- 1Centro Interdisciplinario de Neurociencias de Valparaíso, Kitivo cha Sayansi, Universidad de Valparaíso, Valparaíso, Chile
- 2Departamento de Neurociencia, Kitivo cha Ufundi, Universidad de Chile, Santiago, Chile
- 3Núcleo Milenio NUMIND Biolojia ya Shida za Neuropsychiatric, Universidad de Valparaíso, Valparaíso, Chile
- 4Idara ya Baiolojia ya Masi na Masi, Kitivo cha Sayansi ya Baiolojia, Pontifica Universidad Católica de Chile, Santiago, Chile
- 5Idara ya maduka ya dawa na kitengo cha kitaifa cha Neuroscience, Kitivo cha Kemia, Pontifica Universidad Católica de Chile, Santiago, Chile
Nguvu ya tabia inayoelekezwa kwa lengo inadhibitiwa na neuroni za midbrain dopamine. Dysfunctions ya dopaminergic mizunguko huzingatiwa katika madawa ya kulevya na machafuko-ya kulazimisha. Tabia ya kulazimisha ni sifa ambayo shida zote mbili hushiriki, ambayo inahusishwa na neurotransuction ya dopamine iliyoinuliwa. Shughuli ya neurop ya dopamine ya neuropu imedhibitiwa hasa na hatua ya homeopatic ya dopamine kupitia receptors D2 (D2R) ambayo hupunguza kurusha kwa neurons pamoja na awali ya dopamine na kutolewa. Utoaji wa dopamine pia umewekwa na mifumo ya heterologous ya neurotransmitter kama mfumo wa opaidi ya kappa, kati ya wengine. Ujuzi wetu wa sasa wa mfumo wa opaidi ya kappa na ushawishi wake juu ya maambukizi ya dopamine hutoka kwa mifano ya wanyama wa mapema wa magonjwa ya ubongo. Mnamo 1988, kwa kutumia virutubishi vya ubongo, ilionyeshwa kuwa uanzishaji wa papo hapo wa Kappa Opioid Receptors (KOR) unapungua viwango vya daptamini ya dopamine kwenye striatum. Athari ya kinga ya KOR inapinga ushawishi unaowezesha wa madawa ya kulevya kwenye kutolewa kwa dopamine, na kusababisha pendekezo la utumiaji wa agonist wa KOR kama tiba ya dawa kwa ulaji wa lazima wa dawa. Kwa kushangaza, miaka 30 baadaye, wapinzani wa KOR wanapendekezwa kutibu madawa ya kulevya. Je! Nini kingeweza kutokea wakati wa miaka hii ambayo ilibadilisha mabadiliko haya ya dhana? Ushahidi uliokusanywa ulionyesha kuwa athari ya KOR kwenye viwango vya dopamine ya synaptic ni ngumu, kulingana na frequency ya uanzishaji wa KOR na wakati na uchochezi mwingine unaokuja wa neuropu za dopamine, pamoja na tofauti za kijinsia na aina. Kinyume chake kwa athari yake ya papo hapo, uanzishaji sugu wa KOR unaonekana kuwezesha dopamine neurotransication na tabia ya dopamine-Mediated. Vitendo vya kupinga vilivyotolewa na uanzishaji wa papo hapo dhidi ya sugu ya KOR vimehusishwa na athari ya kwanza ya kuhujumu na kuchelewesha, wakati wa kufichua madawa ya dhuluma. Tabia za kulazimishwa zinazosababishwa na uanzishaji wa mara kwa mara wa D2R pia huathiriwa na ushirikiano ulio endelevu wa KOR, ambao unalingana na viwango vya kupunguka vya dopamine na unyeti wa D2R. Kwa hivyo, uhamishaji unaotegemea wakati wa athari za KOR moja kwa moja kwenye viwango vya dopamine vinaathiri kugeuzwa kwa tabia iliyohamasishwa. Uchambuzi huu unachambua mchango wa mfumo wa kappa opioid kwa uingilianaji wa dopaminergic ya tabia ya kulazimisha.
kuanzishwa
Mfumo wa Dopaminergic katika Tabia za Kulazimisha
Kulazimishwa ni kutowezekana kwa kujizuia kutekeleza hatua ya kawaida na matokeo inayojulikana, licha ya athari mbaya (Robbins et al., 2012). Tabia za kulazimisha ni ishara za shida ya kutilia maanani (OCD) na ulevi wa madawa ya kulevya, kati ya magonjwa mengine ya akili. Tabia ya kukagua ni ya kawaida sana katika shida za wigo zinazoonekana zinazoonekana kwa kurudiwa mara kwa mara kwa utaratibu fulani, kwa njia ya kimakusudi au ya kiibada (Williams et al., 2013). Aina anuwai ya tabia ya kawaida (kwa mfano, kukagua, kusafisha, kunawa mikono, n.k) inaweza kugeuka kuwa ya kulazimisha kwa wagonjwa wa OCD na kwa ujumla, hujitokeza kwa kujibu mawazo yanayokazia na yanayosababisha wasiwasi. Vivyo hivyo, kutafuta na kuteketeza dawa za dhuluma kunakuwa lazima kwa watumizi wa dawa za kulevya Kama ilivyo kwa OCD, wasiwasi una jukumu muhimu kusababisha uchukuzi wa utumiaji wa dawa za kulevya kwa wanyonyaji wenye uzoefu. Uharibifu huo huo katika usindikaji wa malipo na adhabu huzingatiwa katika hali zote mbili (Figee na wenzake, 2016), ambayo imesababisha waandishi wengine kujadili OCD kama tabia ya tabia (Holden, 2001).
Mbinu moja inayowezekana inayoongoza kwa tabia ya kulazimishwa imeandaliwa ndani ya nadharia ya kuhamasisha motisho ambayo ni kwamba motisha iliyoimarishwa ("kutaka") kwa dawa inakua wakati wa ulevi bila kuendeleza athari ya kufurahishwa ("liking") (Berridge et al., 1989; Berridge na Robinson, 2016). Kuhimiza uhamasishaji wa mshahara / msukumo wa motisha unahusika katika uchukuaji wa uhamasishaji-uhamasishaji unaohusiana na utaftaji wa dawa za kulevya. Mzunguko wa ujira / msukumo umeundwa na neuroni za tumbo za dopamine za substantia nigra (SN) na eneo la kuvuta vizio (VTA), ambalo linalenga mihemko ya ndani na ya ndani ya mteremko, mtawaliwa. Dopamine neurons kwamba mradi kwa striatum ya cyral au nukta ya kiinitete (jenet) imekuwa jadi kuhusiana na tabia inayoelekezwa kwa malengo, wakati dopamine neurons ambayo mradi huo kwa dorsal striatum umehusishwa na upatikanaji wa tabia (Everitt na Robbins, 2005; Mwenye hekima, 2009; Yager et al., 2015; Volkow et al., 2017).
Sensitization ya malipo / msukumo mzunguko ni kuzingatiwa katika panya kama ongezeko la polepole la shughuli za indomotor inayotokana na utawala unaorudiwa wa kipimo kikali cha dawa inayowezeshwa na dawa (Pierce na Kalivas, 1997; Robinson na Berridge, 2001). Usikivu wa locomotor ni jambo la kudumu kwani linaonekana baada ya wiki, miezi na hata mwaka baada ya uondoaji wa dawa (Robinson na Berridge, 1993). Ilipendekezwa mapema kuwa uhamasishaji wa mzunguko wa malipo / msukumo unachangia utaftaji wa madawa ya lazima ((Robinson na Berridge, 1993). Ipasavyo, uhamasishaji wa locomotor kuwezesha utawala wa kibinafsi wa kokeini unatafuta kurudishwa tena (De Vries et al., 2002). Kwa kuongezea, panya zilizo na ufikiaji mkubwa wa tawala za kahawa zinaonyesha mwitikio mkubwa wa cocaine kuliko panya na ufikiaji mdogo (Ferrario et al., 2005). Kwa kuongezea, mabadiliko ya kimhemko ya msingi wa uhamasishaji wa sauti kwa psychostimulants pia huzingatiwa katika utaftaji wa lazima wa dawa za kulevya (Steketee na Kalivas, 2011; Giuliano et al., 2019). Hizi data zinaunga mkono mawasiliano ya mapema yaliyopendekezwa kati ya uhamasishaji wa sauti na utaftaji wa utaftaji wa madawa unaotekelezwa kwa wanadamu (Robinson na Berridge, 1993; Vanderschuren na Kalivas, 2000). Mechanistically, kurudiwa kwa utawala wa dawa za unyanyasaji kuhisi mizunguko ya dopamine ya mesolimbic inayoongeza dopaminergic neurotransuction. Psychostimulants, kama cocaine au amphetamines, ambayo huzuia transporter ya dopamine ya plasma (DAT), husababisha ongezeko kubwa la dopamine katika nafasi ya synaptic katika striatum na NAc, na hivyo kuamsha uchochezi (Steketee na Kalivas, 2011). Kama ilivyo kwa madawa ya kulevya, uhamasishaji wa malipo ya dopamine / mzunguko wa motisha huchangia kwa tabia za kulazimisha zinazoonekana katika OCD. Kwa kweli, uanzishaji wa kurudiwa wa dopamine D2 receptors (D2Rs) ni wa kutosha kushawishi hisia za tabia na kuangalia tabia katika panya na panya wote (Szechtman et al., 1998; Szechtman et al., 1999; Sun et al., 2019). Utawala unaorudiwa wa quinpirole, agonist ya D2R / D3R, ni mfano uliokubaliwa wa OCD unapoonyesha uhalali wa uso, kupitia nyongeza ya tabia ya kukagua na tabia mbaya, uhalisi wa utabiri, kama inavyoonekana na kupungua kwa tabia ya kulazimisha baada ya matibabu sugu na matibabu ya serotonin Inhibitors (SRI) na huunda uhalali kama miundo ya ubongo inayohusika katika mfano huu inashirikiwa na wale walio kwenye ugonjwa wa magonjwa ya akili (Stuchlik et al., 2016; Szechtman et al., 2017). Kwa muhtasari, uanzishaji unaorudiwa wa maambukizi ya dopamine, ama kwa pre-synaptic (kutolewa kwa dopamine) au utaratibu wa post-synaptic (uanzishaji wa D2R) unaongoza kwa usikivu wa tabia na tabia ya kulazimisha.
Mfumo wa kappa opioid ni moja wapo ya mifumo inayofaa zaidi kudhibiti maambukizi ya dopamine kwenye mzunguko wa malipo / msukumo. Ushahidi unaonyesha kwamba maambukizi ya kappa-opioid inapingana na athari za dopamine; uanzishaji mkubwa wa receptors za kappa opioid (KORs) unashughulikia shughuli za locomotor zilizosababishwa na psychostimulants (Grey et al., 1999). Kwa upande mwingine, uanzishaji wa kurudia wa KOR unadumisha na huongeza utaftaji wa madawa ya lazima na ya kawaida ((Koob, 2013). Matumizi ya dawa za kulevya husababisha ugonjwa unaosababishwa zaidi wa maambukizi ya kappa opioid, labda inachangia hali mbaya za kihemko za dysphoria (Koob, 2013) inasababisha utumiaji wa dawa za dharura (Chavkin na Koob, 2016). Kwa kweli, blockade ya KOR ilizuia kufadhaika- lakini sio kutekelezwa kwa dawa ya nikotini (Jackson et al., 2013), cocaine (Beardsley et al., 2005) na ethanol (Sperling et al., 2010). Sambamba na utaftaji huu, KOR blockade inarudisha mabadiliko ya dopaminergic katika dorsolateral striatum ya amphetamine panya zilizosisitizwa, bila kurekebisha majibu yao ya densi ya kuboreshwa kwa dawa (Azocar et al., 2019). Kwa hivyo, mfumo wa KOR unaonekana kuongeza nguvu haswa inayoongeza thamani ya dawa. Katika OCD, uimarishaji hasi husababishwa na uzingatiaji, ambao huimarisha kulazimishwa kwa nguvu ili kuepusha umati huo. Ingawa haijajaribiwa moja kwa moja, kuimarisha hasi kunaweza kuchukua jukumu kwenye uhamasishaji wa quinpirole. Hakika, D2R inahusika katika kizazi cha uimarishaji hasi. Kwa mfano, uepukaji wa eneo la kujiondoa kwa morphine- halijatengenezwa kwa panya kukosa upungufu wa muda mrefu wa D2R (Smith et al., 2002) na matibabu yanayorudiwa ya quinpirole wakati wa kukomesha yanaimarisha tena cocaine na heroin anayetafuta kwenye dhana ya utawala wa kiotomatiki, athari inayohusiana na uvumbuzi wa kuhamasishwa kwa quinpirole (De Vries et al., 2002), kupendekeza njia zilizoshirikiwa kati ya kisaikolojia na uhamasishaji unaosababishwa na quinpirole. Kwa kuongeza, kuanzishwa kwa ngome ya nyumbani, lakini sio ngome ya riwaya, kwa uwanja wa uwanja-wazi hupunguza uhamasishaji wa tabia na tabia ya kuangalia kwa nguvu (Szechtman et al., 2001), ikionyesha kuwa usalama / usalama unaofahamika unaweza kushindana na tabia mbaya za mazingira zinazopendelea uhamasishaji. Vile vile kwa usisitizo wa kisaikolojia, uhamasishaji unaorudiwa wa KOR kuwezesha uhamasishaji wa locomotor (Escobar et al., 2017) na tabia ya kuangalia ya lazima (Perreault et al., 2007) inayochochewa na utawala unaorudiwa wa quinpirole. Ikiwa athari hii inayoleta athari ni matokeo ya uimarishaji hasi ulioboreshwa kufanywa wazi.
Mchanganuo kamili uliofanywa hivi karibuni unaonyesha kuwa athari ya mfumo wa kappa-opioid kwenye maambukizi ya dopaminergic ni ngumu: inategemea njia ya dopamine inayohusika (Margolis et al., 2006; Margolis et al., 2008), na kwa wakati kati ya uanzishaji wa receptor ya KOR na uanzishaji wa recopor ya dopamine (Chartoff et al., 2016). Sanjari na ugumu huu, matumizi ya matibabu ya nguvu ya KOR yamejadiliwa sana. Imependekezwa kuwa agonist ya KOR inaweza kuwa muhimu kliniki wakati wa awamu ya matumizi ya dawa, ikipatia dawa iliyosababisha hyperdopaminergia (Shippenberg et al., 2007). Kwa upande mwingine, mpinzani wa KOR anaweza kuwa muhimu katika kutibu ugonjwa wa kujiondoa unaosababishwa na kuongezeka kwa usemi wa dynorphin baada ya matumizi ya mara kwa mara ya dawa za kulevya (Wee na Koob, 2010). Ipasavyo, imependekezwa kuwa KOR agonist ya sehemu (Béguin et al., 2012) inaweza kuwa chaguo la matibabu ya kutibu ulaji wa madawa ya kulevya na dalili za kujiondoa kwa watu waliolazwa (Chartoff et al., 2016; Callaghan et al., 2018). Katika hakiki hii, tunachambua modulisho la wakati unaodhamiri / wa mazingira wa dopaminergic ya uingilianaji wa tabia na uvumilivu.
Anatomical na Kazi ya Crosstalk Kati ya Kappa Opioid na Mifumo ya Dopaminergic katika Mikoa ya Striatal na Midbrain
Mikoa ya Jiji
KOR ni vifaa vya receptors vya Gi / o pamoja na protini vilivyoonyeshwa sana katika mfumo wa dopamine ya midbrain (Mansour et al., 1996). Receptors hizi ni za familia ya receptors opioid linajumuisha mu (MOR), delta (DOR) na kappa (KORs). Wanajeshi wa endo asili ya receptors hizi ni endorphins, enkephalin na dynorphin, mtawaliwa. Katika striatum, dynorphin imeundwa na dopamine D1receptor (D1R) - inayojumuisha seli za kiwango cha kati (MSNs) ambazo zina axons za kawaida zinazowezesha KOR kutoka kwa kiini sawa (Mansour et al., 1995). Picha za elektroni za elektroni ya panya NAc zinaonyesha kuwa KOR hupatikana katika miundo ya kifahari yenye vijiti vya DAT wakati sehemu ndogo ya KOR inarifiwa kwenye dendrites katika utekelezwaji wa DAT (Svingos et al., 2001; Kivell et al., 2014). Uchunguzi wa Immunofluorescent unaoonyesha maandalizi ya presynaptic-synaptosomal kutoka NAc yanaonyesha kuwa KORs na D2R hupendelea kwa usawa katika synoptosisi zilizo na dopamine synthetizing enzyme, tyrosine hydroxylase (TH) (Escobar et al., 2017). Kwa kuongezea, KOR ni nyingi katika miili ya seli ya NAc na striatum, na inahusika na D2R katika sehemu ndogo ya seli (Escobar et al., 2017). Kwa ufahamu wa maumbile na Masi, imependekezwa kuwa 20% ya jumla ya kufunga KOR katika striatum inazingatiwa katika vituo vya DA ((Van't Veer et al., 2013). Aidha, Tejeda et al. (2017) ilionyesha kuwa D1R na D2R MSN zinaelezea KOR na upendeleo wa juu kwa D1R iliyo na MSNs (Tejeda et al., 2017). Takwimu hii ya anatomiki inaonyesha kuwa KOR zinakuwepo kabla na postynaptically, kudhibiti dopamine neurotransuction katika mzunguko wa malipo / msukumo.
Njia kadhaa za majaribio zinaonyesha kuwa uanzishaji wa KORs huzuia kutolewa kwa dopaminee. Uanzishaji wa papo hapo wa KOR na sindano ya kimfumo au infusion ya ndani ya agonists hupunguza viwango vya nje vya dopamine katika NAc (Di Chiara na Imperato, 1988; Spanagel et al., 1992; Fuentealba et al., 2006) na dorsal striatum (Gehrke et al., 2008). Kuunga mkono hatua ya kuzuia tonic ya KORs juu ya dotamine neurotransuction, infusion ya moja kwa moja ya antagonist ya muda mrefu na ya kuchagua KOR au binaltorphimine (wala-BNI) (Urembo na al., 1994) inaongeza kiwango cha chini cha dopamine katika NAc (Spanagel et al., 1992) na kutolewa kwa dopamine kwenye dorsal striatum (Azocar et al., 2019). Ushahidi wa mwisho wa kizuizi cha tonic ya KOR ya dopamine ilionyeshwa kwenye panya za KOR, ambayo ilionyesha viwango vya ziada vya dopamine kwenye striatum na NAc (Chefer et al., 2005). Njia zinazohusika na kizuizi cha KOR cha kutolewa kwa dopamine hazijafafanuliwa kabisa. Walakini, inajulikana kuwa uanzishaji wa KORs husababisha kuongezeka kwa K + na kupungua kwa mwenendo wa Ca2 +, na hivyo kushawishi hyperpolarization ya seli na kizuizi cha kutolewa kwa mishipa ya damu (Bruchas na Chavkin, 2010; Margolis na Karkhanis, 2019).
Zaidi ya hayo, katika vitro na katika vivo data ya utendaji unaonyesha kwamba KORs hurekebisha viwango vya dopamine vya nje kwa kurekebisha shughuli za DAT. Kwa mfano, uanzishaji wa KORs kwenye seli za EM4 ambazo zinaelezea kORs na DAT, husababisha kuongezeka kwa dopamine iliyopimwa na voltammetry (Kivell et al., 2014) An ex vivo uchanganuzi pia kwa kutumia voltammetry katika tishu ambazo hazikugawanywa, ilionyesha kuwa sindano ya kimfumo ya KOR agonist U-69593 iliongeza utaftaji wa dopamine katika NAc (Thompson et al., 2000). Kifungu kama hicho cha hivi karibuni kinaonyesha kwamba B -I-BNI haizui ongezeko la uporaji wa dopamine kwenye hali ya hewa ya ndani na ya ndani, iliyosababishwa na sindano kali ya kimfumo ya MP1104, mchanganyiko wa mapokezi ya Kappa / Delta opioid agonist (Atigari na wenzake, 2019). Walakini, athari ya uanzishaji wa KOR juu ya matumizi ya dopamine bado haijaainishwa kikamilifu. Utawala wa kimfumo wa kOR sehemu ya agonist agonist nalmefene ilipungua kipimo cha dopamine ya kuchukua dokiti kwa kutegemewa, imesimamishwa na voltammetry ya kasi ya skirini (FSCV) (Rose et al., 2016). Kutumia kipaza sauti ya flux ya no-net katika panya za kiume za watu wazima, kuzuia KOR kuliambatana na kuongezeka kwa sehemu ya uchimbaji (Ed), ambayo ni hatua isiyo ya moja kwa moja ya dopamine ya kuchukua (Chefer et al., 2006; Azocar et al., 2019), ikionyesha kwamba uanzishaji wa tonic wa KOR unadhibiti udhibiti wa shughuli za DAT (dopamine up). Matokeo haya yanaonyesha jukumu ngumu ya shughuli za KOR za kiasili juu ya matumizi ya dopamine kudhibiti viwango vya nje vya dopamine. Mbinu za juu za utatuzi wa kidunia kama FSCV zimeshindwa kuonyesha athari ya KOR juu ya matumizi ya dopamine (Ebner na wenzake, 2010; Ehrich et al., 2015; Hoffman et al., 2016), ikionyesha kwamba KOR inaongeza shughuli za DAT katika maeneo ya matabaka inahitaji kipindi cha incubat. mistari (Kivell et al., 2014).
Mikoa ya Midbrain
Kujiuliza kwa Autoradiografia iliyofanywa katika kitambara cha panya huonyesha kufungwa muhimu kwa KORs kwenye mhimili wa rostrocaudal wa SN na VTA (Mtaalam et al., 1993). Kwa upande mwingine, data ya microscopy ya elektroni inaonyesha kuwa vituo vyenye dynorphin vyenye hubadilika moja kwa moja kwenye dendrites za TH kwenye SN na VTA (Sesack na Pickel, 1992), na kupendekeza kwamba KORs hubadilika katika vitengo vya somatodendritic ya dopamine neurons. Striatal D1R zenye MSN ni moja ya pembejeo dynorphin kwa midbrain dopamine neurons. Kwa kufurahisha, blockage ya KORs haibadilisha athari ya kuzuia ya D1R-MSNs kwa VTA dopamine neurons, ikionyesha kuwa kizuizi hiki kimepatanishwa na GABA (Edward et al, 2017). KORs hurekebisha majibu ya somatodendritic ya neuropu ya dopamine ya katikati. Uchunguzi wa elektrolojia huonyesha kuwa uanzishaji wa KOR katika hyperpolarizes ya VTA na hupungua kiwango cha kurusha kwa spontaneus ya dopamine neurons (Margolis et al., 2003). Kwa hivyo, infusion ya agonists ya KOR inapungua somatodendritic dopamine efflux (Smith et al., 1992; Dalman na O'Malley, 1999). Walakini, athari hii ya inhibitory ya KORs kwenye neuropu ya dopamine inaonekana kuwa ya utegemezi wa mzunguko. Uingizaji wa agonisi za kappa-opioid katika VTA hupunguza kutolewa kwa dopamine kwenye gamba la tezi za mapema (mPFC) (Margolis et al., 2006) lakini sio katika NAc (Devine et al., 1993; Margolis et al. 2006). Aidha, Margolis na wengine. (2006) iligundua kuwa KORs inazuia VTA dopamine neurons kwamba mradi kwa mPFC na amygdala basol, lakini sio wale ambao mradi wa NAc. Katika mwaka huo huo, Ford na wengine. (2006) ilionyesha kuwa matumizi ya bafu ya wanaweolojia wa KOR katika vipande vya panya ya VTA ilichochea hali ya juu zaidi ya sasa katika dopamine neuron ambayo mradi huo umeenda kwa NAc ukilinganisha na mradi huo kwa amygdala ya basolateral, ikionyesha kuwa KORs ina kizuizi kikubwa zaidi cha neuropu ya dopamine mradi huo wa NAc kuliko kwa amygdala. Kwa kuongezea, uanzishaji wa KOR unapunguza kiwango cha kufurahisha (Margolis et al., 2005) na kizuizi (Ford et al., 2007) mikondo ya postynaptic ndani ya neuroni ya dopamine ya katikati. Tofauti kati ya spishi na projensi ngumu za VTA kwa mPFC na NAc (Van Bockstaele na Pickel, 1995; Carr na Sesack, 2000) ifanye iwezekane kujua ikiwa KORs huzuia kwa hiari baadhi ya idadi ya dopamine ya neuronal katika VTA. Walakini, data iliyofupishwa hapa inaonyesha kwamba KORs ziko kwenye soma na vituo vya neuropu ya dopamine, na vile vile pembejeo ambazo zinasimamia, kwa hivyo zinawekwa katika nafasi nzuri ya kudhibiti shughuli za upatanishi wa neuropu ya dopamine.
Jukumu la KOR Kudhibiti Uwasilishaji wa Dopamini katika Psychostimulants-Ushawishi wa Ushawishi na Tabia za Kulazimisha.
Ulevi wa dawa za kulevya ni mchakato ambao unahusisha utaftaji wa dawa za mwanzoni zinazohusiana na athari zao za kuimarisha-nguvu. Kwa upande mwingine, kulazimishwa ni tabia inayoonekana katika watumizi wa dawa za kulevya. Neuroadaptations kadhaa katika njia za dopaminergic zimependekezwa ili kushughulikia utaftaji wa madawa ya kulevya na ulaji kufuatia udhihirisho wa mara kwa mara wa dawa za kulevya (Everitt na Robbins, 2005; Koob na Volkow, 2016). Mojawapo ya nadharia zilizopendekezwa zinazoongoza ulaji wa madawa ya kulevya ni uhamasishaji wa athari zake za kuimarisha-nguvu (Koob, 2013). Udhibiti wa kizuizi cha mfumo wa opaidi ya kappa kwenye kutolewa kwa dopamine inaweza kuchangia mali isiyoimarisha ya mali ya madawa ya unyanyasaji. Walakini, matokeo ya uanzishaji wa KOR kwenye dopamine neurotransization na madawa ya kulazimisha kutafuta yanaonekana kuwa ngumu na dhahiri ya kupingana. Hakika, kutolewa kwa dopamine kunasababishwa na amphetamine na cocaine kunapatikana kwa usimamizi unaofanana wa agonists wa KOR (Heidbreder na Shippenberg, 1994; Maisonneuve et al., 1994; Thompson et al., 2000) na hata kupungua kujiendesha kwa kokaini (Negus na wenzake, 1997). Kwa kuongeza, KORs hutoa maoni ya kuzuia juu ya kutolewa kwa dopamine ya njia ya mesolimbic kukabiliana na uanzishaji endelevu wa post-synaptic D1R kama inavyotokea na mfiduo wa kurudia wa psychostimulants (Cole et al., 1995; Nestler, 2001). Kwa kushangaza, uanzishaji wa KOR pia unaweza kuwezesha kutolewa kwa dopamine kwenye njia ya malipo / motisha (Fuentealba et al., 2006; Fuentealba et al., 2007) na matumizi ya kisaikolojia (Wee et al., 2009). Fuentealba et al. (2007) ilionyesha kuwa baada ya siku nne kusimamia U69593, agonist wa KOR, iliongezeka amphetamine iliyotolewa kutolewa dopamine katika NAc. Hivi karibuni, ilionyeshwa kuwa kuzuia KORs kurudisha nyuma mabadiliko katika kutolewa kwa dopamine na kuchukua katika hali ya dorsal ambayo hufanyika wakati wa uhamasishaji wa locomotor unaosababishwa na amphetamine (Azocar et al., 2019). Kwa jumla, data hizi zinaonyesha kwamba uanzishaji wa KOR unaweza pia kuchangia katika kuimarisha mali inayofaa ya dawa ya unyanyasaji (Chartoff et al., 2016).
Kwa kuongezea, uanzishaji wa KOR pia unaonekana kuchangia kutafuta nguvu kwa dawa; KORs blockade inapunguza cocaine (Wee et al., 2009), heroin (Schlosburg na wenzake, 2013) na methamphetamine (Whitfield et al., 2015ulaji wa panya na ufikiaji usio na kikomo wa dawa (Wee et al. 2009). Athari hii pia inadhihirishwa katika utaftaji wa dawa unaosababisha mafadhaiko. Kwa mfano, panya wa kugonga KOR hakuonyesha upendeleo wa mahali pa cocaine baada ya kulazimishwa kuogelea kwa shida ya kuogelea (McLaughlin na wenzake, 2006a). Uzuiaji wa KOR hupata upendeleo wa mahali pa nikotini unaosababishwa na udhihirisho wa shida ya kuogelea (Smith et al., 2012). Kwa kufurahisha, kuzuia kwa KOR kunaonyesha kokeini na nikotini inayotokana na mafadhaiko lakini haikuathiri kutafuta ikiwa na changamoto ya dawa (Beardsley et al., 2005; Jackson et al., 2013). Athari za uwezeshaji KOR zilizosababishwa na mafadhaiko zinaonekana kupatanishwa na mzunguko / malipo ya msukumo (Shippenberg et al., 2007; Wee na Koob, 2010). Katika utafiti wa kifahari uliofanywa na Dk. Kauer na kikundi chake, ilionyeshwa kuwa kuzuia KORs katika VTA, labda hapo awali au baada ya mkazo mkubwa, inazuia kujuana tena kwa utaftaji wa cocaine, athari inayohusiana na uokoaji wa muda mrefu- Uwezo wa visukuku vya inhibitory katika neuropu ya dopamine (Graziane et al., 2013; Polter na wenzake, 2014).
Uwezeshaji wa ulaji wa psychostimulants uliotolewa na KORs hutegemea wakati wa dirisha kuhusu utaftaji wa dawa za kulevya. Usimamizi wa KOR agonist U50488 1 h kabla ya mfiduo wa kokaini uwekaji upendeleo wa mahali pa kupikia na kutolewa kwa dopamine ya jamaa aliyetolewa na cocaine katika NAc, wakati athari zingine zinazingatiwa wakati wa kupewa dakika 15 kabla (McLaughlin na wenzake, 2006a; Ehrich et al., 2014). Kutumia msukumo wa kibinafsi Chartoff na wengine. (2016) aligundua kuwa mtaalam wa kOR Salvinorin A, ana athari ya mwanzo na ya kuchelewesha, iliyoambatana na kupungua na ongezeko la kutolewa kwa dopamine katika NAc, mtawaliwa. Hizi data zote pamoja zinaonyesha athari inayotegemea wakati ya uanzishaji wa KOR juu ya mali yenye thawabu ya cocaine, na inaangazia uanzishaji wa upatanishi wa KOR kama mchezaji muhimu wa maendeleo ya utaftaji wa madawa ya dharura.
Quinpirole-Iliyosababisha Sensitization ya Usinifu na Tabia ya Kulazimisha
Ukweli kwamba mfumo wa dopamine unahusika katika kizazi cha uhamasishaji na uvumilivu huimarishwa na tabia inayoonekana katika panya iliyotibiwa na agonist wa D2R, quinpirole. Kwa kifupi, D2Rs ni pamoja na receptors zilizojumuishwa sana katika mzunguko / malipo ya motisha; zinaonyeshwa somatodendritically na kwenye vituo vya axon vya dopamine neurons (Sesack et al., 1994), na uanzishaji wake unapunguza viwango vya dopamine vya nje (Imperato na Di Chiara, 1988). Katika striatum, D2R pia ziko kwenye postynaptically kwenye neurons ya kati ya spiny (Sesack et al., 1994) na uanzishaji wake huzuia njia ya moja kwa moja ikiruhusu shughuli za locomotor.
Dk. Henry Szechtman alianza kusoma athari za quinpirole juu ya tabia ya panya wanaomaliza miaka kumi ya 1980. Matokeo yao ya awali yalionyesha kuwa usimamizi mkali wa quinpirole una athari inayotegemea kipimo kwenye shughuli za locomotor. Kwa viwango vya chini (0.03 mg / kg) hupunguza shughuli za locomotor, wakati kwa viwango vya juu (> 0.5 mg / kg), inaongezeka. (Eilam na Szechtman, 1989). Athari hizi zinahusishwa na uanzishaji wa D2R za umoja wa juu na D2R zilizo chini ya ushirika, mtawaliwa (Usiello et al., 2000). Bila kutarajia, usimamizi unaorudiwa (kila siku nyingine) huongeza ongezeko la polepole na endelevu ya fahari, inafanana na usikivu wa locomotor unaosababishwa na psychostimulants (Szechtman et al., 1993; Szechtman et al., 1994). Athari ya kusisimua ya injini ilionyeshwa kutegemea D2R, kwani upungufu wa panya kwa receptor hii haukuzi uhamasishaji wa alama ya quinpirole (Escobar et al., 2015).
Mwanzoni mwa miaka ya 90, Szechtman na Eilam waliripoti kwamba pamoja na uhamasishaji wa locomotor, panya walikua na tabia inayopendelewa, ambayo inaimarishwa na kila utawala wa quinpirole (Eilam na Szechtman, 1989; Szechtman et al., 1993). Leo, utawala unaorudiwa wa quinpirole ni mfano uliothibitishwa kwa OCD (Szechtman et al., 1999; Szechtman et al., 2001; Eilam na Szechtman, 2005; Stuchlik et al., 2016; Szechtman et al., 2017), kwa kuzingatia uchunguzi kwamba tabia ya panya inazidi kuwa muundo na usibadilikaji, kumbukumbu ya tabia ya kitamaduni tabia ya tabia ya kuangalia kulazimishwa (Szechtman et al., 1998; Szechtman et al., 2017). Uchunguzi wa hivi karibuni unaonyesha kuwa quinpirole inayojirudia pia huchochea tabia ya kulazimisha katika panya, kama vile kuangalia kwa kulazimisha (Sun et al., 2019), tabia ya kubadilika na kutafuna kwa nguvu (Asaoka et al., 2019), mwisho hurudiwa na kizuizi cha D2R kwenye striatum, ikiunga mkono zaidi kwamba uanzishaji wa D2R unaohitajika unahitajika kushawishi tabia za kulazimisha. Pamoja data inaashiria jukumu muhimu la D2Rs katika njia za dopamine ya katikati ya kuhamasisha hisia za uvumbuzi na uvumilivu. Tabia ya kurudia ya quinpirole iliyorudiwa mara kwa maraThompson et al., 2010) na athari za athari za amphetamine (Cope et al., 2010), ikiimarisha wazo kwamba uanzishaji wa D2Rs unasababisha uhamasishaji wa kisaikolojia na kupendekeza utaratibu ulioshirikiwa kati ya uhamishaji wa hisia za quinpirole na psychostimulants. Kwa kufurahisha, athari inayofurahisha ya uanzishaji wa mara kwa mara wa D2R inaonekana kuwa na nguvu kuliko ile inayosababishwa na wanasaikolojia, kwani kila panya linashughulikiwa na hisia za kukuza sauti (Escobar et al., 2015), wakati karibu asilimia sitini ya panya wanahamasisha kupata amphetamine (Escobar et al., 2012; Casanova et al., 2013).
Usikivu wa mwenendo unaosababishwa na uanzishaji wa kurudia wa D2R unaambatana na marekebisho katika mzunguko / malipo. Panya zilizosisitizwa na quinpirole zina sauti ya chini ya dopaminergic katika NAc, inazingatiwa kama basal iliyopungua (Koeltzow na wenzake, 2003) na kutolewa kwa dopamine ya tonic na phasic dopamine (Escobar et al., 2015), kuonyesha kupungua kwa kutolewa kwa dopamine ya mzunguko wa dopamine midbrain. Viwango vya dopamine ya Synaptic katika NAc inadhibitiwa na shughuli za shughuli zote mbili, DAT na dopamine neurons (Goto na Grace, 2008), ambayo katika vivo lina tonic na kupasuka kurusha (Wilson et al., 1977; Neema na Bunney, 1980). Ripoti za hapo awali zinaonyesha kuwa panya zilizo na unyevu wa nukta zinaonyesha idadi ndogo ya neuropu ya dopamine katika tonic na kupasuka kwa risasi katika VTA (Sesia et al., 2013). Pamoja data hizi zinaonyesha kuwa kupungua kwa kutolewa kwa dopamine inayoonekana baada ya unyeti wa quinpirole ni matokeo ya kupungua kwa shughuli ya jumla ya dopamine neurons. Tabia ya kulazimisha na shughuli za kuhamasisha zilizosababishwa na matibabu yanayorudiwa na quinpirole inaweza kuwa matokeo ya uhamasishaji wa D2R, kwa sababu ya kupungua kwa sauti ya dopaminergic katika NAc. Kwa kweli, panya zilizo na hisia za quinpirole zinaonyesha kuongezeka kwa kufungwa kwa dopamine D2R (Culver et al., 2008) na kuongezeka kwa hali ya ushirika wa receptors hizi (Perreault et al., 2007), kuunga mkono dhana hii.
Ushirikiano wa KOR-Dopamine katika Beinviors iliyosukuma Quinpirole
Masomo ya awali kuhusu jukumu la KOR katika tabia za kulazimisha zinazosababishwa na D2R pia zilitoka kwa maabara ya Szechtman. Kikundi hiki kilichunguza usimamizi wa pamoja wa KOR agonist U69593 na quinpirole juu ya shughuli za locomotor. Hasa, waandishi walisimamia sindano za ngozi kwa panya na mchanganyiko U69593 na quinpirole, hadi sindano 8 hadi 10 zilikamilishwa. Kinyume na athari ya hypolocomotor ya U69593 peke yake, hyperlocomotion ilizingatiwa wakati inasimamiwa wakati mmoja na kipimo cha chini (presynaptic) na cha juu (postsynaptic) cha quinpirole. U69593 ilibadilisha athari ya hypolocomotor ya kipimo cha presynaptic cha quinpirole kuwa hyperlocomotion na kuongeza athari ya hyperlocomotor ya kipimo cha postynaptic cha quinpirole (Perreault et al., 2006). Uanzishaji wa KORs pia uliharakisha uhamishaji wa hisia za locomotor na kuathiri athari ya uanzishaji wa D2R, kwa kuwa kiwango cha juu cha uvumbuzi kilichopatikana na kurudia mara mbili ya matibabu athari ya locomotor iliyosababishwa na quinpirole pekee (Perreault et al., 2006; Escobar et al., 2017). Uanzishaji wa KORs pia huharakisha kupatikana kwa tabia ya ukaguzi wa kulazimisha (Perreault et al., 2007). Athari hizi zinazowezekana za KORs juu ya tabia zilizochochewa za quinpirole zinahitaji uanzishaji unaorudiwa wa KOR. Kwa kweli, sindano kali ya KOR agonist U69593 haikurekebisha zaidi shughuli ya locomotor katika panya zilizohamishwa na quinpirole (Escobar et al., 2017). Utaratibu wa uhamishaji wa KOR wa kuchochea D2R haujulikani. Uwezo mmoja ni kwamba mfumo wa kappa opioid wa kienyeji yenyewe ni upatanishi wa uhamasishaji unaotegemea D2R. Walakini, uwezekano huu ulitupwa kwa kuonyesha kwamba usimamizi wa mapema wa NKNI haikurekebisha usisitishaji wa hali ya hewa, ikipendekeza kwamba dynorphin haijatolewa uanzishaji wa D2R (Escobar et al., 2017). Takwimu hii haitoi kwamba dynorphin inaweza kuwa na jukumu la kuhimiza tabia ya kulazimisha, kwa mfano, mkazo unachochea kutolewa kwa dynorphin na uanzishaji wa KOR ambayo inawezesha tabia ya kulazimisha (McLaughlin et al., 2003; McLaughlin na wenzake, 2006a; McLaughlin na wenzake, 2006b).
Crosstalk kati ya D2Rs na KOR ni ngumu na inaonekana hutegemea ikiwa uanzishaji wa receptors zote mbili ni sawa au umetengwa kwa muda. Takwimu za anatomical zinaonyesha kuwa crosstalk kati ya D2Rs na KORs zinaweza kutokea kwa kizazi kwenye axons na soma ya dopamine neurons, na pia postynaptically katika MSNs ya striatum. Ingawa haitoi jukumu la KORs ziko kwenye axons za mifumo mingine ya neva, data ya anatomiki inaangazia jukumu moja kwa moja la KORs kusimamia D2R. Uwezo wa papo hapo au unaorudiwa, uanzishaji wa KOR hupunguza kazi ya D2R ya inhibitory kwenye neuropu ya dopamine. Uchunguzi wa elektronisi ulionyesha kuwa uanzishaji wa papo hapo wa KOR katika dopamine neurons ya VTA na SN huzuia kizuizi cha kizuizi cha muda wa D2R, athari iliyoingiliana na utaratibu wa kabla na wa postynaptic wakati KOR inapunguza kutolewa kwa dopamine na dynorphin inazuia athari ya kuzuia ya kutumika kwa dopamine (Ford et al., 2007). Uchunguzi wa Neurochemical ulionyesha kuwa uanzishaji unaorudiwa wa KORs huzuia kizuizi kilichosababisha D2R kinachochochewa cha kutolewa kwa dopamine katika NAc (Fuentealba et al., 2006). Kwa kuongezea, uanzishaji wa D2R unaofanana na uanzishaji wa papo hapo wa KOR hupunguza kizuizi cha kutolewa kwa dopamine katika NAc ikilinganishwa na athari ya kila receptor peke yake (Escobar et al., 2017). Kwa hivyo, KORs za presynaptic hazifanyi kazi kwa nguvu au kwa kushirikiana na D2R za presynaptic, kwa upande wake, KORs ama kuzuia au athari ya athari ya kinga ya D2R. Njia hii inaweza kuelezea athari ya uanzishaji wa locomotor ya kipimo cha papo hapo cha wanaharakati wa KOR wanaofanana na kipimo cha chini cha quinpirole (Perreault et al., 2006).
Utafiti wa hivi karibuni unaonyesha kuwa uanzishaji wa KOR katika VTA huingiliana tabia ya kulazimishwa iliyopimwa kama tabia ya kuzuia na kuzika kwa marumaru (Abraham et al., 2017), kuimarisha wazo kwamba uanzishaji wa KOR kwa kweli ni kichocheo cha kulazimishwa. Takwimu iliyochapishwa na Margolis na wengine. (2006; 2008) zinaonyesha kuwa mwingiliano wa KOR na D2R unapaswa kuchukua nafasi ya dopamine neurons inayolenga mPFC (Margolis et al., 2006; Margolis et al., 2008). Bila kujali, Ford na wengine. (2006; 2007) iligundua kuwa kizuizi cha KORs cha IP2 kilichoingiliana na IPSC hufanyika kwenye dopamine neurons inayolenga NAc (Ford et al., 2006; Ford et al., 2007). Pamoja data hizi zinaonesha kuwa mwingiliano wa KOR na D2R kwenye eneo la somopodendritic ya neuropu ya dopamine inaweza kutokea kama matokeo ya mgongo katika neuron ya dopamine ileile. Ikiwa hii inatokea kwa makadirio ya mesolimbic au mesocortical bado ni ya ubishani.
Kwa kushangaza, KOR ilipatikana katika MSNs ya NAc (Escobar et al., 2017; Tejeda et al., 2017), na hivyo kuashiria kwamba uwezekano wa tabia ya kulazimisha ya D2R-pia inaweza kutokea kwa hatua moja kwa moja kwenye seli zinazolenga za neuropu ya dopamine. Katika suala hili, inafaa kutaja kuwa utawala unaorudiwa wa U69593 unaongeza kiwango cha D2R katika hali ya juu ya ushirika (Perreault et al., 2007). Takwimu za Neurochemical zinaonyesha kuwa viwango vya dopamine vya ziada vya dopamine vinahusishwa na unyeti wa D2R. Uanzishaji wa KOR haupunguzi zaidi viwango vya nje vya dopamine katika NAc tayari imepungua kwa uanzishaji unaorudiwa wa D2Rs (Escobar et al., 2017), kutoa jukumu la kuharakisha KOR kuongeza kasi au kuwezesha uhamasishaji wa D2R katika NAc kupitia utaratibu huu. Kwa hivyo, KORs inasababisha taratibu za Masi polepole zaidi kuhisi athari za neva na tabia ya D2R, na kupendekeza kuwa uboreshaji wa uhamasishaji wa encomotion inaweza kuwa ni kwa sababu ya adaptynaptic inayoweza kubadilika badala ya athari ya kizazi. Kwa hali hii uanzishaji unaorudiwa wa KORs unaweza kusababisha kizuizi cha njia ya moja kwa moja ya D2R kubadili usawa wa D1R / D2R hadi D1R ikichochea uvumilivu (Kielelezo 1).
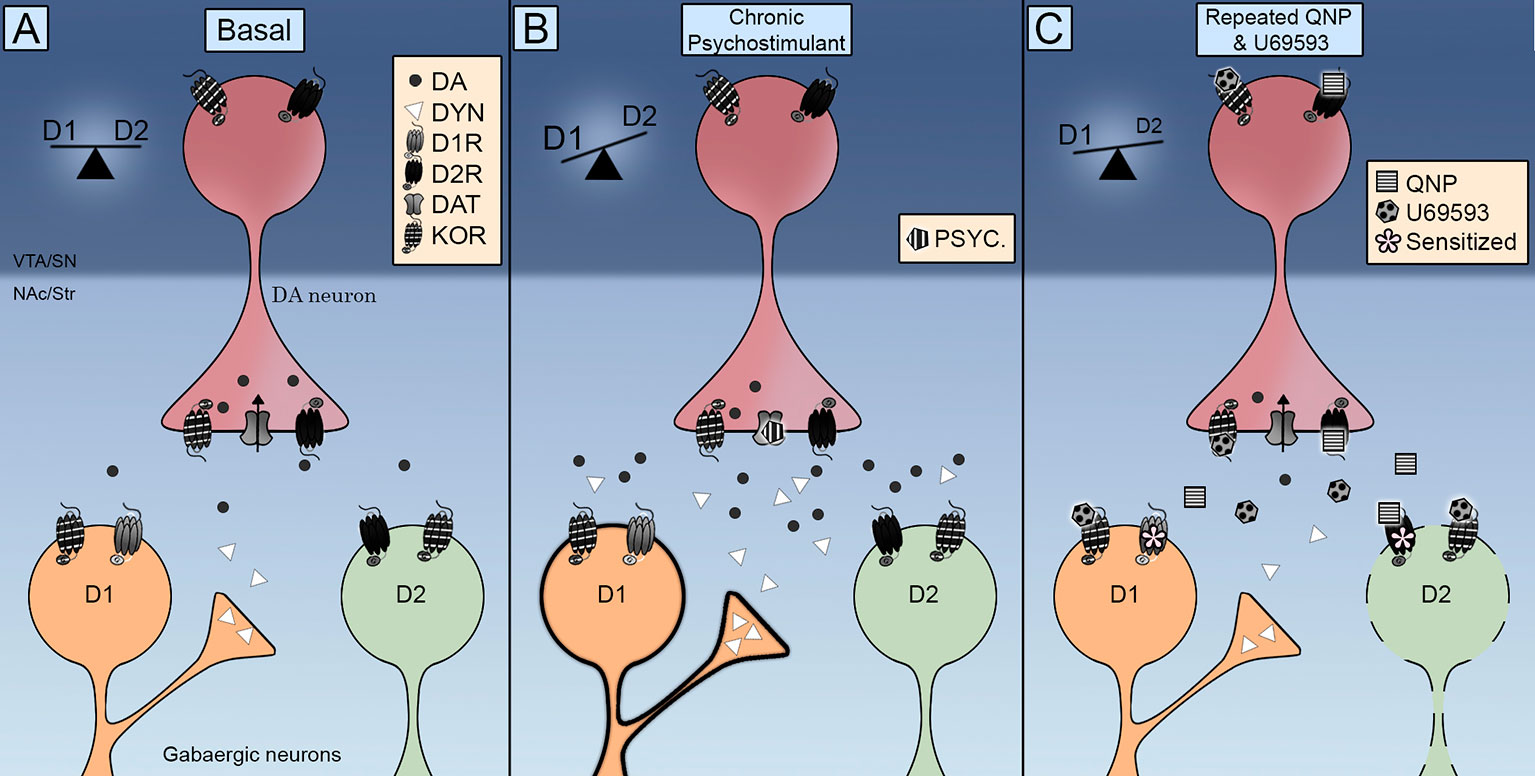 Kielelezo 1 Mpango wa ujumuishaji wa udhibiti wa Kappa Opioid Receptors (KOR) kwenye moja kwa moja (D1R) na moja kwa moja (D2R). (A) KOR ziko kabla ya dhambi kwenye vituo vya dopamine na huchapisha sinaptiki katika seli za kiwango cha kati (MSNs). Uanzishaji wake unadhibiti viwango vya nje vya dopamine na ujanibishaji wake unakuza mwingiliano na dopamine transporter (DAT) na dopamine D2 receptors. (B) Mfiduo wa kurudia wa psychostimulant unaambatana na kuongezeka kwa viwango vya dopamine vya nje na dynorphin. Uanzishaji wa receptors za D1 na D2 hubadilisha usawa kwenye njia ya moja kwa moja ya D1R kukuza uhamasishaji wa locomotor. (C) Usimamizi wa ushirikiano wa quinpirole na U69593 unaambatana na kupungua kwa kiwango cha dopamine extracellular. Uanzishaji wa pamoja wa receptors za KOR na D2 huboresha njia ya moja kwa moja ya D2 inayoongoza tabia ya kulazimisha.
Kielelezo 1 Mpango wa ujumuishaji wa udhibiti wa Kappa Opioid Receptors (KOR) kwenye moja kwa moja (D1R) na moja kwa moja (D2R). (A) KOR ziko kabla ya dhambi kwenye vituo vya dopamine na huchapisha sinaptiki katika seli za kiwango cha kati (MSNs). Uanzishaji wake unadhibiti viwango vya nje vya dopamine na ujanibishaji wake unakuza mwingiliano na dopamine transporter (DAT) na dopamine D2 receptors. (B) Mfiduo wa kurudia wa psychostimulant unaambatana na kuongezeka kwa viwango vya dopamine vya nje na dynorphin. Uanzishaji wa receptors za D1 na D2 hubadilisha usawa kwenye njia ya moja kwa moja ya D1R kukuza uhamasishaji wa locomotor. (C) Usimamizi wa ushirikiano wa quinpirole na U69593 unaambatana na kupungua kwa kiwango cha dopamine extracellular. Uanzishaji wa pamoja wa receptors za KOR na D2 huboresha njia ya moja kwa moja ya D2 inayoongoza tabia ya kulazimisha.
Tofauti za Kijinsia za Mwingiliano wa KOR-Dopamine katika Behaviors zinazosababisha
Uchunguzi wa kliniki umeonyesha tofauti za kijinsia katika tabia ya kulazimisha pamoja na utaftaji wa madawa ya kulevya. Mwanzo wa mapema wa dalili za OCD unazingatiwa kwa wanaume ikilinganishwa na wanawake (Mathis na wenzake, 2011), na wanawake wanaonyesha kiwango cha kuongezeka kwa uchafu na dalili za kusafisha (Labad et al., 2008). Kuhusu tofauti za kijinsia katika ulevi wa dawa za kulevya, ushahidi wa kliniki unaonyesha kwamba wakati utumiaji wa dawa unapatikana sana kwa wanaume, wanawake wanaonyesha maendeleo ya haraka kuliko wanaume katika kutafuta madawa ya dhabiti (Hernandez-Avila na wenzake, 2004; Fattore na Melis, 2016).
Hivi majuzi, ushahidi wa kliniki ya hapo awali umeangazia misingi ya neurobiolojia inayosisitiza tofauti za kijinsia katika unyanyasaji wa dawa za kulevya zinazoonekana katika masomo ya kliniki (Becker na Chartoff, 2019). Uchunguzi wa mapema usio na virutubishi vya flux ya non-net ilionyesha kuwa dopamine extracellular mkusanyiko katika dorsal striatum hutofautiana wakati wa mzunguko wa estrous na viwango vya juu katika proestrus na estrus ikilinganishwa na diestrus. Kwa kuongezea, wakati ovariectomy itapungua densi ya dopamine ya nje ya mkusanyiko katika panya wa kike, kutupwa kwa panya wa kiume hakurekebisha dopamine striatal extracellular mkusanyiko (Xiao na Becker, 1994), kupendekeza jukumu muhimu la homoni za ovari kwenye shughuli za dopamine. Kwa kuongeza, homoni za kike zinasimamia majibu kwa psychostimulants. Mapema vitro majaribio yalionyesha kuwa estradiol pamoja na progesterone inarejeza kutolewa kwa dopamine ya amphetamine kutoka kwa tishu za striatal zilizopatikana kwa panya wa kike ovariectomized (Becker na Ramirez, 1981). Hivi karibuni, uchunguzi wa haraka wa mzunguko wa volclic umeonyesha kuwa wanawake wanaonyesha kutolewa kwa dopamini ya umeme na kuchochea zaidi ikilinganishwa na wanaume (Walker et al., 2000). Tofauti hizi za kimapenzi katika dopamine neurotransuction zinaweza kusababisha cocaine ya juu na amphetamine inayotazamwa katika wanawake. (Roberts et al., 1989; Cox et al., 2013).
Udhibiti wa KOR juu ya viwango vya dopamine vya nje pia unaonyesha tofauti za kijinsia (Chartoff na Mavrikaki, 2015). Kutumia kujisisimua kwa ndani na volteli ya volclic, Conway et al. (2019) ilionyesha kuwa unyeti wa chini wa athari ya anhedoni ya papo hapo ya agonist ya KOR inayotazamwa katika panya wa kike ukilinganisha na panya wa kiume, inaambatana na kizuizi kilichowasilishwa cha kutolewa kwa dopamine katika NAc (Conway et al., 2019). Imependekezwa kuwa estradiol inachangia kizuizi kilichotiwa wazi cha kutolewa kwa dopamine kinachozingatiwa katika panya za kike baada ya uanzishaji wa KOR (Abraham et al., 2018). Wakati crosstalk kati ya KOR na ishara dopamine imesomwa kwa wanaume (Tejeda na Bonci, 2019), utafiti juu ya mwingiliano huu na athari zake katika mchakato wa ulevi katika wanawake ni kukosa (Chartoff na Mavrikaki, 2015). Katika panya za kike, utawala wa papo hapo wa KOR agonist U69593 umepata hyperlocomotion ya cocaine iliyoingiliana katika panya zote mbili, kudhibiti na panya ovari. Kwa kufurahisha, Utawala unaorudiwa wa U69593 ulipata hyperlocomotion iliyochochewa na koa kwa njia inayotegemea estradiol (Puig-Ramos et al., 2008). Hizi data zinaonyesha kwamba estradiol inachukua hatua za KOR katika panya za kike, athari ambayo inaweza kuhusishwa na tofauti za kijinsia katika kukabiliana na mafadhaiko (Puig-Ramos et al., 2008). Ikiwa katika panya za kike kuamsha mara kwa mara kwa KORs kuwezesha kutolewa kwa dopamini ya dri kama inavyoonekana katika kiume ni swali lisilojibiwa.
Ingawa uwezeshaji katika kutolewa kwa dopamine ya psychostimulant huzingatiwa kwa kike ukilinganisha na panya wa kiume, tofauti za kimapenzi katika mifumo ya dopamini iliyo chini ya usisitizo wa amphetamine locomotor haujainishwa kikamilifu (Becker, 1999). Mfiduo wa mara kwa mara wa amphetamine huchochea shughuli kubwa ya uvumbuzi katika wote wawili, ujana (Mathews na McCormick, 2007) na panya wa kike watu wazima (Media-Hallé et al., 2007), na panya za ujana wa kike zinazoonyesha unyeti wa nguvu zaidi baada ya kufichua mara kwa mara na amphetamine. Uamsho wa neonatal wa receptor ya D2 uliathiri hisia za tabia ya amphetamine tu katika panya za kike (Brown et al., 2011). Kama ilivyosemwa hapo awali, imeonekana katika panya za kiume udhihirisho wa mara kwa mara wa D2 agonist induces sensitization locomotor na tabia ya kulazimisha-kama (Dvorkin et al., 2006). Kwa kuongezea, uanzishaji wa KOR unaangazia usisitizo wa locomotor unaosababishwa na kufunuliwa mara kwa mara kwa quinpirole, kuwezesha udhibiti wa kizuizi cha receptors D2 juu ya kutolewa kwa DA katika NAc (Escobar et al., 2017). Tofauti za kijinsia kama vile unyeti wa chini wa athari ya athari ya KOR ya kutolewa kwa dopamini katika wanawake (Conway et al., 2019) inaweza kutoa hesabu ya mchango tofauti wa KOR juu ya utaftaji wa dawa ngumu.
Hitimisho
Je! KORs zinarekebishaje ishara ya dopamine ili kufafanua tabia za kuhamasishwa na ni wakati gani husababisha tabia ya kuhimiza? Takwimu za anatomical zinaonyesha kuwa KOR zina nafasi nzuri kudhibiti shughuli za synaptic ya neuropu ya dopamine ya katikati. Takwimu za kazi zinaonyesha kuwa KORs zinadhibiti DAT na D2R inafanya kazi na kiwango cha kurusha kwa neuropu ya dopamine. Ushahidi wa awali unaoonyesha kuwa uanzishaji wa papo hapo wa KORs unapungua kutolewa kwa dopamine kwa dawa za dhuluma kumetekelezwa na data inayoonyesha kuwa uhamasishaji unaorudiwa wa KOR unawezesha kutolewa kwa dopamine na utaftaji wa madawa ya kulevya. Dopamine kuashiria njia za moja kwa moja na za moja kwa moja za pato kutoka maeneo ya striatal (Kielelezo 1A). Kuchochea sugu na psychostimulants ambayo huongeza kutolewa kwa dopamine kwa kutumia D1R na D2R (Kielelezo 1B) au quinpirole inayoanzisha D2R tu (Kielelezo 1C) husababisha hisia za locomotor na tabia ya kulazimishwa na njia iliyo wazi ya D2R, na hivyo kubadili usawa kwenye njia ya moja kwa moja ya D1R. Uwasilishaji wa KOR huimarishwa wakati wa ulaji sugu wa kisaikolojia na kuongezeka kwa dynorphin katika neuroni za striatal D1 (Kielelezo 1B). Uwasilishaji wa KOR ulioimarishwa umewekwa katika mfano wa kifamasia wa OCD kwa kuisimamia U69593. Uanzishaji huu wa KOR unazidisha zaidi njia ya moja kwa moja ya D2 (Kielelezo 1C). Utafiti wa siku zijazo unapaswa kufanywa ili kufafanua kikamilifu athari za uanzishaji wa KOR juu ya shughuli ya DAT, kuelewa jukumu la mfumo wa KOR endoge katika uhasama wa quinpirole na kuamua mchango wa mfumo wa KOR kwa tendo la ngono linaloonekana katika tabia ya kulazimisha.
Msaada wa Mwandishi
AE, MA, na JF walichangia ubunifu wa muswada huo. AE na JF waliandika rasimu ya kwanza ya muswada huo na pembejeo kutoka kwa MA. MA na JC walichangia uhakiki muhimu na uhariri wa muswada. Waandishi wote waliidhinisha kwa kuchapishwa.
Fedha
Kazi ya waandishi waliotajwa katika hakiki hii imeungwa mkono na nambari za ruzuku za FONDECYT: 1110352 na 1150200 hadi MA; 1141088 kwa JF; DIPOG kutoa 391340281 kwa JF; FONDECYT Postdoctoral wenzake 3170497 kwa JC na 3190843 kwa AE.
Mgogoro wa Maslahi
Waandishi wanatangaza kuwa utafiti ulifanyika bila kutokuwepo na uhusiano wowote wa biashara au wa kifedha ambao unaweza kuitwa kama mgogoro wa maslahi.
Mhariri anayeshughulikia kwa sasa anaandaa Mada ya Utafiti na mmoja wa waandishi JF, na anathibitisha kutokuwepo kwa ushirikiano wowote mwingine.
Marejeo
Abraham, AD, Fontaine, HM, Wimbo, AJ, Andrews, MM, Baird, MA, Kieffer, BL, et al. (2017). Uanzishaji wa receptor ya Kappa opioid katika dopamine neurons inasumbua kizuizi cha tabia. Neuropsychopharmacology 43 (2), 362-372. kufanya: 10.1038 / npp.2017.133
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Wimbo, AJ, et al. (2018). Udhibiti wa estrogeni wa GRK2 inactivates kappa opioid receptor kuashiria analgesia ya upatanishi, lakini sio chuki. J. Neurosci. 38 (37), 8031-8043. Doi: 10.1523 / JNEUROSCI.0653-18.2018
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019). Mpinzani wa adenosine A2A receptor inaboresha dalili nyingi za psychin iliyoandaliwa mara kwa mara ya quinpirole. eNeuro 6 (1), 1-16. ENEURO.0366-18.2019. Doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, agonist ya mapokezi ya kappa-delta opioid iliyochanganywa ina mali ya kupambana na cocaine na athari mbaya za panya. Neuropharmacology 150, 217-228. do: 10.1016 / j.neuropharm.2019.02.010
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, MIMI, Fuentealba, JA (2019). Uzuiaji wa receptor ya kappa-opioid unabadilisha mabadiliko katika mienendo ya dorsolateral striatum dopamine wakati wa uhamasishaji wa amphetamine. J. Neurochem. 148, 348-358. Doi: 10.1111 / jnc.14612
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE, et al. (2012). Tofauti za kuashiria mali katika receptor ya kappa opioid ya 12-epi-salvinorin A na analogues zake. Bioorg. Med. Chem. Barua. 15; 22 (2), 1023-1026. Doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Athari tofauti za riwaya ya kappa opioid receptor antagonist, JDTic, juu ya kurudishwa tena kwa utaftaji wa-cocaine unaosababishwa na mafadhaiko wa muda mrefu wa mchezo wa cocaine na athari zake za kukinga kama vile kwenye panya. Psychopharmacol. (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Tofauti za kijinsia katika njia za neural kupatanishi thawabu na ulevi. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Becker, JB, Ramirez, VD (1981). Tofauti za kijinsia katika amphetamine ilichochea kutolewa kwa katekisimu kutoka kwa tishu za laini za vitunguu. Resin ya ubongo. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Becker, JB (1999). Tofauti za kijinsia katika kazi ya dopaminergic katika striatum na mkusanyiko wa kiini. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Berridge, KC, Robinson, TE (2016). Kuipenda, kutaka, na nadharia ya uhamasishaji ya uhamasishaji. Am. Kisaikolojia. 71, 670-679. doi: 10.1037 / amp0000059
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Berridge, KC, Venier, IL, Robinson, TE (1989). Onja uchambuzi wa kufanya kazi upya wa 6-hydroxydopamine-indenediagia: athari za hisia za kuamka na anhedonia ya kazi ya dopamine. Behav. Neurosci. 103, 36-45. Je: 10.1037 / 0735-7044.103.1.36
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Athari tofauti za utaratibu zilizosimamiwa kwa utaratibu wala- binaltorphimine (wala-BNI) juu ya agonist za kappa-opioid kwenye assay ya panya. Psychopharmacology 115, 311-319. do: 10.1007 / BF02245071
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Amphetamine uvumbuzi wa mahali na upendeleo mahali pa upendeleo katika panya wa kiume na wa kike kutibiwa kwa neinatiro. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Bruchas, MR, Chavkin, C. (2010). Kinase kasino na ishara zilizoelekezwa na ligand kwenye receptor ya kappa opioid. Psychopharmacol. (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Jukumu linalowezekana la vipokezi vya opioid katika motisha na shida kuu ya unyogovu. Pembeza. Resin ya ubongo. 239, 89–119. Doi: 10.1016 / bs.pbr.2018.07.009
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Carr, DB, Sesack, SR (2000). Gaba zenye neuroni katika mradi wa eneo la barabara ya tezi ya pembeni hadi gombo la utangulizi. Sinepsi 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Usikitishaji wa locomotor ya amphetamine unaambatana na kutolewa kwa juu zaidi ya K +-iliyosisitishwa ya Dopamine kwenye gamba la panya la pre-preal. Behav. Resin ya ubongo. 237, 313-317. do: 10.1016 / j.bbr.2012.09.052
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Chartoff, EH, Mavrikaki, M. (2015). Tofauti za ngono katika kazi ya receptor ya kappa opioid na athari yao katika athari. Mbele. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME, et al. (2016). Wakati wa uhusiano kati ya uanzishaji wa kappa opioid receptor na cocaine huamua athari kwenye malipo na kutolewa kwa dopamine. Neuropsychopharmacology 41, 989-1002. toa: 10.1038 / npp.2015.226
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Chavkin, C., Koob, GF (2016). Dynorphin, dysphoria, na utegemezi: mkazo wa ulevi. Neuropsychopharmacology 41, 373-374. toa: 10.1038 / npp.2015.258
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Mifumo ya upokeaji wa kappa-opioid ya asili hudhibiti mienendo ya machoaccumbal dopamine na mazingira magumu ya cocaine. J. Neurosci. 25, 5029-5037. toa: 10.1523 / JNEUROSCI.0854-05.2005
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Vipimo vya virutubisho vya hakuna-net-flux ya kiboreshaji huruhusu ugunduzi wa kuongezeka na hupungua kwa kuchukua dopamine katika kipenyo cha kipenyo cha panya. J. Neurosci. Njia 155, 187-193. toa: 10.1016 / j.jneumeth.2005.12.018
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Cole, RL, Konradi, C., Douglass, J., Hyman, SE (1995). Marekebisho ya Neuronal ya amphetamine na dopamine: mifumo ya Masi ya kanuni ya geni ya prodynorphin katika striatum ya panya. Neuron 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). Wanawake hawajali sana kuliko wanaume kwa athari za kusukuma-na kukandamiza dopamine ya uanzishaji wa kappa opioid receptor. Neuropharmacology 146, 231-241. do: 10.1016 / j.neuropharm.2018.12.002
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Cope, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, brown, RW (2010). Matibabu ya neinatal quinpirole huongeza uanzishaji wa injini na kutolewa kwa dopamine kwenye kiini hujilimbikiza msingi wa kukabiliana na matibabu ya amphetamine kwa kuwa watu wazima. Sinepsi 64, 289-300. do: 10.1002 / syn.20729
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Cox, BM, Mdogo, AB, Tazama, RE, Reichel, CM (2013). Tofauti za ngono katika kutafuta methamphetamine katika panya: athari ya oxytocin. Psychoneuroendocrinology 38, 2343-2353. toa: 10.1016 / j.psyneuen.2013.05.005
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Culver, KE, Szechtman, H., Levant, B. (2008). Ilibadilika kama dopamine D2-kama receptor inayofunga kwenye panya na unyeti wa tabia kwa quinpirole: athari za matibabu ya kabla na Ro 41-1049. Eur. J. Pharmacol. 592, 67-72. toa: 10.1016 / j.ejphar.2008.06.101
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Dalman, FC, O'Malley, KL (1999). uvumilivu wa kappa-Opioid na utegemezi katika tamaduni za neva za ubongo za ubongo za dopaminergic. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Kurudisha kwa tabia ya kokeini na tabia ya kutafuta-heroin inayoingiliana na dopamine D2 receptors inategemea wakati na inahusishwa na uhamasishaji wa tabia. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Devine, DP, Leone, P., Pocock, D., Hekima, RA (1993). Ushirikishwaji tofauti wa vipunguzi vya tezi ya sehemu ya chini, delta na kappa opioid katika mabadiliko ya kutolewa kwa dopamine ya basal mesolimbic: katika masomo ya vivo microdialysis. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Athari zinazopingana za mu na kappa opiate agonists juu ya kutolewa kwa dopamine kwenye mkusanyiko wa nukta na kwenye duru ya dorsal ya panya zinazoenda kwa uhuru. J. Pharmacol. Exp. Ther. 244, 1067-1080. Nenda: 10.1073 / pnas.85.14.5274
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Maendeleo na shirika la muda la kuangalia kwa bidii inayosababishwa na sindano zilizorudiwa za dopamine agonist quinpirole katika mfano wa mnyama wa machafuko-ya kulazimisha. Behav. Resin ya ubongo. 169, 303-311. do: 10.1016 / j.bbr.2006.01.024
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Athari za unyogovu kama kappa opioid receptor agonist salvinorin A zinahusishwa na kupungua kwa kutolewa kwa dopamine ya phasic katika kiunga cha mkufu. Psychopharmacol. (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edward, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., et al. (2017). Ukweli wa mzunguko katika usanifu wa inhibitory wa VTA inasimamia tabia iliyochochewa ya kokaini. Nat. Neurosci. 20 (3), 438-448. doi: 10.1038 / nn.4482
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). Uanzishaji wa receptor ya Kappa opioid inasababisha kuongezeka kwa-cococaine iliyosababisha kutolewa kwa kutolewa kwa dopamine iliyorekodiwa katika vivo kwenye mkusanyiko wa panya wa panya. Neuropsychopharmacology 39, 3036-3048. toa: 10.1038 / npp.2014.157
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, et al. (2015). Ubadilishaji wa Kappa Opioid Receptor-Imesababishwa na Inahitaji Uanzishaji wa p38 MAPK katika VTA Dopamine Neurons. J. Neurosci. 35, 12917-12931. toa: 10.1523 / JNEUROSCI.2444-15.2015
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Eilam, D., Szechtman, H. (1989). Athari ya Biphasic ya quinpirole ya agonist ya D-2 juu ya kutuliza na harakati. Eur. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Eilam, D., Szechtman, H. (2005). Tabia iliyosababishwa na kisaikolojia kama mfano wa wanyama wa machafuko-ya kulazimisha: njia ya kiadili kwa aina ya mila zinazolazimisha. Mtazamaji wa CNS. 10, 191-202. doi: 10.1017 / S109285290001004X
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Escobar, AP, Cornejo, FA, Andrés, MIMI, Fuentealba, JA (2012). Matibabu yaliyorudiwa na kappa opioid receptor agonist U69593 inarudisha kutolewa kwa dokamini iliyosababishwa ya K + iliyoingizwa kwenye densi za kiinitete, lakini sio usemi wa hisia za locomotor katika panya zilizo na hisia za amphetamine. Neurochem. Int. 60 (4), 344–349. Doi: 10.1016 / j.neuint.2012.01.014
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). Kupunguza dopamine na neurotransuction ya glutamate kwenye kiini cha mkusanyiko wa vidokezo vya panya-kuhisiwa kwa panya kwa kazi ya inhibitory D2 autoreceptor. J. Neurochem. 134, 1081-1090. Doi: 10.1111 / jnc.13209
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Escobar, AP, González, mbunge, Meza, RC, Noches, V., Henny, P., Gysling, K., et al. (2017). Mbinu za kupokelewa kwa kappa opioid receptor ya dopamine D2 receptor kazi katika uhamasishaji wa quinpirole-ikiwa ndani ya panya. Int. J. Neuropsychopharmacol. 20, 660-669. doi: 10.1093 / ijnp / pyx042
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Everitt, BJ, Robbins, TW (2005). Mifumo ya Neural ya kuimarisha madawa ya kulevya: kutoka kwa vitendo hadi tabia hadi kulazimishwa. Nat. Neurosci. 8, 1481-1489. toa: 10.1038 / nn1579
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Fattore, L., Melis, M. (2016). Tofauti za kijinsia katika tabia isiyo na msukumo na ya kulazimisha: kuzingatia ulevi wa madawa ya kulevya. Udhaifu. Biol. 21 (5), 1043-1051. doi: 10.1111 / adb.12381
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Plastiki ya asili na ya tabia inayohusiana na mpito kutoka kwa kudhibitiwa hadi kuongezeka kwa matumizi ya kokaini. Bioli. Psy. 58 (9), 751-9.
Figee, M., Pattij, T., Willuhn, mimi, Luigjes, J., van den Brink, W., Goudriaan, A., et al. (2016). Kulazimishwa katika machafuko-ya kulazimisha-ulaji na ulevi. Eur. Neuropsychopharmacol. 26, 856-868. doi: 10.1016 / j.euroneuro.2015.12.003
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ford, CP, Marko, GP, Williams, JT (2006). Mali na kizuizi cha opioid cha mesolimbic dopamine neurons hutofautiana kulingana na eneo la shabaha. J. Neurosci. 26, 2788-2797. toa: 10.1523 / JNEUROSCI.4331-05.2006
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Ford, CP, Beckstead, MJ, Williams, JT (2007). Kappa opioid inhibition ya somatodendritic dopamine inhibitory postynaptic currents. J. Neurophysiol. 97, 883-891. toa: 10.1152 / jn.00963.2006
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). Utawala unaorudiwa wa agonist ya kuchagua kappa-opioid receptor agonist U-69593 huongeza viwango vya dopamine vya seli za nje katika msongamano wa panya wa panya. J. Neurosci. Res. 84, 450-459. Dhana: 10.1002 / jnr.20890
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Kuongeza majibu ya locomotor kwa amphetamine iliyochochewa na utawala unaorudiwa wa kuchagua kappa-opioid receptor agonist U-69593. Sinepsi 61, 771-777. do: 10.1002 / syn.20424
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Athari za utawala wa papo hapo na mara kwa mara wa salvinorin A juu ya kazi ya dopamine kwenye storati ya dorsal. Psychopharmacol. (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Kulazimisha ulewaji wa pombe hulazimika kutokana na kutofaulu kwa udhibiti wa tabia ya dorsolateral ya tabia. J. Neurosci. 39 (9), 1744-1754. Doi: 10.1523 / JNEUROSCI.2615-18.2018
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Goto, Y., Neema, AA (2008). Usindikaji wa habari wa limbic na cortical katika kiini cha mkusanyiko. Mwelekeo wa Neurosci. 31, 552-558. Doi: 10.1016 / j.tins.2008.08.002
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Neema, AA, Bunney, BS (1980). Nigral dopamine neurons: kurekodi kwa ndani na kitambulisho na sindano ya L-dopa na histofluorescence. Bilim 210, 654-656. toa: 10.1126 / sayansi.7433992
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Grey, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). Agonist ya κ-opioid, U-69593, inapunguza tabia ya papo hapo ya amphetamine-evoke na viwango vya dialysate vya dialysate ya kalsiamu na glutamate katika striatum ya ventral. J. Neurochem. 73, 1066-1074. toa: 10.1046 / j.1471-4159.1999.0731066.x
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Kappa opioid receptors inasimamia utaftaji wa cocaine uliosisitiza na ubinifu wa synaptic. Neuron 77, 942-954. toa: 10.1016 / j.neuron.2012.12.034
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Heidbreder, CA, Shippenberg, TS (1994). U-69593 inazuia uhamasishaji wa cocaine kwa kurejesha dopamine ya basal. Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Wanawake wa opioid-, bangi- na wanawake wanaotegemea pombe huonyesha maendeleo ya haraka zaidi kwa matibabu ya dhuluma. Dawa ya Dawa Inategemea. 74 (3), 265-272. Doi: 10.1016 / j.drugalcdep.2004.02.001
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Kutolewa kwa dopamine iliyoimarishwa na vizuizi vya usafiri wa dopamine vilivyoelezewa na mfano wa kutengwa wa kizuizi na kasi ya volclic ya skanni. ACS Chem. Neurosci. 7, 700-709. doi: 10.1021 / acschemneuro.5b00277
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Holden, C. (2001). "Tabia" tabia: hizi zipo? Bilim 294, 980-982. toa: 10.1126 / sayansi.294.5544.980
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Imperato, A., Di Chiara, G. (1988). Athari za wagunduzi wa ndani wa D-1 na D-2 wanaotumika na wapinzani walisoma na uchambuzi wa ubongo. Eur. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Athari za kappa opioid receptor antagonist, norbinaltorphimine, juu ya mafadhaiko na kurudishwa tena kwa madawa ya kulevya ya upendeleo mahali pa hali ya nikotini katika panya. Psychopharmacol. (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., et al. (2014). Salvinorin A inasimamia kazi ya kupandikiza dopamine kupitia receptor ya kappa opioid na utaratibu wa kutegemea ERK1 / 2. Neuropharmacology 86, 228-240. do: 10.1016 / j.neuropharm.2014.07.016
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Usikivu wa mwenendo kwa quinpirole hauhusiani na kuongezeka kwa nukta za densi za dopamine zinazoongezeka. Neuropharmacology 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Koob, GF, Volkow, ND (2016). Neurobiolojia ya ulevi: uchambuzi wa neurocircuitry. Lancet Psychiatry 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Koob, GF (2013). Dawa ya kulevya ni upungufu wa thawabu na shida ya kutofautisha mafadhaiko. Mbele. Psychiatry 4, 72. doi: 10.3389 / fpsyt.2013.00072
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., et al. (2008). Tofauti za kijinsia katika vipimo vya dalili vya kulazimisha. Jibu wasiwasi. 25 (10), 832-838. doi: 10.1002 / da.20332
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, kappa opioid receptor agonist, hupata kuongezeka kwa doketi ya kokeini katika dopamine ya nje katika mkusanyiko wa panya wa panya. Neurosci. Barua. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Maoni ya Opioid-receptor mRNA katika panya CNS: athari za anatomiki na kazi. Mwelekeo wa Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Ujanibishaji wa Immunohistochemical ya receptor iliyokatwa ya kappa 1 kwenye CNS ya panya na hali. Neuroscience 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Margolis, EB, Karkhanis, AN (2019). Dopaminergic seli na michango ya mzunguko kwa kappa opioid receptor upatanishi upatanishi. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Margolis, EB, Hjelmstad, GO, Bonci, A., Mashamba, HL (2003). Kappa-opioid agonists huzuia moja kwa moja ugonjwa wa tumbo wa dopaminergic. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Margolis, EB, Hjelmstad, GO, Bonci, A., Mashamba, HL (2005). Wote wawili wa Kappa na Mu Opioid Agonists huzuia Uingizaji wa Glutamatergic kwa Neurons ya Ventral. J. Neurophysiol. 93, 3086-3093. toa: 10.1152 / jn.00855.2004
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Mashamba, HL (2006). Kappa opioids inadhibiti kwa hiari dopaminergic neurons zinazoangazia gamba la utangulizi. Proc. Natl. Chuo. Sci. Marekani 103, 2938-2942. Nenda: 10.1073 / pnas.0511159103
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Mashamba, HL (2008). Midbrain dopamine neurons: Lengo la makadirio huamua muda wa kuchukua hatua na dopamine D (2) kizuizi cha receptor. J. Neurosci. 28, 8908-8913. toa: 10.1523 / JNEUROSCI.1526-08.2008
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Mathews, IZ, McCormick, CM (2007). Panya wa kike na kiume katika ujana wa kuchelewa hutofautiana na watu wazima katika shughuli za upeanaji wa amphetamine, lakini sio kwa upendeleo wa mahali pa amphetamine. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, mimi, AR, T., et al. (2011). Tofauti za kijinsia katika machafuko-ya kulazimisha: ukaguzi wa fasihi. Braz. J. Saikolojia 33 (4), 390–399. Doi: 10.1590 / S1516-44462011000400014
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Kappa opioid receptor antagonism na prodynorphin usumbufu wa gene huzuia majibu ya tabia yanayosababisha mafadhaiko. J. Neurosci. 23 (13), 5674-5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
McLaughlin, JP, Ardhi, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Kabla ya uanzishaji wa mapokezi ya kappa opioid na mifano ya U50,488 mara kwa mara ililazimishwa kuogelea kwa shinikizo la kuogelea kwa upendeleo mahali pa upendeleo wa cocaine. Neuropsychopharmacology 31, 787-794. toa: 10.1038 / sj.npp.1300860
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). Majibu ya kitabia ya kukabiliana na mafadhaiko yanayosababishwa na jamii hubadilishwa na mfumo wa opaidi ya kappa ya asili. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Media-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Tofauti za kijinsia katika (+) - amphetamine- na (+) - majibu ya tabia ya methamphetamine-iliyochochea katika panya wa kiume na wa kike wa Sprague-Dawley. Pharmacol. Biochem. Behav. 86, 140-149. doa: 10.1016 / j.pbb.2006.12.018
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Athari za opioid za kappa juu ya kujiendesha kwa kokaini na nyani wa rhesus. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Msingi wa kimasiasa wa udhabiti wa muda mrefu wa udhihirisho. Nat. Mchungaji Neurosci. 2, 119-128. toa: 10.1038 / 35053570
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Kappa-opioid agonist U69593 potentiates sensitization sensor kwa D2 / D3 agonist quinpirole: pre- na postynaptic mifumo. Neuropsychopharmacology 31, 1967-1981. toa: 10.1038 / sj.npp.1300938
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Mchekeshaji, ML, Seeman, P., Szechtman, H. (2007). Kappa-opioid receptor ya kuchochea inafanya haraka pathogenesis ya kuangalia kwa lazima katika mfano wa uhamishaji wa quinpirole wa machafuko-ya kulazimisha (OCD). Behav. Neurosci. 121, 976-991. Je: 10.1037 / 0735-7044.121.5.976
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Pierce, RC, Kalivas, PW (1997). Mfano wa mzunguko wa usemi wa uhamasishaji wa tabia kwa psychostimulants kama amphetamine. Resin ya ubongo. Resin ya ubongo. Mchungaji. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Polter, AM, Askofu, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). Vizuizi vya Posta ya Kappa Opioid Receptors Huokoa Uwezo wa Muda mrefu wa Sinodi za Kuzuia na Inazuia Kurudishwa tena kwa Utaftaji wa Cocaine. Biol. Psychiatry 76, 785-793. Nenda: 10.1016 / j.biopsych.2014.04.019
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). U-69593, agonist ya kappa opioid receptor, hupunguza hisia za tabia za cocaine-ikiwa katika panya za kike. Behav. Neurosci. 122, 151-160. Je: 10.1037 / 0735-7044.122.1.151
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Endophenotypes ya Neococitiveitive ya impulivity na kulazimishwa: kuelekea mwelekeo wa akili. Njia ya Utambuzi. Sayansi 16, 81-91. toa: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Mzunguko wa estrous unaathiri kujiendesha kwa cocaine kwenye ratiba ya uwiano inayoendelea katika panya. Psychopharmacol. (Berl) 98, 408-411. do: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). Msingi wa neural wa kutamani madawa ya kulevya: nadharia ya uhamasishaji ya uhamasishaji. Resin ya ubongo. Resin ya ubongo. Mchungaji. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Robinson, TE, Berridge, KC (2001). Kuhamasisha-uhamasishaji na madawa ya kulevya. Kulevya 96, 103-114. toa: 10.1046 / j.1360-0443.2001.9611038.x
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Madhara ya kutofautisha ya nalmefene juu ya viwango vya kuchukua vya dopamine na shughuli za mapokezi ya kappa opioid katika kiini cha mkusanyiko kufuatia mfiduo sugu wa ethanol wa muda mrefu. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, et al. (2013). Upinzani wa muda mrefu wa receptors za vera opioid huzuia kuongezeka na kuongezeka kwa motisha kwa ulaji wa heroin. J. Neurosci. 33 (49), 19384-19392. Doi: 10.1523 / JNEUROSCI.1979-13.2013
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Sesack, SR, Pickel, VM (1992). Ujanibishaji wa eneo mbili ya upumuaji wa enkephalin na tyrosine hydroxylase immunoreac shughuli katika eneo la sehemu ya sehemu ya panya: sehemu ndogo za mwingiliano wa opiate-dopamine. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Sesack, SR, Aoki, C., Pickel, VM (1994). Ujanibishaji wa Ultrasta wa ujanibishaji wa d2 receptor-kama shughuli za neuropu ya dopamine na malengo yao ya striatal. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Sesia, T., Bizup, B., Neema, AA (2013). Tathmini ya mifano ya wanyama ya machafuko-ya kulazimisha: uunganisho na shughuli za neuroni ya dopamine. Int. J. Neuropsychopharmacol. 16, 1295-1307. doi: 10.1017 / S146114571200154X
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dynorphin na pathophysiology ya madawa ya kulevya. Pharmacol. Hapo. 116, 306-321. Doi: 10.1016 / j.pharmthera.2007.06.011
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Smith, JA, Loughlin, SE, Leslie, FM (1992). kizuizi cha kappa-Opioid cha [3H] kutolewa kwa dopamine kutoka kwa tamaduni za seli za seli zilizo na pembeni. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Dopamine D2L receptor panya ya kutoza panya zinaonyesha upungufu katika mali chanya na hasi za kuimarisha mali za morphine na katika kujifunza kujizuia. Neuroscience 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Uamsho unaosababisha mafadhaiko ya mfumo wa receptor wa dynorphin / κ-opioid katika upendeleo wa nafasi ya nikotini ya eneo la upendeleo. J. Neurosci. 32, 1488-1495. toa: 10.1523 / JNEUROSCI.2980-11.2012
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Spanagel, R., Herz, A., Shippenberg, TS (1992). Kupingana na mifumo endio asili ya opioid inayounda njia ya mesolimbic dopaminergic. Proc. Natl. Chuo. Sci. 89, 2046-2050. Nenda: 10.1073 / pnas.89.6.2046
Mtaalam, SG, Manaye, KF, Sadeq, M., Ujerumani, DC (1993). Vipokezi vya Opioid katika mikoa ya dopaminergic ya punda ya panya. II. Kappa na delta receptor autoradiography. J. Neural Transm. Sehemu ya Mwanzo. 91, 53-66. do: 10.1007 / BF01244918
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Upatanishi wa asili wa kappa-opioid wa utaftaji-uliochochea wa upendeleo wa mahali pa ethanol na hali ya kujisimamia. Psychopharmacol. (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Kutafuta Dawa ya Kulenga: uhamasishaji wa tabia na kurudi tena kwa tabia ya kutafuta-dawa ya kuongoa DR, ed. Pharmacol. Mchungaji. 63, 348–365. Doi: 10.1124 / pr.109.001933
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., et al. (2016). Uhalisia wa mfano wa usawa wa usawa wa quinpirole ya OCD: unganisho la ushahidi kutoka kwa masomo ya wanyama na ya kliniki. Mbele. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Jua, T., Wimbo, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., et al. (2019). Uingizaji wa msingi wa amygdala kwa njia ya upimaji wa njia ya mbele ya korti inadhibiti tabia ya kuangalia-ya kulazimisha-kama ya kuangalia. Proc. Natl. Chuo. Sci. Marekani 116, 3799-3804. Nenda: 10.1073 / pnas.1814292116
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Ushirikiano mkubwa wa receptors kubwa za opioid na "dopamine transporter" katika nukta hukusanya maelezo mafupi ya axonal. Sinepsi 42, 185-192. do: 10.1002 / syn.10005
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Sehemu ya mazingira na tabia ya uhamasishaji inayosababishwa na dopamine agonist quinpirole. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Nguvu za uhamasishaji wa tabia zinazosababishwa na dopamine agonist quinpirole na utaratibu wa kati wa kudhibiti nishati. Psychopharmacol. (Berl) 115, 95-104. do: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Quinpirole inachochea tabia ya kuangalia kulazimishwa katika panya: kielelezo kinachowezekana cha mnyama wa shida inayozingatia (OCD). Behav. Neurosci. 112, 1475-1485. Je: 10.1037 / 0735-7044.112.6.1475
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Szechtman, H., Culver, K., Eilam, D. (1999). Jukumu la mifumo ya dopamine katika machafuko-ya kulazimisha (OCD): maana kutoka kwa mfano wa riwaya ya riwaya ya wanyama. Pol. J. Pharmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, C a, JZ, M., et al. (2001). Tabia ya kuangalia ya kulazimishwa kwa panya zilizo na unyeti kama mfano wa wanyama wa shida ya Obsessive-Compulsive Disorder (OCD): fomu na udhibiti. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D., Harvey, BH, Edemann-Callesen, H., et al. (2017). Machafuko yanayozingatia-macho: Maoni kutoka kwa mifano ya wanyama. Neurosci. Biobehav. Mchungaji. 76, 254-279. toa: 10.1016 / j.neubiorev.2016.04.019
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Tejeda, HA, Bonci, A. (2019). Udhibiti wa receptor ya dynorphin / kappa-opioid ya mienendo ya dopamine: Matokeo kwa nchi hasi na shida ya akili. Resin ya ubongo. 1713, 91-101. toa: 10.1016 / j.brainres.2018.09.023
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., et al. (2017). Njia na kiini maalum cha kappa-opioid receptor moduli ya usawa wa uchochezi-uzuiaji tofauti milango d1 na d2 kusanyiko la shughuli za neuron. Neuron 93 (1), 147-163. Doi: 10.1016 / j.neuron.2016.12.005
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Thompson, AC, Zapata, A., haki, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Uanzishaji wa receptor ya Kappa-opioid hurekebisha utaftaji wa dopamine kwenye mkusanyiko wa kiini na unapinga athari za cocaine. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Thompson, D., Martini, L., Whistler, JL (2010). Kiwango kilichobadilishwa cha receptors za D1 na D2 dopamine kwenye striatum ya panya inahusishwa na uhamasishaji wa tabia kwa cocaine. PloS One 5, e11038. Doi: 10.1371 / journal.pone.0011038
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., et al. (2000). Kazi za kutofautisha za isoforms mbili za dopamine D2 receptors. Nature 408, 199-203. toa: 10.1038 / 35041572
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Van Bockstaele, EJ, Pickel, VM (1995). Neurusi zenye Gaba kwenye mradi wa eneo la kuvuta pumzi hadi kiini cha seli. Resin ya ubongo. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Van't Veer, A., Bechtholt, AJ, Onvani, S., Mfinyanzi, D., Wang, Y., Liu-Chen, LY, et al. (2013). Utoaji wa vipokezi vya kappa-opioid kutoka kwa ubongo wa dopamine neurons ina athari kama za wasiwasi na huongeza plastiki inayosababishwa na cocaine. Neuropsychopharmacology 38 (8), 1585-1597. kufanya: 10.1038 / npp.2013.58
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Vanderschuren, LJ, Kalivas, PW (2000). Mabadiliko katika maambukizi ya dopaminergic na glutamatergic katika induction na usemi wa hisia za tabia: uhakiki muhimu wa masomo ya preclinical. Psychopharmacol. (Berl) 151 (2-3), 99-120. Doi: 10.1007 / s002130000493
Volkow, ND, Hekima, RA, Mtoaji, R. (2017). Mfumo wa nia ya dopamine: athari kwa madawa ya kulevya na chakula. Nat. Mchungaji Neurosci. 18, 741-752. Doi: 10.1038 / nrn.2017.130
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Kutolewa kwa dopamine na kuchukua ni kubwa zaidi kwa wanawake kuliko striatum za punda za kiume kama inavyopimwa na kasi ya mzunguko wa volclic. Neuroscience 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Wee, S., Koob, GF (2010). Jukumu la mfumo wa opioid dynorphin-κ katika athari za utiaji nguvu za madawa ya kulevya. Psychopharmacol. (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Uzuiaji wa receptors za kappa opioid uliongezeka kwa ulaji zaidi wa kahawa katika panya na ufikiaji wa kupikia wa cocaine. Psychopharmacol. (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., et al. (2015). Vinjari vya Opioid receptors katika kiini hujumisha kuongezeka kwa upatanishi wa ulaji wa methamphetamine. J. Neurosci. 35 (10), 4296-4305. Doi: 10.1523 / JNEUROSCI.1978-13.2015
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Vipimo vya dalili katika machafuko-ya kulazimisha: phenomenology na matokeo ya matibabu na yatokanayo na kuzuia ibada. Psychopathology 46, 365-376. toa: 10.1159 / 000348582
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Wilson, CJ, mchanga, SJ, Groves, PM (1977). Sifa ya kitakwimu ya treni za spike za neuronal katika nigra ya aina: aina za seli na mwingiliano wao. Resin ya ubongo. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Hekima, RA (2009). Majukumu ya nigrostriatal-sio tu mesocorticolimbic-dopamine katika malipo na madawa ya kulevya. Mwelekeo wa Neurosci. 32, 517-524. Doi: 10.1016 / j.tins.2009.06.004
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Xiao, L., Becker, JB (1994). Uainishaji wa virutubishi wa upimaji wa nje ya mkusanyiko wa dopamine ya nje katika panya za kiume na za kike: athari za mzunguko wa estrous na gonadectomy. Neurosci. Barua. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Yager, LM, Garcia, AF, Wunsch, AM, Ferguson, SM (2015). Ins na nje ya striatum: jukumu la madawa ya kulevya. Neuroscience 301, 529-541. toa: 10.1016 / j.neuroscience.2015.06.033
Kitambulisho cha PubMed | Nakala Kamili ya CrossRef | Google
Keywords: kappa opioid receptor, dopamine, uvumilivu, amphetamine, quinpirole, uhamasishaji wa locomotor
Utunzaji: Escobar AdP, Casanova JP, Andrés ME na Fuentealba JA (2020) Crosstalk Kati ya Kappa Opioid na Mifumo ya Dopamine katika Behaviors zinazosababisha. Mbele. Pharmacol. 11: 57. Dhana: 10.3389 / fphar.2020.00057
Iliyopokelewa: 16 Oktoba 2019; Iliyopokelewa: 22 Januari 2020;
Ilichapishwa: 18 Februari 2020.
Mwisho na:
Gonzalo E. Yevenes, Chuo Kikuu cha Concepcion, Chile
Upya na:
Luis Gerardo Aguayo, Chuo Kikuu cha Concepcion, Chile
Hugo Tejeda, Taasisi ya Kitaifa ya Dawa za Kulehemu (NIDA), Marekani
Cecilia Scorza, Instituto de Inacaciones Biológicas Clemente Estable (IIBCE), Urugwai
Hati miliki © 2020 Escobar, Casanova, Andrés na Fuentealba. Hii ni nakala ya ufikiaji wazi iliyosambazwa chini ya masharti ya License Attribution License (CC BY). Matumizi, usambazaji au uzazi katika vyuo vikuu vingine inaruhusiwa, ikiwapa waandishi wa awali na mmiliki wa hakimiliki ni sifa na kwamba kuchapishwa kwa awali katika gazeti hili kunasemekana, kwa mujibu wa mazoezi ya elimu ya kukubalika. Hakuna matumizi, usambazaji au uzazi inaruhusiwa ambayo haitii sheria hizi.
* Mawasiliano: José Antonio Fuentealba, [barua pepe inalindwa]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675