Neurosci Biobehav Rev. 2011 Tháng 1; 35 (3): 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Tháng 11 2.
- 1Khoa Khoa học Thần kinh Bernard B. Brodie, Đại học Cagliari, Cittadella Universitaria, 09042 Monserrato, CA, Italy. [email được bảo vệ]
TRỪU TƯỢNG
Oxytocin là một chất gây cảm ứng cương dương mạnh khi tiêm vào hệ thần kinh trung ương. Ở chuột đực, khu vực não nhạy cảm nhất đối với tác dụng cương dương của oxytocin là nhân trung tâm của vùng dưới đồi. Hạt nhân này và các vùng lân cận chứa các tế bào của tất cả các tế bào thần kinh oxytocinergic chiếu đến các vùng não ngoài vùng dưới đồi và tủy sống. Tổng quan này cho thấy rằng oxytocin cũng gây ra sự cương cứng dương vật khi được tiêm vào một số khu vực này (ví dụ, vùng não thất, vùng dưới đồi của vùng đồi thị, nhân vỏ não sau của amygdala và cột sống thắt lưng). Các nghiên cứu vi tiêm kết hợp với vi lọc trong não và nghiên cứu miễn dịch huỳnh quang kép cho thấy oxytocin ở những khu vực này kích hoạt trực tiếp hoặc gián tiếp (chủ yếu thông qua axit glutamic) tế bào thần kinh dopaminergic mesolimbic. Dopamine được giải phóng trong nhân accumbens lần lượt kích hoạt các con đường thần kinh dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic incothalamic trong nhân paraventricular. Điều này kích hoạt không chỉ các tế bào thần kinh oxytocinergic chiếu vào tủy sống và trung gian cương dương vật, mà còn kích hoạt các khu vực ngoài vùng dưới đồi, điều chỉnh trực tiếp hoặc gián tiếp (thông qua axit glutamic) hoạt động của các tế bào thần kinh trung mô. Cùng với các con đường thần kinh này có thể tạo thành một mạch giả thuyết phức tạp, đóng vai trò không chỉ trong giai đoạn hoàn thành hoạt động tình dục (chức năng cương dương và giao hợp), mà còn trong các khía cạnh thúc đẩy và bổ ích của giai đoạn dự đoán hành vi tình dục.
1.Giới thiệu
Cương cứng dương vật là một phản ứng tình dục của nam giới đóng vai trò chính trong việc sinh sản của động vật có vú bao gồm cả con người và cũng có thể được quan sát trong các bối cảnh khác với những bối cảnh liên quan chặt chẽ đến sinh sản. Tùy thuộc vào bối cảnh xảy ra tình trạng cương dương vật, các cơ chế thần kinh trung ương và ngoại biên và / hoặc thể dịch khác nhau tham gia vào quy định của nó (xem Meisel và Sachs, 1994; Argiolas và Melis, 1995, 2004, 2005; Sachs, 2000, 2007; 2000; Giuliano và Rampin, 2000, 2004; Andersson, 2001; Melis và Argiolas, 1995a, 2003; Hull và đồng sự, 2002). Trong số các chất dẫn truyền thần kinh trung ương và neuropeptide kiểm soát sự cương cứng của dương vật, được biết đến nhiều nhất là dopamine, serotonin, axit amin kích thích, oxit nitric, adrenocorticotropin, oxytocin và peptide opioid. Họ có thể tạo điều kiện hoặc ức chế cương dương vật bằng cách hành động trong một số khu vực não, tức là vùng tiền sản trung gian, nhân trung tâm của vùng dưới đồi, vùng não thất, vùng đồi thị, vùng amygdala, nhân giường của stria terminalis, nhân não, ống tủy (Bảng 1) (xem Meisel và Sachs, 1994; Witt và Insel, 1994; Stancampiano et al., 1994; Argiolas và Melis, 1995, 2005; Argiolas, 1999; Bancila et al., 2002 McKenna, 2000; Andersson, 2000; Hull và đồng sự, 2001; Coolen và cộng sự, 2002).
Oxytocin, peptide neurohypophyseal nổi tiếng với vai trò nội tiết tố trong việc tiết sữa và đẻ, có mặt ở nữ giới và nam giới, không chỉ trong các tế bào thần kinh với các tế bào thần kinh nằm trong các tế bào thần kinh và tế bào thần kinh của vùng dưới đồi. từ nhân paraventricular và các cấu trúc xung quanh đến các khu vực não ngoài màng cứng (tức là vùng kín, vùng não thất, vùng đồi thị, amygdala, tủy não và tủy sống). Những tế bào thần kinh này được cho là có liên quan đến nhiều chức năng trung tâm, như trí nhớ, học tập, hành vi liên kết và tình dục xã hội, bao gồm cương dương vật và hành vi điều phối (xem Bùijs, 1978; Sofroniew, 1983; Argiolas và Gessa, 1991; Pedersen et ., 1992; Carter, 1992; Wagner và Clemens, 1993; Ivell và Russel, 1995; Carter et al., 1997; Tang et al., 1998; Veronneau-Longueville et al., 1999). Thật vậy, oxytocin tạo điều kiện cho chức năng cương dương và hành vi tình dục của nam giới ở chuột, chuột, thỏ và khỉ (xem Argiolas và Gessa, 1991; Carter, 1992; Pedersen et al., 1992; Argiolas và Melis, 1995, 2004 Điều này cũng có thể xảy ra ở người, vì oxytocin trong huyết tương tăng lên do kích thích tình dục, đặc biệt là khi xuất tinh (Carmichael và cộng sự, 1999; Murphy và cộng sự, 1987) và do thao tác của vú và cơ quan sinh dục, thường xảy ra khi quan hệ tình dục giao hợp (Tindall, 1987).
Tác dụng thuận lợi của oxytocin đối với hành vi tình dục của nam giới lần đầu tiên được chứng minh bằng khả năng oxytocin tiêm tĩnh mạch làm giảm độ trễ cho lần xuất tinh đầu tiên và làm chậm sự cạn kiệt tình dục của thỏ đực kết hợp với con cái dễ tiếp nhận (Melin và Kihlstrom, 1963). Tuy nhiên, tác dụng tình dục của oxytocin chỉ được công nhận chắc chắn vào những năm tám mươi. Oxytocin được cung cấp tập trung ở số lượng nanogram sau đó đã được tìm thấy có thể gây ra sự cương cứng dương vật (Argiolas và cộng sự, 1985, 1986) và để cải thiện hành vi điều phối (Arletti et al., 1985) ở chuột đực và làm tăng tình trạng nhiễm mỡ ở chuột cái (Arletti và Bertolini, 1985; Caldwell và cộng sự, 1986), dường như bằng cách tác động lên các thụ thể oxytocinergic loại tử cung (xem Argiolas và Melis, 1995, 2004; Argiolas, 1999; Melis và Argiolas, 2003). Oxytocin cải thiện hành vi tình dục không chỉ ở chuột đực mạnh về tình dục (Arletti et al., 1985) mà còn ở chuột đực già (Arletti et al., 1990), và ở trội, nhưng không phải ở khỉ, sóc đực (Winslow và Insel , 1991).
Tác dụng chống cương dương của oxytocin phụ thuộc vào testosterone, vì nó bị loại bỏ bằng cách cắt bỏ và cắt bỏ, và được phục hồi bằng cách bổ sung testosterone hoặc các chất chuyển hóa của nó, estradiol và 5_-dihydro-testosterone (Melis và cộng sự, 1994a). Vùng não nhạy cảm nhất để tạo ra sự cương cứng dương vật bởi oxytocin là nhân trung tâm của vùng dưới đồi (Melis et al., 1986), từ đó tất cả các dự đoán oxytocinergic ngoài vùng dưới đồi bắt nguồn (xem ở trên). Ở đây, oxytocin đã được tìm thấy có thể gây ra sự cương cứng dương vật (và ngáp) khi được tiêm với liều thấp như 3 pmol (xem Phần 2.1 bên dưới). Oxytocin cũng gây ra sự cương cứng dương vật khi được tiêm hai bên vào trường CA1 của vùng đồi thị, nhưng không phải ở vùng dưới màng cứng (xem Phần 2.3 bên dưới), vách ngăn bên, nhân trung tâm, vùng tiền đình hạt nhân siêu âm (Melis et al., 1986). Đối với cơ chế mà oxytocin hoạt động trong nhân trung tâm để tạo ra phản ứng tình dục này, nhiều nghiên cứu cho thấy oxytocin kích hoạt tế bào thần kinh của chính nó. Theo giả thuyết này, sự tương tác tình dục làm tăng FOS, sản phẩm gen của gen c-fos ngay lập tức trong các tế bào thần kinh oxytocinergic paraventricular chiếu vào tủy sống, có liên quan đến việc kiểm soát cương dương vật (xem Witt và Insel, 1994 trong đó), và bất lực về tình dục (ví dụ, sự bất lực của một con chuột đực trưởng thành để giao hợp với một con cái tiếp nhận estrogen-progesteroneprimed ovariectomized) đã được liên kết ở chuột đực với nồng độ oxytocin mRNA thấp trong nhân paraventular al., 1997).
Hiện tại liệu oxytocin có ảnh hưởng đến giai đoạn dự đoán hay giai đoạn hoàn thành hành vi tình dục không rõ ràng hay không. Vì oxytocin gây ra sự cương cứng dương vật và tác dụng chính của oxytocin đối với hành vi điều phối là làm giảm khoảng thời gian sau xuất tinh ở chuột đực (Arletti et al., 1985), nên có lý khi cho rằng peptide cải thiện hiệu suất tình dục. Tuy nhiên, vì oxytocin cũng làm tăng tương tác tình dục xã hội (xem Pedersen và cộng sự, 1992; Carter và cộng sự, 1997; Ivell và Russel, 1995) và các chất đối kháng thụ thể oxytocin ngăn ngừa sự cương cứng không tiếp xúc (Melis et al., 1999a). được coi là một chỉ số của hưng phấn tình dục (xem Sachs, 1997, 2000, 2007; Melis và cộng sự, 1998, 1999b và các tài liệu tham khảo trong đó), không thể loại trừ vai trò của oxytocin trong hưng phấn tình dục và động lực tình dục.
Đánh giá này tóm tắt các kết quả được công bố và chưa được công bố của các nghiên cứu gần đây, cho thấy rằng Oxytocin gây ra sự cương cứng dương vật không chỉ khi được tiêm vào nhân trung tâm của vùng dưới đồi, mà còn ở các vùng não ngoài vùng dưới đồi khác, chẳng hạn như vùng não thất (Melis và cộng sự, 2007, 2009a; Succu và cộng sự, 2008), tanh ta đặt bụng dưới đồi thị và nhân sau của amygdala (Melis và cộng sự, 2009b, 2010), là những thành phần quan trọng của hệ thống limbic và được cho là đóng vai trò chính trong quá trình thúc đẩy và khen thưởng. Các nghiên cứu này tiết lộ rằng oxytocin tham gia vào các mạch thần kinh, bao gồm các chất dẫn truyền thần kinh khác, chẳng hạn như dopamine và axit glutamic, và các khu vực não khác ngoài nhân não, ví dụ, vùng não thất, vùng não, vùng đồi thị và vùng đồi thị. xác định. Các mạch này có khả năng làm trung gian cho sự tương tác giữa hệ thống dopaminergic mesolimbic và incothalamic, và đóng vai trò không chỉ trong giai đoạn hoàn thành hành vi tình dục của nam giới (ví dụ, cương dương vật và giao hợp), mà còn trong động lực tình dục và tình dục kích thích, do đó cung cấp một chất nền thần kinh để giải thích các thuộc tính động lực và bổ ích của hoạt động tình dục.
KHAI THÁC. Oxytocin ảnh hưởng đến sự cương cứng dương vật bằng cách hành động ở các vùng não khác nhau
KHAI THÁC. Nhân trung tâm của vùng dưới đồi
Như đã nhắc lại ở trên, nhân trung tâm của vùng dưới đồi đã sớm được xác định là vùng não nhạy cảm nhất đối với tác dụng cương dương của oxytocin. Khi được tiêm đơn phương vào nhân này, oxytocin đã được tìm thấy hoạt động ở liều thấp như 3 ng (3 pmol) (Melis et al., 1986). Các nghiên cứu về mối quan hệ hoạt động cấu trúc cho thấy sự cương cứng dương vật do oxytocin gây ra được trung gian bởi các thụ thể oxytocin tử cung, kết hợp với một dòng Ca2 + vào các tế bào của các tế bào thần kinh oxytocinergic chiếu vào các vùng não ngoại vi. Lần lượt oxit nitric bằng cách hoạt động như một chất truyền tin nội bào với cơ chế chưa được biết đến (không liên quan đến guanylate cyclase) dẫn đến việc kích hoạt các tế bào thần kinh oxytocinergic chiếu vào tủy sống và các vùng não ngoài vùng dưới đồi, tạo ra sự cương cứng dương vật (Hình. 1) bên dưới và Argiolas và Melis, 1995, 2004, 2005 và các tài liệu tham khảo trong đó). Khả năng oxytocin kích hoạt tế bào thần kinh của chính nó
Hình. 1. (MISSING) Biểu diễn sơ đồ của các tế bào thần kinh oxytocinergic, bắt nguồn từ nhân paraventricular của vùng dưới đồi và dự án cho các khu vực não ngoài vùng dưới đồi, như tủy sống, VTA, đồi thị, amygdala, v.v. dopamine, axit amin kích thích, chính oxytocin, peptide tương tự hexarelin và peptide có nguồn gốc VGF dẫn đến cương dương vật, có thể giảm và / hoặc bị hủy bỏ do kích thích thụ thể GABAergic, opioid và cannabinoid CB1. Sự kích hoạt của các tế bào thần kinh oxytocinergic là thứ yếu để kích hoạt nitric oxide-synthase có trong các tế bào thần kinh này. Thật vậy, oxit nitric nội sinh được hình thành do sự kích thích của dopamine, axit amin kích thích hoặc thụ thể oxytocin hoặc oxit nitric ngoại sinh, do có nguồn gốc từ các nhà tài trợ oxit nitric được đưa trực tiếp vào nhân paraventricular, kích hoạt các tế bào thần kinh oxytocinergic. kích thích guanylate cyclase. Điều này gây ra sự giải phóng oxytocin trong tủy sống và ở các vùng não ngoài vùng dưới đồi. Một số chi tiết về các cơ chế mà oxytocin gây ra sự cương cứng dương vật khi được giải phóng ở những khu vực này, ví dụ, VTA, tiểu não thất và amygdala được mô tả trong các phần não tương ứng. Ở đây, oxytocin tác động lên các thụ thể của chính nó và làm tăng sản xuất NO, dẫn đến sự cương cứng dương vật như được tìm thấy trong PVN. Tuy nhiên, không phù hợp với PVN, trong VTA NO, kích hoạt cyclase guanylate. Điều này gây ra sự gia tăng nồng độ cGMP dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic và cương cứng dương vật. Trong VS NO kích hoạt các tế bào thần kinh glutamatergic chiếu tới các khu vực ngoài đồi thị, bao gồm cả VTA. Axit glutamic trong VTA kích hoạt lần lượt các tế bào thần kinh dopaminergic mesolimbic như được tìm thấy với oxytocin. Các cơ chế tương tự như được mô tả ở trên cũng có khả năng hoạt động khi cương cứng dương vật xảy ra trong bối cảnh sinh lý, cụ thể là khi chuột đực được đặt trong sự hiện diện của một con cái tiếp nhận không thể tiếp cận (ví dụ, cương cứng không tiếp xúc) hoặc trong quá trình giao hợp.
trong nhân paraventricular được hỗ trợ bởi các nghiên cứu cho thấy: (1) Các thụ thể oxytocin có trong nhân ở vùng dưới đồi này (Freund-Mercier và cộng sự, 1987; Freund-Mercier và Stoeckel, 1995); (2) oxytocin tạo điều kiện phát hành riêng in vitro và in vivo (Freund-Mercier và Richard, 1981, 1984; Moos et al., 1984); và (3) oxytocin kích thích tế bào thần kinh của chính nó bằng cách hoạt động trong nhân paraventricular (Yamashita và cộng sự, 1987). Hơn nữa, các khớp thần kinh oxytocinergic nằm trên cơ thể tế bào của các tế bào thần kinh oxytocinergic phóng đại cũng đã được xác định trong nhân paraventricular và supraoptic của vùng dưới đồi (Theodosis, 1985). Cuối cùng, phá hủy các tế bào thần kinh oxytocinergic trung ương bằng các tổn thương kích thích điện hóa hoặc hóa học của nhân paraventricular, làm suy giảm hoàn toàn hàm lượng oxytocin trên hệ thống thần kinh trung ương và tủy sống, loại bỏ không chỉ tác dụng cương dương của oxytocin. cương cứng dương vật và cương cứng không tiếp xúc (xem bên dưới và Argiolas và cộng sự, 1987a, b; Liu và cộng sự, 1997 và tài liệu tham khảo trong đó). Kết quả tương tự với kết quả tìm thấy với các tổn thương của nhân paraventricular được tìm thấy với chất đối kháng thụ thể oxytocin mạnh và chọn lọc. Thật vậy, các hợp chất này được tiêm vào nhân paraventricular với số lượng nanogram đã ngăn chặn sự cương cứng dương vật hoàn toàn do oxytocin gây ra, trong khi khi đưa vào tâm thất bên cạnh, không chỉ cương cứng dương vật do chính oxytocin gây ra. và Argiolas và Melis, 3, 1995, 2004 và các tài liệu tham khảo trong đó) và cương cứng không tiếp xúc (Melis et al., 2005a), và hơn nữa rất hiệu quả trong việc làm giảm hành vi điều phối của chuột đực mạnh về tình dục (Argiolas et al., XNUM). Hơn nữa, tương tác tình dục làm tăng FOS, sản phẩm gen của c-fos gen ngay lập tức trong các tế bào thần kinh oxytocinergic paraventricular chiếu vào tủy sống liên quan đến việc kiểm soát cương dương vật (xem Witt và Insel, 1999 và tài liệu tham khảo ở đó). Cuối cùng, bất lực tình dục (ví dụ, sự bất lực của một con chuột đực trưởng thành để giao hợp với một con cái tiếp nhận estrogen-progesteroneprimed) hcũng được liên kết ở chuột đực với nồng độ oxytocin mRNA thấp và synthase oxit nitric trong nhân paraventricular của vùng dưới đồi (Benelli et al., 1995; Arletti et al., 1997) (để đánh giá toàn diện các nghiên cứu này, hãy xem Argiolas, 1999; Argiolas và Melis, 2004, 2005).
KHAI THÁC. Vùng bụng
Vùng não thất được phát hiện gần đây là một vị trí não trong đó oxytocin gây ra sự cương cứng dương vật. Khu vực này chứa các đầu dây thần kinh oxytocinergic có nguồn gốc từ nhân paraventricular và thụ thể oxytocin (Freund-Mercier et al., 1987; Vaccari et al., 1998). Chính xác hơn, oxytocin được tìm thấy có khả năng của tôicương cứng dương vật khi tiêm đơn phương vào đuôi, nhưng không phải trong khu vực vây bụng theo cách phụ thuộc vào liều (Melis et al., 2007). Các liều hoạt động cao hơn so với liều cần thiết khi tiêm vào nhân trung tâm và tương tự như khi cương cứng dương vật gây ra khi tiêm vào tiểu não thất của đồi thị hoặc vào nhân vỏ não sau của amygdala (xem bên dưới). Rõ ràng, hiệu ứng cương dương được trung gian bởi sự kích hoạt của các tế bào thần kinh dopaminergic mesolimbic chiếu vào vỏ của các hạt nhân accumbens, từ đó kích hoạt các con đường thần kinh chưa được biết đến chiếu vào tế bào thần kinh dopaminergic (Melis và cộng sự, 2007, 2009a).
Đối với cơ chế oxytocin kích hoạt dẫn truyền thần kinh dopaminergic ở vùng não thất, dữ liệu có sẵn cho thấy oxytocin kích thích các thụ thể oxytocinergic nằm trong cơ thể tế bào của các tế bào thần kinh dopaminergic mesolimbic. Điều này làm tăng dòng Ca2 + bên trong cơ thể tế bào của các tế bào thần kinh dopaminergic, do đó kích hoạt nitric oxide-synthase (Succu et al., 2008). Khác với hạt nhân paraventricular (xem Phần 3 bên dưới), oxit nitric lần lượt kích hoạt guanylate cyclase, do đó làm tăng nồng độ của GMP theo chu kỳ. Theo cơ chế này, hoặc d (CH2) 5Tyr (Me) 2-Orn8-vasotocin, một chất đối kháng oxytocin mạnh, hoặc S-methyl-thio-l-citrulline, một chất ức chế mạnh của thần kinh khu vực vây bụng trước oxytocin, bãi bỏ dương vật cương cứng và sự gia tăng nồng độ dopamine ngoài tế bào trong vỏ của nhân được bồi tụ bởi oxytocin. Hơn nữa, 8-bromo-cyclic GMP, một chất tương tự theo chu kỳ hoạt động kháng phosphodiesterase hoạt động, gây ra sự cương cứng dương vật khi được tiêm vào vùng não của vây đuôi và làm tăng nồng độ dopamine ngoài tế bào trong vỏ của hạt nhân. vùng vây bụng phía sau (Succu et al., 2008; Melis et al., 2009a) (xem thêm hình 2).
Luôn phù hợp với cơ chế này, haloperidol, một chất đối kháng thụ thể dopamine D2 mạnh, được tiêm vào vỏ của hạt nhân accumbens làm giảm sự cương cứng dương vật do oxytocin tiêm vào vùng não thất (Melis et al., 2007). Cơ chế trên cũng được hỗ trợ bởi các nghiên cứu miễn dịch huỳnh quang kép, cho thấy các sợi oxytocin nằm trên cơ thể tế bào của các tế bào thần kinh dopaminergic ở vùng đuôi não, được dán nhãn trước đó bằng chất đánh dấu ngược dòng Fluorogold được tiêm vào vỏ của hạt nhân. Melis và cộng sự, 2007; Succu và cộng sự, 2008). Sự kích hoạt của các tế bào thần kinh dopaminergic và các thụ thể dopamine trong nhân accumbens dẫn đến việc kích hoạt các con đường thần kinh chưa được xác định, kích thích các tế bào thần kinh dopaminergic dopaminergic giải phóng dopamine trong tế bào thần kinh paropentamic. tủy sống và trung gian cương dương vật (xem ở trên và Melis và cộng sự, 2007; Succu và cộng sự, 2007, 2008). Thật vậy, oxytocin được tiêm vào vùng phía sau của vây đuôi với liều gây ra sự cương cứng dương vật, tăng nồng độ dopamine ngoài tế bào trong thẩm tách không chỉ thu được từ nhân accumbens, mà còn từ nhân paraventricular (Succu et al.
KHAI THÁC. Hồi hải mã
Trường CA1 của đồi hải mã là vùng não khác giàu sợi oxytocinergic và các thụ thể được xác định bởi các nghiên cứu trước đó trong đó tiêm oxytocin gây ra sự cương cứng dương vật (xem Bujis, 1978; Sofroniew, 1983). Tuy nhiên, khác với nhân paraventricular, ở đây oxytocin được tìm thấy có khả năng gây cương cứng dương vật khi tiêm hai bên và ở liều cao hơn so với những người tìm thấy hoạt động trong nhân paraventricular (Melis et al., 1986; Chen et al., 1992). Tiêm oxytocin vào subiculum được tìm thấy không hoạt động trong các nghiên cứu trước đó. Tuy nhiên, các nghiên cứu vi tiêm gần đây và cẩn thận hơn dẫn đến việc xác định một khu vực của tiểu tâm thất trong đó việc tiêm oxytocin có khả năng gây cương cứng dương vật theo cách phụ thuộc vào liều (Melis et al., 2009b). Tác dụng chống cương dương của oxytocin được tiêm vào vùng não này được quan sát thấy ở các liều tương tự như hoạt chất ở vùng não thất sau khi tiêm đơn phương (Melis et al., 2007), như được tìm thấy trong nhân não. Rõ ràng, oxytocin được tiêm vào tiểu não gây ra sự cương cứng dương vật bằng cách kích hoạt thụ thể oxytocinergic trong tế bào thần kinh có chứa nitric oxide-synthase, gây ra sự gia tăng sản xuất oxit nitric. Lần lượt oxit nitric đóng vai trò là chất truyền tin nội bào kích hoạt dẫn truyền thần kinh axit glutamic, dẫn đến cương dương vật, có thể thông qua các dự báo tràn dịch thần kinh (glutamatergic) từ tiểu não đến các vùng não ngoài vùng đồi thị điều chỉnh hoạt động của các tế bào thần kinh dopaminergic mesolimbic (ví dụ, vùng não thất, vỏ não trước trán, nhân paraventricular) (xem bên dưới và Melis, 2007, 2009b;
Cơ chế hoạt động này được hỗ trợ bởi các thí nghiệm vi lọc trong não, cho thấy oxytocin được tiêm vào tiểu não thất ở liều gây ra sự cương cứng dương vật, làm tăng sản xuất oxit nitric và nồng độ axit glutamic ngoại bào trong màng lọc từ màng bụng (Melis. , 2010) và của dopamine ngoại bào trong nhân accumbens (Melis et al., 2007). Những phản ứng này được đối kháng không chỉ bởi chất đối kháng thụ thể oxytocin d (CH2) 5Tyr (Me) 2-Orn8-vasotocin, mà còn bởi chất ức chế nitric oxide-synthase của tế bào thần kinh. vào tiểu não thất vài phút trước khi oxytocin (Melis và cộng sự, 2010).
Hơn nữa, theo cơ chế hoạt động này, kích hoạt dẫn truyền thần kinh glutamatergic bằng cách tiêm NMDA vào tiểu não thất gây ra sự cương cứng dương vật (Melis et al., 2010). Kiểu hình của các dự báo tràn ra từ tiểu não thất, gây ra sự kích hoạt của các tế bào thần kinh dopaminergic mesolimbic và sự gia tăng của dopamine ngoài tế bào trong nhân accumbens, hiện chưa rõ. Tuy nhiên, do sự cương cứng dương vật do oxytocin gây ra được tiêm vào tiểu não thất xảy ra đồng thời với sự gia tăng của axit glutamic ngoài tế bào trong thẩm tách từ vùng não thất, nhưng không phải từ nhân tế bào và được đối kháng bởi (+). một chất đối kháng mạnh không cạnh tranh của các thụ thể axit amin kích thích của phân nhóm NMDA (Woodruff et al., 801), được tiêm vào vùng não bụng, nhưng không được tiêm vào nhân (xem hình. 1987 và Melis et al., 2) , có khả năng những dự báo này dẫn đến việc kích hoạt dẫn truyền thần kinh glutamatergic ở vùng não thất, từ đó kích hoạt các tế bào thần kinh dopaminergic mesolimbic chiếu vào nhân tế bào. Cho dù nồng độ axit glutamic tăng lên được tìm thấy ở vùng não thất sau khi tiêm oxytocin vào tiểu não thất được giải phóng từ các tế bào thần kinh có nguồn gốc từ tiểu não hoặc ở các vùng não khác (ví dụ, vỏ não trước trán) hiện chưa rõ. Tuy nhiên, điều này gây ra sự kích hoạt các tế bào thần kinh dopaminergic mesolimbic và tăng giải phóng dopamine trong nhân accumbens. Ở đây, việc kích hoạt các thụ thể dopamine dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic incothalamic, giải phóng dopamine trong nhân paraventricular, do đó kích hoạt các tế bào thần kinh oxytocinergic chiếu vào tủy sống và trung gian cương dương vật (xem ở trên và Melis và cộng sự, 2007, 2009a; Succu et al., 2008).
KHAI THÁC. Các amygdala
Amygdala là một vùng não khác giàu sợi oxytocin và các thụ thể (xem Freund-Mercier và cộng sự, 1987; Vaccari et al., 1998; Uhl-Bronner et al., 2005). Oxytocin ở đây được cho là có liên quan đến các chức năng khác nhau, từ giải phẩu, trí nhớ xã hội và nhận thức, học tập được củng cố về mặt xã hội, sự đồng cảm về cảm xúc, xử lý khuôn mặt cảm xúc và nỗi sợ hãi ở con người để cương cứng và hành vi tình dục (xem Kondo et al., 1998; al., 2001; Ebner và cộng sự, 2005; Huber và cộng sự, 2005; Domes et al., 2007; Petrovic et al., 2008; Lee và cộng sự, 2009; Donaldson và Young, 2009; Hurlemann et al. , 2010). Tuy nhiên, khả năng oxytocin gây ra sự cương cứng dương vật ở chuột đực khi được tạo ra trong nhân vỏ não sau của amygdala chỉ được phát hiện gần đây (Melis et al., 2009b). Phản ứng này xảy ra đồng thời với sự gia tăng nồng độ dopamine ngoài tế bào trong chất thẩm phân thu được từ vỏ của hạt nhân accumbens, như được tìm thấy sau khi tiêm oxytocin vào tiểu não thất (Melis et al., 2009b). Cơ chế mà oxytocin tiêm vào nhân vỏ não sau của amygdala gây ra sự cương cứng dương vật hiện chưa rõ. Các dữ liệu có sẵn cho thấy cả sự cương cứng dương vật và sự gia tăng nồng độ dopamine ngoài tế bào trong chất thẩm phân thu được từ nhân accumbens đều qua trung gian bởi sự kích hoạt các thụ thể oxytocinergic, vì cả hai phản ứng đều bị loại bỏ bởi chất đối kháng thụ thể oxytocin d (CHXNUM). Tôi) 2-Orn5-vasotocin được tiêm vào nhân amygdala vài phút trước khi oxytocin (Melis et al., 2b).
Bất cứ cơ chế nào oxytocin kích hoạt trong nhân vỏ não sau của amygdala, phản ứng tình dục gây ra bởi peptide đều bị hủy bỏ bởi sự phong tỏa của tất cả các thụ thể dopaminergic với cis-flupenthixol được tiêm vào vỏ của hạt nhân. +) MK-801 được tiêm vào vùng não thất, nhưng không vào vùng chứa nhân, như được tìm thấy khi cương cứng dương vật do oxytocin tiêm vào tiểu não thất (Melis et al., 2009b). Điều này cho thấy rằng oxytocin được tiêm vào nhân sau của amygdala kích hoạt sự dẫn truyền thần kinh axit glutamic ở vùng não thất. Điều này gây ra lần lượt kích hoạt các tế bào thần kinh dopaminergic mesolimbic, dẫn đến cương dương vật. Theo quan điểm của các nghiên cứu cho thấy các con đường thần kinh liên kết hạt nhân này của amygdala với tiểu não thất (Canteras et al., 1995; French và Totterdell, 2003), những phát hiện này cho thấy khả năng tương tác có thể tồn tại giữa hai vùng não này, mặc dù trực tiếp các con đường từ amygdala hoặc đến nhân accumbens hoặc đến vùng não thất đã được mô tả (Kelley và Domesick, 1982; Witter, 2006).
KHAI THÁC. Tủy sống
Tủy sống là một khu vực khác của hệ thần kinh trung ương có chứa các sợi và thụ thể oxytocinergic (Freund-Mercier et al., 1987; Uhl-Bronner et al., 2005), trong đó oxytocin gây cương cứng dương vật (Tang et al., 1998 ; Veronneau-Longueville và cộng sự, 1999; Giuliano và Rampin, 2000; Giuliano và cộng sự, 2001). Như đã nhắc lại ở trên, các sợi oxytocinergic này bắt nguồn từ nhân trung tâm của vùng dưới đồi và góp phần làm giảm các con đường kiểm soát các tế bào thần kinh tự chủ cột sống làm trung gian cương dương vật. Thật vậy, các sợi này tạo ra các tiếp xúc synap trong các cột tế bào giao cảm và giao cảm sừng sừng phía trước ở ống ngực và thắt lưng với các tế bào thần kinh cột sống có liên quan đến hang động dương vật (Marson và McKenna, 1996, Giuliano, Rul , 2000). Những tiếp xúc synap này đã được chứng minh bằng cách dán nhãn các tế bào thần kinh cột sống có nguồn gốc từ dương vật và đến tủy sống bằng các chất đánh dấu ngược cụ thể được tiêm vào hang cavernous, kết hợp với nghiên cứu kính hiển vi huỳnh quang kép và nghiên cứu kính hiển vi laser đồng tiêu (Tang et al., 1998; et al., 1999). Theo các nghiên cứu này, ở những con chuột đực được gây mê đã tiêm một liều oxytocin tích lũy ở lumbo-sacral, nhưng không phải ở mức độ thắt lưng ngực, áp lực nội tiết được khơi dậy tăng theo cách phụ thuộc liều. Những tác dụng này đã bị xóa bỏ bởi sự phong tỏa thụ thể oxytocinergic với d (CH2) 5Tyr (Me) 2-Orn8-vasotocin và theo phần của các dây thần kinh vùng chậu (Giuliano và Rampin, Những kết quả này chứng minh rằng oxytocin, hoạt động ở tủy sống lumbo, làm tăng áp lực nội sọ và gợi ý rằng oxytocin, được giải phóng trong quá trình kích hoạt sinh lý của nhân paraventricular là một chất kích hoạt mạnh mẽ của các tế bào thần kinh cương cứng cột sống. Điều thú vị là, các tế bào thần kinh tủy sống cương dương mà oxytocin hoạt động để phát huy tác dụng cương dương của nó, cũng nhận được các tiếp xúc synap từ các tế bào thần kinh serotoninergic có nguồn gốc từ paragigantocellularis của sự hình thành võng mạc của tế bào tủy ., 1992). Sự phá hủy các tế bào thần kinh serotoninergic tạo điều kiện cho sự xuất tinh và phản xạ dương vật ở chuột đực (Marson và McKenna, 1992; Yells et al., 1992). Vì thuốc kích thích thụ thể 5HT2C tạo điều kiện thuận lợi cho việc cương dương vật khi được tiêm thuốc vào cơ thể, nhưng không được tiêm thuốc vào cơ thể, nhưng không được tiêm thuốc vào cơ thể, không được tiêm thuốc vào nhau. xem Stancampiano và cộng sự, 5 và tài liệu tham khảo trong đó), người ta cũng đề xuất rằng oxytocin tạo điều kiện cho hoạt động của các thụ thể 2HT5C ở mức độ của tủy sống lumbo (Stancampiano et al., XN). Ngoài ra, oxytocin có thể ảnh hưởng đến hoạt động của các tế bào thần kinh serotoninergic giảm dần bằng cách tác động trực tiếp vào nhân paragigantocellularis, nơi các tế bào thần kinh này bắt nguồn (xem Stancampiano và cộng sự, 1994).
KHAI THÁC. Tương tác giữa oxytocin, dopamine và axit glutamic trong hệ thần kinh trung ương và cương dương vật
Như đã nhắc lại trong Phần 1, tất cả các tế bào thần kinh oxytocinergic có trong hệ thống thần kinh trung ương đều bắt nguồn từ nhân paraventricular và các cấu trúc xung quanh. Hoạt động của các tế bào thần kinh này nằm dưới sự kiểm soát của các chất dẫn truyền thần kinh và / hoặc neuropeptide khác nhau. Trong số các nghiên cứu nhiều nhất ở cấp độ paraventricular là dopamine, axit glutamic, axit gamma-aminobutyric (GABA), oxit nitric, endocannabinoids, peptide opioid, peptide giải phóng hormone tăng trưởng, peptide liên quan đến VGF. Dopamine, axit glutamic, peptide giải phóng hormone tăng trưởng, peptide có nguồn gốc VGF và oxytocin là chất kích thích, ví dụ, các hợp chất này và chất chủ vận của chúng tạo điều kiện cho dương vật cương cứng khi được tiêm vào nhân paraventricular, trong khi GABA, peptide opioid hoặc chất chủ vận của họ ức chế cương dương vật (xem Meisel và Sachs, 1994; Witt và Insel, 1994; Argiolas và Melis, 1995, 2004, 2005; Giuliano và Rampin, 2000, 2004; McKenna, 2000; Andersson, 2001;
Một số dòng bằng chứng thực nghiệm cho thấy các tế bào thần kinh oxytocinergic này và các chất dẫn truyền thần kinh và neuropeptide nói trên có liên quan đến việc kiểm soát chức năng cương dương và hành vi tình dục trong các bối cảnh sinh lý khác nhau. Hơn nữa, oxytocin được giải phóng ở các vùng não ngoài vùng dưới đồi, chẳng hạn như vùng não thất, vùng đồi thị và các vùng của nó, amygdala và tủy sống có thể ảnh hưởng đến hoạt động của các tế bào thần kinh mà oxytocinergic đồng bộ. Tại thời điểm này, các tế bào thần kinh duy nhất quan trọng cho sự cương cứng dương vật mà các khớp thần kinh oxytocinergic impinge, được xác định một cách chắc chắn, là các cơ quan tế bào của các tế bào thần kinh dopaminergic mesolimbic của vùng vỏ não của vỏ não. ; Succu và cộng sự, 2007), và các tế bào thần kinh tủy sống cương cứng chiếu từ ống lumbo đến xương ức (xem Giuliano và Rampin, 2008; Giuliano et al., 2000) (xem thêm Đoạn 2001) . Thật vậy, mặc dù các khớp thần kinh và thụ thể oxytocinergic cũng đã được xác định trong tiểu não thất, amygdala và tủy sống, những khu vực đều quan trọng đối với sự cương cứng dương vật (xem ở trên), ở những khu vực này có loại chất dẫn truyền thần kinh / s mà kết thúc thần kinh oxytocinergic impinge, vẫn chưa được biết.
Phần tổng quan này tóm tắt ngắn gọn các tài liệu gần đây về các cơ chế ảnh hưởng đến khả năng cương dương của oxytocin được tiêm vào vùng vây bụng của đuôi, vùng dưới đồi của vùng đồi thị và ở tủy sống. Đặc biệt chú ý đến sự tương tác của peptide với dopamine và axit glutamic ở những khu vực này và về vai trò tương tác này có thể đóng vai trò kiểm soát trung tâm chức năng cương dương. Một bản tóm tắt ngắn gọn về tác dụng của dopamine và axit glutamic đối với các tế bào thần kinh oxytocinergic trong nhân paraventricular, cũng đóng vai trò chính trong chức năng cương dương được đưa ra trước tiên, để người đọc nhận thức được tình trạng nghiên cứu ban đầu trong lĩnh vực này, như để người đọc nhận thức được tình trạng nghiên cứu sớm trong lĩnh vực này, như những nghiên cứu này đã được xem xét rộng rãi (xem Argiolas và Melis, 1995, 2004, 2005; Melis và Argiolas, 2003). Cũng trong trường hợp này, sự chú ý đặc biệt được dành cho các kết quả gần đây nhất, điều này cho thấy vai trò quan trọng đối với cả liên kết dopamine-oxytocin và liên kết axit glutamic-oxytocin không chỉ trong hoạt động tình dục (cương dương vật và giao hợp), mà còn trong tình dục kích thích và động lực tình dục.
KHAI THÁC. Tương tác Dopamine-oxytocin trong nhân paraventricular
Sản phẩm khả năng của chất chủ vận dopamine để gây cương cứng dương vật bằng cách kích hoạt các tế bào thần kinh oxytocinergic trung ương đã được đề xuất ngay lập tức sau khi phát hiện ra rằng apomorphin gây ra sự cương cứng dương vật khi được tiêm vào nhân tế bào (Melis et al., 1987) khi đối kháng thụ thể oxytocin d (CH2) 5Tyr (MeX). có thể làm giảm sự cương cứng gần như hoàn toàn do dương vật gây ra không chỉ bởi oxytocin được cung cấp, mà còn bởi apomorphin, được tiêm dưới da (Argiolas et al., 2b). Những kết quả này được theo sau bởi những nghiên cứu khác cho thấy kết quả tương tự khi d (CH8) 1987Tyr (Me) 2-Orn5-vasotocin được tiêm icv và apomorphin được tiêm trực tiếp vào nhân paraventricular (Melis et al.đề nghị rằng các chất chủ vận dopamine gây ra sự cương cứng dương vật bằng cách kích hoạt các tế bào thần kinh oxytocinergic paraventricular chiếu vào các khu vực não ngoài vùng dưới đồi và đặc biệt là tủy sống (xem Argiolas và Melis, 1995, 2004, 2005). Theo giả thuyết này, ở những con chuột bị gây mê, sự phong tỏa các thụ thể oxytocinergic lumbo-sacral bằng một chất đối kháng thụ thể oxytocin không peptide đã được tìm thấy gần đây có khả năng loại bỏ apomorine gây ra bởi sự ức chế apomorine gây ra bởi áp lực nội bào. Con đường oxytocinergic paraventriculo-cột sống liên quan đến cương dương vật (Baskerville et al., 2009).
Các nghiên cứu nhằm vào Xác định thụ thể dopamine chịu trách nhiệm cho việc gây cương cứng dương vật, tiết lộ rằng cũng trong các chất chủ vận thụ thể dopamine nhân paraventricular gây ra cương cứng dương vật bằng cách tác động lên thụ thể dopamine của gia đình D2, như được tìm thấy với các chất chủ vận thụ thể dopamine được đưa ra một cách có hệ thống (xem Melis và cộng sự, 1987; Eaton và cộng sự, 1991; Melis và Argiolas, 1995a). Theo đó, apomorphin, một chất chủ vận thụ thể D1 / D2 hỗn hợp mạnh, và quinpirole, một chất chủ vận chọn lọc D2receptor mạnh, nhưng không phải là chất đồng vận của thụ thể D38393 chọn lọc, mà không phải là thuốc gây ung thư thụ thể D1 chọn lọc. và phản ứng tình dục gây ra bởi các chất chủ vận thụ thể D2 này đã bị bãi bỏ bởi các chất đối kháng thụ thể D2, như haloperidol và l-sulpiride, nhưng không phải bởi SCH 23390, một chất đối kháng thụ thể D1 chọn lọc (Melis et al. Khả năng của apomorphin gây ra sự cương cứng dương vật khi được tiêm vào nhân paraventricular cũng được xác nhận bởi các nghiên cứu từ xa cho thấy chất chủ vận dopamine được đưa vào nhân paraventricular có thể làm tăng áp lực nội bào ở chuột đực. , 1987; Giuliano và Allard, 1999), như được tìm thấy sau khi tiêm hệ thống (Bernabè et al., 2001). Những nghiên cứu này cũng khẳng định vai trò chính của các thụ thể D2, vì các chất chủ vận thụ thể D1 thường được tìm thấy là không thể làm tăng áp lực nội bào khi được tiêm vào nhân paraventricular (Chen và cộng sự, 1999).
Một số dòng bằng chứng thực nghiệm sau đó đã cho thấy rằng các thụ thể D2 ở hai bên, có sự kích thích gây ra sự cương cứng dương vật, nằm trên các tế bào của các tế bào thần kinh oxytocinergic. Đầu tiên, nhân paraventricular chứa các đầu dây thần kinh dopaminergic thuộc về các tế bào thần kinh dopaminergic incertohypothalamic. Các tế bào của các tế bào thần kinh này nằm trong nhóm A13 và A14 của Dahlstrom và Fuxe (1964), cung cấp rộng rãi và bẩm sinh một số cấu trúc vùng dưới đồi, bao gồm các tế bào thần kinh oxytocinergic paraventricular chiếu vào phân tích thần kinh và / hoặc đến các vùng não ngoài vùng dưới đồi (Bùijs et al.
Sự tham gia của các tế bào thần kinh dopaminergic này ở cấp độ paraventricular trong việc kiểm soát sự cương cứng và giao hợp của dương vật được hỗ trợ bởi các nghiên cứu vi phân cho thấy nồng độ của dopamine ngoại bào và axit 3,4-dihydroxyphenylacetic (DOPAC). nhân paraventricular của chuột đực mạnh mẽ tình dục cho thấy sự cương cứng không tiếp xúc khi đặt trong sự hiện diện của một phụ nữ tiếp nhận estrogen + progesterone không thể tiếp cận (Melis et al., 2003).
Sự gia tăng nồng độ dopamine và DOPAC thậm chí còn cao hơn khi cho phép giao hợp với nữ tiếp nhận (Melis và cộng sự, 2003), như được tìm thấy ở khu vực tiền sản trung gian (Hull và cộng sự, 1995) và trong nhân tế bào (Pfaus và Everitt, 1995). Thứ hai, một số nghiên cứu cho thấy sự cương cứng dương vật gây ra bởi sự kích thích của thụ thể D2 paraventricular, được trung gian bởi oxytocin được giải phóng ở những khu vực này. Theo đó, apomorphin được dùng với liều gây cương cứng dương vật, đã được tìm thấy có thể làm tăng nồng độ oxytocin, không chỉ trong huyết tương của chuột và khỉ (Melis et al., 1989a; Cameron et al., 1992), mà còn ở não ngoài vùng dưới đồi các khu vực, chẳng hạn như hải mã (Melis et al., 1990). Để phù hợp với những kết quả này, apomorphin được tiêm vào nhân paraventricular với liều gây ra sự cương cứng dương vật gần đây đã được chứng minh là có thể làm tăng nồng độ dopamine ngoài tế bào trong nhân accumbens, tác dụng làm giảm chất đối kháng thụ thể oxytocin d (CHX) 2Tyr (Me) 5-Orn2-vasotocin được tiêm vào vùng não bụng (Succu et al., 8; Melis et al., 2007a) (xem thêm Phần 2009). Thứ ba, các tổn thương điện giải hai bên của nhân paraventricular, loại bỏ gần như hoàn toàn oxytocin khỏi các vùng não ngoài vùng dưới đồi (Hawthorn et al., 4), loại bỏ sự cương cứng của dương vật do apomorphin gây ra (Argiolas et al. được đưa vào tâm thất bên, nhưng không vào nhân paraventricular, làm giảm sự cương cứng dương vật phụ thuộc liều lượng với một tiềm năng song song với các hợp chất này trong việc ngăn chặn các thụ thể oxytocin (Melis et al., 1985b). Thuốc đối kháng thụ thể Oxytocin cũng cực kỳ mạnh trong việc làm giảm sự thuận lợi của hành vi tình dục nam gây ra không chỉ bởi oxytocin, mà còn bởi apomorphin (Argiolas et al., 1987, 1989).
Theo cơ chế mà các thụ thể D2 được kích hoạt bởi dopamine hoặc bởi các chất chủ vận thụ thể dopamine, làm tăng hoạt động của các tế bào thần kinh oxytocinergic, do đó giải phóng oxytocin ở vùng não ngoài màng cứng và trong tủy sống, nhiều dữ liệu thực nghiệmđưa ra giả thuyết rằng sự kích thích thụ thể D2 làm tăng nồng độ các ion Ca2 + nội bào bên trong cơ thể tế bào của các tế bào thần kinh oxytocinergic, dẫn đến việc kích hoạt nitric oxide-synthase, một loại enzyme phụ thuộc Ca2 + -calmodulin, có trong các tế bào này (Vincent và Kimura, 1992; Torres và cộng sự, 1993; Sanchez và cộng sự, 1994; Sato-Suzuki và cộng sự, 1998). Việc sản xuất oxit nitric tăng gây ra lần lượt kích hoạt các tế bào thần kinh oxytocinergic. Theo đó, sự cương cứng dương vật bị apomorineine gây ra đã bị ngăn chặn bởi các thuốc chẹn kênh canxi hữu cơ và by_-conotoxin GVIA, một chất chặn mạnh mẽ và chọn lọc của các kênh Ca1 + phụ thuộc vào điện áp của loại N (McCleskey et al., 2). hạt nhân (xem Argiolas và cộng sự, 1987 và các tài liệu tham khảo trong đó); (1990) cương dương vật do apomorphin gây ra đã bị ngăn chặn bởi các chất ức chế nitric oxide-synthase được đưa vào nhân paraventricular (Melis et al., 2c); và (1994) apomorphin và các chất chủ vận thụ thể D3 khác được đưa ra ở liều gây ra sự cương cứng dương vật làm tăng sản xuất oxit nitric trong phương pháp thẩm tách paraventricular thu được bằng cách tiêm vi-rút trong não. Thụ thể D2 cương cứng dương vật cương cứng (Melis et al., 2). Cơ chế bằng cách sử dụng oxit nitric kích hoạt các tế bào thần kinh oxytocinergic nội bào, vẫn chưa được biết, mặc dù dữ liệu có sẵn cho thấy oxit nitric hoạt động như một chất truyền tin nội bào và không liên quan đến cyclase guanylate. Thật vậy, chất tương tự phosphodiesteraseresistant hoạt động của GMP theo chu kỳ, 1996-bromo-cyclic, đã được tìm thấy không thể gây ra sự cương cứng dương vật khi được đưa vào nhân paraventricular (Hình. 8) (xem Melis và Argiolas, 2b).
Giải thích trên thường được coi là không thuyết phục, chủ yếu là do sự kích thích thụ thể dopamine D2 thường được kết hợp với sự ức chế hơn là kích thích cơ thể tế bào của các tế bào thần kinh có chứa các thụ thể này thông qua các cơ chế kết hợp protein G khác nhau (xem Sokoloff và Schwartz, 1995). Tuy nhiên, một lời giải thích có thể cho sự khác biệt này, phù hợp với sự kích thích trực tiếp của các tế bào thần kinh oxytocinergic paraventricular bởi dopamine, đã được đề xuất gần đây bởi một khám phá về một Dopamine kết hợp protein G thụ thể, một thành viên của họ thụ thể D2 (D2, D3 và D4), sự kích thích làm tăng dòng Ca2 + trong các chế phẩm tế bào chứa phiên bản nhân bản của phân nhóm thụ thể này (Moreland et al., 2004). Quan trọng hơn, một chất chủ vận thụ thể D4 chọn lọc (ví dụ ABT 724) (N-methyl-4- (2-cyanophenyl) piperazynil-3methylbenzamide maleate) đã được tìm thấy có khả năng gây cương dương vật 2004). Hiệu ứng này không được tìm thấy với chất chủ vận phụ thụ thể D2 chọn lọc PNU-95666E (R-5,6-dihydro-N, N-dimethyl-4Himidazo [4,5,1-i] quinolin-5-amine) cũng không thể tăng dòng Ca2004 + trong các chế phẩm tế bào chứa phiên bản nhân bản của phân nhóm thụ thể D2 (Brioni et al., 4; Moreland et al., 2004). Phù hợp với giả thuyết và phát hiện trên, PD 2004 (N-methyl-168,077- (4-cyanophenyl) piperazynil-2methylbenzamide maleate, PIP-3EA (3-xy-xê-rê-xê-rê-rê-rê-rê [2-a] pyridine) và các chất chủ vận thụ thể D4 chọn lọc khác (Heier et al., 2; Melis et al., 1b; Löber et al., 1,2), đã được tìm thấy có thể gây cương cứng dương vật khi tiêm vào hệ thống, ic hạt nhân paraventricular, mặc dù kém hiệu quả hơn apomorphin. Tác dụng chống cương dương của các chất chủ vận thụ thể D4 này đã bị ngăn chặn bởi L-1997 (2006- (2009- [chlorophenyl] piperazin-4-yl) -methyl-745,870H-pyrrolo [3-pyrroline nhân vật phản diện (Patel et al., 4; Melis và cộng sự, 1, 1b; Löber et al., 2,3).
Cuối cùng, tác dụng chống cương dương của các chất chủ vận thụ thể D4 ở trên cũng bị giảm bởi các chất ức chế nitric oxide-synthase, được đưa vào nhân paraventricular, và bởi d (CH2) 5Tyr (Me) 2Tyr cho icv nhưng không có trong nhân paraventricular. Những kết quả này phù hợp với giả thuyết rằng các chất chủ vận thụ thể D8 cũng kích thích các tế bào thần kinh oxytocinergic bằng cách kích hoạt nitric oxide-synthase, và giải phóng oxytocin trong các khu vực não ngoại vi, điều này tạo điều kiện cho sự cương cứng của dương vật, như thể cho thấy sự cương cứng của dương vật. ., 4, 2b; Löber et al., 2005).
Những phát hiện trên cũng ủng hộ giả thuyết rằng dopamine gây ra sự cương cứng dương vật bằng cách tác động lên các thụ thể D4 nằm trên cơ thể tế bào của các tế bào thần kinh oxytocinergic paraventricular, và gây ra sự gia tăng Ca2 + vào các tế bào của tế bào thần kinh oxytocinergic. Oxit nitric lần lượt kích hoạt các tế bào thần kinh oxytocinergic để giải phóng oxytocin ở các vùng não ngoài vùng dưới đồi và trong tủy sống, như đã thảo luận. Về vấn đề này, Đáng chú ý là các thụ thể dopamine đã được xác định trong các tế bào của các tế bào thần kinh oxytocinergic trong nhân paraventricular chỉ gần đây bởi các nghiên cứu miễn dịch huỳnh quang kép với các kháng thể thụ thể D2, D3 và D4 chọn lọc cao và với các kháng thể oxytocin. Các nghiên cứu này đã cho thấy sự biểu hiện của cả ba phân nhóm D2receptor (D2, D3 và D4), cùng nằm cục bộ trong các tế bào của các tế bào thần kinh oxytocinergic trong nhân tế bào trung tâm (và cả trong tế bào nhân) và Douglas, 2008; Baskerville và cộng sự, 2009).
Điều này cung cấp hỗ trợ thần kinh mạnh mẽ cho khả năng các chất chủ vận thụ thể dopamine và dopamine của loại D2 gây ra sự cương cứng dương vật bằng cách kích hoạt các tế bào thần kinh oxytocinergic chiếu trực tiếp vào các vùng não ngoài màng cứng, ví dụ, tủy sống, vùng não thất, hồi hải mã và amygdala. Tuy nhiên, những phát hiện này không cung cấp bất kỳ trợ giúp nào cho việc xác định phân nhóm thụ thể D2 / s, mà sự kích thích gây ra phản ứng cương dương. Thật không may, không có sự giúp đỡ nào có thể nhận được ngay cả từ các nghiên cứu nhằm xác định các tế bào thần kinh oxytocinergic được kích hoạt bởi chất chủ vận thụ thể dopamine trong nhân paraventricular. Thật vậy, mặc dù hoạt động khác nhau trên các phân nhóm thụ thể dopamine khác nhau, hoặc các chất chủ vận thụ thể dopamine hỗn hợp (ví dụ, apomorphin), hoặc chất chủ vận thụ thể D2 chọn lọc (ví dụ, quinpirole, tác dụng trên tất cả các tiểu thụ thụ thể D2) sự kích hoạt của các tế bào thần kinh oxytocinergic, được đo bằng sự gia tăng protein FOS trong các tế bào thần kinh oxytocinergic parvocellular của nhân paraventricular (Bitner et al., 4). Tuy nhiên, phát hiện này gần đây đã được đặt câu hỏi, vì sự gia tăng protein FOS trong các tế bào thần kinh oxytocinergic paraventricular chỉ được tìm thấy khi sự cương cứng dương vật được tạo ra bởi quinerolane, hoạt động chủ yếu trên các tiểu phần thụ thể D2006 và D2, chứ không phải bởi PDPUMX mặc dù khả năng của cả hai hợp chất gây ra phản ứng tình dục (Baskerville và cộng sự, 3).
Các thí nghiệm tiếp theo với các chất chủ vận chọn lọc của các phân nhóm thụ thể D2 khác (chủ yếu là D2 và D3) sau đó là cần thiết để xác định vai trò chính xác của từng phân nhóm thụ thể dopamine trong việc kiểm soát chức năng cương dương ở cấp độ bán cầu. Về vấn đề này, như đã được nhắc lại trước đây, apomorphin, hoạt động mạnh mẽ trên tất cả các phân nhóm thụ thể dopamine (xem Brioni và cộng sự, 2004, và các tài liệu tham khảo trong đó), có hiệu quả hơn nhiều so với thụ thể D4 trong việc gây cương dương vật khi tiêm thuốc vào dương vật. Trung tâm. Điều này có thể được giải thích bằng ái lực cao hơn của apomorphin trên các thụ thể D4 khi so sánh với các chất chủ vận thụ thể D4 được thử nghiệm, hoặc cách khác, các chất chủ vận thụ thể D4 có thể đóng vai trò là chất chủ vận thụ thể D4, hoặc chất đồng vận của thụ thể D2. tạo ra sự kích hoạt cao hơn các tế bào thần kinh oxytocinergic làm trung gian cương dương vật, so với việc kích hoạt bởi các chất chủ vận thụ thể D4 của phân nhóm thụ thể DXNUMX.
Tương tác giữa các thụ thể dopamine D1 và D2 đã được mô tả trong việc kiểm soát hành vi tình dục ở cấp độ của khu vực tiền sản trung gian (xem Hull và cộng sự, 1989). Trong trường hợp không có khả năng các chất chủ vận thụ thể D2 chọn lọc gây ra sự cương cứng dương vật (Hsieh et al., 2004) sẽ được xác nhận (nhưng xem Depoortère et al., 2009), ví dụ, ngay cả sau khi tiêm các hợp chất này vào hạt nhân , một vai trò chính cho các thụ thể D3 một mình hoặc cùng với các thụ thể D4 trong việc kích hoạt các tế bào thần kinh oxytocinergic làm trung gian cương dương vật nên được phân tích chi tiết (xem Baskerville và cộng sự, 2009). Thật không may, các chất chủ vận thụ thể D2 và D3 chọn lọc (ví dụ, khác nhau về ái lực của chúng đối với hai phân nhóm thụ thể này trong ít nhất bốn / năm bậc cường độ trong ống nghiệm) hiện không có sẵn. Vì lý do này, đề xuất gần đây rằng các thụ thể D3 làm trung gian sự cương cứng dương vật gây ra bởi các chất chủ vận thụ thể D2 cổ điển, chủ yếu dựa vào khả năng của các chất đối kháng thụ thể D3 đặc trưng trong các thí nghiệm in vitro, để làm giảm sự cương cứng dương vật do các thí nghiệm in vitro gây ra. apomorphin, quinpirole và pramipexole, có khả năng kích hoạt tất cả các phân nhóm thụ thể dopamine D2 (Collins et al., 2), chắc chắn cần phải được xác nhận với các thí nghiệm khác. Việc xác nhận này cũng cần thiết vì không có tác dụng của chất chủ vận thụ thể D4 đối với sự cương cứng dương vật trong nghiên cứu này, trái ngược với kết quả của các nghiên cứu được trích dẫn ở trên, chứng minh tác dụng cương dương của chất chủ vận thụ thể D4. Thật vậy, ngay cả khả năng của apomorphin gây ra sự cương cứng dương vật (và ngáp) ở những con chuột bị loại bỏ thụ thể D4 với một tiềm năng giống hệt như ở chuột bị loại bỏ thụ thể D4 hoang dã và khả năng của thuốc đối kháng thụ thể D3 để loại bỏ các chất đối kháng thụ thể D2009 Collins và cộng sự, 3) không thể được coi là một bằng chứng rõ ràng cho vai trò chọn lọc của phân nhóm thụ thể D2 trong cương cứng dương vật do thụ thể DXNUMX gây ra. Các loài khác nhau, các nghiên cứu với chất dẫn truyền thần kinh / neuropeptide và / hoặc dẫn truyền thần kinh / cắt bỏ gen thụ thể neuropeptide (dẫn truyền thần kinh / neuropeptide và / hoặc dẫn truyền thần kinh / động vật kích thích thụ thể thần kinh) thường gây thêm nhầm lẫn và biến chứng của động vật kích thích thụ thể thần kinh / hoặc neuropeptide và các thụ thể của chúng. Cụ thể, quá trình cắt bỏ gen oxytocin tạo ra oxytocin hạ gục những con chuột giao phối và giao phối bình thường, như thể oxytocin không cần thiết cho việc giao phối và giao phối. Ngoài ra, chuột đồng loại oxytocin đồng hợp tử cho thấy giao phối và sinh sản bình thường, mặc dù có sự suy yếu rõ rệt về sự suy giảm sữa (Nishimori et al., 1996; Young et al., 1996). Sự cắt bỏ gen mã hóa tế bào thần kinh nitric oxide synthase, cũng tạo ra nitric oxide synthase hạ gục những con chuột giao phối và giao phối bình thường (Huang và cộng sự, 1993). Tuy nhiên, những phát hiện này có thể chỉ ra một tính năng quan trọng của sinh lý sinh sản, tức là sự dư thừa của các hệ thống liên quan đến sự kiểm soát của nó ở cấp trung ương và ngoại vi.
Sự dư thừa như vậy chắc chắn có nguồn gốc tiến hóa, vì nó đảm bảo việc truyền gen cho thế hệ tiếp theo cho sự tồn tại của loài. Do đó, thực tế rằng việc cắt bỏ gen thụ thể D4 không làm thay đổi tác dụng cương dương của apomorphin, cho thấy các thụ thể D4, như oxytocin và oxit nitric, chỉ là một vài trong số các chất trung gian hoạt động trong hệ thống kiểm soát chức năng cương dương, thay vì cho thấy rằng không có vai trò cho các thụ thể này trong việc kiểm soát cương dương vật và hành vi tình dục. Một nghiên cứu khác gần đây đã được báo cáo bởi một nghiên cứu khác (Depoortère et al., 4). Tuy nhiên, trái ngược với công việc của Collins et al. (2009), và để làm cho bức tranh về vai trò của các phân nhóm thụ thể D2009 khác nhau trong việc kiểm soát sự cương cứng của dương vật thậm chí còn khó hiểu hơn, nghiên cứu này cũng cho thấy các chất đối kháng thụ thể D2 chọn lọc được đưa ra một cách có hệ thống không thể làm giảm sự cương dương của thụ thể. chuột đực của chủng nhạy cảm hơn với tác dụng cương dương của apomorphin, trong khi phản ứng tình dục (và ngáp) được đối kháng bởi chất đối kháng D3 chọn lọc L-2 (741,626 - [[3- (4-chlorophenyl) -4-yl] methyl-4H-indole), dẫn đến các tác giả đề xuất rằng thụ thể D1, chứ không phải là thụ thể D1 và D2, là những người đóng vai trò chính trong cương dương vật gây ra bởi chất chủ vận D3 (Depoortère et. Cuối cùng, khả năng tác dụng kích thích của chất chủ vận thụ thể dopamine đối với tế bào thần kinh oxytocinergic làm trung gian cương dương vật, ít nhất là một phần, là gián tiếp chứ không phải trực tiếp, ví dụ, qua trung gian hoặc bị ảnh hưởng bởi sự thay đổi hoạt động của các chất dẫn truyền thần kinh khác có thể điều chỉnh hoạt động của các chất dẫn truyền thần kinh khác Các tế bào thần kinh oxytocinergic trong nhân paraventricular, không thể loại trừ hoàn toàn.
KHAI THÁC. Tương tác axit glutamic-oxytocin trong nhân paraventricular
Nhân paraventricular của vùng dưới đồi rất giàu các khớp thần kinh có chứa một axit amin kích thích như một chất dẫn truyền thần kinh (ví dụ, axit glutamic và axit aspartic) (Văn Đen Pol, 1991). Các axit amin kích thích trong nhân này có liên quan đến nhiều chức năng, bao gồm cương dương vật và hành vi tình dục (Roeling et al., 1991; Melis et al., 1994b, 2000, 2004b). Theo đó, axit N-methyl-d-aspartic (NMDA), một chất chủ vận chọn lọc của phân nhóm thụ thể NMDA, nhưng không (±) -_- amino-3-hydroxy-5-methyl-isoxazole-4-propionic acid (AMPA) , một chất chủ vận chọn lọc của phân nhóm thụ thể AMPA hoặc (±) -trans (1) -amino-1,3-cyclopentane dicarboxilic acid (ACPD), một chất chủ vận chọn lọc của phân nhóm thụ thể metabotropic, được tìm thấy trong khả năng của thụ thể metabotropic hạt nhân paraventricular của chuột di chuyển tự do (Melis et al., 1994b). Hiệu ứng cương dương của 948 MR Melis, A. Argiolas / Neuroscience và Biobehavioral Nhận xét 35 (2011) 939-955 NMDA đã bị ngăn chặn bởi (+) MK-801, một chất đối kháng thụ thể NMDA không cạnh tranh. ), tiêm vào nhân paraventricular (Melis et al., 1987b). Để phù hợp với những kết quả này, trong các nghiên cứu đo từ xa nhằm theo dõi áp lực nội bào, NMDA đã được tìm thấy hoạt động mạnh hơn nhiều so với các chất chủ vận của các phân nhóm thụ thể axit amin kích thích khác khi tiêm vào nhân paraventricular trong tăng áp lực nội bào trong thức tỉnh hoặc gây mê al., 1994; Chen và Chang, 2000).
Như đã đề xuất ở trên đối với oxytocin và dopamine, có khả năng các thụ thể NMDA làm trung gian cương dương vật nằm trong cơ thể tế bào của các tế bào thần kinh oxytocinergic, vì các đầu dây thần kinh axit amin kích thích nằm trên cơ thể tế bào oxytocinergic (Văn Đen Pol, 1991). Tương tự như những gì được tìm thấy với chất chủ vận thụ thể dopamine, tác dụng cương dương của NMDA rõ ràng là qua trung gian bởi sự kích hoạt dẫn truyền thần kinh oxytocinergic, bị hủy bỏ bởi chất đối kháng oxytocin d (CH2) 5Tyr (Me) không vào nhân trung tâm (xem Argiolas và Melis, 2, 8, 1995 và các tài liệu tham khảo trong đó). Tương tự như vậy, việc kích hoạt dẫn truyền thần kinh oxytocinergic do NMDA gây ra cũng là thứ yếu so với việc kích hoạt nitric oxide-synthase, do việc cương dương vật do NMDA gây ra bị ngăn chặn bởi các chất ức chế nitric oxide-synthase (N-Nitro-N-methyll-arginine methyl-thio-l-citrulline) được đưa vào nhân paraventricular, và NMDA được tiêm vào nhân paraventricular ở liều gây ra sự cương cứng dương vật, làm tăng sản xuất oxit nitric trong nhân hypothalamic (xem Argiolas và Melis, 1995 ). Đối với các chất chủ vận thụ thể dopamine, NMDA gây ra sự kích hoạt nitric oxidesynthase cũng có thể là thứ yếu đối với dòng Ca2 + tăng trong các tế bào tế bào oxytocinergic thông qua các thụ thể NMDA được ghép kênh, như đã thấy trong một số chế phẩm thần kinh Southam và Garthwaite, 2; Schuman và Madison, 1992 và các tài liệu tham khảo trong đó). Oxit nitric lần lượt kích hoạt dẫn truyền thần kinh oxytocinergic (xem ở trên). Nguồn gốc của các phép chiếu glutamatergic kích hoạt các tế bào thần kinh oxytocinergic trung gian cương cứng dương vật cương cứng là không rõ, mặc dù một số bằng chứng về thần kinh học và điện sinh lý cho thấy chúng có thể bắt nguồn, ít nhất là một phần, trong hippocampus . Mặc dù công việc tiếp theo là cần thiết để mô tả rõ hơn nguồn gốc của các phép chiếu glutamatergic đối với nhân paraventricular, sự tham gia của axit glutamic trong nhân paraventricular trong kiểm soát cương dương vật và hành vi tình dục được hỗ trợ rõ ràng bằng các nghiên cứu vi phân. Theo đó, nồng độ ngoài tế bào của axit glutamic và axit aspartic đã tăng lên trong chất thẩm phân thu được từ nhân paraventricular của chuột đực cho thấy sự cương cứng không tiếp xúc khi đưa vào sự hiện diện của chuột cái không thể tiếp cận được estrogen + progesterone. 2004b), cương dương vật cũng được trung gian bởi sự kích hoạt truyền oxytocinergic trung ương (Melis et al., 1999a, b). Sự gia tăng này được tìm thấy thậm chí còn cao hơn khi giao hợp với nữ tiếp nhận được cho phép (Melis và cộng sự, 2004a). Phù hợp với giả thuyết rằng sự gia tăng hoạt động của các axit amin kích thích xảy ra trong nhân paraventricular trong quá trình cương dương và giao hợp, cả sự cương cứng không tiếp xúc và hành vi điều hòa (trong đó xảy ra sự cương cứng của dương vật copula) và sự giảm này được theo sau bởi sự gia tăng sản xuất oxit nitric xảy ra trong hạt nhân dưới đồi này trong các bối cảnh sinh lý (Melis et al., 2000). Sự gia tăng nồng độ axit glutamic ngoại bào thứ phát do giảm giải phóng GABA từ các đầu dây thần kinh GABAergic trên các tế bào thần kinh GABAergic được đặt trên các tế bào của axit amin kích thích được gắn vào các tế bào của tế bào oxytocinergin, cũng được tìm thấy trong các tế bào của tế bào oxytocin. được đưa vào tâm thất bên hoặc trực tiếp vào nhân trung tâm với liều gây ra sự cương cứng dương vật (xem Succu et al., 1; Castelli et al., 1). Sự gia tăng này dẫn đến việc kích hoạt nitric oxide-synthase trong cơ thể tế bào của các tế bào thần kinh oxytocinergic, làm tăng sản xuất oxit nitric. Oxit nitric lần lượt kích hoạt các tế bào thần kinh oxytocinergic làm trung gian cương dương vật như mô tả ở trên. Theo cơ chế như vậy, SR 141761A gây ra sự cương cứng dương vật đã bị giảm do sự phong tỏa các thụ thể NMDA và bởi các chất ức chế nitric oxide-synthase, nhưng không bị phong tỏa bởi các thụ thể dopamine hoặc oxytocin trong nhân tế bào. thụ thể oxytocin trung ương bởi chất đối kháng thụ thể oxytocin cho icv
KHAI THÁC. Tương tác oxytocin-dopamine ở vùng não thất
Oxytocin gây ra sự cương cứng dương vật khi được tiêm vào phần đuôi của vùng phía bụng bụng theo cách phụ thuộc vào liều (Melis và cộng sự, 2007). Liều hoạt động thấp nhất được tiêm đơn phương là 20 ng, trong khi liều cao nhất được thử nghiệm là 100 ng. Hiệu ứng oxytocin được trung gian bởi sự kích hoạt các thụ thể oxytocinergic, vì phản ứng tình dục bị hủy bỏ bằng cách tiêm chất đối kháng oxytocin d (CH2) 5Tyr (Me) 2-Orn8-vasotocal. Những thụ thể này được định vị trong các tế bào của các tế bào thần kinh dopaminergic, dự án chủ yếu đến vỏ của các hạt nhân accumbens. Theo đó, các nghiên cứu miễn dịch huỳnh quang kép (1) cho thấy rằng trong các sợi oxytocinergic vùng vây bụng có liên hệ chặt chẽ với các tế bào của các tế bào thần kinh dopaminergic, phần lớn trong số đó được dán nhãn dương tính với tyrosine-hydroxylase tiêm vào vỏ của hạt nhân accumbens (Melis et al., 2007) và (2) vùng bụng của vùng dương vật do oxytocin gây ra xảy ra đồng thời với sự gia tăng nồng độ của dopamine ngoài tế bào trong vỏ lọc của tế bào. các hạt nhân accumbens (Melis et al., 2007). Sự cương cứng dương vật do oxytocin gây ra cũng xảy ra đồng thời với sự gia tăng sản xuất oxit nitric ở vùng não thất, cả hai phản ứng đối kháng không chỉ bởi d (CH2) 5Tyr (Me) 2-Orn8-vas methyl-thio-l-citrulline, mà còn bởi _-conotoxin, chất ức chế kênh Ca2 + phụ thuộc vào điện áp và bởi ODQ (1H- [1,2,4] oxadiazole [4,3-a] quinoxalin-1-one) cyclase, tất cả được đưa ra trong khu vực vây bụng trước oxytocin (Succu et al., 2008). Vì nhiều cơ quan tế bào dopaminergic được dán nhãn Fluorogold tiếp xúc với các sợi oxytocinergic, được phát hiện dương tính với tyrosine hydroxylase ở vùng đuôi não, cũng được dán nhãn tích cực cho nitric oxide-synthase và guanylate cyclase (Succ. cương cứng dương vật gây ra có thể được trung gian theo cơ chế sau đây. Sự kích hoạt các thụ thể oxytocinergic trong cơ thể tế bào dopaminergic bởi peptide làm tăng dòng Ca2 + bên trong cơ thể tế bào của các tế bào thần kinh dopaminergic. Điều này kích hoạt nitric oxide-synthase, một loại enzyme phụ thuộc Ca2 + -calmodulin, do đó làm tăng sản xuất oxit nitric. Oxit nitric lần lượt kích hoạt guanylate cyclase, dẫn đến tăng nồng độ của chu kỳ GMP. Cyclic GMP kích hoạt các tế bào thần kinh dopaminergic chiếu vào nhân accumbens. Vai trò của GMP theo chu kỳ trong quá trình cương dương vật được tạo ra bởi oxytocin được tiêm vào vùng phía sau của vây đuôi cũng được hỗ trợ bởi khả năng của 8-bromo-cyclic GMP, một chất tương tự kháng phosphodiesterase hoạt động của thuốc cyclic. khu vực vây bụng và để tăng dopamine ngoài tế bào trong thẩm tách từ nhân accumbens (Succu et al., 2008; Melis et al., 2009a). Điều này không đúng với cơ chế mà oxit nitric kích hoạt các tế bào thần kinh oxytocinergic trong nhân paraventricular, là 8-bromo-cyclic GMP không thể gây cương cứng dương vật khi được tiêm vào nhân này (Melis và Argiolas, 1995b). 2). Đối với các con đường thần kinh được kích hoạt bởi dopamine trong nhân accumbens dẫn đến cương dương vật, những điều này vẫn chưa được biết. Tuy nhiên, dữ liệu có sẵn cho thấy những con đường này kích hoạt sự dẫn truyền thần kinh dopamine trong nhân paraventricular của vùng dưới đồi. Theo đó, sự cương cứng dương vật do oxytocin gây ra xảy ra đồng thời với sự gia tăng dopamine ngoài tế bào không chỉ trong hạt nhân accumbens, mà còn trong nhân paraventricular, và được đối kháng bởi chất đối kháng thụ thể dopamine haloperidol tiêm vào nhân paraventricular (Melis et al., 2007). MộtKết hợp lại với nhau, những kết quả này hỗ trợ cho ý tưởng rằng các tế bào thần kinh oxytocinergic có nguồn gốc từ nhân paraventricular và chiếu đến vùng phía sau của vây đuôi, khi được kích hoạt giải phóng oxytocin ở khu vực này, do đó kích hoạt hệ thống tín hiệu GMP NO-cyclic, từ đó kích hoạt các tế bào thần kinh dopaminergic mesolimbic (Melis và cộng sự, 2007, 2009a; Succu và cộng sự, 2008). Dopamine được giải phóng trong nhân accumbens lần lượt kích hoạt các con đường thần kinh dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic incothalamic, kích thích các tế bào thần kinh oxytocinergic paralentularular chiếu vào dương vật cương cứng. Đồng thời, dopamine được giải phóng trong nhân paraventricular cũng có thể kích hoạt các tế bào thần kinh oxytocinergic chiếu đến các khu vực não ngoài vùng dưới đồi như vùng não thất, vùng đồi thị, vùng đồi thị và có lẽ các vùng não khác.
Như đã nhắc lại ở trên, phù hợp với giả thuyết này, apomorphin được tiêm vào nhân paraventricular với liều gây ra sự cương cứng dương vật cũng làm tăng nồng độ dopamine ngoài tế bào trong nhân accumbens, tác dụng làm giảm chất đối kháng thụ thể oxytocin d (CHX) ) 2-Orn5-vasotocin được tiêm vào vùng não thất (Succu et al., 2; Melis et al., 8a). Cùng với nhau, các con đường thần kinh ở trên có thể tạo thành một mạch thần kinh giả thuyết liên quan đến dopamine, oxytocin và các chất dẫn truyền thần kinh khác (ví dụ, axit glutamic, xem bên dưới) ảnh hưởng không chỉ đến hiệu suất tình dục, mà còn cả động lực tình dục và bổ ích tình dục (xem Phần 4).
KHAI THÁC. Tương tác axit oxytocin-glutamic trong tiểu não thất của đồi hải mã
Oxytocin gây ra sự cương cứng dương vật khi được tiêm vào tâm thất, nhưng không phải ở phần dưới lưng, theo cách phụ thuộc liều (Melis et al., 2009b). Phản ứng tình dục được trung gian bởi sự kích thích các thụ thể oxytocin, bị bãi bỏ bằng cách tiêm d (CH2) 5Tyr (Me) 2-Orn8-vasotocin được đưa vào cùng một vị trí của oxytocin. . Đối với việc bản địa hóa các thụ thể này, dữ liệu có sẵn cho thấy rằng chúng được định vị trong các tế bào của các tế bào thần kinh giàu nitric oxide-synthase.
Theo đó, các nghiên cứu vi phân cho thấy sự cương cứng dương vật do oxytocin gây ra xảy ra đồng thời với sự gia tăng sản xuất oxit nitric trong tiểu não thất, và sự gia tăng này không chỉ được loại bỏ bằng cách tiêm thuốc ức chế nitric oxidesynthase trước đó. và bởi hemavebin nitric oxide scavenger, mà còn bởi d (CH2) 5Tyr (Me) 2-Orn8-vasotocin, tất cả được đưa vào cùng một vị trí của oxytocin với liều đối kháng dương vật (Melis et. Quan trọng hơn, sự cương cứng dương vật do oxytocin gây ra cũng xảy ra đồng thời với sự gia tăng nồng độ axit glutamic ngoài tế bào trong tiểu não thất, chỉ được đối kháng một phần bởi chất đối kháng thụ thể NMDA không cạnh tranh (+) MK-2010 được đưa vào tiểu não thất (Melis et al., 801). Cùng với nhau, những kết quả này cho thấy rằng oxit nitric mới được hình thành, bằng cách hoạt động như một chất truyền tin nội bào, kích hoạt sự dẫn truyền thần kinh axit glutamic dẫn đến cương dương vật, có thể thông qua các dự báo chất lỏng thần kinh từ tâm thất subiculum đến các khu vực não ngoài đồi thị. Phù hợp với giả thuyết này, NMDA được tiêm vào tiểu não thất gây ra sự cương cứng dương vật theo cách phụ thuộc vào liều và tác dụng này được đối kháng hoàn toàn bằng cách tiêm trước vào cùng một vị trí (+) MK-801, nhưng không phải bởi S-methyl-thio-l-citrulline , huyết sắc tố hoặc d (CH2) 5Tyr (Me) 2-Orn8-vasotocin (Melis et al., 2010). Đối với các con đường chất lỏng thần kinh dự phóng đến các khu vực não ngoài đồi thị được kích hoạt bởi các axit amin kích thích (ví dụ, axit glutamic) trong tiểu não thất, có khả năng đây là các glutamatergic, như là phần lớn của các dự báo tràn máu vùng đồi thị. Hiện tại, có thể chỉ đề xuất rằng những dự đoán này điều chỉnh hoạt động của các tế bào thần kinh dopaminergic mesolimbic, từ đó điều chỉnh hoạt động của các tế bào thần kinh dopaminergic incothalamic trong nhân paraventricular, dẫn đến việc kích hoạt các tế bào thần kinh oxy hóa. trung gian cương cứng dương vật như đã thảo luận (xem ở trên).
Theo đó, sự cương cứng dương vật gây ra bởi tiểu não thất oxytocin xảy ra đồng thời với sự gia tăng nồng độ của dopamine ngoài tế bào trong vỏ của các hạt nhân, và sự gia tăng này, như sự cương cứng của dương vật, bị hủy bỏ bởi d (CH2) 5 Orn2-vasotocin được đưa vào tiểu não thất trước oxytocin (Melis et al., 8b). Ngoài ra, do tiểu não do oxytocin gây ra sự cương cứng dương vật cũng bị giảm bởi (+) MK-2009 được tiêm vào vùng não thất, nhưng không đi vào nhân tế bào (Melis et al., 801b) và xảy ra đồng thời với sự gia tăng của tế bào Axit glutamic trong vùng não thất, nhưng không phải trong vùng tụ nhân, cả hai phản ứng đều bị hủy bỏ bởi d (CH2009) 2Tyr (Me) -Orn5-vasotocin, được tiêm vào tiểu não trước khi tiêm oxy. có khả năng kích hoạt các tế bào thần kinh dopaminergic mesolimbic là thứ yếu để tăng dẫn truyền thần kinh glutamatergic trong khu vực não thất. Điều này cho thấy rằng một tương tác axit glutamic-dopamine kiểm soát sự cương cứng dương vật tồn tại trong khu vực não thất. Các nghiên cứu tiếp theo là cần thiết để xác định xem liệu các con đường glutametergic tràn vào tiền liệt tuyến từ tiểu khung đến vùng não thất là trực tiếp hay gián tiếp, ví dụ, thông qua vỏ não trước trán hoặc các vùng não khác (xem Melis et al., 2009b). Vì nhân paraventricular cũng nhận được các phép chiếu glutamatergic từ vùng đồi thị (xem ở trên và Saphier và Feldman, 1987), và Axit glutamic kích hoạt các tế bào thần kinh oxytocinergic paraventricular bao gồm cả những người chiếu đến khu vực não thất (xem Argiolas và Melis, 2005 và các tài liệu tham khảo trong đó), và Oxytocin ở vùng não thất gây ra sự cương cứng dương vật và làm tăng hoạt động của tế bào thần kinh dopaminergic mesolimbics (xem ở trên), thật hấp dẫn khi suy đoán rằng pCác tế bào thần kinh oxytocinergic araventricular cũng có thể được tham gia, ít nhất là một phần, trong việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic bằng cách tiêm oxytocin vào tiểu não thất (xem Phần 4).
KHAI THÁC. Kết luận
Các nghiên cứu được xem xét ở trên đã xác nhận và mở rộng những phát hiện ban đầu cho thấy ở chuột đực oxytocin đóng vai trò chính trong sự kiểm soát trung tâm của sự cương cứng dương vật ở cấp độ của nhân paraventricular của vùng dưới đồi và của tủy sống. Đặc biệt, nhiều nhấtcent nghiên cứu cho thấy oxytocin ảnh hưởng đến sự cương cứng dương vật bằng cách hoạt động ở các khu vực não khác, ví dụ, khu vực não thất, tiểu não thất và nhân vỏ não sau của amygdala.
Ở cấp độ paraventricular, phát hiện mới quan trọng nhất có lẽ là phát hiện ra sự biểu hiện của tất cả các thụ thể dopamine của họ D2 (D2, D3 và D4) trong các tế bào của tế bào thần kinh oxytocinergic trong nhân tế bào khu vực tiền sản trung gian) (Baskerville và Douglas, 2008; Baskerville và cộng sự, 2009). Điều này cung cấp thần kinh mạnh mẽ ủng hộ giả thuyết rằng các chất chủ vận thụ thể dopamine và dopamine có thể kích hoạt các tế bào thần kinh oxytocinergic trực tiếp liên quan đến chức năng cương dương và chiếu không chỉ vào tủy sống, mà còn đến các vùng não ngoài vùng dưới đồi. Về vấn đề này, điều quan trọng nữa là phát hiện ra rằng sự gia tăng áp lực của chất chủ vận thụ thể dopamine gây ra do áp lực nội bào bị giảm do sự phong tỏa các thụ thể oxytocinergic trong tủy sống lumbo (Baskerville et al., 2009). Thật vậy, mặc dù những bằng chứng như vậy đã thu được ở những con chuột đực được gây mê, phát hiện này xác nhận sự kích hoạt của một con đường giảm dần oxytocinergic paraventriculo-cột sống liên quan đến cương dương vật thụ thể dopamine gây ra. HTuy nhiên, vẫn chưa xác định được liệu cương cứng dương vật gây ra bởi sự kích thích thụ thể dopamine có trong cơ thể tế bào oxytocinergic là thứ yếu để kích hoạt một phân nhóm thụ thể dopamine cụ thể của họ D2 (D2, D3) hoặc nếu các phân nhóm thụ thể này hợp tác trong việc điều chỉnh phản ứng cương dương, có thể theo các cách khác nhau tùy thuộc vào bối cảnh xảy ra cương dương vật (xem Moreland et al., 2004; Enguehard-Gueiffier et al., 2006; Melis et al., 2006a, b; Löber et al., 2009; Collins và cộng sự, 2009; Depoortère và cộng sự, 2009; Baskerville và cộng sự, 2009).
Một phát hiện mới quan trọng khác là oxytocin gây ra sự cương cứng dương vật khi được tiêm không chỉ vào nhân paraventricular hoặc trường CA1 của vùng đồi thị, mà còn vào khu vực phía trước của não thất, vỏ não thất và vỏ não sau. Những vùng não này đã không được thử nghiệm trong các nghiên cứu trước đó cho thấy oxytocin làm tăng tình trạng cương dương vật tự phát ở chuột đực, mặc dù chúng nhận được như các dự báo oxytocinergic tủy sống từ hạt nhân. Oxytocin thực sự đã được tìm thấy có khả năng làm tăng các giai đoạn cương dương vật tự phát, xảy ra ở chuột đực trưởng thành trong trường hợp không có bất kỳ kích thích tình dục nào, chẳng hạn như bắt nguồn từ sự hiện diện của một con chuột có thể tiếp cận hoặc không thể tiếp cận được (estrogen-progesterone) Thao tác của cơ quan sinh dục, khi được tiêm vào nhân paraventricular và trường CA1 của đồi hải mã, nhưng không phải trong tiểu khung lưng, vách ngăn bên, nhân caudate, vùng tiền đình trung thất, nhân trung thất. , 1986). Trong tất cả các nghiên cứu, sự cương cứng dương vật thường được tính khi dương vật nhô ra khỏi vỏ dương vật bởi một người quan sát không biết về các phương pháp điều trị được đưa ra trực tiếp trong thí nghiệm hoặc sau đó bằng cách quan sát thí nghiệm được ghi trên băng video bằng thiết bị máy quay video. Mỗi giai đoạn cương dương vật kéo dài trong tối thiểu 0.5-1 và thường đi kèm với việc chải chuốt dương vật và / hoặc uốn cong hông. Không có thí nghiệm nào thường được thực hiện ở những con chuột này để xác định hiệu quả của trải nghiệm tình dục, tuổi tác hoặc nếu những con chuột này có thể được chia thành những người đáp ứng thấp hoặc cao với tác dụng cương dương của oxytocin được tiêm vào các vùng não khác nhau. Điều này cũng áp dụng cho phần lớn các nghiên cứu về tác dụng cương dương của các neuropeptide khác và các loại thuốc làm tăng các giai đoạn cương dương vật tự phát, bao gồm các chất chủ vận dopamine, axit amin kích thích, ACTH-MSH, hexarelin và VGF peptide. Tuy nhiên, tác dụng cương dương của các hợp chất này đã được xác nhận nhiều lần bằng các phương pháp đo từ xa, xác định sự xuất hiện của cương dương vật bằng cách tăng áp lực nội bào xảy ra một cách tự nhiên hoặc sau khi sử dụng các hợp chất này theo các tuyến khác nhau, ví dụ, theo hệ thống, nội sọ hoặc trực tiếp vào các hạt nhân não cụ thể, sau khi cấy một microtransducer áp lực trực tiếp vào khoang hang (xem Bernabè et al., 1999). Ở vùng não thất, tiểu não thất và nhân sau của amygdala cũng oxytocin gây cương cứng dương vật bằng cách tác động lên các thụ thể oxytocinergic. Điều này dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic bắt nguồn từ vùng não thất và chiếu vào vỏ của các hạt nhân accumbens, được đo bằng sự gia tăng nồng độ dopamine ngoài tế bào trong chất thẩm phân thu được từ vỏ hạt nhân và bằng cách giảm phản ứng cương dương gây ra bởi peptide được tiêm vào các khu vực ngoại vi này, được tìm thấy sau khi phong tỏa thụ thể dopaminergic trong hạt nhân accumbens (xem bên dưới). Đối với các cơ chế được kích hoạt bởi sự kích thích các thụ thể oxytocinergic ở những vùng não này, dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic và cương cứng dương vật, điều được làm rõ nhất là những trường hợp xảy ra ở vùng vây bụng. Thật vậy, ở đây, kết quả dược lý và miễn dịch huỳnh quang cho thấy các đầu dây thần kinh oxytocin nằm trên cơ thể tế bào của các tế bào thần kinh dopaminergic chiếu vào vỏ của hạt nhân accumbens (Melis et al., 2007a, 2009a; Succu et al. Nhiều trong số các tế bào thần kinh này rất giàu nitric oxide synthase và guanylate cyclase. Sự kích thích các thụ thể oxytocinergic trong cơ thể tế bào của các tế bào thần kinh dopaminergic này gây ra sự kích hoạt của nitric oxide synthase dẫn đến tăng sản xuất oxit nitric. Oxit nitric lần lượt kích hoạt guanylate cyclase, do đó làm tăng nồng độ của chu kỳ GMP, Điều này dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic và giải phóng dopamine trong nhân accumbens, được đo bằng sự gia tăng của dopamine ngoài tế bào trong thẩm tách từ nhân tế bào thu được từ vi lọc trong não (Succu et al. ). Dopamine được giải phóng trong nhân accumbens lần lượt kích hoạt các con đường thần kinh dẫn đến cương dương vật. Điều này được hỗ trợ bởi khả năng của thuốc đối kháng thụ thể dopamine haloperidol và / hoặc cis-flupentixol được tiêm vào nhân accumbens để làm giảm sự cương cứng của vùng bụng do oxytocin gây ra (Succu et al., 2008). Một trong những con đường pro-erectile dường như kích hoạt các tế bào thần kinh dopaminergic incothalamic, đặc biệt là các dự án đến các cơ quan tế bào của các tế bào thần kinh oxytocinergic paraventricular. Thật vậy, oxytocin được tiêm vào vùng phía sau của vây đuôi làm tăng dopamine ngoại bào không chỉ trong nhân accumbens mà còn ở nhân paraventricular, và sự phong tỏa của thụ thể dopamine trong nhân paralental , 2008, 2007; Melis và cộng sự, 2008, 2007a). Sự tồn tại của các hạt nhân accumbens dopamine-paraventricular dopamine-paraventricular oxytocin-ventral area tegmental liên kết oxytocin-dopamine cũng được đề xuất bởi khả năng của một liều apomorphin pro-erectile và của chất chủ vận thụ thể D4 PD 168077 được tiêm vào nhân paraventricular để tăng dopamine ngoài tế bào trong vỏ của hạt nhân (Succu et al., XNUM). được bãi bỏ bởi d (CH2007) 2Tyr (Me) -Orn5-vasotocin được đưa vào vùng não bụng (Melis et al., 8a, xem thêm bên dưới). Tuy nhiên, công việc tiếp theo là cần thiết để xác định các con đường thần kinh kết nối các hạt nhân với hệ thống dopaminergic incothalamic.
Cơ chế mà oxytocin gây ra sự cương cứng dương vật và kích hoạt các tế bào thần kinh dopaminergic mesolimbic khi được tiêm vào tiểu não thất hoặc vào nhân sau của amygdala chỉ được hiểu một phần tại thời điểm này. Ở những khu vực này cũng vậy, oxytocin kích hoạt các thụ thể của chính nó dẫn đến việc kích hoạt nitric oxide-synthase, do đó làm tăng sản xuất oxit nitric. Oxit nitric lần lượt kích hoạt các dự báo không rõ nguồn gốc, điều này rõ ràng làm tăng dẫn truyền thần kinh glutamatergic ở vùng não thất. Axit glutamic sau đó kích thích các tế bào thần kinh dopaminergic mesolimbic dẫn đến cương dương vật. Giả thuyết này được hỗ trợ chủ yếu bởi khả năng oxytocin được tiêm vào tiểu não thất để tăng axit glutamic ngoài tế bào ở vùng não bụng (Hình. 3) và của chất đối kháng thụ thể NMDA không cạnh tranh (+) MK-801 được tiêm vào vùng não thất, nhưng không phải trong các hạt nhân, để làm giảm sự cương cứng của dương vật do oxytocin tiêm vào trong màng não thất. của amygdala (Melis et al., 2009b). Hiện tại, có nhiều thông tin chi tiết hơn cho sự cương cứng dương vật do oxytocin gây ra. Ở đây cương cứng dương vật do oxytocin gây ra dường như thứ yếu để kích hoạt các thụ thể oxytocinergic nằm trong cơ thể tế bào của các tế bào thần kinh chứa nitric oxide-synthase. Điều này gây ra sự gia tăng sản xuất oxit nitric, kích hoạt sự dẫn truyền thần kinh glutamatergic bằng cách hoạt động như một chất truyền tin nội bào với cơ chế tương tự như mô tả cho sự tạo ra lâu dài (xem Snyder, 1992; Southam và Garthwaite, 1993; Schuman và Madison, 2004; . Theo giả thuyết này, sự cương cứng dương vật do oxytocin gây ra xảy ra đồng thời với sự gia tăng axit glutamic ngoại bào trong thẩm tách từ tiểu não thất, và kích thích thụ thể axit amin kích thích trong tiểu não thất do NMDA gây ra. Axit glutamic lần lượt kích hoạt các dự báo chất lỏng thần kinh, dẫn đến việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic ở vùng não thất, như đã báo cáo ở trên. Nếu các cơ chế này cũng hoạt động trong hạt nhân sau của amygdala hiện chưa rõ. Hơn nữa, các nghiên cứu tiếp theo là cần thiết để chứng minh rằng (1) các đầu dây thần kinh và thụ thể oxytocinergic trong tiểu não thất và trong nhân sau của amygdala được định vị trong các tế bào của các tế bào thần kinh có chứa nitric oxide-synthase, (2) dưới sự kiểm soát của các axit amin kích thích (glutamatergic) và (3) để xác định hệ thống tín hiệu được kích hoạt bởi oxit nitric. Về vấn đề này, đáng chú ý là khả năng oxytocin tăng hoạt động của nitric oxide-synthase trong cơ thể tế bào của các tế bào thần kinh dopaminergic ở vùng đuôi não và của các tế bào thần kinh chưa được biết đến trong tiểu não thất và nhân tế bào sau của tế bào thần kinh đệm. khả năng oxytocin kích hoạt nitric oxide-synthase trong cơ thể tế bào của các tế bào thần kinh oxytocinergic trong nhân paraventricular (Melis et al., 1997). Tuy nhiên, trong khi ở vùng não thất, việc sản xuất oxit nitric trong cơ thể tế bào của các tế bào thần kinh dopaminergic dẫn đến việc kích hoạt các tế bào thần kinh này bằng cách kích hoạt guanylate cyclase và tăng GMP theo chu kỳ, điều này không xảy ra trong nhân tế bào. Theo đó, 8-bromo-cyclic được tiêm vào nhân paraventricular không gây cương cứng dương vật, trong khi nó được tiêm vào vùng não thất. Một con đường truyền tín hiệu khác với hệ thống GMP nitric oxyecyclic sau đó được tham gia ở cấp độ paraventricular trong việc kích hoạt các tế bào thần kinh oxytocinergic trung gian cương dương vật bằng oxit nitric nội sinh và / hoặc ngoại sinh (Melis và Argiolas, 1995b; . 1). Mặt khác, GMP theo chu kỳ ở vùng não thất dường như cũng đóng một vai trò quan trọng trong việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic và trong sự gia tăng của dopamine ngoài tế bào xảy ra trong màng lọc của hạt nhân của chuột đực được chọn để hiển thị hoặc không hiển thị cương cứng dương vật không tiếp xúc khi đặt trong sự hiện diện của một con chuột cái không thể tiếp cận buồng trứng (estrogen + progesterone được điều trị). Trong những điều kiện thí nghiệm này, ở những con chuột đực có biểu hiện cương dương vật không tiếp xúc, sự gia tăng nồng độ dopamine ngoài tế bào được tìm thấy như mong đợi, và sự gia tăng này được tăng thêm, mặc dù chỉ một cách khiêm tốn, bởi các chất ức chế phosphodiesterase được đưa vào vùng vây bụng (Sanna et al., 2009).
Khả năng oxytocin được tiêm vào vùng não thất, trong tiểu não thất và trong nhân vỏ não sau của amygdala, cùng với chất chủ vận dopamine được tiêm vào nhân tế bào thần kinh đệm. . Đầu tiên, các cơ chế tương tự như các cơ chế được nhắc lại ở trên có thể hoạt động khi cương dương vật xảy ra trong bối cảnh sinh lý, chẳng hạn như trong quá trình giao hợp (khi xảy ra cương dương vật copula) hoặc trong quá trình cương dương vật không tiếp xúc. Những sự cương cứng này là sự cương cứng dương vật qua trung gian pheromone không thể phân biệt với những người bị gây ra bởi thuốc hoặc oxytocin, xảy ra khi chuột đực có khả năng tình dục được đưa vào sự hiện diện của một con chuột không thể tiếp cận được (chuột bị cắt bỏ estrogen + progesterone) kích thích (Sachs, 1997, 2007). Thật vậy, mặc dù những kết quả này không chứng minh rằng oxytocin ở những khu vực này đóng vai trò trong việc cương dương vật xảy ra trong bối cảnh sinh lý hoặc sau khi dùng thuốc, họ tiếp thêm sức mạnh cho những phát hiện ban đầu cho thấy những vùng não này thuộc về những nơi mà oxytocin hoạt động tập trung để tăng không chỉ các giai đoạn cương dương vật tự phát thấy sau khi dùng thuốc cương dương, mà còn cải thiện hành vi tình dục của nam (và nữ) (xem Argiolas và Melis, 2004 và các tài liệu tham khảo ở đó). Theo đó, nồng độ oxytocin tăng ở vùng đồi thị của chuột đực được điều trị bằng một liều apomorphin cương dương, một chất chủ vận dopamine cổ điển (Melis et al., 1990) và d (CH2) 5Tyr (Me) Các thụ thể, cực kỳ hiệu quả không chỉ trong việc làm suy yếu hành vi điều phối (Argiolas và cộng sự, 8a) mà còn có tác dụng hỗ trợ của apomorphin đối với hành vi điều phối của con đực (Argiolas et al., 1987b) ở chuột đực. xảy ra d (CH1987) 2Tyr (Me) -Orn5-vasotocin cũng cực kỳ mạnh trong việc làm giảm sự cương cứng không tiếp xúc ở chuột đực có khả năng tình dục, khi được đưa vào số lượng nanogram vào tâm thất bên, nhưng không vào PVN (Melis et al.
Thứ hai, các tế bào thần kinh dopaminergic mesolimbic đóng một vai trò quan trọng trong các đặc tính thúc đẩy và bổ ích của các kích thích củng cố tự nhiên, như thực phẩm, nước và hoạt động tình dục (Fibiger và Phillips, 1988; Wise và Rompre, 1989; Everitt, 1990). Cụ thể, dopamine được giải phóng từ các tế bào thần kinh này được cho là làm trung gian cho sự chuyển đổi các khía cạnh động lực của các kích thích tự nhiên thành các hành vi hướng đến mục tiêu, ví dụ trong trường hợp hoạt động tình dục, tìm kiếm bạn tình và quan hệ tình dục để đạt được sự thỏa mãn và thỏa mãn tình dục (Goto và ân sủng, 2005). Theo đó, nồng độ dopamine ngoại bào tăng trong thẩm tách từ nhân tế bào của chuột đực có khả năng tình dục khi tiếp xúc với chuột cái có khả năng tiếp nhận estrogen + progesterone không thể tiếp cận được, và mức tăng này thậm chí còn cao hơn khi chuột đực được cho phép đối phó với chuột đực (Pfaus và Everitt, 1995).
Thứ ba, các kết quả hiện tại ủng hộ giả thuyết rằng một mạch thần kinh kết nối nhân trung tâm với vùng não thất trực tiếp hoặc gián tiếp (thông qua tiểu não thất và / hoặc nhân vỏ não sau của amygdala) và nhân não con đường trở lại một lần nữa đến nhân paraventricular để kiểm soát hoạt động của các tế bào thần kinh oxytocinergic chiếu đến tủy sống trung gian cương dương vật và của các tế bào thần kinh oxytocinergic chiếu vào khu vực phía trước của não thất, phần dưới của não thất. hoạt động của các tế bào thần kinh dopaminergic mesolimbic (Hình 4). Mạch thần kinh phức tạp này có thể đóng một vai trò trong việc tích hợp các hoạt động thần kinh liên quan đến việc kiểm soát các khía cạnh tiêu dùng (cương dương-xuất tinh) và dự đoán (động lực và bổ ích) của hành vi tình dục nam trong bối cảnh sinh lý. Thật vậy, dopamine ngoài tế bào tăng trong nhân accumbens (Pfaus và Everitt, 1995) và trong nhân paraventricular của chuột đực có khả năng tình dục khi tiếp xúc với chuột cái không thể tiếp cận được, khi sự cương cứng không được tiếp cận, và thậm chí nhiều hơn khi cho phép giao hợp, ví dụ: khi xảy ra tình trạng cương dương vật copula (Melis et al., 2003). Do đó, mặc dù các nghiên cứu tiếp theo là cần thiết để làm rõ vai trò của oxytocin nội sinh ở vùng não thất, tiểu não thất và amygdala trong quá trình cương cứng và hành vi tình dục, nhưng có thể hợp lý khi cho rằng mạch thần kinh giả định này, trong khi đóng góp vào các khía cạnh tiêu dùng của hành vi tình dục, đồng thời cũng có thể kích hoạt hệ thống dopaminergic mesolimbic cung cấp chất nền thần kinh để giải thích các đặc tính bổ ích của hoạt động tình dục (Everitt, 1990; Pfaus và Everitt, 1995). Về vấn đề này, đáng chú ý là hệ thống dopaminergic mesolimbic được kích hoạt bởi oxytocin được tiêm vào vùng não thất được kích hoạt tương tự bởi các loại thuốc lạm dụng như opiates, cannabinoids, amphetamine, cocaine và rượu (Tanda et al. rằng oxytocin đã được tìm thấy có khả năng làm giảm sự dung nạp và phụ thuộc vào cocaine, morphin, rượu và cannabinoids (Kovacs et al., 1997; Cui et al., 1998). Tóm lại, có vẻ như oxytocin được giải phóng không chỉ ở vùng não thất, mà còn ở tiểu não thất và nhân vỏ não sau của amygdala, có thể kích hoạt các tế bào thần kinh dopaminergic mesolimbic, có thể liên quan đến hoạt động của các tế bào thần kinh dopaminergic . Việc kích hoạt các tế bào thần kinh dopaminergic mesolimbic có thể trực tiếp, thông qua các thụ thể oxytocinergic trong cơ thể tế bào của các tế bào thần kinh dopaminergic mesolimbic, hoặc gián tiếp thông qua việc kích hoạt dẫn truyền thần kinh axit glutamic ở vùng não thất.
Dopamine được giải phóng trong vỏ hạt nhân accumbens điều chỉnh lần lượt hoạt động của các tế bào thần kinh dopaminergic incothalamic trong nhân paraventricular gây ra sự cương cứng dương vật (thông qua kích hoạt các tế bào thần kinh oxytocinergic chiếu vào tủy sống) tế bào thần kinh chiếu đến vùng não thất, tiểu não thất hoặc nhân vỏ não sau của amygdala). Vì dopamine cũng được giải phóng trong vỏ accumbens của hạt nhân và trong nhân paraventricular khi sự cương cứng dương vật xảy ra trong bối cảnh sinh lý (ví dụ, cương cứng và giao hợp không tiếp xúc) (Succu et al., 2007; Melis et al., 2003, XNUM) có khả năng các tế bào thần kinh oxytocinergic trung ương tham gia vào các mạch thần kinh làm trung gian cho sự tương tác giữa mesolimbic và hệ thống dopaminergic incothalamic. Các mạch thần kinh này có thể đóng một vai trò không chỉ trong giai đoạn hoàn thành hành vi tình dục (ví dụ, cương dương vật và giao hợp), mà còn trong động lực tình dục, hưng phấn tình dục và thưởng cho tình dục.
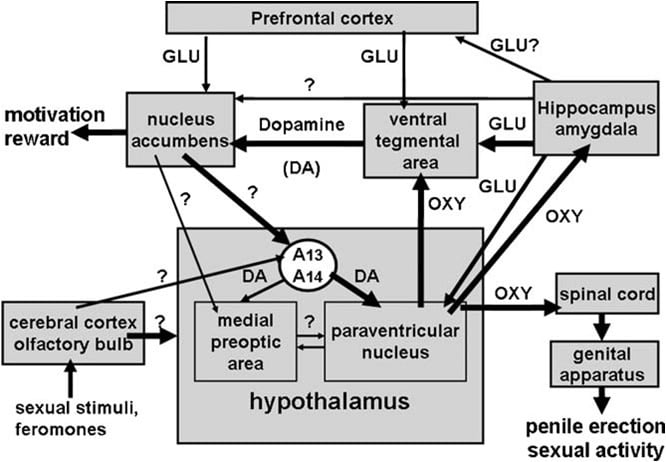
Sơ đồ biểu diễn của một mạch thần kinh giả thuyết liên quan đến oxytocin ảnh hưởng đến động lực tình dục, phần thưởng và hiệu suất tình dục, như được đề xuất bởi kết quả của chương này và các báo cáo trước đó. Các tế bào thần kinh oxytocinergic có nguồn gốc từ nhân paraventricular và chiếu vào tủy sống khi được kích hoạt ví dụ bởi dopamine và axit glutamic (nhưng cũng bởi các chất dẫn truyền thần kinh khác và / hoặc neuropeptide), tạo điều kiện cho hoạt động cương dương vật và hoạt động tình dục bằng cách kích hoạt dương vật. Dopamine và axit glutamic (nhưng cũng là chất dẫn truyền thần kinh và neuropeptide) trong nhân paraventricular cũng kích hoạt các tế bào thần kinh oxytocinergic chiếu vào khu vực não thất, do đó kích hoạt các tế bào thần kinh trung mô dopaminergic để kích thích tế bào thần kinh dopaminergic. Dopamine được giải phóng trong nhân accumbens (NA) kích hoạt lần lượt các con đường thần kinh chưa được biết đến, làm tăng hoạt động của các tế bào thần kinh dopaminergic incopalamic (có nguồn gốc từ các nhóm thần kinh A13-A14 của Dahlstrom và Fuxe). tủy sống, dẫn đến cương dương vật. Mạch này cũng có thể được kích hoạt bởi oxytocin được tiêm không chỉ vào vùng phía sau của vây đuôi, mà còn trong tiểu não thất và trong amygdala, cũng nhận được một chất bảo vệ oxytocinergic trực tiếp từ nhân trung tâm hoặc trực tiếp. khu vực tegmental, dẫn đến sự điều chỉnh của cả động lực tình dục và cương dương vật. Cuối cùng, các mạch trên cũng có thể được kích hoạt bởi các kích thích tình dục và pheromone, vì dopamine ngoài tế bào và axit glutamic tăng trong nhân paraventricular (và trong khu vực tiền sản trung gian) trong quá trình cương dương và điều trị không qua trung gian pheromone Danh sách tài liệu tham khảo).
Lời cảm ơn Công việc này được hỗ trợ một phần bởi một khoản trợ cấp từ Bộ Đại học Ý và Nghiên cứu cho AA và MRM
dự án
Andersson, KE, 2001. Dược lý cương dương vật. Dược điển. Rev. 53, 417-450. Argiolas, A., 1994. Oxit nitric là một trung gian trung gian của cương dương vật. Thần kinh học 33, 1339-1344. Argiolas, A., 1999. Neuropeptide và hành vi tình dục. Thần kinh. Biobehav. Rev. 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Chức năng trung tâm của oxytocin. Thần kinh. Biobehav. Rev. 15, 217-231. Argiolas, A., Melis, MR, 1995. Thần kinh điều trị cương dương vật: tổng quan về vai trò của dẫn truyền thần kinh và neuropeptide. Ăn xin. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. Vai trò của oxytocin và nhân paraventricular trong hành vi tình dục của động vật có vú nam. Vật lý trị liệu. Hành vi. 83, 309-317. Argiolas, A., Melis, MR, 2005. Kiểm soát trung tâm cương dương vật: vai trò của nhân trung tâm của vùng dưới đồi. Ăn xin. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. Chất đối kháng oxytocin d (CH2) 5Tyr (Me) -Orn8-vasotocin đối kháng với hành vi điều phối của con đực ở chuột. Á Âu J. Dược điển. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Sự kích thích apomorphin của hành vi điều phối đực bị ngăn chặn bởi chất đối kháng oxytocin d (CH2) 5Tyr (Me) -Orn8-vasotocin ở chuột. Dược điển. Sinh hóa. Hành vi. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. Oxytocin tiêm tĩnh mạch gây ra ngáp và cương dương vật ở chuột. Á Âu J. Dược điển. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Oxytocin: một chất cảm ứng cực kỳ mạnh mẽ của sự cương cứng dương vật và ngáp ở chuột đực. Á Âu J. Dược điển. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Tổn thương nhân paraventricular ngăn chặn ngáp và cương dương vật gây ra bởi apomorphin và oxytocin nhưng không phải do ACTH ở chuột. Brain Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vasotocin, một chất đối kháng oxytocin mạnh, đối kháng cương dương vật và ngáp do oxytocin và apomorphin gây ra, nhưng không phải bởi ACTH 1 Á Âu J. Dược điển. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. _-Conotoxin ngăn chặn sự cương cứng dương vật do apomorphin và oxytocin gây ra và ngáp ở chuột đực. Dược điển. Sinh hóa. Hành vi. 37, 253-257. Arletti, R., Bertolini, A., 1985. Oxytocin kích thích hành vi lordosis ở chuột cái. Neuropeptide 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. Oxytocin cải thiện hiệu suất điều phối của đàn ông ở chuột. Horm. Hành vi. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. Hành vi tình dục của chuột đực bị lão hóa được kích thích bởi oxytocin. Á Âu J. Dược điển. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. Bất lực tình dục có liên quan đến việc giảm sản xuất oxytocin và tăng sản xuất peptide opioid trong nhân paraventricular của vùng dưới đồi. Thần kinh. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Bằng chứng cho một hình chiếu trực tiếp từ nhân paraventricular của vùng dưới đồi đến tế bào thần kinh serotoninergic giả định của nhân paragigantocellularis liên quan đến việc kiểm soát sự cương cứng ở chuột. Á Âu J. Thần kinh. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Oxit nitric: một chất trung gian sinh lý của sự cương cứng dương vật. Khoa học 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Tương tác giữa dopamine và oxytocin trong việc kiểm soát hành vi tình dục. Ăn xin. Brain Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Tương tác Dopamine-oxytocin trong cương dương vật. Á Âu J. Thần kinh. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. Oxit nitric có liên quan đến hành vi tình dục của chuột. Á Âu J. Dược điển. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Áp lực nội bào trong quá trình cương cứng ở chuột: một cách tiếp cận tích hợp dựa trên ghi từ xa. Là. J. Vật lý trị liệu. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MW, Moreland, RB, 2006. Tín hiệu thụ thể Dopamine D4 trong nhân tế bào vùng dưới đồi chuột: bằng chứng của sự ghép đôi tự nhiên liên quan đến cảm ứng gen ngay lập tức và phosphoryl protein kinase hoạt hóa mitogen. Thần kinh học 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch 3rd., J., Kolasa, T., Polakowski , JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. Kích hoạt thụ thể dopamine D4 bằng ABT-724 gây ra sự cương cứng dương vật ở chuột. Proc Natl. Học viện Khoa học Hoa Kỳ 101, 6758-6763. Bùijs, RM, 1978. Con đường vasopressin và oxytocin trong và ngoài vùng dưới đồi ở chuột. Cell Tissue Res. 192, 423-435. Bùijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984. Sự bảo tồn dopaminergic của hạt nhân siêu âm và paraventricular. Một nghiên cứu kính hiển vi ánh sáng và điện tử. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Oxytocin tạo điều kiện cho sự tiếp thu tình dục của chuột cái được điều trị bằng estrogen. Neuropeptide 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Dopaminergic kích thích nồng độ oxytocin trong huyết tương của khỉ đực và khỉ cái bằng apomorphin và chất chủ vận thụ thể D2. J. Lâm sàng. Nội tiết. Metab. 75, 855-860. Máy ảnh, NS, Simily, RB, Swanson, LW, 1995. Tổ chức các dự báo từ hạt nhân trung gian của amygdala: một nghiên cứu PHAL trên chuột. J. Comp. Thần kinh. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Oxytocin huyết tương tăng trong phản ứng tình dục của con người. J. Lâm sàng. Nội tiết. Metab. 64, 27-31. Carter, CS, 1992. Oxytocin và hành vi tình dục. Thần kinh. Biobehav. Rev. 16, 131-144. Carter, CS, Lederhendler, II, Kirkpatrick, B., 1997. Sinh học tương tác của liên kết, Biên niên sử của Viện Khoa học New York, tập. 807. Viện hàn lâm Khoa học New York, New York. Castelli, MP, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. , XUẤT KHẨU. Các thụ thể CB1 Cannabinoid trong nhân paraventricular và kiểm soát trung tâm của cương dương vật: miễn dịch hóa, miễn dịch tự động và nghiên cứu hành vi. Khoa học thần kinh 147, 197-206. Chen, KK, Chang, LS, 2003. Tác dụng của chất chủ vận thụ thể axit amin kích thích lên sự cương cứng dương vật sau khi dùng vào nhân trung tâm của vùng dưới đồi ở chuột. J. Nước tiểu. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Gợi ý cương cứng dương vật sau khi kích hoạt sự hình thành vùng đồi thị ở chuột. Thần kinh. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Dẫn truyền thần kinh Dopaminergic tại nhân paraventricular của vùng dưới đồi trong điều hòa trung tâm của cương cứng dương vật ở chuột. J. Nước tiểu. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Rice, KC, Chồng, SM, Greedy, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen , J., Wang, S., Katz, JL, Grandy, DK, Sunahara, RK, Rừng, JH, 2009. Tác dụng chống cương dương của chất chủ vận tương tự dopamineD2 được trung gian bởi thụ thể D3 ở chuột và chuột. J. Dược điển. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Quy định trung ương của xuất tinh. Vật lý trị liệu. Hành vi. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Ngăn ngừa hội chứng cai cannabinoid bằng lithium: liên quan đến hoạt hóa tế bào thần kinh oxytocinergic. J. Thần kinh. 21, 9867-9876. Dahlstrom, A., Fuxe, K., 1964. Bằng chứng cho sự tồn tại của các tế bào thần kinh chứa monoamin trong hệ thống thần kinh trung ương. I. Trình diễn các monoza trong các tế bào của các tế bào thần kinh não. Actaol. Vụ bê bối. 62 (Bổ sung. 232), 1-54. Depoortère, R., Bardin, L., Coleues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Cương cứng dương vật và ngáp gây ra bởi chất chủ vận thụ thể giống dopamine D2 ở chuột: ảnh hưởng của chủng và sự đóng góp của dopamine D2, nhưng không phải là thụ thể D3 và D4. Hành vi. Dược điển. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Oxytocin làm giảm phản ứng amygdala đối với khuôn mặt cảm xúc bất kể hóa trị. Biol. Tâm thần 62, 11871190. Toduez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Điều chỉnh bởi amygdala trung gian của giao hợp và giải phóng dopamine trung gian tiền sản. J. Thần kinh. 21, 349-355. Donaldson, ZR, Trẻ, LJ, 2009. Oxytocin, vasopressin và các tế bào thần kinh của xã hội. Khoa học 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Thompson, JT, Moses, J., Hull, EM, 1991. Các thụ thể D2 trong nhân paraventricular điều chỉnh các phản ứng sinh dục và giao hợp ở chuột đực. Dược điển. Sinh hóa. Hành vi. 39, 177-181. Ebner, K., Bosch, OJ, Krömer, SA, Singewald, N., Neumann, ID, 2005. Sự giải phóng oxytocin trong amygdala trung tâm chuột điều chỉnh hành vi đối phó với căng thẳng và giải phóng các axit amin kích thích. Thần kinh thực vật 30, 223-230. Enguehard-Gueiffier, C., Hzigner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-Phenylpiperazin-1-yl) methyl] imidazo (di) azines như các phối tử D4 chọn lọc. Cảm ứng cương dương vật bằng 2- [4- (2-methoxyphenyl) piperazin-1-ylmethyl] imidazo [1,2-a] pyridine (PIP3EA), một chất mạnh mẽ và chọn lọc. J. Med. Hóa. 49, 3938-3947. Everitt, BJ, 1990. Động lực tình dục: một phân tích thần kinh và hành vi của các cơ chế dựa trên phản ứng thèm ăn và điều phối của chuột đực. Thần kinh. Biobehav. Rev. 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Hệ thống dopamine Mesocorticolimbic và phần thưởng. Ann. N. Y. Học viện Khoa học 5, 206-215. Tiếng Pháp, SJ, Totterdell, S., 2003. Các tế bào thần kinh phóng xạ hạt nhân riêng lẻ nhận được cả amygdala cơ bản và các liên kết dưới màng cứng ở chuột. Khoa học thần kinh 119, 19-31. 954 MR Melis, A. Argiolas / Neuroscience và Biobehavioral Nhận xét 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Tác dụng kích thích của tiêm oxytocin tiêm vào não thất đối với phản xạ tống máu sữa ở chuột. Thần kinh. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Bằng chứng điện sinh lý để kiểm soát thuận lợi các tế bào thần kinh oxytocin bằng oxytocin trong quá trình bú ở chuột. J. Vật lý trị liệu. (Thích.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Somorodendritic autoreceptors trên tế bào thần kinh oxytocin. Trong: Ivell, R., Nga, JA (Eds.), Oxytocin, Phương pháp tế bào và phân tử trong y học và nghiên cứu. Adv. Exp. Med. Biol., 365. Hội nghị toàn thể, New York và London, trang. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Đặc điểm dược lý và phân bố giải phẫu của các vị trí gắn kết oxytocin 3H trong não chuột Wistar được nghiên cứu bằng kỹ thuật autoradiography. Khoa học thần kinh 20, 599-614. Giuliano, F., Rampin, O., 2000. Kiểm soát trung tâm cương cứng dương vật. Thần kinh. Biobehav. Rev. 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamine và chức năng tình dục. Int. J. Bất lực Res. 13 (Bổ sung. 3), 18-28. Giuliano, F., Rampin, O., 2004. Kiểm soát thần kinh cương cứng. Vật lý trị liệu. Hành vi. 83, 189-201. Giuliano, F., Bernabè, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Tác dụng kích thích cột sống của oxytocin ở chuột gây mê. Am J. Vật lý trị liệu. Regul. Tích. Comp. Vật lý trị liệu. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Dopaminergic điều chế limbic và vỏ não của accumbens hạt nhân trong hành vi hướng mục tiêu. Nat. Thần kinh. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Ảnh hưởng của các tổn thương ở vùng hạ đồi, siêu nhân và siêu hạt nhân trên vasopressin và oxytocin trong não chuột và tủy sống. Brain Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Moon, MW, 1997. Tổng hợp và hoạt động sinh học của (R) -5,6-dihydro-N, N-dimethyl-4H-imidazo [4,5,1-ij] quinolin-5-amin) và các chất chuyển hóa của nó. J. Med. Hóa. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Các cơ chế trung tâm điều chỉnh sự cương cứng dương vật ở chuột có ý thức: các hệ thống dopaminergic liên quan đến tác dụng proerectile của apomorphin. J. Dược điển. Exp. Ther. 308, 330-338. Hoàng, PL, Dawson, TM, Bredt, DS, Snyder, SH, Người cá, MC, 1993. Sự phá vỡ mục tiêu của gen tổng hợp oxit nitric oxide. Ô 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Vasopressin và oxytocin kích thích các quần thể tế bào thần kinh khác biệt ở vùng trung tâm amygdala. Khoa học 308, 245-248. Hull, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. Tỷ lệ D2 / D1 ở khu vực tiền sản trung gian ảnh hưởng đến sự giao hợp của chuột đực. J. Dược điển. Exp. Ther. 251, 422-427. Hull, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Dopamine ngoài tế bào ở vùng tiền sản trung gian: ngụ ý cho động lực tình dục và kiểm soát nội tiết tố của sự giao hợp. J. Thần kinh. 15, 7465-7471. Thân tàu, EM, Meisel, RL, Sachs, BD, 2002. Hành vi tình dục của nam giới. Trong: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Eds.), Hormone, Não và Hành vi. Nhà xuất bản Học thuật, New York, trang. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, KM, 2010. Oxytocin tăng cường phụ thuộc amygdala, học tập được củng cố xã hội và sự đồng cảm về cảm xúc ở người. J. Thần kinh. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oxytocin: Phương pháp tiếp cận tế bào và phân tử trong y học và nghiên cứu. Những tiến bộ trong y học thực nghiệm và sinh học, tập. 395. Hội nghị toàn thể, New York. Kelley, AE, Domesick, VB, 1982. Sự phân bố của hình chiếu từ sự hình thành vùng đồi thị đến nhân tế bào ở chuột: một nghiên cứu về peroxidase trước và ngược dòng - horseradish peroxidase. Khoa học thần kinh 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. Kích hoạt thụ thể 5-HT (2C) là một cơ chế phổ biến về tác dụng proerectile của apomorphin, oxytocin và melanotan-II ở chuột. Á Âu J. Dược điển. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Tầm quan trọng của amygdala trung gian trong cương cứng dương vật chuột gợi lên bởi các kích thích từ xa từ con cái động dục. Hành vi. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Oxytocin và nghiện: một đánh giá. Tâm lý học tâm thần 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Oxytocin: người hỗ trợ tuyệt vời cho cuộc sống. Ăn xin. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Trình diễn hóa học chọn lọc của các hệ thống đầu cuối dopamine trong chuột di- và telencephalon: bằng chứng mới cho việc bảo tồn dopaminergic của hạt nhân thần kinh vùng dưới đồi. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Đáp ứng tình dục bị suy giảm sau các tổn thương của nhân paraventricular của vùng dưới đồi ở chuột đực. Hành vi. Thần kinh. 111, 1361-1367. Löber, S., Tschammer, N., Hzigner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Khung azulene như một bioisostere mới: thiết kế các thụ thể dopamine D4 mạnh tạo ra sự cương cứng dương vật. Hóa. Med. Hóa. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. _-Conotoxin: phong tỏa trực tiếp và liên tục các loại kênh canxi cụ thể trong tế bào thần kinh nhưng không phải cơ bắp. Bảo vệ. Nat. Học viện Khoa học Hoa Kỳ 84, 4327-4331. McKenna, KE, 2000. Một số đề xuất liên quan đến việc tổ chức kiểm soát hệ thần kinh trung ương của cương dương vật. Thần kinh. Biobehav. Rev. 24, 535-540. Marson, L., McKenna, KE, 1992. Một vai trò cho 5-hydroxytryptamine trong việc ức chế giảm dần các phản xạ tình dục cột sống. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. Các nhóm tế bào CNS liên quan đến việc kiểm soát các cơ ischiocavernosus và tubospongiosus: một nghiên cứu truy tìm transneuronal sử dụng virus pseudorabies. J. Comp. Thần kinh. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Sinh lý học của hành vi tình dục nam. Trong: Knobil, E., Neil, J. (Eds.), Sinh lý học của sinh sản, tập. 2, ấn bản thứ hai. Báo Raven, New York, trang. 3-96. Melin, P., Kihlstrom, JE, 1963. Ảnh hưởng của oxytocin đến hành vi tình dục ở thỏ đực. Nội tiết 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopamine và hành vi tình dục. Thần kinh. Biobehav. Rev. 19, 19-38. Melis, MR, Argiolas, A., 1995b. Các nhà tài trợ oxit nitric gây cương cứng dương vật và ngáp khi tiêm vào hệ thống thần kinh trung ương của chuột đực. Á Âu J. Dược điển. 294, 1-9. Melis, MR, Argiolas, A., 2003. Dẫn truyền thần kinh oxytocinergic trung ương: một mục tiêu thuốc để điều trị rối loạn cương dương psycogen. Curr. Mục tiêu ma túy 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Ngáp do oxytocin gây ra và cương dương vật: vị trí tác động trong não. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Ngáp Apomorphin gây ra và cương dương vật: vị trí của hành động trong não. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Apomorphin làm tăng nồng độ oxytocin huyết tương ở chuột. Thần kinh. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Bằng chứng là apomorphin gây ra sự cương cứng dương vật và ngáp bằng cách giải phóng oxytocin trong hệ thống thần kinh trung ương. Á Âu J. Phạmacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Tác dụng của apomorphin đối với nồng độ oxytocin ở các vùng não và huyết tương khác nhau của chuột đực. Á Âu J. Dược điển. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Sự cương cứng dương vật do Apomorphin và oxytocin gây ra và ngáp ở những con chuột đực còn nguyên vẹn và bị thiến: tác dụng của steroid tình dục. Thần kinh học 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. Cương cứng dương vật và ngáp gây ra bởi tiêm NMDA paraventricular được trung gian bởi oxytocin. Dược điển. Sinh hóa. Hành vi. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Ngăn ngừa bằng methyl ester NG-nitro-l-arginine của apomorphin- và oxytocin gây cương dương vật và ngáp: vị trí của hành động trong não. Dược điển. Sinh hóa. Hành vi. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Chất chủ vận dopamine làm tăng sản xuất oxit nitric trong nhân trung tâm của vùng dưới đồi: tương quan với cương dương vật và ngáp. Á Âu J. Thần kinh. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Oxytocin làm tăng sản xuất oxit nitric trong nhân paraventricular của vùng dưới đồi: tương quan với cương dương vật và ngáp. Reg. Peptide 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Sản xuất oxit nitric được tăng lên trong nhân paraventricular của vùng dưới đồi của chuột đực trong quá trình cương dương vật không tiếp xúc và giao hợp. Á Âu J. Thần kinh. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Chất đối kháng oxytocin d (CH2) 5Tyr (Me) 2-Orn8-vasotocin làm giảm sự cương cứng dương vật không tiếp xúc ở chuột đực. Thần kinh. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Morphine được tiêm vào nhân paraventricular của vùng dưới đồi ngăn ngừa sự cương cứng không tiếp xúc và làm suy yếu sự giao hợp: sự tham gia của oxit nitric. Á Âu J. Thần kinh. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Tác dụng của thuốc đối kháng thụ thể axit amin, dopamine và oxytocin đối với sự cương cứng dương vật không tiếp xúc và sản xuất oxit nitric paraventricular ở chuột đực. Hành vi. Thần kinh. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Dopamine ngoài tế bào tăng trong nhân paraventricular của chuột đực trong hoạt động tình dục. Á Âu J. Thần kinh. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Sự đối kháng của CB1receptors cannabinoid trong nhân paraventricular của chuột đực gây ra cương cứng dương vật. Thần kinh. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Các axit amin kích thích ngoài tế bào tăng trong nhân paraventricular của chuột đực trong hoạt động tình dục: vai trò chính củaNMDAreceptors trong chức năng cương dương. Á Âu J. Thần kinh. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, một chất chủ vận thụ thể dopamine D4 chọn lọc, gây ra sự cương cứng dương vật khi tiêm vào nhân paraventricular của chuột đực. Thần kinh. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. Sự cương cứng dương vật do SR 141716A gây ra ở chuột đực: có sự tham gia của axit glutamic paraventricular và oxit nitric. Thần kinh học 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, A., 2006b. PIP3EA và PD168077, hai chất chủ vận thụ thể dopamine D4 chọn lọc, gây ra sự cương cứng dương vật ở chuột đực: vị trí và cơ chế hoạt động trong não. Á Âu J. Thần kinh. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Oxytocin được tiêm vào vùng não thất gây ra sự cương cứng dương vật và làm tăng dopamine ngoài tế bào trong nhân accumbens MR Melis, A. Argiolas / Neuroscience và Biobehavioral Nhận xét 35 (2011) 939-955 955 và nhân paraventricular của vùng dưới đồi của chuột đực. Á Âu J. Thần kinh. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Vai trò của oxytocin trong các giai đoạn dự đoán và hoàn thành hành vi tình dục của chuột đực. Trong: Jastrow, H., Feuerbach, D. (Eds.), Sổ tay nghiên cứu Oxytocin: Tổng hợp, lưu trữ và phát hành, hành động và các dạng thuốc. Nhà xuất bản Nova, New York, Hoa Kỳ, trang. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Oxytocin được tiêm vào tiểu não thất hoặc nhân vỏ não sau của amygdala gây ra sự cương cứng dương vật và làm tăng dopamine ngoại bào trong nhân tế bào của chuột đực. Á Âu J. Thần kinh. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. Oxytocin gây cương cứng dương vật khi tiêm vào tiểu não thất: vai trò của oxit nitric và axit glutamic. Thần kinh học 58, 1153-1160. Moreland, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, EJ, Helfrich, RJ, Namovic, MT, El-Kouhen, OF, Thạc sĩ, JN, Brioni, JD, 2004. Dược lý so sánh của các dòng tế bào ổn định giống thụ thể dopamine D (2) của con người kết hợp với thông lượng canxi thông qua Galpha (qo5). Sinh hóa. Dược điển. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Giải phóng oxytocin và vasopressin bằng nhân tế bào trong ống nghiệm: tác dụng hỗ trợ cụ thể của oxytocin trên bản phát hành riêng của nó. J. Nội tiết. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Thay đổi bài tiết oxytocin và vasopressin trong hoạt động tình dục ở nam giới. J. Lâm sàng. Nội tiết. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Oxytocin là cần thiết cho điều dưỡng nhưng không cần thiết cho hành vi sinh sản hoặc sinh sản. Proc Natl. Học viện Khoa học Hoa Kỳ 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, F., Fletcher, AE, Knowles, M., Marwood, R., Mccallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI , XUẤT KHẨU. Hồ sơ sinh học của L 745,870, một chất đối kháng chọn lọc có ái lực cao với thụ thể dopamine D4. J. Dược điển. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Oxytocin trong các hành vi bà mẹ, tình dục và xã hội, Biên niên sử của Viện hàn lâm khoa học New York, tập. 652. Viện hàn lâm Khoa học New York, New York. Petrovic, P., Kalisch, R., Ca sĩ, T., Dolan, RJ, 2008. Oxytocin làm suy yếu các đánh giá tình cảm của khuôn mặt có điều kiện và hoạt động amygdala. J. Thần kinh. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Tâm sinh lý của hành vi tình dục. Trong: Knobil, FE, Kupfer, DJ (Eds.), Tâm lý học: Thế hệ tiến bộ thứ tư. Báo Raven, New York, trang. 742-758. Rajfer, J., Aronson, WJ, Bush, PA, Dorey, FJ, Ignarro, LJ, 1992. Oxit nitric như một chất trung gian thư giãn của corpus cavernosum để đáp ứng với dẫn truyền thần kinh nonadrenergic, noncholinergic. N. Tiếng Anh J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. Tác dụng hành vi của NMDA được tiêm vào nhân trung tâm vùng dưới đồi của chuột. Brain Res. 550, 220-224. Sachs, BD, 1997. Sự cương cứng gợi lên ở chuột đực bằng mùi hương trong không khí từ con cái động dục. Vật lý trị liệu. Hành vi. 62, 921-924. Sachs, BD, 2000. Phương pháp tiếp cận bối cảnh để sinh lý và phân loại chức năng cương dương, rối loạn cương dương và hưng phấn tình dục. Thần kinh. Biobehav. Rev. 24, 541-560. Sachs, BD, 2007. Một định nghĩa theo ngữ cảnh của hưng phấn tình dục nam. Horm. Hành vi. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. Sự cùng tồn tại của NADPH-diaphorase với vasopressin và oxytocin trong nhân tế bào thần kinh tế bào thần kinh vùng dưới đồi của chuột. Cell Tissue Res. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Các chất ức chế 5 loại Phosphodiesterase tạo điều kiện cho sự cương cứng không tiếp xúc ở chuột đực: vị trí tác động trong não và cơ chế hoạt động. J. Sex. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Tác dụng của kích thích vùng kín và vùng đồi thị đối với các tế bào thần kinh nhân paraventricular. Khoa học thần kinh 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Phản ứng ngáp rập khuôn gây ra bởi sự kích thích điện và hóa học của nhân paraventricular của chuột. J. Thần kinh. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Oxit nitric và chức năng synap. Ann. Rev. Thần kinh. 17, 153-183. Snyder, SH, 1992. Oxit nitric: đầu tiên trong một lớp dẫn truyền thần kinh mới? Khoa học 254, 494-496. Sofroniew, MV, 1983. Vasopressin và oxytocin trong não và tủy sống của động vật có vú. Xu hướng Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Các thụ thể dopamine tiểu thuyết nửa thập kỷ sau. Xu hướng Pharmacol. Khoa học 16, 270-275. Southam, E., Garthwaite, J., 1993. Con đường truyền tín hiệu nitric oxide-cyclic trong não chuột. Thần kinh học 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Cương cứng dương vật và ngáp gây ra bởi chất chủ vận 5-HT1c ở chuột đực: mối quan hệ với truyền dopaminergic và oxytocinergic. Á Âu J. Dược điển. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. Chất đối kháng thụ thể CB1 cannabinoid SR 141716A gây ra sự cương cứng dương vật bằng cách tăng axit glutamic ngoài tế bào trong nhân paraventricular của chuột đực. Hành vi. Brain Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Kích thích thụ thể dopamine trong nhân paraventricular của vùng dưới đồi của chuột đực gây ra sự cương cứng dương vật và làm tăng dopamine ngoài tế bào trong nhân accumbens: sự tham gia của oxytocin trung tâm. Thần kinh học 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Oxytocin gây ra sự cương cứng dương vật khi được tiêm vào vùng não thất của chuột đực: vai trò của oxit nitric và GMP theo chu kỳ. Á Âu J. Thần kinh. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Cannabinoids và heroin kích hoạt truyền dopamine mesolimbic bằng cơ chế thụ thể opioid mu1 phổ biến. Khoa học 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Bảo tồn oxytocinergic và serotoninergic của hạt nhân lumbosacral xác định kiểm soát cương dương vật ở chuột đực. Khoa học thần kinh 82, 241-254. Theodosis, DT, 1985. Thiết bị đầu cuối miễn dịch oxytocin đồng bộ trên các tế bào thần kinh oxytocin trong hạt nhân siêu âm. Thiên nhiên (Luân Đôn) 313, 682-684. Tindall, JS, 1974. Kích thích gây ra sự giải phóng oxytocin. Trong: Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Eds.), Sổ tay Sinh lý học. Giáo phái. 7, Nội tiết, tập. IV. Hiệp hội Sinh lý học Hoa Kỳ, Washington DC, trang. 257-267. Torres, G., Lee, S., Rivier, C., 1993. Bản chất của synthase oxit hypothalamic chuột và colocalization với neuropeptide. Mol. Ô. Thần kinh. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Biểu hiện dị hình giới tính của các vị trí gắn oxytocin ở tiền đình và tủy sống của chuột. Khoa học thần kinh 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Phân phối so sánh của vasopressin V1b và các axit ribonucleic thụ thể oxytocin trong não. Nội tiết 139, 5015-5033. Văn Đen Pol, A., 1991. Glutamate và aspartate miễn dịch trong các sợi trục thần kinh dưới đồi. J. Thần kinh. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G., Giuliano, F. , XUẤT KHẨU. Bảo tồn oxytocinergic của hạt nhân tự trị kiểm soát cương dương vật ở chuột. Khoa học thần kinh 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Lập bản đồ mô học của nitric oxide synthase trong não chuột. Khoa học thần kinh 46, 755-784. Wagner, CK, Clemens, LG, 1993. Con đường chứa Neurophysin từ nhân paraventricular của vùng dưới đồi đến một hạt nhân vận động lưỡng hình tình dục trong tủy sống thắt lưng. J. Comp. Thần kinh. 336, 106-116. Winslow, JT, Insel, TR, 1991. Địa vị xã hội trong các cặp khỉ sóc đực quyết định phản ứng hành vi với chính quyền oxytocin trung tâm. J. Thần kinh. 11, 2032-2038. Khôn ngoan, RA, rompre, P.-P., 1989. Dopamine não và phần thưởng. Ann. Rev. Thần kinh. 40, 191-225. Witt, DM, Insel, TR, 1994. Hành vi tình dục của nam giới kích hoạt protein giống như c-fos trong các tế bào thần kinh oxytocin trong nhân paraventricular của vùng dưới đồi. J. Neuroendocrinol. 6, 13-18. Witter, MP, 2006. Các kết nối của subiculum của chuột: địa hình liên quan đến tổ chức cột và tầng. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. Sự tương tác giữa MK-801 và các thụ thể đối với N-methyl-d-aspartate: hậu quả chức năng. Thần kinh học 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Oxytocin chủ yếu kích thích các tế bào thần kinh oxytocin giả định trong nhân siêu chuột trong ống nghiệm. Brain Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992. Các tổn thương của nhân paragigantocellularis: ảnh hưởng đến hành vi giao phối ở chuột đực. Brain Res. 596, 73-79. Trẻ, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Sự thiếu hụt oxytocin trên chuột ngăn cản việc tống sữa, nhưng không gây ra khả năng sinh sản hoặc đẻ. J. Neuroendocrinol. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Người vận chuyển, S., 2000. Tăng áp lực dương vật nội bào sau khi tiêm chất chủ vận thụ thể axit amin kích thích trong nhân trung tâm vùng dưới đồi của chuột gây mê. J. Nước tiểu.