Психіатрія біолів. 2008 грудень 1; 64 (11): 941-50. Epub 2008 лип. 26.
Teegarden SL, Nestler EJ, Bale TL.
Source
Кафедра біології тварин, Пенсильванський університет, Філадельфія, Пенсільванія 19104-6046, США.
абстрактний
передумови:
Чутливість до винагороди залучена як фактор, що впливає на поведінку, пов'язану з зловживанням наркотиками, а також як переїдання. Однак основні механізми, що сприяють чутливості до винагород, невідомі. Ми припустили, що порушення регуляції в сигналізації дофаміну може бути основною причиною підвищеної чутливості винагороди, завдяки чому корисні стимули можуть діяти для нормалізації системи.
МЕТОДИ:
Ми використовували генетичну модель миші з підвищеною чутливістю до винагороди, мишею Delta FosB, що перенапружує, щоб вивчити зміни шляху винагороди у відповідь на приємну дієту з високим вмістом жиру. Маркери нагородних сигналів у цих мишей були досліджені як в основному, так і після наступних 6 тижнів приємного впливу дієти. Мишей досліджували в поведінковому тесті після відміни дієти з високим вмістом жиру, щоб оцінити вразливість цієї моделі до усунення корисних стимулів.
РЕЗУЛЬТАТИ:
Наші результати демонструють змінену активацію шляху винагороди по схемі ядерних гіпоталамо-вентральних ділянок тегментальної області, що виникає внаслідок надмірної експресії Delta FosB в прилеглих ядрах і смугастих областях. Рівні білка, що зв'язує фосфорильований циклічний аденозинмонофосфат (цАМФ) (pCREB), нейротрофічний фактор, отриманий від мозку (BDNF), а дофамін та циклічний регульований фосфопротеїном монофосфат аденозинового монофосфату з молекулярною масою 32 kDa (DARPP-32) у ядрах ядер знижувались у мишей Delta FosB, що свідчить про зменшення сигналу дофаміну. Шість тижнів впливу дієти з високим вмістом жиру повністю покращили ці відмінності, виявивши потужну корисну дієту дієти. Миші Delta FosB також показали значне збільшення опорно-рухової активності та відповіді, пов'язані з тривожністю 24 годин після виведення жиру.
ВИСНОВКИ:
Ці результати встановлюють основну чутливість до змін у винагороді, пов’язаних з порушенням регуляції Delta FosB та дофамінової сигналізації, які можуть бути нормалізовані за допомогою дієт, що піддаються смаку, і можуть бути схильним фенотипом при деяких формах ожиріння..
Вступ
Незважаючи на все більші знання про нейронні системи, які контролюють апетит і ситості, показники ожиріння продовжують зростати в Сполучених Штатах. Поточні методи лікування лікарськими засобами мають обмежену ефективність, а модифікації поведінки страждають від мінімальної довгострокової відповідності (1). Споживання калорійно-густої, смачної їжі пов'язане із зміною шляху стресу та винагород у мозку, що дозволяє припустити, що корисні властивості таких продуктів можуть перекривати сигнали енергетичного балансу (2-4). Їжа з високим вмістом жиру діє як природна винагорода, активізуючи центри нагородження мозку таким чином, як зловживання наркотиками, і як таку використовували в парадигмах самовведення (5-8). Таким чином, цілком ймовірно, що поведінка та мотивація до переїдання та зловживання наркотиками мають спільні основні механізми, що може відкрити нові шляхи лікування для обох станів.
Вивчаючи взаємозв'язок між смачною їжею та шляхами, що регулюють винагороду та стрес у мозку, Ми раніше визначали молекулярні та біохімічні маркери зниженої винагороди та підвищеного стресу після відмови від приємної дієти з високим вмістом жиру (HF). Подібно до зловживань наркотиками, в наших дослідженнях вплив сприятливої дієти призвів до підвищення рівня фактора транскрипції ΔFosB в ядерних ядрах (NAc), центральної структури винагороди мозку (9, 10). Миші, які індуктивно перенапружують ΔFosB, демонструють посилений інструментальний реагування на харчову винагороду (11), що робить їх цінним інструментом для вивчення ролі чутливості до винагороди та тривалої дирегуляції системи винагород у молекулярних та біохімічних реакціях на приємну дієту.
У цьому дослідженні ми використовували мишей, що експресують ΔFosB, для дослідження довгострокових змін маркерів нагороди в нейроциркуляції NAc-гіпоталамус-вентральний ділянка (ВТА) у відповідь на приємну дієту HF. Спираючись на попередні дослідження цих мишей, чутливих до нагород, ми висунули гіпотезу, що індуковані ΔFosB зміни чутливості до винагороди включають порушення регуляції дофамінової сигналізації внаслідок зворотного зв'язку NAc до VTA. Крім того, ми висловлювали гіпотезу, що вплив природного винагороди за енергетично щільну дієту HF нормалізує дофамінергічну систему у цих мишей, що призведе до перебільшеної реакції на стрес, що відходить від цієї дієти з ВЧ. Унікальний аспект використання приємної дієти як корисної речовини дозволяє нам включати гіпоталамічні вкладення для винагородження схеми у фенотипі, який може бути передбачувальним для населення, схильного до стійкого до лікування ожиріння. Щоб вивчити цю гіпотезу, ми вивчали маркери нейромедіації дофаміну, включаючи pCREB, BDNF та DARPP-32 в NAc та тирозин гідроксилазу та транспортер дофаміну в VTA, після впливу HF. Ми також вивчали конкретні маркери енергетичного балансу, як відомо, що впливають на дофаміновий викид, включаючи рецептори лептину та орексину у ВТА та експресію орексину в латеральному гіпоталамусі.
Матеріали та методи
Звірята
Чоловічі бітрансгенні миші, які індуктивно переекспресують ΔFosB у динорфін-позитивних нейронах NAA та дорсального стриатуму (Kelz et al., 1999), генерувались на змішаному тлі (ICR: C57Bl6 / SJL) у Техаському університеті Південно-Західного медичного центру та підтримували та випробуваний в університеті Пенсільванії. Всіх мишей підтримували на доксицикліні (100 мкг / мл у питній воді) до прибуття до університету Пенсільванії. Для індукції надмірної експресії доксициклін було видалено (n = 23) (12). Контрольні миші (n = 26) продовжували отримувати препарат. Мишей були призначені групам дієти через вісім тижнів після видалення доксициліну, в цей час було показано, що експресія досягає максимальних рівнів (13). Мишей підтримували на циклі 12: 12 світло-темний (світильники на 0700) із наявністю їжі та води ad libitum. Усі дослідження проводилися згідно з експериментальними протоколами, затвердженими Комітетом з питань догляду та використання тварин Університету Пенсільванії, а всі процедури проводились відповідно до інституційних вказівок.
Дієтичне опромінення
Мишей підтримували на домашньому чау (n = 16) або поміщали на HF (n = 16-17) протягом шести тижнів. Домашня чау (Purina Lab Diet, Сент-Луїс, Міссурі) містила 4.00 ккал / г, що складається з білка 28%, жиру 12% та вуглеводів 60%. HF дієта (Research Diets, New Brunswick, NJ) містила 4.73 ккал / г, що складається з білка 20%, жиру 45% та вуглеводів 35%.
Біохімія та експресія генів
Мишей аналізували після шести тижнів впливу дієти. Мозок видаляли з черепа і заморожували цілими на сухому льоду або NAc розсікали (приблизно 0.5 - 1.75 мм від брегми, на глибині 3.5 - 5.5 мм) і заморожували у рідкому азоті. Тканина зберігалася при температурі -80 ° C до аналізу.
Біохімічні аналізи
Методи вестерн-блотів описані в додаткових матеріалах. Використовувані антитіла були: Cdk5, CREB і BDNF (1: 500, Santa Cruz Biotechnology, Santa Cruz, CA) і фосфо-CREB (pCREB) (Ser 133) (1: 500, Cell Signaling Technology, Danvers, MA).
Авторедіографія рецепторів
Детальні методи авторадіографії описані в додаткових матеріалах. Використовувані ліганди були 2 nM H3 - SCH 23390 і 5 nM H3 - спіперон (PerkinElmer, Boston, MA).
Гібридизація in situ
Обробку тканин та гібридизацію проводили, як описано раніше (14). Зонд DARPP-32 був люб’язно наданий П. Грінґардом (Університет Рокфеллера), а зонд орексину - Дж. Елмквістом (Південно-Західний медичний центр Техаського університету). Слайди, проаналізовані на DARPP-32, були призначені для знімання протягом 3 днів, а слайди, проаналізовані на орексин, були призначені для знімання протягом 4 днів. Кількісне визначення плівкових зображень проводили, як описано раніше (10).
QRT-PCR
РНК виділяли з VTA та експресії окремих генів, оцінених за допомогою аналізу експресії генів TaqMan (Applied Biosystems, Foster City, CA). Детальні методи та статистичний аналіз можуть бути знайдені в додаткових матеріалах.
Поведінкові аналізи
Для того, щоб вивчити ефекти чутливості до винагороди на зміни поведінки, спричинені дієтою, підмножину мишей було вилучено з HF через чотири тижні впливу та повернуто до домашньої чау (n = контроль 9, n = 8 ΔFosB). Через двадцять чотири години після виведення мишей піддавали випробуванню під відкритим полем відповідно до нашої раніше опублікованої дієтичної парадигми виведення (10). Якщо коротко, мишу помістили в центр апарату під відкритим полем і стежили протягом п'яти хвилин. Виміряли загальні хрестові лінії, фекальні болі, час у центрі та хрести в центрі.
Статистика
Всі дані, крім Вестерн-блот, аналізували за допомогою двостороннього аналізу ANOVA, а потім тесту PLSD Фішера з лікуванням доксицикліном (експресія ΔFosB) та дієтичним станом як незалежними змінними. Для аналізів RT-PCR використовували знижене значення P для корекції багаторазового порівняння в групах споріднених генів (див. Додаткові матеріали). Вестерн-блот аналізували за допомогою t-критерію Стьюдента з обробкою доксицикліном як незалежною змінною, порівнюючи оптичні щільності всередині тієї ж плями. Усі дані представлені як середнє значення ± SEM.
результати
Базальні біохімічні відмінності
Для з’ясування молекулярних шляхів, що лежать в основі підвищеної чутливості до винагороди у мишей із надмірною експресією ΔFosB, у NAc досліджували рівні кількох ключових сигнальних молекул. Спостерігалася тенденція до підвищення рівня Cdk5 у NAc мишей ΔFosB порівняно з контрольними тваринами, що вивозили сміття, що утримувались на доксицикліні (F = 5.1, P = 0.08; рис. 1А). Миші ΔFosB виражали суттєво знижені рівні pCREB (F = 7.4, P <0.05; рис. 1B), а також загальні рівні CREB (F = 5.4, P = 0.05; рис. 1C). Значне зменшення BDNF спостерігалося також у NAc мишей ΔFosB (F = 10.6, P <0.05; рис. 1D).
малюнок 1
У мишей, що експресують ΔFosB, виявлені біохімічні маркери зниженої дофамінової сигналізації в NAc
Споживання їжі та маса тіла на дієті з високим вмістом жиру
Далі ми дослідили вплив природно корисної ВЧ дієти на зміни сигнальних молекул у мишей із надмірним вираженням ΔFosB. Не було відмінностей між мишами ΔFosB та контролем у споживанні їжі як у будинку, так і у СН. Однак спостерігалося загальне зниження споживання калорій, нормоване до маси тіла під впливом ВЧ, яке було специфічним для мишей ΔFosB (F = 11.2, P <0.01; рис. 2А). Наприкінці шести тижнів дієти, миші, які отримували СН, важили значно більше, ніж ті, що були на дієті чау (F = 17.2, P <0.001), а миші ΔFosB важили менше в порівнянні з контролем (F = 5.6, P <0.05; Рис. 2Б). Цей ефект був специфічним для відмінностей між групами на дієті чау (Р <0.05).
малюнок 2
ΔFosB миші, що перенапружують, не виявляли відмінностей у прийомі їжі на дієтах із чаую чи жиру (HF)
Біохімічні відмінності в дієті з високим вмістом жиру
Щоб визначити, як базальні відмінності в сигналі NAc можуть бути змінені HF дієтою, ті ж сигнальні білки, що вивчалися за базовою лінією, були досліджені на тварин, які отримували шість тижнів ВЧ. Суттєвих відмінностей у рівнях Cdk5 не було (рис. 3A). Рівні pCREB і загального CREB більше не відрізнялися після шести тижнів ВЧ (рис. 3B, C). Рівні BDNF були значно підвищені у мишей ΔFosB через шість тижнів впливу HF (F = 6.5, P = 0.05; рис. 3D).
малюнок 3
Поліпшена сигнальна різниця в дієті з високим вмістом жиру (HF), що спостерігається у NAc мишей, що перенапружують ΔFosB
Авторадіографія рецепторів дофаміну
Ми використовували авторадиографію рецепторів, щоб оцінити, чи викликані ΔFosB зміни сигналізації дофаміну в NAc пов'язані зі змінами в експресії рецепторів дофаміну (рис. 4А). Дієта з високим вмістом жиру, як виявилося, дещо збільшила щільність зв'язування дофамінових рецепторів D1 (P = 0.14), і ця різниця була більшою у мишей ΔFosB (рис. 4B). Також спостерігалась тенденція до збільшення площі зв'язування D1 після HF (P = 0.06), і post hoc тестування показало, що це є значущим у мишей ΔFosB (P <0.05; рис. 4C). На відміну від рецепторів D1, відсутність змін щільності зв'язування рецепторів D2 (контроль чау = 97.6 ± 6.9, контроль HF = 101.1 ± 8.2, ΔFosB чау = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) або області зв'язування (контроль чау = 47.3 ± 3.4, контроль HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) в NAc спостерігались.
малюнок 4
Дієта з високим вмістом жиру (HF) призвела до зміни зв'язування рецепторів дофаміну D1 та експресії DARPP-32 в ядрах ядер (NAc) мишей, що експресують ΔFosB, надвиражаючими
Вираз DARPP-32 в NAc
Гібридизацію in situ використовували для визначення рівнів експресії DARPP-32 в NAc (рис. 4D). Дієта з високим вмістом жиру значно збільшила експресію DARPP-32 у цій області мозку (F = 5.1, P <0.05), і спостерігалася значна взаємодія між дієтою та експресією ΔFosB (F = 8.9, P <0.05), причому миші ΔFosB демонстрували більший індуковані дієтою зміни (рис. 4Е). Базальна різниця в експресії DARPP-32 між контрольними та мишами ΔFosB була виявлена шляхом post hoc тестування (P <0.01), а також значне збільшення експресії DARPP-32 у мишей ΔFosB на HF (P <0.01).
Експресія генів у VTA
QRT-PCR використовували для оцінки змін у експресії генів у VTA, націлюючи кілька ключових генів, раніше залучених до регулювання винагороди. Всі зразки нормалізувались до β-актину. Щоб переконатися, що експресія β-актину не була змінена внаслідок лікування, було проведено окремий аналіз для порівняння β-актину з другим внутрішнім контролем, GAPDH. Суттєвих відмінностей у експресії β-актину не було (значення ΔCT, β-актин - GAPDH: контроль чау = 2.29 ± 0.21, контроль HF = 2.01 ± 0.04, ΔFosB чау = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
Тенденція до взаємодії між експресією ΔFosB та дієтичним спостереженням спостерігалась для експресії тирозингідроксилази (F = 3.6, P <0.06; рис. 5А). Шість тижнів впливу HF, здається, зменшували експресію тирозингідроксилази у контрольних мишей та збільшували експресію у мишей ΔFosB. Суттєва взаємодія між експресією ΔFosB та дієтою спостерігалась для експресії транспортера дофаміну (F = 6.7, P <0.03; рис. 5B). Подібно до тирозингідроксилази, вплив HF знижував експресію транспортера дофаміну у контрольних мишей та значно збільшував експресію у мишей ΔFosB (P <0.05). Базальна різниця в експресії транспортера дофаміну між мишами контролю та ΔFosB не досягла значущості (P = 0.16), але після 6 тижнів HF миші ΔFosB виражали суттєво підвищений рівень транспортера дофаміну порівняно з контролем (P <0.05).
малюнок 5
Вплив дієти з високим вмістом жиру (HF) та експресія ΔFosB призвели до зміни експресії ряду ключових молекул у VTA
Існувала тенденція, що вказує на вплив підвищеної експресії ΔFosB на зниження рівня TrkB у VTA (F = 5.7, P <0.04; рис. 5C). Хоча не було основних ефектів на експресію κ-опіоїдних рецепторів, спостерігалася тенденція до зниження експресії у мишей ΔFosB (P = 0.08; рис. 5D). Експресія рецептора лептину також визначалася у VTA. Було виявлено значний ефект від дієтичного впливу (F = 6.1, P <0.03), при цьому СН значно знижувала рівні рецептора лептину в VTA як у ΔFosB, так і у контрольних мишей (рис. 5Е). Також була досліджена експресія рецептора орексину 1 у VTA. Спостерігався значний вплив дієти на експресію рецептора орексину (F = 9.0, P <0.02), причому миші, що зазнавали дії ВЧ, експресували більш високі рівні VTA (рис. 5F). Також спостерігалася тенденція до мишей ΔFosB виражати загальні вищі рівні рецептора орексину 1 у цій області мозку (Р <0.05).
Експресія орексину в бічному гіпоталамусі
Ми вимірювали рівні орексину в латеральному гіпоталамусі, походження орексинергічної іннервації VTA, шляхом гібридизації in situ (рис. 6А). Існувала значна взаємодія між експресією ΔFosB та дієтичним впливом на експресію орексину (F = 9.1, P <0.01), при цьому HF значно підвищував рівень орексину у контрольних мишей (P <0.05) та зменшував експресію у мишей ΔFosB (рис. 6B). Хоча не було значущих відмінностей у експресії орексину в базальному стані, після 6 тижнів СН, миші ΔFosB виражали суттєво знижені рівні орексину порівняно з контролем (Р <0.05).
малюнок 6
Дієта з високим вмістом жиру (HF) мала різний вплив на експресію орексину у контрольних (Ctrl) та ΔFosB мишей, що перенапружували
Beповедінкові аналізи
Щоб оцінити зміни збудження та емоційності внаслідок зміни дієти, мишей піддавали відкритому тесту через 24 години після відмови від ВЧ дієти (10). Загальний перехід ліній, який оцінювали як міру збудження, суттєво впливав на експресію ΔFosB (F = 6.6, P <0.05) та дієту (F = 4.6, P <0.05; рис. 7А). Миші ΔFosB були більш активними в новому середовищі, ніж контрили, і post hoc тестування показало, що миші, виведені з HF, були значно активнішими, ніж ті, що зазнали чау (P <0.05). Калові болі зараховували як міру поведінки, подібної до тривоги (10). Був основний ефект експресії ΔFosB (F = 10.2, P <0.01), коли миші з надмірною експресією ΔFosB виробляли більше фекальних болі в новому середовищі, особливо в домашніх чау-групах та групах, що виводили HF (рис. 7B). Миші ΔFosB, які перебувають на ВЧ-дієті, виробляли менше калових болі, ніж ті, що утримувались на чау-їжі, і тих, кого вилучили за 24 години до тесту. Здавалося, контрольні миші не впливали на дієту. Суттєвого впливу експресії ΔFosB або дієти на час перебування в центрі відкритого поля не спостерігалося (контроль чау = 14.5 ± 3.1 сек, контроль ВЧ = 18.0 ± 3.2 сек, контроль Ш / Г = 15.4 ± 1.9 сек, ΔFosB чау = 16.9 ± 2.4 сек, ΔFosB HF = 13.1 ± 3.9 сек, ΔFosB Ш / Г = 19.8 ± 2.6 сек).
малюнок 7
Миші, що експресують ΔFosB, були більш чутливими до ефектів відміни дієти з високим вмістом жиру (HF)
Обговорення
У лікуванні ожиріння існує критична потреба у виявленні факторів, які впливають на сприйнятливість до переїдання та збільшення ваги. Шляхи нагородження мозку відіграють важливу роль у мотивації та реагуванні на смачні страви та дієтичні зміни (6, 10, 15, 16). Оскільки орексигенні та анорексигенні сигнали можуть безпосередньо впливати на нагородну сигналізацію через ланцюг гіпоталамус-VTA-NAc, з'ясування генів, що реагують на багаті енергією смачні дієти в центрах нагородження, може забезпечити нові терапевтичні цілі при лікуванні ожиріння (17, 18). Таким чином, ми розглянули біохімічні та молекулярні маркери сигналу нагородження та енергетичного балансу по ланцюгу гіпоталамус-VTA-NAc у відповідь на дієту HF у мишей, що переважають ΔFosB, як надмірна експресія як модель підвищеної чутливості до змін у винагороді (13, 19, 20) та поведінкова чутливість після відмови від дієти. Ми висловлювали гіпотезу, що базальна дисрегуляція дофамінової сигналізації у мишей ΔFosB буде нормалізована корисними ефектами ВЧ дієти, охоплюючи перетин сигналів енергетичного балансу та дофамінової системи.
Для дослідження маркерів, що вказують на порушення регуляції дофамінової сигналізації в NAc, ми дослідили рівні D1-рецепторів та ефектори нижче за течією. Хоча не було суттєвих відмінностей у зв'язуванні D1 рецепторів, спостерігалася тенденція впливу HF на збільшення площі зв’язування у мишей ΔFosB. Це цікаво, оскільки індукція ΔFosB лікарськими засобами, а природні винагороди переважають у динорфіно-позитивному підтипі середніх колючих нейронів, які в першу чергу експресують D1 рецептори (9, 21). У мишей ΔFosB значно знижувались рівні мішені pCREB сигналізації дофаміну, що підтримує знижену активацію D1-рецепторів у цій області мозку (22, 23). Цікаво, що ми також виявили значне зниження загального рівня CREB у мишей ΔFosB, що дозволяє припустити подальше зниження здатності до передачі дофамінового сигналу, яке може бути вторинним для зворотного зв’язку внаслідок тривалого зниження pCREB (24). Експресія BDNF регулюється pCREB, підвищується при активації D1 і є важливим посередником нейропластичності, пов'язаної з нагородою, в NAc (25, 26). Відповідно, ми виявили значне зниження білка BDNF у NAc мишей ΔFosB.
Всі середні колючі нейрони в NAc експресують DARPP-32 (27). Її численні ефективники в нижній течії роблять його життєво важливим гравцем у шляху винагороди (28), і це стосується наркоманії та інших розладів, пов’язаних із системою дофаміну, включаючи афективні розлади та шизофренію. (27, 29). Ми виявили глибокі базальні скорочення експресії DARPP-32 в NAc мишей ΔFosB. Експресія DARPP-32 регулюється BDNF, і тому зменшена експресія може бути безпосередньо пов'язана зі зменшенням рівнів BDNF, виявлених у мишей ΔFosB (27, 29, 30). Навіть помірні зміни стану фосфорилювання DARPP-32 можуть призвести до значних змін у внутрішньоклітинній сигналізації в межах NAc (27). Попередні дослідження не повідомили про зміну білка DARPP-32 у мишей ΔFosB після видалення 12-wk з доксицикліну, коли проводилася більш широка смугаста оцінка (31), припускаючи, що вплив ΔFosB на DARPP-32 може бути залежним від часу та регіону.
Ми припускали, що різке зниження показників дофамінової сигналізації у NAc мишей ΔFosB, ймовірно, пов'язане із змінами нейронів проекції дофаміну VTA, навіть якщо ΔFosB не надмірно виражений у цих нейронах. Тому ми вивчали експресію дофамінових генів у ВТА, включаючи тирозин гідроксилазу та транспортер допаміну. Рівні тирозин гідроксилази та транспортера дофаміну позитивно корелюють із виходом дофаміну. Існувала тенденція у мишей ΔFosB виявляти знижену тирозин гідроксилазу та значне зниження транспортера дофаміну, відповідно до порушення регуляції дофамінової сигналізації в NAc. Оскільки ці базальні скорочення генів, пов'язаних з дофаміном, у VTA мишей ΔFosB, імовірно, відображають змінений зворотній зв'язок з NAc під час тривалої надмірної експресії ΔFosB, ми розглянули експресію рецептора BDNF, TrkB, як можливий механізм зворотного зв’язку NAc до VTA (32). Подібно до тирозин гідроксилази та транспортера дофаміну, експресія TrkB також показала тенденцію до зниження в основному у мишей ΔFosB, які не досягли значущості при корекції для багаторазового порівняння. Комплекс BDNF-TrkB можна ретроградно транспортувати і діяти в межах VTA, щоб впливати на місцеву експресію генів та сприяти росту та підтримці клітин (33). Крім того, активація BDNF пресинаптичного TrkB в межах NAc може безпосередньо стимулювати нейромедіацію дофаміну (32), підтримуючи основне зменшення дофамінової сигналізації у цих мишей.
Активація динорфіну к-опіоїдних рецепторів регулює дофамінову сигналізацію і є ще одним механізмом, завдяки якому NAc забезпечує зворотний зв'язок з VTA (34). Ми виявили, що експресія κ-опіоїдних рецепторів у VTA демонструвала тенденцію до зниження у мишей ΔFosB. Оскільки було показано, що перенапруження ΔFosB знижує експресію динорфіну в NAc (20), миші ΔFosB, ймовірно, мають глибоке зниження чистих VTA κ-активація опіоїдів. Хоча сигналізація про динорфін зазвичай справляє інгібуючу дію на нейрони дофаміну (35), щури, які демонструють посилене самовведення наркотиків, що вживають наркотики, демонструють знижений рівень динорфіну в NAc, вказуючи на роль зниженого сигналу динорфіну в підвищенні чутливості до винагороди (36). , 37). Дисрегуляція системи динорфін - κ-опіоїдів була пов’язана із придбанням та постійним зловживанням наркотиками, підтримуючи критичний баланс сигналів опіоїдів при нормалізації дофамінових шляхів. (38).
Виходячи з корисної здатності енергетично щільної дієти HF, ми припустили, що порушення регуляції дофамінової та опіоїдної сигналізації нагород у мишей ΔFosB схилятиме цих мишей до посилених реакцій нагородження такою дієтою, тим самим нормалізуючи систему винагороди за допомогою активації гіпоталамуса -VTA-NAc ланцюг. Протягом шести тижневого впливу дієти не спостерігалось відмінностей у прийомі їжі між ΔFosB та контрольними мишами, що дозволяє припустити, що зміни, виявлені в біохімічних та молекулярних маркерах нагородного сигналу у мишей ΔFosB, не були обумовлені різницею в споживаних калоріях. Як і передбачалося, базальні відмінності, виявлені в pCREB, загальному рівні CREB, BDNF, DARPP-32 та κ-опіоїдних рецепторах між ΔFosB та контрольними мишами, були ослаблені, ймовірно, через збільшення виходу дофаміну у мишей ΔFosB на HF (29, 39-41) .
Дослідження як тирозин гідроксилази, так і транспортера дофаміну в VTA виявило дивовижні протилежні реакції ΔFosB та контрольних мишей після HF. Контрольні миші показали зменшення експресії тирозин-гідроксилази та дофаміну, тоді як миші ΔFosB виявляли посилену експресію обох цих генів, пов’язаних з дофаміном. Цікаво, що експресія тирозин гідроксилази змінюється у ВТА шляхом хронічного введення кокаїну чи метамфетаміну (42-44), що дозволяє припустити, що миші ΔFosB можуть знайти природну винагороду HF більш помітною, ніж контрольні миші.
Для того, щоб вивчити, яким потенційним гіпоталамічним входом до ВТА можуть бути передані сигнали, що відображають енергетичний баланс, експресію рецептора лептину та рецептора орексину-1 також досліджували. Рівень циркулюючого лептину збільшується на ВЧ, і лептин може, в свою чергу, діяти на VTA, щоб змінити дофамінову сигналізацію (18, 45). Експресія рецепторів лептину VTA аналогічно знижувалась HF як у ΔFosB, так і у контрольних мишей, у відповідності з аналогічним збільшенням ваги та споживаним раціоном під час ВЧ. Також високий вміст жиру збільшував експресію рецептора орексину-1 у VTA обох ΔFosB та контрольних мишей. Орексин активує дофамінові нейрони в VTA, сприяє пластичності VTA та підвищує рівень дофаміну в NAc (46-48). Показано, що дієта з високим вмістом жиру збільшує експресію орексину у мишей відповідно до наших спостережень (49, 50). Таким чином, посилена експресія рецептора орексину, а також зміни сигналу лептину в VTA можуть сприяти нагородженню дієти як у ΔFosB, так і у контрольних мишей, підтримуючи дисоціацію між шляхами, що передають сигнали енергетичного балансу, та тими, які пов'язані безпосередньо з винагородою.
Для дослідження стресогенних наслідків зняття винагороди мишей досліджували у тесті на відкритому полі 24 год після видалення HF. Миші ΔFosB були більш чутливими до гострих ефектів кращого відміни дієти, демонструючи підвищену активність збудження та вироблення фекальних болів на новому відкритому арені порівняно з усіма іншими групами контролю та дієти. Миші ΔFosB також показали цікаву поведінкову схему в цьому тесті, що свідчить про винагороду та чутливість до стресу, при цьому HF дієта спочатку зменшила вироблення фекальних болів відносно чау, а виведення знову посилило цю реакцію, пов’язану з тривожністю. Це спостережуване збільшення активності у відкритому полі не корелювало із змінами експресії орексину, що свідчить про відношення до збудженого стресом збудження, яке не є лише впливом змін сигналізації, опосередкованої орексином. В цілому ці дані підтверджують нашу гіпотезу про те, що миші ΔFosB були б чутливішими до гострих наслідків переважного відмови від дієти завдяки підвищеній чутливості до нагородження.
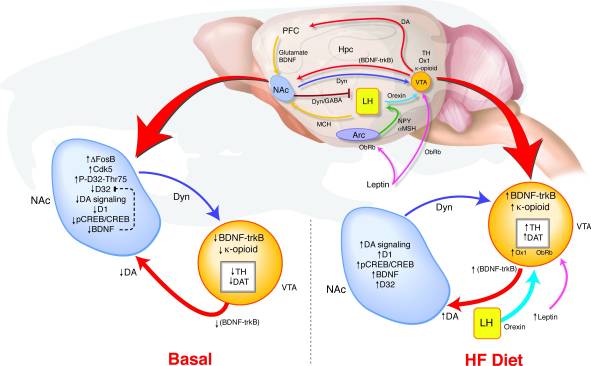
Як довготривала переекспресія ΔFosB в NAc призводить до таких змін у поведінці та нагородній сигналізації? Ми запропонували модель виявлення збігу ВТА, де змінений зворотний зв'язок від NAc та гіпоталамуса ретранслює сигнали щодо стану винагороди для визначення регуляції дофамінової системи, яка може підтримувати зв’язок між порушенням регуляції шляху винагороди та схильністю до ожиріння (рис. 8). Під час ВЧ-експозиції на VTA сходяться багато входів, що відображають як енергетичний баланс, так і стан винагороди. Збільшення сигналу лептину та орексину, а також змінений зворотний зв'язок від NAc до бічного гіпоталамусу може впливати на те, як ці орексигенні сигнали реагують на HF у мишей ΔFosB (17, 18, 45, 47, 51-53). Підвищена дієта з високим вмістом жиру в BDNF може забезпечити зворотний зв'язок з VTA, додатково сприяючи змінам експресії генів, пов’язаних з дофаміном.
малюнок 8
Дієта з високим вмістом жиру (HF) нормалізує дисрегульовану сигналізацію нагородження у мишей ΔFosB
Ці результати окреслюють молекулярні маркери чутливості до винагороди та вказують на те, що тривала дисрегуляція дофамінової системи може схилити людину до звикання та ожиріння. Крім того, ці дані забезпечують важливий крок до визначення конкретних нових терапевтичних цілей у лікуванні та профілактиці ожиріння та інших порушень, які можуть зосереджуватися на системі винагород. Надалі буде важливо дослідити, як ця система реагує на зняття раціону з ВЧ, а також дослідити будь-які статеві відмінності у чутливості до винагороди та дієті з високим вмістом жиру.
Додатковий матеріал
Суп. Методи
Натисніть тут, щоб переглянути. (61K, док.)
Подяки
Автори хочуть подякувати Кеті Штеффен за допомогу у розведенні та передачі тварин. Ця робота була підтримана грантом від Університету Пенсільванії діабетичного центру (DK019525) та грантами Національного інституту психічного здоров’я (R01 MH51399 та P50 MH66172) та Національного інституту зловживання наркотиками (R01 DA07359).
Виноски
Фінансові розкриття інформації: Усі автори заявляють, що у них немає біомедичних фінансових інтересів або потенційного конфлікту інтересів.
посилання
1. Wadden TA, Berkowitz RI, LG Womble, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Kaplan R, Stunkard AJ. Довільне випробування модифікації способу життя та фармакотерапії проти ожиріння. N Engl J Med. 2005; 353 (20): 2111 – 20. [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Знижена нікотинова нагорода при ожирінні: перехресне порівняння у людини та миші. Психофармакологія (Берл) 2005
3. Franken IH, Muris P. Індивідуальні відмінності у чутливості до винагороди пов'язані з тягою до їжі та відносною масою тіла у здорових жінок. Апетит. 2005; 45 (2): 198 – 201. [PubMed]
4. Kelley AE, Berridge KC. Нейрологія природних нагород: відповідність наркотичним засобам. J Neurosci. 2002; 22 (9): 3306 – 11. [PubMed]
5. Cagniard B, Balsam PD, Brunner D, Zhuang X. Миші з хронічно підвищеним дофаміном виявляють посилену мотивацію, але не навчаючись, за харчову винагороду. Нейропсихофармакологія. 2006; 31 (7): 1362 – 70. [PubMed]
6. Liang NC, Hajnal A, Norgren R. Sham, що харчується кукурудзяною олією, збільшує кількість дофаміну у щурів. Am J Physiol Regul Integr Comp Physiol. 2006; 291 (5): R1236 – 9. [PubMed]
7. Mendoza J, Angeles-Castellanos M, Escobar C. Захоплення приємним прийомом їжі викликає активність, що передбачає їжу, та експресію c-Fos у ділянках мозку, пов'язаних з нагородами. Неврознавство. 2005; 133 (1): 293 – 303. [PubMed]
8. Schroeder BE, Binzak JM, Kelley AE. Загальний профіль префронтальної кортикальної активації після впливу нікотинових або шоколадних контекстних сигналів. Неврознавство. 2001; 105 (3): 535 – 45. [PubMed]
9. Nestler EJ, Barrot M, Self DW. DeltaFosB: постійний молекулярний комутатор для залежності. Proc Natl Acad Sci США A. 2001; 98 (20): 11042 – 6. [Безкоштовна стаття PMC] [PubMed]
10. Teegarden SL, Bale TL. Зниження дієтичних переваг викликає підвищену емоційність та ризик рецидиву харчування. Психіатрія біолів. 2007; 61 (9): 1021 – 9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR. dFosB в накопиченні ядра регулює харчову активізацію поведінки та мотивацію. Журнал нейрознавства. 2006; 26 (36): 9196 – 9204. [PubMed]
12. Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ. Трансгенні тварини з індукованою, цільовою експресією генів у мозку. Mol Pharmacol. 1998; 54 (3): 495 – 503. [PubMed]
13. Kelz MB, Chen J, Carlezon WA, Jr, Whisler K, Gilden L, Beckmann AM, Steffen C, Zhang YJ, Marotti L, Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Duman RS, Picciotto MR, Nestler EJ. Експресія фактора транскрипції deltaFosB в мозку контролює чутливість до кокаїну. Природа. 1999; 401 (6750): 272 – 6. [PubMed]
14. Бейл TL, Дорса ДМ. Статеві відмінності та ефекти естрогену на експресію рибонуклеїнової кислоти рецептора окситоцину в Вентромедіальному гіпоталамусі. Ендокринологія 1995; 136 (1): 27 – 32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. Цукрозалежні щури виявляють посилене реагування на цукор після утримання: свідчення ефекту позбавлення цукру. Фізіол Бехав. 2005; 84 (3): 359 – 62. [PubMed]
16. Will MJ, Franzblau EB, Kelley AE. Му-опіоїди нуклеусів регулюють прийом дієти з високим вмістом жиру шляхом активації розподіленої мозкової мережі. J Neurosci. 2003; 23 (7): 2882 – 8. [PubMed]
17. Чжен Х, Паттерсон Л.М., Бертуд HR. Сигналізація орексину в вентральній тегментальній області необхідна для апетиту з високим вмістом жиру, викликаного опіоїдною стимуляцією ядра ядра. J Neurosci. 2007; 27 (41): 11075 – 82. [PubMed]
18. Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, Gao XB, Thurmon JJ, Marinelli M, DiLeone RJ. Сигналізація рецепторів лептину в нейронах середнього мозку дофаміну регулює годування. Нейрон. 2006; 51 (6): 801 – 10. [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. Перенапруження DeltaFosB, характерне для типу стриатальних клітин, посилює стимул кокаїну. J Neurosci. 2003; 23 (6): 2488 – 93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. Важлива роль DeltaFosB в ядрі, що накопичується в дії морфіну. Nat Neurosci. 2006; 9 (2): 205 – 11. [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. Формування дендриту хребта, спричинене кокаїном, у середніх колючих нейронах, що містять рецептори D1 та D2, містять рецептори дофаміну. Proc Natl Acad Sci США A. 2006; 103 (9): 3399 – 404. [Безкоштовна стаття PMC] [PubMed]
22. Бленді Дж. А., Мальдонадо Р. Генетичний аналіз наркоманії: роль білка, що зв'язує цАМФ. J Mol Med. 1998; 76 (2): 104 – 10. [PubMed]
23. Nestler EJ. Молекулярні механізми наркоманії. Нейрофармакологія. 2004; 47 1: 24 – 32. [PubMed]
24. Tanis KQ, Duman RS, Newton SS. Зв’язування та активність CREB в головному мозку: регіональна специфіка та індукція електроконвульсивним припадком. Психіатрія біолів. 2007
25. Kumar A, Choi KH, Renthal W, Цанкова Н.М., Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ. Ремоделювання хроматином є ключовим механізмом, що лежить в основі пластичності, викликаної кокаїном, у стриатумі. Нейрон. 2005; 48 (2): 303 – 14. [PubMed]
26. Graham DL, Edwards S, Bachtell RK, Dileone RJ, Rios M, Self DW. Динамічна активність BDNF в ядрах, що споживають кокаїн, збільшує самовведення та рецидив. Nat Neurosci. 2007; 10 (8): 1029 – 37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 опосередковує дії численних зловживань наркотиками. Aaps J. 2005; 7 (2): E353 – 60. [Безкоштовна стаття PMC] [PubMed]
28. Palmer AA, Verbitsky M, Suresh R, Kamens HM, Reed CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, Phillips TJ. Різниці в експресії генів у мишей, що підбираються з урахуванням чутливості до метамфетаміну. Геном мами 2005; 16 (5): 291 – 305. [PubMed]
29. Богуш А, Педріні S, Пельта-Хеллер Дж, Чан Т, Ян Q, Мао Z, Слузас Е, Гірінгер Т, Ерліх МЕ. AKT і CDK5 / p35 опосередковують індукцію нейтротрофічного фактора мозку DARPP-32 у колючих нейронах середнього розміру in vitro. J Biol Chem. 2007; 282 (10): 7352 – 9. [PubMed]
30. Benavides DR, Bibb JA. Роль Cdk5 у зловживанні наркотиками та пластичності. Енн Нью-Йорк Акад. Наук. 2004; 1025: 335 – 44. [PubMed]
31. Bibb JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Ефекти хронічного впливу кокаїну регулюються протеїном нейрона Cdk5. Природа. 2001; 410 (6826): 376 – 80. [PubMed]
32. Blochl A, Sirrenberg C. Нейротрофіни стимулюють вивільнення дофаміну з мезенцефальних нейронів щурів через рецептори Trk та p75Lntr. J Biol Chem. 1996; 271 (35): 21100 – 7. [PubMed]
33. Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Цанкова Н.М., Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. Важлива роль BDNF у мезолімбічному дофаміновому шляху при стресі соціального ураження. Наука. 2006; 311 (5762): 864 – 8. [PubMed]
34. Nestler EJ, Carlezon WA., Jr. Мезолімбічна схема нагородження дофаміну при депресії. Психіатрія біолів. 2006; 59 (12): 1151 – 9. [PubMed]
35. Ford CP, Beckstead MJ, Williams JT. Інгібування опіоїдних капп соматодендритних дофамінових інгібіторів постсинаптичних струмів. J Нейрофізіол. 2007; 97 (1): 883 – 91. [PubMed]
36. Найландер I, Власковська М, Тереніус Л. Мозкові динорфінові та енкефалінові системи у щурів Фішера та Льюїса: ефекти толерантності до морфіну та виведення. Мозок Рез. 1995; 683 (1): 25 – 35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Відмінність між щурами, що віддають перевагу алкоголю (AA) та щурам, які уникають алкоголю (ANA), у системах продинорфіну та проенкефліну. Клініка алкоголю Exp Res. 1994; 18 (5): 1272 – 9. [PubMed]
38. Крик MJ. Кокаїн, дофамін та ендогенна опіоїдна система. J Наркоман Діс. 1996; 15 (4): 73 – 96. [PubMed]
39. Carlezon WA, Jr, Duman RS, Nestler EJ. Багато облич CREB. Тенденції Neurosci. 2005; 28 (8): 436 – 45. [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Taher M, Barczak A, Kameyama K, Huganir R, Konradi C. Дофамінові D1 рецептори опосередковують фосфорилювання CREB за допомогою фосфорилювання NMDA-рецептора на Ser897-NR1. J Neurochem. 2003; 87 (4): 922 – 34. [PubMed]
41. Самостійний DW. Регулювання прийому наркотиків та їх поведінки шляхом нейроадаптації в мезолімбічній дофаміновій системі. Нейрофармакологія. 2004; 47 1: 242 – 55. [PubMed]
42. Beitner-Johnson D, Nestler EJ. Морфін та кокаїн чинять загальні хронічні дії на тирозин-гідроксилазу в дофамінергічних регіонах мозку. J Neurochem. 1991; 57 (1): 344 – 7. [PubMed]
43. Lu L, Grimm JW, Shaham Y, Hope BT. Молекулярні нейроадаптації в околицях та вентральній тегментальній області протягом перших днів 90 примусового утримання від кокаїну при самостійному введенні щурів. J Neurochem. 2003; 85 (6): 1604 – 13. [PubMed]
44. Шепард Ж.Д., Чуанг Д.Т., Шахам Ю., Моралес М. Вплив самовведення метамфетаміну на рівень тирозин гідроксилази та допаміну в мезолімбічному та нігростриальному дофамінових шляхах щурів. Психофармакологія (Berl) 2006; 185 (4): 505 – 13. [PubMed]
45. Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, Pothos EN, Maratos-Flier E, Flier JS. Регулювання лептину дофамінового шляху мезоаккуменів. Нейрон. 2006; 51 (6): 811 – 22. [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Implication of протеїнкіназа C у підвищенні орексину підвищення рівня позаклітинного дофаміну та його корисний ефект. Eur J Neurosci. 2007; 25 (5): 1537 – 45. [PubMed]
47. Наріта М, Нагумо Y, Хашимото S, Хотиб Дж, Міятаке М, Сакурай Т, Янагісава М, Накамачі Т, Шиода С, Сузукі Т. Пряме залучення орексинергічних систем в активацію мезолімбічного дофамінового шляху та пов'язані з ними поведінки, індуковані морфіном. J Neurosci. 2006; 26 (2): 398 – 405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, Fields HL, Bonci A. Orexin A у VTA є критично важливим для індукції синаптичної пластичності та сенсибілізації поведінки до кокаїну. Нейрон. 2006; 49 (4): 589 – 601. [PubMed]
49. Park ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS. Зміни в експресії орексину-А та нейропептиду Y у гіпоталамусі голодуючих щурів, які годувались голодним та високим вмістом жиру. J Vet Sci. 2004; 5 (4): 295 – 302. [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF. Пептиди, які регулюють споживання їжі: експресія гена орексину підсилюється під час гіпертригліцеридемії. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1454 – 65. [PubMed]
51. Чжен Н, Коркерн М, Стоянова I, Паттерсон Л.М., Тіан Р, Бертуд HR. Пептиди, які регулюють прийом їжі: маніпуляція, що викликає апетит, активізує нейрони гіпоталамічного орексину та пригнічує нейрони POMC. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1436 – 44. [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, Daniel RA, Landry CF, Kelley AE. Активізація субпопуляції гіпоталамічних нейронів, що містять орексин / гіпокретин, за допомогою інгібування GABAA-рецептора, опосередкованого рецептором оболонки ядра, але не шляхом впливу нового середовища. Eur J Neurosci. 2004; 19 (2): 376 – 86. [PubMed]
53. Харріс Г.К., Віммер М, Астон-Джонс Г. Роль латеральних гіпоталамічних нейронів орексину у пошуку винагород. Природа. 2005; 437 (7058): 556 – 9. [PubMed]