 Фабріціо Санна1*,
Фабріціо Санна1*,  Джессіка Братзу1,
Джессіка Братзу1,  Марія А. Пілуду2,
Марія А. Пілуду2,  Марія Г. Корда2,
Марія Г. Корда2,  Марія Р. Меліс1,
Марія Р. Меліс1,  Освальдо Джорджі2 та
Освальдо Джорджі2 та  Антоніо Арджолас1,3
Антоніо Арджолас1,3- 1Кафедра біомедичних наук, відділ нейронауки та клінічної фармакології, Центр передового досвіду нейробіології залежностей, Університет Кальярі, Кальярі, Італія
- 2Кафедра медико-біологічних наук, відділ фармацевтичних, фармакологічних і медичних наук, Університет Кальярі, Кальярі, Італія \ t
- 3Інститут неврології, Національна дослідницька рада, відділ Кальярі, Cittadella Universitaria, Кальярі, Італія
Риби високого (RHA) і низького уникнення (RLA) безпородних щурів, які відрізняються відповідно швидким або поганим придбанням активної реакції уникнення в човникові, показують відмінності в сексуальній активності при положенні в присутності сексуально сприйнятливі жіночі щури. Дійсно, щури RHA показують більш високий рівень сексуальної мотивації та копуляторної продуктивності, ніж щури RLA, які зберігаються також після повторної сексуальної активності. Ці відмінності корелювали з більш високим тоном мезолімбічної допамінергічної системи щурів RHA проти щурів RLA, що виявлялося більш високим збільшенням дофаміну, знайденого в диализате, отриманому з ядра акушерів RHA, ніж щурів RLA під час статевої активності. Дана робота показує, що позаклітинний дофамін і норадреналін (NA) також збільшують діалізат з медіальної префронтальної кори (mPFC) самців щурів RHA і RLA, які знаходяться в присутності недоступної жіночої щури і більш помітно під час безпосередньої статевої взаємодії. Таке збільшення дофаміну (та його основного метаболіту 3,4-дигідроксифенилуксусной кислоти, DOPAC) та NA було виявлено як у статевозрілих, так і у досвідчених тварин, але вони були вищими: (i) у RHA, ніж у щурів RLA; і (ii) у щурів, що мають досвід сексуально RHA і RLA, ніж у їхніх наївних аналогів. Нарешті, відмінності в дофаміні і НС у mPFC відбувалися одночасно з тими, які мали сексуальну активність, оскільки щури RHA проявляли більш високий рівень сексуальної мотивації та копуляторної продуктивності, ніж щури RLA як в статевих, так і в досвідчених умовах. Ці результати дозволяють припустити, що більш високий дофамінергічний тонус також виникає в mPFC, разом з підвищеним норадренергічним тоном, який може бути залучений в різні копуляторні структури, знайдені в щурах RHA і RLA, як було запропоновано для мезолімбічної допамінергічної системи.
Вступ
Римські лінії високої (RHA) і низької уникнення (RLA) безпородні лінії щурів, спочатку вибрані для відповідно швидкого або поганого отримання активної реакції уникнення в човниковій коробці (Bignami, 1965; Бродхерст і Бігнамі, 1965; Driscoll і Bättig, 1982; Fernández-Teruel et al., 2002; Giorgi et al., 2007) виявляють значні відмінності в сексуальній активності при введенні в присутності статево сприйнятливого жіночого щура. Дійсно, щури RHA показують більш високі рівні сексуальної мотивації, ніж щури RLA, що виявляється за рахунок більшої кількості неконтрактних ерекцій пеніса, показаних цими щурами, при наявності недоступної сприйнятливої жінки і кращих копуляторних показників, ніж виявлено у RLA щурів. значні відмінності в численних копуляторних параметрах виявлені, головним чином, у першому копуляторному тесті між двома лініями щурів, але також після п'яти копуляторних тестів, хоча й ослаблені (Sanna et al., 2014a). Серед найбільш виражених відмінностей, виявлених між щурами RHA і RLA, є відсоток щурів, які ініціюють монтаж і інтромітінг, і що еякулюють у першому копуляторному тесті, 80% щурів RHA проти 40% щурів RLA, відповідно (Sanna et al., 2014a). Велика кількість експериментальних даних свідчить про те, що функціонально різні дофамінергічні тони беруть участь у відмінностях копуляції між щурами RHA і RLA. Фактично, різні копуляторні структури щурів RHA та RLA виявилися різним чином модифіковані апоморфіном, змішаним агоністом дофамінових рецепторів D1 / D2, що вводяться в дозах, що полегшують сексуальну поведінку, і галоперидолом, антагоністом дофамінових рецепторів D2 низькі дози, які пригнічують сексуальну поведінку (Sanna et al., 2014b). Відповідно, було виявлено, що щури RLA є більш чутливими до полегшення та інгібування статевої поведінки, індукованої апоморфіном і галоперидолом, відповідно, про що свідчать більші модифікації, викликані двома препаратами, головним чином, при введенні в менших дозах при різних копуляторних параметрах. Щури RLA щодо щурів RHA (\ tSanna et al., 2014b). Можливо, більш важливим для даного дослідження є різні копуляторні структури щурів RHA і RLA, також пов'язані з відмінностями в активності мезолімбічних допамінергічних нейронів, активність яких добре відома під час сексуальної поведінки (Pfaus et al., 1990; Pleim et al., 1990; Pfaus і Phillips, 1991; Damsma et al., 1992; Wenkstern et al., 1993; Balfour et al., 2004; Pitchers et al., 2010, 2013; Beloate et al., 2016). Дійсно, незважаючи на те, що щури RHA та RLA, котрі мають сексуальну наївність і статеві контакти, мають подібні базальні значення позаклітинного допаміну в діалізаті від nucleus accumbens, концентрації позаклітинного дофаміну і 3,4-дигідроксифенилуксусной кислоти (DOPAC, один з його основних метаболітів). ), виявлено, що вони дифференциально збільшуються в диализатах, отриманих з оболонки nucleus accumbens наївних і статево досвідчених щурів RHA і RLA як у передчасному, так і в кінцевому періодах сексуальної поведінки. Перераховані вище відмінності були більш вираженими між сексуальними неінфекційними щурами RHA і RLA, але зберігалися між статево досвідченими RHA і RLA щурами, хоча вони мали тенденцію до зменшення у цих щурів, як було виявлено з відмінностями в сексуальній поведінці (Sanna et al., 2015).
Функціональна роль підвищення дофамінергічної активності в ядрі accumbens, що спостерігається під час сексуальної активності (або в апетитній, і в конмативній фазах сексуальної поведінки), все ще є предметом дискусій, а також тим, що спостерігається під час годування, що, як і сексуальна активність, має сильну активність. мотиваційна валентність. Таким чином, хоча дофамін в nucleus accumbens бере участь у мотивації, а мезолімбічні допамінергічні нейрони зазвичай називаються корисними нейронами, в останніх 15 років останні дослідження підтверджують гіпотезу, що мезолімбічний допамін не залучений до первинного вираження мотивованої або корисної поведінки, скоріше з вивченням та пам'яттю асоціацій стимулів-винагород (Agmo et al., 1995; Berridge і Robinson, 1998; Ikemoto та Panksepp, 1999; Pitchers et al., 2013, 2014; Beloate et al., 2016; Salamone et al., 2016). Відповідно до цієї гіпотези, блокада дофамінових рецепторів в ядрі accumbens або інактивація дофамінергічних нейронів в вентральній тегментальній області нещодавно виявилися неспроможними змінити експресію апетитних і консервативних аспектів копуляторної поведінки у самців щурів (Pitchers et al., 2013, 2014; Beloate et al., 2016).
Однак неможливо виключити роль інших областей мозку, що містять дофамін у вищезгаданих відмінностях в сексуальній поведінці між щурами RHA і RLA. Відповідно, добре відомо, що дофамін надає полегшуючий вплив на попередні та консервативні фази сексуальної поведінки у лабораторних тварин, а також на людях не тільки в nucleus accumbens (Everitt, 1990; Pfaus et al., 1990; Hull et al., 1991; Pfaus і Everitt, 1995; Меліс і Арджолас, 2011), але також і в інших областях головного мозку, таких як медіальний преоптичний ділянку, гіпоталамус і його ядра (тобто паравентрикулярне ядро (PVN); Pfaus і Phillips, 1991; Argiolas і Melis, 1995, 2005, 2013; Hull et al., 1995, 1999; Меліс і Арджолас, 1995; Melis et al., 2003; Succu et al., 2007; Pfaus, 2010). Ще однією областю, яка містить допамін і може відігравати певну роль у сексуальній поведінці, є медіальна префронтальна кора (mPFC; Fernández-Guasti et al., 1994; Agmo і Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama et al., 2003; Balfour et al., 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011). Насправді, в цій області мозку виявляються нервові закінчення мезокортикальних нейронів дофаміну, їхні клітинні тіла локалізовані в вентральній тегментальной області як мезолімбічні дофамінові нейрони. Що стосується nucleus accumbens, точна роль цієї ділянки мозку в сексуальній поведінці далека від ясного. Дійсно, ураження mPFC зазвичай виявляються нездатними змінювати сексуальну поведінку щурів-самців статево сприйнятливими жінками (Fernández-Guasti et al., 1994; Agmo і Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama et al., 2003; Balfour et al., 2006; Afonso et al., 2007), а також вираження умовного місцеположення для сексуальної винагороди (Davis et al., 2010). Однак поразки mPFC, які не змінювали появу умовного переваги місця для сексуальної винагороди, скасували у тих самих тварин здатність формувати обумовлену неприйняття до сексуальної активності в парі з аверсивними стимулами (Davis et al., 2010) і вибіркове випалювання клітин під час наближення поведінки самців щура до недоступної статево сприйнятливої жінки вимірювали в mPFC самців щурів (Febo, 2011). Ці результати показали, що активація mPFC під час сексуальної поведінки відіграє певну роль у інтеграції зовнішньої та внутрішньої інформації для виконання та контролю цілеспрямованої поведінки, а не у вираженні вроджених відповідей на природні підсилювачі (див. Goto і Grace, 2005). Відповідно, разом з ядром accumbens, mPFC є частиною складної нейронної системи, що бере участь у модуляції мотивованої поведінки (цілеспрямованої поведінки), яка вимагає інтеграції когнітивної інформації від mPFC, емоційної інформації з мигдалини та контексту- відповідну інформацію з гіпокампу, в nucleus accumbens (Goto і Grace, 2005).
Експериментальні дані свідчать про те, що вивільнення дофаміну в mPFC бере участь у адаптивному регулюванні мотивованої поведінки, і вважається, що дерегуляція цих механізмів відіграє певну роль у патологічних або дезадаптивних станах, таких як психічні розлади як шизофренія, дефіцит уваги і гіперактивні розлади ( СДУГ), депресія (Dunlop і Nemeroff, 2007; Masana et al., 2011), або поведінка зловживання та азартних ігор (Everitt і Robbins, 2005). Цікаво, що на активність дофаміну, що вивільняється в mPFC, може впливати норадреналін (NA), який присутній в mPFC на більш високих рівнях, ніж дофамін, і, зокрема, транспортером NA (NET), який не тільки більш багатий, ніж транспортер допаміну (DAT) у mPFC (Carboni et al., 1990, 2006; Gresch et al., 1995; Westernik et al., 1998), але також показує спорідненість до допаміну навіть вище, ніж для NA (Хорн, 1973). Оскільки допамін і НС часто співпрацюють у багатьох функціях mPFC, від формування робочої пам'яті та набору уваги та переходу до оберненого навчання, інгібування відповіді та відповіді на стрес (див. Роббінс і Арнстен, 2009), це підвищує ймовірність того, що дофамін, окремо або разом з NA, в mPFC може також відігравати роль у поведінкових відмінностях між RHA і RLA щурами, включаючи ті, які виявляються при сексуальній поведінці.
Для того, щоб перевірити цю гіпотезу, активність мезокортикальної допамінергічної системи і норадренергічної системи в mPFC досліджували в двох лініях щурів RHA і RLA за допомогою внутрішньомозкового мікродіалізу. Коротко, дофамін (і його основний метаболіт DOPAC) і NA вимірювали в диализатах, отриманих з прелімбічних (PrL) і інфралімбових (IL) компартментів mPFC статевозрілої (наприклад, ніколи не піддавалися рецептивної жінки) і статево досвідчених RHA і Щури RLA (наприклад, які пройшли п'ять попередніх тестів копуляції і демонструють постійний рівень копуляторної активності) при введенні в присутності недоступної сприйнятливої жінки і під час безпосередньої статевої взаємодії рідинною хроматографією високого тиску в поєднанні з електрохімічним детектуванням (HPLC-ECD).
Матеріали та методи
Звірята
Самородні щури RHA і RLA (N = 30 для кожної лінії, вагою ≈300 g на початку експериментальної роботи) були всі з колонії, заснованої в 1998 в Університеті Кальярі, Італія (Giorgi et al., 2007). Процедури, що використовуються для селективного розмноження сардинської колонії, вже детально описані (Giorgi et al., 2005).
Овариектомированние стимулюючі SD-щури (250-300 g на початку експериментальної роботи), використані у всіх експериментах, були отримані від Envigo (San Pietro al Natisone, Італія). Тварини акліматизувалися чотири на клітку (38 см × 60 см × 20 см) до житлових приміщень кафедри біомедичних наук Університету Кальярі протягом мінімум 10 днів до початку експериментів, при 24 ° C, вологість 60%, зворотний цикл 12 h світло / темрява (вимикається світло з 08: 00 h до 20: 00 h), вода і стандартне лабораторне харчування ad libitum. Упродовж всього періоду звикання тварини щодня оброблялися для 1 – 2 хв, щоб обмежити стрес під час експериментів; крім того, контакт з обслуговуючим персоналом у будинку для тварин був обмежений однією особою, а постільна білизна в домашніх клітинах ніколи не змінювалася ні напередодні, ні на день експериментів. Всі експерименти проводили між 10: 00 h і 18: 00 h. Це дослідження проводилося відповідно до рекомендацій керівних принципів Європейських Співтовариств, Директиви від вересня 22, 2010 (2010 / 63 / EU) та італійського законодавства (DL March 4, 2014, n. 26). Протокол був схвалений Етичним комітетом для експериментів на тваринах Університету Кальярі (№ авторизації 361 / 2016-PR, квітень 08, 2016 до FS).
Експериментальні групи
Використовувалися щури RHA та RLA, які не мали сексуальної активності, а також щурів, що мають статевий досвід. Щури, які не були сексуальними, були щурами, які ніколи не піддавалися сексуально-сприйнятливо-овариэктомированному і естрадіолу + прогестерону-праймерам; статево досвідчені щури були щурами, які вже пройшли п'ять послідовних копуляційних тестів 60 min з інтервалами 3 днів з рецептивною жінкою (Sanna et al., 2014a,b). Самців вносили в еструс шляхом обробки підшкірним бензоатом естрадіолу (200 мкг / щур в арахісовому маслі) і прогестерону (0.5 мг / щур в арахісовому маслі), 48 і 6 год. Еструс був підтверджений забарвленням May-Grunwald-Giemsa та мікроскопічним дослідженням вагінальних мазків 1 h перед експериментами. У згоді з попередніми дослідженнями (Sanna et al., 2014a,b, 2015) було виявлено, що п'ять попередніх копуляційних тестів є достатніми для того, щоб у щурів-римлянок обох ліній спостерігався постійний рівень копуляторної активності: наприклад, статево досвідчені щури RHA і RLA задовольняли критерію щонайменше однієї еякуляції, досягнутої в кожному з двох останніх тестів; (одна щура RHA і дві щури RLA, які не задовольняли даному критерію, були відкинуті в цій фазі). Через два дні після цих попередніх копуляторних тестів статево досвідчені римські щури піддавалися стереотаксичній операції для імплантації зонда мікродіалізу в mPFC, як описано нижче (Sanna et al., 2015).
Мікродіаліз в mPFC під час сексуального поведінки
За день до того, як мікродиаліз, щури, які не були ратифікованими статевим шляхом, або досвідчені щури RHA і RLA, були розташовані в стереотаксичному апараті (Stoelting Co., Wood Dale, IL, USA) під анестезією ізофлурана (1.5% –2%; Harvard Apparatus, Holliston, MA, USA) і імплантують вертикальним саморобним зондом мікродіалізу (діалізна мембрана ≈3 мм вільної поверхні; Melis et al., 2003), і спрямовані в односторонньому порядку в компартментах mPFC, PrL і IL (координати: 3.0 мм передня і 0.7 мм латерально до брегма, і 5.5 мм вентрально до твердої оболонки); Паксинос і Уотсон, 2004). У день експерименту тварин переносили під час темної фази циклу до спарову клітку (45 см × 30 см × 24 см), яка була розташована в звукоізоляційному приміщенні, освітленому тьмяним червоним світлом і містить всередині іншого маленька клітка з оргскла (15 см × 15 см × 15 см) з отворами 25 (Ø 2 мм) у вертикальних стінах, що дозволяють візуальні, нюхові та акустичні, але не прямі взаємодії. Після періоду звикання 2 h зонд мікродіалізу був з'єднаний з мікрофонним насосом CMA / 100 (Harvard Apparatus, Holliston, MA, USA) з поліетиленовою трубкою і перфузований розчином Рінгера (147 mM NaCl, 3 mM KCl і 1.2 mM). CaCl2, рН 6.5), при швидкості потоку 2.5 мкл / хв. Після періоду врівноваження 2 h перфузійного середовища з позаклітинною рідиною, аліквоти диализата 37.5 мкл збирали кожну 15 хв під час експерименту в охолоджених льодом поліетиленових пробірках для вимірювання концентрації допаміну, DOPAC і NA, як описано нижче. Після збору, щонайменше, чотирьох аликвот диализата, в маленьку клітку, розташовану всередині спарювальної клітини, вводили рецептивну жіночу щура за 30 хв. Протягом цих 30 хв збирали інші два аліквоти диализата. У цих умовах самці щури не можуть безпосередньо взаємодіяти з самкою, але показують неконтактні ерекції (див. Нижче). Після закінчення цього періоду невелику клітку видаляли, копуляції дозволяли протягом 75 хв, і інші п'ять аліквоти диализата були зібрані. Наприкінці цього періоду самку витягували з спарювальної клітини і збирали додаткову аликвоту диализата (Pfaus і Everitt, 1995; Melis et al., 2003; Sanna et al., 2015). Статеві параметри, пов'язані з попередньою та консервативною фазами статевої поведінки, реєструвалися протягом всього експерименту (див. Нижче).
Сексуальне поведінка
Кілька параметрів статевої мотивації та копуляторних показників, пов'язаних з попередньою та консервативною фазами сексуальної поведінки, було зафіксовано спостерігачем, який не знав конкретних експериментальних умов, наприклад, хто не знав лінії та рівень сексуального досвіду. тварин, використаних у цьому експерименті (див. нижче). Коротко, затримка до першої неконтактної ерекції (NCPEL, приурочена від введення сприйнятливої жінки у внутрішню маленьку клітку) та їх частота (NCPEF, кількість неконтактних ерекцій пеніса, які відбуваються в період, в якому знаходиться самка у внутрішній клітці). Ці феромон-опосередковані ерекції статевого члена, які виникають у статево-потужних щурів-самців у присутності недоступної сприйнятливої жінки, вважаються важливим показником сексуального збудження (Sachs et al., 1994; Sachs, 2000; Melis et al., 2003). При дозволі статевої взаємодії, наприклад, під час копуляції, латентність і інтроміт (ML і IL, приурочені від видалення внутрішньої невеликої клітини до першого монтування або першого втручання, відповідно); частота монтажу і интромитинга (MF і IF, кількість монтувань і інтромісій відповідно у першій серії копуляторної активності і протягом усього періоду копуляції); латентність до еякуляції (EL, приурочена від першого втручання першої серії до еякуляції); реєстрували частоту еякуляції (EF, загальну кількість еякуляцій під час копуляційного тесту) і пост-еякуляційний інтервал (PEI, приурочений від першої еякуляції до наступного втручання). Крім того, ефективність копуляції (CE; кількість інтромісій даного ряду, поділена на суму кількості гойдань і інтромісій у тій же серії) і інтервал міжтрансмісії (III; співвідношення між латентністю еякуляції даної серії) і кількість інтромісій у цій серії) також були розраховані для першої серії копуляторної активності (Sachs і Barfield, 1976; Мейсел і Сакс, 1994; Melis et al., 2003; Sanna et al., 2014a,b, 2015).
Визначення концентрацій дофаміну, допаку та норадреналіну в диализате з mPFC
Концентрації дофаміну, DOPAC і NA вимірювали в 20 мкл тієї ж аліквоти диализата з mPFC рідинною хроматографією високого тиску (ВЕРХ), пов'язаної з електрохімічним детектуванням з використанням подвійної комірки 4011 (Coulochem II, ESA, Cambridge, MA, USA) вже описано (Melis et al., 2003). Виявлення проводили в режимі відновлення при + 350 і -180 mV. ВЕРХ оснащували колоною Supelcosil C18 (розмір частинок 7.5 см × 3.0 мм, розмір частинок 3 мкм; Supelco, Supelchem, Milan, Італія), элюируя цитратом 0.06 М / ацетат рН 4.2, що містить метанол 20% v / v, 0.1 mM EDTA, 1 мкМ триетиламін і 0.03 мМ додецилсульфат натрію в якості рухомої фази при швидкості потоку 0.6 мл / хв і кімнатної температури. Чутливість аналізу була 0.125 pg для допаміну, 0.1 pg для DOPAC та 0.2 pg для NA.
Гістологія
Наприкінці експериментів щурів умертвляли декапітацією, мозок негайно видаляли з черепа і занурювали в 4% водний формальдегід протягом 12 – 15 днів. Після закінчення цього періоду корональні ділянки головного мозку 40 мкм готували з заморожуючим мікротомом, фарбували нейтральним червоним і перевіряли на фазово-контрастному мікроскопі. Положення кінчика зонда потім локалізувалося в mPFC, слідуючи тракту зонда через ряд ділянок мозку (див. Малюнок). 1). Для статистичної оцінки результатів розглядалися тільки щури, для яких встановлено, що вони мають активну частину діалізуючої мембрани, правильно розташовану в компартментах PrL і IL mPFC (на цій фазі одна щура від кожної експериментальної групи була відкинута).
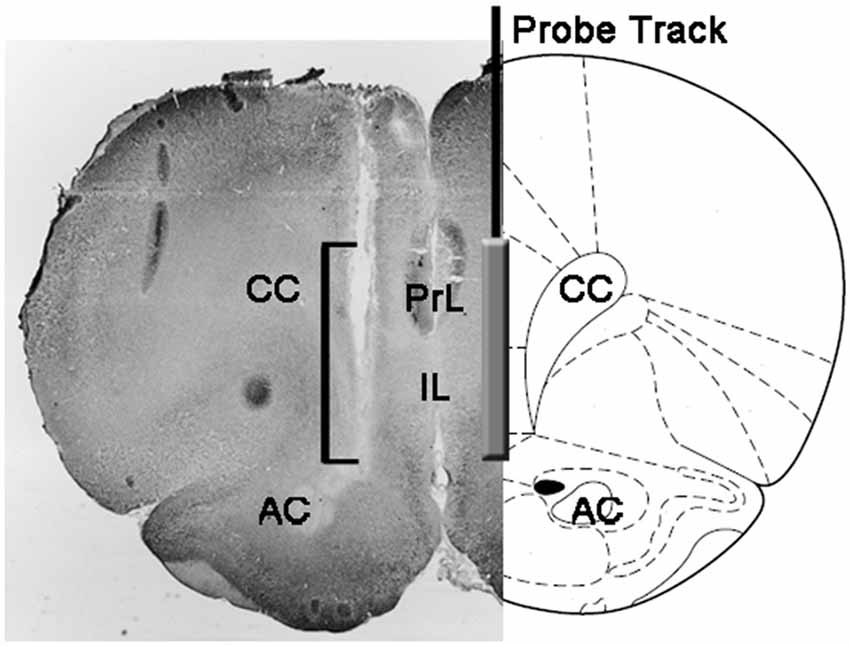 малюнок 1. Схематичне зображення коронального ділянки мозку щурів, що показує доріжку зонда мікродіалізу в PrL і IL частинах медіальної префронтальної кори (mPFC; Паксинос і Уотсон, 2004). Квадратна дужка на мікро-фотографії вказує на ділянку розділу нейтрального червоного кольору, що показує активну частину діалізуючої мембрани зонда мікродіалізу. Скорочення: PrL, попередня область; ІЛ, инфлибимная область; AC, передня комісура; CC, corpus callosum.
малюнок 1. Схематичне зображення коронального ділянки мозку щурів, що показує доріжку зонда мікродіалізу в PrL і IL частинах медіальної префронтальної кори (mPFC; Паксинос і Уотсон, 2004). Квадратна дужка на мікро-фотографії вказує на ділянку розділу нейтрального червоного кольору, що показує активну частину діалізуючої мембрани зонда мікродіалізу. Скорочення: PrL, попередня область; ІЛ, инфлибимная область; AC, передня комісура; CC, corpus callosum.
Статистика
Статистичні аналізи біохімічних (допамінів, допак та NA) та поведінкових (NCPEL та NCPEF, ML, IL та EL, MF, IF та EF, та PEI) даних були проведені або у всіх експериментальних суб'єктах (як це традиційно робилося в дослідженнях самців щурів). копуляторної поведінки) або виключаючи тих суб'єктів, які не копулировали з еякуляцією під час експерименту мікродіалізу. У першому випадку, коли всі тварини були включені в аналізи, тваринам, які не виявили NCPE протягом часу, коли жінка була недоступною або не могла або не інтромітувати або еякулювати з наявними жінками, були присвоєні відповідні повні діапазони результатів: 1800 s самець не показав NCPE в хв. хв. хв., в якому жінка була недоступна; 30 s для ML і IL, якщо самці не змонтувалися або не інтромітували наявну жінку; 900 s для EL, якщо самці не досягали еякуляції і 1800s для PEI, якщо самці не інтромітували після першої еякуляції. У другому випадку ті тварини, які не копулювали до еякуляції, були виключені з аналізів. Це було зроблено для оцінки того, чи можуть відмінності в сексуальній поведінці співвідноситися з відмінностями в концентраціях позаклітинного допаміну, DOPAC і NA в mPFC римських ліній щурів під час двох фаз сексуальної активності, що виключає можливі заплутані втручання через включення суб'єктів, які не виявляли сексуальної поведінки, призначаючи їм фіксовані значення для аналізованих статевих параметрів.
Було проведено кілька статистичних аналізів з ANOVA, щоб виявити і краще охарактеризувати можливі відмінності між використаними експериментальними умовами. Перед виконанням ANOVA, набори даних кожної з різних експериментальних змінних були перевірені на однорідність дисперсій між чотирма експериментальними групами (зробленими з усіма тваринами або з щурами, які копулювали тільки до еякуляції) з тестом Бартлетта або Левіна залежно від випадку. Коли були виявлені суттєві відмінності в варіаціях, набори даних були логарифмічно трансформованими (тобто експериментальні значення Y, змінені на значення Log Y), повторно перевірили на однорідність відхилень і потім аналізували за допомогою ANOVA. Коротко, поведінкові параметри, отримані раніше (наприклад, з недоступною жінкою) і під час першої серії копуляторної активності (від першого монтування / інтромісії до першого монтування / інтромії після першої еякуляції) наївних і статево досвідчених щурів RHA і RLA під час мікродіалізу аналізували двома шляхами ANOVA, використовуючи лінію щурів і рівень статевого досвіду між факторами суб'єктів (середні значення поведінкових параметрів наведені в таблиці 1 і F значення і рівні значимості аналізів ANOVA в табл 2).
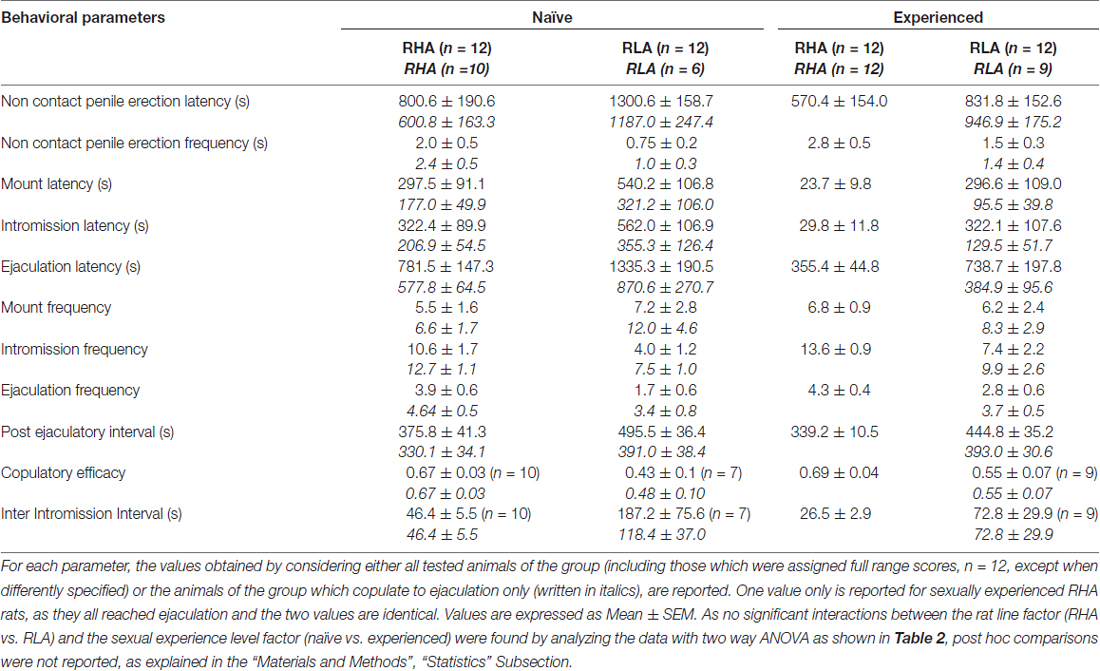 Таблиця 1. Копуляторні параметри, виміряні в першій серії копуляторної активності (яка починається з першого монтування / вторгнення, коли самка стає доступною для самця і закінчується після інтервалу після еякуляції з першим монтуванням / втручанням другого ряду) і не контактує пенілем ерекції статевозрілих і досвідчених щурів RHA і RLA.
Таблиця 1. Копуляторні параметри, виміряні в першій серії копуляторної активності (яка починається з першого монтування / вторгнення, коли самка стає доступною для самця і закінчується після інтервалу після еякуляції з першим монтуванням / втручанням другого ряду) і не контактує пенілем ерекції статевозрілих і досвідчених щурів RHA і RLA.
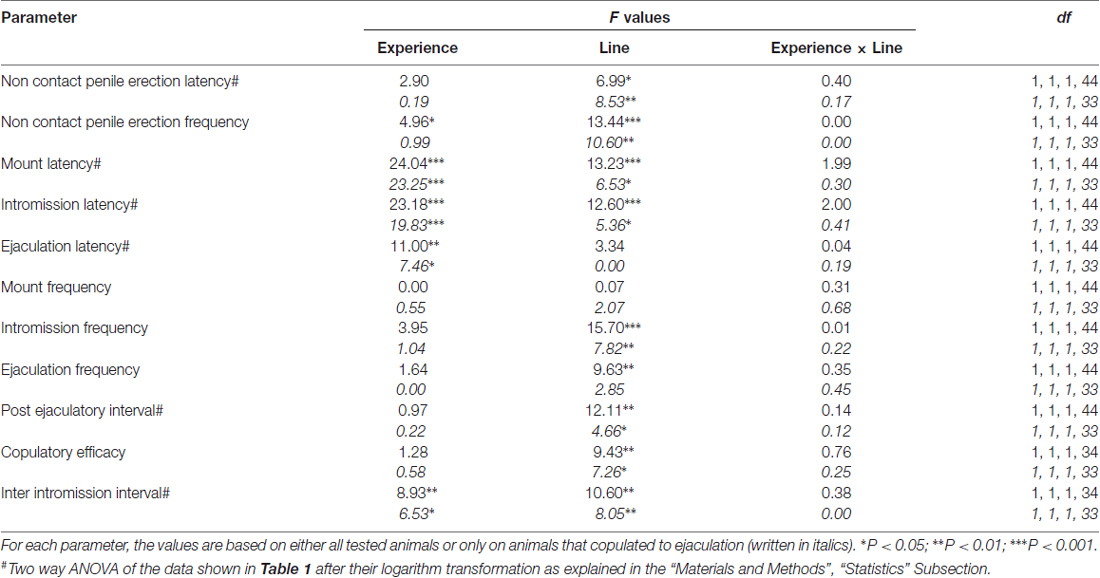 Таблиця 2. F значення і рівні значущості двостороннього ANOVA, виконані за даними, наведеними в табл 1 за допомогою лінії щурів (RHA проти RLA) і рівня сексуального досвіду (наївні проти досвідчених) як між суб'єктними факторами.
Таблиця 2. F значення і рівні значущості двостороннього ANOVA, виконані за даними, наведеними в табл 1 за допомогою лінії щурів (RHA проти RLA) і рівня сексуального досвіду (наївні проти досвідчених) як між суб'єктними факторами.
Крім того, загальний аналіз даних, отриманих від кожної щури під час мікродіалізу, проводили шляхом розрахунку спочатку AUC, отриманих нанесенням значень концентрацій допаміну, DOPAC або NA або кількості неконтактних ерекцій, гомілок, інтромісій та еякуляцій час (починаючи з 1 h після введення самця щури в спарову клітку до кінця експерименту -180 min, розділеної на фракції 15 min для нейрохімічних значень - або в той час, коли самку вводили в спарову клітку). для поведінкових параметрів), а потім шляхом порівняння розрахункових значень двома шляхами ANOVA, використовуючи лінію щурів і рівень сексуального досвіду між факторами суб'єктів (середні значення AUC нейрохімічних і поведінкових параметрів наведені в таблиці 4 і F значення і рівні значимості аналізів ANOVA в табл 5). Нарешті, більш детальний аналіз кожної сукупності даних з точки зору до точки (тобто, значення концентрацій допаміну, DOPAC, NA та кількості неконтактних ерекцій, монтувань, інтромісій та еякуляцій у порівнянні з часом) виконувалися факторним ANOVAs для повторних заходів, використовуючи лінію щурів і рівень сексуального досвіду між суб'єктними факторами і часом (тобто фракціями діалізату), як в межах суб'єктного фактора ( F Значення та рівні значущості цих аналізів ANOVA наведено в табл 6). Аналогічні аналізи були проведені також з урахуванням лише базальних значень нейрохімічних показників (останні чотири аліквоти диализата, зібрані перед введенням у спарову клітку рецептивної жінки), середні значення яких наведені в табл. 3. В якості перших, але не другого порядку взаємодії були виявлені при виконанні загальних факторних ANOVAs для повторних заходів, зокрема ліній × Час і Досвід × Час взаємодії для нейрохімічних параметрів (див. Таблицю) 6), двосторонні ANOVAs з лінією або рівнем сексуального досвіду як між суб'єктом фактора і часу, як в межах суб'єкт-фактора були виконані на цих наборах даних шляхом безпосереднього порівняння сексуальних наївних або досвідчених RHA проти RLA щурів або статевих контактів наївні щури RHA або статево досвідчені щури проти RIA. Результати Постфактум пара-мудрі контрасти, що виконуються за допомогою тесту HSK Tukey на значні взаємодії, виявлені цими двома шляхами ANOVA, наведені на малюнку 2. У всіх інших випадках, Постфактум порівняння не повідомлялося, оскільки ANOVA не виявила жодної істотної взаємодії між лінією щурів, рівнем сексуального досвіду та часом. Статистичні аналізи були проведені з Graph Pad 5 (PRISM, Сан-Дієго, Каліфорнія, США) і STATISTICA 12 (Statsoft, Tulsa, OK, США) з рівнем значущості, встановленим на P < 0.05.
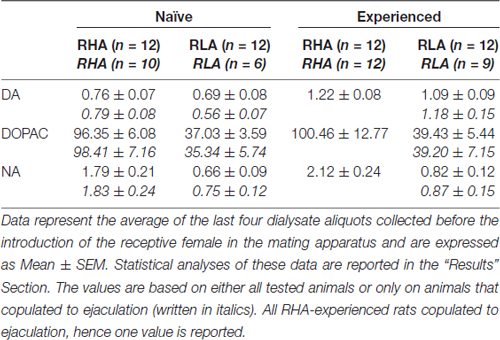 Таблиця 3. Концентрації базальних допамінів (DA), 3,4-дигідроксифенилуксусной кислоти (DOPAC) і норадреналіну (NA) в диализате з медіальної префронтальної кори (mPFC) статевозрілих і досвідчених щурів RHA і RLA.
Таблиця 3. Концентрації базальних допамінів (DA), 3,4-дигідроксифенилуксусной кислоти (DOPAC) і норадреналіну (NA) в диализате з медіальної префронтальної кори (mPFC) статевозрілих і досвідчених щурів RHA і RLA.
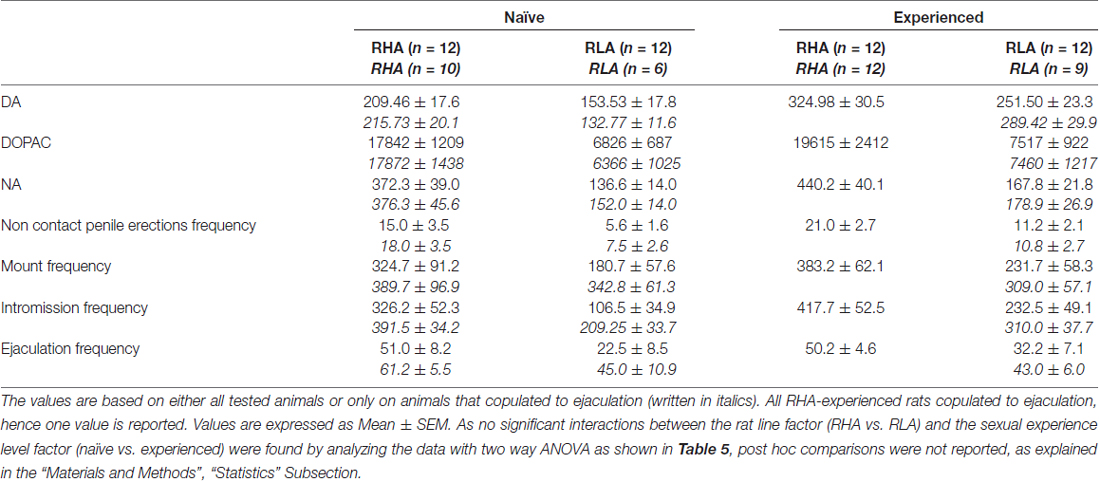 Таблиця 4. Загальна оцінка відмінностей концентрацій DA, DOPAC і NA, а також сексуальної поведінки між сексуальними наївними і досвідченими щурами RHA і RLA шляхом аналізу усереднених AUC, отриманих з результатів, показаних на фігурах 2 та 3.
Таблиця 4. Загальна оцінка відмінностей концентрацій DA, DOPAC і NA, а також сексуальної поведінки між сексуальними наївними і досвідченими щурами RHA і RLA шляхом аналізу усереднених AUC, отриманих з результатів, показаних на фігурах 2 та 3.
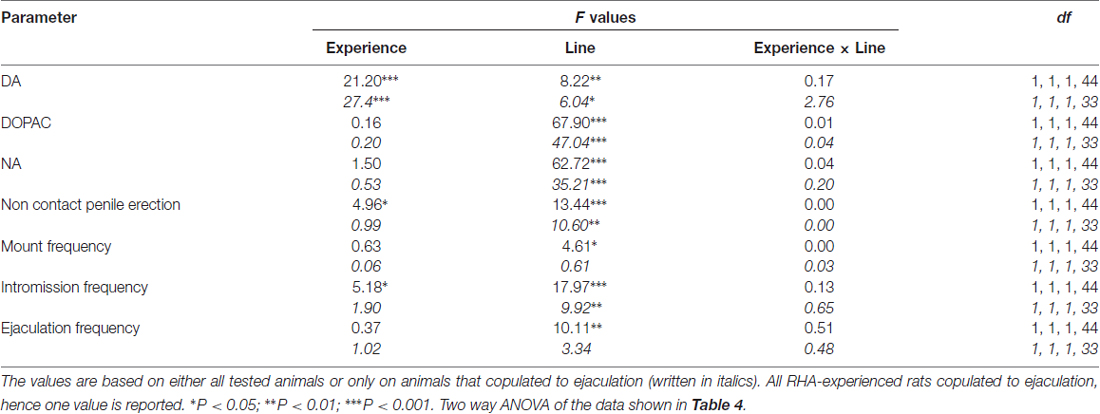 Таблиця 5. F значення і рівні значущості двосторонніх ANOVA, виконаних за даними, наведеними в табл 4 за допомогою лінії щурів (RHA проти RLA) і рівня сексуального досвіду (наївні проти досвідчених) як між суб'єктними факторами.
Таблиця 5. F значення і рівні значущості двосторонніх ANOVA, виконаних за даними, наведеними в табл 4 за допомогою лінії щурів (RHA проти RLA) і рівня сексуального досвіду (наївні проти досвідчених) як між суб'єктними факторами.
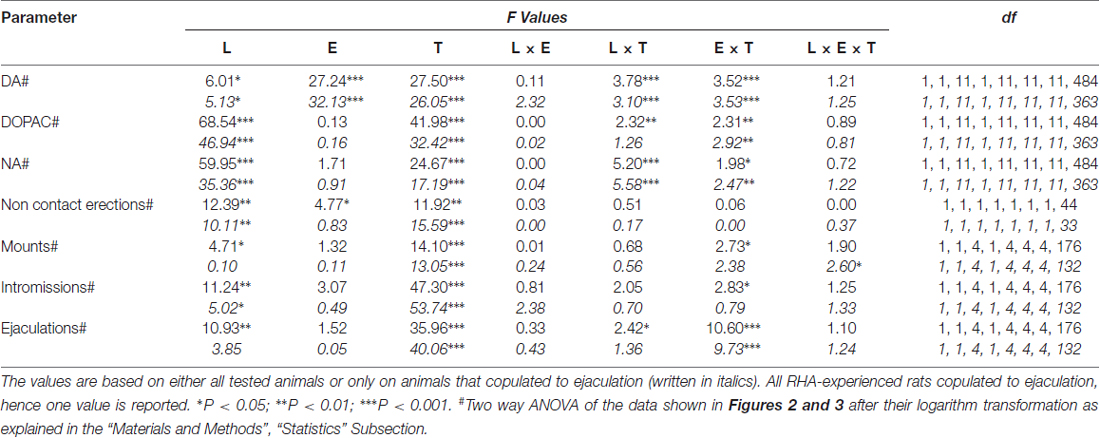 Таблиця 6. F значення і рівні значущості загальних факторних ANOVAs для повторних заходів, виконаних за результатами, показаними на Фігурах 2 та 3 за допомогою лінії щурів (L; RHA проти RLA) і рівня сексуального досвіду (E) (наївка проти досвідченого) як між факторами суб'єкта, так і за часом (T) (фракції диализата) як у суб'єктному факторі.
Таблиця 6. F значення і рівні значущості загальних факторних ANOVAs для повторних заходів, виконаних за результатами, показаними на Фігурах 2 та 3 за допомогою лінії щурів (L; RHA проти RLA) і рівня сексуального досвіду (E) (наївка проти досвідченого) як між факторами суб'єкта, так і за часом (T) (фракції диализата) як у суб'єктному факторі.
 малюнок 2. Концентрації дофаміну (DA), DOPAC і норадреналіну (NA) в дисперсному розчині mPFC від статевих (A, C, E) і досвідчені RHA і RLA самці щурів (B, D, F) під час сексуальної активності з сприйнятливою жінкою. Статеві наївні (тобто ніколи не піддані сексуальному сприйняттю жінок) і статеві особи (які пройшли п'ять копуляційних тестів) обох ліній з мікродомішковим зондом, стереотаксично імплантованим в mPFC, поміщали індивідуально в спарову клітку. Зонд мікродіалізу перфузували буфером для діалізу, як описано в розділі «Матеріали і методи». Після збору чотирьох аліквот для діалізату для визначення базальних значень, недоступну сприйнятливу самку помістили всередину невеликої клітки пристрою для спарювання (час = 0, inaccess.fe). Після 30 хв невелика клітка була вилучена і копуляція була дозволена для 75 хв (копуляція), після чого самка була вилучена з спарювальної клітини (fe.remov). Під час експерименту вимірювали неконтактні ерекції і копуляторні параметри, а алігони диализата збирали кожні хв. НМХХ хв і аналізували на допамін, DOPAC і NA, як описано в розділі «Матеріали і методи». Значеннями є значення ± SEM значень, отриманих всіма щурами 15 на групу (RHA_all = суцільні зелені лінії, RLA_all = тверді коричневі лінії) або ті, які копулюються тільки для еякуляції (RHA_cop = пунктирні червоні лінії; RLA_cop = пунктирні сині лінії). У досвідчених щурів RHA всі тварини досягали копуляції і значення ідентичні. *P <0.05 щодо базових значень (немає жінки) групи (зелений для RHA_all; червоний для RHA_cop, коричневий для RLA_all, синій для RLA_cop); #P <0.05 щодо відповідних значень групи RLA (червоний, RHA_all проти RLA_all; зелений, RHA_cop проти RLA_cop); §P <0.05 щодо значень часу, що відповідають статевим наївним щурам (зелений, досвідчений RHA_all проти наївного RHA_all; червоний, досвідчений RHA_cop проти наївного RHA_cop; коричневий, досвідчений RLA_all проти наивного RLA_all; синій, досвідчений RLA_cop проти наївного RLA_cop) (двосторонні ANOVA, зроблені на показаних даних після їх логарифмічного перетворення, як пояснюється в "Матеріали та методи", підрозділ "Статистика", після чого проводяться тести HSD Тукі).
малюнок 2. Концентрації дофаміну (DA), DOPAC і норадреналіну (NA) в дисперсному розчині mPFC від статевих (A, C, E) і досвідчені RHA і RLA самці щурів (B, D, F) під час сексуальної активності з сприйнятливою жінкою. Статеві наївні (тобто ніколи не піддані сексуальному сприйняттю жінок) і статеві особи (які пройшли п'ять копуляційних тестів) обох ліній з мікродомішковим зондом, стереотаксично імплантованим в mPFC, поміщали індивідуально в спарову клітку. Зонд мікродіалізу перфузували буфером для діалізу, як описано в розділі «Матеріали і методи». Після збору чотирьох аліквот для діалізату для визначення базальних значень, недоступну сприйнятливу самку помістили всередину невеликої клітки пристрою для спарювання (час = 0, inaccess.fe). Після 30 хв невелика клітка була вилучена і копуляція була дозволена для 75 хв (копуляція), після чого самка була вилучена з спарювальної клітини (fe.remov). Під час експерименту вимірювали неконтактні ерекції і копуляторні параметри, а алігони диализата збирали кожні хв. НМХХ хв і аналізували на допамін, DOPAC і NA, як описано в розділі «Матеріали і методи». Значеннями є значення ± SEM значень, отриманих всіма щурами 15 на групу (RHA_all = суцільні зелені лінії, RLA_all = тверді коричневі лінії) або ті, які копулюються тільки для еякуляції (RHA_cop = пунктирні червоні лінії; RLA_cop = пунктирні сині лінії). У досвідчених щурів RHA всі тварини досягали копуляції і значення ідентичні. *P <0.05 щодо базових значень (немає жінки) групи (зелений для RHA_all; червоний для RHA_cop, коричневий для RLA_all, синій для RLA_cop); #P <0.05 щодо відповідних значень групи RLA (червоний, RHA_all проти RLA_all; зелений, RHA_cop проти RLA_cop); §P <0.05 щодо значень часу, що відповідають статевим наївним щурам (зелений, досвідчений RHA_all проти наївного RHA_all; червоний, досвідчений RHA_cop проти наївного RHA_cop; коричневий, досвідчений RLA_all проти наивного RLA_all; синій, досвідчений RLA_cop проти наївного RLA_cop) (двосторонні ANOVA, зроблені на показаних даних після їх логарифмічного перетворення, як пояснюється в "Матеріали та методи", підрозділ "Статистика", після чого проводяться тести HSD Тукі).
результати
Щури RHA та RLA показують різну кількість не контактних ерекцій та різних моделей копуляторного поведінки
Відповідно до попередніх досліджень (Sanna et al., 2014a,b, 2015), інша кількість щурів RHA і RLA, котрі стають сексуальними, не залучені до статевої активності, коли вони об'єднані з сприйнятливою жінкою під час збору аліквот диализата з mPFC шляхом внутрішньомозкового мікродіалізу. Коротко, в цьому дослідженні 10 з 12 статевозрілих щурів RHA (83%) копулювали до еякуляції у своєму першому копуляторному тесті, проти тільки 6 з 12 наївних RLA щурів (50%) під час експерименту мікродиалізу. Ця різниця була виявлена також у статево досвідчених щурів RHA і RLA (після п'яти копуляторних тестів), хоча і ослаблені, при цьому всі щури RHA 12 досягали еякуляції проти 9 з щурів 12 RLA під час мікродіалізу. Завжди у згоді з попередніми дослідженнями, також у цьому дослідженні дві римські лінії щурів демонстрували різні латентності і частоти не контактних ерекцій статевого члена, коли вони піддавалися недоступній сприйнятливій жінці, а також різним моделям копуляторної поведінки під час сексуальної взаємодії. Ці відмінності були виявлені або при розгляді даних всіх експериментальних тварин даної групи, незалежно від того, чи вони копулювалися до еякуляції, або ні (наприклад, шляхом присвоєння повних балів тим тваринам, які не показували поведінку, тобто щурів, які не копулювали до еякуляції) або при розгляді даних про тварин, які показували тільки поведінку даної групи (наприклад, тварин, які копулювали до еякуляції; 1). Відповідно, статистичний аналіз значень неконтактних ерекцій (NCPE) і параметрів копуляції, виміряних у першій серії копуляторної активності двостороннім ANOVAs, виявив значні відмінності між двома лініями щурів або при розгляді всіх експериментальних щурів або тільки копулюючих щурів. у чотирьох експериментальних групах (див. табл 2). Фактично, коли розглядалися дані, отримані від всіх експериментальних тварин, кількість неконтактних ерекцій була вищою, а NCPEL, ML, IL і PEI були значно коротшими у щурів RHA порівняно з щурами RLA. Крім того, EF і CE були вищими, тоді як III був значно коротшим у щурів RHA порівняно з щурами RLA. Деякі з наведених вище відмінностей між двома римськими лініями щурів мали тенденцію до зменшення або зникнення при повторних копуляторних тестах. Тим не менш, деякі з цих відмінностей все ще існували після стабілізації сексуальної поведінки шляхом повторного сексуального досвіду. Аналогічні результати були отримані при розгляді тільки значень тварин чотирьох експериментальних груп, які копулювали до еякуляції, за винятком EF (див. Таблиці). 1, 2).
Базальні концентрації позаклітинного допаміну, допаку та норадреналіну в диализах mPFC від сексуальних наївних та досвідчених щурів RHA та RLA
За нинішніх експериментальних умов кількість допаміну, DOPAC і NA в диализатах, отриманих з mPFC всіх (наприклад, незалежно від того, що вони копулювали до еякуляції, чи ні) щурів RHA і RLA, що не мали статевого життя, становила 2.32 pg і 2.11 pg для допаміну, 322.56 pg і 124.42 pg для DOPAC та 6.05 pg та 2.24 pg для NA, відповідно, у 20 μl діалізату. Аналогічні кількості вимірювали в диализатах, отриманих з mPFC щурів, що мають досвід сексуально RHA і RLA (допамін: xNUMX pg і 3.73 pg, DOPAC: xNUMX pg і xNUMX pg, NA: 3.33 pg і 353.62 pg у щурів RHA і RLA, відповідно) . Ці значення вказують на концентрацію N133.32 – 7.17 nM та N2.78 – 0.8 nM для позаклітинного дофаміну та DOPAC відповідно, а N1.2 – 95 nM для позаклітинного NA, у mPFC щурів RHA та концентрації N100 – 1.8 nM і N2.2 – 0.8 nM для позаклітинного дофаміну і DOPAC, відповідно, і N1.2 – 35 nM для позаклітинного NA, у mPFC щурів RLA (таблиця) 3). Зазначені вище значення були отримані після періоду 2 h рівноваги буфера для діалізу з позаклітинною рідиною mPFC. Оскільки відновлення автентичних допамінів, DOPAC і NA діалізних зондів було оцінено як близьке до 20%, концентрації позаклітинного дофаміну, DOPAC і NA можуть бути оцінені як близькі до N4 – 5 nM в обох лініях для допаміну, N500 ≅ 200 nM для DOPAC і N10 і N4 nM для NA у mPFC щурів RHA і RLA відповідно. Факторіальні ANOVAs для повторних заходів, проведених на вищевказаних значеннях після логарифмічної трансформації, виявили значні відмінності в базальних рівнях дофаміну між сексуальними наївними і статево досвідченими щурамиF(1,44,132) = 26.05, P <0.001), а в базальних рівнях DOPAC (F(1,44,132) = 63.36, P <0.001) та НС (F(1,44,132) = 42.14, P <0.001) між щурами RHA та RLA (останні чотири зразки, зібрані перед введенням самки в маленьку клітку). Подібні значення були виявлені при розгляді лише щурів, копулировавшихся до еякуляції під час експерименту мікродіалізу (табл 3). Також у цьому випадку факторні ANOVAs для повторних заходів, проведених на значеннях після логарифмічної трансформації, виявили значні відмінності в базальних рівнях дофаміну між сексуальними наївними і статево досвідченими щурами (F(1,33,99) = 26.97, P <0.001), а в базальних рівнях DOPAC (F(1,33,99) = 42.95, P <0.001) та НС (F(1,33,99) = 27.63, P <0.001) між щурами RHA та RLA.
Концентрації позаклітинного дофаміну, допаку і норадреналіну в мДФК диализатах від статевонеологічних та досвідчених щурів RHA та RLA змінюються диференційно під час статевої активності
Наявність недоступної сприйнятливої жінки у внутрішній маленькій клітці та подальшої безпосередньої статевої взаємодії збільшувала концентрації позаклітинного допаміну, DOPAC та NA у диализатах mPFC, отриманих як від статевозрілої, так і від досвідчених щурів RHA і RLA, хоча зі значними відмінностями між дві лінії щурів і рівень умов досвіду, або при розгляді всіх експериментальних тварин, або тільки тих, які копулировали до еякуляції (Фігура 2). Насправді, двосторонні ANOVAs AUCs концентрацій допаміну, DOPAC і NA показали, що загальний вміст дофаміну, DOPAC і NA всього тесту, були значно вищими у щурів RHA порівняно з RLA щурами, і в сексуально у порівнянні з наївними щурами (див. табл 4 для значень AUC та таблиці 5 та цінності F значення та рівень значимості). Ці відмінності були додатково підтверджені факторними ANOVAs з точки до точки для повторних вимірювань кількості дофаміну, DOPAC та NA двох статевих наївних та досвідчених ліній щурів вздовж експерименту, які виявили значні основні ефекти лінії, досвіду, часу та значної лінії × час та Досвід × Час першого порядку взаємодії (див. табл 6 та цінності F значення та рівень значимості).
Концентрації позаклітинного дофаміну, допаку і норадреналіну в мДФК-диализатах, отриманих від сексуальних наївних щурів RHA і RLA, змінюються диференційно під час статевої активності
У сексуальних наївних щурів RHA і RLA позаклітинний дофамін, але не DOPAC, збільшувалися спочатку при введенні в присутності недоступної жінки, тоді як NA збільшувалася тільки у щурів RHA. У наївних щурів RHA позаклітинний дофамін додатково збільшувався в першій 15 хв. Статевої взаємодії, як було виявлено з DOPAC і NA. Збільшення екстрацелюлярного дофаміну, DOPAC і NA тривало протягом усього часу копуляції, з дофаміном, DOPAC і NA досягали пікового значення в хв. XMUM, 60 хв і 75 хв відповідно, потім позаклітинний допамін і, меншою мірою, DOPAC і NA , знизившись до значень, схожих з базальними після видалення жінки.
З іншого боку, перше значне збільшення і пікове значення позаклітинного дофаміну, DOPAC і NA відбувалося в нефабрикованих щурів RLA протягом першого хв. Однак при відхиленні від щурів RHA у RLA щурів позаклітинний допамін, DOPAC і NA мали тенденцію до повернення до значень, схожих на базальні, протягом першої 15 хв безпосередньої взаємодії з самкою (див. Фігури 2A, C, E).
Нарешті, під час копуляції збільшення екстрацелюлярного дофаміну було більшим і більш стійким у сексуальних наївних RHA, ніж у щурів RLA (див. Малюнок) 2Aв той час як у відсутності жінок їх концентрації були подібними, хоча і не ідентичними, на двох лініях щурів (див. вище та табл 3). Цікаво, хоча не було виявлено різниці між двома римськими лініями у відсотках збільшення DOPAC і, в меншій мірі, концентрацій NA протягом присутності сприйнятливої жінки, різниця більш ніж у два рази в абсолютних значеннях концентрацій DOPAC і NA спостерігали між двома лініями щурів, як у базальних умовах (див. табл 3) і протягом усього тесту з сприйнятливою жінкою (див. малюнки 2C, E).
Концентрації позаклітинного допаміну, допаку і норадреналіну в диализатах мПФК, отриманих від статево досвідчених щурів RHA і RLA, змінювалися диференційно під час сексуальної активності
Як було виявлено у сексуальних наївних щурів RHA і RLA, присутність недоступних жінок і подальша безпосередня статева взаємодія призвели до збільшення позаклітинного дофаміну і DOPAC в mPFC-диализате, отриманому від статево досвідчених щурів RHA і RLA. Однак у цьому випадку були виявлені відмінності, пов'язані з лінією щурів, або при розгляді всіх експериментальних тварин, або, в меншій мірі, тільки тих, які копулювали до еякуляції (ілюстрація 2). У статево досвідчених щурів RHA і RLA перше збільшення позаклітинного дофаміну відбувалося з недоступною жінкою. Після цього концентрація дофаміну збільшувалася під час копуляції в обох лініях щурів. Концентрації дофаміну досягали пікових значень після копуляції 45 – 60 і повільно зменшувалися до базових значень наприкінці копуляторного тесту (див. Малюнок) 2B). Подібно до статевозрілих щурів, концентрації позаклітинного дофаміну були вищими у статево досвідчених щурів RHA порівняно з щурами RLA, головним чином, в аликвотах диализата, зібраних у центральній частині фази копуляції (після копуляції 30 – 45). Концентрації DOPAC зростали під час присутності недоступної жінки тільки у щурів RHA, тоді як вона збільшувалася до аналогічної міри в обох лініях щурів під час копуляції (пікові значення при 75 – 90 min) і, як правило, поверталися до базальних значень наприкінці копуляторний тест (див. рис 2D). Концентрації NA також збільшувалися в обох лініях, що зазнали статевого поширення, після введення жіночої статі, досягнувши пікових значень у хв. 90 у щурів RHA і в хв. 2F). При відхиленні від дофаміну (див. Вище), аналогічні відмінності були виявлені в підвищенні концентрацій DOPAC і NA у статево досвідчених щурів з обох ліній при розгляді всіх тварин проти тварин, які досягли тільки еякуляції. Проте, як було виявлено у щурів, які не мали сексуального характеру, хоча відсоток збільшення концентрацій DOPAC і NA був подібним, істотні відмінності в абсолютних значеннях DOPAC і NA були виявлені між щурами, які мають досвід RHA та RLA під час проведення тесту (тобто до, під час і після присутність сприйнятливої жінки), коли щури RHA показують більш високі значення, ніж щури RLA (див. Таблицю 3 для базових значень і цифр 2D, F для значень по всьому тесту).
Статевий досвід впливає на зміни концентрацій дофаміну та норадреналіну в диализатах mPFC, отриманих від щурів RHA та RLA під час сексуальної активності
Порівняння концентрацій позаклітинного дофаміну та NA у статевозрілих щурів з RHA та RLA показало, що сексуальний досвід змінив концентрацію позаклітинного дофаміну і, в меншій мірі, NA на обох лініях щурів, або при розгляді всіх експериментальних тварин або ті, які копулюються тільки до еякуляції (Фігури 2A, B, E, F). Відповідно, суттєві відмінності спостерігалися в базальних значеннях дофаміну між сексуальними наївними і досвідченими щурами обох ліній (див. Таблицю). 3), а значення дофаміну під час копуляторної активності в цілому були вищими у статево досвідчених щурів RHA та RLA порівняно з їхніми наївними аналогами (див. малюнок) 2B). Більш того, у щурів, які переживали статевий випадок, хоча збільшення дофаміну, виявлене під час присутності недоступної жінки, відбувалося за тією ж тимчасовою схемою, що спостерігалася у щурів, які не мали сексуального характеру, при копуляції були виявлені відмінності у тимчасовій картині. Насправді, в цьому випадку, при відхиленні від того, що спостерігалося у сексуальних наївних щурів RHA (один головний пік при 60 хв), було виявлено два основних пікових значення, перший після 15 хв і другий після 45 хв копуляції. Після цього останнього збільшення значення дофаміну мали тенденцію до повернення до базальних значень (див. Малюнок 2B).
Аналогічна картина була виявлена у сексуальних наївних щурів проти RLA. Фактично, як і у щурів RHA, також у досвідчених щурів RLA була виявлена тимчасова картина, яка характеризується трьома основними збільшеннями позаклітинного дофаміну: перша з недоступною жінкою та двома іншими під час копуляції, тобто після 15 min і 60 хв копуляції (див. малюнки 2A, B). Загалом, сексуальний досвід, здається, призводить до більш тривалих змін у тимчасовій картині вивільнення дофаміну в обох лініях щурів при переході від статевого наївного до пережитого стану.
Як показано на малюнках 2C – Fвиявлено лише невеликі відмінності в концентраціях DOPAC і NA між наївними і досвідченими щурами обох ліній. Однак незначна тенденція у позаклітинному НС до більш високих значень у досвідчених у порівнянні з наївними щурами спостерігалася у базальних (до введення рецептивної жінки, див. Таблицю) 3) і в загальних кількостях НС, як було виявлено за значеннями AUC (див. таблицю) 4) в розрахунку на позаклітинні концентрації, отримані з усього тесту (до і під час присутності сприйнятливої жінки). Нарешті, тимчасова картина концентрацій НА була дуже схожою при порівнянні досвідчених щурів обох ліній щурів з їхніми наївними аналогами, що вказує на те, що сексуальний досвід може призвести до загального збільшення позаклітинної НА, ніж до змін у її зростанні (див. Фігури) 2E, F).
Зміни концентрацій позаклітинного дофаміну та норадреналіну в диализатах mPFC, отриманих від сексуальних наївних та досвідчених щурів RHA та RLA, відбувалися одночасно зі змінами в статевому поведінці
Відмінності в концентраціях позаклітинного допаміну, DOPAC і NA в диализатах, отриманих з mPFC, знайдені як у статевозрілих, так і у досвідчених щурів RHA і RLA під час випробування (ілюстрація 2) відбувалися разом з модифікаціями різних статевих параметрів, виміряних протягом двох основних фаз експериментів (тобто, жінки недоступні і доступні для чоловіків, відповідно), які включають, крім іншого, не контактні ерекції пеніса (рис. 3A, B), кріплення (Фігури) 3C, D), intromissions (Цифри 3E, F) і еякуляції (рис 3G, H). Ці відмінності були виявлені або при розгляді всіх експериментальних тварин, або тих, які копулювали тільки до еякуляції. Фактично попередній аналіз значень AUCs цих параметрів здійснюється за допомогою двосторонніх ANOVA (див. Табл 4 для значень AUC та таблиці 5 та цінності F Значення та рівень значущості), за якими слідує точковий аналіз значень цих параметрів вздовж експерименту шляхом факторного ANOVA для повторних вимірювань виявили значні основні ефекти лінії, досвіду, часу, значущої лінії першого порядку × Час та Досвід × часу взаємодії, і значну взаємодію другого порядку для кріплення (див. Таблицю 6 та цінності F значення та рівень значимості). Відповідно, відповідно до попередніх досліджень (Sanna et al., 2015): (i) щури, які не піддавалися сексуальному вжитку, показали більшу не контактну ерекцію статевого члена, інфекції та еякуляції, ніж їхні колеги RLA; (ii) статево досвідчені щури RHA показали більшу кількість неконтрактних ерекцій пеніса, інтромісій та еякуляцій, ніж їхні колеги RLA; і (iii) щурів, що мають статевий досвід обох ліній, і, зокрема, щурів RLA, показали більш неконтрактну ерекцію статевого члена, інтромезії та еякуляції, ніж їхні сексуально наївні аналоги. Насправді, в той час як у щурів RHA більша частина змін, що відбуваються внаслідок сексуального досвіду, відбувалася в часі, а не в частоті поведінкових параметрів (наприклад, досвідчені щури вживалися в основному в першій половині тесту, тоді як наївні щури показали аналогічна копуляторная тенденція протягом усього тесту), у щурів RLA більш виражене загальне поліпшення сексуальної поведінки, головним чином у статевих показниках, було вироблено статевим досвідом, як виявлено змінами значень копуляторних параметрів у досвідчених щурів RLA порівняно з їх наївні колеги.
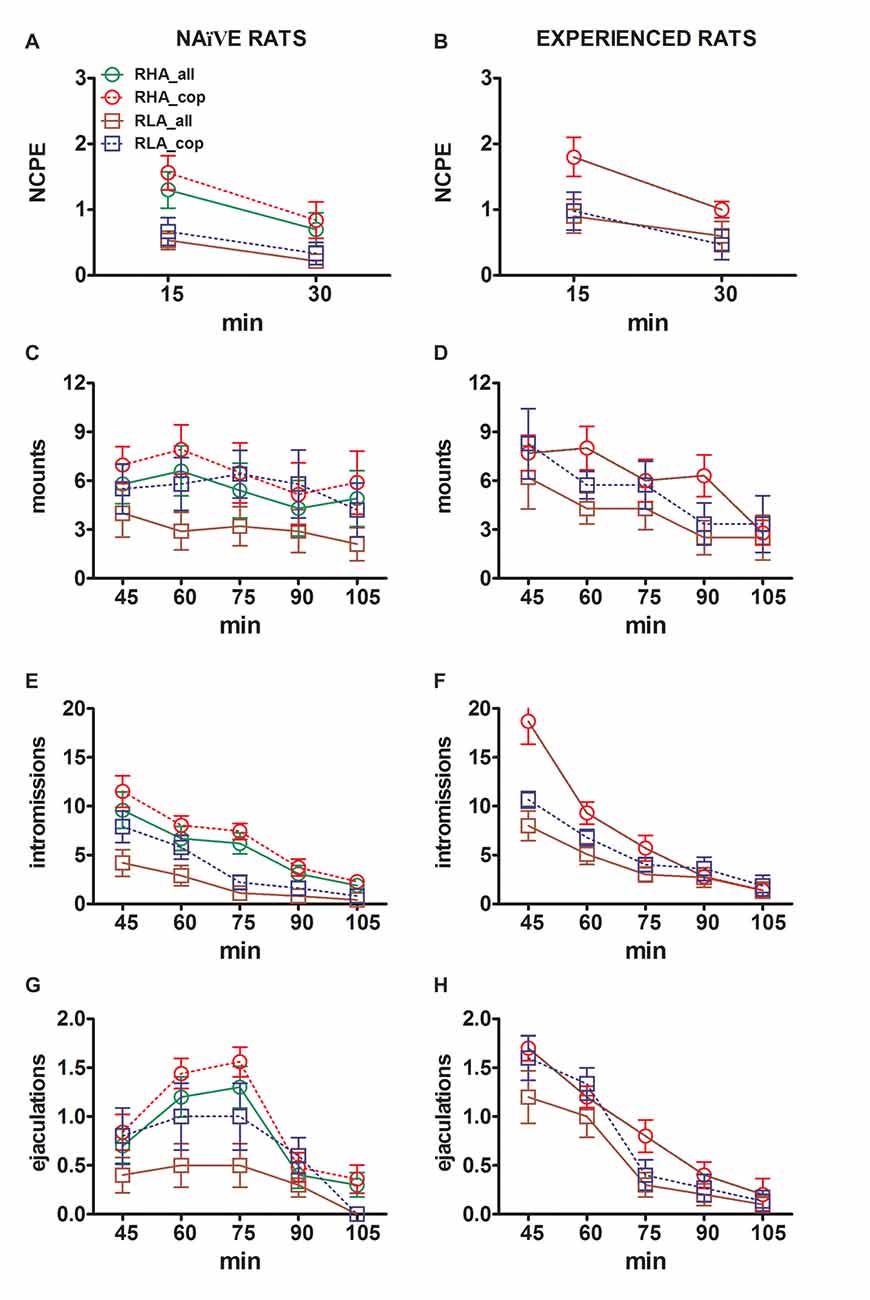 малюнок 3. Зміни в кількості неконтактних ерекцій статевого члена (NCPE), опіків, інтроміусів і еякуляцій, зареєстрованих з сексуальної наївності (A, C, E, G) і досвідчені щури RHA і RLA (B, D, F, H) під час експериментів з мікродіалізу, наведених на рис 2. Всі експериментальні умови були ідентичні описаним в легенді на Фіг 2. Значеннями є значення ± SEM значень, отриманих усіма щурами 12 на групу (включаючи ті, які були призначені для повних балів; RHA = суцільні зелені лінії, RLA = тверді коричневі лінії) або ті, які копульовані тільки до еякуляції (RHA = пунктирні червоні лінії) RLA = пунктирні сині лінії). У досвідчених щурів RHA всі тварини досягали копуляції і значення ідентичні. Оскільки не було виявлено значних взаємодій між коефіцієнтом лінії щурів (RHA проти RLA), рівень факторів статевого досвіду (наївний чи досвідчений) та час були виявлені шляхом аналізу даних двовимірними ANOVA, як показано в таблиці 6, Постфактум не було повідомлено про порівняння, як пояснюється в підрозділі "Матеріали та методи", "Статистика".
малюнок 3. Зміни в кількості неконтактних ерекцій статевого члена (NCPE), опіків, інтроміусів і еякуляцій, зареєстрованих з сексуальної наївності (A, C, E, G) і досвідчені щури RHA і RLA (B, D, F, H) під час експериментів з мікродіалізу, наведених на рис 2. Всі експериментальні умови були ідентичні описаним в легенді на Фіг 2. Значеннями є значення ± SEM значень, отриманих усіма щурами 12 на групу (включаючи ті, які були призначені для повних балів; RHA = суцільні зелені лінії, RLA = тверді коричневі лінії) або ті, які копульовані тільки до еякуляції (RHA = пунктирні червоні лінії) RLA = пунктирні сині лінії). У досвідчених щурів RHA всі тварини досягали копуляції і значення ідентичні. Оскільки не було виявлено значних взаємодій між коефіцієнтом лінії щурів (RHA проти RLA), рівень факторів статевого досвіду (наївний чи досвідчений) та час були виявлені шляхом аналізу даних двовимірними ANOVA, як показано в таблиці 6, Постфактум не було повідомлено про порівняння, як пояснюється в підрозділі "Матеріали та методи", "Статистика".
Обговорення
Це дослідження підтверджує та розширює результати попередніх досліджень, які показують, що щури RHA та RLA, які демонструють різний стиль стимуляції у відповідь на аверсивні умови (Driscoll і Bättig, 1982; Giorgi et al., 2003a; Steimer і Driscoll, 2003), демонструють чіткі поведінкові закономірності в присутності недоступних сприйнятливих жінок і під час класичних копуляторних тестів (Sanna et al., 2014a,b, 2015). Відповідно, це дослідження також показує, що сексуальні наївні щури RHA (ніколи не піддавалися рецептивній жіночій групі) виявляли більш високу сексуальну мотивацію і кращу сексуальну продуктивність у порівнянні з сексуальними наївними щурами RLA, наприклад, щури RHA показують більш високу кількість неконтактних ерекцій пеніса (феромон- індуковані ерекції статевого члена вважаються показником статевого збудження; Sachs et al., 1994; Sachs, 2000) і різні про-статеві зміни в кількох копуляторних параметрах, які були виявлені в попередніх дослідженнях (Sanna et al., 2014a,b, 2015). Ці відмінності як і раніше зберігалися в обох римських лініях щурів після придбання статевого досвіду, тобто після стабілізації сексуальної поведінки з п'ятьма експозиціями та копуляційними тестами з сприйнятливою жінкою. Це дослідження також вперше показує, що як у лінії RHA, так і у RLA, у щурів, як у ненадійному, так і в досвідченому стані, не контактна ерекція пеніса та статева взаємодія, відбувалися одночасно із збільшенням концентрації позаклітинного допаміну (та його основного метаболіту DOPAC). ) і NA в диализатах, отриманих шляхом внутрішньомозкового мікродіалізу з mPFC обох римських ліній щурів. Цікаво, що ці збільшення позаклітинного дофаміну і NA в mPFC пов'язані з відмінностями або в фенотипі щурів, або в рівні сексуального досвіду. Відповідно, як показано в розділі «Результати», як сексуальні наївні, так і досвідчені щури RHA демонстрували більш високе і тривале збільшення екстрацелюлярного дофаміну і NA, у порівнянні з їхніми RLA аналогами як в апетитній, так і в консервативній фазі сексуальної поведінки. Більш того, щури-статеві особи обох римських ліній виявляли більш високе і тривале збільшення екстрацелюлярного дофаміну і NA в mPFC в обох фазах сексуальної поведінки порівняно з їхніми наївними аналогами. Нарешті, всі вищезазначені відмінності в позаклітинному дофаміні і НС у mPFC були, очевидно, пов'язані з відмінностями в поведінкових параметрах сексуальної поведінки, наприклад, статево досвідчені щури обох римських ліній показали вищу сексуальну мотивацію і кращі копуляторні показники порівняно з їхніми наївними аналогами. і сексуальні наївні і досвідчені щури RHA проявляли вищу сексуальну мотивацію і кращі копуляторні показники у порівнянні зі своїми сексуальними наївними та досвідченими колегами RLA.
Що стосується збільшення екстрацелюлярного дофаміну, знайденого в диализате mPFC під час сексуальної активності, перше збільшення відбулося в першій 15 хв після введення сексуально сприйнятливої жінки в спарювальному апараті, коли самка була недоступна для чоловіка, що зазвичай показує в цьому стані неконтактні ерекції пеніса. Другий і більший приріст позаклітинного дофаміну відбувався протягом першої ХНУМХ хв після того, як жінка стала доступною для копуляції самцям. Цікаво, що тоді, коли в наївному стані позаклітинний дофамін збільшувався переважно в першій частині тесту з доступною жінкою з тенденцією до повернення до базальних значень у другій частині, у статевому стані дофамін залишався підвищеним протягом всього тесту, досягаючи пікових значень після 15 – 45 min копуляції в обох римських лініях щурів.
Крім зазначених вище відмінностей у позаклітинному дофаміну під час різних фаз статевої активності між щурами RHA та RLA в статевому адекватному стані, це дослідження також показує несподівані важливі і дуже значні відмінності в концентрації позаклітинного DOPAC між двома лініями римських щурів до ( , базальні значення) і після введення сприйнятливої жінки в спарювальний апарат. Відповідно, базальні значення позаклітинного DOPAC в диализате mPFC були виявлені у щурів RHA приблизно в 60 разів вище, ніж у щурів RLA. Ця різниця між щурами RHA і RLA спостерігалася і зберігалася у статевих випадках, незважаючи на дуже скромні відмінності в базальних значеннях позаклітинного дофаміну в диализатах mPFC, отриманих з двох римських ліній щурів в цих експериментальних умовах. Взяті разом, ці відмінності можуть вказувати на різний оборот допаміну на рівні mPFC між двома римськими лініями щурів. Відповідно, концентрації DOPAC зазвичай відображають кількість дофаміну, що вивільняється, і потім відбирається дофамінергічними нервовими кінцями і перетворюється в DOPAC моноамінооксидазою (MAO; Carlsson, 1975). Отже, більш високі базальні концентрації DOPAC, виявлені у щурів RHA, свідчать про те, що більш високі кількості дофаміну вивільняються і повторно залучаються дофамінергічними нервовими закінченнями у mPFC щурів RHA, наприклад, більш висока базальна активність мезокортикальної допамінергічної системи щурів RHA порівняно з RLA щури. Більш високий мезокортикальний дофамінергічний тонус у щурів RHA, ніж у щурів RLA, також підтримується вищими концентраціями позаклітинного дофаміну, виявленими під час статевої активності в диализате mPFC як з самих статевозрілих, так і з досвідчених щурів RHA по відношенню до щурів RLA. У зв'язку з цим, доречно нагадати, що більш високий дофамінергічний тон вважається відповідальним, принаймні частково, багатьма різними і навіть протилежними поведінковими ознаками, присутніми у щурів RHA і RLA. Відповідно, щури RHA є активними коператорами, сильно імпульсивними, новизнами і шукачами відчуття і схильні до прийому і зловживань кількома класами наркотичних засобів, а щури RLA - це реактивні коппери, гіперемоційні і схильні до розвитку депресивних симптомів (Zeier et al., 1978; Giorgi et al., 1994, 2003b, 2007; Corda et al., 1997, 2014; Escorihuela et al., 1999; Steimer і Driscoll, 2003; Lecca et al., 2004; Giménez-Llort et al., 2005; Carrasco et al., 2008; Fattore et al., 2009; Moreno et al., 2010; Coppens et al., 2012; Díaz-Morán et al., 2012; Sabariego et al., 2013; Manzo et al., 2014a,b; Oliveras et al., 2015), включаючи більш високу сексуальну мотивацію і кращу копуляторну продуктивність сексуальних наївних і досвідчених щурів RHA проти щурів RHA (Sanna et al., 2015). Більш високе збільшення концентрації позаклітинного дофаміну, знайденого в диализате з mPFC статевозрілої і досвідченої щури RHA, нагадує більш високі концентрації позаклітинного дофаміну в диализате з ядром акумулюючих статевозрілих і досвідчених щурів RHA по відношенню до їх аналогів RLA (Sanna et al., 2015). Проте, при відхиленні від mPFC, базальні рівні позаклітинного дофаміну і DOPAC в диализате з ядром accumbens як статевозрілих, так і досвідчених щурів RHA і RLA істотно не відрізнялися (Sanna et al., 2015). Таким чином, вищий мезолімбічний допамінергічний тонус у ядрах акумулюючих статевозрілих і досвідчених щурів RHA може бути виведений тільки вищими концентраціями позаклітинного дофаміну, знайденими в диализатах з ядра акумулюючих щурів RHA при порівнянні з аналогами RLA в присутності сприйнятливих. жіноча, а не інша базальна допамінергічна активність виявлена більш високими і нижчими базальними рівнями концентрації DOPAC у щурів RHA і щурах RLA, відповідно, як було виявлено в mPFC в цьому дослідженні. Це також відповідає результатам попередніх звітів, які показують, що рівні позаклітинного дофаміну в диализате з mPFC щурів RHA були виявлені вище, ніж у щурів RLA, в той час як подібні рівні дофаміну були знайдені в діалізаті з оболонки nucleus accumbens як щурів RHA, так і RLA у різних експериментальних умовах (D'Angio et al., 1988; Scatton et al., 1988; Willig et al., 1991; Giorgi et al., 2003a, 2007). Незважаючи на вищезазначені відмінності між mPFC і nucleus accumbens, сексуальний досвід, здається, у mPFC призводить до тривалого збільшення тимчасової картини вивільнення дофаміну в обох римських лініях щурів при переході від статевого до неспокійного стану, хоча таке збільшення були зазвичай вище у щурів RHA, ніж щури RLA. Це дещо відрізняється від того, що було знайдено в ядрі accumbens, де сексуальний досвід мав тенденцію зміщувати екстрацелюлярні збільшення допаміну до першої частини копуляторного тесту у щурів RHA, а збільшував його протягом усього тесту копуляції у щурів RLA (Sanna et al., 2015).
Наскільки нам відомо, це дослідження також вперше показує, що не тільки позаклітинний дофамін, але і позаклітинна NA зростає під час сексуальної активності в диализате з mPFC щурів RHA та RLA, як було виявлено з дофаміном, але також і в цьому випадку з відмінності, пов'язані з римською лінією щурів і сексуальним досвідом. Зокрема, при відхиленні від базальних рівнів дофаміну, які були виявлені дуже подібними в діалізаті з mPFC щурів RHA і RLA, базальні рівні позаклітинного NA у діалізаті у mPFC щурів RHA були значно вищими (приблизно 2,5-кратно) ), ніж у щурів RLA. Причини цих відмінностей невідомі, але, як обговорювалося вище для дофаміну і DOPAC, ці результати можуть вказувати на те, що більш високий базальний норадренергічний тонус існує принаймні у корі mPFC щурів RHA порівняно з щурами RLA. Такий більш високий норадренергічний тонус також може бути залучений до різних і часто протилежних поведінкових ознак, наявних у щурів RHA і RLA, як обговорювалося вище для дофаміну. Подальша підтримка більш високого норадренергічного тонусу у щурів RHA, ніж у щурів RLA у mPFC, випливає з результатів, які показують, що відмінності в базальних рівнях позаклітинного NA зберігалися також під час сексуальної активності як у статевих, так і в досвідчених умовах, наприклад, позаклітинні NA рівні збільшилися Діалізат mPFC у щурів RHA при впливі недоступних жінок і протягом усього періоду копуляції значно більше, ніж у щурів RLA, у яких рівні NA збільшилися тільки в першій 15 хв періоду копуляції, після чого рівні NA повернулися до базальних значень . Ці зміни у позаклітинних рівнях NA спостерігалися як у статевому, так і в досвідченому стані, але підвищення рівня NA було більш високим у статево досвідчених щурів RHA та RLA у порівнянні з їхніми сексуально неактивними аналогами. Таким чином, сексуальний досвід, мабуть, індукує подальше потенціювання норадренергічної активності у mPFC щурів RHA і, в меншій мірі, щурів RLA, як вже обговорювалося для дофаміну. Супутні вищі норадренергічні і дофамінергічні тони у mPFC щурів RHA, порівняно з щурами RLA, заслуговують на деякі зауваження. Фактично, повідомлялося, що норадренергічна активність у mPFC може впливати на вивільнення дофаміну через різні механізми (наприклад, див. Carboni et al., 1990, 2006; Gresch et al., 1995; Westernik et al., 1998 і посилання в ньому). Один з них пов'язаний з активністю NET, для якого було показано, що зв'язує дофамін з спорідненістю ще вище до такої для NA у синаптосомах, отриманих з гомогенатів PFC (Хорн, 1973). Якщо це має відбутися в mPFC в природних умовахДопамін, що виділяється в mPFC, може бути відновлений не тільки дофамінергічними, але і норадренергічними нервовими терміналами зі швидкістю, навіть вищою, ніж у NA. Це може призвести, в свою чергу, до збільшення вивільнення допаміну з дофамінергічних терміналів (наприклад, підвищеного допамінергічного тонусу) для компенсації кількості допаміну, видаленого з синаптичної щілини; таким чином, більш високий норадренергічний тонус, знайдений у щурів RHA, може сприяти, принаймні частково, вищому дофамінергічному тону, знайденому у mPFC щурів RHA, у порівнянні з щурами RLA. Необхідні подальші дослідження для перевірки цієї можливості. Примітно, що різниця в норадренергічної активності, виявленої в даному дослідженні, в mPFC між щурами RHA і RLA може також відігравати певну роль у їх різній реакції на лікування антидепресантами. Насправді, щури RHA вважаються моделлю стійкості і RLA щури є моделлю вразливості до депресії. Відповідно, гостре або хронічне лікування антидепресантами, такими як дезипрамін, флуоксетин і хлориміпрамін, значно покращило поведінкові реакції при випробуванні на примусове плавання у щурів RLA, не впливаючи на відповіді щурів RHA (Piras et al., 2010, 2014).
Нарешті, це дослідження підтверджує та поширює попередні результати, які показують, що mPFC бере участь у контролі за статевою поведінкою, хоча точна роль цієї області мозку в сексуальній поведінці далека від ясного (Fernández-Guasti et al., 1994; Agmo і Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles et al., 1998, 2007; Kakeyama et al., 2003; Balfour et al., 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011). Нещодавно було показано селективне випалювання клітин у mPFC під час наближення поведінки самців щура до недоступної статево-сприйнятливої жінки з використанням парадигми запису однієї клітини.Febo, 2011). Цікаво, що нейрони, які не відповіли під час першої експозиції чоловіка недоступній сприйнятливій жінці, стають активними під час другої експозиції, як якщо б попередній досвід був здатний викликати пластичні зміни, що призводять до зміни поведінкових реакцій. Тим не менше, виявлено, що ураження іботенічною кислотою mPFC не здатні змінювати сексуальну поведінку щурів-самців статево сприйнятливими жінками, а також вираженням сприятливого місця для сексуальної винагороди. Однак здатність формувати обумовлену аверсію до статевої активності в парі з аверсивними стимулами була виявлена повністю скасованою у цих уражених тварин (Davis et al., 2010). Це говорить про те, що активація mPFC під час сексуальної поведінки відіграє певну роль у інтеграції зовнішньої та внутрішньої інформації для виконання та контролю цілеспрямованої поведінки, а не у вираженні вроджених реакцій на природні підсилювачі (див. Goto і Grace, 2005). Більш важливе для цієї роботи декілька досліджень показали, що ділянки головного мозку, що беруть участь у сексуальній мотивації та сексуальній поведінці, такі як ядро accumbens, вентральна область тегментальної, медіальній зоні преоптики, ядро лімбистого ядра, базолатеральна мигдалина та паркоцелюлярне субпарафакулярне ядро таламуса отримують входи з mPFC під час сексуальної активності (див Balfour et al., 2006). Багато з цих областей мозку беруть участь у складному нейронному ланцюзі, який бере участь у контролі за сексуальною поведінкою, від сексуальної мотивації та винагороди до сексуальної діяльності. Ця схема включає окситоцинергічні нейрони, які походять з PVN гіпоталамуса (PVN) і проектуються до вентральної тегментальної області, nucleus accumbens, гіпокампу, мигдалині, PFC, ядра ліжка Stria terminalis, довгастого мозку і спинного мозку, мезолімбічної / мезокортикальної допамінергічної нейрони, що проектуються з вентральної тегментальної ділянки до ядра accumbens і PFC, вгіпоталамічні допамінергічні нейрони і глутаматергічні нейрони, які беруть участь на локальному і системному рівні в декількох областях схеми (див. Меліс і Арджолас, 1995, 2011; Melis et al., 2003, 2007, 2009, 2010; Succu et al., 2007, 2008, 2011). Таким чином, збільшення позаклітинного дофаміну (але також NA) в диализатах з mPFC, що спостерігається під час сексуальної активності, може вказувати на те, що mPFC є активною частиною цієї схеми, тобто мезокортикальні дофамінергічні нейрони беруть участь у контролі за статевою активністю разом з норадренергічними, мезолімбічними допамінергічними, окситоцинергічними і глутаматергічними нейронами, хоча це не є строго необхідним для вираження сексуальної поведінки (Davis et al., 2010). У відповідності з цією гіпотезою, це дослідження показує, що відмінності у вивільненнях дофаміну та NA у mPFC під час сексуальної активності відбувалися одночасно з відмінностями в кількості неконтактних ерекцій статевого члена та змінах параметрів копуляції, з більш високим вивільненням дофаміну і NA протягом фази апетиту та континентації, пов'язані з вищою сексуальною мотивацією та кращими копуляторними діями як у римських лініях щурів, так і в сексуальних наївних умовах.
Вищенаведене обговорення базується, головним чином, на відомому збільшенні позаклітинного дофаміну, який виникає в ядрах щурів-самців щурів під час сексуальної активності (Pfaus et al., 1990; Pleim et al., 1990; Pfaus і Phillips, 1991; Damsma et al., 1992; Wenkstern et al., 1993). Однак дофамін і інші нейротрансмітери можуть вивільнятися в mPFC (і в nucleus accumbens) в інших експериментальних умовах, таких як під час стресу або справлятися з ним (Thierry et al., 1976; Fadda et al., 1978), які не суворо пов'язані з сексуальною поведінкою. Таким чином, можна стверджувати, що збільшення дофаміну виявляється у позаклітинному дофаміну в mPFC (це дослідження) або в nucleus accumbens (Sanna et al., 2015), більше пов'язана зі стресом, або до співпраці з наявністю та / або взаємодією з сексуально сприйнятливою жінкою, а не з сексуальною активністю. Це може мати певну актуальність у щурів RHA та RLA, які демонструють різний стиль стимуляції та різну активацію гіпоталамо-гіпофізарно-наднирної осі у відповідь на аверсивні умови (щури RLA показують активацію цієї осі набагато вище, ніж у щурів RHA; Carrasco et al., 2008; Díaz-Morán et al., 2012). Це також підвищує ймовірність того, що гормони стресу (тобто кортикостероїди) можуть впливати на дофамінергічну активність у mPFC та інших областях мозку. Дійсно, щури RLA є реакційноздатними коперами і показують гіперемоційну поведінку, що характеризується гіпомотильністю і заморожуванням, тоді як щури RHA показують проактивну поведінку, спрямовану на отримання контролю над стресором (Driscoll і Bättig, 1982; Willig et al., 1991; Escorihuela et al., 1999; Steimer і Driscoll, 2003; Giorgi et al., 2007). Хоча неможливо повністю виключити, що це може траплятися у сексуально наївних щурів (які раніше ніколи не взаємодіяли з сприйнятливою самкою) та зокрема у сексуально наївних щурів RLA (див. Вище), це малоймовірно. В наших експериментальних умовах робиться все можливе, щоб уникнути будь-якого стресу (див. Розділ «Матеріали та методи»), а експерименти організовуються таким чином, що самці не взаємодіють відразу з самкою, а лише через 30 хв впливу на неї , під час якого вимірюване збільшення дофаміну набагато нижче, ніж те, що відбувається під час статевої взаємодії. Якщо підвищення рівня дофаміну, виявлене в mPFC (і в nucleus accumbens) римських щурів (сексуально нерозвинені RHA та RLA щури мають базальний рівень позаклітинного дофаміну в діалізаті з mPFC та nucleus accumbens, дуже схожих) були справді вторинними щодо стрес через або боротьбу з присутністю / взаємодією з жінкою: (i) підвищення дофаміну було б виявлено набагато вищим у перший період часу, коли чоловіки потрапляють у присутність жінки, а не пізніше, як виявлено під час копуляції; та (ii) збільшення дофаміну, як очікується, зникне або сильно зменшиться після придбання стабільної сексуальної активності, тобто коли процеси навчання (включаючи подолання новизни) будуть завершені і відіграють лише незначну роль у здійсненні сексуальної діяльності активність, тоді як на відміну від цього вона продовжує відбуватися при повторній сексуальній активності. Іншими словами, позаклітинний дофамін у mPFC (і в nucleus accumbens) завжди збільшується під час копуляції, навіть після вивчення сексуальної поведінки. Потрібні подальші дослідження для з'ясування функціональної ролі цього збільшення активності дофаміну в mPFC (і в nucleus accumbens; див. Розділ "Вступ та посилання").
На закінчення, це дослідження вперше показує, що присутність, і навіть більше, взаємодія з сексуально сприйнятливою жінкою призводить до збільшення позаклітинного дофаміну і NA в диализате з mPFC статевозрілої (ніколи не піддається сексуальним подразникам). ) та досвідчені статевим шляхом (які пройшли п'ять попередніх тестів копуляції та показали стабільні статеві показники) щурів RHA та RLA, які демонструють різний стиль стимуляції у відповідь на аверсивні стани та різні закономірності копулятивної поведінки з сексуально сприйнятливою жінкою. Відповідно, концентрації дофаміну і NA виявилися вищими в діалізаті від статевозрілих і досвідчених щурів RHA у порівнянні з їхніми аналогами RLA як в передчасному, так і в кінцевому періодах сексуальної активності. Ці результати можуть бути частково обумовлені дофамінергічним та норадренергічним тоном у mPFC щурів RHA вище, ніж у щурів RLA, як повідомляється для дофаміну в nucleus accumbens (Sanna et al., 2015). Більш того, позаклітинні дофамін mPFC та NA також були виявлені вище у досвідчених RHA та у меншій мірі у щурів RLA у порівнянні з їхніми сексуальними наївними аналогами в базальних умовах, а також під час апетитної та консервативної фаз сексуальної поведінки. Це свідчить про те, що сексуальний досвід індукує пластичні процеси, які додатково потенціюють нейротрансмісію дофаміну і NA в mPFC, як встановлено для допаміну в ядрі accumbens обох римських ліній щурів (Sanna et al., 2015). Наведені вище відмінності в допамінергічній і норадренергічній нейротрансмісії в mPFC двох римських ліній можуть грати роль не тільки в різних моделях сексуальної поведінки, але і в різних і часто протилежних поведінкових ознаках, що характеризують ці дві лінії щурів.
Внески автора
FS, AA, MRM, OG та MGC розробили проект. FS і JB розробили, провели та проаналізували дані від сексуальної поведінки та експериментів з мікродіалізу. MAP, OG і MGC відбирали і розводять римських щурів. FS, AA, MRM, OG і MGC контролювали дослідження. Рукопис написали FS, AA, MRM, OG і MGC. Всі автори обговорювали результати та коментували рукопис.
Фінансування
Ця робота була частково підтримана грантами від італійського міністерства університету та досліджень (MIUR) (наукова незалежність молодих дослідників, SIR 2014, код № RBSI14IUX7) до FS, з Автономної області Сардинія (ARS) (LR 7 / 2007) “Просування наукових досліджень та технологічних інновацій у Сардинії”, Європейський соціальний фонд, 2007 – 2013, проектний код № CRP-59842) до OG, а також з університету Кальярі до AA, MRM, FS, OG та MGC.
Заява про конфлікт інтересів
Автори заявляють, що дослідження проводилося за відсутності будь-яких комерційних або фінансових відносин, які могли б бути витлумачені як потенційний конфлікт інтересів.
Подяки
Барбара Тувері, Університет Кальярі, для житла та догляду за тваринами.
посилання
Афонсо, В. М., Сісон, М., Лович В., Флемінг А.С. (2007). Медіальні ураження префронтальної кори у самки пацюки впливають на статеву та материнську поведінку та їх послідовну організацію. Behav. Neurosci. 121, 515 – 526. doi: 10.1037 / 0735-7044.121.3.515
Agmo, A., і Villalpando, A. (1995). Центральні нервові стимулятори полегшують сексуальну поведінку у щурів-самців з медіальними ураженнями префронтальної кори. Мозок Рес. 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A., Villalpando, A., Picker, Z., і Fernández, H. (1995). Поразки медіальної префронтальної кори і сексуальної поведінки у самців щура. Мозок Рес. 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A., і Melis, MR (1995). Нейромодуляція ерекції статевого члена: огляд ролі нейротрансмітерів і нейропептидів. Prog. Neurobiol. 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A., і Melis, MR (2005). Центральний контроль ерекції статевого члена: роль паравентрикулярного ядра гіпоталамуса. Prog. Neurobiol. 76, 1 – 21. doi: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A., і Melis, MR (2013). Нейропептиди і центральний контроль сексуальної поведінки від минулого до сьогодення: огляд. Prog. Neurobiol. 108, 80 – 107. doi: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L., і Coolen, LM (2006). Потенційний внесок еферентів з медіальної префронтальної кори до нервової активації після сексуальної поведінки у самців щура. Неврологія 137, 1259 – 1276. doi: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L., і Coolen, LM (2004). Сексуальне поведінка і пов'язані з сексом екологічні сигнали активізують мезолімбічну систему у самців щурів. Нейропсіхофармакологіі 29, 718 – 730. doi: 10.1038 / sj.npp.1300350
Beloate, LN, Omrani, A., Adan, RA, Webb, IC, і Coolen, LM (2016). Активація дофамінової клітини вентральної області під час сексуальної поведінки самців щурів регулює нейропластичність і перехресну сенсибілізацію d-амфетаміну після статевого стримання. J. Neurosci. 36, 9949 – 9961. doi: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K., Robinson, T. (1998). У чому полягає роль допаміну в нагороді: гедонічний вплив, нагорода навчання або стимулююче відзначення? Brain Res. Rev. 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965). Відбір для високих показників і низькі показники уникнення кондиціонування у щурів. Anim. Behav. 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL, і Bignami, G. (1965). Корелятивні ефекти психогенетичного відбору: дослідження римських високих і низьких штамів уникнення щурів. Behav. Res. Ther. 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E., Silvagni, A., Vacca, C., і Di Chiara, G. (2006). Кумулятивний ефект блокади норадреналіну і дофаміну на позаклітинний дофамін збільшується в оболонці nucleus accumbens, ядрі щілини stria terminalis і префронтальній корі. J. Neurochem. 96, 473 – 481. doi: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E., Tanda, GL, Frau, R., і Di Chiara, G. (1990). Блокада норадреналінового носія підвищує концентрацію позаклітинного дофаміну в префронтальній корі: свідчить про те, що дофамін розкривається в природних умовах за допомогою норадренергічних терміналів. J. Neurochem. 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975). «Рецепторно-опосередкований контроль метаболізму дофаміну», в До і після синаптичних рецепторів, ред. Е. Усдін і В. Е. Банні (Нью-Йорк, Нью-Йорк: Марсель Декер), 49 – 65.
Carrasco, J., Marquez, C., Nadal, R., Tobeña, A., Fernández-Teruel, A., і Armario, A. (2008). Характеристика центральних і периферичних компонентів гіпоталамусно-гіпофізарно-надниркової осі в інбредних ромських штамах щурів. Психоневроендокринологія 33, 437 – 445. doi: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T., і Koolhaas, JM (2012). Імпульсивність та агресивна поведінка у римських щурів з високим та низьким рівнем уникнення: базові відмінності та зміни соціального стресу підлітків. Physiol. Behav. 105, 1156 – 1160. doi: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D., Piras, G., Di Chiara, G., і Giorgi, O. (1997). Біохімічні показники дофамінергічної і ГАМКергіческій нейротрансмісії в ЦНС римського високого уникнення і римського низького уникнення щурів. Behav. Genet. 27, 527 – 536. doi: 10.1023 / A: 1021452814574
Corda, MG, Piras, G., Piludu, MA, і Giorgi, O. (2014). Диференціальні ефекти добровільного споживання етанолу на вихід дофаміну в оболонці nucleus accumbens римських щурів з високим і низьким рівнем уникнення: дослідження поведінки і мікродіалізу мозку. World J. Neurosci. 4, 279 – 292. doi: 10.4236 / wjns.2014.43031
Dahlstrom, A., і Fuxe, K. (1964). Докази існування моноамінсодержащих нейронів в центральній нервовій системі. I. Демонстрація моноамінів в клітинній тілі нейронів стовбура мозку. Acta Physiol. Scand. Suppl. 62, 1-54.
Damsma, G., Pfaus, J., Wenkstern, D., Phillips, A., і Fibiger, H. (1992). Статева поведінка збільшує передачу допаміну в nucleus accumbens і striatum самців щурів: порівняння з новизною і локомоції. Behav. Neurosci. 106, 181 – 191. doi: 10.1037 / 0735-7044.106.1.181
D'Angio, M., Serrano, A., Driscoll, P., і Scatton, B. (1988). Стресові екологічні подразники збільшують рівні позаклітинного DOPAC у префронтальній корі гіпоемоційного (римського високого уникнення), але не гіперемоційного (римського) низького рівня уникнення щурів. a в природних умовах вольтамперометрическое дослідження. Мозок Рес. 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M., Di Sebastiano, AR, Brown, JL, Lehman, MN, і Coolen, LM (2010). Ураження медіальної префронтальної кори викликають неадаптивну сексуальну поведінку у самців щурів. Biol. Психіатрія 67, 1199 – 1204. doi: 10.1016 / j.biopsych.2009.12.029
Díaz-Morán, S., Palència, M., Mont-Cardona, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., et al. (2012). Стиль боротьби та відповіді на гормони стресу у генетично гетерогенних щурів: порівняння з романськими штамами щурів. Бехав Мозок Рес. 228, 203 – 210. doi: 10.1016 / j.bbr.2011.12.002
Driscoll, P., і Bättig, K. (1982). "Поведінкові, емоційні та нейрохімічні профілі щурів, вибраних для екстремальних відмінностей в активній, двосторонній ефективності уникнення", Генетика головного мозку, вид. I. Lieblich (Амстердам: Elsevier), 95 – 123.
Dunlop, BW і Nemeroff, CB (2007). Роль дофаміну в патофізіології депресії. Арка. Психіатрія 64, 327 – 337. doi: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A., Gil, L., Aguilar, R., Tobeña, A., і Driscoll, P. (1999). Інбредні римські щури з високим і низьким рівнем уникнення: відмінності в поведінці тривоги, пошуках новизни та шаттл-боксах. Physiol. Behav. 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990). Сексуальна мотивація: нейронний і поведінковий аналіз механізмів, що лежать в основі апетитних і копуляторних відповідей самців щурів. Неврологія. БіоБехав. Rev. 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ і Robbins, TW (2005). Нейронні системи підкріплення для наркоманії: від дій до звичок до примусу. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fadda, F., Argiolas, A., Melis, MR, Tissari, AH, Onali, PL та Gessa, GL (1978). Стрес-індуковане підвищення рівня 3,4-дигідроксифенилуксусной кислоти (DOPAC) в корі головного мозку і n.accumbens: розворот діазепамом. Наук про життя 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L., Piras, G., Corda, MG, і Giorgi, O. (2009). Римські лінії з високим і низьким рівнем уникнення щурів відрізняються придбанням, підтримкою, вимиранням та відновленням внутрішньовенного самоврядування кокаїну. Нейропсіхофармакологіі 34, 1091 – 1101. doi: 10.1038 / npp.2008.43
Febo, M. (2011). Випадки префронтальних клітин у щурів-самців під час підходу до статево сприйнятливих жінок: взаємодія з кокаїном. Синапс 65, 271 – 277. doi: 10.1002 / syn.20843
Fernández-Guasti, A., Omaña-Zapata, I., Luján, M., і Condés-Lara, M. (1994). Дії лігатури сідничного нерва на сексуальну поведінку статево досвідчених і недосвідчених щурів-самців: наслідки деконтації фронтального полюса. Physiol. Behav. 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobeña, A., і Escorihuela, RM (2002). Незмінні ефекти збагачення навколишнього середовища на пошуки новизни, прийом сахарину та етанолу в двох лініях щурів (RHA / Verh та RLA / Verh), що відрізняються в поведінці, що вимагає стимулів. Фармакол. Біохімія. Бехав 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L., Cañete, T., Guitart-Masip, M., Fernández-Teruel, A., і Tobeña, A. (2005). Два відмінних індукованих апоморфіном фенотипів у римських щурів з високим і низьким рівнем уникнення. Physiol. Behav. 86, 458 – 466. doi: 10.1016 / j.physbeh.2005.07.021
Giorgi, O., Lecca, D., Piras, G., і Corda, MG (2005). Диференціальна активація вивільнення дофаміну в ядрі і оболонці nucleus accumbens після гострих або повторних ін'єкцій амфетаміну: порівняльне дослідження римських ліній високої та низької щільності. Неврологія 135, 987 – 998. doi: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O., Lecca, D., Piras, G., Driscoll, P., і Corda, MG (2003a). Дисоціація між мезокортикальним вивільненням допаміну і поведінкою, пов'язаною з страхом, у двох психогенетично відібраних лініях щурів, які відрізняються в стратегіях подолання аверсивних станів. Євро. J. Neurosci. 17, 2716 – 2726. doi: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O., Piras, G., Lecca, D., Hansson, S., Driscoll, P., і Corda, MG (2003b). Диференціальні нейрохімічні властивості центральної серотонінергічної передачі у римських щурів високого та низького рівня. J. Neurochem. 86, 422 – 431. doi: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O., Orlandi, M., Escorihuela, RM, Driscoll, P., Lecca, D., і Corda, MG (1994). ГАМКергічна і допамінергічна передача в мозку римських високих ухилень і римських щурів з низьким рівнем уникнення. Мозок Рес. 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O., Piras, G., і Corda, MG (2007). Психогенетично відібрані римські лінії з високим і низьким рівнем уникнення щурів: модель для вивчення індивідуальної вразливості до наркоманії. Неврологія. БіоБехав. Rev. 31, 148 – 163. doi: 10.1016 / j.neubiorev.2006.07.008
Goto, Y., і Грейс, AA (2005). Допамінергічна модуляція лімбічного і кортикального приводу ядра акумулює цілеспрямовану поведінку. Nat. Neurosci. 8, 805 – 812. doi: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ, і Finlay, JM (1995). Локальний вплив ендогенного норадреналіну на позаклітинний дофамін в медіальній префронтальній корі щура. J. Neurochem. 65, 111 – 116. doi: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, М., Guevara, MA, Cervantes, M., Morali, G., і Corsi-Cabrera, M. (1998). Характерні смуги частот кортико-лобової ЕЕГ під час статевої взаємодії самця щура в результаті факторного аналізу. J. Фізіологія 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M., Prieto-Beracoechea, CA, Arteaga-Silva, М., і Guevara, MA (2007). Різна функціональність медіальної та орбітальної префронтальної кори під час статево-мотивованого завдання у щурів. Physiol. Behav. 90, 450 – 458. doi: 10.1016 / j.physbeh.2006.10.006
Horn, AS (1973). Структурно-активні зв'язки для інгібування поглинання катехоламіну в синаптосоми з норадренергічних і дофамінергічних нейронів у гомогенатах мозку щурів. Бр. J. Pharmacol. 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Халл, Е. М., Du, J., Lorrain, DS, і Matuszewich, L. (1995). Позаклітинний дофамін в медіальній преоптической області: наслідки для сексуальної мотивації і гормонального контролю копуляції. J. Neurosci. 15, 7465-7471.
Hull, EM, Lorrain, DS, Du, J., Matuszewich, L., Lumley, LA, Putnam, SK, et al. (1999). Взаємодія гормональних нейромедіаторів у контролі за сексуальною поведінкою. Бехав Мозок Рес. 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Hull, EM, Weber, MS, Eaton, RC, Dua, R., Markowski, VP, Lumley, L., et al. (1991). Рецептори дофаміну в вентральній тегментальной області впливають на рухові, але не мотиваційні або рефлекторні компоненти копуляції у самців щурів. Мозок Рес. 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S., і Panksepp, J. (1999). Роль nucleus accumbens дофаміну в мотивованій поведінці: уніфікуюча інтерпретація зі спеціальним посиланням на пошук винагороди. Brain Res. Rev. 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, М., Sone, H., Miyabara, Y., і Tohyama, C. (2003). Перинатальне вплив 2,3,7,8-тетрахлордібензо-p-діоксином змінює залежну від активності експресію мРНК BDNF у статевій поведінці неокортексу і самців щурів у зрілому віці. Нейротоксикологія 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D., Piras, G., Driscoll, P., Giorgi, O., і Corda, MG (2004). Диференціальна активація виходу дофаміну в оболонці і ядрі nucleus accumbens пов'язана з руховими реакціями на наркотичні засоби: дослідження діалізу мозку у римських щурів високого та низького рівня. Нейрофармакологія 46, 688 – 699. doi: 10.1016 / j.neuropharm.2003.11.011
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Donaire, R., Sabariego, M., Fernández-Teruel, A., et al. (2014a). Зв'язок між перевагою етанолу та пошуком новинки. Physiol. Behav. 133, 53 – 60. doi: 10.1016 / j.physbeh.2014.05.003
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A., Papini, MR, і Torres, C. (2014b). Анти-тривожна самолікування, індукована стимулюючими втратами у щурів. Physiol. Behav. 123, 86 – 92. doi: 10.1016 / j.physbeh.2013.10.002
Masana, M., Bortolozzi, A., і Artigas, F. (2011). Селективне поширення мезокортикальної допамінергічної передачі норадренергічними препаратами: терапевтичні можливості при шизофренії. Int. J. Neuropsychopharmacol. 14, 53 – 68. doi: 10.1017 / s1461145710000908
Meisel, RL і Sachs, BD (1994). "Фізіологія чоловічої сексуальної поведінки" в Росії Фізіологія репродукції, Vol. 2, 2nd Edn., Eds E. Knobil та J. Neil (Нью-Йорк, Нью-Йорк: Raven Press), 3 – 96.
Melis, MR, і Argiolas, A. (1995). Допамін і сексуальна поведінка. Неврологія. БіоБехав. Rev. 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR, і Argiolas, A. (2011). Центральний контроль ерекції статевого члена: повторне відвідування ролі окситоцину та його взаємодії з дофаміном і глутаміновою кислотою у самців щурів. Неврологія. БіоБехав. Rev. 35, 939 – 955. doi: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, Т., Cocco, C., Succu, S., Sanna, F., Pillolla, G., et al. (2007). Окситоцин, що вводиться в вентральну тегментальную область, індукує ерекцію статевого члена і збільшує позаклітинний допамін в nucleus accumbens і паравентрикулярном ядрі гіпоталамуса самців щурів. Євро. J. Neurosci. 26, 1026 – 1035. doi: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., et al. (2010). Окситоцин індукує ерекцію статевого члена при введенні в вентральний сублікум: роль оксиду азоту та глутамінової кислоти. Нейрофармакологія 58, 1153 – 1160. doi: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S., Mascia, MS, Cortis, L., і Argiolas, A. (2003). Надклітинний дофамін збільшується в паравентрикулярном ядрі щурів-самців під час сексуальної активності. Євро. J. Neurosci. 17, 1266 – 1272. doi: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S., Sanna, F., Boi, A., і Argiolas, A. (2009). Окситоцин, що вводиться в вентральний субикулум або постромедіальне кортикальне ядро мигдалини, індукує ерекцію статевого члена і підвищує рівні позаклітинного дофаміну в ядрі accumbens самців щурів. Євро. J. Neurosci. 30, 1349 – 1357. doi: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M., Cardona, D., Gómez, MJ, Sánchez-Santed, F., Tobeña, A., Fernández-Teruel, A., et al. (2010). Характеристика імпульсивності у римських штамів щурів з високим і низьким рівнем уникнення: поведінкові та нейрохімічні відмінності. Нейропсіхофармакологіі 35, 1198 – 1208. doi: 10.1038 / npp.2009.224
Oliveras, I., Río-Álamos, C., Cañete, Т., Blázquez, G., Martínez-Membrives, E., Giorgi, O., et al. (2015). Інгібування препульсу прогнозує продуктивність просторової робочої пам'яті у інбредних щурів з високим і низьким рівнем уникнення, а також у генетично гетерогенних щурах NIH-HS: значимість для вивчення попередніх і когнітивних аномалій шизофренії. Фронт. Behav. Neurosci. 9: 213. doi: 10.3389 / fnbeh.2015.00213
Paxinos, G., і Watson, C. (2004). Мозок щура в стереотаксичних координатах, 4th Edn, Нью-Йорк: Академічна преса.
Pfaus, JG (2010). Допамін: допомагає чоловікам копулювати щонайменше 200 мільйонів років: теоретичний коментар про Kleitz-Nelson et al. (2010). Behav. Neurosci. 124, 877 – 880. doi: 10.1037 / a0021823
Pfaus, JG, Damsma, G., Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG, et al. (1990). Статева поведінка посилює центральну передачу допаміну у самців щура. Мозок Рес. 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG, і Everitt, BJ (1995). «Психофармакологія сексуальної поведінки» в Росії Психофармакологія: Четверте покоління прогресу, ред. Ф. Е. Блум і DJ Купфер (Нью-Йорк, Нью-Йорк: Raven Press), 742 – 758.
Pfaus, JG, і Phillips, AG (1991). Роль дофаміну в очікувальних і консервативних аспектах сексуальної поведінки у самців щурів. Behav. Neurosci. 105, 727 – 747. doi: 10.1037 / 0735-7044.105.5.727
Piras, G., Giorgi, O., і Corda, MG (2010). Вплив антидепресантів на працездатність у вимушеному плавальному тесті двох психогенетично відібраних ліній щурів, що відрізняються стратегіями копінгу від аверсивних станів. Психофармакология 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G., Piludu, MA, Giorgi, O., і Corda, MG (2014). Вплив хронічних антидепресантних методів лікування в передбачуваній генетичній моделі вразливості (римські щури з низьким рівнем уникнення) і стійкість (римські щури з високим рівнем уникнення) до депресії, викликаної стресом. Психофармакология 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V., Mouzon, K., Nestler, EJ, Lehman, MN, et al. (2010). ΔFosB в nucleus accumbens має вирішальне значення для посилення ефекту сексуальної винагороди. Гени Brain Behav. 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J., Van, S., Frohmader, KS, et al. (2014). Ендогенна індукована опіоїдами нейропластичність дофамінергічних нейронів в вентральній тегментальній ділянці впливає на природну і опіатичну винагороду. J. Neurosci. 34, 8825 – 8836. doi: 10.1523 / JNEUROSCI.0133-14.2014
Глечики, KK, Vialou, V., Nestler, EJ, Laviolette, SR, Lehman, MN, і Coolen, LM (2013). Натуральні і лікарські винагороди діють на загальні механізми нейронної пластичності з ΔFosB як ключовим медіатором. J. Neurosci. 33, 3434 – 3442. doi: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E., Matochik, J., Barfield, R., і Auerbach, S. (1990). Кореляція вивільнення дофаміну в nucleus accumbens з сексуальною поведінкою чоловіків у щурів. Мозок Рес. 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW, і Arnsten, AFT (2009). Нейропсихофармакологія фронто-виконавчої функції: моноамінергічна модуляція. Ann. Neurosci. 32, 267 – 287. doi: 10.1146 / annurev.neuro.051508.135535
Sabariego, M., Morón, I., Gómez, MJ, Donaire, R., Tobeña, A., Fernández-Teruel, A., et al. (2013). Втрата стимулів і експресія гена гіпокампа в інбредних римських щурах з уникненням високого (RHA-I) і римського низького рівня (RLA-I). Бехав Мозок Рес. 257, 62 – 70. doi: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000). Контекстуальні підходи до фізіології та класифікації еректильної функції, еректильної дисфункції та статевого збудження. Неврологія. БіоБехав. Rev. 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K., Citro, JK, Daniels, SB, і Natoli, JH (1994). Безконтактна стимуляція у жінок самки викликає ерекцію статевого члена у щурів. Physiol. Behav. 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD, і Barfield, RJ (1976). Функціональний аналіз чоловічої копулятивної поведінки у щурів. Adv. Шпилька. Behav. 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M., Yohn, SE, López-Cruz, L., San Miguel, N., і Correa, M. (2016). Мезолімбічний дофамін і регулювання мотивованої поведінки. Curr. Угору. Behav. Neurosci. 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F., Corda, MG, Melis, MR, Piludu, MA, Giorgi, O., і Argiolas, A. (2014a). У самців щурів з високим і низьким рівнем уникнення виявляються різні моделі копуляторного поведінки: порівняння з щурами sprague dawley. Physiol. Behav. 127, 27 – 36. doi: 10.1016 / j.physbeh.2014.01.002
Sanna, F., Piludu, MA, Corda, MG, Argiolas, A., Giorgi, O., і Melis, MR (2014b). Допамін бере участь у різних моделях копулятивної поведінки щурів з високим і низьким рівнем уникнення римського роду: дослідження з апоморфіном і галоперидолом. Фармакол. Біохімія. Бехав 124, 211 – 219. doi: 10.1016 / j.pbb.2014.06.012
Sanna, F., Piludu, MA, Corda, MG, Melis, MR, Giorgi, O., і Argiolas, A. (2015). Залучення дофаміну в відмінності в сексуальній поведінці між римськими щурами високого та низького рівня: дослідження внутрішньомозкового мікродіалізу. Бехав Мозок Рес. 281, 177 – 186. doi: 10.1016 / j.bbr.2014.12.009
Scatton, B., D'Angio, M., Driscoll, P., and Serrano, A. (1988). An в природних умовах Вольтамперометрическое дослідження відповіді мезокортикальних і мезоаккумних дофамінергічних нейронів на екологічні подразники у штамів щурів з різними рівнями емоційності. Ann. NY Acad. Sci. 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T., і Driscoll, P. (2003). Дивергентні реакції стресу та стилю стикування у психогенно відібраних римських високих (RHA) та низьких (RLA) уникненнях щурів: поведінкові, нейроендокринні та розвиваючі аспекти. стрес 6, 87 – 100. doi: 10.1080 / 1025389031000111320
Succu, S., Sanna, F., Argiolas, A., і Melis, MR (2011). Окситоцин, який вводиться в вентральний сублікум гіппокампа, індукує ерекцію статевого члена у щурів-самців, збільшуючи глутаматергічну нейротрансмісію в вентральній тегментальной області. Нейрофармакологія 61, 181 – 188. doi: 10.1016 / j.neuropharm.2011.03.026
Succu, S., Sanna, F., Cocco, C., Melis, Т., Boi, A., Ferri, GL, et al. (2008). Окситоцин індукує ерекцію статевого члена при введенні в область вентрального тегментального щура: роль оксиду азоту та циклічного GMP. Євро. J. Neurosci. 28, 813 – 821. doi: 10.1111 / j.1460-9568.2008.06385.x
Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., і Melis, MR (2007). Стимуляція дофамінових рецепторів в паравентрикулярном ядрі гіпоталамуса самців щурів викликає ерекцію статевого члена і підвищує надклітинний дофамін в nucleus accumbens: залучення центрального окситоцину. Нейрофармакологія 52, 1034 – 1043. doi: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G., і Glowinski, J. (1976). Селективна активація мезокортикальної системи ДА стресом. природа 263, 242 – 244. doi: 10.1038 / 263242a0
Wenkstern, D., Pfaus, J., і Fibiger, H. (1993). Передача дофаміну збільшується в ядрах щурів-самців щурів під час їх першого впливу на статево сприйнятливих щурів-самців. Мозок Рес. 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Вестернік, БХК, Де Бур, П., Де Фріз, Дж. Б., Лонг, С. (1998). Антипсихотичні препарати індукують подібні ефекти на вивільнення дофаміну і норадреналіну в медіальній префронтальній корі головного мозку щура. Євро. J. Pharmacol. 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F., M'Harzi, М., Bardelay, C., Viet, D., і Delacour, J. (1991). Римські штами як психогенетична модель для дослідження робочої пам'яті: поведінкові та біохімічні дані. Фармакол. Біохімія. Бехав 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
Ключові слова: сексуальна поведінка, дофамінові, норадреналінові, mPFC, мікродіалогічні, RHA і RLA щури
Цитата: Санна Ф, Братзу Дж., Пілуду М.А., Корда М.Г., Меліс М.Р., Джорджі О. і Арджолас А (2017) допамін, норадреналін і відмінності в статевому поведінці між високим і низьким уникненням риб чоловічої статі. . Фронт. Behav. Neurosci. 11: 108. doi: 10.3389 / fnbeh.2017.00108
Отримано: 22 грудень 2016; Прийнято: 19 травня 2017;
Опубліковано: 07 Червень 2017.