 Martin Zack
Martin Zack- 1Phòng thí nghiệm tâm sinh lý nhận thức, Khoa khoa học thần kinh, Trung tâm nghiện và sức khỏe tâm thần, Toronto, ON, Canada
- 2Chương trình Khoa học thần kinh tịnh tiến, Khoa Tâm thần học, Trường Y, Đại học Pennsylvania, Philadelphia, PA, Hoa Kỳ
- 3Khoa Sinh học, Khoa Thần kinh, Trung tâm Nghiện và Sức khỏe Tâm thần, Toronto, ON, Canada
Nghiện được coi là một bệnh não do tiếp xúc lâu dài với thuốc. Nhạy cảm hệ thống dopamine não (DA) một phần làm trung gian hiệu ứng này. Đánh bạc bệnh lý (PG) được coi là nghiện hành vi. Do đó, PG có thể được gây ra bởi tiếp xúc lâu dài với cờ bạc. Xác định một sự nhạy cảm do cờ bạc gây ra bởi các hệ thống DA sẽ hỗ trợ khả năng này. Phần thưởng cờ bạc gợi lên việc phát hành DA. Một phần của trò chơi xèng chuyển đổi phản ứng DA từ phân phối phần thưởng sang khởi phát tín hiệu (guồng quay) để nhận phần thưởng, phù hợp với các nguyên tắc học tập khác biệt theo thời gian. Do đó, các kích thích có điều kiện (CS) đóng vai trò chính trong các phản ứng của DA đối với cờ bạc. Ở loài linh trưởng, phản ứng DA với CS mạnh nhất khi xác suất thưởng là 50%. Theo lịch trình này, CS gợi ra một kỳ vọng về phần thưởng nhưng không cung cấp thông tin nào về việc liệu nó có xảy ra trong một thử nghiệm nhất định hay không. Trong quá trình đánh bạc, lịch biểu 50% sẽ gợi ra việc phát hành DA tối đa. Điều này phù hợp chặt chẽ với tần suất phần thưởng (46%) trên máy đánh bạc thương mại. Phát hành DA có thể góp phần gây mẫn cảm, đặc biệt đối với amphetamine. Phơi nhiễm mãn tính với một CS dự đoán phần thưởng 50% có thể bắt chước hiệu ứng này. Chúng tôi đã thử nghiệm giả thuyết này trong ba nghiên cứu với chuột. Các động vật đã nhận được các mức phơi nhiễm 15 × 45-min cho một CS dự đoán phần thưởng với xác suất là 0, 25, 50, 75 hoặc 100%. CS là một ánh sáng; phần thưởng là một giải pháp sucrose% 10. Sau khi đào tạo, chuột nhận được một chế độ nhạy cảm với năm liều riêng biệt (1 mg / kg) của d-amphetamine. Cuối cùng, họ đã nhận được thử thách 0.5 hoặc 1 mg / kg trước khi thử nghiệm hoạt động vận động 90-min. Trong cả ba nghiên cứu, nhóm 50% cho thấy hoạt động lớn hơn các nhóm khác để đáp ứng với cả hai liều thử thách. Kích thước hiệu ứng rất khiêm tốn nhưng nhất quán, được phản ánh bởi một nhóm đáng kể × liên kết xếp hạng (= 0.986, p = 0.025). Phơi nhiễm mãn tính với một lịch trình giống như cờ bạc của các kích thích dự đoán có thể thúc đẩy sự nhạy cảm với amphetamine giống như tiếp xúc với chính amphetamine.
Giới thiệu
Nghiện đã được đặc trưng như một bệnh não do tiếp xúc lâu dài với các loại thuốc lạm dụng (Leshner, 1997). Thần kinh được cho là trung gian ảnh hưởng của việc tiếp xúc như vậy (Nestler, 2001). Nhạy cảm với hệ thống dopamine não (DA) là một dạng dẻo dai thần kinh liên quan đến phản ứng siêu phản ứng với các kích thích có điều kiện (CS) đối với thuốc và tìm kiếm thuốc bắt buộc (Robinson và Berridge, 2001). Nhạy cảm đã được xác định một cách vận hành bằng cách tăng giải phóng DA để đáp ứng với CS để nhận phần thưởng và bằng cách tăng phản ứng vận động đối với thách thức DA dược lý (Robinson và Berridge, 1993; Xỏ và Kalivas, 1997; Vanderschuren và Kalivas, 2000). Mặc dù sự nhạy cảm chỉ là một trong nhiều thay đổi não liên quan đến nghiện (x. Robbins và Everitt, 1999; Koob và Le Moal, 2008), những thay đổi trong việc giải phóng dopamine trước khi sinh đã được đề xuất để đại diện cho các phản ứng thần kinh phổ biến liên quan đến tìm kiếm ma túy dựa trên nghiện (ví dụ, tái nghiện), trong đó các thuốc gây ra sự nhạy cảm vận động với thuốc phiện (ví dụ, morphine) hoặc thử thách kích thích , cũng gây ra sự phục hồi của các phản ứng của người vận hành bị dập tắt đối với việc tự quản lý heroin hoặc cocaine. Một mô hình động vật tái nghiện (Vanderschuren và cộng sự, 1999). Bằng chứng là sự nhạy cảm khuyến khích (tăng giá trị của phần thưởng thuốc) rõ rệt nhất sau khi tiếp xúc ban đầu với thuốc gây nghiện cho thấy thêm rằng sự nhạy cảm cũng có thể liên quan đến giai đoạn đầu của nghiện (Vanderschuren và Xỏ, 2010).
Cờ bạc bệnh lý (PG) đã được mô tả là nghiện hành vi và gần đây được phân loại lại cùng loại với các rối loạn phụ thuộc chất trong phiên bản 5th của Cẩm nang Chẩn đoán và Thống kê Rối loạn Tâm thần (Frascella và cộng sự, 2010; APA, 2013). Điều này ngụ ý rằng PG có thể được gây ra do tiếp xúc lâu dài với hoạt động giống như đánh bạc, rằng các cơ chế phổ biến có thể làm trung gian tác động của cờ bạc và phơi nhiễm ma túy (Zack và Poulos, 2009; Leeman và Potenza, 2012); và sự nhạy cảm của các con đường DA não có thể là một yếu tố quan trọng của quá trình này.
Bằng chứng lâm sàng gián tiếp hỗ trợ khả năng này: Sử dụng phương pháp chụp cắt lớp phát xạ positron (PET) Boileau và các đồng nghiệp đã phát hiện ra rằng các đối tượng PG nam thể hiện sự phóng thích DA lớn hơn đáng kể khi đáp ứng với amphetamine (0.4 mg / kg) so với đối chứng nam khỏe mạnh (Boileau và cộng sự, 2013). Sự khác biệt nhóm nói chung có ý nghĩa trong nhóm nghiên cứu liên kết và somatosensory. Trong các chi limbic, bao gồm các hạt nhân accumbens, các nhóm không khác nhau. Tuy nhiên, trong các đối tượng PG, DA phát hành trong limbic striatum tương quan trực tiếp với mức độ nghiêm trọng của các triệu chứng PG. Những phát hiện này phù hợp với sự nhạy cảm của các con đường DA não trong PG, nhưng cũng cho thấy một số khác biệt quan trọng với các cá nhân phụ thuộc vào chất người và với mô hình động vật cổ điển của sự nhạy cảm với amphetamine. Không giống như các đối tượng PG và động vật tiếp xúc với liều thấp amphetamine (x. Robinson và cộng sự, 1982), những người bị lệ thuộc chất liên tục thể hiện sự giải phóng DA xuống một thách thức kích thích (ROLow và cộng sự, 1997; Martinez và cộng sự, 2007), và bằng chứng từ động vật cho thấy rằng điều này có thể phản ánh sự thiếu hụt chức năng DA trong các giai đoạn kiêng khem ban đầu theo mô hình lạm dụng chất gây nghiện (Mateo và cộng sự, 2005). Trong các nghiên cứu trong đó sự nhạy cảm với chất kích thích được thể hiện ở động vật, sự phóng thích DA tăng cường thường được quan sát thấy ở các chi trên chứ không phải là vây lưng (kết hợp, somatosensory) (Vezina, 2004). Tuy nhiên, việc tìm kiếm ma túy do cue (nghĩa là có điều kiện) ở động vật liên tục tiếp xúc với cocaine có liên quan đến việc phóng thích DA tăng cường ở vây lưng, kết quả được cho là cho thấy một dạng hành vi có động cơ thường xuyên hơn (Ito và cộng sự, 2002). Do đó, độ cao tổng thể trong phát hành DA ở các vùng lưng trong các đối tượng PG có thể liên quan đến việc tìm kiếm phần thưởng dựa trên thói quen (không linh hoạt, thường xuyên) liên quan đến việc tiến triển từ vùng bụng sang vùng lưng khác của vùng vây lưng (Everitt và Robbins, 2005, tr. 1481), trong khi việc phát hành DA phụ thuộc vào mức độ nghiêm trọng trong thể vân limbic ở những đối tượng này có thể tương ứng chặt chẽ hơn với sự nhạy cảm khuyến khích như mô hình điển hình ở động vật. Các phát hiện của PET không thể tiết lộ liệu phản ứng siêu phản ứng DA là một đặc điểm có sẵn của các đối tượng PG này, hậu quả của việc tiếp xúc với cờ bạc hoặc là kết quả của một số quá trình khác hoàn toàn. Để giải quyết câu hỏi này, cần chứng minh cảm ứng nhạy cảm bằng phơi nhiễm cờ bạc mãn tính ở những đối tượng bình thường trước khi tiếp xúc. Điều này đặt ra câu hỏi là những tính năng nào của cờ bạc có nhiều khả năng gây ra sự nhạy cảm.
Skinner lưu ý rằng lịch trình tăng cường thay đổi là cơ bản cho sức hấp dẫn của cờ bạc (hoặc ít nhất là tính bền bỉ của nó) (Skinner, 1953). Hành vi đặt cược trong trò chơi máy đánh bạc tuân thủ tốt các nguyên tắc cơ bản của điều hòa công cụ, được phản ánh bởi mối tương quan tiềm năng giữa mức chi trả tiền và kích thước đặt cược trên các vòng quay liên tiếp (Tremblay và cộng sự, 2011). Do đó, đáp ứng của người vận hành tỷ lệ biến có vẻ như cung cấp một mô hình cờ bạc máy đánh bạc hợp lệ bên ngoài.
Nghiên cứu gần đây với động vật cung cấp hỗ trợ ban đầu mạnh mẽ cho tác động nguyên nhân của việc tiếp xúc với cờ bạc đối với sự nhạy cảm. Ca sĩ và đồng nghiệp đã kiểm tra tác động của 55 1, h các phiên cố định saccharin cố định (FR20) hoặc biến đổi (VR20) hàng ngày trong mô hình nhấn đòn bẩy của người vận hành đối với phản ứng vận động tiếp theo với liều thấp Sprague Dawley) chuột (Ca sĩ và cộng sự, 2012). Họ đưa ra giả thuyết rằng, nếu đánh bạc dẫn đến sự nhạy cảm, những con chuột tiếp xúc với lịch trình thay đổi, bắt chước đánh bạc, sẽ thể hiện phản ứng với amphetamine nhiều hơn so với những con chuột tiếp xúc với lịch trình cố định. Theo dự đoán, nhóm VR20 hiển thị phản ứng vận động lớn hơn so với amphetamine so với nhóm FR50. Ngược lại, các nhóm hiển thị đầu máy tương đương sau khi tiêm nước muối. Những phát hiện này xác nhận rằng việc tiếp xúc lâu dài với sự gia cố thay đổi là đủ để gây ra phản ứng siêu đối với một thách thức DA ở động vật khỏe mạnh ngẫu nhiên theo lịch trình tương ứng.
Một số câu hỏi phát sinh từ kết quả này: Đầu tiên, đến mức độ nào thì sự phụ thuộc về nhận thức, hoặc thiếu sự khác biệt giữa phản ứng của người vận hành và kết quả của nó làm trung gian cho các hiệu ứng này? Trong các thuật ngữ học tập, hiệu ứng này có liên quan đến một kết quả mong đợi về kết quả của hồi giáo hay không, có thể thấy một hiệu ứng tương tự trong trường hợp không có phản hồi của người vận hành, tức là, một sự mong đợi về kết quả kích thích trong một mô hình Pavlovian (x. Xu hướng, 1972)? Thứ hai, mức độ dự phòng giữa sự kiện tiền đề (phản ứng hoặc kích thích) và kết quả của nó có ảnh hưởng đến mức độ nhạy cảm?
Câu hỏi thứ hai liên quan đến vai trò của sự không chắc chắn trong sự nhạy cảm. Ví dụ: các trò chơi có kết quả thực sự ngẫu nhiên, hoàn toàn không thể đoán trước được, có khả năng gây ra sự nhạy cảm cao hơn các trò chơi có tỷ lệ thắng được xác định rõ ràng nhưng không ngẫu nhiên, ngay cả khi tỷ lệ phần thưởng tuyệt đối thấp? Các nghiên cứu hiện nay giải quyết những câu hỏi này.
Thiết kế thí nghiệm đã được thông báo bởi một nghiên cứu tinh tế về kỳ vọng thưởng và phản ứng nơ-ron DA ở khỉ (Fiorillo và cộng sự, 2003). Các động vật trong nghiên cứu đó đã nhận được phần thưởng nước trái cây (Hoa Kỳ) theo lịch biểu tỷ lệ biến số 0, 25, 50, 75 hoặc 100%. Lịch trình được chỉ định bởi 1 của 4 CS (biểu tượng) khác nhau. Lịch biểu 0% được phân phối thường xuyên như lịch biểu 100%, nhưng đã bỏ qua CS. Tốc độ bắn của các tế bào thần kinh DA trong khoảng thời gian giữa lúc khởi phát CS và giao hàng hoặc thiếu sót của Hoa Kỳ là biện pháp phụ thuộc chính. Nghiên cứu cho thấy phản ứng DA tăng lên như một chức năng của sự không chắc chắn của việc trao thưởng. Do đó, theo lịch trình 100%, CS gợi lên ít hoạt động, theo lịch trình 25 và 75%, CS gợi lên mức độ hoạt động vừa phải và tương tự, và theo lịch trình 50%, CS gợi lên hoạt động tối đa. Trong mỗi trường hợp, tốc độ bắn leo thang trong suốt khoảng thời gian CS-US, tức là, khi kỳ vọng tiếp cận kết quả.
Những phát hiện này chỉ ra rằng hoạt động DA không chỉ thay đổi theo việc phần thưởng có chắc chắn hay không (Tỷ lệ cố định) hoặc không chắc chắn (Tỷ lệ biến), mà còn thay đổi tỷ lệ nghịch với lượng thông tin về việc trao phần thưởng được truyền tải bởi CS. Trong điều kiện 100%, CS gợi lên sự mong đợi phần thưởng và cũng dự đoán hoàn hảo việc giao hàng của nó. Trong điều kiện 25 và 75%, CS gợi lên sự mong đợi và dự đoán việc trao phần thưởng ba trong bốn lần. Trong điều kiện 50%, CS gợi lên sự mong đợi nhưng không cung cấp thông tin nào về việc trao phần thưởng ngoài cơ hội. Dựa trên những phát hiện của họ, Fiorillo et al. kết luận: Sự gia tăng không chắc chắn này gây ra bởi dopamine có thể đóng góp vào các đặc tính bổ ích của cờ bạc, (trang 1901).
Tác động của 50% phần thưởng thay đổi trong một phiên duy nhất sẽ không thay đổi trong quá trình nhiều phiên vì khả năng phần thưởng vẫn hoàn toàn không thể đoán trước trong mỗi lần thử. Do đó, khi xem xét các điều kiện có thể tối đa hóa sự kích hoạt mãn tính của tế bào thần kinh DA qua các đợt đánh bạc lặp đi lặp lại, lịch trình 50% sẽ tạo ra hiệu quả lâu dài nhất cũng như mạnh mẽ nhất. Điều này đáng chú ý là tỷ lệ phần thưởng trong thời gian dài (tiền thưởng> 0) được quan sát qua hàng nghìn lần quay trên máy đánh bạc thương mại là 45.8% (Tremblay và cộng sự, 2011). Do đó, phần thưởng biến số 50% dường như phản ánh chính xác lịch thanh toán được quản lý bởi các thiết bị đánh bạc thực tế.
Nghiên cứu này sử dụng cùng một lịch trình điều hòa như Fiorillo et al. trong tình trạng phơi nhiễm mãn tính, thiết kế giữa các nhóm với chuột. Động vật trải qua ~ 3 tuần điều kiện hàng ngày, trong đó CS (ánh sáng) được ghép nối với US (một lượng nhỏ sucrose). Sau giai đoạn huấn luyện, động vật nghỉ ngơi trước khi đánh giá mức độ nhạy cảm được lập chỉ mục bằng phản ứng vận động cơ địa với amphetamine. Dựa trên các tài liệu, người ta dự đoán rằng những con chuột tiếp xúc với các lịch trình thưởng khác nhau sẽ không khác nhau về hành vi vận động không dùng thuốc của chúng nhưng sẽ biểu hiện các mức độ vận động khác nhau đáng kể sau khi dùng amphetamine, với nhóm 50% có phản ứng vận động cơ địa cao hơn với thuốc so các nhóm khác trong quá trình sử dụng liều, một mô hình sẽ được mong đợi nếu 50% động vật trước đó đã tiếp xúc với các liều bổ sung của bản thân amphetamine (tức là nhạy cảm chéo).
Thử nghiệm 1
Vật liệu và phương pháp
Đối tượng
Bốn nhóm (n = 8 / nhóm) chuột trưởng thành (300, 350 g) chuột đực Sprague-Dawley (Charles River, St. Constant, Quebec, Canada) được đặt riêng trong các hộp polycarbonate rõ ràng (20 × 43 × 22 cm) dưới một 12 ngược: Chu kỳ tối-tối 12. Họ đã nhận được quảng cáo tự do truy cập vào thực phẩm và nước, và xử lý hàng ngày bởi một người thí nghiệm trong 2 tuần trước khi nghiên cứu. Mỗi nhóm được điều hòa theo một trong bốn lịch thưởng thay đổi: 0, 25, 50 hoặc 100%. Nhóm 75% đã bị bỏ qua trong nghiên cứu ban đầu này, như Fiorillo và cộng sự. (2003) đã tìm thấy bản phát hành DA sau CS tương đương theo lịch thưởng 25 và 75%, do đó cả hai điều kiện dẫn đến việc phát hành DA lớn hơn so với điều kiện 100% CS-US, nhưng ít hơn điều kiện 50%.
Thiết bị
Truy cập vào các bài thuyết trình sucrose và CS được cung cấp riêng trong các hộp điều hòa hoạt động (33 × 31 × 29 cm). Mỗi hộp được trang bị một tạp chí gia cố, nằm trên bức tường phía trước. Một ánh sáng trong đầu tạp chí phục vụ như CS. Một cái gáo chất lỏng điều khiển bằng điện từ có thể được nâng lên sàn của tạp chí. Các sự kiện trong hộp được điều khiển bởi thiết bị và phần mềm của Med Associates, sử dụng chương trình nội bộ được viết bằng MED-PC. Thử nghiệm vận động được tiến hành riêng lẻ trong các lồng Plexiglas (27 × 48 × 20 cm). Mỗi lồng được trang bị một hệ thống giám sát bao gồm sáu tế bào chùm ảnh để phát hiện chuyển động ngang.
Thủ tục
Hội thảo. Nghiên cứu được thực hiện tuân thủ các nguyên tắc đạo đức do Hội đồng Chăm sóc Động vật Canada quy định. Chuột bị giới hạn thức ăn ở mức 90% trọng lượng cơ thể trong suốt thời gian nghiên cứu và được nuôi riêng lẻ. Mỗi con chuột được huấn luyện phần thưởng sucrose ngày (giải pháp nước 15% ở mức 10 ml mỗi phần thưởng): 0.06 ngày liên tiếp × 5 tuần, với ngày nghỉ cuối tuần. Động vật được duy trì trên chow tiêu chuẩn trước và sau giai đoạn huấn luyện; tiếp xúc với sucrose bị giới hạn trong mười lăm buổi tập luyện 3-phút. Mỗi phiên hàng ngày bao gồm các bài thuyết trình kích thích 40 (một ánh sáng; CS), mỗi phần được phân tách bằng một khoảng thời gian thử nghiệm của 15 s. Ánh sáng được đặt ở bảng trên cùng của tạp chí, và vẫn duy trì cho các 120, với sucrose có sẵn trong các 25 cuối cùng. Trong trường hợp của nhóm 5, cái gáo sucrose được nâng lên mỗi lần 0 (đối với 140) nhưng ánh sáng kích thích không được chiếu sáng. Điều này đánh đồng khoảng thời gian giữa các bài thuyết trình của cái môi trong nhóm 5 và các nhóm khác (0 + 120 s). Mỗi đợt điều trị kéo dài ~ 25 phút. Trung bình, nhóm 40 đã nhận được sucrose một lần cho mỗi bốn bài thuyết trình CS; nhóm 25 đã nhận được sucrose một lần cho mỗi hai bài thuyết trình CS và nhóm 50 nhận được sucrose sau mỗi bài thuyết trình CS.
Kiểm tra. Hai tuần sau phiên truy cập sucrose cuối cùng (hoặc điều hòa trực tiếp), phản ứng vận động với d-amphetamine (AMPH; ip) đã được đánh giá. Chuột được cho ba phiên 2-h để làm quen với các hộp vận động, sau đó là sáu phiên kiểm tra AMPH. Ngày thử nghiệm AMPH xảy ra tại các khoảng thời gian 1-wk. Vào những ngày thử nghiệm, chuột được cho dùng 30 tối thiểu để làm quen với các hộp sau đó nhận được một liều AMPH 0.5 / kg duy nhất theo sau, vào các phiên hàng tuần riêng biệt, bằng năm liều 1.0 mg / kg (một liều mỗi ngày) vào những ngày thử nghiệm . Đầu máy sau AMPH được đánh giá cho 1 tối thiểu trên mỗi phiên.
Phương pháp phân tích dữ liệu
Phân tích thống kê được thực hiện với SPSS (v. 16 và v. 21; SPSS Inc., Chicago IL). Phản ứng hành vi ngay lập tức đối với CS được đánh giá dưới dạng chọc mũi vào khẩu độ nơi sucrose được phân phối. Số lần chọc mũi trung bình trong khoảng thời gian này (5 s trên mỗi thử nghiệm) sau đó được so sánh với số lần chọc mũi trung bình trong cùng thời gian (5 s) tính trung bình theo thời gian khi CS vắng mặt. Nhóm × Phiên ANOVAs chọc mũi với CS có mặt và vắng mặt đã theo dõi việc mua lại phản ứng phân biệt đối xử với các câu trả lời chọc mũi và bừa bãi theo các lịch trình khác nhau trong các khóa đào tạo sucrose của 15.
Hiệu quả của điều trị đối với đáp ứng vận động được đánh giá với ANOVAs Nhóm × Phiên đối với giai đoạn sử dụng thuốc không có thuốc (ba phiên), thử thách 0.5 mg / kg nhạy cảm trước (một phiên) và trong giai đoạn 1 mg / kg AMPH năm phiên Phác đồ mẫn cảm, khi các nhóm được dự kiến sẽ khác nhau khi đáp ứng với liều AMPH lặp đi lặp lại. Nhóm × Phiên ANOVAs cũng đánh giá các phản ứng vận động không có thuốc trong giai đoạn tập trung trước khi tiêm 30-min từ mỗi phiên kiểm tra AMPH. So sánh có kế hoạch đã đánh giá sự khác biệt về hiệu suất trung bình của nhóm 50 so với nhóm 0 (không kiểm soát kỳ vọng) và nhóm 100 (không kiểm soát độ không đảm bảo), bằng phương tiện t-test (Howell, 1992), sử dụng lỗi MS và các điều khoản lỗi df cho hiệu ứng có liên quan (nghĩa là tương tác nhóm hoặc nhóm × phiên) từ ANOVA (Người chiến thắng, 1971). Phân tích xu hướng đa thức đã kiểm tra hồ sơ của các thay đổi trong quá trình phiên.
Để xác định xem các phản ứng tiếp cận trong sự hiện diện và vắng mặt của CS trong các buổi đào tạo sucrose của 15 đã góp phần thay đổi phản ứng vận động với AMPH, hoặc sự khác biệt của nhóm qua trung gian trong phản ứng AMPH, phân tích theo dõi hiệp phương sai (ANCOVAs) đã được thực hiện trên AMPH dữ liệu vận động, bao gồm tổng số mũi chọc (tổng cho các phiên 15) khi CS vắng mặt dưới dạng hiệp phương sai. Một tác động đáng kể của hiệp phương sai sẽ chỉ ra rằng các phản ứng tiếp cận không có thuốc được kiểm duyệt (ảnh hưởng đến sức mạnh) của các tác động của nhóm hoặc phiên. Sự suy giảm về tầm quan trọng của các tác động của nhóm hoặc phiên với sự có mặt của hiệp phương sai có ý nghĩa cho thấy các phản ứng tiếp cận qua trung gian (chiếm) các tác động của nhóm hoặc phiên. Sự suy giảm về tầm quan trọng của hiệu ứng nhóm hoặc phiên trong trường hợp không có hiệu ứng hiệp biến đáng kể chỉ đơn giản phản ánh sự mất năng lực thống kê do việc phân bổ lại df từ thuật ngữ lỗi sang hiệp phương sai và sẽ không ảnh hưởng đến việc giải thích ảnh hưởng của nhóm hoặc phiên.
Kết quả
Mũi chọc trong phiên điều hòa sucrose
CS hiện tại. Nhân vật 1A cho thấy các mũi chọc trung bình cho các nhóm 25, 50 và 100 trong khi CS có mặt trên các phiên điều hòa sucrose của 15 (các mũi chọc không được mã hóa cho nhóm 0, không nhận được CS). Một nhóm 3 × 15 Phiên ANOVA mang lại hiệu quả chính đáng kể của Nhóm, F(2, 21) = 5.63, p = 0.011 và Phiên, F(14, 294) = 14.00, p <0.001, cùng với tương tác Nhóm × Phiên đáng kể, F(28, 294) = 2.93, p <0.001. Nhân vật 1A chỉ ra rằng tác động chính của Phiên phản ánh sự gia tăng chọc mũi qua các phiên trong cả ba nhóm và hiệu ứng chính của Nhóm phản ánh điểm tổng thể cao hơn trong nhóm 100 so với nhóm 25 với điểm trung gian trong nhóm 50. Một nhóm quan trọng × Tương tác phiên cho xu hướng hình khối, F(2, 21) = 4.42, p = 0.030, biểu thị sự gia tăng nhanh chóng, nhúng và chững lại trong việc chọc mũi qua các phiên trong nhóm 100, so với mức tăng tuyến tính qua các phiên trong nhóm 50 và tăng tuyến tính hơn so với các phiên trong nhóm 25.
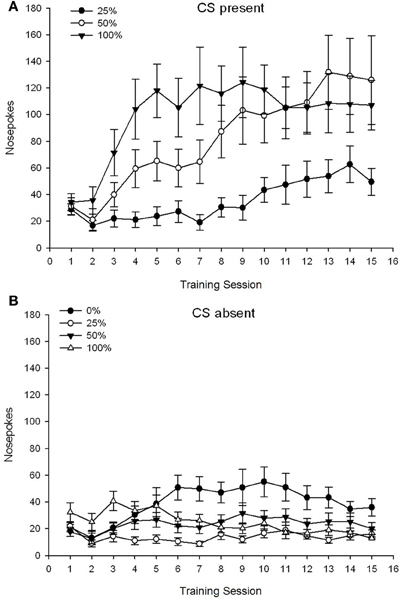
Hình 1. Các phản ứng tiếp cận trung bình (SE) (chọc mũi) trong các buổi huấn luyện sucrose sucrose trong các nhóm chuột Sprague Dawley (n = 8 / nhóm) tiếp xúc với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch biểu biến đổi 0, 25, 50 hoặc 100%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. (A) Điểm khi CS có mặt (thử nghiệm 5 s × 15). (B) Điểm khi CS vắng mặt (trung bình cho 5 × 15 s trong khi đèn tắt).
CS vắng mặt. Nhân vật 1B cho thấy các mũi chọc trung bình cho cả bốn nhóm trong một thời gian tương đương (các thử nghiệm 5 s × 15) tính trung bình theo thời gian khi CS vắng mặt. Một nhóm 4 × 15 Phiên ANOVA mang lại hiệu quả chính đáng kể của Nhóm, F(3, 28) = 7.06, p = 0.001 và Phiên F(14, 392) = 2.84, p <0.001, cùng với tương tác Nhóm × Phiên đáng kể, F(42, 392) = 3.93, p <0.001. Tương tác Nhóm × Phiên đáng kể cho xu hướng bậc hai, F(3, 28) = 3.91, p = 0.019, cùng với không có tương tác cho xu hướng hình khối, F(3, 28) <0.93, p > 0.44, phản ánh cấu hình “chữ U ngược” của mũi chọc vào các phiên trong nhóm 0, so với cấu hình nhìn chung ổn định so với các phiên trong các nhóm khác.
Thói quen với buồng đầu máy
Một nhóm 4 × 3 Phiên ANOVA mang lại hiệu ứng chính của Phiên, F(2, 56) = 5.67, p = 0.006 và không có hiệu ứng đáng kể nào khác, F(3, 28) <1.60, p > 0.21. Số lần ngắt tia trung bình (SE) mỗi 2 giờ trong các hộp định vị là 1681 (123) trong phiên 1, 1525 (140) trong phiên 2 và 1269 (96) trong phiên 3. Các so sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100 vào buổi sinh hoạt đầu tiên hoặc cuối cùng, t(84) <1.69, p > 0.05. Do đó, trong trường hợp không có AMPH, việc tiếp xúc lặp lại với các hộp thử nghiệm có liên quan đến sự suy giảm nhất quán trong hoạt động vận động vị trí tự phát trong bốn nhóm (tức là hiệu ứng Phiên) và không có phản ứng khác biệt như một chức năng của lịch đào tạo sucrose (không có tương tác) .
Buổi kiểm tra
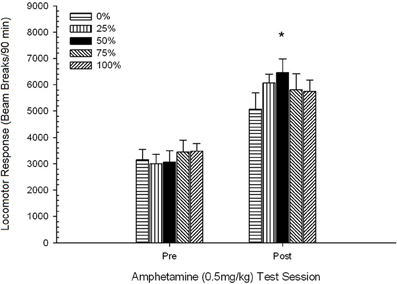
Ảnh hưởng của thử thách nhạy cảm 0.5 mg / kg AMPH.
Đầu máy trước tiêm. Một ANOVA nhóm một chiều ANOVA của phản ứng vận động trong giai đoạn tập trung trước khi tiêm 4-min không mang lại hiệu quả đáng kể, F(3, 28) <1.05, p > 0.38. So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100, t(32) <0.87, p > 0.40. Do đó, sự khác biệt cơ bản về vận động trước khi tiêm không giải thích cho sự khác biệt nhóm về phản ứng của vận động cơ với AMPH. Điểm gãy chùm trung bình (SE) đối với mẫu là 559 (77).
Đầu máy sau tiêm so với phiên làm quen không có thuốc cuối cùng. ANOVA 4 Nhóm × 2 Phiên so sánh các phản ứng vận động cơ địa của các nhóm trong phiên tạo thói quen cuối cùng và ngay sau thử thách gây mẫn cảm trước 0.5 mg / kg AMPH. Điểm cho phiên làm quen (120 phút) được chia tỷ lệ để tương ứng với thời lượng của phiên kiểm tra AMPH (90 phút) (điểm thói quen thô × 90/120). Phân tích mang lại hiệu quả chính đáng kể của Phiên, F(1, 28) = 34.16, p <0.001, và không có ảnh hưởng đáng kể nào khác, F(3, 28) <2.26, p > 0.10. Hiệu ứng Session phản ánh sự gia tăng các điểm gãy chùm trung bình (SE) theo phản ứng với liều, từ 952 (72) lên 1859 (151). So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100 về phản ứng với liều lượng, t(56) <1.72, p > 0.10. Tuy nhiên, thứ tự xếp hạng của điểm phá vỡ chùm (M; SE) phù hợp với giả thuyết: nhóm 50 (2205; 264)> nhóm 0 (2025; 203)> nhóm 100 (1909; 407)> nhóm 25 (1296; 299) .
Tác dụng của 1 mg / kg AMPH.
Đầu máy trước tiêm. Một nhóm 4 × 5 Phiên ANOVA của phản ứng vận động trong giai đoạn sử dụng trước khi tiêm 30-min trên các phiên kiểm tra XNPHX mg / kg AMPH mang lại hiệu quả chính của Phiên, F(4, 112) = 43.64, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(3, 28) <0.97, p > 0.42. So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100 trong phiên kiểm tra đầu tiên hoặc cuối cùng, t(140) <0.84, p > 0.30. Do đó, sự khác biệt cơ bản về vận động không giải thích cho sự khác biệt của nhóm về phản ứng của vận động cơ với AMPH. Điểm phá vỡ chùm tia trung bình (SE) cho giai đoạn làm quen trước khi dùng liều trong các phiên 1–5 là: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
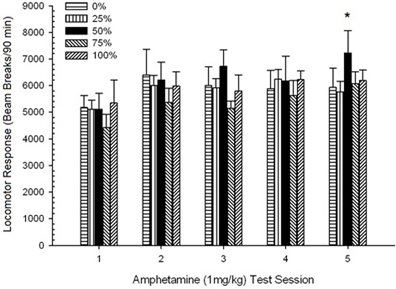
Đầu máy sau tiêm. Nhân vật 2 cho thấy tác động của năm lần tiêm 1 mg / kg AMPH (một lần mỗi tuần) đối với điểm hoạt động vận động ở bốn nhóm. Một nhóm 4 × 5 Phiên ANOVA mang lại hiệu ứng chính của Phiên, F(4, 112) = 8.21, p <0.001, một tác động chính biên của Nhóm, F(2, 45) = 3.28, p = 0.085 và không có tương tác quan trọng, F(12, 122) <0.77, p > KHAI THÁC.
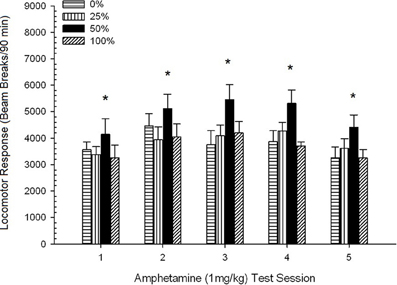
Hình 2. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) đến 1 mg / kg d-amphetamine (ip) trên các phiên hàng tuần của 5 trong các nhóm chuột Sprague Dawley (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50 hoặc 100%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. *p <0.05 cho sự khác biệt trung bình giữa nhóm 50 và nhóm 0 cũng như nhóm 100, dựa trên các so sánh theo kế hoạch.
So sánh có kế hoạch cho thấy điểm số 50 của nhóm khác biệt đáng kể so với nhóm 0, t(14) = 2.19, p = 0.037 và nhóm 100, t(14) = 2.36, p = 0.025 [và khác biệt nhỏ so với nhóm 25, t(14) = 2.03, p = 0.051]. Do đó, trong nhóm 50, phản ứng vận động với 1 mg / kg AMPH đáng tin cậy vượt quá ba nhóm còn lại trong tất cả năm phiên kiểm tra. Phân tích xu hướng đa thức phát hiện xu hướng bậc hai đáng kể qua các phiên, F(1, 28) = 32.47, p <0.0001 và không có xu hướng quan trọng nào khác, F(1, 28) <1.78, p > 0.19. Nhân vật 2 cho thấy kết quả này phản ánh một mẫu hình chữ U ngược trong các phiên.
Kiểm soát sự thay đổi trong phản ứng chọc mũi trong quá trình đào tạo sucrose
ANCOVA tiếp theo của các phản ứng vận động với 1 mg / kg AMPH, với các mũi chọc (hiện tại CS) là hiệp phương sai, trong ba nhóm đã nhận CS, mang lại hiệu ứng chính cận biên của Nhóm, F(2, 20) = 3.07, p = 0.069 và không có tác động liên quan đến đồng biến quan trọng, F(4, 80) <0.05, p > 0.85. Do đó, phản ứng theo phương pháp tiếp cận được sắp xếp trong quá trình đào tạo không giải thích được sự thay đổi đáng kể trong phản ứng vị trí vận động đối với 1 mg / kg AMPH ở nhóm 25, 50 hoặc 100.
ANCOVA tiếp theo của các phản ứng vận động với 1 mg / kg AMPH, với các mũi chọc (CS vắng mặt) như một hiệp phương sai, mang lại hiệu quả rõ rệt của hiệp phương sai, F(1, 27) = 6.17, p = 0.020, một hiệu ứng chính đáng kể của Nhóm, F(3, 27) = 4.13, p = 0.016, một phiên cận biên × Tương tác đồng biến, p = 0.080 và không có hiệu ứng đáng kể nào khác, F(4, 108) <1.48, p > 0.21. Do đó, phương pháp tiếp cận không được sắp xếp (bừa bãi) đáp ứng trong quá trình đào tạo đã giải thích sự thay đổi đáng kể trong phản ứng vị trí vận động đối với 1 mg / kg AMPH. Tuy nhiên, sự thay đổi này không trùng lặp với phương sai liên quan đến nhóm, bởi vì việc bao gồm hiệp biến trong phân tích làm tăng hơn là giảm ý nghĩa của hiệu ứng nhóm.
Thảo luận
Dữ liệu chọc mũi trong khi CS có mặt cho thấy các nhóm có được mối liên hệ giữa CS và sucrose phân phối như được phản ánh bởi sự gia tăng các phản ứng cued trong các buổi đào tạo. Hồ sơ trả lời qua các phiên trong khi CS có mặt cho thấy lịch trình 100 và 50% CS-US có hiệu quả tương đương trong việc khơi gợi cách tiếp cận, trong khi lịch trình 25% gợi ra sự gia tăng khiêm tốn hơn trong cách tiếp cận do cue gây ra. Mũi chọc dữ liệu trong khi CS vắng mặt cho thấy các nhóm nhận được bất kỳ lịch trình đào tạo CS-sucrose nào (nhóm 25, 50, 100) đã nhanh chóng học cách giảm mũi của họ khi không có CS, trong khi các động vật trong nhóm 0 , không nhận được CS, chỉ học cách giảm hành vi tiếp cận của họ xuống một mức độ hạn chế sau khi được đào tạo mở rộng.
Dữ liệu về thói quen cho thấy các nhóm không khác nhau trước AMPH và việc tiếp xúc nhiều lần với các hộp thử nghiệm có liên quan đến việc giảm đáp ứng vận động không có thuốc. Do đó, sự khác biệt giữa các nhóm và tăng đáp ứng với liều AMPH lặp đi lặp lại có thể được quy cho sự khác biệt tồn tại từ trước trong hành vi vận động.
Kết quả của thử thách tiền mẫn cảm với 0.5 mg / kg AMPH đã xác nhận rằng thuốc làm tăng hoạt động vận động so với ngày cuối cùng không có thuốc. Theo giả thuyết, nhóm 50 được xếp hạng cao hơn nhóm 0 hoặc 100 (cũng như nhóm 25) về mức độ đáp ứng trung bình với liều, mặc dù sự khác biệt trung bình giữa các nhóm là không đáng kể.
Đối với các phiên điều trị nhạy cảm, so sánh theo kế hoạch giữa các nhóm cho thấy rằng việc tiếp xúc trước với phần thưởng 50% sucrose có điều kiện dẫn đến sự gia tăng đáng kể trong phản ứng vận động đối với liều amphetamine 1.0 mg / kg so với ba lịch trình còn lại. Hiệu quả này rõ ràng ngay từ liều đầu tiên và không thay đổi đáng kể qua các liều lặp lại. Phân tích xu hướng chỉ ra phản ứng hai pha (đối với mẫu đầy đủ) với các liều lặp lại của AMPH, tăng lên đến liều thứ ba và giảm sau đó. Kết quả của ANCOVA tiếp theo có chọc ngoáy mũi (không có CS) làm hiệp biến xác nhận rằng sự khác biệt trong phản ứng vị trí vận động của bốn nhóm đối với AMPH 1 mg / kg không được trung gian bởi phương pháp tiếp cận không được kiểm soát phản ứng trong các buổi huấn luyện đường sucrose.
Hiệu ứng nhóm trong các phiên nhạy cảm phù hợp với giả thuyết của chúng tôi. Hiệu ứng phiên bi-phasic không phù hợp với sự leo thang tiếp tục dự kiến trong các phản ứng vận động với liều AMPH lặp đi lặp lại. Điều này có thể liên quan đến khoảng thời gian dùng thuốc. Để giải quyết vấn đề này, nên áp dụng một quy trình (liều thay thế hàng ngày) để tạo ra sự leo thang nhất quán trong đáp ứng vận động với liều XNPHX / kg liều AMPH (nghĩa là nhạy cảm hành vi). Tác động của chế độ nhạy cảm với AMPH đối với phản ứng tiếp theo đối với thử thách 1.0 mg / kg thứ hai sẽ hỗ trợ thêm cho tính tổng quát của hiệu ứng này. Việc đưa vào thử thách nước muối trước AMPH sẽ xác định vai trò của các tác động mong đợi hoặc liên quan đến tiêm (ví dụ: căng thẳng) đối với phản ứng vận động đối với AMPH. Việc đưa vào một nhóm sucrose có điều kiện 0.5% sẽ giúp làm rõ vai trò của sự không chắc chắn về phần thưởng so với sự không thường xuyên về phần thưởng đối với mô hình trả lời cho các nhóm 75 và nhóm 50. Ngoài ra, để cho phép đánh giá (bởi ANCOVA) về sự đóng góp của các phản ứng tiếp cận không có ma túy đối với sự vận động theo AMPH (sử dụng mũi chọc với CS là đồng biến), các mũi chọc cũng được mã hóa cho nhóm 25 trong khoảng thời gian khi CS đã có mặt trong bốn nhóm khác (ví dụ, do đó, mũi chọc từ cả năm nhóm, bao gồm cả nhóm 0 không nhận CS CS có thể được đưa vào phân tích hiệp phương sai với CS có mặt như hiệp phương sai). Những tinh chỉnh này được kết hợp trong thử nghiệm 0.
Thử nghiệm 2
Vật liệu và phương pháp
Phương pháp của thí nghiệm 2 tương tự như thí nghiệm 1 nhưng được sửa đổi để gần đúng hơn một chế độ được tìm thấy để tạo ra sự nhạy cảm AMPH một cách đáng tin cậy (Fletcher và cộng sự, 2005). Các thay đổi như sau: (a) Nhóm 75% CS-sucrose (n = 8) đã được bao gồm; (b) Trong quá trình huấn luyện sucrose, chuột (trừ nhóm 0) đã nhận được các bài thuyết trình 20 CS (ánh sáng) (trái ngược với 15 trong thí nghiệm 1); (c) Các bài thuyết trình CS được phân tách bằng khoảng thời gian thử nghiệm trung bình của 90 s; phạm vi: 30 triệt 180 s (so với 120 trong thử nghiệm 1), bù cho sự gia tăng trong các thử nghiệm đào tạo để đánh đồng thời lượng của mỗi buổi đào tạo với thử nghiệm 1; (d) thời lượng của mỗi trong ba phiên tập sự đã giảm từ 120 xuống 90 tối thiểu để tương ứng với thời lượng của các phiên kiểm tra; (e) Một thử thách nước muối (ip, 1 ml / kg) (90 phút) đã được thêm vào (ngày đào tạo sau sucrose 8), để đánh giá hiệu quả vận động của tiêm cho mỗi gia nhập (ví dụ, kỳ vọng, căng thẳng); (f) Các phiên nhạy cảm 1 mg / kg được tổ chức vào các ngày trong tuần thay thế (ngày sau đào tạo 12 XN 21) thay vì vào các khoảng thời gian hàng tuần như trong thử nghiệm 1; (g) Cùng với thử thách 0.5 mg / kg nhạy cảm trước (ngày đào tạo 9), thử thách 0.5 mg / kg sau nhạy cảm thứ hai đã được thêm vào (ngày đào tạo sau sucrose), để kiểm tra tính tổng quát của tác dụng mẫn cảm trên các liều; (h) chọc mũi trong khi CS có mặt được mã hóa cho tất cả các nhóm (bao gồm cả nhóm 28); (i) chọc mũi trong khi CS vắng mặt đã được ghi lại cụ thể từ khoảng 0-s ngay trước khi bắt đầu CS để chỉ số phản ứng tiếp cận sớm.
Kết quả
Mũi chọc trong phiên điều hòa sucrose
Một nhóm 5 × Phiên 15 × Giai đoạn 2 (hiện tại không có CS) ANOVA của mũi chọc mang lại hiệu quả chính đáng kể của Nhóm, F(4, 19) = 2.89, p = 0.050, Phiên F(14, 266) = 2.28, p = 0.006 và Pha, F(1, 19) = 14.72, p = 0.001, cũng như tương tác ba chiều đáng kể, F(56, 266) = 1.38, p = 0.050. Tấm (A, B) của hình 3 vẽ biểu đồ điểm số chọc mũi trung bình của các nhóm cho các pha CS có mặt và CS vắng mặt, tương ứng. So sánh giữa hai bảng cho thấy hiệu ứng chính của Pha phản ánh nhiều phản ứng chọc mũi tổng thể hơn khi CS có mặt so với khi vắng mặt. Do đó, các câu trả lời đã được kiểm soát xảy ra thường xuyên hơn đáng kể so với các câu trả lời không được kiểm soát sớm. Các tác động chính của Nhóm và Phiên không được giải thích dễ dàng do tương tác thứ tự cao hơn. Kết quả thứ hai này phản ánh sự hội tụ của điểm số cho năm nhóm ở mức thấp tương đối ổn định qua các phiên khi CS vắng mặt (Hình 3B), cùng với sự phân kỳ điểm số thành các mức cao (nhóm 75, nhóm 100), trung cấp (nhóm 50) và mức độ thấp (nhóm 0, nhóm 25) phản ứng qua các phiên khi CS có mặt (Hình 3A). Trong số các xu hướng đa thức bậc thấp (tuyến tính, bậc hai, khối), chỉ có tương tác ba chiều cho xu hướng tuyến tính có ý nghĩa tiếp cận, F(4, 19) = 2.32, p = 0.094, phản ánh sự gia tăng nói chung đơn điệu trong việc chọc mũi qua các phiên trong nhóm 75 và ổn định tương đối nhanh hơn ở mức độ cao, trung bình và mức độ đáp ứng thấp trong các nhóm khác khi CS có mặt.
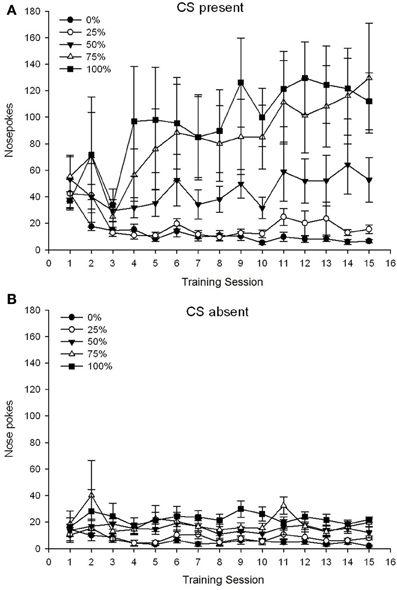
Hình 3. Các phản ứng tiếp cận trung bình (SE) (chọc mũi) trong các buổi huấn luyện sucrose sucrose trong các nhóm chuột Sprague Dawley (n = 8 / nhóm) tiếp xúc với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch biểu biến đổi 0, 25, 50, 75 hoặc 100%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. (A) Điểm khi CS có mặt (thử nghiệm 5 s × 20). (B) Điểm khi CS vắng mặt (trung bình cho 5 × 20 s trong khi đèn tắt).
Thói quen với hộp đầu máy
Một nhóm 5 × 3 Phiên ANOVA của các phản ứng vận động không có thuốc mang lại hiệu quả chính đáng kể của Phiên, F(2, 70) = 60.01, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <0.70, p > 0.60. So sánh có kế hoạch của nhóm 50 với nhóm 0 và với nhóm 100 trong các buổi sinh hoạt đầu tiên và cuối cùng không mang lại hiệu quả đáng kể, tcủa <0.84, p > 0.40. Do đó, phản ứng định vị không dùng thuốc trung bình ở các nhóm chính không khác nhau trước khi thử nghiệm. Số lần ngắt xà trung bình (SE) trong 90 phút là 2162 (118) trong phiên 1, 1470 (116) trong phiên 2 và 1250 (98) trong phiên 3.
Buổi kiểm tra
Nước muối. Một nhóm 5 × 2 Phiên ANOVA đã so sánh phản ứng vận động trong phiên tập sự cuối cùng và phiên thử thách nhiễm mặn. ANOVA mang lại hiệu ứng chính của Phiên, F(1, 35) = 62.46, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <0.65, p > 0.64. Nhân vật 4 vẽ sơ đồ nhóm có nghĩa và cho thấy hiệu ứng Phiên phản ánh sự giảm tổng thể phản ứng vận động từ phiên làm quen không có thuốc cuối cùng sang phiên nước muối, không thay đổi theo nhóm. Do đó, sự suy giảm trong phản ứng vận động được thấy trong ba phiên tập sự tiếp tục ở lần tiếp xúc thứ tư không có thuốc với các hộp thử nghiệm.
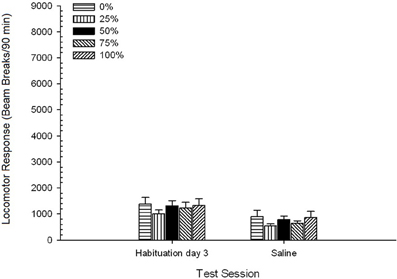
Hình 4. Phản ứng vận động trung bình (SE) trung bình (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) vào lần cuối của các phiên sử dụng thuốc không có ma túy và trong phiên tiếp theo sau khi tiêm nước muối (ip, 3 ml / kg) trong các nhóm Sprague Dawley chuộtn = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện.
Tác dụng của 0.5 mg / kg AMPH.
Đầu máy trước tiêm. Một nhóm 5 × 2 Phiên ANOVA của đầu máy tiêm trước (30-min) trong các ngày thử nghiệm 0.5 mg / kg nhạy cảm trước mang lại hiệu quả chính đáng kể của Phiên, F(1, 35) = 13.39, p = 0.001 và không có hiệu ứng đáng kể nào khác, F(4, 35) <1.79, p > 0.15. So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100 trong phiên đầu tiên, t(70) <1.00, p > 0.30. Tuy nhiên, trong phiên thứ hai (sau nhạy cảm), nhóm 50 (1203; 121) hiển thị nhiều hơn đáng kể các đứt đoạn trước khi tiêm (M; SE) so với nhóm 100 (756; 103), t(70) = 5.11, p <0.001, nhưng không khác với nhóm 0 (1126; 211), t(7) <0.88, p > 0.40. Do đó, sự khác biệt cơ bản về vận động không tính đến sự khác biệt giữa các nhóm về phản ứng vận động với liều AMPH 0.5 mg / kg đầu tiên nhưng có thể góp phần vào sự khác biệt giữa nhóm 50 và nhóm 100 về phản ứng vận động với liều 0.5 mg / kg thứ hai của AMPH. . Điểm phá vỡ chùm tia trung bình (SE) đối với giai đoạn trước khi tiêm vào phiên thử nghiệm AMPH 0.5 mg / kg đầu tiên và lần thứ hai là 757 (41) và 974 (59).
Đầu máy sau tiêm. Một nhóm 5 × 2 Phiên ANOVA của phản ứng vận động với 0.5 mg / kg AMPH trước và sau chế độ nhạy cảm với liều 5 mang lại hiệu quả chính của Phiên, F(1, 35) = 76.05, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <1.10, p > 0.37. Nhân vật 5 hiển thị điểm trung bình cho mỗi nhóm và phiên.
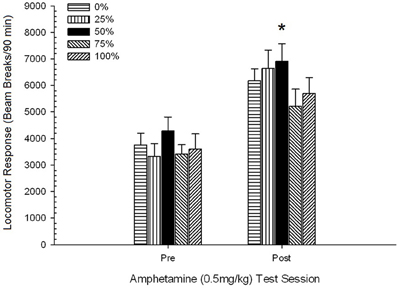
Hình 5. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) thành 0.5 mg / kg d-amphetamine trong các phiên riêng biệt trước và sau chế độ nhạy cảm với phiên 5 của d-amphetamine (1.0 mg / kg; mỗi phiên) trong các nhóm chuột Sprague Dawley (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. *p <0.05 cho sự khác biệt trung bình giữa nhóm 50 và nhóm 0 cũng như nhóm 100, dựa trên các so sánh theo kế hoạch.
Hình vẽ cho thấy hiệu ứng Phiên liên quan đến sự gia tăng đáng kể trong các chùm tia trung bình (SE) trung bình trên mỗi 90 tối thiểu từ 0.5 mg / kg liều 1, 3674 (216) đến 0.5 mg / kg liều 2, 6123 (275). Việc thiếu tương tác hoặc hiệu ứng nhóm cho thấy độ nhạy cảm với AMPH không thay đổi đáng tin cậy giữa các nhóm. Mặc dù thiếu các tác động đáng kể liên quan đến nhóm trong ANOVA, kiểm tra số liệu cho thấy nhóm 50 cho thấy phản ứng lớn nhất đối với cả liều 0.5 mg / kg thứ nhất và thứ hai. So sánh có kế hoạch về đáp ứng với liều 0.5 mg / kg đầu tiên cho thấy không có sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100, t'S(35) <0.48, p > 0.50. Tuy nhiên, phản ứng với liều thứ hai (sau mẫn cảm) 0.5 mg / kg, nhóm 50 thể hiện sự vận động mạnh hơn đáng kể so với nhóm 0, t(35) = 2.00, p <0.05, cũng như nhóm 100, t(35) = 3.29, p <0.01.
Trước sự khác biệt đáng kể của nhóm về sự vận động trước tiêm trong phiên XNPHX mg / kg thứ hai được báo cáo ở trên, một phiên 0.5 tiếp theo × 5 Phiên ANCOVA của phản ứng vận động với 2 mg / kg AMPH đã được tiến hành, kiểm soát trước tiêm đầu máy vào phiên thứ hai. Phân tích này mang lại một hiệu ứng đáng kể của hiệp phương sai, F(1, 34) = 8.65, p = 0.006, tác dụng chính của Phiên F(1, 34) = 10.83, p = 0.002 và không có hiệu ứng đáng kể nào khác, F(4, 34) <0.85, p > 0.50. Quan trọng là, các so sánh theo kế hoạch dựa trên lỗi MS và sai số df từ ANCOVA đã xác nhận rằng phản ứng vị trí vận động trung bình với liều 0.5 mg / kg thứ hai của AMPH vẫn lớn hơn đáng kể ở nhóm 50 so với nhóm 100, t(34) = 3.09, p <0.01 và nhóm 0, t(34) = 1.88, p <0.05 (một bên), khi sự thay đổi trước khi tiêm từ phiên 2 được kiểm soát. Do đó, nhóm 50 thể hiện phản ứng vận động sau nhạy cảm với 0.5 mg / kg AMPH cao hơn đáng kể so với nhóm 100 hoặc nhóm 0, và những khác biệt của nhóm này không được trung gian bởi sự vận động trước khi tiêm vào những ngày thử nghiệm.
Tác dụng của 1.0 mg / kg AMPH.
Đầu máy trước tiêm. Một nhóm 5 × 5 Phiên ANOVA của điểm trước khi tiêm 30-min cho các phiên nhạy cảm với 1 mg / kg AMPH mang lại hiệu quả chính của Phiên, F(4, 140) = 16.70, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <0.94, p > 0.45. So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể về vận động trước khi tiêm giữa nhóm 50 và nhóm 0 hoặc nhóm 100 trong buổi đầu tiên, t(175) <1.66, p > 0.10. Tuy nhiên, vào phiên cuối cùng, nhóm 50 (1167; 140) hiển thị nhiều ngắt tia (M; SE) hơn đáng kể so với nhóm 100 (1000; 99), t(175) = 2.35, p <0.05, nhưng không khác với nhóm 0 (1085, 120), t(175) <1.16, p > 0.20. Do đó, sự khác biệt về vận động trước khi tiêm đã góp phần vào sự khác biệt giữa nhóm 50 và 100 về phản ứng vận động với liều AMPH 1 mg / kg cuối cùng. Điểm gãy chùm trung bình (SE) tổng thể cho mẫu trong giai đoạn trước khi tiêm cho các Phiên từ 1 đến 5 là: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Đầu máy sau tiêm. Một nhóm 5 × 5 Phiên ANOVA của các phản ứng với 1 mg / kg AMPH mang lại hiệu quả chính đáng kể của Phiên, F(4, 140) = 6.72, p <0.001, một nhóm nhỏ × Tương tác phiên, F(16, 140) = 1.57, p = 0.085 và không có tác dụng chính của Nhóm, F(4, 35) <0.44, p > 0.77. Các phân tích xu hướng đa thức cho thấy một xu hướng tuyến tính quan trọng, F(1, 35) = 9.19, p = 0.005 và xu hướng hình khối, F(1, 35) = 21.63, p <0.001, qua các phiên từ 1 đến 5. Hình 6 hiển thị điểm số vận động trung bình cho mỗi nhóm và phiên.
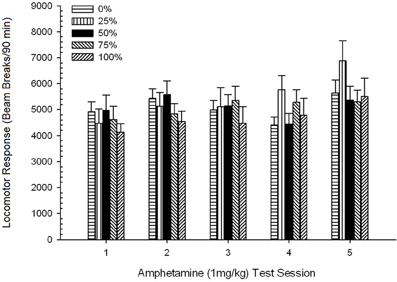
Hình 6. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) đến 1 mg / kg d-amphetamine (ip) trên các phiên hàng tuần của 5 trong các nhóm chuột Sprague Dawley (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện.
Hình vẽ cho thấy hiệu ứng Phiên phản ánh sự gia tăng đáng kể về phá vỡ chùm trung bình (SE) cho toàn bộ mẫu từ phiên 1, 4624 (213) đến phiên 5, 5736 (272), xác nhận sự xuất hiện của độ nhạy cảm với AMPH. Xu hướng hình khối biểu thị cực đại tương đối trên các phiên 1, 3 và 5, với sự sụt giảm trên các phiên 2 và 4, đặc biệt đối với các nhóm 0 và 50. Hình này cũng cho thấy, mặc dù thiếu tương tác đáng kể, nhóm 25 hiển thị phản ứng vận động lớn hơn dần dần qua các phiên và khác biệt đáng kể so với các nhóm khác trong phiên 4 và 5 (lần lượt là 9 và 22%, so với nhóm cao nhất tiếp theo). So sánh có kế hoạch cho thấy nhóm 50 không khác biệt đáng kể so với nhóm 0 hoặc 100, t(175) <0.89, p > 0.40 trong lần thử nghiệm AMPH 1 mg / kg đầu tiên hoặc cuối cùng.
Kiểm soát sự thay đổi trong phản ứng chọc mũi trong quá trình đào tạo sucrose
Hai nhóm 5 × 2 ANCOVAs về phản ứng vận động đối với 0.5 mg / kg AMPH trước và sau chế độ mẫn cảm, bao gồm cả việc chọc mũi trong quá trình luyện tập sucrose với CS hiện diện và không có CS như là đồng biến riêng biệt F(1, 18) <1.03, p > 0.31. Do đó, cách tiếp cận đáp ứng trong quá trình đào tạo không làm trung gian cho sự khác biệt giữa các nhóm trong phản ứng với 0.5 mg / kg AMPH.
Hai nhóm 5 × 5 ANCOVAs về phản ứng vận động với 1 mg / kg trong các phiên nhạy cảm với tổng số mũi chọc (hiện tại CS, vắng mặt CS) vì các đồng biến riêng biệt không mang lại tác dụng đáng kể nào cho đồng biến trong khi CS có mặt, F(4, 104) <1.04, p > 0.38 và ảnh hưởng chính biên của hiệp biến khi không có CS, F(1, 18) = 3.32, p = 0.085.
Thảo luận
Kết quả của nghiên cứu này không nhất quán ủng hộ giả thuyết rằng nhóm 50 sẽ chứng minh phản ứng vận động cao hơn qua các phiên so với các nhóm khác. Dữ liệu XNPHX mg / kg AMPH đã xác nhận sự xuất hiện của sự nhạy cảm với chế độ dùng thuốc thay thế trong ngày. Mô hình giữa các nhóm chỉ ra xu hướng nhạy cảm hơn trong các phiên sau trong nhóm 1, không có bằng chứng nào cho nhóm 25. Ngược lại, kết quả liều 50 mg / kg cho thấy xu hướng nhạy cảm cao hơn ở nhóm 0.5, đồng thời xác nhận sự gia tăng đáng kể về đáp ứng vận động giữa các nhóm với liều thứ hai so với liều AMPH đầu tiên / kg AMPH. Hiệu quả vô hiệu của tiêm nước muối đã khẳng định rằng sự mong đợi hoặc căng thẳng liên quan đến tiêm không đóng góp vào hiệu ứng AMPH.
Dữ liệu chọc mũi một lần nữa cho thấy sự gia tăng tổng thể trong cách tiếp cận đáp ứng trong suốt các buổi huấn luyện khi CS có mặt, không có sự gia tăng tương ứng khi CS vắng mặt. Do đó, các động vật dường như có được sự liên kết giữa CS và triển vọng của phần thưởng sucrose. Sự khác biệt của nhóm về tần suất chọc mũi khi CS có mặt phù hợp với tần suất phân phối phần thưởng theo lịch tương ứng, với các nhóm 75 và 100 hiển thị hầu hết các mũi chọc, nhóm 50 hiển thị số lần chọc mũi trung gian và các nhóm 0 và 25 hiển thị ít chọc mũi nhất. Những kết quả này cho thấy CS đã đến để kiểm soát cách tiếp cận đáp ứng theo cách phù hợp với xác suất thưởng chung. Mặc dù suy đoán, một lời giải thích khả dĩ cho tỷ lệ chọc mũi thấp hơn với CS có trong nhóm 50 trong thí nghiệm 2 so với thử nghiệm 1 có thể là rút ngắn khoảng thời gian giữa các thử nghiệm, vì các khoảng thời gian thử nghiệm dài hơn (thử nghiệm 1) xuất hiện để khuyến khích sự bốc đồng xu hướng và điều này có liên quan đến việc tăng doanh thu của DA trong các vỏ não trước, vỏ trước và vỏ ngoài (Dalley và cộng sự, 2002). Do đó, việc giảm 30% trong khoảng thời gian thử nghiệm trong thử nghiệm 2 (và 3) có thể đã thay đổi mức DA vỏ não và thúc đẩy chọn lọc hơn (nghĩa là được hướng dẫn bởi tần suất thưởng tương đối) so với cách tiếp cận (không được hướng dẫn bởi tần suất thưởng) đáp ứng trong nhóm 50 trong các thử nghiệm đào tạo trong thí nghiệm 2 so với thử nghiệm 1.
Việc thiếu các tác động liên quan đến đồng biến đáng kể đối với các mũi chọc trong điều kiện hiện tại của CS trong ANCOVAs chỉ ra rằng cách tiếp cận đáp ứng trong quá trình huấn luyện sucrose không làm trung gian tác động của các lịch trình CS-sucrose khác nhau đối với các phản ứng với AMPH. Hiệu quả rõ rệt của hiệp phương sai đối với tình trạng vắng mặt CS trong ANCOVA của phản ứng vận động với 1 mg / kg AMPH cho thấy xu hướng đáp ứng không có thuốc sớm đã giải thích một số thay đổi về hiệu ứng vận động của AMPH trong các phiên nhạy cảm.
Cùng với nhau, các bằng chứng cho thấy rằng các tác động của lịch sử điều hòa có thể thấy rõ hơn với 0.5 AMPH so với 1 mg / kg AMPH, và một giao thức tạo ra sự nhạy cảm trong trường hợp không có bất kỳ thao tác nào khác có thể che khuất hoặc tạo ra các hiệu ứng dư thừa nhạy cảm - thúc đẩy hành vi thao túng hành vi (nghĩa là phần thưởng biến mãn tính).
Nhạy cảm hành vi với AMPH là một hiệu ứng mạnh mẽ trong phòng thí nghiệm. Tuy nhiên, bên ngoài phòng thí nghiệm, chỉ có một số ít các cá nhân đánh bạc leo thang mạn tính đến mức bệnh lý. Mặc dù nguy cơ mẫn cảm có liên quan đến nguy cơ nghiện (hoặc tìm kiếm ma túy), đặc biệt là đối với thuốc kích thích tâm thần (Vezina, 2004; Flagel và cộng sự, 2008), nhiều yếu tố ngoài nguy cơ mẫn cảm có thể khiến người ta nghiện (ví dụ: Verdejo-Garcia và cộng sự, 2008; Conversano và cộng sự, 2012; ROLow và cộng sự, 2012). Tuy nhiên, các yếu tố đặc điểm gây tổn thương cho sự nhạy cảm có thể tương tác với lịch sử điều hòa để làm nổi bật các tác động của phần thưởng không thể đoán trước (ví dụ, lịch trình 50% CS-US) đối với phản ứng của hệ thống DA. Để điều tra khả năng này, thí nghiệm 3 đã sử dụng quy trình tương tự như thí nghiệm 2 nhưng đã sử dụng chủng Lewis thay vì chuột Sprague Dawley.
Chuột Sprague Dawley hiển thị mức độ trung gian của các nhà vận chuyển DA, với mức độ thấp hơn so với chuột biến dạng Wistar (Zamudio và cộng sự, 2005), nhưng mức độ cao hơn so với chuột Wistar-Kyoto (một chủng giống như trầm cảm của người Hồi giáo) trong các hạt nhân accumbens, amygdala, vùng não bụng và vùng đồi mồi (provia nigra (Jiao và cộng sự, 2003). Hồ sơ này có thể khiến chuột Sprague Dawley chỉ nhạy cảm ở mức độ vừa phải đối với các thao tác dược lý hoặc môi trường của chức năng DA. Ngược lại, chuột Lewis thể hiện mức độ thấp của chất vận chuyển DA cũng như thụ thể D2 và D3 DA trong nhân accumbens và vây lưng so với các chủng khác (ví dụ, F344) (Flores và cộng sự, 1998). Những khác biệt về hình thái này có thể góp phần vào phản ứng khác biệt của chuột Lewis với các thao tác DA. Chuột Lewis cũng thể hiện một loạt các phản ứng nổi bật đối với các thao tác sử dụng thuốc thử nghiệm so với các chủng khác (ví dụ, F344). Quan trọng nhất, chuột Lewis có biểu hiện nhạy cảm hơn với methamphetamine, được đặc trưng bởi phản ứng thấp với liều ban đầu nhưng phản ứng cao hơn với liều sau (Trại và cộng sự, 1994). Chuột Lewis cũng thể hiện sự nhạy cảm vận động lớn hơn với một loạt liều cocaine (Kosten và cộng sự, 1994; Haile và cộng sự, 2001). Dựa trên mô hình hiệu ứng này, chúng tôi phỏng đoán rằng chuột Lewis sẽ cho phép chúng tôi điều tra xem liệu tính nhạy cảm đối với sự nhạy cảm có khuếch đại tác động của lịch điều hòa đối với phản ứng tiếp theo với AMPH hay không.
Thử nghiệm 3
Vật liệu và phương pháp
Phương pháp này giống như trong thí nghiệm 2, ngoài việc sử dụng chuột Lewis (200 mộc 225 g khi đến, Charles River, Quebec, Canada).
Kết quả
Mũi chọc trong phiên điều hòa sucrose
Một nhóm 5 × Phiên 15 × Giai đoạn 2 (hiện tại không có CS) ANOVA của mũi chọc mang lại hiệu quả chính đáng kể của Nhóm, F(4, 34) = 6.12, p = 0.001, Phiên, F(14, 476) = 3.42, p <0.001 và Pha, F(1, 34) = 20.83, p <0.001, cũng như tương tác ba chiều đáng kể, F(56, 476) = 1.56, p = 0.008. Tấm (A, B) của hình 7 vẽ biểu đồ điểm số chọc mũi trung bình của các nhóm cho các pha CS có mặt và CS vắng mặt, tương ứng. So sánh hai bảng điều khiển cho thấy hiệu ứng chính của Pha phản ánh nhiều phản ứng chọc mũi tổng thể hơn khi CS có mặt so với khi vắng mặt. Do đó, các câu trả lời được kiểm tra xảy ra thường xuyên hơn đáng kể so với các câu trả lời trước khi trưởng thành. Các tác động chính của Nhóm và Phiên không được giải thích dễ dàng do tương tác thứ tự cao hơn. Tương tác ba chiều phản ánh sự hội tụ của điểm số cho năm nhóm ở mức thấp tương đối ổn định trong các phiên khi CS vắng mặt [Ban điều hành (B)], cùng với sự khác biệt về điểm số khi CS có mặt trong các hồ sơ tương đối riêng biệt cho từng nhóm song song với thứ tự tần suất thưởng của họ: từ cao nhất (nhóm 100) đến thấp nhất (nhóm 25) [Bảng điều khiển (A)]. Chỉ có xu hướng tuyến tính cho sự tương tác là đáng kể, F(4, 34) = 4.03, p = 0.009, phản ánh sự gia tăng thường xuyên của việc chọc mũi qua các phiên trong nhóm 100 khi CS có mặt so với hồ sơ tương đối không nhất quán về việc tăng mũi chọc qua các phiên trong các nhóm khác trong giai đoạn này.
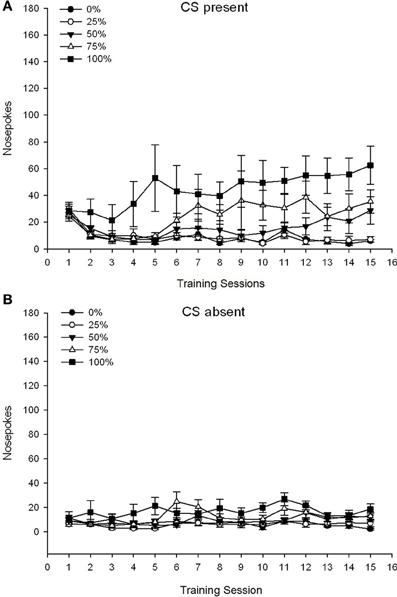
Hình 7. Các phản ứng tiếp cận trung bình (SE) (chọc mũi) trong các buổi huấn luyện sucrose sucrose trong các nhóm chuột Lewis (n = 8 / nhóm) tiếp xúc với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch biểu biến đổi 0, 25, 50, 75 hoặc 100%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. (A) Điểm khi CS có mặt (thử nghiệm 5 s × 20). (B) Điểm khi CS vắng mặt (trung bình cho 5 × 20 s trong khi đèn tắt).
Thói quen với hộp đầu máy
Một nhóm 5 × 3 Phiên ANOVA mang lại hiệu ứng chính của Phiên, F(2, 70) = 23.07, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(8, 70) <1.47, p > 0.18. Mô hình đường cong của điểm vận động trung bình (SE) xuất hiện từ phiên 1, 1076 (74), qua phiên 2, 644 (48), đến phiên 3, 762 (59). So sánh theo kế hoạch của nhóm 50 với nhóm 0 và với nhóm 100 trong các buổi sinh hoạt đầu tiên và cuối cùng cho thấy gãy chùm ít hơn đáng kể trong nhóm 50 (M = 911; SE = 109) so với nhóm 0 (M = 1103; SE = 176) trên phiên làm quen 1, t(105) = 2.02, p <0.05, nhưng không có sự khác biệt giữa nhóm 50 và nhóm 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, vào phiên này. Nhóm 50 không có sự khác biệt đáng kể so với nhóm 0 hoặc nhóm 100 trong lần định cư cuối cùng, t(105) <0.93, p > 0.30. Do đó, phản ứng định vị không dùng thuốc trung bình ở các nhóm chính không có sự khác biệt nhất quán trước khi thử nghiệm.
Buổi kiểm tra
Nước muối. Một nhóm 5 × 2 Phiên ANOVA của các phản ứng vận động trong phiên tập sự cuối cùng và phiên kiểm tra nước muối mang lại hiệu quả chính đáng kể của Phiên, F(1, 35) = 50.12, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <0.57, p > 0.68. Nhân vật 8 cho thấy điểm trung bình của nhóm trong hai phiên và chỉ ra rằng hiệu ứng Phiên phản ánh sự suy giảm đáng kể từ thói quen sử dụng đến xét nghiệm nước muối. Như vậy, nhận được tiêm cho mỗi gia nhập (ví dụ, kỳ vọng, căng thẳng) không tăng cường đáp ứng vận động.
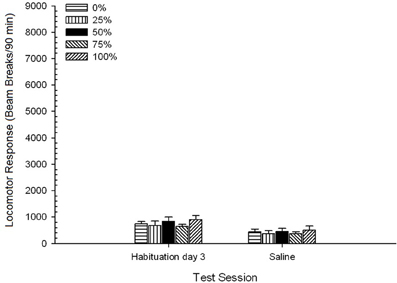
Hình 8. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) vào lần cuối của phiên làm việc không có thuốc 3 và vào phiên tiếp theo sau khi tiêm nước muối (ip, 1 ml / kg) trong các nhóm chuột Lewis (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện.
Tác dụng của 0.5 mg / kg AMPH.
Đầu máy trước tiêm. Một nhóm 5 × 2 Phiên ANOVA của đầu máy tiêm trước mang lại hiệu quả chính đáng kể của Phiên, F(1, 35) = 15.04, p <0.001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <1.19, p > 0.33. So sánh theo kế hoạch không tìm thấy sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100 trong cả hai phiên kiểm tra, t(70) <0.99, p > 0.30. Do đó, sự khác biệt cơ bản về vận động trước khi tiêm không tính đến sự khác biệt giữa các nhóm về phản ứng vận động với 0.5 mg / kg AMPH. Điểm phá vỡ chùm tia trung bình (SE) đối với giai đoạn trước khi tiêm cho phiên đầu tiên và thứ hai (sau mẫn cảm) 0.5 mg / kg là 325 (25) và 473 (36).
Đầu máy sau tiêm. Một nhóm 5 × 2 Phiên ANOVA của phản ứng vận động với liều 0.5 mg / kg được phân phối trước và sau khi dùng 1 mg / kg AMPH mãn tính mang lại hiệu quả chính của Phiên, F(1, 34) = 87.44, p <0.0001, và không có ảnh hưởng đáng kể nào khác, F(4, 34) <0.94, p > 0.45. Nhân vật 9 vẽ sơ đồ điểm số vận động trung bình cho từng nhóm và phiên và cho thấy hiệu ứng Phiên phản ánh mức tăng đáp ứng tổng thể với liều 0.5 mg / kg thứ hai, phù hợp với độ nhạy. Hình cũng cho thấy các nhóm thực hiện rất giống nhau trên phiên 1, nhưng nhóm 50 đó hiển thị nhiều hoạt động vận động hơn các nhóm khác trên phiên 2. So sánh có kế hoạch để đáp ứng với liều 0.5 mg / kg đầu tiên cho thấy không có sự khác biệt đáng kể giữa nhóm 50 và nhóm 0 hoặc nhóm 100, t(35) <1.28, p > 0.20. Tuy nhiên, nhóm 50 cho thấy phản ứng vận động cơ địa cao hơn đáng kể với liều 0.5 mg / kg thứ hai so với nhóm 0, t(35) = 4.32, p <0.001, hoặc nhóm 100, t(35) = 2.24, p <0.05.
Hình 9. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) thành 0.5 mg / kg d-amphetamine trong các phiên riêng biệt trước và sau chế độ nhạy cảm với phiên 5 của d-amphetamine (1.0 mg / kg; mỗi phiên) trong các nhóm chuột Lewis (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. *p <0.05 cho sự khác biệt trung bình giữa nhóm 50 và nhóm 0 cũng như nhóm 100, dựa trên các so sánh theo kế hoạch.
Tác dụng của 1 mg / kg AMPH.
Đầu máy trước tiêm. Một nhóm 5 × 5 Phiên ANOVA của điểm trước khi tiêm 30-min cho các phiên nhạy cảm mang lại hiệu quả chính của Phiên, F(4, 140) = 4.10, p = 0.004 và không có hiệu ứng đáng kể nào khác, F(4, 35) = 1.25, p > 0.31. So sánh theo kế hoạch cho thấy rằng đứt chùm tia trong giai đoạn trước khi tiêm (M; SE) ở nhóm 50 (395; 62) thấp hơn đáng kể so với nhóm 100 (508; 62), t(175) = 2.58, p <0.01, nhưng không phải nhóm 0, t(175) <1.83, p > 0.10, ở phiên 1 mg / kg AMPH 1. Vào phiên 1 mg / kg AMPH cuối cùng, so sánh theo kế hoạch cũng cho thấy rằng vận động trước khi tiêm ở nhóm 50 (378; 60) thấp hơn đáng kể so với nhóm 100 (650; 75 ), t(175) = 6.17, p <0.001, nhưng không thuộc nhóm 0, t(175) <1.84, p > 0.10. Do hướng của những khác biệt nhóm này (nhóm chứng = nhóm 50) ngược lại với mô hình giả thuyết, sự khác biệt của nhóm về vận động sau tiêm phù hợp với giả thuyết không thể được quy cho những khác biệt cơ bản trước khi tiêm. Số lần ngắt tia trung bình (SE) tổng thể trong giai đoạn trước khi tiêm cho các Phiên từ 1 đến 5 là: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Đầu máy sau tiêm. Một nhóm 5 × 5 phiên ANOVA phản ứng với liều 1 mg / kg mang lại hiệu quả chính đáng kể của Phiên, F(4, 140) = 6.15, p <0.001, và không có ảnh hưởng đáng kể nào khác, F(4, 35) <0.57, p > 0.68. Các phân tích xu hướng đa thức cho thấy một xu hướng tuyến tính quan trọng, F(1, 35) = 9.34, p = 0.004 và xu hướng hình khối, F(1, 35) = 5.08, p = 0.031, kết quả sau biểu thị cực đại tương đối trên các phiên 3 và 5. Nhân vật 10 vẽ các điểm số này và cho thấy, mặc dù thiếu sự tương tác đáng kể trong ANOVA, nhóm 50 thể hiện sự vận động lớn hơn đáng kể so với bốn nhóm còn lại để đáp ứng với liều 1 mg / kg cuối cùng. Theo đó, các so sánh theo kế hoạch cho thấy điểm trung bình lớn hơn đáng kể trên phiên 5 trong nhóm 50 so với tất cả các nhóm khác, t(35) > 3.68, p <0.001.
Hình 10. Phản ứng vận động trung bình (SE) (số lần phá vỡ chùm tia trong một mảng điện tử trên 90 phút) đến 1 mg / kg d-amphetamine (ip) trên các phiên hàng tuần của 5 trong các nhóm chuột Lewis (n = 8 / nhóm) trước đây đã tiếp xúc với các phiên điều hòa hàng ngày 15 với phần thưởng sucrose (giải pháp 10%) được phân phối theo lịch trình biến đổi 0, 25, 50, hoặc 75%. Các kích thích có điều kiện là một ánh sáng (120 s). Nhóm 0 nhận được số phần thưởng tương đương với nhóm 100 trong trường hợp không có các kích thích có điều kiện. *p <0.05 cho sự khác biệt trung bình giữa nhóm 50 và nhóm 0 cũng như nhóm 100, dựa trên các so sánh theo kế hoạch.
Kiểm soát sự thay đổi trong phản ứng chọc mũi trong quá trình đào tạo sucrose
Hai nhóm 5 × 2 ANCOVAs về phản ứng vận động đối với 0.5 mg / kg AMPH trước và sau chế độ mẫn cảm, bao gồm cả việc chọc mũi trong quá trình luyện tập sucrose với CS hiện diện và không có CS như là đồng biến riêng biệt F(1, 32) <0.44 p > 0.51. Hai ANCOVA 5 Nhóm × 5 Phiên của phản ứng vị trí vận động đối với 1 mg / kg AMPH trong các phiên nhạy cảm với tổng số lần chọc mũi (có CS, không có CS) khi các biến số riêng biệt không mang lại tác động đáng kể nào của đồng biến khi CS có hoặc không có, F(1, 33) <0.14, p > 0.71. Do đó, phương pháp tiếp cận không dùng thuốc đáp ứng không tính đến sự khác biệt giữa các nhóm về đáp ứng vận động cơ địa với một trong hai liều AMPH.
Thảo luận
Sự nhạy cảm đã phát triển với tác dụng của amphetamine 1.0 mg / kg lặp lại. Dữ liệu về môi trường sống và nước muối xác nhận rằng tác động này không phải do sự khác biệt từ trước, tuổi thọ hoặc phản ứng liên quan đến căng thẳng khi tiêm. Các ANCOVA với chọc mũi xác nhận rằng những tác động này không phải do hành vi tiếp cận không dùng thuốc. Bản thân dữ liệu chọc ngoáy mũi chỉ ra rằng các nhóm có được mối liên hệ giữa CS và triển vọng thưởng đường sucrose. Mức xếp hạng của các nhóm về phản ứng ngoáy mũi khi kết thúc đào tạo khớp với tần suất khen thưởng tổng thể theo các lịch trình khác nhau từ cao nhất (nhóm 100) đến thấp nhất (nhóm 0), giống như trong thử nghiệm 2. Trung bình tổng thể tương đối thấp hơn Mức độ chọc mũi trong thí nghiệm này so với thí nghiệm 1 và 2 có thể phản ánh cách tiếp cận chọn lọc hơn để đáp ứng các tín hiệu về phần thưởng ở chuột Lewis (Kosten và cộng sự, 2007).
Dữ liệu liều 0.5 mg / kg cho thấy phản ứng vận động ban đầu với AMPH ở chuột Lewis (Hình 9) đã phần nào bị triệt tiêu so với chuột Sprague Dawley (thí nghiệm 2; Hình 5), nhưng sự gia tăng trong nhóm đáp ứng với liều thứ hai ở chuột Lewis là đáng kể (gần gấp đôi đáp ứng với liều 0.5 mg / kg đầu tiên) theo chế độ AMPH phiên 5 Đáng chú ý nhất là nhóm 50 cho thấy phản ứng vận động lớn hơn so với tất cả các nhóm trừ nhóm 25 đến liều 0.5 thứ hai (nghĩa là nhạy cảm sau) và đáp ứng vận động lớn hơn tất cả các nhóm khác, bao gồm cả nhóm 25, đến liều 1 mg / kg AMPH cuối cùng (phiên nhạy cảm cuối cùng) .
Phân tích tóm tắt bảng xếp hạng nhóm qua các thí nghiệm
Để xác định độ tin cậy của sự khác biệt nhóm trong độ nhạy cảm, một phân tích không tham số đã đánh giá mức độ phụ thuộc giữa nhóm và thứ hạng của đáp ứng vận động trung bình với liều 0.5 mg / kg sau mãn tính thứ hai và liều AMPHX / kg cuối cùng của liều AMPH từ các thí nghiệm 1.0. Phân tích mang lại hiệu quả rõ rệt, = 3, p = 0.025, phản ánh thực tế rằng nhóm 50 xếp thứ nhất trong tất cả trừ một trong những so sánh. Thứ hạng cao hơn của nhóm 50 so với tất cả các nhóm khác để đáp ứng với liều 0.5 mg / kg sau mãn tính thứ hai được mô tả trong hình 5 (thử nghiệm 2) và hình 9 (thí nghiệm 3). Thứ hạng cao hơn của nhóm 50 so với các nhóm khác để đáp ứng với liều 1.0 mg / kg cuối cùng được mô tả trong hình 2 (thử nghiệm 1) và hình 10 (thí nghiệm 3). Ngoại lệ duy nhất cho mô hình này là phản ứng với liều 1.0 mg / kg cuối cùng ở chuột Sprague-Dawley trong thí nghiệm 2.
thảo luận chung
Một loạt các thí nghiệm hiện nay đã kiểm tra giả thuyết rằng việc tiếp xúc lâu dài với lịch thưởng giống như đánh bạc có thể làm nhạy cảm các con đường DA giống như phơi nhiễm mãn tính với các loại thuốc lạm dụng. Bằng chứng cho một hiệu ứng như vậy sẽ cho thấy rằng dẻo dai thần kinh, cùng loại được cho là góp phần gây nghiện ma túy, có thể được gây ra do tiếp xúc mãn tính với lịch trình thưởng không thể đoán trước. Phù hợp với các tài liệu về nghiện ma túy, đáp ứng vận động với liều 0.5 và 1.0 mg / kg liều phản ứng hệ thống DA được lập chỉ mục AMPH, với sự vận động lớn hơn để đáp ứng với liều sau đó xác định độ nhạy của hoạt động (x. Robinson và Berridge, 1993; Xỏ và Kalivas, 1997; Vanderschuren và Kalivas, 2000).
Nhìn chung, kết quả phù hợp với giả thuyết của chúng tôi. Tuy nhiên, họ cũng chỉ ra sự thay đổi đáng kể trong các hiệu ứng thử nghiệm do các yếu tố thủ tục. Tác động của lịch điều hòa rất khiêm tốn nhưng phù hợp, với nhóm 50 cho thấy phản ứng tốt hơn so với bốn nhóm còn lại đối với cả hai liều sau chế độ 5 liều. Mặc dù tổng thể F-giá trị cho các hiệu ứng liên quan đến nhóm trong các phân tích phương sai thường không có ý nghĩa, sự khác biệt nhóm chính được xác nhận bằng các so sánh theo kế hoạch theo cặp. Về vấn đề này, cần lưu ý rằng, tuy nhiên, suy nghĩ hiện tại là ý nghĩa tổng thể [cho F trong ANOVA] là không cần thiết. Trước hết, các giả thuyết được kiểm tra bằng thử nghiệm tổng thể và thử nghiệm đa so sánh khá khác nhau, với mức độ sức mạnh khá khác nhau. Ví dụ, tổng thể F thực sự phân phối sự khác biệt giữa các nhóm trên số lượng mức độ tự do cho các nhóm. Điều này có tác dụng làm loãng tổng thể F trong trường hợp một số nhóm có nghĩa là bằng nhau nhưng khác với một số nghĩa khácHowell, 1992, tr. 338). Đây chính xác là tình huống được áp dụng trong các thử nghiệm hiện tại, trong đó nhóm 50 được dự kiến sẽ khác với nhóm điều khiển 0 và nhóm 100 nhưng không có sự khác biệt giữa các nhóm kiểm soát này được dự đoán cho nhóm 25 hoặc nhóm 75.
Dữ liệu chọc mũi xác nhận rằng, trong mọi thí nghiệm, các động vật có được mối liên hệ giữa CS và triển vọng của phần thưởng sucrose. Sự tương ứng giữa tần suất chọc mũi cho các nhóm khác nhau và tần suất thưởng chung theo lịch trình đào tạo tương ứng của họ cho thấy tỷ lệ trung bình của phương pháp tiếp cận không có thuốc hướng dẫn sử dụng sucrose đáp ứng. Tuy nhiên, việc thiếu tác dụng trung gian của việc chọc mũi vào các phản ứng vận động liên quan đến nhóm với AMPH trong ANCOVAs chỉ ra rằng các quy trình riêng biệt làm nền tảng cho hai hành vi.
Trong một số trường hợp, hiệu quả của lịch điều hòa là rõ ràng khi đáp ứng với liều AMPH đầu tiên; trong các trường hợp khác, nó chỉ nổi lên sau khi dùng liều lặp lại. Sự khác biệt của nhóm trong phản ứng vận động với liều AMPH đầu tiên cho thấy rằng việc tiếp xúc với lịch trình thưởng giống như đánh bạc là đủ để tự gây ra sự nhạy cảm. Sự khác biệt của nhóm về sự vận động sau nhiều liều AMPH cho thấy một hiệu ứng tinh tế hơn có thể được đặc trưng là tính mẫn cảm, và chỉ biểu hiện khi kết hợp với tiếp xúc với chất nhạy cảm chính (ví dụ, amphetamine).
Sự khác biệt trong mô hình phản ứng giữa các thí nghiệm cho thấy rằng khoảng thời gian dài hơn giữa đào tạo và thử thách AMPH ban đầu có thể tối đa hóa cơ hội phát hiện hiệu quả nhạy cảm vốn có của điều trị điều hòa. Điều này cho thấy rằng tác động của việc tiếp xúc với phần thưởng có điều kiện có thể ủ theo thời gian, một hiện tượng cũng được thấy với sự nhạy cảm kích thích (Grimm và cộng sự, 2006). Mô hình đáp ứng với hai liều amphetamine cho thấy liều 0.5 mg / kg có thể hiệu quả hơn trong việc tiết lộ những ảnh hưởng của lịch sử điều hòa. Đến lượt nó, điều này cho thấy các hiệu ứng điều hòa theo giao thức đào tạo hiện tại có phần kém tinh tế và có thể được ngụy trang bằng hiệu ứng trần dưới liều AMPH và các điều kiện tạo ra de novo mẫn cảm.
Trong thí nghiệm 3, kiểu phản ứng hai pha với liều 0.5 mg / kg và sự xuất hiện tiến bộ của ưu thế trong nhóm 50 phù hợp với hồ sơ dự kiến cho chuột Lewis khi đáp ứng với methamphetamine (Trại và cộng sự, 1994). Điều này hỗ trợ cho tính hợp lệ của những phát hiện hiện tại và gợi ý sự chồng chéo giữa các yếu tố dễ bị tổn thương vừa phải đối với sự nhạy cảm về tâm thần và với lịch trình thưởng giống như đánh bạc.
Qua các thí nghiệm, phản ứng vận động sau nhạy cảm của nhóm 50 thường vượt quá các nhóm khác dưới các liều amphetamine khác nhau và ở các chủng động vật khác nhau. Tuy nhiên, độ biến động trong nhóm cao và kích thước hiệu ứng giữa các nhóm khiêm tốn cho thấy vai trò của các yếu tố khác trong phản ứng của hệ thống DA với amphetamine sau khi tiếp xúc với các lịch trình khác nhau của phần thưởng sucrose có điều kiện. Mặc dù phản ứng của các tế bào thần kinh DA đối với các tín hiệu khen thưởng có thể cung cấp một mô hình cờ bạc thô thiển (Fiorillo và cộng sự, 2003), giống như tất cả các mô hình, có sự mất thông tin vì lợi ích của Parsimony, tức là, để chứng minh một quy trình chính. Kết quả là, mô hình hiệu ứng trên các điều kiện CS-US trong Fiorillo et al. nghiên cứu không khái quát đầy đủ để đáp ứng vận động với amphetamine. Các tinh chỉnh tiếp theo của mô hình được kêu gọi để nắm bắt đầy đủ các khía cạnh của cờ bạc ảnh hưởng đến chức năng hệ thống DA.
Kết hợp lại với nhau, kết quả của loạt thí nghiệm này cung cấp hỗ trợ tạm thời cho giả thuyết rằng việc tiếp xúc lâu dài với lịch trình thưởng giống như đánh bạc giúp tăng cường khả năng phản ứng của hệ thống DA não trước thách thức về tâm thần. Như vậy, họ mở rộng những phát hiện của Ca sĩ và cộng sự. (2012) người đã chứng minh rằng, liên quan đến một lịch trình cố định, trước khi tiếp xúc với lịch trình tăng cường thay đổi trong mô hình hoạt động sẽ tăng cường phản ứng vận động tiếp theo với amphetamine. Cụ thể hơn, những phát hiện hiện tại chỉ ra sự không chắc chắn của việc trao phần thưởng là yếu tố quan trọng làm cơ sở cho tác động của phần thưởng thay đổi. Tầm quan trọng của các hiệu ứng trong mô hình toán tử lớn hơn đáng kể so với các hiệu ứng được tìm thấy trong các thí nghiệm hiện tại. Điều này có thể phản ánh mức độ phơi nhiễm kinh niên hơn đối với hoạt động giống như đánh bạc (55 so với 15 ngày); nó có thể phản ánh các tác động của việc yêu cầu một phản ứng của người vận hành để khơi gợi phần thưởng (nghĩa là vai trò của cơ quan) thay vì tiếp xúc thụ động, như trong nghiên cứu hiện tại. Tăng thời lượng đào tạo trong mô hình hiện tại sẽ giúp giải quyết những câu hỏi này.
Tính hợp lệ của các phần thưởng và lịch trình tăng cường thay đổi khi các mô hình đánh bạc không thể được lượm lặt từ các thí nghiệm này. Nghiên cứu trong tương lai xem xét tác động của lịch sử điều hòa đối với hành vi chấp nhận rủi ro trong các nhiệm vụ đánh bạc gặm nhấm có thể giải quyết vấn đề này. Tương tự, sự tương ứng giữa độ nhạy cảm hành vi được tìm thấy ở đây và phản ứng DA nổi bật với amphetamine gần đây được tìm thấy trong các con bạc bệnh lý phải chờ điều tra thêm (Boileau và cộng sự, 2013). Phương pháp lọc máu vi mô có thể giải quyết câu hỏi này, và dự đoán dựa trên dữ liệu của con người sẽ là việc phát hành DA lớn hơn trong nhóm kiểu hình cờ bạc 50, sẽ được quan sát rõ ràng nhất trong các vây lưng (cảm biến) chứ không phải là vây bụng (limbic) . Xác nhận phơi nhiễm phần thưởng CS + biến %% trong các mô hình khác này sẽ hỗ trợ tiện ích của nó như là một mô hình thử nghiệm chân thực của PG.
Trong khi một số hình thức cờ bạc rõ ràng đòi hỏi phản ứng bằng công cụ (ví dụ: máy đánh bạc), thì trong các hình thức cờ bạc khác (ví dụ: xổ số), mối liên hệ giữa hành động (mua vé, tức là đặt cược), tín hiệu nhận thưởng (tức là , số xổ số) và bản thân phần thưởng (số chiến thắng và phần thưởng bằng tiền) có tính lan tỏa hơn nhiều. Tuy nhiên, việc kích hoạt DA trong khoảng thời gian CS-US cũng có thể xảy ra. Điều này có thể giải thích tại sao, khi “số trúng thưởng” được công bố, sự chú ý sẽ được chú ý khi mỗi quả bóng xổ số riêng lẻ rơi liên tiếp để tạo ra một chuỗi các chữ số cụ thể trong số trúng thưởng. Mặc dù xác suất xuất hiện của một chữ số cụ thể được xác định bằng toán học, nhưng kết quả của mỗi quả bóng xổ số riêng lẻ là nhị phân — trúng (khớp với số của người chơi) hoặc trượt (không khớp với số của người chơi) —và kết quả của bất kỳ lần thử nhất định nào là không xác định. Tình huống như vậy có thể mô tả tốt hơn kinh nghiệm của nhóm 50 trong các thử nghiệm hiện tại, nơi phần thưởng được cung cấp không ngẫu nhiên nhưng cũng không thể đoán trước được và CS chỉ đơn thuần chỉ ra khả năng phần thưởng mà không tiết lộ liệu nó có xảy ra trong một thử nghiệm nhất định hay không. Máy đánh bạc có liên kết chặt chẽ với PG hơn là vé số (Cox và cộng sự, 2000; Bakken và cộng sự, 2009), cho thấy một vai trò quan trọng đối với các yếu tố công cụ (và tính trực tiếp) trong các khía cạnh bổ ích của cờ bạc cho dân số này (Loba và cộng sự, 2001). Tuy nhiên, quy trình Pavlovian được mô hình hóa trong các thử nghiệm hiện tại (phần thưởng không chắc chắn của CS +) dường như là một điều cần thiết nếu không đủ yếu tố của trải nghiệm đánh bạc.
Cùng với việc thiếu một yêu cầu công cụ rõ ràng, một số tính năng thiết kế khác có thể đã góp phần vào mô hình thử nghiệm tương đối khiêm tốn và thay đổi. Các nhóm khác nhau về phơi nhiễm sucrose tổng thể cũng như sự dự phòng giữa CS và phần thưởng sucrose. Mặc dù điều này có thể đã góp phần vào sự biến đổi giữa các nhóm, nhưng nó không thể giải thích dễ dàng tại sao động vật có phơi nhiễm sucrose lớn nhất (nhóm 100) hiển thị ít nhạy cảm hơn so với nhóm 50. Ngoài ra, nhóm 0 không nhận được kích thích trước khi tiếp xúc với sucrose trên mỗi thử nghiệm. Mặc dù điều này ngăn cản sự mong đợi về phần thưởng, nhưng nó không kiểm soát được sự hiện diện của một kích thích trước khi trao phần thưởng, tồn tại trong tất cả các nhóm khác. Để giải quyết vấn đề này, nghiên cứu trong tương lai nên bao gồm một điều kiện mà động vật nhận được phần thưởng cho mỗi thử nghiệm sau khi tiếp xúc ngẫu nhiên với một kích thích trung tính (nghĩa là sự hiện diện của chúng không báo hiệu tiềm năng cho phần thưởng).
Một hạn chế thiết kế khác là sự xuất hiện tiềm năng của hành vi bổ trợ có thể ảnh hưởng đến tác động của lịch trình đào tạo. Khi đối mặt với sự không chắc chắn, động vật có thể phát triển các hành vi mê tín được thiết kế để tăng cường kiểm soát nhận thức và giảm kích hoạt DA không chắc chắn gây ra (x. Harris và cộng sự, 2013). Do đó, có thể các khía cạnh không được kiểm soát của thiết kế thí nghiệm đã cho phép các động vật bù đắp các tác động của lịch trình điều hòa. Một hiệu ứng như vậy có thể góp phần vào phản ứng tương đối khiêm tốn và thay đổi đối với amphetamine trong nhóm 50 sau khi đào tạo CS + sucrose. Nghiên cứu trong tương lai nên ghi lại hành vi tự phát, ngoài việc chọc mũi, trong các buổi đào tạo để kiểm tra khả năng này và kiểm soát nó theo thống kê nếu nó xuất hiện. Bởi vì hành vi như vậy sẽ được dự kiến sẽ chống lại hoặc làm giảm tác động của sự không chắc chắn do lịch trình gây ra, phản ứng vận động với amphetamine trong nhóm 50 nên được tăng cường khi nó được kiểm soát (theo thủ tục hoặc theo thống kê). Do đó, thiết kế hiện tại (không được kiểm soát) cung cấp một thử nghiệm bảo thủ về tác động của phần thưởng 50% CS + đối với sự nhạy cảm với amphetamine.
Về tính hợp lệ bên ngoài, việc sử dụng chuột đực cũng hạn chế tính tổng quát của kết quả. Việc thiếu một điều kiện trừng phạt rõ ràng của người Viking cũng khác với đánh bạc, trong đó tổn thất tiền tệ lớn là phổ biến và gây ra các hiệu ứng động lực quan trọng (Nieuwenhuis và cộng sự, 2005; Singh và Khan, 2012). Khả năng tích lũy phần thưởng cũng không có trong mô hình hiện tại và tiền thưởng tích lũy trong một trò chơi máy đánh bạc đã được tìm thấy để tương tác với các thao tác DA ở người (Tremblay và cộng sự, 2011; Thông minh và cộng sự, 2013). Tương tự, cơ hội cho giải độc đắc là một sự khác biệt quan trọng giữa mô hình hiện tại và cờ bạc thực tế.
Mặc dù có những hạn chế này, các kết quả hiện tại cho thấy rằng phần thưởng CS + có thể thay đổi% có thể tham gia vào các lộ trình DA có liên quan đến các tác động củng cố của cờ bạc (Fiorillo và cộng sự, 2003; Anselme, 2013). Nhạy cảm chéo với đáp ứng với AMPH theo lịch trình giống như đánh bạc này phù hợp với vai trò then chốt của DA trong cờ bạc và các tác dụng thuốc kích thích tâm thần (Zack và Poulos, 2009) và mở rộng các nghiên cứu trước đó về việc tạo ra động lực để đánh bạc bằng AMPH trong các con bạc bệnh lý (Zack và Poulos, 2004). Các kết quả hiện tại cũng gián tiếp đề xuất rằng liều AMPH khiêm tốn, không gây ra sự phóng thích DA siêu sinh lý, có thể mô hình hóa hoạt động của não tốt hơn để đáp ứng với các tín hiệu thưởng gián đoạn (ví dụ, trong khi đánh bạc) so với phơi nhiễm ở mức cao liều thuốc kích thích (x. Vanderschuren và Xỏ, 2010). Có thể nhận được hỗ trợ trực tiếp cho sự tương ứng này bằng cách đánh giá việc phát hành DA để đáp ứng với lịch trình biến CS-US% 50% và các liều AMPH khác nhau bằng phương pháp vi phân.
Từ quan điểm thực nghiệm, mô hình Pavlovian hiện tại và mô hình hoạt động trước đây của cốt thép biến đều xuất hiện để tạo ra một kiểu hình giống như con bạc bệnh lý của con người. Do đó, chúng cung cấp một bổ sung có giá trị cho các nhiệm vụ đánh bạc gặm nhấm, mô hình hóa hành vi đánh bạc (như một biện pháp phụ thuộc), nhưng cho đến nay, chỉ sử dụng động vật khỏe mạnh, tương đương với các con bạc xã hội loài người. Dựa trên tài liệu, các động vật thường xuyên tiếp xúc với phần thưởng thay đổi cũng có thể khác nhau trong các nhiệm vụ này, đặc biệt là trong phản ứng với thuốc DA-ergic. Kết hợp kiểu hình đánh bạc của chuột với các nhiệm vụ đánh bạc có thể cho phép phát triển một cách có hệ thống các loại thuốc để điều trị PG, có thể không được thực hiện đầy đủ với các động vật khỏe mạnh. Những cải tiến thêm trong thiết kế thí nghiệm và chế độ huấn luyện, như được mô tả ở trên, sẽ cải thiện sự tương ứng giữa các động vật được huấn luyện trong mô hình này và các con bạc bệnh lý thực tế.
Từ quan điểm xã hội học lâm sàng, phát hiện tiếp xúc với phần thưởng CS + biến đổi% +, rất khớp với lịch thưởng trên máy đánh bạc thương mại (Tremblay và cộng sự, 2011), thay đổi hệ thống DA não theo những cách đáng tin cậy và bền bỉ cho thấy, trong một số trường hợp, hoạt động cờ bạc, như ma túy lạm dụng, có thể là một mầm bệnh của Hồi giáo có khả năng gây nghiện. Tuy nhiên, quy mô hiệu ứng khiêm tốn và độ biến động cao để đáp ứng với phần thưởng 50% CS + cho thấy, giống như ma túy lạm dụng, xu hướng lịch trình thưởng giống như đánh bạc để thúc đẩy nghiện sẽ phụ thuộc rất lớn vào hồ sơ rủi ro hiện có của con bạc. Tuy nhiên, để tránh những cá nhân có nguy cơ cao tiếp xúc với các tác động bất lợi tiềm ẩn liên quan đến cờ bạc, có vẻ hợp lý rằng các chính sách áp dụng để ngăn chặn việc sử dụng và giảm thiểu tác hại từ các loại thuốc lạm dụng cũng có thể được mở rộng sang đánh bạc.
Xung đột về tuyên bố lãi suất
Các tác giả tuyên bố rằng nghiên cứu được thực hiện trong trường hợp không có bất kỳ mối quan hệ thương mại hoặc tài chính nào có thể được hiểu là xung đột lợi ích tiềm năng.
Lời cảm ơn
Nghiên cứu này được tài trợ bởi các khoản tài trợ từ Hội đồng nghiên cứu khoa học tự nhiên và kỹ thuật Canada cho Paul J. Fletcher. Chúng tôi xin chân thành cảm ơn cô Djurdja Djordjevic đã chuẩn bị các số liệu.
dự án
Anselme, P. (2013). Dopamine, động lực và ý nghĩa tiến hóa của hành vi giống như đánh bạc. Hành vi. Não. 256, 1 lên 4. doi: 10.1016 / j.bbr.2013.07.039
APA (2013). Cẩm nang Chẩn đoán và Thống kê Rối loạn Tâm thần, 5th Edn. Arlington, VA: Nhà xuất bản Tâm thần Hoa Kỳ.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG và Oren, A. (2009). Hành vi đánh bạc và các vấn đề đánh bạc ở Na Uy 2007. Vụ bê bối. J. Tâm thần. 50, 333 lên 339. doi: 10.1111 / j.1467-9450.2009.00713.x
Boileau, I., Người trả tiền, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA, et al. (2013). In vivo bằng chứng cho sự giải phóng dopamine do amphetamine lớn hơn trong cờ bạc bệnh lý: nghiên cứu chụp cắt lớp phát xạ positron với [C] - (+) - PHNO. Mol. Tâm thần học doi: 10.1038 / mp.2013.163. [Epub trước khi in].
Dải, RC (1972). Củng cố, kỳ vọng và học tập. Thần kinh. Rev. 79, 394 lên 409. doi: 10.1037 / h0033120
Trại, DM, Browman, KE và Robinson, TE (1994). Tác dụng của methamphetamine và cocaine đối với hành vi vận động và dopamine ngoại bào ở vùng bụng của Lewis so với chuột Fischer 344. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G., và Dell'Osso, L. (2012). Đánh bạc bệnh lý: đánh giá một cách có hệ thống các phát hiện sinh hóa, hình ảnh thần kinh và tâm thần kinh. Thu hoạch. Rev. Tâm thần học 20, 130 Gian 148. doi: 10.3109 / 10673229.2012.694318
Cox, BJ, Kwong, J., Michaud, V. và Enns, MW (2000). Vấn đề và cờ bạc bệnh lý có thể xảy ra: cân nhắc từ một cuộc khảo sát cộng đồng. Có thể. J. Tâm thần học 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. và Robbins, TW (2002). Sự thiếu hụt trong kiểm soát xung lực liên quan đến chức năng serotonergic tăng cao trong vỏ não trước trán của chuột. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Everitt, BJ và Robbins, TW (2005). Hệ thống thần kinh củng cố cho nghiện ma túy: từ hành động đến thói quen để bắt buộc. Nat. Thần kinh. 8, 1481 Gian 1489. doi: 10.1038 / nn1579
Fiorillo, CD, Tobler, PN và Schultz, W. (2003). Mã hóa rời rạc của xác suất thưởng và sự không chắc chắn của các tế bào thần kinh dopamine. Khoa học 299, 1898 lên 1902. doi: 10.1126 / khoa học.1077349
Flagel, SB, Watson, SJ, Akil, H. và Robinson, TE (2008). Sự khác biệt cá nhân trong việc quy kết sự khuyến khích đối với một gợi ý liên quan đến phần thưởng: ảnh hưởng đến sự nhạy cảm với cocaine. Hành vi. Não. 186, 48 lên 56. doi: 10.1016 / j.bbr.2007.07.022
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V. và Kapur, S. (2005). Nhạy cảm với amphetamine, nhưng không phải là PCP, làm suy yếu sự thay đổi tập trung chú ý: đảo ngược bởi một chất chủ vận thụ thể D1 được tiêm vào vỏ não trước trán trung gian. Tâm sinh lý học (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Flores, G., Wood, GK, Barbeau, D., Quirion, R. và Srivastava, LK (1998). Chuột Lewis và Fischer: so sánh mức độ vận chuyển và thụ thể dopamine. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Frascella, J., Potenza, MN, Brown, LL và Childress, AR (2010). Các lỗ hổng não được chia sẻ mở đường cho những người nghiện không quan tâm: khắc nghiện nghiện ở khớp mới? Ann. Học viện NY Khoa học. 1187, 294 lên 315. doi: 10.1111 / j.1749-6632.2009.05420.x
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A., và Wells, B. (2006). Sự phân ly phụ thuộc vào thời gian của liều cocaine - tác dụng đáp ứng đối với sự thèm thuốc và sự vận động của sucrose. Hành vi. Dược điển. 17, 143 lên 149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Haile, CN, Hiroi, N., Nestler, EJ, và Kosten, TA (2001). Phản ứng hành vi khác biệt với cocaine có liên quan đến động lực học của protein dopamine mesolimbic ở chuột Lewis và Fischer 344. Khớp thần kinh 41, 179 lên 190. doi: 10.1002 / syn.1073
Harris, JA, Andrew, BJ và Kwok, DW (2013). Cách tiếp cận tạp chí trong một tín hiệu cho thực phẩm phụ thuộc vào Pavlovian, không phải công cụ, điều hòa. J. Exp. Thần kinh. Hoạt hình. Hành vi. Quá trình. 39, 107 lên 116. doi: 10.1037 / a0031315
Howell, DC (1992). Phương pháp thống kê tâm lý học. Boston, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW và Everitt, BJ (2002). Dopamine giải phóng ở vây lưng trong hành vi tìm kiếm cocaine dưới sự kiểm soát của một cue liên quan đến ma túy. J. Neurosci. 22, 6247 lên 6253.
Jiao, X., Pare, WP và Tejani-Mông, S. (2003). Sự khác biệt về sự phân bố các vị trí vận chuyển dopamine trong não chuột. Ăn xin. Neuropsychopharmacol. Biol. Tâm thần học 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Koob, GF và Le Moal, M. (2008). Ôn tập. Cơ chế sinh học thần kinh cho các quá trình động lực đối thủ trong nghiện. Triết gia Xuyên. R. Sóc. Thích B Biol. Khoa học. 363, 3113 lên 3123. doi: 10.1098 / rstb.2008.0094
Kosten, TA, Miserendino, MJ, Chi, S. và Nestler, EJ (1994). Các chủng chuột Fischer và Lewis cho thấy hiệu ứng cocaine khác biệt trong sở thích và điều kiện nhạy cảm hành vi nhưng không hoạt động vận động hoặc ác cảm vị giác. J. Dược điển. Exp. Có. 269, 137 lên 144.
Kosten, TA, Zhang, XY và Haile, CN (2007). Căng thẳng sự khác biệt trong việc duy trì tự quản cocaine và mối quan hệ của họ với các phản ứng hoạt động mới lạ. Hành vi. Thần kinh. 121, 380 lên 388. doi: 10.1037 / 0735-7044.121.2.380
Leeman, RF và Potenza, MN (2012). Điểm tương đồng và khác biệt giữa cờ bạc bệnh lý và rối loạn sử dụng chất gây nghiện: tập trung vào sự bốc đồng và cưỡng chế. Tâm sinh lý học (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Leshner, AI (1997). Nghiện là một bệnh về não, và nó quan trọng. Khoa học 278, 45 lên 47. doi: 10.1126 / khoa học.278.5335.45
Loba, P., Stewart, SH, Klein, RM và Blackburn, JR (2001). Thao tác các tính năng của trò chơi xổ số video tiêu chuẩn (VLT): hiệu ứng trong các con bạc bệnh lý và không bệnh lý. J. Gambl. Nghiên cứu. 17, 297 lên 320. doi: 10.1023 / A: 1013639729908
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., et al. (2007). Giải phóng dopamine do amphetamine: bị giảm sút rõ rệt trong sự phụ thuộc cocaine và dự đoán về sự lựa chọn để tự điều trị cocaine. Là. J. Tâm thần học 164, 622 Gian 629. doi: 10.1176 / appi.ajp.164.4.622
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC và Jones, SR (2005). Giảm chức năng đầu cuối của dopamine và không nhạy cảm với cocaine sau khi tự uống cocaine và thiếu thốn. Neuropsychopharmacology 30, 1455 Gian 1463. doi: 10.1038 / sj.npp.1300687
Nestler, EJ (2001). Cơ sở phân tử của nghiện nhựa cơ bản lâu dài. Nat. Mục sư thần kinh. 2, 119 lên 128. doi: 10.1038 / 35053570
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB, và Yeung, N. (2005). Hoạt động trong các khu vực não nhạy cảm với phần thưởng của con người phụ thuộc mạnh vào bối cảnh. Neuroimage 25, 1302 Gian 1309. doi: 10.1016 / j.neuroimage.2004.12.043
Xỏ, RC và Kalivas, PW (1997). Một mô hình mạch của sự biểu hiện của sự nhạy cảm hành vi với chất kích thích tâm thần giống như amphetamine. Não Res. Não Res. Rev. 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Robbins, TW và Everitt, BJ (1999). Nghiện ma túy: thói quen xấu cộng lại. Thiên nhiên 398, 567 Gian 570. doi: 10.1038 / 19208
Robinson, TE, Becker, JB, và Uy tín, SK (1982). Tạo điều kiện lâu dài cho hành vi quay do amphetamine gây ra và giải phóng dopamine trong giai đoạn đầu được tạo ra bởi một lần tiếp xúc với amphetamine: sự khác biệt giới tính. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Robinson, TE và Berridge, KC (1993). Cơ sở thần kinh của sự thèm thuốc: một lý thuyết kích thích sự nhạy cảm của nghiện. Não Res. Não Res. Rev. 18, 247 lên 291. doi: 10.1016 / 0165-0173 (93) 90013-P
Robinson, TE và Berridge, KC (2001). Nhạy cảm và nghiện. Nghiện 96, 103 lên 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Ca sĩ, BF, Scott-Railton, J. và Vezina, P. (2012). Tăng cường saccharin không thể đoán trước tăng cường vận động đáp ứng với amphetamine. Hành vi. Não. 226, 340 lên 344. doi: 10.1016 / j.bbr.2011.09.003
Singh, V. và Khan, A. (2012). Ra quyết định trong các biến thể của phần thưởng và hình phạt của nhiệm vụ đánh bạc iowa: bằng chứng về tầm nhìn xa tầm hay hay khung hình? Trước mặt. Thần kinh. 6: 107. doi: 10.3389 / fnins.2012.00107
Skinner, BF (1953). Khoa học và hành vi của con người. New York, NY: Báo chí miễn phí.
Thông minh, K., Desmond, RC, Poulos, CX và Zack, M. (2013). Modafinil tăng khả năng thưởng trong một trò chơi máy đánh bạc trong các con bạc bệnh lý có tính bốc đồng thấp và cao. Khoa học thần kinh 73, 66 lên 74. doi: 10.1016 / j.neuropharm.2013.05.015
Tremblay, AM, Desmond, RC, Poulos, CX và Zack, M. (2011). Haloperidol sửa đổi các khía cạnh công cụ của cờ bạc máy đánh bạc trong các con bạc bệnh lý và kiểm soát lành mạnh. Con nghiện. Biol. 16, 467 lên 484. doi: 10.1111 / j.1369-1600.2010.00208.x
Vanderschuren, LJ và Kalivas, PW (2000). Thay đổi trong truyền dopaminergic và glutamatergic trong cảm ứng và biểu hiện của sự nhạy cảm hành vi: một đánh giá quan trọng của các nghiên cứu tiền lâm sàng. Tâm sinh lý học (Berl.) 151, 99 lên 120. doi: 10.1007 / s002130000493
Vanderschuren, LJ và Pierce, RC (2010). Quá trình nhạy cảm trong nghiện ma túy. Curr. Hàng đầu. Hành vi. Thần kinh. 3, 179 lên 195. doi: 10.1007 / 7854_2009_21
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH và De Vries, TJ (1999). Các cơ chế dopaminergic làm trung gian cho sự biểu hiện lâu dài của sự nhạy cảm vận động sau khi tiếp xúc trước với morphin hoặc amphetamine. Tâm sinh lý học (Berl.) 143, 244 lên 253. doi: 10.1007 / s002130050943
Verdejo-Garcia, A., Lawrence, AJ và Clark, L. (2008). Tính bốc đồng như một điểm đánh dấu lỗ hổng cho các rối loạn sử dụng chất: xem xét các phát hiện từ nghiên cứu rủi ro cao, các con bạc có vấn đề và nghiên cứu liên kết di truyền. Thần kinh. Biobehav. Rev. 32, 777 lên 810. doi: 10.1016 / j.neubiorev.2007.11.003
Vezina, P. (2004). Nhạy cảm với phản ứng thần kinh dopamine midbrain và tự quản lý thuốc kích thích tâm thần. Thần kinh. Biobehav. Rev. 27, 827 lên 839. doi: 10.1016 / j.neubiorev.2003.11.001
ROLow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R., et al. (1997). Giảm đáp ứng dopaminergic trong giai đoạn đầu ở các đối tượng phụ thuộc cocaine giải độc. Thiên nhiên 386, 830 lên 833. doi: 10.1038 / 386830a0
ROLow, ND, Wang, GJ, Fowler, JS và Tomasi, D. (2012). Nghiện mạch trong não người. Annu. Rev. Pharmacol. Chất độc. 52, 321 lên 336. doi: 10.1146 / annurev-Pharmtox-010611-134625
Người chiến thắng, B. (chủ biên). (1971). Nguyên tắc thống kê trong thiết kế thí nghiệm. New York, NY: McGraw-Hill.
Zack, M. và Poulos, CX (2004). Amphetamine thúc đẩy động lực để đánh bạc và các mạng ngữ nghĩa liên quan đến cờ bạc trong các con bạc có vấn đề. Neuropsychopharmacology 29, 195 Gian 207. doi: 10.1038 / sj.npp.1300333
Zack, M. và Poulos, CX (2009). Vai trò song song của dopamine trong cờ bạc bệnh lý và nghiện thuốc kích thích tâm thần. Curr. Lạm dụng ma túy Rev. 2, 11 lên 25. doi: 10.2174 / 1874473710902010011
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F. và Flores, G. (2005). Căng thẳng sự khác biệt của mức độ thụ thể dopamine và hành vi liên quan đến dopamine ở chuột. Não Res. Bò đực. 65, 339 lên 347. doi: 10.1016 / j.brainresbull.2005.01.009
Từ khóa: cờ bạc bệnh lý, mẫn cảm, amphetamine, dopamine, không chắc chắn
Trích dẫn: Zack M, Featherstone RE, Mathewson S và Fletcher PJ (2014) Phơi nhiễm mãn tính với lịch trình giống như đánh bạc của các kích thích dự đoán có thể thúc đẩy sự nhạy cảm với amphetamine ở chuột. Trước mặt. Hành vi. Thần kinh. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Đã nhận: 01 tháng 11 2013; Được chấp nhận: 23 tháng 1 2014;
Xuất bản trực tuyến: 11 tháng 2 2014.
Sửa bởi:
Ca sĩ Bryan F., Đại học Michigan, Hoa Kỳ
Đanh gia:
Louk Vanderschuren, Đại học Utrecht, Hà Lan
Ruud Van Den Bos, Đại học Radboud Nijmegen, Hà Lan
Patrick Anselme, Đại học Liège, Bỉ
Bản quyền © 2014 Zack, Featherstone, Mathewson và Fletcher. Đây là một bài viết truy cập mở được phân phối theo các điều khoản của Giấy phép ghi nhận tác giả Creative Commons (CC BY). Việc sử dụng, phân phối hoặc sao chép trong các diễn đàn khác được cho phép, miễn là tác giả gốc hoặc người cấp phép được ghi có và rằng ấn phẩm gốc trong tạp chí này được trích dẫn, phù hợp với thực tiễn học thuật được chấp nhận. Không được phép sử dụng, phân phối hoặc sao chép mà không tuân thủ các điều khoản này.
* Thư tương ứng: Martin Zack, Phòng thí nghiệm tâm sinh lý nhận thức, Khoa thần kinh học, Trung tâm nghiện và sức khỏe tâm thần, 33 Russell Street, Toronto, TRÊN M5S 2S1, Canada e-mail: [email được bảo vệ]