Фронт Behav Neurosci. 2015 травень 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Андреатета М1, Паулі П1.
абстрактний
У класичному кондиціонуванні спочатку нейтральний стимул (умовний стимул, CS) асоціюється з біологічно суттєвою подією (безумовний стимул, УЗД), який може бути болем (неприйнятне кондиціонування) або їжею (апетитне кондиціонування). Після кількох асоціацій CS може ініціювати або захисні, або спонукальні відповіді, відповідно. На відміну від аверсивної обумовленості, апетитна обумовленість рідко досліджується у людей, хоча її значення для нормальної та патологічної поведінки (наприклад, ожиріння, звикання) незаперечне. Це дослідження має на меті перекласти знахідки тварин щодо апетитного кондиціонування для людей, які використовують їжу як США. Тридцять три учасники були розслідувані між 8 і 10 ранку без сніданку, щоб переконатися, що вони відчували голод. Протягом двох фаз придбання одна геометрична фігура (avCS +) передбачала неприємні УС (болісний удар струмом), інша форма (appCS +) - апетитну США (шоколад або солоний крендель відповідно до уподобань учасників), а третя фігура (CS– ) прогнозував ні США. На фазі вимирання ці три фігури плюс нова форма (НОВА) були представлені знову без доставки в США. Оцінки валентності та збудження, а також відповіді на здивування та провідність шкіри (SCR) були зібрані як показники навчання. Ми виявили успішне відверте та апетитне обумовлення. З одного боку, avCS + був оцінений як більш негативний та більш збудливий, ніж CS–, і викликав здригнення та посилення SCR. З іншого боку, appCS + був оцінений більш позитивно, ніж CS–, та викликав ослаблення здригування та більший SCR. Підводячи підсумок, ми успішно підтвердили знахідки тварин у (голодних) людей, продемонструвавши апетитне навчання та нормальне неприйнятне навчання.
Вступ
Прогнозування загрози та їжі має вирішальне значення для виживання будь-якого організму. У класичному кондиціонуванні (Павлов 1927), кию передує неприємна подія, така як легкий хворобливий ураження електричним струмом (відвертий безумовний стимул, США), або апетитна подія, наприклад харчова грануля (апетитна США), кілька разів. Згодом лише цей стимул (тепер він позначений як CS, CS +) здатний викликати відповідно оборонні чи спонукальні відповіді. Перший вид асоціативного навчання називається аверсивним обумовленням, тоді як останній називається апетитним обумовленням.
Хоча прогнозування апетитних подій настільки ж важливо для виживання, як і прогнозування аверсивних подій, апетитна обумовленість надзвичайно менш досліджена у тварин (Bouton and Peck, 1989; Koch et al., 1996; McDannald та ін., 2011, для огляду див Martin-Soelch et al., 2007), а також у людей (Klucken et al., 2009, 2013; Остін і Дука, 2010; Delgado et al., 2011; Леві та Глімчер, 2011). Цей брак досліджень, можливо, пояснюється складністю апетитної парадигми порівняно з відвертою. Наприклад, їжа в якості основного зміцнювача повинна бути доставлена, коли організм голодний, щоб бути корисним (для останнього огляду див. Dickinson і Balleine, 1994; Clark et al., 2012). У людських дослідженнях цю складність було подолано за допомогою грошей (Остін і Дука, 2010; Delgado et al., 2011; Леві та Глімчер, 2011) або еротичні фотографії (Klucken et al., 2009, 2013). Однак нейровізійні дослідження показали, що первинні (тобто закуски чи напої) та вторинні (тобто гроші) підсилювачі активізують деякі загальні ділянки мозку (наприклад, стриатум), але також тягнуть за собою чіткі схеми активації (Delgado et al., 2011; Леві та Глімчер, 2011).
Наскільки нам відомо, лише декілька досліджень щодо кондиціонування людини вивчали дію первинних апетитних підсилювачів, таких як запах (Gottfried et al., 2002), вода (Kumar et al., 2008) або їжу (Prévost et al., 2012). У першому дослідженні Готфрід та ін. (2002) пов'язані нейтральні обличчя (КС) з неприємним, приємним або нейтральним запахом. Цікаво, що вони виявили більшу активацію в орбітофронтальній корі (OFC) та в вентральній смузі у відповідь на апетитний CS + проти неприязного CS +. Вони дійшли висновку, що OFC обробляє значення запаху і бере участь у перенесенні афективного значення з нюхової (US) в зорову (CS) систему. Активація (вентро) смугастої інтерпретації інтерпретується як відображення апетитного CR, що викликається апетитним CS +. У другому дослідженні Kumar et al. (2008) запросив своїх учасників (здорові контролі та пацієнти з великою депресією) до лабораторії рано вранці і попросив утриматися від пиття протягом ночі, щоб переконатися, що вони спрагли перед скануванням. На фрактальних знімках були КС та 0.1 мл води США. Цікаво, що серед інших активацій здорові учасники (але не пацієнти з депресією) виявили більшу активацію в вентральному стриатумі до апетитного CS +, що дозволяє припустити, що цей стимул розглядався як корисний. У третьому дослідженні Prévost та ін. (2012) також представив фрактальні фотографії як CS та солодкі або солоні закуски як США відповідно до уподобань учасників. CS був представлений протягом 6 с, і протягом останньої секунди додатково була представлена картина їжі. Щоразу, коли з’являлася картина з їжею, експериментатор клав шматок їжі в руки учасників, яким дозволялося негайно споживати закуску. На жаль, автори не повідомляли про активацію мозку апетитному CS + під час класичної фази кондиціонування; однак вони спостерігали менше серцевого уповільнення до CS +, що отримав винагороду, порівняно з CS без винагороди, що свідчить про диференціальні ефекти кондиціонування.
Відсутність досліджень, що використовують реакцію враження як показник апетитного кондиціонування, викликає подив, особливо зважаючи на широке його використання в умовах неприємного кондиціонування. Реактивна реакція - це споконвічна та автоматична захисна реакція на раптові, несподівані та сильні відрази (Кох, 1999). Ця захисна реакція опосередковується відносно простим нейронним шляхом, що включає нейрони кохлеарного кореня, каудальне понтінове ядро ретикулярної формації (PnC) та спинальні мотонейрони (Fendt і Fanselow, 1999; Кох, 1999). Дослідження на тваринах виявили, що потенціал стрелів залежить від проекцій від мигдалини до PnC (Fendt і Fanselow, 1999; Кох, 1999), тоді як ослаблення пуску залежить від непошкодженого ядра ядра (NAcc, Кох, 1999). Така модуляція реакції на стрес є корисним неявним заходом для валентності представлених подразників переднього плану. Таким чином, потенціювання вказує на негативну валентність, тоді як послаблення вказує на позитивну валентність, без того, щоб сильно впливати когнітивні процеси (Хамм і Вейке, 2005; Andreatta та ін., 2010). Наскільки нам відомо, лише одне дослідження на тваринах досліджувало апетитне кондиціонування шляхом вимірювання реакцій стресу як залежної міри. Цікаво, що обстежувані щури демонстрували ослаблення стресу до CS + (тобто світла), пов'язаного з доставкою розчину сахарози. Помітно, таке ослаблення було порушено у тварин з ураженнями NAcc, але не у тих, у кого були ураження амігдали, що свідчить про те, що NAcc відіграє особливу роль у виведенні апетитних КР та в ослабленні реакцій на стрес (Koch et al., 1996).
Поточне дослідження, наскільки нам відомо, першим перевело цю апетитну умовну умову на людей, використовуючи первинні підсилювачі як американські, тобто солодкі (шоколадні смарки®) або солона (невелика солона крендель) їжа, і стягувальна модуляція як міра КР. Ми очікували, що appCS + спровокує сильну апетитну КР, що відображається на згасанні стрелів, посиленому SCR та позитивному валентному рейтингу порівняно з іншими стимулами, тобто avCS + та CS–.
Матеріали та методи
Учасниками
Сорок два добровольці прийняли участь у дослідженні та отримали залікові курси. Дев'ять учасників були виключені з аналізу, двоє через технічні проблеми, троє, оскільки вони були закодовані як невідповідачі (середня амплітуда пуску <5 мкВ), і четверо, оскільки вони не виявляли достатньої реакції на страх за умови (мінімум = 2; для подробиці, див. Матеріали та метод). Врешті-решт, в аналізі було враховано 33 учасника (16 чоловіків; середній вік: 22.09 років, SD: 2.84; діапазон: 18 – 29 років). Чотири учасники не були рідними німцями, а шість - лівшами. Один учасник впродовж експерименту не знав про асоціації CS-USs (див. Порядок), але ми вирішили не виключати цього учасника, оскільки його відповіді були нормальними і не впливали на результати.
Матеріали
Безумовні стимули (США)
Були використані два види США. Як противники США, ми застосували легкий болісний ураження електричним струмом на домінантне передпліччя учасників. Електричний удар подавався за допомогою двох електродів діаметром 9 мм та відстані 30 см. Електричний стимул складався з імпульсного стимулу з частотою 50 Гц і тривалістю 200 мс, породженого струмовим стимулятором (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Великобританія, 400 V, максимум 9.99 mA). Інтенсивність ураження електричним струмом визначалася індивідуально за допомогою порогової процедури, описаної раніше (Andreatta та ін., 2010). Коротко, учасники пройшли дві серії зростаючої та низхідної інтенсивності в інтервалах 0.5 mA. Їм доводилося оцінювати кожен стимул у візуальній шкалі, починаючи від 0 (взагалі нічого не відчуваючи) до 10 (дійсно інтенсивний біль), використовуючи 4 як прив'язку до порога (просто помітний біль). Середня інтенсивність електричного подразника становила 2.12 мА (SD = 0.56), і вона була оцінена як болюча (M = 6.45, SD = 1.73). Апетитні США складалися або з шоколаду (Smarties®) або невеликий солоний крендель. Вибір апетитного США залежав від індивідуальних уподобань учасника, про що повідомлялося під час попереднього співбесіди. А саме, учасники мали повідомити, чи їдять вони зазвичай солену чи солодку їжу під час сніданку. Вони також могли вільно вибирати, чи віддають перевагу шоколаду чи солоному кренделю під час самого експерименту. Врешті-решт 22 учасники обрали шоколад та 11 маленький солоний крендель.
Умовні стимули (CS)
Геометричні фігури (синій квадрат, жовтий круг, зелений трикутник, червоний шестикутник) з діагоналлю 8 см були представлені у вигляді CS. Фігури були представлені посеред чорного екрану комп'ютера для 8 s. Одна форма (avCS +) завжди асоціювалася з відразою США (хворобливий ураження електричним струмом), одна форма (appCS +) завжди асоціювалася з апетитними США (шоколад або солоний крендель), одна форма (CS–) ніколи не асоціювалася ні з противником або апетитні US, а четверта форма (NEW) була представлена у фазі вимирання, але не під час фази придбання, щоб забезпечити її нейтральність.
Зонд запуску
Білий шум 103 дБ тривалістю 50 мс був використаний як зонд для здивування. Акустичні подразники подавались бінаурально через навушники і виникали випадково через 4–6 с після початку фігури.
Анкети
До і після експерименту учасники повинні були заповнити німецькі версії Інвентаризації тривоги штату (STAI, Laux та ін., 1981) та графік позитивних негативних впливів (PANAS, Krohne та ін., 1996). STAI - це опис для оцінки ознак учасників та / або стану тривожності та складається з 20 предметів як для ознаки, так і для версій штату. Рівень тривожності учасників до (M = 37.06, SD = 7.80) і після (M = 39.33, SD = 9.16) експеримент суттєво не змінився [t(32) = 1.61, p = 0.117]. Оцінки тривожності в поточній вибірці склали від 20 до 58 (M = 36.6, SD = 8.98), що порівняно з опублікованим нормальним діапазоном дорослих (Laux та ін., 1981). ПАНАС (Krohne та ін., 1996) - показник позитивного та негативного настрою. Особи, які мають високі бали за шкалою позитивних афектів (PAS), схильні до таких емоцій, як ентузіазм, тоді як особи, які мають високі бали за шкалою негативних афектів (NAS), схильні до таких емоцій, як дистрес. Кожен предмет складається з прикметника, і учасники вказують за шкалою від 1 (дуже незначно) до 5 (надзвичайно), наскільки прикметник відображає їхні почуття в той конкретний момент. Істотних відмінностей у негативному впливі учасників в результаті експерименту не виявлено [початок: M = 11.67, SD = 2.29; кінець: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Чомусь учасники значно змінили свій позитивний настрій з самого початку (M = 26.72, SD = 4.70) до кінця (M = 23.66, SD = 7.02) експерименту [t(31) = 3.11, p = 0.004]. Це зниження позитивного настрою учасників могло бути пов’язане з неприємністю парадигми (були представлені болісні ураження електричним струмом, а також неприємний білий шум).
Процедура
Після прибуття до лабораторії учасники зачитали та підписали бланк інформованої згоди, затверджений комітетом з етики кафедри психології Вюрцбурзького університету. Їх не поінформували про надзвичайні ситуації між ЦС та США. Після заповнення анкети електроди були прикріплені та процедура порогового болю була проведена, як описано вище.
Під час фаза звикання, чотири геометричні фігури були представлені двічі з міжпробним інтервалом (ITI), що змінюється між 18 та 25 s (середнє: 21.5 s). У цій фазі не було доставлено жодних зондів США або здивування.
Перед початком етапу придбання подавалось сім баттів білого шуму кожні 7 – 15 с, щоб знизити початкову реактивність пуску.
Наступні два фази придбання були однаковими (мал 1). Кожна фаза придбання складалася з випробувань 24: випробування 8 CS–, випробування 8 avCS + та випробування 8 appCS +. Послідовність CS була псевдовипадковою, єдиним обмеженням було те, що один і той же стимул не міг бути представлений більше двох разів поспіль. Зокрема, avCS + був представлений разом з блискавкою як символом ураження електричним струмом, і хворобливі УЗД були доставлені на компенсації. Програма appCS + була представлена в поєднанні із зображенням Smarties або солоної кренделі, і учасник міг вибрати Смарти або кренделі з банки. CS– був представлений разом із символом заборони, і США не було доставлено. Під час трьох презентацій 8 CS кожного типу, після настання подразника було передано зонд для запуску між 4 та 6 s. Три додаткових зонди для запуску були представлені під час ІТІ, щоб переконатися в їх непередбачуваності та зменшити звикання. ITI, що складається з чорного екрана, змінювався між 18 та 25 s із середнім значенням 21.5 s.
МАЛЮНОК 1
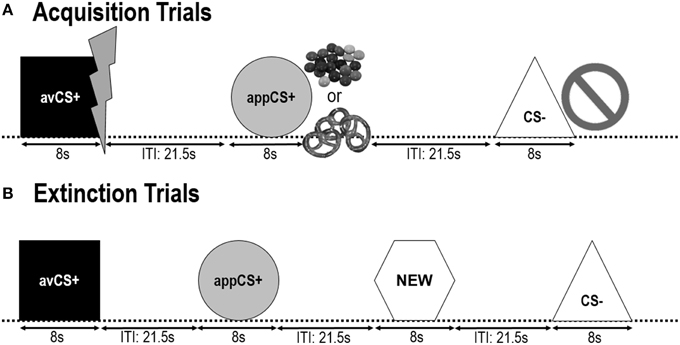
Рисунок 1. Випробування протягом двох фаз придбання (А) та фази вимирання (В). Учасники дізналися, що одна форма (avCS +) передбачила легкий болісний ураження електричним струмом, одна форма (appCS +) передбачила або шматочок шоколаду, або невеликий солоний крендель (відповідно до їх переваг), а третя форма (CS–) не передбачала жодної біологічно яскрава подія. Кожна форма була представлена разом із малюнком, що зображав ураження електричним струмом, шматки / солоний крендель або нічого, що залежить від асоціації зі США. Під час фази вимирання три геометричні фігури були представлені ще раз, але США не були доставлені. Крім того, четверта геометрична форма (NEW) була представлена як нейтральне управління.
Під час фаза вимирання, учасники побачили три геометричні фігури (тобто avCS +, appCS + та CS–) знову плюс нову нейтральну форму (NEW). США не були доставлені, і фігури не були представлені разом із зображеннями спалахів, шоколадних / солоних кренделей або заборон. Кожен стимул був представлений вісім разів у псевдовипадковому порядку (тобто той самий стимул не був представлений більше двох разів поспіль), створюючи випробування 32. Подразники зондового зондування були представлені під час 4 з подання стимулів 8 кожного типу CS. Як і на етапі придбання та звикання, ІТІ варіювався між 18 і 25 s, а додаткові зонди для запуску 4 були поставлені непередбачувано під час ITI.
Після кожної фази учасники оцінювали валентність (приємність) та збудження (збуджувальну) ЦС з візуальними аналоговими шкалами (VAS), що варіюється від 1 до 9. Шкала валентності варіювалася від "1", що вказує на "дуже неприємно" до "9", вказуючи на "дуже приємно"; шкала збудження варіювалася від "1", що вказує на "спокій" до "9", що вказує на "хвилююче". оцінювали після двох фаз придбання та фази вимирання. Учасники побачили геометричну форму для 1 s, і тоді їм довелося вказати, чи пов’язана ця форма з електричним струмом, з шоколадною / солоною кренделями, ні з чим, чи вони не змогли створити ніяких асоціацій. Зокрема, всі учасники (крім одного) знали про надзвичайні ситуації після придбання 2.
Зменшення даних
Фізіологічні відповіді реєстрували за допомогою підсилювача V-Amp 16 та програмного забезпечення Vis-Recorder V-Amp Edition (версія 1.03.0004, BrainProducts Inc., Мюнхен, Німеччина). Були застосовані частота дискретизації 1000 Гц та фільтр 50 Гц. Офлайн-аналізи проводили за допомогою аналізатора мозкового зору (версія 2.0; BrainProducts Inc., Мюнхен, Німеччина).
Страхітна відповідь
Стартову реакцію вимірювали за допомогою електроміографії (ЕМГ) зліва orbicularis oculi м'яз з двома електродами Ag / AgCl 5 мм. Відповідно до вказівок (Blumenthal та ін., 2005), один електрод був розміщений під зіницею, а другий - 1 см бічно. Заземлюючі та опорні електроди були розміщені відповідно на правій та лівій мастоидах. Перед прикріпленням електродів шкіру злегка розтирали та очищали спиртом, щоб зберегти опір нижче 10 kΩ. Електроміографічний сигнал відфільтрований в автономному режимі за допомогою фільтра низького відсічення 28 Гц та фільтра високого відсікання 500 Гц. Потім сигнал EMG був виправлений і застосовано ковзаюче середнє значення 50 мс. Ми використовували 50 ms перед початком запуску зонда як базову лінію (Grillon et al., 2006). Відповіді на зондові враження оцінювали вручну, а випробування з надмірними зрушеннями базової лінії (± 5 мкВ) або артефактами руху були виключені з подальшого аналізу. Відповіді на пуск нижче 5 мкВ кодуються як нуль і враховуються для обчислення величини пуску (Blumenthal та ін., 2005). Загалом, відхилено 10.4% випробувань, і мінімум 2 із відповідей на стрімкість 3 у фазах придбання та 4 з реакцій на стрімкість 8 у фазі вимирання для кожної умови потрібно було тримати учасника в пулі аналізу. З цієї причини чотирьох учасників було виключено. Амплітуда піку була визначена як максимальний пік щодо базової лінії протягом вікна часу 20 – 120 мс після початку зонду пуску. Потім вихідні дані були нормалізовані в рамках суб'єктів, використовуючи z-оцінки, а потім Т-бали з метою зменшення впливу індивідуальної мінливості та кращого виявлення психологічних процесів. Оцінки T були усереднені для кожної умови (avCS +, appCS +, CS–, NEW та ITI). Для того, щоб дослідити потенціал запуску або ослаблення пуску, оцінки для відповідей на стрімкий час ІТІ віднімали від відповідей на спущення для кожної умови.
Відповідь провідності шкіри (SCR)
Відповідь на шкірну провідність (SCR) реєстрували за допомогою двох електродів 5 мм Ag / AgCl, розміщених на долоні домінуючої руки. Гальванічну відповідь відфільтрували в автономному режимі за допомогою фільтра високого відсіку 1 Гц. SCR визначали як різницю (у мкС) між початком відповіді (1 – 3 s після настання стимулу) та піком відповіді (Транель і Дамасіо, 1994; Delgado et al., 2011). Випробування, що містять пускові зонди, не враховувались в аналізі SCR. Відповіді нижче 0.02 μS були кодовані як нуль. П'ять інших учасників були виключені з аналізу SCR, оскільки середній показник SCR був нижчим, ніж 0.02 μS. Дані про сирість шкірної провідності трансформували квадратний корінь з метою нормалізації розподілу, а середні оцінки оцінювались для кожної умови окремо за дві фази придбання (avCS +, appCS +, CS–) та фазу згасання (avCS +, appCS +, CS– та NEW) ).
Статистичний аналіз
Всі дані були проаналізовані за допомогою SPSS для Windows (версія 20.0, SPSS Inc.). Для фізіологічних відповідей було розраховано роздільний багатофакторний аналіз дисперсії (ANOVA) для двох фаз придбання та фази вимирання. Фази придбання ANOVA для стимулюючих факторів мали стимул (avCS +, appCS +, CS–) та фазу (придбання 1, придбання 2) як фактори, що підпадають під предмет. ANOVA для фази вимирання мав лише стимул (avCS +, appCS +, CS–, NEW) як фактор всередині суб'єктів. Рейтинги валентності, збудження та надзвичайних ситуацій були проаналізовані за допомогою окремих ANOVA, що містять стимул факторів всередині суб'єктів (avCS +, appCS +, CS– та NEW) та фази. Цей фактор мав чотири рівні для оцінок валентності та збудження (T1: після фази звикання, T2: після першої фази придбання, T3: після другої фази придбання, T4: після фази вимирання), але три рівні для рейтингів надзвичайних ситуацій (T1: після перша фаза придбання, T2: після другої фази придбання, T3: після фази вимирання).
Для всіх аналізів встановлено рівень альфа (α) на 0.05. Розмір ефекту повідомляється як частковий η2.
результати
Рейтинг валентності та збудження для кожної фази зображено на малюнку 2; відповіді на стрес і SCR зображені на малюнку 3.
МАЛЮНОК 2
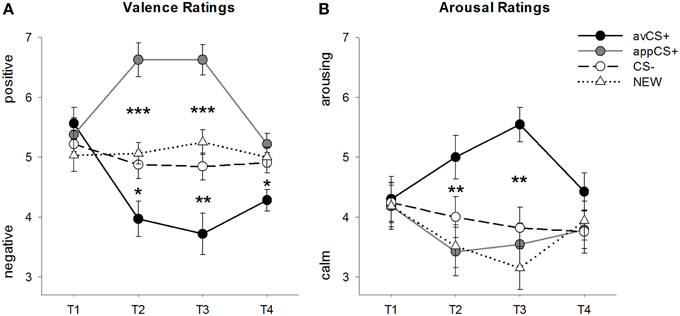
Рисунок 2. Рейтинг валентності (A) та збудження (B). Рядки (зі стандартними помилками) відображають рейтинги після фази звикання (T1), придбання 1 (T2), придбання 2 (T3) та фази вимирання (T4). Аверсивний CS + (чорна суцільна лінія) отримав негативну валентність та високе збудження після двох фаз придбання порівняно з CS– (чорна пунктирна лінія) та НОВА (чорна пунктирна лінія). Важливо, що апетитний CS + (сіра суцільна лінія) набув позитивної валентності порівняно з CS– та NEW. *p <0.05, **p > 0.01, ***p < 0.001.
МАЛЮНОК 3
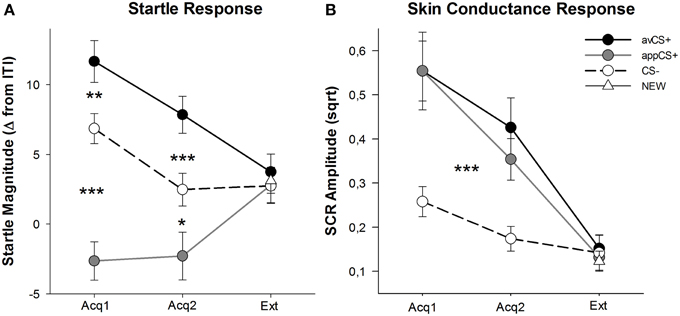
Рисунок 3. Стартові реакції (A) та шкірна провідність (B) (зі стандартними помилками) під час першої фази придбання (Acq1), другої фази придбання (Acq2) та фази вимирання (Ext). Стартові відповіді були суттєво потенційовані до аверсивного CS + (чорна суцільна лінія) та значно ослаблені до апетитного CS + (сіра суцільна лінія) порівняно з CS– (чорна пунктирна лінія) під час фаз придбання. SCR був значно більшим за avCS + та appCS + порівняно з CS–. Не було виявлено відмінностей під час фази вимирання. *p <0.05, **p > 0.01, ***p < 0.001.
Рейтинги
ANOVA для рейтинги валентності під час придбання виявлено значні основні ефекти стимулу [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, частковий η2 = 0.358] і фаза [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, частковий η2 = 0.096], а також значна взаємодія між стимулом і фазою [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, частковий η2 = 0.235]. Слідувати t-тести показують, що валентності чотирьох геометричних фігур на початку експерименту були однаковими (ps> 0.19), тоді як після придбання 1 та придбання 2 avCS + було оцінено як особливо негативне, а appCS + - як особливо позитивне. Зокрема, avCS + було оцінено як більш негативно валентне порівняно з CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], НОВИЙ [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] та appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + оцінено як значно позитивніший, ніж CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] та НОВИЙ [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Відмінності між CS– та NEW ніколи не були суттєвими (ps> 0.18).
Важливо, що порівняння валентних оцінок для appCS +, пов’язаних із шоколадом та солоним кренделем, не було виявлено жодних суттєвих відмінностей [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Після фази вимирання avCS + все ще був оцінений як більш негативний порівняно з CS– [t(31) = 2.40, p = 0.023], НОВИЙ [t(31) = 3.40, p = 0.002], а також додатокCS + [t(31) = 3.35, p = 0.002]. Навпаки, валентність appCS + вже не відрізнялася від CS– [t(31) = 1.77, p = 0.086] або НОВИЙ [t(31) = 1.07, p = 0.293].
ANOVA для рейтинги збудження під час придбання виявлено значний основний ефект стимулу [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, частковий η2 = 0.181], але не фази [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, частковий η2 = 0.038], і значна взаємодія між стимулом і фазою [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, частковий η2 = 0.124]. Слідувати t-тести не показали суттєвих відмінностей між подразниками щодо їх початкового збудження (ps> 0.74). Однак після двох фаз придбання avCS + було оцінено як більш збудливе, ніж НОВЕ [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] та appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] і незначно більш збудливий, ніж CS - після першої фази придбання [t(32) = 1.96, p = 0.058], але значно більше збуджує після другої фази придбання [t(32) = 3.65, p = 0.001]. На відміну від рейтингів валентності, збудження appCS + не відрізнялося від збудження CS - та NEW (ps > 0.13) після фаз придбання.
Так само, як і валентні рейтинги, CS– та NEW не відрізнялися за своїм збудженням (ps> 0.07).
Як і у випадку з валентністю, не було виявлено відмінностей для appCS +, асоційованого з шоколадом, і appCS +, пов’язаного з солоним кренделем [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Після фази вимирання суттєвих відмінностей у оцінках збудження не виявлено (ps> 0.08).
Для додаткового post-hoc t-тести, що порівнюють рейтинги після фази звикання, першої та другої фази придбання та фази вимирання, див. Додаткові матеріали.
Страхітна відповідь
ANOVA для фаз придбання повертає основний ефект стимулу [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, частковий η2 = 0.609], але не фаза [F(1, 32) = 3.16, p = 0.085, частковий η2 = 0.090], і істотний стимул взаємодії × Фаза [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, частковий η2 = 0.095]. Слідувати t-тести виявили значну потенційну здатність до avCS + порівняно з CS– під час обох перших [t(32) = 3.27, p = 0.003] і другий [t(32) = 4.00, p <0.001] фази придбання. Здивовані відповіді на avCS + також були значно посилені порівняно з відповідями на appCS +, знову ж таки після обох придбань 1 [t(32) = 8.20, p <0.001] та придбання 2 [t(32) = 5.74, p <0.001]. Важливо, що здивована величина appCS + була значно ослаблена порівняно з CS– під час обох придбань 1 [t(32) = 6.34, p <0.001] та придбання 2 [t(32) = 2.91, p = 0.007]. Знову і відповідно до рейтингів, для програмиCC + не було виявлено диференціальних реакцій враження, коли вони асоціювалися з шоколадом або солоною кренделями [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Під час фази вимирання значного ефекту не виявлено [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, частковий η2 = 0.008].
Відповідь провідності шкіри (SCR)
Із ANOVA для SCR протягом двох фаз придбання основний стимул впливу [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, частковий η2 = 0.401] і фаза [F(1, 27) = 20.91, p <0.001, частковий η2 = 0.436] виявилося важливим, але не їх взаємодія [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, частковий η2 = 0.024]. Post-hoc t-тести вказували на значно більшу SCR до avCS + [t(27) = 6.46, p <0.001] і до програмиCS + [t(27) = 4.84, p <0.001] порівняно з CS–, тоді як учасники продемонстрували порівнянний SCR з avCS + та appCS + [t(27) = 0.64, p = 0.527]. Помітно, не було виявлено відмінностей у SCR до програми для шоколадуCCS + та до програми для солоного кренделіCc + [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Як це стосується рейтингів та вражаючої реакції, значущих наслідків для фази вимирання не було виявлено [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, частковий η2 = 0.010].
Обговорення
Метою цього дослідження було перевести знахідки тварин на людей, використовуючи класичну апетитну умовну парадигму з первинним підсилювачем як безумовний стимул (США). Для цього учасники прийшли до лабораторії рано вранці без сніданку, щоб переконатися, що вони голодні, і відповідно до їх уподобань, шматочки шоколаду чи солоні кренделі використовувались як привабливі американці. Під час фази придбання одна геометрична форма (avCS +) стала асоціюватися з м'яко болісним електричним струмом (відразна США), інша форма (appCS +) із апетитними США та третя форма (CS–), що не має афективних США та не неприязні США. Результати вказують на успішне неприйнятне та апетитне обумовлення на явному вербальному рівні (тобто рейтингах), на рівні неявної поведінки (тобто стрімкої реакції) та на фізіологічному рівні (тобто, SCR). Зокрема, avCS + порівняно з CS - виявив більше негативних валентних рейтингів, вищі рейтинги збудження, потенціал стресу та більший SCR. Найголовніше, що appCS + порівняно з CS– викликало більше позитивних валентних оцінок, ослаблення стресу та більш високу SCR. Наші висновки щодо неприязного кондиціонування відповідали очікуванням, оскільки попередні дослідження виявили, що стимул, що прогнозує загрозу (avCS +), оцінюється як відраза, викликає більший рівень страху та посилює фізіологічне збудження (Fendt і Fanselow, 1999; Хамм і Вейке, 2005; Andreatta та ін., 2010, 2013). Наші результати щодо апетитного кондиціонування також відповідали попереднім дослідженням на людях і тваринах, показуючи, що стимул, що передбачає винагороду (appCS +), оцінюється як позитивний, гальмує реакції на страх та посилює фізіологічне збудження (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Остін і Дука, 2010; Prévost et al., 2012). Наскільки нам відомо, це перше дослідження, що демонструє умовне ослаблення стрелу у людини до стимулу, що передбачає первинну винагороду. Важливо, що нам вдалося перенести та підтвердити результати дослідження на тваринах (Koch et al., 1996). Це дослідження на тваринах продемонструвало, що ослаблення стрелів у щурів залежить від прогнозів від NAcc (частини вентральної смуги) до PnC. Отже, ослаблення стресу в нашому дослідженні може означати активність NAcc, що також відповідає результатам fMRI (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Леві та Глімчер, 2011). Отже, ми робимо висновок, що наша парадигма з привабливою умовою була успішною, оскільки вказували як явна (рейтинги), так і неявна позитивна валентність (ослаблення стресу).
Окрім цієї нової, але досить передбачуваної знахідки, варто згадати ще два цікаві результати. По-перше, вербальні та фізіологічні відповіді на збудження на додатокCS + дисоціювали. По-друге, ми виявили більш швидке вимирання апетитних КР порівняно з протилежними КР.
SCR вказує на активацію симпатії, яка збільшується як до стимулюваного стимулу (appCS +), так і до погрози, пов'язаного із загрозою (avCS +). Цей результат відповідає попередньому дослідженню кондиціонування, в якому еротичні фотографії використовувались як апетитні США (Klucken et al., 2013), і дослідження, що виявляє порівнянний SCR - кия, що прогнозує гроші, і кия, що передбачає неприязний шум (Остін і Дука, 2010). Зокрема, SCR є орієнтуючою реакцією, пов'язаною з активацією симпатичної системи. Ця відповідь пропонується відображати підготовку поведінкової реакції на мотиваційно яскраві події (Бредлі, 2009). Виходячи з цього, ми вважаємо, що стимули, пов'язані із загрозою, та пов'язані з винагородою, викликали підготовчу відповідь для послідовних поведінкових реакцій. Іншими словами, їжа та больові сигнали ініціювали підготовку відповідно до поведінки та уникання поведінки. На відміну від високого фізіологічного збудження, словесні відповіді вказували на низьке збудження щодо стимулу, пов’язаного з нагородою. Можливо, що на словесне збудження може впливати більше збудливий характер США, а не фізіологічна активація сам по собі. Насправді, ми вважаємо, що appCS + проти avCS + були оцінені як низькі порівняно з сильно збудливими, оскільки вони були пов'язані відповідно з низькими та сильно збуджуючими США. На жаль, ми не зібрали рейтингів збудження для американців, і тому майбутні дослідження повинні перевірити цю гіпотезу чітко.
Під час фази вимирання США не доставляли. Це, можливо, спонукало до нового гальмівного навчання, званого вимирання (огляд див Мілад і Кирк, 2012), причому як відхилення, так і апетитні КР зменшуються як наслідок. Очевидно, що на поведінковому (тобто реакції на стрес) та на фізіологічному (тобто SCR) рівні не було виявлено жодних диференціальних реакцій на avCS +, appCS + та CS–. Аналогічно, явне збудження умовних подразників, оцінених після фази вимирання, вирівнюється на низькому рівні, що говорить про успішне навчання вимирання. Однак avCS + все ще був оцінений значно більш негативно, ніж CS–, тоді як валентність appCS + вже не відрізнялася від валентності CS. Більш повільне згасання відвертої явної відповіді може бути наслідком еволюційного консерватизму, що означає, що сигнали про загрозу особливо важко забути, оскільки невідповідь на сигнал загрози може бути небезпечною для життя.
Залишається одне питання: чому тоді реалізація стресу (тобто неявна валентність) повністю згасла під час фази вимирання? По-перше, ці відповіді були обчислені протягом усього курсу фази. Тому можливо, що дискримінаційні КР все ще виявлятимуться під час перших випробувань фази вимирання. У дослідний спосіб ми дотримувались цієї гіпотези і розглядали як стрімкі реакції, так і SCR протягом усієї фази вимирання (див. Додатковий матеріал). Хоча ми не виявили суттєвих відмінностей, ми спостерігали дещо вищу величину пуску для avCS + порівняно з CS– та новим стимульним контролем. Ми також спостерігали дещо більше ослаблення стресу до appCS + порівняно з CS– та новим стимулом контролю на самому початку фази вимирання, яке, однак, зникло протягом декількох випробувань. SCR до appCS + знизився вже після випробування вимирання 2nd, тоді як SCR до avCS + залишився вище майже для всього вимирання порівняно з SCR до NEW. Хоча слід трактувати ці результати з великою обережністю, здається, що вражаючі реакції паралельно оцінювали валентність. Більше того, і страхітливі відповіді, і SCR надалі підтримують ідею еволюційного консерватизму щодо стимулів погрози.
Нарешті, ми маємо визнати деякі обмеження цього дослідження. По-перше, через технічну проблему ми не змогли повідомити про оцінку приємності (і збудження) для шоколаду та солоної кренделі. Тим не менш, апетитні КР припускають, що учасники дійсно відчували двох США як апетитні. По-друге, тривалість аверсивної та апетитної США сильно відрізнялася. Таким чином, хворобливий ураження електричним струмом було доставлено під час компенсації avCS + і тривало рівно 200 мс, тоді як шоколад та солоний кренделі були представлені учасникам близько 2 s після початку appCS +, а тривалість була невизначена, оскільки це залежало від швидкості людина їх їв. Вибір доставити апетитні США таким чином базувався на попередньому дослідженні на людях (Prévost et al., 2012). Однак методологічно було б більш елегантно доставити апетитну США, порівняннішу з відразовою США, наприклад, ковток соку або імбирний ель при компенсації appCS +. Для компенсації цієї великої перцептивної різниці ми представили геометричні фігури в поєднанні з візуальним стимулом, що символізує США. По-третє, ми не можемо однозначно виключити можливість того, що швидко згасаючі фізіологічні реакції у фазі вимирання зумовлені методологічним аспектом. Насправді візуальні стимули в цій фазі не були представлені разом із символом США, як це було у фазах придбання. Цілком можливо, що представлення лише КС, можливо, вплинуло на КР, і з цієї причини суттєвих відмінностей не було видно під час перших випробувань на вимирання. Однак наші швидкі погашені відповіді узгоджуються з погашеними реакціями в попередньому дослідженні, в якому CS + (обличчя) було представлено в поєднанні з США (крик) під час придбання, але не під час згасання (Ліссек та ін., 2008).
На закінчення ми знайшли успішні відверті та апетитні обумовлені реакції на стимул, пов'язаний із загрозою, та на стимул, пов’язаний із винагородою відповідно. Цікаво, що явний (рейтинги) та неявний (рефлекторний стрес) рівень відповідей працювали синергічно, оскільки avCS + повідомлялося як про негативне та індуковане потенційне запуску, а про appCS + повідомляли як про позитивне та індуковане загасання пуску. Крім того, чітке (рейтингове) та фізіологічне (SCR) збудження appCS + роз'єднане, що відображає два різних процеси.
Заява про конфлікт інтересів
Автори заявляють, що дослідження проводилося за відсутності будь-яких комерційних або фінансових відносин, які могли б бути витлумачені як потенційний конфлікт інтересів.
Подяки
Цю роботу підтримав Центр спільних досліджень «Страх, тривога та тривожні розлади», SFB-TRR 58, проект B1.
Додатковий матеріал
Додатковий матеріал для цієї статті можна знайти в Інтернеті за адресою: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
посилання
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. and Pauli, P. (2013). Передбачуваність болю скасовує валентність рельєфу, пов'язаного з полегшенням. Передня. Сист. Невросці. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Анотація | CrossRef Повний текст | Google Scholar
Андреатета, М., Мюльбергер, А., Яралі, А., Гербер, Б. і Паулі, П. (2010). Розрив між неявною та явною умовною валентністю після навчання болю у людей. Зб. Біол. Наук. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Анотація | CrossRef Повний текст | Google Scholar
Остін, Джорджія та Дука, Т. (2010). Механізми уваги щодо апетитних та неприязних результатів в умовах павловського кондиціонування. Behav. Brain Res. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Анотація | CrossRef Повний текст | Google Scholar
Блюменталь, Т.Д., Катберт, Б.Н., Філіон, Д.Л., Хаклі, С., Ліпп, О. В. та Ван Бокстель, А. (2005). Доповідь Комітету: вказівки для електроміографічних досліджень очного моменту, що вражають людину. Психофізіологія 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Анотація | CrossRef Повний текст | Google Scholar
Bouton, ME та Peck, Каліфорнія (1989). Контекстний вплив на кондиціонування, вимирання та відновлення в апетитному препараті для кондиціонування. Тварина. Дізнайтеся. Бехав. 17, 188 – 198. doi: 10.3758 / BF03207634
CrossRef Повний текст | Google Scholar
Бредлі, М.М. (2009). Природна вибіркова увага: орієнтація та емоція. Психофізіологія 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Анотація | CrossRef Повний текст | Google Scholar
Кларк, Дж. Дж., Холлон, НГ та Філіпс, PEM (2012). Павловські системи оцінювання в навчанні та прийнятті рішень. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Анотація | CrossRef Повний текст | Google Scholar
Delgado, MR, Jou, RL та Phelps, EA (2011). Нейронні системи, що лежать в основі аверсивного кондиціонування у людини з первинними та вторинними підсилювачами. Фронт. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Анотація | CrossRef Повний текст | Google Scholar
Дікінсон, А. і Баллен, Б. (1994). Мотиваційний контроль цільових дій. Тварина. Дізнайтеся. Бехав. 22, 1 – 18. doi: 10.3758 / BF03199951
CrossRef Повний текст | Google Scholar
Fendt, M. та Fanselow, MS (1999). Нейроанатомічна та нейрохімічна основа обумовленого страху. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Анотація | CrossRef Повний текст | Google Scholar
Готфрід, JA, О'Дохерті, J., і Dolan, RJ (2002). Апетитивне та аверсивне нюхове навчання у людей досліджували за допомогою функціональної магнітно-резонансної томографії. J. Neurosci. 22, 10829 – 10837. Доступний онлайн за адресою: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B., and Johnson, L. (2006). Контекстне обумовлення та уникнення поведінки у середовищі віртуальної реальності: ефект передбачуваності. Biol. Психіатрія 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Анотація | CrossRef Повний текст | Google Scholar
Hamm, AO та Weike, AI (2005). Нейропсихологія навчання страху та регулювання страху. Int. Ж. Психофізіол. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Анотація | CrossRef Повний текст | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Нейрові активізації набуття умовного сексуального збудження: наслідки усвідомлення непередбачених ситуацій та сексу. J. Секс. Мед. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Анотація | CrossRef Повний текст | Google Scholar
Klucken, Т., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). Поліморфізм 5-HTTLPR пов'язаний зі зміненими гемодинамічними реакціями під час апетитного кондиціонування. Hum. Мозок Мапп. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Анотація | CrossRef Повний текст | Google Scholar
Кох, М. (1999). Нейробіологія стресу. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Анотація | CrossRef Повний текст | Google Scholar
Кох, М., Шмід, А. та Шніцлер, Х.-У. (1996). Задоволення-ослаблення пуску порушується ураженням ядер ядер. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Анотація | CrossRef Повний текст | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W., and Tausch, A. (1996). Універсальна версія програми "Позитивний та негативний вплив" (PANAS). Діагностика 42, 139-156.
Кумар, П., Офіціант, Г., Ахерн, Т., Мілдерс, М., Рейд, І. та Стіл, Дж. Д. (2008). Ненормальна часова різниця нагороджує сигнали при великій депресії. Brain 131, 2084 – 2093. doi: 10.1093 / brain / awn136
PubMed Анотація | CrossRef Повний текст | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P., and Spielberger, CD (1981). Das State-Trait Angstinventar. Вайнхайм: Тест Бельца.
Levy, DJ та Glimcher, PW (2011). Порівняння яблук і апельсинів: використання специфічного для винагороди та загального суб'єктивної цінності подання в мозку. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Анотація | CrossRef Повний текст | Google Scholar
Ліссек, С., Левенсон, Дж., Біггс, А. Л., Джонсон, Л. Л., Амелі, Р., Пайн, Д.С. та ін. (2008). Підвищений страх обумовлює соціально значущі безумовні стимули при соціальному тривожному розладі. Ам. J. Психіатрія 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Анотація | CrossRef Повний текст | Google Scholar
Martin-Soelch, C., Linthicum, J., та Ernst, M. (2007). Апетитивна обумовленість: нервові основи та наслідки для психопатології. Neurosci. Biobehav. Rev. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Анотація | CrossRef Повний текст | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y., and Schoenbaum, G. (2011). Вентральний стриатум і орбітофронтальна кора необхідні для вивчення підкріплення, що базується на моделі, але не є модельним. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Анотація | CrossRef Повний текст | Google Scholar
Мілад, MR та Quirk, G. (2012). Вимирання страху як модель для трансляційної нейронауки: десять років прогресу. Анну. Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Анотація | CrossRef Повний текст | Google Scholar
Павлов, IP (1927). Умовні рефлекси: дослідження фізіологічної активності кори головного мозку. Лондон: Oxford University Press.
Превост К., Лілєхольм М., Тишка Дж. М., О'Доерті, JP (2012). Нейронні кореляти специфічного та загального перекладу Павлова на інструментальний пристрій в субрегіонах мигдалин людини: дослідження fMRI з високою роздільною здатністю. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Анотація | CrossRef Повний текст | Google Scholar
Tranel, D. та Damasio, H. (1994). Нейроанатомічні кореляти електродермальних реакцій шкірної провідності. Психофізіологія 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Анотація | CrossRef Повний текст | Google Scholar
Ключові слова: класичне кондиціонування, винагорода, покарання, рефлекс стресу, реакція на шкірну провідність
Citation: Andreatta M and Pauli P (2015) Апетитивний та неприйнятний стан у людини. Фронт. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Отримано: 11 лютого 2015; Прийнято: 05 травня 2015;
Опубліковано: 19 травень 2015.
Редаговано:
Нільс Бірбаумер, Університет Тюбінген, Німеччина
Відгук:
Тім Клюкен, Університет Юстаса Лібіга, Гіссен, Німеччина
Алессандро Ангріллі, Університет Падуї, Італія
Copyright © 2015 Andreatta і Pauli. Ця стаття з відкритим доступом розповсюджується за умовами Ліцензія на використання Creative Commons (CC BY). Використання, розповсюдження або відтворення на інших форумах дозволяється за умови зарахування оригінального автора (авторів) або ліцензіара, а також цитування оригінальної публікації в цьому журналі відповідно до прийнятої академічної практики. Не дозволяється використання, розповсюдження або відтворення, які не відповідають цим умовам.
* Листування: Марта Андреата, кафедра психології (біологічна психологія, клінічна психологія та психотерапія), Вюрцбургський університет, Маркусстрассе 9-11, D-97070 Вюрцбург, Німеччина, [захищено електронною поштою]