Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
абстрактний
Підлітковий вік - це період розвитку, який характеризується субоптимальними рішеннями та діями, що призводять до збільшення випадків ненавмисних травм і насильства, зловживання алкоголем та наркотиками, ненавмисної вагітності та захворювань, що передаються статевим шляхом. Традиційні нейробіологічні та когнітивні пояснення поведінки підлітків не зуміли врахувати нелінійні зміни в поведінці, які спостерігалися в підлітковому віці, щодо дитинства та дорослого віку. Цей огляд забезпечує біологічно правдоподібну концептуалізацію нейронних механізмів, що лежать в основі цих нелінійних змін у поведінці, як підвищеної чутливості до стимулів, тоді як контроль імпульсу все ще залишається відносно незрілим протягом цього періоду. Нещодавні дослідження людини і дослідження на тваринах дають біологічну основу для цього погляду, пропонуючи диференційований розвиток лімбічних систем винагороди відносно систем управління зверху вниз під час підліткового віку щодо дитинства і дорослого віку. Ця модель розвитку може загостритися у тих підлітків, які мають схильність до ризику, збільшуючи ризик поганих результатів.
За даними Національного центру статистики охорони здоров'я, у Сполучених Штатах щорічно відбувається загибель підлітків 13,000. Приблизно 70% цих смертей виникли внаслідок аварій на транспортних засобах, ненавмисних травм, вбивств і самогубств (Eaton et al., 2006). Результати Національного обстеження поведінки молодіжного ризику 2005 (YRBS) показують, що підлітки залучаються до поведінки, яка підвищує ймовірність смерті або хвороби, керуючи автомобілем після пиття або без ременя безпеки, нести зброю, використовуючи незаконні речовини, і займатися незахищеним сексом що призводить до ненавмисної вагітності та ЗПСШ, включаючи ВІЛ-інфекцію (Eaton et al., 2006). Ці статистичні дані підкреслюють важливість розуміння ризикового вибору та дій у підлітків.
Ряд когнітивних і нейробіологічних гіпотез постулювали, чому підлітки вступають в субоптимальну поведінку вибору. У недавньому огляді літератури про розвиток людського мозку підлітків, Юргелун-Тодд (2007) припускає, що когнітивний розвиток протягом підліткового віку пов'язаний з прогресивною ефективністю когнітивних здібностей. Ця ефективність описується як залежне від дозрівання префронтальної кори, про що свідчить підвищена активність у фокальних префронтальних областях (Rubia et al., 2000; Тамм, Менон та Рейсс, 2002) і зменшена активність у нерелевантних областях мозку (Brown et al., 2005; Durston et al., 2006).
Ця загальна закономірність поліпшення когнітивного контролю з дозріванням префронтальної кори свідчить про лінійне зростання розвитку від дитинства до дорослого віку. Проте субоптимальний вибір та дії, що спостерігаються під час підліткового віку, представляють собою нелінійну зміну поведінки, яку можна відрізнити від дитинства та дорослого віку, про що свідчить Національний центр статистики здоров'я про поведінку та смертність підлітків. Якщо когнітивний контроль і незріла префронтальна кора були основою для субоптимального вибору поведінки, то діти повинні виглядати дивно схожі або навіть гірше, ніж підлітки, враховуючи їх менш розвинені префронтальні кори і когнітивні здібності. Таким чином, незріла префронтальна функція сама по собі не може враховувати поведінку підлітків.
Точна концептуалізація когнітивних і нейробіологічних змін підлітка повинна розглядати підлітковий період як перехідний період розвитку (Спис, 2000), а не один знімок часу (Кейсі, Тоттенхем, Лістон та Дурстон, 2005). Іншими словами, щоб зрозуміти цей період розвитку, переходи в підлітковий і вихідний вік є необхідними для розрізнення окремих ознак цього етапу розвитку. Встановлення траєкторій розвитку для когнітивних і нервових процесів має важливе значення для характеристики цих переходів і обмежуючих інтерпретацій про зміни в поведінці протягом цього періоду. На когнітивному або поведінковому рівні підлітки характеризуються як імпульсивні (тобто відсутність когнітивного контролю) і прийняття ризику за допомогою цих конструкцій, що використовуються як синоніми і без вдячності за різними траєкторіями розвитку кожного з них. На нейробіологічному рівні, зображення людини і дослідження на тваринах свідчать про різні нейробіологічні основи і траєкторії розвитку нейронних систем, які лежать в основі цих окремих конструкцій імпульсного контролю і ризикованих рішень.
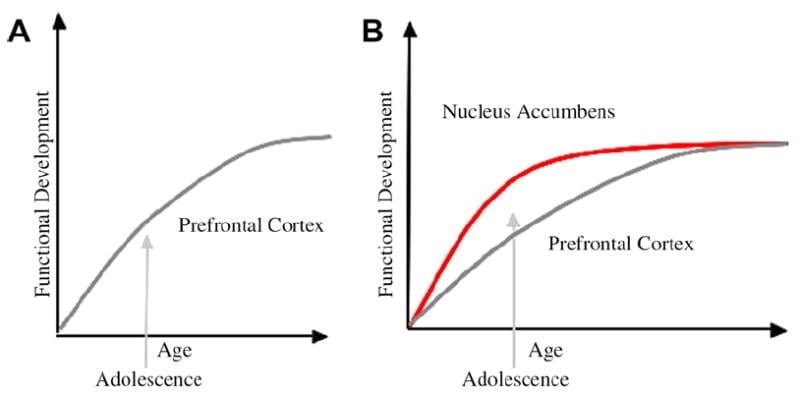
Ми розробили нейробіологічну модель розвитку підлітків в рамках цієї системи, яка базується на моделях гризунів (Лавіола, Адріані, Теранова та Герра, 1999; Спис, 2000) і останні дослідження візуалізації підліткового віку (Ernst et al., 2005; Гальван, Заєць, Восс, Гловер та Кейсі, 2007; Galvan et al., 2006). Рис. 1 нижче зображена ця модель. Ліворуч - традиційна характеристика підліткового віку, пов'язана майже виключно з незрілістю префронтальної кори. Праворуч - запропонована нами нейробіологічна модель, яка ілюструє, як лімбічні субкортикальні і префронтальні зверху вниз контрольні регіони повинні розглядатися разом. Мультфільм ілюструє різні траєкторії розвитку цих систем, причому лімбічні системи розвиваються раніше, ніж префронтальні контрольні області. Згідно з цією моделлю, індивідуум більш упереджено функціонально зрілими лімбічними регіонами під час підліткового віку (тобто дисбалансом лімбічного відносно префронтального контролю), порівняно з дітьми, для яких обидві ці системи (тобто лімбічні та префронтальні) ще розвиваються; і порівняно з дорослими, для яких ці системи повністю зрілі. Ця перспектива є основою для нелінійних зрушень у поведінці в усьому розвитку, завдяки більш ранньому дозріванню цієї лімбічної відносно менш зрілої префронтальної контрольної області згори. З розвитком і досвідом, функціональна зв'язок між цими регіонами забезпечує механізм для управління зверху вниз цих регіонів (Заєць, Восс, Гловер та Кейсі, 2007а). Далі, модель узгоджує протиріччя статистики охорони здоров'я з ризикованою поведінкою підліткового віку з проникливим спостереженням Рейна та Фарлі (2006) що підлітки здатні розуміти і розуміти ризики поведінки, в яких вони беруть участь. Згідно з нашою моделлю, в емоційно важливих ситуаціях лімбічна система перемагає системи управління, враховуючи її зрілість відносно системи префронтального управління. Докази з поведінкових і людських досліджень візуалізації для підтримки цієї моделі надаються в контексті дій у корисному та емоційному контексті (Galvan et al., 2006, 2007; Заєць, Восс, Гловер та Кейсі, 2007б; Hare et al., 2007a). Крім того, ми спекулюємо, чому мозок може розвиватися таким чином і чому деякі підлітки можуть опинитися в більшому ризику для прийняття субоптимальних рішень, що призводять до бідніших довгострокових результатів (Galvan et al., 2007; Hare et al., 2007b).
Запропоновано традиційне пояснення поведінки підлітків через тривалий розвиток префронтальної кори (А). Наша модель враховує розвиток префронтальної кори разом з субкортикальними лімбічними областями (наприклад, nucleus accumbens), які були залучені до ризикованого вибору та дій (B).
Розвиток цілеспрямованої поведінки
Наріжним каменем когнітивного розвитку є здатність придушувати невідповідні думки та дії на користь цілеспрямованих, особливо за наявності переконливих стимулів (Кейсі, Гальван та Заєць, 2005; Casey et al., 2000b; Кейсі, Томас, Девід-син, Кунц та Францен, 2002а; Кейсі, Тоттенхем і Фосселла, 2002б). Ряд класичних досліджень розвитку показали, що ця здатність розвивається протягом дитинства та юності (Корпус, 1972; Flavell, Feach, & Chinsky, 1966; Кітінг і Боббіт, 1978; Паскуаль-Леоне, 1970). Декілька теоретиків стверджували, що когнітивний розвиток відбувається за рахунок збільшення швидкості та ефективності обробки, а не через збільшення розумової спроможності (наприклад, Bjorkland, 1985; Bjorkland, 1987; Корпус, 1985). Інші теоретики включили конструкт «інгібуючих» процесів у свій розгляд когнітивного розвитку (Harnishfeger & Bjorkland, 1993). Відповідно до цього, незрілі пізнання характеризуються схильністю до втручання з боку конкуруючих джерел, які повинні бути придушені (наприклад, Brainerd & Reyna, 1993; Кейсі, Томас, Девідсон, Кунц та Францен, 2002а; Dempster, 1993; Алмаз, 1985; Munakata & Yerys, 2001). Таким чином, цілеспрямована поведінка вимагає контролю імпульсів або затримки задоволення для оптимізації результатів, і ця здатність, здається, зріла у дитинстві та юності.
Поведінка підлітків описується як імпульсивна і ризикована, майже синонімно, але ці конструкції покладаються на різні когнітивні і нервові процеси, які передбачають різні конструкції з різними траєкторіями розвитку. Зокрема, огляд літератури свідчить про те, що імпульсивність зменшується з віком у дитинстві та підлітковому віці (Casey et al., 2002a; Кейсі, Гальван та ін., 2005; Galvan et al., 2007) і пов'язаний з тривалим розвитком префронтальної кори (Кейсі, Гальван та ін., 2005), хоча існують відмінності в тій мірі, в якій дана особа імпульсивна чи ні, незалежно від віку.
На відміну від імпульсного / когнітивного контролю, прийняття ризику зростає протягом підліткового віку по відношенню до дитинства і дорослого віку і пов'язане з підкірковими системами, які, як відомо, беруть участь у оцінці винагород. Вивчення людських зображень, які будуть переглянуті, свідчать про збільшення підкоркової активації (наприклад, аккумбенс) при прийнятті ризикових виборів (Кухнен і Натсон, 2005; Matthews & et al., 2004; Montague & Berns, 2002), що перебільшено у підлітків, у порівнянні з дітьми та дорослими (Ernst et al., 2005; Galvan et al., 2006). Ці результати свідчать про різні траєкторії поведінки, що базується на винагороді або стимулюванні, з більш раннім розвитком цих систем щодо систем управління, які демонструють затяжний і лінійний розвиваючий курс, з точки зору вибору неприйнятних варіантів і дій на користь цілеспрямованих.
Докази з нейровізуальних досліджень розвитку людського мозку
Недавні дослідження розвитку підліткового мозку базувалися на досягненнях у методах нейровізуалізації, які можна легко використовувати для розвитку людських популяцій. Ці методи покладаються на методи магнітно-резонансної томографії (МРТ) (див Рис. 2) і включають: структурні МРТ, які використовуються для вимірювання розмірів і форми структур; функціональна МРТ, яка використовується для вимірювання структури мозкової діяльності; і томографа дифузії (DTI), який використовується для індексування зв'язності трактів волокон білої речовини. Докази нашої моделі розвитку конкуренції між кортикальними та підкірковими областями підтримуються незрілою структурною та функціональною зв'язком, виміряним відповідно DTI та fMRI.
МРТ дослідження розвитку людського мозку
Кілька досліджень використовували структурний МРТ для відображення анатомічного ходу нормального розвитку мозку (див. Огляд) Durston et al., 2001). Хоча загальний розмір мозку становить приблизно 90% від його дорослого розміру до шести років, сірі і білі субкомпоненти мозку продовжують динамічно змінюватися протягом підліткового віку. Дані недавніх досліджень MRI показують, що об'єм сірої речовини має інвертований U-образний малюнок, з більшою регіональною варіацією, ніж біла речовина (Giedd, 2004; Gogtay et al., 2004; Sowell et al., 2003; Соуелл, Томпсон і Тога, 2004). Загалом, регіони, які обслуговують первинні функції, такі як моторні та сенсорні системи, зрілі раніше; області асоціації вищого порядку, які інтегрують ці основні функції, зрілі пізніше (Gogtay et al., 2004; Соуелл, Томпсон і Тога, 2004). Наприклад, дослідження, що використовують заходи на основі МРТ, показують, що втрата кіркової сірої речовини відбувається раніше в первинних сенсомоторних областях, а останні - у дорсолатеральній префронтальній і латеральній скроневій корі (Gogtay et al., 2004). Ця закономірність узгоджується з нелюдськими приматами і людськими посмертними дослідженнями, які показують, що префронтальна кора є однією з останніх ділянок головного мозку, зрілу (Буржуа, Голдман-Ракіч і Ракіч, 1994; Huttenlocher, 1979). На відміну від сірої речовини, об'єм білої речовини збільшується приблизно лінійно, збільшуючись протягом усього розвитку і до дорослого віку (Gogtay et al., 2004). Ці зміни, ймовірно, відображають поточну мієлінізацію аксонів олігодендроцитами, що підвищують провідність і зв'язок нейронів.
Хоча при дослідженні структурних змін субкортикальним областям приділяється менше уваги, деякі з найбільших змін у розвитку мозку спостерігаються в цих регіонах, зокрема в базальних гангліях (Sowell et al., 1999См. Рис. 3) і особливо у чоловіків (Giedd et al., 1996). Зміни в структурному обсязі в базальних гангліях і префронтальних регіонах цікаві в світлі відомих процесів розвитку (наприклад, дендритне розведення, загибель клітин, синаптична обрізка, мієлінізація), що відбуваються в дитинстві та підлітковому віці. Ці процеси дозволяють тонко налаштувати і посилити зв'язки між префронтальними і підкірковими областями з розвитком і навчанням, які можуть збігатися з більшим когнітивним контролем. Як ці структурні зміни стосуються когнітивних змін? Ряд досліджень пов'язаний з структурним дозріванням лобової частки і когнітивною функцією за допомогою нейропсихологічних і когнітивних заходів (наприклад, Sowell et al., 2003). Зокрема, повідомлялося про асоціації між регіонарними обсягами МРТ на основі префронтальних коркових і базальних гангліїв на основі МРТ та заходи когнітивного контролю (тобто здатність перевизначати невідповідну реакцію на користь іншого або пригнічувати увагу до атрибуту нерелевантних стимулів на користь відповідного атрибуту стимулу (Casey, Trainor et al., 1997). Ці дані свідчать про те, що когнітивні зміни відображені в структурних змінах мозку і підкреслюють важливість субкортикальних (базальних гангліїв), а також розвитку кортикального (наприклад, префронтального кори).

DTI дослідження розвитку людського мозку
Розглянуті дослідження МРТ на основі морфометрії свідчать про те, що коркові сполуки підбудовуються з усуненням надлишковості синапсів і посилення відповідних зв'язків з розвитком і досвідом. Останні досягнення в області технології МРТ, такі як DTI, є потенційним інструментом для вивчення ролі конкретних трактів білої речовини для розвитку мозку і поведінки з більш детальною інформацією. Відповідними для цієї статті є дослідження нейровізуалізації, які пов'язують розвиток волокнистих шляхів із поліпшенням когнітивних здібностей. Зокрема, показані зв'язки між показниками префронтального розвитку білої речовини на основі DTI та когнітивним контролем у дітей. В одному дослідженні розвиток цієї здатності позитивно корелював з префронтально-тім'яними волокнами (Nagy, Westerberg, & Klingberg, 2004) узгоджується з функціональними нейровизуализационними дослідженнями, що демонструють диференційований набір цих регіонів у дітей щодо дорослих.
Використовуючи аналогічний підхід, Liston et al. (2005) показали, що тракти білої речовини між префронтально-базальними гангліями і -засобами нижніх волокон продовжують розвиватися в дитячому віці до дорослого віку, але тільки ті тракти між префронтальною корою головного мозку і базальними гангліями корелюють з імпульсним контролем, що вимірюється продуктивністю на ходу / nogo завдання. Префронтальні волокнисті шляхи були визначені регіонами інтересів, визначеними в дослідженні fMRI з використанням того ж самого завдання. В обох дослідженнях ДТІ розвитку показники волокнистого тракту корелювали з розвитком, але специфічність окремих волокнистих шляхів з когнітивною діяльністю виявлялася шляхом дисоціації окремого тракту (Liston et al., 2005) або пізнавальні здібності (Nagy et al., 2004). Ці знахідки підкреслюють важливість вивчення не тільки регіональних, але і пов'язаних із схемою змін при формулюванні тверджень про вікові зміни в нейронних субстратах когнітивного розвитку.
Функціональні МРТ дослідження поведінкового та розвитку мозку
Хоча структурні зміни, виміряні за допомогою МРТ та ДТІ, були пов'язані з поведінковими змінами під час розвитку, більш прямим підходом для вивчення структурно-функціональної асоціації є вимірювання змін головного мозку та поведінки одночасно, як і з fMRI. Здатність вимірювати функціональні зміни в розвитку мозку за допомогою МРТ має значний потенціал для галузі розвитку науки. У контексті поточної статті fMRI надає засоби для обмеження інтерпретації поведінки підлітків. Як зазначалося раніше, вважається, що розвиток префронтальної кори відіграє важливу роль у дозріванні вищих когнітивних здібностей, таких як прийняття рішень і когнітивний контроль (Кейсі, Тоттенхем і Фосселла 2002b; Casey, Trainor et al., 1997). Багато парадигми були використані, разом з fMRI, для оцінки нейробіологічної основи цих здібностей, включаючи флангові, Stroop і go / nogo завдання (Casey, Castellanos et al., 1997; Кейсі, Гідд та Томас, 2000а; Durston et al., 2003). У сукупності ці дослідження показують, що діти набирають різні, але часто більші, більш дифузні префронтальні області при виконанні цих завдань, ніж дорослі. Структура діяльності в областях мозку центральна для виконання завдань (тобто, що корелює з когнітивною продуктивністю) стає більш фокусною або тонкою з віком, тоді як регіони, які не співвідносяться з виконанням завдань, зменшуються в діяльності з віком. Ця закономірність спостерігається в обох перерізах (Brown et al., 2005) і поздовжні дослідження (Durston et al., 2006) і в різних парадигмах. Хоча нейровизуализирующие дослідження не можуть остаточно охарактеризувати механізм таких змін у розвитку (наприклад, дендритне обростання, синаптичне обрізання), результати відображають розвиток і вдосконалення проекцій в і від активованих областей мозку з дозріванням і дозволяють припустити, що ці зміни відбуваються протягом тривалого часу проміжок часу (Brown et al., 2005; Бунге, Дудукович, Томасон, Вайдя та Габріелі, 2002; Casey, Trainor et al., 1997; Casey et al., 2002a; Крона, Донохью, Гономіхль, Венделькен та Бунге, 2006; Luna et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro, & Eden, 2003).
Як ця методологія може повідомити нас про те, чи дійсно у підлітків бракує достатнього когнітивного контролю (імпульсивний) або є ризикованими у своєму виборі та дії? Контроль імпульсів, виміряний за допомогою когнітивних завдань управління, таких як завдання go / nogo, демонструє лінійну структуру розвитку в дитинстві та юності, як описано вище. Проте, нещодавні дослідження нейровізуалізації почали вивчати пов'язану з винагородою обробку, характерну для ризику у підлітків (Bjork et al., 2004; Ernst et al., 2005; May et al., 2004). Ці дослідження були зосереджені в основному на області аккумбенса, частина базальних гангліїв, що беруть участь у прогнозуванні винагороди, а не характеризують розвиток цього регіону в поєднанні з регуляторними областями зверху вниз (префронтальна кора). Незважаючи на те, що нещодавній звіт про меншу вентральну префронтальну активність у підлітків щодо дорослих під час грошового рішення щодо прийняття рішень щодо ризикованої поведінки було показано (Ешель, Нельсон, Блер, Пайн та Ернст, 2007).
В цілому, деякі дослідження досліджували, як розвиток схеми винагороди в субкортикальних областях (наприклад, accumbens) змінюється разом з розвитком коркових префронтальних областей. Більш того, як ці нейронні зміни збігаються з пошуком винагороди, імпульсивністю та поведінкою ризику залишаються відносно невідомими. Наша нейробіологічна модель пропонує, що поєднання підвищеної чутливості до нагороди та незрілості в областях поведінкового контролю може ухилити підлітків шукати негайних, а не довгострокових прибутків, можливо, пояснюючи їх збільшення ризикованим прийняттям рішень та імпульсивною поведінкою. Відстеження субкортикального (наприклад, accumbens) і коркового (наприклад, префронтального) розвитку прийняття рішень в дитинстві до зрілого віку, надає додаткові обмеження щодо того, чи зміни, зазначені в підлітковому віці, є специфічними для цього періоду розвитку, або відображають дозрівання, яке постійно відбувається в Дещо лінійний малюнок від дитинства до дорослого віку.
Емпіричні дані нещодавнього дослідження fMRI допомагають підтримувати нашу нейробіологічну модель і використовують перехідний підхід до розуміння підліткового віку шляхом вивчення змін до і після підліткового віку. У цьому дослідженні (Galvan et al., 2006), ми розглянули поведінкові та нервові відповіді на винагороду за маніпуляції в процесі розвитку, зосередившись на областях мозку, пов'язаних з навчанням та поведінкою, пов'язаною з винагородоюХікосака та Ватанабе, 2000; Pecina, Cagniard, Berridge, Aldridge, & Zhuang, 2003; Шульц, 2006) і дослідження дорослих зображень (наприклад, Натсон, Адамс, Фонг, і Гоммер, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Ендрюс, 2001; Zald et al., 2004) і в дослідженнях залежності (Hyman & Malenka, 2001; Volkow & Li, 2004). На основі моделей гризунів (Laviola et al., 1999; Спис, 2000) та попередня робота з зображеннями (Ernst et al., 2005), ми припустили, що у відношенні дітей і дорослих підлітки показуватимуть перебільшену активізацію аккумбенс, узгоджено з менш зрілим набором зверху вниз префронтальних контрольних областей. Нещодавня робота, що демонструє затримку функціонального зв'язку між цими префронтальними та лімбічними субкортикальними областями у підлітковому віці щодо дорослих, забезпечує механізм відсутності контролю над цими регіонами зверху вниз (Hare et al., 2007a).
Наші результати узгоджуються з моделями гризунів (Лавіола, Макрі, Морлі-Флетчер та Адріані, 2003) та попередні дослідження зображень (Ernst et al., 2005) припускають, що активність аккумсмена підвищується до нагороди в підлітковому віці. Дійсно, у порівнянні з дітьми і дорослими, підлітки показали перебільшений відгук в очікуванні винагороди. Однак і діти, і підлітки показали менш зрілу реакцію у префронтальних контрольних регіонах, ніж дорослі. Ці дані свідчать про те, що різні траєкторії розвитку для цих регіонів можуть лежати в основі посилення активності в дітей, або дорослих, що, в свою чергу, може стосуватися підвищеної імпульсної і ризикованої поведінки, що спостерігається протягом цього періоду розвитку (див. Рис. 4).

Диференціальний рекрутмент префронтальних і підкіркових областей був зареєстрований у багатьох дослідженнях fMRI розвитку (Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Зазвичай ці висновки трактуються з точки зору незрілих префронтальних регіонів, а не дисбалансу між префронтальним і субкортикальним регіональним розвитком. Наведені дані про префронтальні регіони в направленні відповідних дій у різних контекстах (Міллер і Коен, 2001) незріла префронтальна діяльність може перешкоджати належній оцінці майбутніх результатів і оцінці ризикованого вибору, і, таким чином, може бути менш впливовим на оцінку винагород, ніж аккумбенс. Ця закономірність узгоджується з попередніми дослідженнями, які показують підвищену підкоркову, відносно кортикальної активності, коли рішення змінюються безпосередньо на довгострокові вигоди (McClure, Laibson, Loewenstein, & Cohen, 2004). Крім того, активність accumbens показала, що fMRI позитивно корелює з подальшою поведінкою ризику (Кухнен і Натсон, 2005). У підлітковому віці, відносно дитинства або дорослого віку, незріла вентральна префронтальна кора може не забезпечити достатній верхній-вниз контроль над сильно активованими областями обробки винагороди (наприклад, accumbens), що призводить до меншого впливу префронтальних систем (орбітофронтальна кора) відносно accumbens в винагороді оцінка.
Чому мозок буде запрограмований таким чином?
Підлітковий вік - це перехідний період між дитинством і дорослим, часто зустрічається з пубертатом. Статеве дозрівання означає початок статевого дозрівання (Graber & Brooks-Gunn, 1998) і можуть бути визначені біологічними маркерами. Підлітковий вік можна охарактеризувати як прогресивний перехід до дорослого віку з туманним онтогенетичним курсом (Спис, 2000). Еволюційно кажучи, підлітковий вік - це період, коли набувають навичок незалежності, щоб збільшити успіх у відділенні від захисту сім'ї, але збільшують шанси на шкідливі обставини (наприклад, травми, депресії, тривоги, вживання наркотиків і наркоманії (Келлі, Шокет і Лендрі, 2004). Поведінки, що шукають незалежність, поширені серед різних видів, наприклад, збільшення соціальної взаємодії, спрямованої на однолітків, і посилення інновацій та поведінки, що приймає ризики. Психосоціальні фактори впливають на схильність підлітків до ризикованої поведінки. Однак, ризикована поведінка є продуктом біологічно обумовленого дисбалансу між підвищеною новизною та пошуком сенсацій у поєднанні з незрілою “саморегулівною компетентністю” (Steinberg, 2004). Наші нейробіологічні дані свідчать про те, що це відбувається через диференційну розробку цих двох систем (лімбічні та контрольні).
Спекуляція дозволить припустити, що ця модель розвитку є еволюційною особливістю. Ви повинні займатися ризикованою поведінкою, щоб залишити родину і село, щоб знайти партнера і взяти на себе ризик у той самий час, коли гормони підштовхують підлітків до пошуку сексуальних партнерів. У сучасному суспільстві, коли підлітковий вік може продовжуватися нескінченно, з дітьми, які живуть з батьками і мають фінансову залежність і обирають товаришів пізніше в житті, ця еволюція може вважатися недоцільною.
Існують дані про види видів для посиленого пошуку новизни та прийняття ризику під час підліткового віку. Пошук інших однолітків і боротьба з батьками, які допомагають підліткам вийти з домашньої території для спарювання, спостерігається в інших видах, включаючи гризунів, нечеловецьких приматів і деяких птахів (Спис, 2000). У порівнянні з дорослими, щури-підлітки показують підвищену поведінку, що шукає новизну, у парадигмі новизни для вільного вибору (Laviola et al., 1999). Нейрохімічні дані свідчать про те, що баланс у підлітковому мозку між кортикальними і підкорковими системами дофаміну починає зміщуватися в бік більших рівнів кортикального дофаміну в підлітковому віці (Спис, 2000). Подібне затяжне дофамінергічне збереження в підлітковому віці до дорослого віку було показано і в префронтальній корі нечеловецьких приматів (Розенберг і Льюїс, 1995). Таким чином, ця підвищена очевидність ризику виявляється у різних видів і має важливі адаптивні цілі.
Біологічні схильності, розвиток і ризик
Індивідуальні відмінності в контролі імпульсів і прийнятті ризиків визнані в психології протягом деякого часу (Бентін, Словіч та Северсон, 1993). Можливо, одним з класичних прикладів індивідуальних відмінностей, про які свідчать ці здібності в соціальній, когнітивній та розвивальної літературі психології, є затримка задоволення (Мішель, Шода та Родрігес, 1989). Затримка задоволення, як правило, оцінюється в 3 - до 4-річних малюків. Малюку запитують, чи вони воліють невелику винагороду (одну печиво) або велику винагороду (два куки). Потім дитині говорять, що експериментатор залишить кімнату, щоб підготуватися до майбутніх заходів, і пояснює дитині, що якщо вона залишиться на своєму місці і не їсть печива, вона отримає велику винагороду. Якщо дитина ні, або не може чекати, вона повинна подзвонити дзвону, щоб викликати експериментатора і тим самим отримати меншу винагороду. Як тільки зрозуміло, що дитина розуміє завдання, вона сидить за столом з двома нагородами і дзвоном. Відволікання в кімнаті мінімізовано, без іграшок, книг або фотографій. Експериментатор повертається після 15 хв або після того, як дитина подзвонив дзвоном, з'їдав нагороди або виявив будь-які ознаки лиха. Мішель показав, що діти, як правило, поводяться одним з двох способів: (1) або вони дзвонять дзвони майже відразу, щоб мати печиво, що означає, що вони отримують тільки один; або (2) вони чекають і оптимізують свої прибутки і отримують обидва файли cookie. Це спостереження свідчить про те, що деякі люди краще за інших у своїй здатності контролювати імпульси перед обличчям надзвичайно важливих стимулів, і ця упередженість може бути виявлена в ранньому дитинстві (Mischel et al., 1989), і вони, здається, залишаються протягом юності та молодості (Eigsti et al., 2006).
Що може пояснити індивідуальні відмінності в оптимальному прийнятті рішень і поведінці? Деякі теоретики постулювали, що дофамінергічна мезолімбічна схема, залучена в обробку винагороди, лежить в основі ризикованої поведінки. Індивідуальні відмінності в цій схемі, такі як алельні варіанти в генів, пов'язаних з дофаміном, що призводять до занадто мало або занадто багато дофаміну в підкіркових областях, можуть стосуватися схильності до ризикованої поведінки (O'Doherty, 2004). Показано, що nucleus accumbens зростає активність безпосередньо перед прийняттям ризикованого вибору на парадигмах монетарного ризику (Кухнен і Натсон, 2005; Matthews et al., 2004; Montague & Berns, 2002) і, як описано раніше, підлітки показують перебільшену активність до нагородження результатів щодо дітей або дорослих (Ernst et al., 2005; Galvan et al., 2006). У сукупності ці дані свідчать про те, що підлітки можуть бути більш схильними до ризикованого вибору як групи (Садівник і Штайнберг, 2005), але деякі підлітки будуть більш схильними, ніж інші, брати участь у ризикованій поведінці, ставлячи їх до потенційно більшого ризику негативних наслідків. Тому важливо враховувати індивідуальну мінливість при дослідженні складних взаємовідносин мозку та поведінки, пов'язаних з прийняттям ризику та обробкою винагороди у популяціях, що розвиваються.
Досліджувати індивідуальні відмінності в ризикованій поведінці Galvan et al. (2007) Нещодавно досліджено зв'язок між діяльністю у пов'язаній з винагородою нейронної схемою в очікуванні великої грошової винагороди з ознаками особистості, які визначають ризик і імпульсивність в підлітковому віці. Функціональна магнітно-резонансна томографія та анонімні рейтингові шкали саморепортажу ризикованої поведінки, сприйняття ризику та імпульсивності були отримані у осіб у віці від 7 до 29 років. Існувала позитивна асоціація між діяльністю accumbens і ймовірністю залучення до ризикованої поведінки в процесі розвитку. Ця діяльність змінювалася залежно від оцінки індивідуумів очікуваних позитивних або негативних наслідків такої поведінки. Ті особи, які сприймали ризиковану поведінку як призводять до тяжких наслідків, активізували аккумсенів менше за винагороду. Цю асоціацію в основному керували діти, а дорослі оцінювали наслідки такої поведінки. Оцінки імпульсивності не були пов'язані з активністю аккумсена, а з віком. Ці дані свідчать про те, що під час підліткового віку деякі люди можуть бути більш схильними до ризикованої поведінки через зміни у розвитку, пов'язані з мінливістю у схильності даного індивіда до ризикованої поведінки, а не до простих змін імпульсивності (див. Рис. 5).

Поведінка підлітків неодноразово характеризувалася як імпульсивна та ризикована (Steinberg, 2004, 2007), проте цей огляд літературних зображень свідчить про різні нейробіологічні субстрати та різні траєкторії розвитку цих поведінків. Зокрема, імпульсивність пов'язана з незрілим вентральним префронтальним розвитком і поступово зменшується від дитинства до дорослого життя (Кейсі, Гальван та ін., 2005). Негативна кореляція між рейтингами імпульсивності та віком у дослідженні Galvan et al. (2007) далі підтримує це поняття. На відміну від цього, прийняття ризику пов'язане зі збільшенням активності аккумів (Кухнен і Натсон, 2005; Matthews et al., 2004; Montague & Berns, 2002), що перебільшено у підлітків, у порівнянні з дітьми та дорослими (Ernst et al., 2005; Galvan et al., 2006). Таким чином, вибір і поведінка підлітків не можна пояснити імпульсивністю або затяжним розвитком самої префронтальної кори, оскільки дітям передбачається, що вони будуть більш ризикованими. Отримані дані забезпечують нейронну основу для того, чому деякі підлітки перебувають під більшим ризиком, ніж інші, але далі забезпечують основу для того, як поведінка підлітків відрізняється від дітей і дорослих при прийнятті ризику.
У сукупності ці дані свідчать про те, що хоча підлітки як група вважаються ризикуючими (Садівник і Штайнберг, 2005), деякі підлітки будуть більш схильними, ніж інші, до ризикованої поведінки, ставлячи їх до потенційно більшого ризику негативних наслідків. Ці результати підкреслюють важливість врахування індивідуальної мінливості при вивченні складних взаємозв'язків мозку та поведінки, пов'язаних з прийняттям ризику та обробкою винагороди у популяціях, що розвиваються. Крім того, ці індивідуальні відмінності та відмінності у розвитку можуть допомогти пояснити вразливість деяких людей до ризику, пов'язаного з вживанням речовин, і, зрештою, з пристрастю.
Висновки
Аналіз людських зображень показує структурні та функціональні зміни у фронтотріальних регіонах (Giedd et al., 1996, 1999; Jernigan et al., 1991; Sowell et al., 1999; для перегляду, Кейсі, Гальван та ін., 2005) що здається паралельним збільшенням когнітивного контролю та саморегуляції (Casey, Trainor et al., 1997; Luna & Sweeney, 2004; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004; Дивіться також Steinberg, 2008, ця проблема). Ці зміни, здається, показують зрушення в активації префронтальних регіонів від дифузного до більш фокусного набору з часом (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor et al., 1997; Durston et al., 2006; Moses et al., 2002) і підвищений підбір підкоркових областей підліткового віку (Casey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Хоча нейровизуализирующие дослідження не можуть остаточно охарактеризувати механізм таких змін у розвитку, ці зміни в обсязі і структурі можуть відображати розвиток і вдосконалення проекцій в ці і з цих областей мозку під час дозрівання, що вказує на точну настройку системи з розвитком.
Взяті разом, отримані тут результати свідчать про те, що підвищена ризикована поведінка в підлітковому віці пов'язана з різними траєкторіями розвитку підкіркових задоволень і коркових контрольних областей. Ці зміни в розвитку можуть бути посилені індивідуальними відмінностями в діяльності систем винагород. Незважаючи на те, що підлітковий вік був визначений як період, який характеризується пошуком винагороди та поведінкою ризику (Садівник і Штайнберг, 2005; Спис, 2000) індивідуальні відмінності в нервових реакціях на винагороду, схиляють деяких підлітків до більшого ризику, ніж інші, ставлячи їм більший ризик негативних наслідків. Ці знахідки забезпечують важливу основу шляхом синтезу різноманітних знахідок, пов'язаних з поведінкою ризику в підлітковому віці та розумінням індивідуальних відмінностей та маркерів розвитку для схильності до негативної поведінки.
Подяки
Цю роботу частково підтримали гранти Національного інституту зловживання наркотиками R01 DA18879 і Національного інституту психічного здоров'я 1P50 MH62196.
посилання
- Benthin A, Slovic P, Severson H. Психометричне дослідження сприйняття ризику підлітків. Журнал юності. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Стимулююча активація мозку у підлітків: подібності та відмінності від молодих дорослих. Журнал Neuroscience. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Роль концептуальних знань у розвитку організації в дитячій пам'яті. В: Brainerd CJ, Pressley M, редактори. Основні процеси розвитку пам'яті: прогрес у дослідженнях когнітивного розвитку. Нью-Йорк: Springer-Verlag; 1985. С. 103 – 142.
- Bjorkland DF. Як вікові зміни в базі знань сприяють розвитку дитячої пам'яті: інтерпретаційний огляд. Розвиток розвитку. 1987;7: 93-130.
- Буржуа JP, Гольдман-Ракич П.С., Ракич П. Синаптогенез в префронтальній корі макак-резусів. Кора головного мозку. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Рейна В.Ф. Незалежність пам'яті і втручання пам'яті в когнітивний розвиток. Психологічний огляд. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Розвиваючі зміни в функціональній організації мозку людини для генерації слів. Кора головного мозку. 2005;15: 275-290. [PubMed]
- Бунге С.А., Дудукоков Н.М., Томасон М.Є., Вайдя С.Я., Габріелі Ю.Д. Внесок незрілих лобових часток до когнітивного контролю у дітей: Докази з fMRI. Neuron. 2002;33: 301-311. [PubMed]
- Випадок Р. Перевірка конструкта неоіагейської ємності. Журнал експериментальної дитячої психології. 1972;14: 287-302.
- Справа Р. Інтелектуальний розвиток: Народження до дорослого життя. Нью-Йорк: Академічна преса; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. та ін. Наслідки правильної фронтостріальної схеми в пригніченні реакції та розладі дефіциту уваги / гіперактивності. Журнал Американської академії дитячої та юнацької психіатрії. 1997;36: 374-383. [PubMed]
- Кейсі Б.Я., Гальван А, Харе Т.А. Зміни функціональної організації мозку під час когнітивного розвитку. Актуальна думка в невробіології. 2005;15: 239-244. [PubMed]
- Кейсі BJ, Giedd JN, Томас KM. Структурно-функціональний розвиток мозку та його зв'язок з когнітивним розвитком. Біологічна психологія. 2000a;54: 241-257. [PubMed]
- Кейсі Б.Я., Томас К.М., Девідсон М.К., Кунц К., Францен П.Л. Дисоціація стриатиальной і гіпокампальної функцій розвитку з завданням стимулювання-відповіді. Журнал Neuroscience. 2002a;22: 8647-8652. [PubMed]
- Кейсі Б.Я., Томас К.М., Валлійська Т.Ф., Бадгайян Р.Д., Еккард CH, Jennings JR, et al. Дисоціація конфлікту реакції, вибір уваги та тривалість з функціональною магнітно-резонансною візуалізацією. Праці Національної академії наук. 2000b;97: 8728-8733.
- Кейсі Б.Я., Тоттенхем Н., Фосселла Дж. Клінічні, візуалізаційні, ураження і генетичні підходи до моделі когнітивного контролю. Психобіологія розвитку. 2002b;40: 237-254. [PubMed]
- Кейсі BJ, Tottenham N, Liston C, Durston S. Imaging розвивається мозку: Що ми дізналися про когнітивний розвиток? Тенденції когнітивної науки. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Дослідження функціональної МРТ щодо розвитку префронтальної активації під час виконання завдання «go-no-go». Журнал когнітивної неврології. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Регіони мозку, що опосередковують використання гнучкого правила під час розвитку. Журнал Neuroscience. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Стійкість до перешкод: зміна основного механізму обробки. В: Хоув М.Л., Паснак Р., редактори. Виникаючі теми в когнітивному розвитку Том 1: Основи. Нью-Йорк: Springer; 1993. С. 3 – 27.
- Алмаз А. Розвиток здатності використовувати нагадування для керівництва діями, як це показано в роботі дітей на АБ. Дитячий розвиток. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Параметричні маніпуляції з конфліктами і конкуренцією за допомогою швидкого змішаного випробування подій, пов'язаних з fMRI. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, et al. Перехід від дифузної до осередкової кортикальної активності з розвитком. Наука розвитку. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Анатомічна МРТ розвивається людського мозку: Що ми дізналися? Журнал Американської академії дитячої психіатрії. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. Моніторинг ризикованої поведінки молоді - Сполучені Штати, 2005, резюме спостереження. Щотижневий Звіт про захворюваність та смертність. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Мішель W, Шода Y, Ayduk O, Dadlani MB, et al. Прогнозування когнітивного контролю від дошкільного до пізнього підліткового та молодого дорослого віку. Психологічна наука. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala і nucleus accumbens в відповідях на отримання і опущення доходів у дорослих і підлітків. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Ешель Н., Нельсон Е.Е., Блер Р.Ю., Пайн Д.С., Ернст М. Нейронні субстрати вибору вибору у дорослих і підлітків: Розвиток вентролатеральних префронтальних і передніх поясних поясів. Нейропсихологія. 2007;45: 1270-1279. [PMC безкоштовна стаття] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Спонтанна вербальна репетиція в пам'яті як функція віку. Дитячий розвиток. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Раніше розвиток аккумбенса щодо орбітофронтальної кори може спричинити ризиковану поведінку у підлітків. Журнал Neuroscience. 2006;26: 6885-6892. [PubMed]
- Гальван А, Харе Т, Восс Н, Гловер Г, Кейсі Б.Я. Ризик та підлітковий мозок: хто ризикує? Наука розвитку. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Вплив однолітків на прийняття ризиків, переваги ризику та ризикований процес прийняття рішень у підлітковому та дорослому віці: експериментальне дослідження. Психологія розвитку. 2005;41: 625-635. [PubMed]
- Giedd JN. Структурна магнітно-резонансна томографія підліткового мозку. Аннали Нью-Йоркської академії наук. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Лю H, Zijdenbos A, et al. Розвиток мозку в дитячому та підлітковому віці: дослідження поздовжнього МРТ. Природознавство. 1999;2: 861-863.
- Giedd JN, Снелл JW, Lange N, Rajapakse JC, Кейсі BJ, Козуч PL, та ін. Кількісна магнітно-резонансна томографія розвитку людського мозку: століття 4 – 18. Кора головного мозку. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Динамічне відображення розвитку людського кори в дитинстві до раннього дорослого віку. Праці Національної академії наук Сполучених Штатів Америки. 2004;101: 8174-8179. [PMC безкоштовна стаття] [PubMed]
- Грабер Я.А., Брукс-Ганн Дж. Пубертет. В: Блехман Е.А., Браунелл К.Д., редактори. Поведінкова медицина і жінки всеосяжний підручник. Нью-Йорк, Нью-Йорк: Guilford Press; 1998. С. 51 – 58.
- Харе Т.А., Восс Х.У., Гловер Г.Х., Кейсі Б.Я. Мозок підлітка і потенційний ризик для тривоги і депресії. 2007a Подано для публікації.
- Харе Т.А., Восс Х.У., Гловер Г.Х., Кейсі Б.Я. Конкуренція між префронтальними і підкірковими лімбічними системами лежить в основі емоційної реактивності в підлітковому віці. 2007b Подано для публікації.
- Harnishfeger KK, Bjorkland F. Онтогенез механізмів гальмування: оновлений підхід до когнітивного розвитку. У: Хоув М.Л., Паснек Р., редактори. Виникаючі теми в когнітивному розвитку. Vol. 1. Нью-Йорк: Springer-Verlag; 1993. С. 28 – 49.
- Hikosaka K, Watanabe M. Затримка активності орбітальних і латеральних префронтальних нейронів мавпи різними з різними нагородами. Кора головного мозку. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Синаптична щільність в лобовій корі людини - розвиваючі зміни і наслідки старіння. Дослідження мозку. 1979;163: 195-205. [PubMed]
- Hyman SE, Маленка РК. Наркоманія і мозок: нейробіологія примусу і її наполегливість. Природи огляду нейронауки. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Аномалії магнітно-резонансної томографії в чотирьох ядрах і корі головного мозку при шизофренії. Архіви загальної психіатрії. 1991;48: 811-823.
- Кітінг Д.П., Боббіт Б.Л. Індивідуальні та розвиваючі відмінності в компонентах когнітивної обробки розумових здібностей. Дитячий розвиток. 1978;49: 155-167.
- Kelley AE, Schochet T, Ландрі С. Аннали Нью-Йоркської академії наук. 2004;1021: 27-32. [PubMed]
- Knutson B, Адамс CM, Fong GW, Hommer D. Передбачення збільшення грошової винагороди вибірково набирає nucleus accumbens. Журнал Neuroscience. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Нейронні основи прийняття фінансових ризиків. Neuron. 2005;47: 763-770. [PubMed]
- Лавіола Г., Адріані У., Терранова М. Л., Герра Г. Психобіологічні фактори ризику уразливості до психостимуляторів у підлітків і моделях тварин. Неврологічні та біологічні огляди. 1999;23: 993-1010. [PubMed]
- Лавіола G, Macri S, Morley-Fletcher S, Adriani W. Анотація ризикованої поведінки у підлітків мишей: психобіологічні детермінанти і ранній епігенетичний вплив. Неврологічні та біологічні огляди. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Фронтострітальна мікроструктура модулює ефективний набір когнітивного контролю. Кора головного мозку. 2005;16: 553-560. [PubMed]
- Luna B, Суїні Я.А. Виникнення спільної функції мозку: FMRI вивчає розвиток інгібування відповіді. Аннали Нью-Йоркської академії наук. 2004;1021: 296-309. [PubMed]
- Луна Б, Тульборн К.Р., Муньос Д.П., Мерріам Е.П., Гарвер К.Е., Мінсеєв Н.Ю. Дозрівання широко поширеної функції мозку підносить когнітивний розвиток. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Селективна активація ядра accumbens під час прийняття рішень. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Травня JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Пов'язана з подіями функціональна магнітно-резонансна візуалізація пов'язаної з винагородами схеми мозку у дітей та підлітків. Біологічна психіатрія. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Окремі нейронні системи цінують негайну грошову винагороду. Наука. 2004;306: 503-507. [PubMed]
- Міллер Е.К., Коен JD. Інтегративна теорія функції префронтальної кори. Щорічний огляд нейронауки. 2001;24: 167-202.
- Мішель W, Шода Y, Родрігес М.І. Затримка задоволення у дітей. Наука. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Незрілість підлітків у залученні уваги мозку до емоційного міміки. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Бернс Г.С. Нейронна економіка та біологічні субстрати оцінки. Neuron. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Франк LR, Stiles J. Функціональна МРТ глобальної та локальної обробки у дітей. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Всі разом зараз: Коли дисоціації між знанням і дією зникають. Психіологічна наука. 2001;12: 335-337.
- Надя З, Вестерберг Х, Клінгберг Т. Дозрівання білої речовини пов'язане з розвитком когнітивних функцій в дитинстві. Журнал когнітивної неврології. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Анотація винагороди і покарання уявлень в людській орбітофронтальній корі. Природа Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Нагороджуйте репрезентації та пов'язані з винагородою навчання в мозку людини. Актуальні думки в невробіології. 2004;14: 769-776.
- Pascual-Leone JA. Математична модель переходу в стадії розвитку Піаже. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Олдрідж JW, Чжуан X. Гіпердопамінергічні мутантні миші мають вищу "бажану", але не "пристрасть" до солодких нагород. Журнал Neuroscience. 2003;23: 9395-9402. [PubMed]
- Рейна В.Ф., Фарлі Ф. Ризик і раціональність у прийнятті рішень підлітками: наслідки для теорії, практики та державної політики. Психологічна наука в суспільному інтересі. 2006;7: 1-44.
- Розенберг Д.Р., Льюїс Д.А. Постнатальне дозрівання дофамінергічної іннервації префронтальних і моторних кори мавпи: імуногістохімічний аналіз тирозингидроксилази. Журнал порівняльної неврології. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Функціональна фронталізація з віком: Візуалізація траєкторій нейроразвития з fMRI. Неврологічні та біологічні огляди. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Вісчер КМ, Miezin FM, Petersen SE. Функціональні нейроанатомічні відмінності між дорослими і дітьми шкільного віку при обробці окремих слів. Наука. 2002;296: 1476-1479. [PubMed]
- Шульц В. Поведінкові теорії та нейрофізіологія винагороди. Щорічні огляди психології. 2006;57: 87-115.
- Соуелл Е.Р., Петерсон Б.С., Томпсон П.М., Вітаємо SE, Henkenius AL, Toga AW. Зіставлення коркових змін через тривалість життя людини. Природознавство. 2003;6: 309-315.
- Соуелл Е.Р., Томпсон П.М., Холмс С.Ю., Джерніган Т.Л., Тога А.В. Докази in vivo для дозрівання головного мозку у фронтальних і стриатних областях. Природознавство. 1999;2: 859-861.
- Соуелл Е.Р., Томпсон П.М., Тога А.В. Картографічні зміни в корі людини протягом всього періоду життя. Нейрон. 2004;10: 372-392. [PubMed]
- Спис LP. Підлітковий мозок і вікові поведінкові прояви. Неврологічні та біологічні огляди. 2000;24: 417-463. [PubMed]
- Штейнберг Л. Прийняття ризику в підлітковому віці: що змінюється і чому? Аннали Нью-Йоркської академії наук. 2004;1021: 51-58. [PubMed]
- Штейнберг Л. Ризик в підлітковому віці: нові перспективи від мозку і поведінкової науки. Поточні напрямки в психологічній науці. 2007;16: 55-59.
- Стейнберг Л. Соціальна перспектива неврології на ризик підлітків. Розвиток розвитку. 2008;28: 78-106. [PMC безкоштовна стаття] [PubMed]
- Тамм Л, Менон В., Рейс А.Л. Дозрівання функції мозку пов'язане з інгібуванням відповіді. Журнал Американської академії дитячої та юнацької психіатрії. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Докази відмінностей у розвитку вивчення неявної послідовності: дослідження FMRI дітей і дорослих. Журнал когнітивної неврології. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Квіти DL, Zeffiro TA, Eden GF. Розробка нейронних механізмів для читання. Природознавство. 2003;6: 767-773.
- Volkow ND, Li TK. Наркоманія: Невробіологія поведінки перейшла в ногу. Природи огляду нейронауки. 2004;5: 963-970.
- Юргелун-Тод Д. Емоційні та пізнавальні зміни підліткового віку. Актуальна думка в невробіології. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Передача дофаміну в стриатуме людини під час грошової винагороди. Журнал Neuroscience. 2004;24: 4105-4112. [PubMed]