo proerektilni učinak oksitocina, već također narušava lijekove erekciju penisa i nekontaktne erekcije (vidi niže i Argiolas et al., 1987a, b; Liu i sur., 1997 i reference u njemu). Rezultati slični onima pronađenim kod lezija paraventricularne jezgre nađeni su s jakim i selektivnim antagonistima receptora oksitocina. Doista, ovi spojevi ubrizgavani u paraventricularnu jezgru u nanogramskim količinama spriječili su potpuno oksitocin-induciranu erekciju penisa, dok su, kada su dani u lateralne komore, spriječili ne samo erekciju penisa induciranu samom oksitocinom, već i erekciju penisa uzrokovanu lijekovima (vidi dio 3 ispod i Argiolas i Melis, 1995, 2004, 2005 i reference u njima) i nekontaktne erekcije (Melis et al., 1999a), a osim toga su bile iznimno učinkovite u pogoršanju kopulacijskog ponašanja seksualno jakih mužjaka štakora (Argiolas et al., 1988). Nadalje, seksualna interakcija povećava FOS, genski produkt neposrednog ranog gena c-fos u paraventricularnim oksitocinergičkim neuronima koji se šire u leđnu moždinu uključenu u kontrolu erekcije penisa (vidi Witt i Insel, 1994 i reference u njemu). Konačno, seksualna nemoć (npr. nemogućnost odraslog mužjaka štakora da se sjedinjuje s estrogen-progesteronceptiranim ženskim)kao i kod mužjaka štakora s niskim nivoima mRNA oksitocina i sintaze dušik-oksida u paraventricularnoj jezgri hipotalamusa (Benelli i sur., 1995; Arletti i sur., 1997) (za opsežan pregled tih studija vidi Argiolas, 1999; Argiolas i Melis, 2004, 2005).
2.2. Ventralno tegmentalno područje
Ventralno tegmentalno područje otkriveno je tek nedavno kao mjesto mozga u kojem oksitocin inducira erekciju penisa, Ovo područje sadrži oksitocinergičke završetke živaca koji potječu iz paraventricularnih jezgri i oksitocinskih receptora (Freund-Mercier et al., 1987; Vaccari et al., 1998). Točnije, nađeno je da oksitocin može ierekcija penisa kada se unese jednostrano u kaudalnu, ali ne u rostralnu ventralnu tegmentalnu površinu na način ovisan o dozi (Melis et al., 2007). Aktivne doze bile su više od onih potrebnih za ubrizgavanje u paraventricularnu jezgru i slične onima koje induciraju erekciju penisa kada se ubrizgaju u ventralni subikulum hipokampusa ili u posteromedijalnu kortikalnu jezgru amigdale (vidi dolje). Očigledno, proerektilni učinak je posredovan aktivacijom mezolimbičkih dopaminergičkih neurona koji se šire u ljusku nukleusa accumbens, što zauzvrat aktivira još nepoznate neuronske puteve koji se projektiraju na in vitro-hipotalamičke dopaminergičke neurone koji udaraju na paraventricularne oksitocinergičke neurone koji posreduju u erekciju penisa (Melis et al., 2007, 2009a).
Što se tiče mehanizma kojim oksitocin aktivira dopaminergičku neurotransmisiju u ventralnom tegmentalnom području, dostupni podaci sugeriraju da Oksitocin stimulira oksitocinergičke receptore koji se nalaze u staničnim tijelima mezolimbičkih dopaminergičkih neurona. To povećava Ca2 + priljev unutar staničnih tijela dopaminergičnih neurona, čime se aktivira sintaza dušikova oksida (Succu et al., 2008). Odstupanje od paraventricularne jezgre (vidi Odjeljak 3 u nastavku), dušikov oksid zauzvrat aktivira gvanilat ciklazu, stoga povećavajući koncentraciju cikličkog GMP. U skladu s ovim mehanizmom, ili d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, snažan antagonist oksitocina ili S-metil-tio-l-citrulin, snažan inhibitor neuronske dušik-oksid-sintaze, ubrizgava se u kaudalnog ventralnog tegmentalnog područja prije oksitocina, ukinuta je erekcija penisa i povećanje ekstra-stanične koncentracije dopamina u ljusci nukleusa accumbens inducirane oksitocinom. Štoviše, 8-bromo-ciklički GMP, aktivni ciklički GMP analog koji je otporan na fosfodiesterazu, inducira erekciju penisa kada se ubrizgava u kaudalnu ventralnu tegmentalnu površinu i povećava izvanstaničnu koncentraciju dopamina u ljusci nukleusa accumbens, kako je pronađeno kod ubrizgavanja oksitocina u kaudalnom ventralnom tegmentalnom području (Succu et al., 2008; Melis et al., 2009a) (vidi također sliku 2).
Uvijek u skladu s ovim mehanizmom, haloperidol, snažan antagonist receptora za dopamin D2, ubrizgan u ljusku nukleusa accumbens smanjuje erekciju penisa uzrokovanu oksitocinom koji se ubrizgava u ventralno tegmentalno područje (Melis et al., 2007). Gornji mehanizam je također potkrijepljen dvostrukim imuno-fluorescentnim studijama, koje pokazuju da oksitocinska vlakna utječu na stanična tijela dopaminergičkih neurona u kaudalnom ventralnom tegmentalnom području, koji su prethodno označeni retrogradnim tragom Fluorogold ubrizganim u ljusku nucleus accumbens ( Melis i sur., 2007; Succu i sur., 2008). Aktivacije ovih dopaminergičnih neurona i dopaminskih receptora u nucleus accumbens vode na aktivaciju neuralnih puteva koji tek trebaju biti identificirani, a koji stimuliraju dopaminergičke neurone hipotalamusa da oslobode dopamin u paraventricularnom jezgru, aktivirajući tako oksitocinergičke neurone leđne moždine i posredovanje erekcije penisa (vidi gore i Melis et al., 2007; Succu et al., 2007, 2008). Doista, oksitocin se ubrizgava u kaudalnu ventralnu tegmentalnu površinu u dozi koja inducira erekciju penisa, povećanu izvanstaničnu koncentraciju dopamina u dijalizatu dobivenu ne samo iz nucleus accumbens, nego i iz paraventricularne jezgre (Succu et al., 2007).
2.3. Hipokampus
CA1 polje hipokampusa bilo je drugo područje mozga bogato oksitocinergičkim vlaknima i receptori identificirani ranijim studijama u kojima je injekcija oksitocina inducirala erekciju penisa (vidi Bujis, 1978; Sofroniew, 1983). Međutim, u odstupanju od paraventricularne jezgre, ovdje je nađeno da je oksitocin sposoban inducirati erekciju penisa samo kada se ubrizgava bilateralno i u višim dozama od onih koji su nađeni aktivni u paraventricularnom jezgru (Melis i sur., 1986; Chen et al., 1992). Injekcije oksitocina u subikulum utvrđene su neaktivnima u ovim ranijim studijama. Međutim, nedavne i pažljivije studije mikroinjekcije dovele su do identifikacije a područje ventralnog subiculuma u kojima je injekcija oksitocina bila sposobna inducirati erekciju penisa na način ovisan o dozi (Melis et al., 2009b). Proerektilni učinak oksitocina koji je ubrizgan u ovo područje mozga opažen je u dozama sličnim onima koje su bile nađene u ventralnom tegmentalnom području nakon unilateralne injekcije (Melis et al., 2007), kao što je pronađeno u paraventricular jezgri. Očigledno, oksitocin koji se ubrizgava u ventralni subikulum izaziva erekciju penisa aktiviranjem oksitocinergičkih receptora u neuronima koji sadrže sintetazu dušikova oksida, uzrokujući povećanje proizvodnje dušikovog oksida. Dušikov oksid koji djeluje kao međustanični glasnik aktivira neurotransmisiju glutaminske kiseline, što dovodi do erekcije penisa, moguće kroz neuralne (glutamatergične) eferentne projekcije iz ventralnog subiculuma na područja izvan hipokampusa koja moduliraju aktivnost mezolimbičkih dopaminergičnih neurona (npr. ventralno tegmentalno područje, prefrontalni korteks, paraventricularna jezgra) (vidi dolje i Melis, 2007, 2009b; Succu et al., 2008).
Ovaj mehanizam djelovanja je podržan eksperimentima intra-cerebralne mikrodijalize, koji pokazuju da je oksitocin ubrizgan u ventralni subikulum u dozama koje induciraju erekciju penisa, povećava proizvodnju dušikovog oksida i koncentraciju izvanstaničnog glutaminske kiseline u dijalizatu iz ventralnog subikuluma (Melis et al. 2010) i izvanstaničnog dopamina u nucleus accumbens (Melis et al., 2007). Ovi odgovori su antagonizirani ne samo antagonistom receptora oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, već i inhibitorom neuronske dušik-oksid-sintaze S-metil-tio-lcitrulinom i danim hemoglobinom koji apsorbira dušikov oksid. u ventralni subiculum nekoliko minuta prije oksitocina (Melis et al., 2010).
Štoviše, u skladu s ovim mehanizmom djelovanja, aktivacija glutamatergične neurotransmisije pomoću NMDA injektiranog u ventralni subikulum uzrokuje erekciju penisa (Melis et al., 2010). Fenotip eferentnih projekcija iz ventralnog subikuluma, koji uzrokuju aktivaciju mezolimbičkih dopaminergičkih neurona i povećanje izvanstaničnog dopamina u nucleus accumbens, trenutno je nepoznat. Međutim, budući da se erekcija penisa izazvana oksitocinom koja se ubrizgava u ventralni subiculum javlja istodobno s povećanjem ekstra-stanične glutaminske kiseline u dijalizatu iz ventralnog tegmentalnog područja, ali ne iz nucleus accumbens i antagonizira se (+) MK-801, potentni nekonkurentni antagonist receptora ekscitatornih aminokiselina podtipa NMDA (Woodruff et al., 1987), injektiran u ventralno tegmentalno područje, ali ne u nucleus accumbens (vidi sliku 2 i Melis et al., 2009b) , Vrlo je vjerojatno da ove projekcije dovode do aktivacije glutamatergične neurotransmisije u ventralnom tegmentalnom području, što zauzvrat aktivira mezolimbičke dopaminergičke neurone koji se šire u nukleus accumbens. Je li povećana koncentracija glutaminske kiseline u ventralnom tegmentalnom području nakon ubrizgavanja oksitocina u ventralni subiculum oslobođena od neurona koji potječu iz subikuluma ili u drugim područjima mozga (npr. Prefrontalni korteks) je nepoznata. Ipak, to uzrokuje aktivaciju mezolimbičkih dopaminergičkih neurona i povećano oslobađanje dopamina u nucleus accumbens. Ovdje aktivacija dopaminskih receptora dovodi do aktivacije inerto-hipotalamičkih dopaminergičkih neurona, oslobađajući dopamin u paraventricularnu jezgru, aktivirajući time oksitocinergičke neurone koji se šire u leđnu moždinu i posredujući erekciju penisa. (vidi gore i Melis et al., 2007, 2009a; Succu et al., 2008).
2.4. Amigdala
Amigdala je drugo područje mozga bogato oksitocinskim vlaknima i receptorima (vidi Freund-Mercier et al., 1987; Vaccari et al., 1998; Uhl-Bronner et al., 2005). Smatra se da je oksitocin uključen u različite funkcije, od anksiolize, društvenog pamćenja i spoznaje, socijalno ojačanog učenja, emocionalne empatije, emocionalne obrade lica i straha kod ljudi do erektilne funkcije i seksualnog ponašanja (vidi Kondo i sur., 1998; Dominguez et al., 2001; 2005; Ebner i sur., 2005; Huber et al., 2007; Domes i sur., 2008; Petrovic i sur., 2009; Lee i sur., 2009; Donaldson i Young, 2010; Hurlemann et al. , 2009). Međutim, sposobnost oksitocina da inducira erekciju penisa kod mužjaka štakora kada je inducirana u posteromedijalnoj kortikalnoj jezgri amigdale otkrivena je tek nedavno (Melis et al., 2009b). Taj se odgovor pojavio istodobno s povećanjem ekstra-stanične koncentracije dopamina u dijalizatu dobivenom iz ljuske nucleus accumbens, kao što je pronađeno nakon injekcije oksitocina u ventralni subikulum (Melis et al., 2b). Mehanizam kojim se oksitocin ubrizgava u posteromedijalnu kortikalnu jezgru amigdale trenutno inducira erekciju penisa. Dostupni podaci pokazuju da je i erekcija penisa i povećanje ekstra-stanične koncentracije dopamina u dijalizatu dobivenom iz nucleus accumbens posredovano aktivacijom oksitocinergičkih receptora, budući da su oba odgovora ukinuta antagonistom receptora oksitocina d (CH5) 2Tyr ( Me) 8-Orn2009-vazotocin ubrizgan u jezgru amigdale nekoliko minuta prije oksitocina (Melis et al., XNUMXb).
Koji god mehanizam oksitocin aktivira u posteromedijalnoj kortikalnoj jezgri amigdale, seksualni odgovor izazvan peptidom je ukinut blokadom svih dopaminergičnih receptora s cis-flupentiksolom ubrizganim u ljusku nucleus accumbens i blokadom NMDA receptora sa ( +) MK-801 se ubrizgava u ventralno tegmentalno područje, ali ne u nucleus accumbens, kao što je pronađeno za erekciju penisa induciranu oksitocinom ubrizganom u ventralni subikulum (Melis et al., 2009b). To sugerira da oksitocin ubrizgan u posteromedijsku jezgru amigdale aktivira neurotransmisiju glutaminske kiseline u ventralnom tegmentalnom području. To uzrokuje aktivaciju mezolimbičkih dopaminergičkih neurona, što dovodi do erekcije penisa. S obzirom na studije koje pokazuju neuronske putove koji spajaju tu jezgru amigdale s ventralnim subikulumom (Canteras i sur., 1995; francuski i Totterdell, 2003), ovi nalazi podižu mogućnost da interakcija može postojati između ta dva područja mozga, iako izravna opisani su putovi od amigdale do jezgre accumbens ili do ventralnog tegmentalnog područja (Kelley i Domesick, 1982; Witter, 2006).
2.5. Kičmena moždina
Kičmena moždina je drugo područje središnjeg živčanog sustava koje sadrži oksitocinergična vlakna i receptore (Freund-Mercier i sur., 1987; Uhl-Bronner i sur., 2005), u kojoj oksitocin inducira erekciju penisa (Tang et al., 1998). Veronneau-Longueville i sur., 1999; Giuliano i Rampin, 2000; Giuliano et al., 2001). Kao što je već spomenuto, ova oksitocinergička vlakna potječu iz paraventricularne jezgre hipotalamusa i pridonose silaznim putovima koji kontroliraju spinalne autonomne neurone koji posreduju u erekciji penisa. Doista, ova vlakna stvaraju sinaptičke kontakte u preganglionskim simpatičkim i parasimpatičkim staničnim stupovima dorzalnog roga u torakalno-lumbalnom i lumbo-sakralnom traktu s spinalnim neuronima koji inerviraju kavernozne korpise penisa (Marson i McKenna, 1996; Giuliano i Rampin, 2000; Giuliano i sur. , 2001). Ovi sinaptički kontakti pokazani su obilježavanjem spinalnih neurona koji potječu iz penisa i dostizanjem kralježnične moždine sa specifičnim retrogradnim tragačima ubrizganim u kavernozne korpuse, u kombinaciji s dvostrukom imuno-fluorescentnom i konfokalnom laserskom mikroskopijom (Tang et al., 1998; Veronneau-Longueville). i sur., 1999). U skladu s tim studijama, kod anesteziranih muških štakora intratekalna injekcija kumulativnih doza oksitocina na lumbo-sakralnoj, ali ne na toroko-lumbalnoj razini, izazvala je porast intrakavernoznog tlaka na način ovisan o dozi. Ovi učinci su ukinuti blokadom oksitocinergičkog receptora s d (CH2) 5Tyr (Me) 2-Orn8-vazotocinom i dijelom zdjeličnih živaca (Giuliano i Rampin, 2000; Giuliano et al., 2001). Ovi rezultati pokazuju da oksitocin, djelujući na lumbo-sakralnu kralježničnu moždinu, povećava intrakavernozni tlak i sugerira da je oksitocin, oslobođen tijekom fiziološke aktivacije paraventricularne jezgre, snažan aktivator spinalnih proerektilnih neurona koji se šire u kavernozni korpus. Zanimljivo je da ti proerektilni spinalni neuroni na kojima oksitocin djeluje na svoj proerektilni učinak, također primaju sinaptičke kontakte od serotoninergičkih neurona koji potječu iz nukleusa paragigantocellularis retikularne formacije medulla oblongata (Marson i McKenna, 1992; Tang et al ., 1998). Uništavanje ovih serotoninergičkih neurona olakšava ejakulaciju i reflekse penisa kod mužjaka štakora (Marson i McKenna, 1992; Yells i sur., 1992). Budući da lijekovi koji stimuliraju receptore 5HT2C olakšavaju erekciju penisa ako se daju intracerebroventrikularno, ali ne i u paraventricularnu jezgru, i lijekovi koji blokiraju receptore 5HT2C smanjuju i erekciju penisa agonistom i oksitocinom, dok antagonisti dopamina ne smanjuju erekciju penisa izazvanu 5HT2C ( vidi Stancampiano et al., 1994 i reference u njemu), također je sugerirano da oksitocin olakšava djelovanje proerektilnih 5HT2C receptora na razini lumbo-sakralne kralježnice (Stancampiano et al., 1994). Alternativno, oksitocin može utjecati na aktivnost spinalnih spinalnih serotoninergičkih neurona, djelujući izravno u jezgri paragigantocellularis, gdje nastaju ti neuroni (vidi Stancampiano et al., 1994).
3. Interakcije između oksitocina, dopamina i glutaminske kiseline u središnjem živčanom sustavu i erekcije penisa
Kao što se spominje u Odjeljku 1, svi oksitocinergički neuroni prisutni u središnjem živčanom sustavu potječu iz paraventricularne jezgre i okolnih struktura. Aktivnost ovih neurona je pod kontrolom različitih neurotransmitera i / ili neuropeptida. Među najviše proučavanim na paraventricularnoj razini su dopamin, glutaminska kiselina, gama-aminobutirna kiselina (GABA), dušikov oksid, endokanabinoidi, opioidni peptidi, peptidi koji oslobađaju hormon rasta, peptidi povezani s VGF-om i sam oksitocin. Dopamin, glutaminska kiselina, peptidi koji oslobađaju hormon rasta, peptidi izvedeni iz VGF-a i oksitocin su stimulativni, npr. Ovi spojevi i njihovi agonisti olakšavaju erekciju penisa kada se ubrizgavaju u paraventricularnu jezgru, dok su GABA, opioidni peptidi i endokanabinoidi inhibitori, npr. Ovi spojevi ili njihovi agonisti inhibiraju erekciju penisa (vidi Meisel i Sachs, 1994; Witt i Insel, 1994; Argiolas i Melis, 1995, 2004, 2005; Giuliano i Rampin, 2000, 2004; McKenna, 2000; Andersson, 2001; Hull et al., 2002).
Nekoliko linija eksperimentalnih dokaza sugerira da su ti oksitocinergički neuroni i gore navedeni neurotransmiteri i neuropeptidi uključeni u kontrolu erektilne funkcije i seksualnog ponašanja u različitim fiziološkim kontekstima. Štoviše, oksitocin koji se oslobađa u izvan-hipotalamičkim područjima mozga, kao što je ventralno tegmentalno područje, hipokampus i njegova područja, amigdala i leđna moždina mogu utjecati na aktivnost onih neurona na koje udaraju oksitocinergični sinapsi. U ovom trenutku, jedini neuroni važni za erekciju penisa na koji udaraju oksitocinergički sinapsi, identificirani sa sigurnošću, su stanična tijela mezolimbičkih dopaminergičkih neurona kaudalnog ventralnog tegmentalnog područja koji se šire u ljusku nucleus accumbens (Melis i sur., 2007). Succu i sur., 2008), i proerektilne spinalne neurone koji projiciraju iz lumbo-sakralnog trakta do kavernoznih korpusa (vidi Giuliano i Rampin, 2000; Giuliano et al., 2001) (vidi također odjeljke 2.2 i 2.5) , Doista, iako su oksitocinergički sinapsi i receptori također identificirani u ventralnom subikulumu, amigdali i kralježničnoj moždini, područja koja su važna za erekciju penisa (vidi gore), u tim područjima tip neurotransmitera prisutan u neuronima na koji neprestano udaraju oksitocinergički završetci živaca.
Ovaj dio pregleda ukratko sažima nedavnu literaturu o mehanizmima na kojima se temelji proerektilni učinak oksitocina koji se ubrizgava u kaudalnu ventralnu tegmentalnu površinu, ventralni subikulum hipokampusa i kičmenu moždinu. Posebna pozornost posvećena je interakciji peptida s dopaminom i glutaminskom kiselinom u tim područjima te o ulozi koju ova interakcija može imati u središnjoj kontroli erektilne funkcije. Prvo se daje kratki sažetak učinaka dopamina i glutaminske kiseline na oksitocinergičke neurone u paraventricularnoj jezgri, koji također igraju ključnu ulogu u erektilnoj funkciji, kako bi čitatelj bio svjestan ranog stanja istraživanja u ovom području. ove studije su već opsežno pregledane (vidi Argiolas i Melis, 1995, 2004, 2005; Melis i Argiolas, 2003). Također u ovom slučaju posebna se pozornost posvećuje najnovijim rezultatima, koji ukazuju na važnu ulogu i za dopaminsko-oksitocinsku vezu i za glutaminsku kiselinu-oksitocinsku vezu ne samo u seksualnoj izvedbi (erekcija penisa i kopulacija), nego iu seksualnoj izvedbi. uzbuđenje i seksualnu motivaciju.
3.1. Interakcija dopamin-oksitocina u paraventricularnoj jezgri
Korištenje električnih romobila ističe Sposobnost dopaminskih agonista da induciraju erekciju penisa aktiviranjem središnjih oksitocerergičkih neurona predloženo je odmah nakon otkrića da apomorfin inducira erekciju penisa kada se ubrizgava u paraventricularnu jezgru (Melis et al., 1987) kada je pronađen antagonist oksitocin receptora d (CH2) 5Tyr (Me) 2-Orn8-vazotocin injektiran intracerebroventrikularno (icv). sposobni smanjiti gotovo potpuno erekciju penisa induciranu ne samo oksitocinom koji se daje icv, nego i apomorfinom, danim subkutano (Argiolas et al., 1987b). Ovi rezultati praćeni su onima drugih studija koje su pokazale slične rezultate kada je d (CH2) 5Tyr (Me) 2-Orn8-vazotocin dobio icv i apomorfin je dan izravno u paraventricularnu jezgru (Melis et al., 1989b), lsugerirajući da dopaminski agonisti induciraju erekciju penisa aktiviranjem paraventricularnih oksitocinergičkih neurona koji se projektiraju u područja izvan hipotalamusa mozga, a posebno u leđnu moždinu. (vidi Argiolas i Melis, 1995, 2004, 2005). U skladu s ovom hipotezom, kod anesteziranih štakora nedavno je otkriveno da blokiranje lumbo-sakralnih oksitocinergičkih receptora od strane nepeptidnog antagonista receptora oksitocina može ukinuti porast apororfnog tlaka induciranog apomorfinom uzrokovan apomorfinom dopaminskog agonista, pružajući dokaze za paraventriculo-spinalni oksitocinergički put uključen u erekciju penisa (Baskerville et al., 2009).
Studije usmjerene na Identifikacija dopaminskog receptora odgovornog za indukciju penisne erekcije, otkrila je da također u paraventricularnom jezgru agonisti dopaminskih receptora induciraju erekciju penisa djelujući na dopaminske receptore obitelji D2kao što je utvrđeno sistemskim agonistima dopaminskih receptora (vidi Melis et al., 1987; Eaton et al., 1991; Melis i Argiolas, 1995a). Prema tome, za apomorfin, snažan miješani agonist D1 / D2 receptora i kvinpirol, snažan selektivni agonist D2receptora, ali ne i SKF 38393, selektivni agonist D1 receptora, injiciran u ovu jezgru hipotalamusa, nađeno je da induciraju erekciju penisa na način ovisan o dozi i seksualni odgovor induciran ovim agonistima D2 receptora je ukinut antagonistima D2 receptora, kao što su haloperidol i l-sulpirid, ali ne SCH 23390, selektivni antagonist D1 receptora (Melis et al., 1987). Sposobnost apomorfina da inducira erekciju penisa kada se ubrizgava u paraventricularnu jezgru također je potvrđena telemetrijskim studijama koje pokazuju da je dopaminski agonist dan u paraventricularnu jezgru sposoban povećati intrakavernozni tlak u budnim mužjacima štakora bez mijenjanja sistemskog krvnog tlaka (Chen et al. 1999, Giuliano i Allard, 2001), kao što je pronađeno nakon sistemske injekcije (Bernabè et al., 1999). Ove studije također potvrdili su glavnu ulogu D2 receptora, budući da se obično utvrdi da agonisti D1 receptora ne mogu povećati intrakavernozni tlak kada se ubrizgavaju u paraventricularnu jezgru (Chen et al., 1999).
Tada je bilo dostupno nekoliko linija eksperimentalnih dokaza koji upućuju na to da su paraventrikularni receptori D2-a, čija stimulacija izaziva erekciju penisa, smješteni na stanicama tijela oksitocinergičkih neurona. Prvo, paraventricularna jezgra sadrži dopaminergičke terminalne živce koji pripadaju takozvanim incertohypothalamic dopaminergičnim neuronima. Stanična tijela tih neurona nalaze se u skupini A13 i A14 Dahlstroma i Fuxe (1964), arboriziraju i inerviraju nekoliko hipotalamičkih struktura, uključujući paraventricularne oksitocinergičke neurone koji se projektiraju na područja neurohipofize i / ili izvan-hipotalamičkog mozga (Buijs i sur., 1984; Lindvall i sur., 1984).
Uključenost ovih dopaminergičkih neurona na paraventricularnoj razini u kontroli erekcije penisa i kopulacije potkrijepljena je mikrodijalizama koje pokazuju da su koncentracije izvanstaničnog dopamina i 3,4-dihidroksifeniloctene kiseline (DOPAC), njegovog glavnog metabolita, povećane u dijalizatu dobivenom iz paraventricular nukleusa seksualno potentnih mužjaka štakora koji pokazuju nekontaktne erekcije kada se stavi u prisustvu nepristupačne ženke s receptorom jajnika ovarijektomiziranog estrogena + progesterona (Melis et al., 2003).
Povećanje koncentracije dopamina i DOPAC bilo je još veće kada je dopuštena kopulacija s receptivnom ženkom (Melis et al., 2003), kao što je pronađeno u medijalnom predoptičkom području (Hull et al., 1995) i u nucleus accumbens (Pfaus). i Everitt, 1995). Drugo, nekoliko studija pokazuje da je erekcija penisa izazvana stimulacijom paraventricularnih D2 receptora posredovana oksitocinom oslobođenim u tim područjima. U skladu s tim, utvrđeno je da apomorfin koji se daje u dozama koje induciraju erekciju penisa povećavaju koncentraciju oksitocina, ne samo u plazmi štakora i majmuna (Melis et al., 1989a; Cameron et al., 1992), ali također u ekstra-hipotalamičkom mozgu područja, kao što je hipokampus (Melis et al., 1990). U skladu s tim rezultatima, nedavno je pokazano da je apomorfin koji je injektiran u paraventricularnu jezgru u dozi koja inducira erekciju penisa u stanju povećati i izvanstaničnu koncentraciju dopamina u nucleus accumbens, učinak smanjen antagonistom receptora oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin ubrizgan u ventralno tegmentalno područje (Succu i sur., 2007; Melis et al., 2009a) (vidi također dio 4). Treće, bilateralne elektrolitičke lezije paraventricularne jezgre, koje gotovo u potpunosti eliminiraju oksitocin iz izvan-hipotalamičkih područja mozga (Hawthorn et al., 1985), ukidaju erekciju penisa izazvanu apomorfinom (Argiolas i sur., 1987a) i selektivni antagonisti oksitocinskog receptora dane u lateralne komore, ali ne u paraventricularnu jezgru, smanjuju erekciju penomom izazvanu ovisnom o dozi s potentnošću paralelnom s onom tih spojeva u blokiranju oksitocinskih receptora (Melis et al., 1989b). Antagonisti receptora oksitocina također su iznimno snažni u smanjenju olakšavanja muškog seksualnog ponašanja izazvanog ne samo oksitocinom, već i apomorfinom (Argiolas et al., 1988, 1989).
Što se tiče mehanizma kojim se receptori D2-a aktiviraju dopaminom ili agonistima dopaminskih receptora, povećavaju aktivnost oksitocinergičkih neurona, oslobađajući tako oksitocin u ekstrahipotalamičkim područjima mozga i kralježničnoj moždini, brojni eksperimentalni podaci.Podupiru hipotezu da stimulacija D2 receptora povećava koncentraciju unutarstaničnih iona Ca2 + unutar staničnih tijela oksitocinergičkih neurona, što dovodi do aktivacije sintaze dušikovog oksida, enzim koji ovisi o Ca2 + -kalmodulinu, koji je prisutan u ovim staničnim tijelima (Vincent i Kimura, 1992; Torres et al., 1993; Sanchez i sur., 1994; Sato-Suzuki i sur., 1998). Povećana proizvodnja dušikovog oksida uzrokuje aktivaciju oksitocinergičkih neurona. Prema tome, (1) erekcija penisa inducirana apomorfinom bila je spriječena organskim blokatorima kalcijevih kanala i by-konotoksinom GVIA, snažnim i selektivnim blokatorom napon-ovisnih Ca2 + kanala N-tipa (McCleskey et al., 1987), danih u paraventricular nukleus (vidi Argiolas et al., 1990, i reference u njemu); (2) erekcija penisa izazvana apomorfinom spriječena je inhibitorima sintetaze dušikovog oksida u paraventricularnom jezgru (Melis et al., 1994c); i (3) apomorfin i drugi agonisti D2 receptora koji se daju u dozama koje induciraju erekciju penisa povećali su proizvodnju dušikovog oksida u paraventricularnom dijalizatu dobivenom intracerebralnom mikrodializom, što je smanjenje koje su smanjene inhibitorima paraventricularne dušik oksid-sintaze dane u dozama koje smanjuju Erekcija penisa izazvana agonistom D2 receptora (Melis et al., 1996). Mehanizam pomoću kojeg dušikov oksid aktivira paraventrikularne oksitocinergičke neurone, još uvijek nije poznat, iako dostupni podaci ukazuju na to da dušikov oksid djeluje kao unutarstanični glasnik i da gvanilat ciklaza nije uključena. Zaista, analozi aktivnog fosfodiesteraze rezistentnog na ciklički GMP, 8-bromo-ciklički GMP, nisu mogli inducirati erekciju penisa kada se daju u paraventricularnu jezgru (Slika 2) (vidi Melis i Argiolas, 1995b i reference u njima).
Gornje tumačenje se često smatra neuvjerljivim, uglavnom zato što je stimulacija receptora dopamina D2 obično povezana s inhibicijom radije nego ekscitacijom staničnih tijela neurona koji sadrže ove receptore kroz različite G mehanizme vezane uz mehanizam (vidi Sokoloff i Schwartz, 1995). Međutim, moguće objašnjenje ovog odstupanja, koje je u skladu s izravnom stimulacijom paraventricularnih oksitocinergičkih neurona dopaminom, nedavno je predloženo otkrićem G dopamin D4 spregnut s proteinima receptor, član obitelji D2 receptora (D2, D3 i D4), čija stimulacija povećava Ca2 + priljev u stanične pripravke koji sadrže kloniranu verziju ovog podtipa receptora (Moreland et al., 2004). Još je važnije da je selektivni agonist D4 receptora (npr. ABT 724) (N-metil-4- (2-cijanofenil) piperazinil-3metilbenzamid maleat) sposoban inducirati erekciju penisa kod mužjaka štakora kada se daje sistemski (Brioni et al., 2004). Ovaj učinak nije pronađen kod selektivnog agonista D2 receptorskog podtipa PNU-95666E (R-5,6-dihidro-N, N-dimetil-4Himidazo [4,5,1-i] kinolin-5-amin) (Hsieh et al., 2004), također nije mogao povećati priljev Ca2 + u stanične pripravke koji sadrže kloniranu verziju podtipa D4 receptora (Brioni et al., 2004; Moreland et al., 2004). U skladu s gornjom hipotezom i nalazima, PD 168,077 (N-metil-4- (2-cijanofenil) piperazinil-3metilbenzamid maleat), PIP-3EA (2- [4- (2-metoksifenil) piperazin-1-ilmetil] imidazo [1,2-a] piridin) i drugi selektivni agonisti D4 receptora (Heier i sur., 1997; Melis et al., 2006b; Löber et al., 2009), nađeni su da mogu inducirati erekciju penisa kada se injektiraju sustavno, icv i paraventricular jezgre, iako manje učinkovito od apomorfina. Pro-erektilni učinak ovih agonista D4 receptora spriječen je s L-745,870 (3- (4- [klorofenil] piperazin-1-il) -metil-1H-pirolo [2,3-B] piridin trihidroklorid), selektivni D4 receptor antagonist (Patel et al., 1997; Melis et al., 2005, 2006b; Löber et al., 2009).
Konačno, proerektilni učinak gore navedenih agonista D4 receptora također je smanjen inhibitorima sintaze dušikovog oksida, danim u paraventricularnu jezgru, te d (CH2) 5Tyr (Me) 2-Orn8-vazotocinom, selektivnim antagonistom oksitocinskog receptora dano icv ali ne u paraventricular jezgri. Ovi rezultati su u skladu s hipotezom da agonisti D4 receptora također stimuliraju oksitocinergičke neurone aktiviranjem sintetaze dušikovog oksida i oslobađanjem oksitocina u ekstrahipotalamičkim područjima mozga, što pak olakšava erekciju penisa, kao što je pokazano za apomorfin i klasične D2 agoniste (Melis i sur. 2005, 2006b, Löber et al., 2009).
Gornji nalazi također podupiru hipotezu da dopamin inducira erekciju penisa djelujući na D4 receptore koji se nalaze na staničnim tijelima paraventricularnih oksitocinergičnih neurona, a koji uzrokuju povećani priliv Ca2 + u stanična tijela oksitocinergičkih neurona, što dovodi do povećane proizvodnje dušikovog oksida. Dušikov oksid zauzvrat aktivira oksitocinergičke neurone kako bi oslobodio oksitocin u moždanim područjima izvan hipotalamusa iu leđnoj moždini, kao što je već spomenuto. U tom pogledu, važno je napomenuti da su dopaminski receptori identificirani u staničnim tijelima oksitocinergičkih neurona u paraventricularnom jezgru tek nedavno dvostrukim imuno-fluorescencijskim studijama s visoko selektivnim antitijelima D2, D3 i D4 receptora i s oksitocin antitijelima. Ove studije su pokazale ekspresiju sva tri podtipa D2receptora (D2, D3 i D4), koji su lokalizirani odvojeno u staničnim tijelima oksitocinergičkih neurona u paraventricularnoj jezgri (i također u supraoptičnom jezgru i medijalnom preoptičnom području) (Baskerville). i Douglas, 2008, Baskerville i sur., 2009).
To osigurava snažnu neuroanatomsku potporu mogućnosti da agonisti dopamina i dopaminskih receptora tipa D2 induciraju erekciju penisa aktiviranjem izravno oksitocinergičkih neurona koji se projektiraju na područja mozga ekstrahipotalamusa koja su navedena gorenpr. kičmena moždina, ventralno tegmentalno područje, hipokampus i amigdala. Međutim, ovi nalazi ne pružaju nikakvu pomoć za identifikaciju podtipa / receptora D2, čija stimulacija uzrokuje erektilni odgovor. Nažalost, niti jedna pomoć se ne može dobiti ni iz studija koje su usmjerene na identificiranje oksitocinergičkih neurona aktiviranih agonistima dopaminskih receptora u paraventricularnoj jezgri. Doista, unatoč različitoj aktivnosti na različite podtipove dopaminskih receptora, bilo miješanih agonista dopaminskih receptora (npr. Apomorfina) ili selektivnih agonista D2 receptora (npr. Kinpirol, koji djeluje na sve podtipove D2 receptora) ili selektivnih agonista D4 receptora aktiviranje oksitocinergičkih neurona, mjereno povećanjem FOS proteina u parvocelularnim oksitocinergičkim neuronima paraventricularne jezgre (Bitner et al., 2006). Međutim, ovaj nalaz je nedavno ispitivan, budući da je povećanje FOS proteina u paraventricularnim oksitocinergičkim neuronima pronađeno samo kada je penisom izazvana erekcija, koja djeluje uglavnom na D2 i D3 receptorske podtipove, ali ne na PD 168077, agonist D4 receptora, usprkos sposobnosti oba spoja da induciraju seksualni odgovor (Baskerville et al., 2009).
Daljnji eksperimenti s selektivnim agonistima drugih D2 receptorskih podtipova (uglavnom D2 i D3) su tada neophodni za identificiranje točne uloge svakog podtipa dopaminskog receptora u kontroli erektilne funkcije na paraventricularnoj razini. U tom smislu, kao što je već ranije spomenuto, apomorfin, koji djeluje snažno na sve podtipove dopaminskih receptora (vidi Brioni et al., 2004, i reference u njemu), je mnogo učinkovitiji od agonista D4 receptora u induciranju erekcije penisa kada se ubrizga u paraventricular jezgra. To se može objasniti većim afinitetom apomorfina na D4 receptore u usporedbi s ispitivanim agonistima D4 receptora, ili alternativno, agonisti D4 receptora mogu djelovati kao djelomični agonisti D4 receptora, ili istodobno aktiviranje različitih podtipova dopaminskih receptora pomoću apomorfina. proizvedu višu aktivaciju oksitocinergičkih neurona koji posreduju erekciju penisa, nego samo aktiviranje agonistima D2 receptora podtipa D4 receptora.
Interakcije između dopaminskih D1 i D2 receptora već su opisane u kontroli seksualnog ponašanja na razini srednjeg preoptičkog područja (vidi Hull et al., 1989). U slučaju da nesposobnost selektivnih agonista D2 receptora da induciraju erekciju penisa (Hsieh et al., 2004) će biti potvrđena (ali vidi Depoortère et al., 2009), na primjer čak i nakon injekcije ovih spojeva u paraventricular nucleus , Glavnu ulogu za D3 receptore same ili zajedno s onom D4 receptora u aktivaciji oksitocinergičkih neurona koji posreduju erekciju penisa treba detaljno analizirati (vidi Baskerville et al., 2009). Nažalost, selektivni agonisti D2 i D3 receptora (npr. Koji se razlikuju u svom afinitetu za ova dva podtipa receptora za najmanje četiri / pet redova veličine in vitro) trenutno nisu dostupni. Iz tog razloga, nedavna sugestija da D3 receptori posreduju erekciju penisa inducirana klasičnim D2 receptor agonistima, koja se temelji uglavnom na sposobnosti pretpostavljenih antagonista D3 receptora karakteriziranih u in vitro eksperimentima, za smanjenje erekcije penisa inducirane klasičnim D2 agonistima, kao što je apomorfin, kvinpirol i pramipeksol, koji snažno aktiviraju sve podtipove receptora dopamina D2 (Collins et al., 2009), svakako se mora potvrditi drugim eksperimentima. Ova validacija je također neophodna jer u ovoj studiji nije nađen učinak agonista D4 receptora na erekciju penisa, što je upečatljiv kontrast rezultatima gore navedenih studija, koje pokazuju proerektilni učinak agonista D4 receptora. Zaista, čak i sposobnost apomorfina da inducira erekciju penisa (i zijevanje) u miševima s nokautom D4 receptora s potentnošću identičnom onom u divljim tipovima D4 receptora nokautiranjem miševa i sposobnosti antagonista D3 receptora da ukinu apomorfni odgovor u tim životinjama ( Collins i sur., 2009) ne mogu se smatrati definitivnim dokazom selektivne uloge podtipa D3 receptora u erekciji penisa izazvanom agonistom D2 receptora. Razlike u vrstama, studije s neurotransmiterskim / neuropeptidnim i / ili neurotransmiterskim / neuropeptidnim receptorskim genima (neurotransmiter / neuropeptid i / ili neurotransmiter / neuropeptidni receptori za nokautiranje životinja) obično su dodavale daljnju konfuziju i komplikacije u potvrđivanju pretpostavljene seksualne uloge neurotransmitera i ili neuropeptidi i njihovi receptori. Naime, oksitocinski gen za ablaciju proizvodi oksitocinske nokautirajuće miševe koji se normalno pare i kopuliraju, kao da je oksitocin nepotreban za parenje i kopulaciju. Također homozigotni ženski miševi s nokautom oksitocina pokazuju normalno parenje i parturaciju, iako s izraženim oštećenjem smanjenja količine mlijeka (Nishimori et al., 1996; Young i sur., 1996). Ablacija gena koji kodira neuronsku sintazu dušikovog oksida također proizvodi miševe koji sintetiziraju dušik oksid koji se normalno pare i kopuliraju (Huang et al., 1993). Međutim, ovi nalazi vjerojatno ukazuju na važnu značajku reproduktivne fiziologije, tj. Redundanciju sustava uključenih u njegovu kontrolu na središnjoj i perifernoj razini.
Takva redundancija svakako ima evolucijsko porijeklo, jer jamči prolazak gena sljedećoj generaciji za opstanak vrste. Prema tome, činjenica da ablacija gena D4 receptora ne mijenja proerektilni učinak apomorfina, sugerira da su receptori D4-a, poput oksitocina i dušikovog oksida, samo neki od medijatora koji rade u sustavima koji kontroliraju erektilnu funkciju, a ne sugerirajući da za ove receptore ne postoji uloga u kontroli erekcije penisa i seksualnog ponašanja. Nedostatak D4 agonista da induciraju erekciju penisa kada se sistemski daju muškim štakorima različitih sojeva nedavno je prijavljen u drugoj studiji (Depoortère et al., 2009). Međutim, za razliku od rada Collinsa i sur. (2009), te kako bi slika o ulozi različitih podtipova D2 receptora u kontroli erekcije penisa bila još zbunjujuća, ova studija također pokazuje da navodni selektivni antagonisti D3 receptora dani sistemski nisu bili u mogućnosti smanjiti erekciju penisa izazvanu apomorfinom u muški štakori soja osjetljiviji na proerektilni učinak apomorfina, dok je seksualni odgovor (i zijevanje) antagoniziran selektivnim D2 antagonistom L-741,626 (3 - [[4- (4-klorofenil) -4-hidroksipiperidin -1-il] metil-1H-indol), što navodi autore da sugeriraju da su D2 receptori, umjesto D3 i D4 receptori, oni koji igraju glavnu ulogu u erekciji penisa izazvanoj D2 agonistom (Depoortère et al., 2009). Konačno, mogućnost da ekscitacijski učinak agonista dopaminskih receptora na oksitocinergičke neurone koji posreduju u erekciji penisa, barem djelomično, je neizravna, a ne izravna, npr. Posredovana ili pod utjecajem promjena u aktivnosti neuropeptida drugih neurotransmitera koji mogu modulirati aktivnost oksitocinergičkih neurona u paraventricularnoj jezgri, ne mogu se potpuno isključiti.
3.2. Interakcija glutaminske kiseline i oksitocina u paraventricularnoj jezgri
Paraventricularna jezgra hipotalamusa je vrlo bogata sinapsama koje sadrže ekscitatorsku aminokiselinu kao neurotransmiter (npr. Glutaminsku kiselinu i asparaginsku kiselinu).) (Van Den Pol, 1991). Uzbudljive aminokiseline u ovoj jezgri su uključene u brojne funkcije, uključujući erekciju penisa i seksualno ponašanje (Roeling et al., 1991; Melis et al., 1994b, 2000, 2004b). Prema tome, N-metil-d-asparaginska kiselina (NMDA), selektivni agonist podtipa NMDA receptora, ali ne i (±) -_-amino-3-hidroksi-5-metil-izoksazol-4-propionska kiselina (AMPA) nađeno je da selektivni agonist AMPA receptorskog podtipa ili (±) -trans (1) -amino-1,3-ciklopentan dikarboksilne kiseline (ACPD), selektivni agonist metabotropnog receptorskog podtipa, može inducirati erekciju penisa kada se ubrizgava u paraventricularna jezgra slobodno pokretnih štakora (Melis et al., 1994b). Pro-erektilni učinak 948 MR Melis, A. Argiolas / Neuroscience i Biobehavioral Reviews 35 (2011) 939-955 NMDA spriječen je s (+) MK-801, nekonkurentnim antagonistom NMDA receptora (Woodruff et al., 1987). ), injektirano u paraventricularnu jezgru (Melis et al., 1994b). U skladu s tim rezultatima, u telemetrijskim studijama s ciljem praćenja intrakavernoznog tlaka, NMDA je nađen puno aktivniji od agonista drugih podtipova receptora ekscitatornih aminokiselina kada se ubrizgava u paraventricularnu jezgru u povećanju intrakavernosnog tlaka kod budnih ili anesteziranih štakora (Zahran et al., 2000, Chen i Chang, 2003).
Kao što je gore rečeno za oksitocin i dopamin, vjerojatno je da su NMDA receptori koji posreduju u erekciji penisa smješteni u staničnim tijelima oksitocinergičkih neurona, jer ekscitatorski završetci živčanih aminokiselina udaraju u tijela oksitocinergičkih stanica u paraventricular jezgri. (Van Den Pol, 1991). Analogno onome što je pronađeno kod agonista dopaminskih receptora, proerektilni učinak NMDA očigledno je posredovan aktivacijom oksitocinergičke neurotransmisije, koja je ukinuta antagonistom oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocinom danim icv, ali ne u paraventricularnu jezgru (vidi Argiolas i Melis, 1995, 2004, 2005 i reference tamo). Slično tome, NMDA-inducirana aktivacija oksitocinergičke neurotransmisije je također sekundarna za aktivaciju dušik-oksid-sintaze, budući da je NMDA-inducirana erekcija penisa spriječena inhibitorima sintetaze dušikovog oksida (N-Nitro-N-metil-arginin metil ester i N- metil-tio-l-citrulin) dano u paraventricularnu jezgru i NMDA ubrizgano u paraventricularnu jezgru u dozama koje induciraju erekciju penisa, povećava proizvodnju dušikovog oksida u jezgri hipotalamusa (vidi Argiolas i Melis, 1995, 2004, 2005 i reference tamo ). Što se tiče agonista dopaminskih receptora, NMDA inducirana aktivacija dušikove okside može također biti sekundarna povećanom Ca2 + ulasku u oksitocinergična stanična tijela putem Ca2 + kanala-NMD receptora, kao što je prikazano u nekoliko neuronskih pripravaka (za pregled vidjeti Snyder, 1992; Southam i Garthwaite, 1993, Schuman i Madison, 1994 i reference u njemu). Dušikov oksid pak aktivira oksitocinergičku neurotransmisiju (vidi gore). Podrijetlo glutamatergičkih projekcija koje aktiviraju paraventricularne oksitocerergičke neurone koje posreduju u erekciji penisa nisu poznate, iako neki neuroanatomski i elektrofiziološki dokazi upućuju na to da oni mogu nastati, barem djelomično, u hipokampusu (Saphier i Feldman, 1987; Chen et al., 1992). , Iako je potreban daljnji rad kako bi se bolje opisalo podrijetlo glutamatergičnih projekcija na paraventricularnu jezgru, uključenost glutaminske kiseline u paraventricularnu jezgru u kontroli erekcije penisa i seksualnog ponašanja jasno je potvrđena mikrodijalizom. Prema tome, ekstra-stanične koncentracije glutaminske kiseline i asparaginske kiseline povećane su u dijalizatu dobivenom iz paraventricularne jezgre muških štakora koji su pokazali nekontaktnu erekciju kada se stavi u prisutnost nepristupačnih ženki štakora primitivnih estrogena i progesterona (Melis et al., 2004b), erekcije penisa koje su također posredovane aktivacijom središnje oksitocinergičke transmisije (Melis et al., 1999a, b). Takva povećanja nađena su još viša kada je dopušteno kopuliranje s receptivnom ženkom (Melis et al., 2004a). U skladu s hipotezom da se povećava aktivnost ekscitatornih aminokiselina u paraventricularnoj jezgri tijekom erekcije penisa i kopulacije, i nekontaktne erekcije i kopulacijsko ponašanje (tijekom kojih se javljaju erekcije penisa) smanjuju se blokadom NMDA receptora u paraventricular nucleus i ovo smanjenje slijedi smanjenje porasta proizvodnje dušikovog oksida koji se javlja u ovoj jezgri hipotalamusa u ovim fiziološkim kontekstima (Melis et al., 2000). Povećanje koncentracije izvanstanične glutaminske kiseline u odnosu na smanjeno oslobađanje GABA iz GABAergičnih živčanih završetaka, koje su djelovale na ekscitatorne aminokiselinske sinapse usporedo s oksitocinergičkim staničnim tijelima, također je pronađeno u paraventricularnoj jezgri nakon blokade CB1 receptora od strane CB1 antagonista SR 141761A, dane u lateralne komore ili izravno u paraventricularnu jezgru u dozama koje induciraju erekciju penisa (vidi Succu et al., 2006; Castelli et al., 2007). Takvo povećanje dovelo je do aktivacije sintaze dušikova oksida u staničnim tijelima oksitocinergičkih neurona, povećavajući proizvodnju dušikovog oksida. Dušikov oksid zauzvrat aktivira oksitocinergičke neurone koji posreduju erekciju penisa kao što je gore opisano. U skladu s takvim mehanizmom, erekcija penisa izazvana SR 141761A smanjena je blokadom NMDA receptora i inhibitorima sintetaze dušikovog oksida, ali ne i blokadom receptora dopamina ili oksitocina u paraventricularnoj jezgri, dok je blokada središnji oksitocinski receptori s antagonistima oksitocinskog receptora s icv
3.3. Interakcija oksitocina i dopamina u ventralnom tegmentalnom području
Oksitocin izaziva erekciju penisa kada se ubrizgava u kaudalni dio ventralnog tegmentalnog područja na način ovisan o dozi (Melis et al., 2007). Najmanja unesena aktivna doza bila je 20 ng, dok je najviša ispitana doza bila 100 ng. Učinak oksitocina je posredovan aktivacijom oksitocinergičkih receptora, budući da je seksualni odgovor ukinut prethodnom injekcijom antagonista oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin u kaudalnom ventralnom tegmentalnom području. Ovi receptori su lokalizirani u staničnim tijelima dopaminergičkih neurona, koji se uglavnom projektiraju u ljusku nucleus accumbens. Prema tome, (1) dvostruke imuno-fluorescentne studije pokazuju da su u kaudalnom ventralnom tegmentalnom području oksitocinergička vlakna u bliskom kontaktu s staničnim tijelima dopaminergičkih neurona, od kojih je većina pozitivno obilježena za tirozin-hidroksilazu i koja sadrži retrogradni tragač Fluorogold ubrizgane u ljusku nukleusa accumbens (Melis et al., 2007) i (2) ventralne tegmentalne zone penisa penisa izazvana oksitocinom dolazi istodobno s povećanjem koncentracije izvanstaničnog dopamina u dijalizatu dobivenom iz ljuske nucleus accumbens (Melis et al., 2007). Erekcija penisa izazvana oksitocinom također se javlja istodobno s povećanjem proizvodnje dušikovog oksida u ventralnom tegmentalnom području, pri čemu su oba odgovora antagonizirana ne samo d (CH2) 5Tyr (Me) 2-Orn8-vazotocinom, već i inhibitorom sintaze dušikovog oksida S- metil-tio-l-citrulin, ali također i _-konotoksinom, blokatorom Ca2 + kanala ovisnim o naponu, i ODQ (1H- [1,2,4] oksadiazol [4,3-a] kinoksalin-1-on), snažan inhibitor guanilata ciklaze, sve dane u kaudalnom ventralnom tegmentalnom području prije oksitocina (Succu et al., 2008). Budući da su mnoga dopaminergična stanična tijela obilježena s Fluorogoldom, koja su bila u kontaktu s oksitocinergičkim vlaknima, za koje je utvrđeno da su pozitivna na tirozinsku hidroksilazu u kaudalnom ventralnom tegmentalnom području, također su pozitivno obilježena za sintazu dušikova oksida i gvanilat ciklazu (Succu et al., 2008), oksitocin erekcija penisa može biti posredovana sljedećim mehanizmom. Aktivacija oksitocinergičkih receptora u tijelima dopaminergičnih stanica peptidom povećava Ca2 + priljev unutar staničnih tijela dopaminergičkih neurona. Time se aktivira sintaza dušikova oksida, enzim koji ovisi o Ca2 + -kalmodulinu, čime se povećava proizvodnja dušikovog oksida. Dušikov oksid opet aktivira gvanilat ciklazu, što dovodi do povećane koncentracije cikličkog GMP. Ciklični GMP aktivira dopaminergične neurone koji se šire u nucleus accumbens. Uloga cikličkog GMP-a u erekciji penisa izazvanoj oksitocinom koji se ubrizgava u kaudalnu ventralnu tegmentalnu površinu također je podržana sposobnošću 8-bromo-cikličkog GMP-a, aktivnog analoga cikličkog GMP-a otpornog na fosfodiesterazu, da inducira erekciju penisa kada se ubrizga u kaudalnog ventralnog tegmentalnog područja, te da se poveća izvanstanični dopamin u dijalizatu iz nucleus accumbens (Succu et al., 2008; Melis et al., 2009a). To je u suprotnosti s mehanizmom kojim dušikov oksid aktivira oksitocinergičke neurone u paraventricularnom jezgru, što je 8-bromo-ciklički GMP koji ne može inducirati erekciju penisa kada se ubrizgava u ovu jezgru (Melis i Argiolas, 1995b) (sl. 2). Što se tiče neuronskih putova aktiviranih dopaminom u nucleus accumbens koji dovode do erekcije penisa, oni su još uvijek nepoznati. Međutim, dostupni podaci upućuju na to da ovi putovi aktiviraju neurotransmisiju dopamina u paraventricularnoj jezgri hipotalamusa. U skladu s tim, erekcija penisa izazvana oksitocinom javlja se istovremeno s povećanjem ekstra-staničnog dopamina ne samo u nucleus accumbens, ali također i u paraventricularnoj jezgri, a antagonizira antagonist dopaminskog receptora haloperidol ubrizgan u paraventricularnu jezgru (Melis et al., 2007). Svi ovi rezultati podupiru ideju da oksitocinergički neuroni potječu iz paraventricularne jezgre i projiciraju se na kaudalnu ventralnu tegmentalnu površinu, kada se aktivira oslobađanje oksitocina u tom području, čime se aktivira NO-ciklički GMP signalni sustav., što zauzvrat aktivira mezolimbičke dopaminergičke neurone (Melis et al., 2007, 2009a; Succu et al., 2008). Dopamin oslobođen u nucleus accumbens zauzvrat aktivira neuronske putove koji dovode do aktivacije in vitro-hipotalamičkih dopaminergičnih neurona, koji stimuliraju paraventricularne oksitocinergičke neurone koji se šire u leđnu moždinu posredujući erekciju penisa. Istovremeno, dopamin oslobođen u paraventricularnoj jezgri također može aktivirati oksitocinergičke neurone koji se projektiraju na područja izvan hipotalamusa kao što su ventralna tegmentalna područja, hipokampus, amigdala i možda druga područja mozga.
Kao što je već spomenuto, u skladu s ovom hipotezom, apomorfin koji se ubrizgava u paraventricularnu jezgru u dozi koja inducira erekciju penisa također povećava izvanstaničnu koncentraciju dopamina u nucleus accumbens, učinak smanjen antagonistom receptora oksitocina d (CH2) 5Tyr (Me ) 2-Orn8-vazotocin ubrizgan u ventralno tegmentalno područje (Succu et al., 2007; Melis et al., 2009a). Zajedno, gornji neuronski putovi mogu predstavljati hipotetski neurološki sklop koji uključuje dopamin, oksitocin i druge neurotransmitere (npr. Glutaminsku kiselinu, vidi dolje) koji utječu ne samo na seksualnu izvedbu, nego i na seksualnu motivaciju i seksualno nagrađivanje. (vidi Odjeljak 4).
3.4. Interakcija oksitocin-glutaminske kiseline u ventralnom subikulumu hipokampusa
Oksitocin inducira erekciju penisa kada se ubrizgava u ventralno, ali ne u dorzalni subikulum, na način ovisan o dozi (Melis et al., 2009b). Seksualni odgovor je posredovan stimulacijom oksitocinskih receptora, ukidanjem prethodne injekcije d (CH2) 5Tyr (Me) 2-Orn8-vazotocina dane na isto mjesto oksitocina, kao što je pronađeno u drugim područjima mozga (vidi gore) , Što se tiče lokalizacije ovih receptora, dostupni podaci sugeriraju da su lokalizirani u staničnim tijelima neurona bogatih sintetazom dušikovog oksida.
Prema tome, studije mikrodijalize pokazuju da se erekcija penisa izazvana oksitocinom događa istodobno s povećanjem proizvodnje dušikovog oksida u ventralnom subikulumu, a to povećanje se ukida ne samo prethodnom injekcijom inhibitora dušične oksidaze S-metil-tio-l-citrulin. i hemoglobinom koji sakuplja dušikov oksid, ali i d (CH2) 5Tyr (Me) 2-Orn8-vazotocinom, sve dane u isto mjesto oksitocina u dozama koje antagoniziraju erekciju penisa (Melis et al., 2010). Još važnije, erekcija penisa izazvana oksitocinom javlja se također istodobno s povećanjem koncentracije ekstra-stanične glutaminske kiseline u ventralnom subikulumu, koji je samo djelomično antagoniziran nekompetentnim antagonistom NMDA receptora (+) MK-801 danim u ventralni subiculum (Melis et al., 2010). Zajedno, ovi rezultati sugeriraju da novoformirani dušikov oksid, djelujući kao međustanični glasnik, aktivira neurotransmisiju glutaminske kiseline koja dovodi do erekcije penisa, moguće kroz neuralne eferentne projekcije iz ventralne subikulum do područja mozga izvan hipokampusa. U skladu s ovom hipotezom, NMDA injektiran u ventralni subikulum inducira erekciju penisa na način ovisan o dozi, a taj se efekt potpuno antagonizira prethodnom injekcijom na isto mjesto (+) MK-801, ali ne i S-metil-tio-l-citrlinom hemoglobin ili d (CH2) 5Tyr (Me) 2-Orn8-vazotocin (Melis et al., 2010). Što se tiče neuronskih eferentnih puteva koji se šire u područja izvan hipokampusa koja se aktiviraju ekscitatornim aminokiselinama (tj. Glutaminskom kiselinom) u ventralnom subikulumu, vjerojatno je da su to glutamatergični, kao i većina hiprentamskih eferentnih projekcija. U ovom trenutku, može se samo sugerirati da ove projekcije moduliraju aktivnost mezolimbičkih dopaminergičkih neurona, što zauzvrat modulira aktivnost in vitro-hipotalamičkih dopaminergičkih neurona u paraventricularnoj jezgri, što dovodi do aktivacije oksitocinergičkih neurona kao što je već spomenuto (vidi gore).
U skladu s tim, erekcija penisa uzrokovana ventralnim subiculum oksitocinom događa se istodobno s povećanjem koncentracije izvanstaničnog dopamina u ljusci nucleus accumbens, a to povećanje, poput erekcije penisa, ukida se d (CH2) 5Tyr (Me) 2- Orn8-vazotocin dan u ventralni subikulum prije oksitocina (Melis et al., 2009b). Štoviše, s obzirom da je ventralna subikulum inducirana erekcija penisa također smanjena s (+) MK-801 ubrizganim u ventralno tegmentalno područje, ali ne u nucleus accumbens (Melis et al., 2009b) i javlja se istodobno s povećanjem izvanstaničnih glutaminsku kiselinu u ventralnom tegmentalnom području, ali ne u nucleus accumbens, oba odgovora ukinuta s d (CH2) 5Tyr (Me) -Orn8-vazotocinom, injektiranim u ventralni subikulum prije oksitocina (vidi sliku 3), vrlo je vjerojatno da je aktivacija mezolimbičkih dopaminergičkih neurona sekundarna zbog povećane glutamatergične neurotransmisije u ventralnom tegmentalnom području. To sugerira da interakcija glutaminske kiseline i dopamina koja kontrolira erekciju penisa postoji u području ventralnog tegmenta. Daljnje studije su potrebne kako bi se utvrdilo jesu li proerektilni eferentni glutametergični putovi od subiculuma do ventralnog tegmentalnog područja izravni ili neizravni, tj. Kroz prefrontalni korteks ili druga područja mozga (vidi Melis et al., 2009b i reference u njemu). Budući da paraventricularna jezgra također prima glutamatergične projekcije iz hipokampusa (vidi gore i Saphier i Feldman, 1987), i glutaminska kiselina aktivira paraventricularne oksitocinergičke neurone, uključujući one koji se šire u ventralno tegmentalno područje (vidi Argiolas i Melis, 2005 i reference u njemu), i oksitocin u ventralnom tegmentalnom području uzrokuje erekciju penisa i povećava aktivnost mezolimbičkog dopaminergičkog neuronas (vidi gore), primamljivo je nagađati da je straraventrikularni oksitocinergički neuroni mogu također biti uključeni, barem djelomično, u aktivaciju mezolimbičkih dopaminergičkih neurona pomoću oksitocina koji se ubrizgava u ventralni subikulum (vidi Odjeljak 4).
4. Zaključne napomene
Prethodno opisane studije potvrđuju i proširuju rana otkrića koja pokazuju da kod mužjaka štakora oksitocin ima ključnu ulogu u središnjoj kontroli erekcije penisa na razini paraventricularne jezgre hipotalamusa i leđne moždine. Konkretno, najvišestudije pokazuju da oksitocin utječe na erekciju penisa također djelovanjem u drugim područjima mozga, tj. ventralnom tegmentalnom području, ventralnom subikulumu i posteromedijalnoj kortikalnoj jezgri amigdale..
Na paraventricularnoj razini, najvažnije novo otkriće je možda otkriće ekspresije svih dopaminskih receptora obitelji D2 (D2, D3 i D4) u staničnim tijelima oksitocinergičkih neurona u paraventricularnoj jezgri (iu supraoptičnom jezgru i srednjeg preoptičkog područja) (Baskerville i Douglas, 2008; Baskerville et al., 2009). To osigurava snažan neuroanatomski podupiru hipotezu da dopaminski i dopaminski receptorski agonisti mogu aktivirati izravno oksitocinergičke neurone uključene u erektilnu funkciju i projektirati ne samo na kičmenu moždinu, nego i na izvan-hipotalamička područja mozga, U tom smislu, također je važno otkriće da je povećanje agonista dopaminskog receptora izazvano povećanjem intkakavernoznog tlaka smanjeno blokadom oksitocinergičkih receptora u lumbo-sakralnoj kralježničnoj moždini (Baskerville et al., 2009). Doista, iako su takvi dokazi dobiveni u anesteziranih mužjaka štakora, nalaz potvrđuje aktivaciju paraventriculo-spinalnog oksitocinergičkog silaznog puta koji je uključen u erekciju penisa izazvanu agonistom dopaminskog receptora. Hunatoč tome, još uvijek se mora utvrditi je li erekcija penisa izazvana stimulacijom dopaminskih receptora prisutnih u tijelima oksitocerergičnih stanica sekundarna u odnosu na aktivaciju specifičnog podtipa dopaminskog receptora obitelji D2 (D2, D3 ili D4) ili ako ti podtipovi receptora sudjeluju u moduliranju erektilnog odgovora, moguće na različite načine, ovisno o kontekstu u kojem se javlja erekcija penisa (vidi Moreland et al., 2004; Enguehard-Gueiffier et al., 2006; Melis i sur., 2006a, b; Löber i sur., 2009; Collins et al., 2009; Depoortère et al., 2009; Baskerville et al., 2009).
Još jedan važan novi nalaz je da oksitocin inducira erekciju penisa kada se ubrizgava ne samo u paraventricularnu jezgru ili CA1 polje hipokampusa, već također u ventralno tegmentalno područje, ventralni subikulum i posteromedijsku kortikalnu jezgru amigdale. Ta područja mozga nisu testirana u ranijim istraživanjima koja pokazuju da je oksitocin povećao broj spontanih epizoda erekcije penisa kod mužjaka štakora, iako primaju poput lumbo-sakralnih oksitocinergičkih projekcija kralježnične moždine iz paraventricularne jezgre. Oksitocin je uistinu bio u stanju povećati spontane epizode erekcije penisa, koje se javljaju u odraslih mužjaka štakora u odsutnosti bilo kakvih seksualnih podražaja, kao što su oni koji potječu od prisutnosti pristupačnog ili nepristupačnog receptivnog (estrogen-progesteron primirani) ovarijektomiranog ženskog štakora ili manipulacija genitalijama, kada se unose u paraventricularnu jezgru i CA1 polje hipokampusa, ali ne u dorzalni subikulum, lateralni septum, kaudatnu jezgru, medijalnu preoptičku površinu, ventromedijalnu jezgru i supraoptičnu jezgru (Melis i sur. , 1986). U svim tim istraživanjima erekcija penisa obično se brojala kada je penis izašao iz omotača penisa promatrač koji nije bio svjestan danih tretmana izravno tijekom eksperimenta ili kasnije promatrajući eksperiment snimljen na video vrpci s aparatom za video kameru. Svaka epizoda erekcije penisa traje min. 0.5-1 i obično je popraćena penisom i / ili fleksijama kukova. U ovih štakora se obično ne radi eksperiment da se utvrdi učinak seksualnog iskustva, starosti ili ako se ovi štakori mogu podijeliti na ispitanike s niskim ili visokim odgovorom na proerektilni učinak oksitocina koji se ubrizgava u različita područja mozga. To vrijedi i za većinu studija o proerektilnom učinku drugih neuropeptida i lijekova koji povećavaju epizode erekcije spina, uključujući dopaminske agoniste, ekscitatorne aminokiseline, ACTH-MSH, heksarelin i VGF peptide. Međutim, pro-erektilni učinak ovih spojeva je opetovano potvrđen telemetrijskim metodama, koje određuju pojavu erekcije penisa povećanjem intrakavernoznog tlaka koji se javlja spontano ili nakon primjene ovih spojeva različitim putevima, tj. Sustavno, intracerebroventrikularno ili izravno u specifične moždane jezgre, nakon implantacije tlačnog mikroprevodnika izravno u kavernozne korpuse (vidi Bernabè et al., 1999). U ventralnom tegmentalnom području, ventralni subikulum i posteromedijalna jezgra amigdale također oksitocin inducira erekciju penisa djelujući na oksitocinergičke receptore. To dovodi do aktivacije mezolimbičkih dopaminergičkih neurona koji potječu iz ventralnog tegmentalnog područja i projiciraju se na ljusku nucleus accumbens, kako je izmjereno povećanjem ekstra-stanične koncentracije dopamina u dijalizatu dobivenom iz ljuske nucleus accumbens i smanjenjem erektilnog odgovora induciranog peptidom ubrizganim u ta izvan-hipotalamička područja, nađen nakon blokade dopaminergičkih receptora u nucleus accumbens (vidi dolje). Što se tiče mehanizama koji se aktiviraju stimulacijom oksitocinergičkih receptora u tim područjima mozga, što dovodi do aktivacije mezolimbičkih dopaminergičkih neurona i erekcije penisa, najbolje su razjašnjena ona koja se pojavljuju u kaudalnom ventralnom tegmentalnom području. Štoviše, rezultati farmakoloških i imuno-fluorescentnih ispitivanja pokazuju da oksitocinski živčani završeci udaraju u stanična tijela dopaminergičkih neurona koji se šire u ljusku nukleusa accumbens (Melis et al., 2007, 2009a; Succu et al., 2008). Mnogi od tih neurona su bogati sintazom dušikovog oksida i gvanilat ciklazom. Stimulacija oksitocinergičkih receptora u staničnim tijelima ovih dopaminergičkih neurona uzrokuje aktivaciju sintaze dušikovog oksida koja dovodi do povećane proizvodnje dušikovog oksida. Dušikov oksid opet aktivira gvanilat ciklazu, čime se povećava koncentracija cikličkog GMP, što dovodi do aktivacije mezolimbičkih dopaminergičnih neurona i oslobađanja dopamina u nucleus accumbens, mjereno povećanjem ekstra-staničnog dopamina u dijalizatu iz nucleus accumbens dobivenog intra-cerebralnom mikrodijalizom (Succu et al., 2008). ). Dopamin koji se oslobađa u nucleus accumbens aktivira neuronske putove koji dovode do erekcije penisa. To je potkrijepljeno sposobnošću antagonista dopaminskih receptora haloperidola i / ili cis-flupentiksola injektiranog u nucleus accumbens da se smanji ventralna tegmentalna površina penisa penisa izazvana erekcijom penisa (Succu et al., 2008). Čini se da jedan od proerektilnih putova aktivira inerto-hipotalamičke dopaminergičke neurone, osobito one koji se projiciraju na stanična tijela paraventrikularnih oksitocinergičkih neurona. Doista, oksitocin koji se ubrizgava u kaudalnu ventralnu tegmentalnu površinu povećava izvanstanični dopamin ne samo u nucleus accumbens nego iu paraventricular jezgri, a blokada dopaminskih receptora u paraventricular jezgri značajno smanjuje ventralnu tegmentalnu područje oksitocin-inducirana penisna erekcija (Succu i sur. Melis et al., 2007, 2008a), 2007, 2009a). Postojanje ovih dopunsko-paraventricularnih dopaminsko-paraventricularnih oksitocin-ventralnih tegmentalnih veza na jezgri Također se sugerira sposobnost pro-erektilne doze apomorfina i agonista PD 4 receptora D168077-a injiciranog u paraventricularnu jezgru da se poveća ekstra-stanični dopamin u ljusci nukleusa accumbens (Succu et al., 2007), odgovor koji je ukinut s d (CH2) 5Tyr (Me) -Orn8-vazotocinom danim u ventralno tegmentalno područje (Melis et al., 2009a, vidi također niže). Međutim, potreban je daljnji rad kako bi se identificirali neuronski putovi koji spajaju nucleus accumbens s in vitro-hipotalamičkim dopaminergičkim sustavom.
Mehanizam kojim oksitocin inducira erekciju penisa i aktivira mezolimbičke dopaminergičke neurone kada se ubrizga u ventralni subikulum ili u posteromedijalnu jezgru amigdale, samo je djelomično shvaćen u ovom trenutku. I u tim područjima oksitocin aktivira vlastite receptore koji dovode do aktivacije sintaze dušikovog oksida, čime se povećava proizvodnja dušikovog oksida. Dušikov oksid zauzvrat aktivira nepoznate eferentne projekcije, koje očito povećavaju glutamatergičnu neurotransmisiju u ventralnom tegmentalnom području. Glutaminska kiselina tada stimulira mezolimbičke dopaminergičke neurone što dovodi do erekcije penisa. Ova hipoteza je podržana uglavnom sposobnošću oksitocina koji se ubrizgava u ventralni subiculum da bi se povećala ekstra-stanična glutaminska kiselina u ventralnom tegmentalnom području (Sl. 3) i nekonkurentnog antagonista NMDA receptora (+) MK-801 ubrizganog u ventralno tegmentalno područje, ali ne u nucleus accumbens, da bi se smanjila erekcija penisa izazvana oksitocinom koji se ubrizgava u ventralni subikulum ili u posteromedijsku jezgru amigdale (Melis et al., 2009b). U ovom trenutku, više detalja je dostupno za ventralnu eroziju penisa uzrokovanu oksitocinom. Erekcija penisa izazvana oksitocinom čini se sekundarnom aktivacijom oksitocinergičkih receptora koji se nalaze u staničnim stanicama neurona koji sadrže sintetazu dušikovog oksida. To uzrokuje povećanje proizvodnje dušikovog oksida, koji aktivira glutamatergičnu neurotransmisiju djelujući kao međustanični glasnik s mehanizmom sličnim onom opisanom za dugoročno pojačavanje (vidi Snyder, 1992; Southam i Garthwaite, 1993; Schuman i Madison, 2004; , U skladu s ovom hipotezom, erekcija penisa izazvana oksitocinom događa se istodobno s povećanjem ekstracelularne glutaminske kiseline u dijalizatu iz ventralnog subikuluma, a stimulacija receptora ekscitatornih aminokiselina u ventralnom subikulumu pomoću NMDA inducira erekciju penisa. Glutaminska kiselina pak aktivira neuralne eferentne projekcije, koje dovode do aktivacije mezolimbičkih dopaminergičkih neurona u ventralnom tegmentalnom području, kao što je gore navedeno. Ako ti mehanizmi djeluju iu posteromedijalnoj jezgri amigdale, trenutno je nepoznata. Nadalje, potrebne su daljnje studije kako bi se dokazalo da su (1) oksitocinergički završetci živaca i receptori u ventralnom subikulumu i posteromedijalnoj jezgri amigdale lokalizirani u staničnim tijelima neurona koji sadrže dušik-oksid-sintazu (2) ako su ti neuroni konačno pod kontrolom ekscitatornih aminokiselinskih (glutamatergičnih) sinapsi i (3) za identifikaciju signalizacijskog sustava aktiviranog dušikovim oksidom. U tom smislu, vrijedno je spomenuti da sposobnost oksitocina da poveća aktivnost nitratnog oksida-sintaze u staničnim tijelima dopaminergičkih neurona u kaudalnom ventralnom tegmentalnom području i još nepoznatim neuronima u ventralnom subikulumu i posteromedijalnoj jezgri amigdale, podsjeća sposobnost oksitocina da aktivira sintetazu dušikova oksida u staničnim tijelima oksitocinergičkih neurona u paraventricularnoj jezgri (Melis et al., 1997). Međutim, dok u ventralnom tegmentalnom području povećana proizvodnja dušikovog oksida u staničnim tijelima dopaminergičkih neurona dovodi do aktivacije tih neurona aktiviranjem gvanilat ciklaze i povećanjem cikličkog GMP-a, to se ne događa u paraventricularnoj jezgri. Prema tome, 8-bromo-ciklički GMP koji se ubrizgava u paraventricularnu jezgru ne inducira erekciju penisa, dok to čini kada se ubrizgava u ventralno tegmentalno područje. Drugi signalni put koji se razlikuje od sustava dušikovog oksidikličnog GMP-a tada je uključen na paraventricularnoj razini u aktivaciji oksitocinergičkih neurona koji posreduju erekciju penisa endogenim i / ili egzogenim dušikovim oksidom (Melis i Argiolas, 1995b; Melis et al., 1997) (Fig , 1). S druge strane, čini se da ciklički GMP u ventralnom tegmentalnom području igra ključnu ulogu u aktivaciji mezolimbičkih dopaminergičnih neurona i povećanju izvanstaničnog dopamina koji se pojavljuje u dijalizatu dobivenom iz ljuske nukleusa accumbens muških štakora. odabrani za prikazivanje ili ne prikazivanje nekontaktne erekcije penisa kada se stavi u prisustvu nepristupačnog ženskog štakora koji je podvrgnut ovarijektomiziranom receptu (estrogen + progesteron). U tim eksperimentalnim uvjetima, kod mužjaka štakora koji su pokazali nekontaktnu erekciju penisa, utvrđeno je da se povećanje ekstra-stanične koncentracije dopamina može očekivati, a to povećanje se dalje povećava, iako samo skromno, inhibitorima fosfodiesteraze koji se daju u kaudalnu ventralnu tegmentalnu površinu (Sanna et al., 2009).
Sposobnost oksitocina koja se ubrizgava u ventralno tegmentalno područje, u ventralni subikulum i u posteromedijalnu kortikalnu jezgru amigdale, zajedno s onom dopaminskih agonista ubrizganih u paraventricularnu jezgru, kako bi se inducirala erekcija penisa i aktivirala mezolimbički dopaminergični neuroni zaslužuje neki komentar. , Prvo, mehanizmi slični onima spomenutim gore mogu biti operativni kada se erekcija penisa odvija u fiziološkom kontekstu, kao što je tijekom kopulacije (kada se dogodi erekcija penisa) ili tijekom nekontaktne erekcije penisa. Ove erekcije su erekcije penisa posredovane feromonom koje se ne mogu razlikovati od onih izazvanih lijekovima ili oksitocinom, koje se javljaju kada se seksualno potentni mužjaci štakora stavljaju u prisustvo nepristupačnog receptivnog ženskog štakora (s ovarijektomiziranim estrogenom i progesteronom) i smatraju se indeksom seksualne aktivnosti. uzbuđenje (Sachs, 1997, 2007). Doista, iako ti rezultati ne pokazuju da oksitocin u tim područjima igra ulogu u erekciju penisa koja se javlja u fiziološkim kontekstima ili nakon primjene lijeka, oni dodatno pojačavaju rana otkrića koja ukazuju na to da ta područja mozga pripadaju onima u kojima se oksitocin daje centralno kako bi se povećao. ne samo spontane epizode erekcije penisa viđene nakon proerektilnih lijekova, već i poboljšanje seksualnog ponašanja muškaraca (i žena) (vidi Argiolas i Melis, 2004 i reference tamo). U skladu s tim, koncentracija oksitocina povećava se u hipokampusu mužjaka štakora tretiranih proerektilnom dozom apomorfina, klasičnog dopaminskog agonista (Melis et al., 1990) i d (CH2) 5Tyr (Me) -Orn8-vazotocin, koji blokira oksitocin Receptori su izuzetno učinkoviti ne samo u pogoršanju ponašanja kopulatora (Argiolas i sur., 1987a), nego i u olakšavajućem učinku apomorfina na muško kopulacijsko ponašanje (Argiolas i sur., 1987b) u spolno snažnih mužjaka štakora, tijekom kojih u kopulama erekcija penisa pojaviti. d (CH2) 5Tyr (Me) -Orn8-vazotocin je također iznimno moćan u smanjenju nekontaktnih erekcija kod seksualno snažnih muških štakora, kada se daje u nanogramskim količinama u lateralne komore, ali ne u PVN (Melis et al., 1999a).
Drugo, mezolimbički dopaminergični neuroni igraju ključnu ulogu u motivacijskim i nagrađivačkim svojstvima prirodnih pojačavajućih podražaja, kao što su hrana, voda i seksualna aktivnost (Fibiger i Phillips, 1988; Wise i Rompre, 1989; Everitt, 1990). Smatra se da dopamin oslobođen iz tih neurona posreduje u transpoziciji motivacijskih aspekata prirodnih stimulansa u ciljana ponašanja, na primjer u slučaju seksualne aktivnosti, traženja seksualnog partnera i seksualnog odnosa kako bi se postigla nagrada i zadovoljstvo. (Goto i Grace, 2005). U skladu s tim, koncentracija izvanstaničnog dopamina povećava se u dijalizatu iz jezgre akumbensa seksualno jakih mužjaka štakora tijekom izlaganja nepristupačnom ženskom štakoru s primarnim ovarijektomiziranim estrogenom i progesteronom, a takvo povećanje bilo je čak i veće kada je dopušteno da muški štakor kopulira s receptivnim ženkama (Pfaus i Everitt, 1995).
Treće, ovi rezultati podupiru hipotezu da neuronski krug spaja paraventricularnu jezgru s ventralnim tegmentalnim područjem izravno ili neizravno (kroz ventralni subikulum i / ili posteromedijalnu kortikalnu jezgru amigdale) i nucleus accumbens, te odavde do nepoznatog. putevi natrag u paraventricularnu jezgru da kontroliraju aktivnost oksitocinergičkih neurona koji se projektiraju na spinalnu moždinu posredujući erekciju penisa i oksitocinergičkih neurona koji se projektiraju na ventralno tegmentalno područje, ventralni subikulum i posteromedijsku kortikalnu jezgru amigdale, modulirajući na taj način aktivnost mezolimbičkih dopaminergičkih neurona (slika 4). Ovaj složeni neuronski krug može igrati ulogu u integraciji neuronskih aktivnosti uključenih u kontrolu konzumatornog (erektilno-ejakulacijskog) i anticipativnog (motivacijskog i nagrađivanog) aspekta muškog seksualnog ponašanja u fiziološkom kontekstu. Doista, izvanstanični dopamin povećava se u nucleus accumbens (Pfaus i Everitt, 1995) i u paraventricularnoj jezgri seksualno jakih mužjaka štakora tijekom izlaganja nepristupačnom receptivnom ženskom štakoru, kada se pojave nekontaktne erekcije, a još više kada je dopušteno kopuliranje npr. kada se u kopulama javljaju erekcije penisa (Melis et al., 2003). Stoga, iako su potrebne daljnje studije kako bi se pojasnila uloga endogenog oksitocina u ventralnom tegmentalnom području, ventralnom subikulumu i amigdali tijekom nekontaktnih erekcija i seksualnog ponašanja, može biti razumno pretpostaviti da je ovaj hipotetički neuronski krug, dok istovremeno doprinosi konzumacijski aspekti seksualnog ponašanja, u isto vrijeme mogu također aktivirati mezolimbički dopaminergički sustav koji osigurava neuralni supstrat za objašnjavanje nagrađivanih svojstava seksualne aktivnosti (Everitt, 1990; Pfaus i Everitt, 1995). U tom smislu, vrijedi spomenuti da je mezolimbički dopaminergički sustav koji se aktivira oksitocinom koji se ubrizgava u ventralno tegmentalno područje isti je aktiviran lijekovima zlouporabe kao što su opijati, kanabinoidi, amfetamin, kokain i alkohol (Tanda et al., 1997), i otkriveno je da oksitocin može smanjiti toleranciju i ovisnost o kokainu, morfinu, alkoholu i kanabinoidima (Kovacs i sur., 1998; Cui et al., 2001). U zaključku, čini se da oksitocin koji se oslobađa ne samo u ventralnom tegmentalnom području, već iu ventralnom subikulumu i posteromedijalnoj kortikalnoj jezgri amigdale, može aktivirati mezolimbičke dopaminergičke neurone, koji mogu biti uključeni u apetitivne i nagrađivane učinke seksualne aktivnosti. , Aktivacija mezolimbičkih dopaminergičkih neurona može biti izravna, preko oksitocinergičkih receptora u staničnim tijelima mezolimbičkih dopaminergičkih neurona, ili neizravno, putem aktivacije neurotransmisije glutaminske kiseline u ventralnom tegmentalnom području.
Dopamin oslobođen u ljusci nucleus accumbens modulira zauzvrat aktivnost in vitro-hipotalamičkih dopaminergičkih neurona u paraventricularnoj jezgri uzrokujući ili erekciju penisa (putem aktivacije oksitocinergičkih neurona koji se šire u leđnu moždinu), ili seksualnu motivaciju i nagradu (putem aktivacije oksitocinergika) neuroni koji se šire u ventralno tegmentalno područje, ventralni subikulum ili posteromedijsku kortikalnu jezgru amigdale). Budući da se dopamin također oslobađa u ljusci nucleus accumbens i u paraventricularnoj jezgri kada se erekcija penisa događa u fiziološkom kontekstu (npr. Nekontaktna erekcija i kopulacija) (Succu et al., 2007; Melis et al., 2003, 2007), vjerojatno da središnji oksitocinergički neuroni sudjeluju u neuronskim krugovima koji posreduju u interakciji između mezolimbičkog i in vitro-hipotalamičkog dopaminergičkog sustava. Ovi neuronski krugovi mogu igrati ulogu ne samo u fazi konzumacije seksualnog ponašanja (npr. Erekcije penisa i kopulacije), već iu seksualnoj motivaciji, seksualnom uzbuđenju i seksualnoj nagradi.
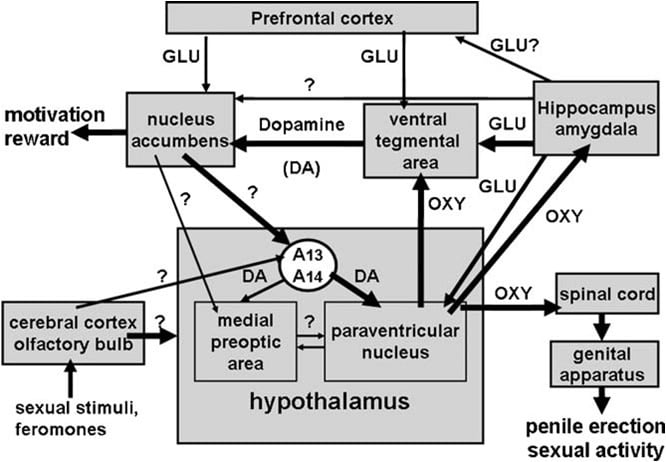
Shematski prikaz hipotetskog živčanog sustava koji uključuje oksitocin koji utječe na seksualnu motivaciju, nagrađivanje i seksualnu izvedbu, što sugeriraju rezultati ovog poglavlja i prethodnih izvješća. Oksitokinergički neuroni koji potječu iz paraventricularne jezgre i projiciraju se na kičmenu moždinu kada se aktiviraju, na primjer, dopaminom i glutaminskom kiselinom (ali i drugim neurotransmiterima i / ili neuropeptidima), olakšavaju erekciju penisa i seksualni učinak aktiviranjem oksitocinergičkih neurona koji se šire u leđnu moždinu. Dopamin i glutaminska kiselina (ali i neurotransmiteri i neuropeptidi) u paraventricularnoj jezgri također aktiviraju oksitocinergičke neurone koji se šire u ventralno tegmentalno područje, aktivirajući tako mezolimbičke dopaminergičke neurone koji se šire u nukleus accumbens, modulirajući seksualnu motivaciju i nagradu. Dopamin oslobođen u nucleus accumbens (NAs) aktivira još uvijek nepoznate neuronske putove, koji povećavaju aktivnost in vitro-hipotalamičkih dopaminergičkih neurona (koji potječu iz skupina A13-A14 Dahlstroma i Fuxe) koji udaraju na oksitocinergičke neurone, uključujući one koji se projektiraju leđne moždine, što dovodi do erekcije penisa. Ovaj sklop može također biti aktiviran oksitocinom koji se ubrizgava ne samo u kaudalnu ventralnu tegmentalnu površinu, nego također u ventralni subikulum i amigdalu, koja također prima oksitocinergičku inervaciju od paraventricularne jezgre, moguće izravnim ili neizravnim glutamatergičnim eferentnim na ventralni tegulacijskom području, što dovodi do modulacije spolne motivacije i erekcije penisa. Konačno, gornji sklop može biti aktiviran i seksualnim podražajima i feromonima, budući da se izvanstanični dopamin i glutaminska kiselina povećavaju u paraventricularnoj jezgri (iu medijalnom preoptičkom području) tijekom feromona posredovane nekontaktne erekcije i kopulacije (za odgovarajuće reference vidi Popis referenci).
Zahvale Ovaj je rad djelomično potpomognut bespovratnim sredstvima od talijanskog ministarstva sveučilišta i istraživanja AA-u i MRM-u
Reference
Andersson, KE, 2001. Farmakologija erekcije penisa. Pharmacol. Rev 53, 417-450. Argiolas, A., 1994. Dušikov oksid je središnji posrednik erekcije penisa. Neurofarmakologija 33, 1339-1344. Argiolas, A., 1999. Neuropeptidi i seksualno ponašanje. Neurosci. Biobehev. Rev 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Središnje funkcije oksitocina. Neurosci. Biobehev. Rev 15, 217-231. Argiolas, A., Melis, MR, 1995. Neuromodulacija erekcije penisa: pregled uloge neurotransmitera i neuropeptida. Prog. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. Uloga oksitocina i paraventrikularne jezgre u seksualnom ponašanju muških sisavaca. Physiol. Behav. 83, 309-317. Argiolas, A., Melis, MR, 2005. Centralna kontrola erekcije penisa: uloga paraventricularne jezgre hipotalamusa. Prog. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. Oksitocin antagonist d (CH2) 5Tyr (Me) -Orn8-vazotocin antagonizira muško kopulacijsko ponašanje kod štakora. E. J. Pharmacol. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Apomorfinska stimulacija muškog kopulacijskog ponašanja spriječena je antagonistom oksitocina d (CH2) 5Tyr (Me) -Orn8-vazotocin u štakora. Pharmacol. Biochem. Behav. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. Intraventrikularni oksitocin izaziva zijevanje i erekciju penisa kod štakora. E. J. Pharmacol. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Oksitocin: iznimno snažan induktor erekcije penisa i zijevanje kod mužjaka štakora. E. J. Pharmacol. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Paraventricularna jezgra lezije sprječava zijevanje i erekciju penisa uzrokovanu apomorfinom i oksitocinom, ali ne i ACTH kod štakora. Brain Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vazotocin, snažan antagonist oksitocina, antagonizira erekciju penisa i zijevanje induciranu oksitocinom i apomorfinom, ali ne ACTH 1-24. E. J. Pharmacol. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. Konotoksin sprječava erekciju penisa izazvanu apomorfinom i oksitocinom i zijeva u mužjaka štakora. Pharmacol. Biochem. Behav. 37, 253-257. Arletti, R., Bertolini, A., 1985. Oksitocin stimulira ponašanje lordoze u ženki štakora. Neuropeptidi 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. Oksitocin poboljšava muške kopulacijske učinke kod štakora. Horm. Behav. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. Seksualno ponašanje starijih štakora stimulirano je oksitocinom. E. J. Pharmacol. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. Seksualna impotencija povezana je sa smanjenom proizvodnjom oksitocina i povećanom proizvodnjom opioidnih peptida u paraventricularnoj jezgri hipotalamusa. Neurosci. Leti. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Dokazi za izravnu projekciju iz paraventricularne jezgre hipotalamusa u pretpostavljene serotoninergičke neurone nukleusa paragigantocelularne koji su uključeni u kontrolu erekcije kod štakora. E. J. Neurosci. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Dušikov oksid: fiziološki posrednik erekcije penisa. Znanost 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Interakcija između dopamina i oksitocina u kontroli seksualnog ponašanja. Prog. Brain Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Interakcija dopamin-oksitocina u erekciji penisa. E. J. Neurosci. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. Dušikov oksid je uključen u muško seksualno ponašanje štakora. E. J. Pharmacol. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Intrakavernozni tlak tijekom erekcije kod štakora: integrativni pristup temeljen na telemetrijskom snimanju. Am. J. Physiol. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MW, Moreland, RB, 2006. Signalizacija receptora dopamina D4 u paraventricularnom jezgru hipotalamusa štakora: dokaz prirodnog povezivanja koji uključuje trenutnu ranu indukciju gena i fosforilaciju protein kinaze aktiviranu mitogenom. Neurofarmakologija 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch 3rd., J., Kolasa, T., Polakowski JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. Aktivacija receptora dopamina D4 pomoću ABT-724-a inducira erekciju penisa kod štakora. Proc. Nat. Acad. Sci. sad 101, 6758-6763. Buijs, RM, 1978. Intra- i ekstra-hipotalamički vazopresin i putevi oksitocina u štakora. Cell Tissue Res. 192, 423-435. Buijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984. Dopaminergička inervacija supraoptičke i paraventricularne jezgre. Studija svjetlosne i elektronske mikroskopije. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Oksitocin olakšava seksualnu receptivnost ženki štakora liječenih estrogenom. Neuropeptidi 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Dopaminergička stimulacija koncentracije oksitocina u plazmi muških i ženskih majmuna apomorfinom i agonistom D2 receptora. J. Clin. Endocrinol. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Organizacija projekcija iz medijske jezgre amigdale: PHAL studija na štakoru. J. Comp. Neural. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Oksitocin u plazmi se povećava u ljudskom seksualnom odgovoru. J. Clin. Endocrinol. Metab. 64, 27-31. Carter, CS, 1992. Oksitocin i seksualno ponašanje. Neurosci. Biobehev. Rev 16, 131-144. Carter, CS, Lederhendler, II, Kirkpatrick, B., 1997. Interaktivna neurobiologija pripadnosti, Annali Akademije znanosti u New Yorku, sv. 807. Njujorška akademija znanosti, New York. Castelli, MP, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. 2007. Kanabinoidni CB1 receptori u paraventricularnoj jezgri i središnja kontrola erekcije penisa: imunocitokemija, autoradiografija i studije ponašanja. Neuroznanost 147, 197-206. Chen, KK, Chang, LS, 2003. Utjecaj agonista receptora ekscitatornih aminokiselina na erekciju penisa nakon primjene u paraventricularnu jezgru hipotalamusa kod štakora. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Elicitacija erekcije penisa nakon aktivacije formiranja hipokampusa kod štakora. Neurosci. Leti. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Dopaminergička neurotransmisija u paraventricularnoj jezgri hipotalamusa u središnjoj regulaciji erekcije penisa kod štakora. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Rice, KC, muževi, SM, Greedy, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen J., Wang, S., Katz, JL, Grandy, DK, Sunahara, RK, Woods, JH, 2009. Proerektilne učinke agonista sličnih dopaminD2-u posreduju receptori D3 u štakora i miševa. J. Pharmacol. Exp. Ter. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Središnja regulacija ejakulacije. Physiol. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Prevencija sindroma povlačenja kanabinoida od litija: uključivanje oksitocinergičke neuronske aktivacije. J. Neurosci. 21, 9867-9876. Dahlstrom, A., Fuxe, K., 1964. Dokazi za postojanje neurona koji sadrže monoamin u središnjem živčanom sustavu. I. Pokazivanje monoamina u staničnim stanicama neurona moždanog debla. Acta Physiol. Scand. 62 (Dodatak. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Erekcija penisa i zijevanje izazvani dopaminskim D2-agonistima u štakorima: utjecaj soja i doprinosa dopaminskog D2-a, ali ne i D3 i D4 receptora. Behav. Pharmacol. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Oksitocin smanjuje odgovore amigdala na emocionalna lica bez obzira na valenciju. Biol. Psihijatrija 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Regulira se medijalnom amigdalom kopulacije i medijalnim preopticnim oslobađanjem dopamina. J. Neurosci. 21, 349-355. Donaldson, ZR, Young, LJ, 2009. Oksitocin, vazopresin i neurogenetika društvenosti. Znanost 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Thompson, JT, Moses, J., Hull, EM, 1991. D2 receptori u paraventricularnoj jezgri reguliraju genitalne odgovore i kopulaciju u mužjaka štakora. Pharmacol. Biochem. Behav. 39, 177-181. Ebner, K., Bosch, OJ, Krömer, SA, Singewald, N., Neumann, ID, 2005. Oslobađanje oksitocina u središnjoj amigdali štakora modulira ponašanje u suočavanju sa stresom i oslobađanje ekscitatornih aminokiselina. Neuropsihofarmakologija 30, 223-230. Enguehard-Gueiffier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-fenilpiperazin-1-il) metil] imidazo (di) azini kao selektivni D4-ligandi. Indukcija erekcije penisa 2- [4- (2-metoksifenil) piperazin-1-ilmetil] imidazo [1,2-a] piridin (PIP3EA), snažan i selektivan D4 agonist. J. Med. Chem. 49, 3938-3947. Everitt, BJ, 1990. Seksualna motivacija: neuronska i bihevioralna analiza mehanizama na kojima se temelje apetitni i kopulatorni odgovori muških štakora. Neurosci. Biobehev. Rev 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Mezokortikolimbički sustav dopamina i nagrada. Ann. N. Y. Acad. Sci. 5, 206-215. French, SJ, Totterdell, S., 2003. Pojedini nukleus akumbens-projekcijski neuroni primaju i bazolateralnu amigdalu i ventralnu subikularnu aferentu u štakora. Neuroznanost 119, 19-31. 954 MR Melis, A. Argiolas / Neuroscience i Biobehavioral Reviews 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Uzbudljivi učinci intraventrikularnih injekcija oksitocina na refleks mlađeg izbacivanja štakora. Neurosci. Leti. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Elektrofiziološki dokazi za olakšavanje kontrole oksitocinskih neurona oksitocinom tijekom sisanja štakora. J. Physiol. (Lond.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Somatodendritički autoreceptori na oksitocin neuronima. U: Ivell, R., Russel, JA (Ur.), Oksitocin, stanični i molekularni pristupi u medicini i istraživanju. Adv. Exp. Med. Biol., 365. Plenum Press, New York i London, str. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Farmakološka svojstva i anatomska raspodjela mjesta vezanja oksitocina 3H u mozgu štakora Wistar proučavana je autoradiografijom. Neuroznanost 20, 599-614. Giuliano, F., Rampin, O., 2000. Centralna kontrola erekcije penisa. Neurosci. Biobehev. Rev 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamin i seksualna funkcija. Int. J. Impotencija Res. 13 (Dodatak. 3), 18-28. Giuliano, F., Rampin, O., 2004. Neuralna kontrola erekcije. Physiol. Behav. 83, 189-201. Giuliano, F., Bernabè, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Spinalni proerektilni učinak oksitocina u anesteziranih štakora. Am J. Physiol. Regul. Integ. Comp. Physiol. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Dopaminergička modulacija limbičkog i kortikalnog pogona nucleus accumbens u ciljno usmjerenom ponašanju. Nat. Neurosci. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Učinci lezija u hipotalamičkim paraventricularnim, supraoptičkim i suprachiasmatic jezgrama na vazopresin i oksitocin u mozgu štakora i leđne moždine. Brain Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Moon, MW, 1997. Sinteza i biološka aktivnost (R) -5,6-dihidro-N, N-dimetil-4H-imidazo [4,5,1-ij] kinolin-5-amina) i njegovih metabolita. J. Med. Chem. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Središnji mehanizmi koji reguliraju erekciju penisa kod svjesnih štakora: dopaminergički sustavi povezani s proerektilnim učinkom apomorfina. J. Pharmacol. Exp. Ter. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Ciljano prekidanje neuronskog gena sintetaze dušikova oksida. Ćelija 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Vazopresin i oksitocin pobuđuju različite neuronske populacije u središnjoj amigdali. Znanost 308, 245-248. Hull, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. Odnos D2 / D1 u medijalnom predoptičkom području utječe na kopulaciju mužjaka štakora. J. Pharmacol. Exp. Ter. 251, 422-427. Hull, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Izvanstanični dopamin u medijalnom predoptičkom području: implikacije za seksualnu motivaciju i hormonsku kontrolu kopulacije. J. Neurosci. 15, 7465-7471. Trup, EM, Meisel, RL, Sachs, BD, 2002. Muško seksualno ponašanje. U: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Ur.), Hormoni, mozak i ponašanje. Academic Press, New York, str. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, KM, 2010. Oksitocin pojačava ovisnost o amigdali, društveno ojačano učenje i emocionalnu empatiju kod ljudi. J. Neurosci. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oksitocin: stanični i molekularni pristupi u medicini i istraživanju. Napredak u eksperimentalnoj medicini i biologiji, sv. 395. Plenum Press, New York. Kelley, AE, Domesick, VB, 1982. Distribucija projekcije iz formacije hipokampusa u nucleus accumbens u štakora: anterogradska i retrogradna studija peroksidaze iz hrena. Neuroznanost 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. Aktivacija receptora 5-HT (2C) uobičajeni je mehanizam za proerektilne učinke apomorfina, oksitocina i melanotana-II kod štakora. E. J. Pharmacol. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Važnost medijske amigdale u erekciji penisa kod štakora izazvana daljinskim podražajima od estrusnih ženki. Behav. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Oksitocin i ovisnost: pregled. Psihoneuroendokrinologija 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Oksitocin: veliki facilitator života. Prog. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Selektivna istokemijska demonstracija dopaminskih terminalnih sustava u di- i telencefalonama štakora: novi dokazi za dopaminergičku inervaciju jezgre neurosekretora hipotalamusa. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Poremećaj seksualnog odgovora nakon lezija paraventricularne jezgre hipotalamusa kod mužjaka štakora. Behav. Neurosci. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Okvir azulena kao novi bioizoster: dizajn snažnih liganda dopaminskog D4 receptora koji induciraju erekciju penisa. Chem. Med. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. Konotoksin: izravna i trajna blokada specifičnih vrsta kalcijevih kanala u neuronima, ali ne i mišića. Prot. Nat. Acad. Sci. sad 84, 4327-4331. McKenna, KE, 2000. Neki prijedlozi u vezi s organizacijom kontrole središnjeg živčanog sustava za erekciju penisa. Neurosci. Biobehev. Rev 24, 535-540. Marson, L., McKenna, KE, 1992. Uloga 5-hidroksitriptamina u silaznoj inhibiciji spinalnih spolnih refleksa. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. Stanične skupine CNS-a su uključene u kontrolu mišića ischiocavernosus i bulbospongiosus: transneuronska studija praćenja pomoću pseudorabies virusa. J. Comp. Neural. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Fiziologija seksualnog ponašanja muškaraca. U: Knobil, E., Neil, J. (Ur.), Fiziologija reprodukcije, sv. 2, drugo izdanje. Raven Press, New York, str. 3-96. Melin, P., Kihlstrom, JE, 1963. Utjecaj oksitocina na seksualno ponašanje u muških kunića. Endokrinologija 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopamin i seksualno ponašanje. Neurosci. Biobehev. Rev 19, 19-38. Melis, MR, Argiolas, A., 1995b. Donori dušikovog oksida izazivaju erekciju penisa i zijevanje kada se ubrizgavaju u središnji živčani sustav mužjaka štakora. E. J. Pharmacol. 294, 1-9. Melis, MR, Argiolas, A., 2003. Središnja oksitocinergička neurotransmisija: cilj lijeka za terapiju psikogene erektilne disfunkcije. Curi. Ciljevi lijeka 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Oksitocin-inducirano zijevanje i erekcija penisa: mjesto djelovanja u mozgu. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Apomorfinsko zijevanje i erekcija penisa: mjesto djelovanja u mozgu. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Apomorfin povećava razinu oksitocina u plazmi kod štakora. Neurosci. Leti. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Dokazi da apomorfin izaziva erekciju penisa i zijevajući otpuštanjem oksitocina u središnjem živčanom sustavu. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Utjecaj apomorfina na koncentracije oksitocina u različitim područjima mozga i plazmi mužjaka štakora. E. J. Pharmacol. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Efekcija penisa izazvana apomorfinom i oksitocinom i zijevanje kod netaknutih i kastriranih mužjaka štakora: učinak spolnih steroida. Neuroendokrinologija 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. Erekcija penisa i zijevanje inducirani paraventricularnim NMDA injekcijama posredovani su oksitocinom. Pharmacol. Biochem. Behav. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Prevencija pomoću NG-nitro-l-arginin metil estera apomorfne i oksitocin-inducirane erekcije penisa i zijevanja: mjesto djelovanja u mozgu. Pharmacol. Biochem. Behav. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Agonisti dopamina povećavaju proizvodnju dušikovog oksida u paraventricularnoj jezgri hipotalamusa: korelaciju s erekciju penisa i zijevanje. E. J. Neurosci. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Oksitocin povećava proizvodnju dušikovog oksida u paraventricularnom jezgru hipotalamusa: korelacija s erekciju penisa i zijevanje. Reg. Peptidi 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Proizvodnja dušikovog oksida povećava se u paraventricularnoj jezgri hipotalamusa muških štakora tijekom beskontaktne erekcije penisa i kopulacije. E. J. Neurosci. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Antagonist oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin smanjuje erekciju penisa bez kontakta kod mužjaka štakora. Neurosci. Leti. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Morfin koji se ubrizgava u paraventricularnu jezgru hipotalamusa sprječava beskontaktnu erekciju i narušava kopulaciju: uključivanje dušikovog oksida. E. J. Neurosci. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Učinak antagonista ekscitatornih aminokiselina, dopamina i oksitocina na nekontaktnu erekciju penisa i proizvodnju paraventricularnog dušikovog oksida u mužjaka štakora. Behav. Neurosci. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Izvanstanični dopamin povećava se u paraventricularnoj jezgri muških štakora tijekom seksualne aktivnosti. E. J. Neurosci. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Antagonizam kanabinoidnih CB1 receptora u paraventricularnoj jezgri muških štakora izaziva erekciju penisa. Neurosci. Leti. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Povećanje ekstracelularnih ekscitatornih aminokiselina u paraventricularnoj jezgri muških štakora tijekom spolne aktivnosti: glavna uloga receptoraNMDA u erektilnoj funkciji. E. J. Neurosci. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, selektivni agonist dopaminskog D4 receptora, inducira erekciju penisa kada se ubrizga u paraventricularnu jezgru muških štakora. Neurosci. Leti. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. Erekcija penisa izazvana SR 141716A kod mužjaka štakora: uključivanje paraventrikularne glutaminske kiseline i dušikovog oksida. Neurofarmakologija 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, A., 2006b. PIP3EA i PD168077, dva selektivna agonista dopaminskog D4 receptora, induciraju erekciju penisa kod mužjaka štakora: mjesto i mehanizam djelovanja u mozgu. E. J. Neurosci. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Oksitocin ubrizgan u ventralno tegmentalno područje inducira erekciju penisa i povećava izvanstanični dopamin u nucleus accumbens MR Melis, A. Argiolas / Neuroscience i Biobehavioral Reviews 35 (2011) 939-955 955 i paraventricularna jezgra hipotalamusa muških štakora. E. J. Neurosci. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Uloga oksitocina u fazama predviđanja i konzumacije spolnog ponašanja mužjaka štakora. U: Jastrow, H., Feuerbach, D. (Eds.), Priručnik za istraživanje oksitocina: Sinteza, skladištenje i otpuštanje, aktivnosti i oblici lijekova. Nova Publishers Inc, New York, SAD, str. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Oksitocin koji se ubrizgava u ventralni subikulum ili posteromedijalnu kortikalnu jezgru amigdale inducira erekciju penisa i povećava izvanstanični dopamin u nukleusu accumbens muških štakora. E. J. Neurosci. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. Oksitocin izaziva erekciju penisa kada se ubrizgava u ventralni subikulum: uloga dušikovog oksida i glutaminske kiseline. Neurofarmakologija 58, 1153-1160. Moreland, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, EJ, Helfrich, RJ, Namovic, MT, El-Kouhen, OF, Masters, JN, Brioni, JD, 2004. Usporedna farmakologija staničnih linija humanog receptora dopamina D (2) stabilnog tipa povezanog s fluksom kalcija preko Galfe (qo5). Biochem. Pharmacol. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Oslobađanje oksitocina i vazopresina pomoću magnocelularnih jezgara in vitro: specifični olakšavajući učinak oksitocina na vlastito oslobađanje. J. Endocrinol. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Promjene u izlučivanju oksitocina i vazopresina tijekom seksualne aktivnosti u muškaraca. J. Clin. Endocrinol. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Oksitocin je potreban za dojenje, ali nije neophodan za porod ili reproduktivno ponašanje. Proc. Nat. Acad. Sci. sad 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, F., Fletcher, AE, Knowles, M., Marwood, R., Mccallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI 1997. Biološki profil L 745,870, selektivni antagonist s visokim afinitetom za dopamin D4 receptor. J. Pharmacol. Exp. Ter. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Oksitocin u majčinskom, seksualnom i društvenom ponašanju, Annali Akademije znanosti u New Yorku, sv. 652. Njujorška akademija znanosti, New York. Petrović, P., Kalisch, R., Singer, T., Dolan, RJ, 2008. Oksitocin slabi afektivne procjene uvjetovanih lica i amigdalne aktivnosti. J. Neurosci. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Psihofarmakologija seksualnog ponašanja. U: Knobil, FE, Kupfer, DJ Psihofarmakologija: četvrta generacija napretka. Raven Press, New York, str. 742-758. Rajfer, J., Aronson, WJ, Bush, PA, Dorey, FJ, Ignarro, LJ, 1992. Dušikov oksid kao medijator opuštanja kavernoznog tijela u odgovoru na nonadrenergičku, ne-holinergičku neurotransmisiju. N. Engl. J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. Bihevioralni učinci NMDA ubrizgani u hipotalamičnu paraventricular jezgru štakora. Brain Res. 550, 220-224. Sachs, BD, 1997. Erekcija izazvana u mužjaka štakora zrakom mirisa iz estrous ženki. Physiol. Behav. 62, 921-924. Sachs, BD, 2000. Kontekstualni pristupi fiziologiji i klasifikaciji erektilne funkcije, erektilne disfunkcije i seksualnog uzbuđenja. Neurosci. Biobehev. Rev 24, 541-560. Sachs, BD, 2007. Kontekstualna definicija muškog spolnog uzbuđenja. Horm. Behav. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. Suživot NADPH-diaphoraze s vazopresinom i oksitocinom u hipotalamičkim magnocelularnim neurosekretornim jezgrama štakora. Cell Tissue Res. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Inhibitori fosfodiesteraze tipa 5 olakšavaju bezkontaktnu erekciju kod mužjaka štakora: mjesto djelovanja u mozgu i mehanizam djelovanja. J. Seks. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Učinci septalnog i hipokampalnog stimulusa na neurone paraventrikularnih jezgara. Neuroznanost 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Stereotipni odgovori zijevanja potaknuti električnom i kemijskom stimulacijom paraventricularne jezgre štakora. J. Neurophysiol. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Dušikov oksid i sinaptička funkcija. Ann. Rev Neurosci. 17, 153-183. Snyder, SH, 1992. Dušikov oksid: prvo u novom razredu neurotransmitera? Znanost 254, 494-496. Sofroniew, MV, 1983. Vazopresin i oksitocin u mozgu i leđnoj moždini sisavaca. Trendovi Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Nove dopaminske receptore pola desetljeća kasnije. Trends Pharmacol. Sci. 16, 270-275. Southam, E., Garthwaite, J., 1993. Signalni put dušik-oksid-ciklički GMP u mozgu štakora. Neurofarmakologija 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Erekcija penisa i zijevanje inducirani agonistima 5-HT1c kod mužjaka štakora: odnos s dopaminergičkim i oksitocinergičkim prijenosom. E. J. Pharmacol. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. Kanabinoidni antagonist receptora CB1 SR 141716A inducira erekciju penisa povećanjem ekstra-stanične glutaminske kiseline u paraventricularnoj jezgri muških štakora. Behav. Brain Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Stimulacija dopaminskih receptora u paraventricularnoj jezgri hipotalamusa muških štakora izaziva erekciju penisa i povećava izvanstanični dopamin u nucleus accumbens: uključenost središnjeg oksitocina. Neurofarmakologija 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Oksitocin izaziva erekciju penisa kada se ubrizgava u ventralno tegmentalno područje mužjaka štakora: uloga dušikovog oksida i cikličkog GMP-a. E. J. Neurosci. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Kanabinoidi i heroinska aktivacija mezolimbičkog prijenosa dopamina zajedničkim mehanizmom opioidnog receptora mu1. Znanost 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Oksitocinergijska i serotoninergička inervacija identificiranih lumbosakralnih jezgri kontrolira erekciju penisa kod mužjaka štakora. Neuroznanost 82, 241-254. Theodosis, DT, 1985. Sinapsa oksitocin-imunoreaktivnog terminala na oksitocinske neurone u supraoptičkim jezgrama. Nature (London) 313, 682-684. Tindall, JS, 1974. Poticaji koji uzrokuju oslobađanje oksitocina. U: Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Ur.), Priručnik za fiziologiju. Sekta. 7, Endocrinology, sv. IV. Američko društvo za fiziologiju, Washington DC, str. 257-267. Torres, G., Lee, S., Rivier, C., 1993. Ontogenija sintaze dušikova oksida hipotalamičnog štakora i kolokalizacija s neuropeptidima. Mol. Stanica. Neurosci. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Spolno dimorfna ekspresija mjesta vezanja oksitocina u prednjem mozgu i kralježničnoj moždini štakora. Neuroznanost 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Usporedna distribucija vazopresina V1b i oksitocin receptora glasnika ribonukleinskih kiselina u mozgu. Endokrinologija 139, 5015-5033. Van Den Pol, A., 1991. Glutamat i aspartatna imunoreaktivnost u hipotalamičkim presinaptičkim aksonima. J. Neurosci. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G., Giuliano, F. 1999. Oksitokinergička inervacija autonomnih jezgri koja kontrolira erekciju penisa kod štakora. Neuroznanost 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Histokemijsko mapiranje sintaze dušikovog oksida u mozgu štakora. Neuroznanost 46, 755-784. Wagner, CK, Clemens, LG, 1993. Put koji sadrži neurofizin od paraventricularne jezgre hipotalamusa do spolno dimorfnog motornog jezgra u lumbalnoj kralježničnoj moždini. J. Comp. Neural. 336, 106-116. Winslow, JT, Insel, TR, 1991. Društveni status u parovima muških vjeverica određuje bihevioralni odgovor na središnju primjenu oksitocina. J. Neurosci. 11, 2032-2038. Wise, RA, Rompre, P.-P., 1989. Mozak dopamina i nagrada. Ann. Rev Psychol. 40, 191-225. Witt, DM, Insel, TR, 1994. Seksualno ponašanje muškarca aktivira protein sličan c-fosu u oksitocin neuronima u paraventricular jezgri hipotalamusa. J. Neuroendocrinol. 6, 13-18. Witter, MP, 2006. Veze subikuluma štakora: topografija u odnosu na stupčastu i laminarnu organizaciju. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. Interakcija između MK-801 i receptora za N-metil-d-aspartat: funkcionalne posljedice. Neurofarmakologija 26, 903-909. Yamashita, H., Shigeru O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Oksitocin pretežno uzbuđuje navodne oksitocinske neurone u supraoptičkom jezgru štakora in vitro. Brain Res. 416, 364-368. Viče, DP, Hendricks, SE, Prendergast, MA, 1992. Lezije nukleusa paragigantocellularis: učinci na ponašanje parenja kod mužjaka štakora. Brain Res. 596, 73-79. Young, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Nedostatak mišjeg oksitocina sprječava izbacivanje mlijeka, ali ne i plodnost ili porod. J. Neuroendocrinol. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000. Povećanje intrakavernoznog pritiska penisa nakon injekcija agonista receptora ekscitatornih aminokiselina u hipotalamičnu paraventricularnu jezgru anesteziranih štakora. J. Urol.