Psicoloxía biolóxica. 2008 Dec 1; 64 (11): 941-50. Epub 2008 xul 26.
Teegarden SL, Nestler EJ, Bale TL.
fonte
Departamento de Bioloxía Animal da Universidade de Pensilvania, Filadelfia, PA 19104-6046, EUA.
Abstracto
TEMA:
A sensibilidade á recompensa estivo implicada como factor predispoñente para comportamentos relacionados co abuso de drogas, así como por comer de máis. Non obstante, os mecanismos subxacentes que contribúen á recompensa da sensibilidade son descoñecidos. Hipótese que unha desregulación na sinalización da dopamina podería ser unha causa subxacente da maior sensibilidade nas recompensas pola cal os estímulos gratificantes poderían actuar para normalizar o sistema.
MÉTODOS:
Usamos un modelo xenético de rato para aumentar a sensibilidade de recompensa, o rato sobreexpresor de FosB Delta, para examinar os cambios nas vías de recompensa en resposta a unha dieta sabrosa e alta. Os marcadores de sinalización de recompensas nestes ratos foron examinados basalmente e seguindo 6 semanas de exposición á dieta agradable. Os ratos foron examinados nunha proba de comportamento tras a retirada de dieta rica en graxas para avaliar a vulnerabilidade deste modelo para eliminar os estímulos gratificantes.
RESULTADOS:
Os nosos resultados demostran unha activación da vía de recompensa alterada ao longo do núcleo dos circuítos da área tegmental hipotálamo-ventral accumbens resultantes da sobreexpresión de Delta FosB no núcleo accumbens e rexións striatal. Niveis de proteínas de unión á resposta de adenosina monofosfato cíclico fosforilado (AMPc) (pCREB), factor neurotrófico derivado do cerebro (BDNF), e a dopamina ea fosfoproteína regulada por adenosina monofosfato cíclico cunha masa molecular de 32 kDa (DARPP-32) no núcleo accumbens reducíronse nos ratos Delta FosB, suxerindo unha reducida sinalización de dopamina. Seis semanas de exposición á dieta rica en graxas melloraron completamente estas diferenzas, revelando a potente capacidade gratificante dunha dieta agradable. Os ratones Delta FosB tamén mostraron un aumento significativo na actividade locomotora e nas respostas relacionadas coa ansiedade. 24 horas despois da retirada de graxa elevada.
CONCLUSIÓNS:
Estes resultados establecen unha sensibilidade subxacente aos cambios de recompensa relacionados coa desregulación da sinalización de Delta FosB e da dopamina que se poden normalizar con dietas saborosas e pode ser un fenotipo predispoñente nalgunhas formas de obesidade.
introdución
A pesar do crecente coñecemento dos sistemas neuronais que controlan o apetito e a saciedade, as taxas de obesidade continúan aumentando nos Estados Unidos. Os tratamentos con fármacos actuais teñen unha eficacia limitada e as modificacións do comportamento sofren un mínimo cumprimento a longo prazo (1). O consumo de alimentos calóricamente densos e saborosos vinculouse a cambios no estrés e vías de recompensa no cerebro, o que suxire que as propiedades gratificantes de devanditos alimentos poden ignorar os sinais de balance de enerxía (2-4). Os alimentos ricos en graxas actúan como recompensas naturais, activando centros de recompensa do cerebro de forma similar ás drogas de abuso e, como tal, utilizáronse en paradigmas de autoadministración (5-8). Así, é probable que os comportamentos e a motivación para comer en exceso e abuso de drogas compartan mecanismos subxacentes comúns, abrindo novas vías de tratamento para ambas condicións.
Ao estudar a relación entre alimentos e vías saborosas que regulan a recompensa eo estrés no cerebro, identificamos previamente marcadores moleculares e bioquímicos de recompensa reducida e incremento da tensión tras a retirada dunha dieta palatábel e rica en graxas (HF). Similar aos fármacos de abuso, a exposición a unha dieta agradable nos nosos estudos deu como resultado un aumento dos niveis do factor de transcrición BFosB no núcleo accumbens (NAc), unha estrutura central de recompensa do cerebro (9, 10). Os ratos que indebidamente sobreexpresan FosB mostran un aumento da resposta instrumental para unha recompensa de alimentos (11), converténdose nunha ferramenta valiosa para examinar o papel da sensibilidade de recompensa e a desregulación a longo prazo do sistema de recompensas nas respostas moleculares e bioquímicas a unha dieta agradable.
No presente estudo utilizamos os ratos toFosB-sobreexpresivos para examinar as alteracións a longo prazo nos marcadores de recompensa no neurocircuitry da área Tegmental (VTA) de NAc-hipotálamo-ventral en resposta a unha dieta HF agradable. Con base en estudos previos destes ratos sensibles á recompensa, fixemos a hipótese de que os cambios inducidos por -FosB na sensibilidade da recompensa implican unha desregulación na sinalización da dopamina resultante da retroalimentación na VTA. Ademais, pensamos que a exposición a unha recompensa natural dunha dieta HF de densidade enerxética normalizaría entón o sistema dopaminérgico nestes ratos, o que resultaría nunha resposta esaxerada ao estrés da retirada desta dieta.. O aspecto único de utilizar unha dieta agradable como sustancia gratificante permítenos incluír os insumos hipotalámicos para recompensar os circuítos nun fenotipo que pode ser predictivo dunha poboación predispuesta á obesidade resistente ao tratamento. Para examinar esta hipótese, estudamos os marcadores da neurotransmisión de dopamina, incluíndo pCREB, BDNF e DARPP-32 no NAc e na tirosina hidroxilase e no transportador de dopamina no VTA, tras a exposición ao HF. Tamén examinamos marcadores específicos de balance enerxético coñecidos por influír na produción de dopamina, incluíndo receptores de leptina e orexina na expresión VTA e orexina no hipotálamo lateral.
Materiais e Métodos
animais
Xa se xeraron ratas bitransxénicas macho que expresa de forma excesiva ΔFosB en neuronas positivas en dinorina no NAc e no estriado dorsal (Kelz et al., 1999) na Universidade de Texas Southwestern Medical Center. probado na Universidade de Pensilvania. Todos os ratos foron mantidos en doxiciclina (57 μg / ml no auga potable) ata a chegada á Universidade de Pensilvania. Para inducir a sobreexpresión, eliminouse a doxiciclina (n = 6) (100). Os ratos de control (n = 23) continuaron a recibir a droga. Os ratos foron asignados a grupos dietéticos oito semanas despois da eliminación da doxiciclina, momento no que se demostrou que a expresión de tempo alcanza os niveis máximos (12). Os ratos mantivéronse nun ciclo de luz e escuridade 26: 13 (as luces de 12) con alimentos e auga dispoñibles ad libitum. Todos os estudos realizáronse de acordo con protocolos experimentais aprobados polo Comité de uso e uso de animais institucionais da Universidade de Pensilvania e todos os procedementos foron realizados de acordo coas directrices institucionais.
Exposición á dieta
Os ratones mantivéronse no chow da casa (n = 16) ou colocados en HF (n = 16-17) durante seis semanas. O chow house (Purina Lab Diet, St. Louis, MO) contiña 4.00 kcal / g, consistente en proteínas 28%, graxa 12% e carbohidratos 60%. A dieta HF (Dietas de investigación, New Brunswick, NJ) contiña 4.73 kcal / g, consistente en proteínas 20%, graxa 45% e carbohidratos 35%.
Bioquímica e expresión xénica
Os ratos foron analizados tras seis semanas de exposición á dieta. Os cerebros foron retirados do cranio e conxelados enteiros sobre xeo seco ou o NAc disecado (aproximadamente 0.5 - 1.75 mm de bregma, a profundidade de 3.5 - 5.5 mm) e conxelado en nitróxeno líquido. O tecido almacenábase en -80 ° C ata o ensaio.
Análise bioquímica
Nos métodos suplementarios descríbense os métodos de transferencia Western. Os anticorpos utilizados foron: Cdk5, CREB e BDNF (1: 500, Santa Cruz Biotechnology, Santa Cruz, CA) e phospho-CREB (pCREB) (Ser 133) (1: 500, Cell Signaling Technology, Danvers, MA).
Autoradiografía do receptor
En materiais suplementarios descríbense métodos detallados para a autoradiografía. Os ligandos usados foron 2 nM H3 - SCH 23390 e 5 nM H3 - spiperona (PerkinElmer, Boston, MA).
Hibridación in situ
O procesamento de tecidos e hibridación realizáronse como se describiu anteriormente (14). A sonda DARPP-32 foi proporcionada xentilmente por P. Greengard (Universidade Rockefeller) e pola sonda orexin por J. Elmquist (Centro médico suroeste da Universidade de Texas). As diapositivas ensaiadas para DARPP-32 foron aptas para filmar para os días 3 e as diapositivas ensaiadas para a orexina foron aptas para rodar para os días 4. A cuantificación das imaxes de película realizouse como se describiu anteriormente (10).
QRT-PCR
O ARN foi illado da VTA e expresión de xenes individuais avaliados usando ensaios de expresión xénica TaqMan (Applied Biosystems, Foster City, CA). Pódense atopar métodos detallados e análises estatísticas en materiais suplementarios.
Análises de comportamento
Para examinar os efectos da sensibilidade de recompensa nos cambios de comportamento inducidos pola dieta, un subconxunto de ratos foi retirado da FH despois de catro semanas de exposición e volveu ao xardín da casa (n = control 9, n = 8 BFosB). Vinte e catro horas despois da retirada, os ratos foron expostos á proba de campo aberto de acordo co noso paradigma de abstención dietética publicado anteriormente (10). Resumidamente, o rato foi colocado no centro do aparello de campo aberto e controlado durante cinco minutos. Medíronse as cruces de liña total, o boli fecal, o tempo no centro e as cruces cara ao centro.
Estatística
Todos os datos excepto Western blot foron analizados usando un ANOVA bidireccional seguido da proba PLSD de Fisher con tratamento con doxiciclina (expresión ΔFosB) e condición da dieta como variables independentes. Para análises RT-PCR, utilizouse un valor P reducido para corrixir múltiples comparacións dentro de grupos de xenes relacionados (ver materiais suplementarios). As transferencias occidentais analizáronse usando a proba t dun estudante con tratamento con doxiciclina como variable independente, comparando densidades ópticas dentro da mesma transferencia. Todos os datos preséntanse como media ± SEM.
Resultados
Diferenzas bioquímicas basais
Para dilucidar as vías moleculares que subxacen á maior sensibilidade á recompensa en ratos con sobreexpresión de osFosB, examináronse niveis de varias moléculas de sinalización clave no NAc. Houbo unha tendencia a un aumento dos niveis de Cdk5 no NAc de ratos ΔFosB en comparación cos animais controlados por camaradas mantidos en doxiciclina (F = 5.1, P = 0.08; Fig. 1A). Os ratos ΔFosB expresaron niveis significativamente reducidos de pCREB (F = 7.4, P <0.05; Fig. 1B), así como niveis totais de CREB (F = 5.4, P = 0.05; Fig. 1C). Tamén se observou unha redución significativa de BDNF no NAc de ratos ΔFosB (F = 10.6, P <0.05; Fig. 1D).
figura 1
Os ratos que sobreexpresan ΔFosB mostraron marcadores bioquímicos de sinalización reducida de dopamina no NAc
A inxestión de alimentos e o peso corporal na dieta rica en graxas
A continuación, examinamos os efectos dunha dieta de IC naturalmente gratificante sobre as alteracións das moléculas de sinalización nos ratos ereFosB que sobreexpresan. Non houbo diferenzas entre os ratos ΔFosB e os controis na inxestión de alimentos nin en casa nin en IC. Non obstante, houbo unha diminución global da inxestión calórica normalizada ao peso corporal cando se expuxo a IC que foi específica para os ratos ΔFosB (F = 11.2, P <0.01; Fig. 2A). Ao final das seis semanas de exposición á dieta, os ratos que recibían IC pesaban significativamente máis que os que estaban na dieta chow (F = 17.2, P <0.001) e os ratos osFosB pesaban menos que os controis (F = 5.6, P <0.05; Fig. 2B). Este efecto foi específico para as diferenzas entre grupos na dieta do chow (P <0.05).
figura 2
Os ratos sobreexpresivos de BFosB non mostraron diferenzas na inxestión de alimentos tanto na dieta chow como na alta graxa (HF)
Diferenzas bioquímicas na dieta rica en graxas
Para determinar como se poderían alterar as diferenzas basais na sinalización de NAc por dieta HF, as mesmas proteínas de sinalización estudadas no inicio foron examinadas en animais que recibiron seis semanas de HF. Non houbo diferenzas significativas nos niveis de Cdk5 (Fig. 3A). Os niveis de pCREB e CREB total xa non eran diferentes despois de seis semanas de HF (Fig. 3B, C). Os niveis de BDNF eleváronse significativamente nos ratos ΔFosB logo de seis semanas de exposición ao HF (F = 6.5, P = 0.05; Fig. 3D).
figura 3
A dieta con alto contido de graxa (HF) mellorou as diferenzas de sinalización observadas no NAc dos ratos xFosB que sobreexpresan
Autoradiografía do receptor de dopamina
Utilizamos autoradiografía de receptores para avaliar se as alteracións inducidas por ΔFosB na sinalización de dopamina no NAc están relacionadas con cambios na expresión do receptor de dopamina (Fig. 4A). A dieta rica en graxas parece aumentar lixeiramente a densidade de unión ao receptor de dopamina D1 (P = 0.14), e esta diferenza foi maior nos ratos ΔFosB (Fig. 4B). Tamén houbo unha tendencia cara a un aumento da área de unión a D1 despois de HF (P = 0.06), e as probas post hoc mostraron que isto era significativo nos ratos ΔFosB (P <0.05; Fig. 4C). En contraste cos receptores D1, non hai cambios na densidade de unión do receptor D2 (control chow = 97.6 ± 6.9, control HF = 101.1 ± 8.2, ΔFosB chow = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) ou área de unión (control chow = 47.3 ± 3.4, control HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) no NAc observáronse.
figura 4
A dieta rica en graxas (HF) levou a cambios na unión do receptor de dopamina D1 e na expresión DARPP-32 no núcleo accumbens (NAc) dos ratos sobreexpresores xFosB
Expresión DARPP-32 no NAc
Utilizouse a hibridación in situ para determinar os niveis de expresión de DARPP-32 no NAc (Fig. 4D). A dieta rica en graxas aumentou significativamente a expresión de DARPP-32 nesta rexión cerebral (F = 5.1, P <0.05) e houbo unha interacción significativa entre a dieta e a expresión de ΔFosB (F = 8.9, P <0.05), con ratos ΔFosB mostrando unha maior cambio inducido pola dieta (Fig. 4E). Unha diferenza basal na expresión de DARPP-32 entre os ratos control e os FosB revelouse mediante probas post hoc (P <0.01), así como un aumento significativo da expresión de DARPP-32 nos ratos ΔFosB en HF (P <0.01).
Expresión xenética no VTA
QRT-PCR utilizouse para avaliar os cambios na expresión xénica no VTA, dirixido a varios xenes clave implicados previamente na regulación da recompensa. Todas as mostras normalizáronse en β-actina. Para garantir que a expresión da β-actina non se alterou polo tratamento, realizouse un ensaio separado para comparar a β-actina cun segundo control interno, GAPDH. Non houbo diferenzas significativas na expresión de β-actina (valores de ΔCT, β-actina - GAPDH: control chow = 2.29 ± 0.21, control HF = 2.01 ± 0.04, ΔFosB chow = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
Observouse unha tendencia á interacción entre a expresión de osFosB e o tratamento da dieta para a expresión de tirosina hidroxilase (F = 3.6, P <0.06; Fig. 5A). Seis semanas de exposición a HF parecen diminuír a expresión de tirosina hidroxilase en ratos control e aumentar a expresión en ratos ΔFosB. Observouse unha interacción significativa entre a expresión de osFosB e a exposición á dieta para a expresión do transportador de dopamina (F = 6.7, P <0.03; Fig. 5B). Semellante á tirosina hidroxilase, a exposición a HF reduciu a expresión do transportador de dopamina en ratos control e aumentou significativamente a expresión en ratos ΔFosB (P <0.05). A diferenza basal na expresión do transportador de dopamina entre os ratos control e os FosB non alcanzou significación (P = 0.16), pero despois de 6 semanas de HF, os ratos ΔFosB expresaron niveis significativamente elevados de transportador de dopamina en comparación cos controis (P <0.05).
figura 5
A exposición á dieta rica en graxas (HF) e a expresión ΔFosB provocaron cambios na expresión dunha serie de moléculas clave no VTA.
Houbo unha tendencia que indica un efecto do aumento da expresión de osFosB para reducir os niveis de TrkB no VTA (F = 5.7, P <0.04; Fig. 5C). Aínda que non houbo efectos principais sobre a expresión do receptor κ-opioide, houbo unha tendencia á expresión reducida en ratos ΔFosB (P = 0.08; Fig. 5D). A expresión do receptor de leptina tamén se determinou no VTA. Atopouse un efecto significativo da exposición á dieta (F = 6.1, P <0.03), con HF que reduciu significativamente os niveis do receptor de leptina no VTA tanto en ratos osFosB como en control (Fig. 5E). Tamén se examinou a expresión do receptor 1 de orexina no VTA. Houbo un efecto significativo da dieta na expresión do receptor de orexina (F = 9.0, P <0.02), con ratos expostos a HF que expresaban niveis máis altos no VTA (Fig. 5F). Tamén houbo unha tendencia para que os ratos ΔFosB expresasen niveis globais máis altos de receptor 1 de orexina nesta rexión cerebral (P <0.05).
Expresión de oríxenas no hipotálamo lateral
Medimos niveis de orexina no hipotálamo lateral, a orixe da inervación orexinérxica do VTA, por hibridación in situ (Fig. 6A). Houbo unha interacción significativa entre a expresión de osFosB e a exposición á dieta na expresión de orexina (F = 9.1, P <0.01), con HF aumentando significativamente os niveis de orexina en ratos control (P <0.05) e diminuíndo a expresión en ratos ΔFosB (Fig. 6B). Aínda que non houbo diferenzas significativas na expresión de orexina no estado basal, tras 6 semanas de HF, os ratos ΔFosB expresaron niveis significativamente reducidos de orexina en comparación cos controis (P <0.05).
figura 6
A dieta con alto contido de graxa (HF) tiña efectos diferenciales na expresión de orexina nos ratos de control (Ctrl) e ΔFosB.
BeAnálise havioral
Para avaliar as alteracións na excitación e emocionalidade debidas ao cambio na dieta, os ratos foron expostos á proba en campo aberto 24 horas despois da retirada da dieta HF (10). Os cruzamentos de liña totais, que se puntuaron como medida de excitación, víronse afectados significativamente pola expresión de osFosB (F = 6.6, P <0.05) e pola dieta (F = 4.6, P <0.05; Fig. 7A). Os ratos ΔFosB foron máis activos no novo ambiente que os contrils e as probas post hoc mostraron que os ratos retirados de HF foron significativamente máis activos que os expostos a chow (P <0.05). Os boli fecais contáronse como unha medida do comportamento similar á ansiedade (10). Houbo un efecto principal da expresión de osFosB (F = 10.2, P <0.01), con ratos con sobreexpresión de osFosB producindo máis boli fecal no ambiente novidoso, particularmente nos grupos de chow domésticos e de retirada de HF (Fig. 7B). Os ratos osFosB mantidos na dieta HF produciron menos boli fecal que os mantidos no chow e os retirados 24 horas antes da proba. Os ratos control non parecen estar afectados pola dieta. Non houbo efectos significativos nin da expresión de osFosB nin da dieta no tempo que pasou no centro do campo aberto (control chow = 14.5 ± 3.1 seg, control HF = 18.0 ± 3.2 seg, control W / D = 15.4 ± 1.9 seg, ΔFosB chow = 16.9 ± 2.4 seg, ΔFosB HF = 13.1 ± 3.9 seg, ΔFosB W / D = 19.8 ± 2.6 seg).
figura 7
Os ratos que sobreexpresan BFosB foron máis sensibles aos efectos da retirada de dieta rica en graxas (HF)
Conversa
No tratamento da obesidade, hai unha necesidade crítica para a identificación de factores que inflúen na susceptibilidade á alimentación excesiva e á ganancia de peso. As vías de recompensa cerebral xogan un papel importante na motivación e resposta a alimentos e cambios dietéticos agradables (6, 10, 15, 16). Como os sinais orexixénicos e anorexixénicos poden influír directamente na sinalización de recompensas a través dun circuíto hipotálamo-VTA-NAc, a elucidación de xenes que responden a dietas ricas en enerxía nos centros de recompensa pode proporcionar novas dianas terapéuticas no tratamento da obesidade (17, 18). Polo tanto, analizamos os marcadores bioquímicos e moleculares de sinal de recompensa e equilibrio enerxético ao longo do circuíto hipotálamo-VTA-NAc en resposta a unha dieta HF en ratos ΔFosB-sobreexpresivos como modelo de sensibilidade mellorada aos cambios de recompensa (13, 19, 20) , ea sensibilidade comportamental tras a retirada da dieta. Hipotetizamos que a desregulación basal da sinalización de dopamina nos ratos ΔFosB normalizaríase polos efectos gratificantes dunha dieta de HF, que englobase a intersección dos sinais de balance de enerxía e do sistema de dopamina.
Para examinar os marcadores indicativos dunha desregulación na sinalización da dopamina no NAc, examinamos os niveis dos receptores D1 e os efectores posteriores. Aínda que non houbo diferenzas significativas na unión do receptor D1, houbo unha tendencia para a exposición a HF para aumentar a área de unión nos ratones BFosB. Isto é interesante porque a inducción de ΔFosB por fármaco e recompensas naturais parece predominar no subtipo de dinorfinas-positivas de neuronas espiñas medianas que expresan principalmente os receptores D1 (9, 21). Os niveis da dianteira de sinalización de dopamina baixo pCREB foron reducidos significativamente nos ratones FosB, que apoian a activación reducida do receptor D1 nesta rexión cerebral (22, 23). Curiosamente, tamén se detectou unha diminución significativa dos niveis de CREB total en ratos BFosB, o que suxire unha nova capacidade reducida para a transdución de sinais de dopamina que pode ser secundaria á retroalimentación resultante dunha diminución prolongada de pCREB (24). A expresión BDNF está regulada por pCREB, é elevada coa activación de D1 e é un importante mediador da neuroplasticidade relacionada coa recompensa no NAc (25, 26). En consecuencia, detectamos unha diminución significativa da proteína BDNF no NAc dos ratones ΔFosB.
Todas as neuronas espiñas medias do NAc expresan DARPP-32 (27). Os seus numerosos efectores posteriores convérteno nun xogador vital nas vías de recompensa (28), e estivo implicado na adicción ás drogas e noutros trastornos que implican o sistema de dopamina incluíndo trastornos afectivos e esquizofrenia (27, 29). Detectamos profundas reducións basais na expresión DARPP-32 no NAc dos ratos ΔFosB. A expresión DARPP-32 está regulada por BDNF e, polo tanto, a expresión reducida pode estar relacionada directamente coas reducións nos niveis BDNF detectados nos ratos ΔFosB (27, 29, 30). Mesmo os cambios moderados no estado de fosforilación de DARPP-32 poden conducir a alteracións substanciais da sinalización intracelular dentro do NAc (27). Estudos anteriores non informaron de cambios na proteína DARPP-32 en ratos ΔFosB tras unha eliminación de 12-wk de doxiciclina cando se realizou unha avaliación estriatal máis ampla (31), suxerindo que os efectos de onFosB en DARPP-32 poden ser específicos do tempo e da rexión.
Cremos que a redución drástica dos índices de sinalización de dopamina no NAc dos ratos ΔFosB probablemente implicou cambios nas neuronas de proxección de dopamina VTA, aínda que ΔFosB non está sobreexpresado nestas neuronas.. Polo tanto, examinamos a expresión de xenes relacionados coa dopamina na VTA, incluíndo a tirosina hidroxilase eo transportista de dopamina. Os niveis de tirosina hidroxilase e transportadora de dopamina están correlacionados positivamente coa produción de dopamina. Houbo unha tendencia para que os ratones FosB mostraban reducida a tirosina hidroxilase e unha redución significativa do transportista de dopamina, de acordo coa disregulación da sinalización de dopamina no NAc. Como estas reducións basais nos xenes relacionados coa dopamina na VTA dos ratones ΔFosB presuntamente reflicten a retroalimentación da NAc durante a sobreexpresión ΔFosB a longo prazo, examinamos a expresión do receptor BDNF, TrkB, como un posible mecanismo de retroalimentación NAc ao VTA (32). Similar ao transportador de tirosina hidroxilase e dopamina, a expresión de TrkB tamén mostrou unha tendencia a ser basicamente reducida nos ratones thatFosB que non alcanzaron importancia cando se corrixiron para comparacións múltiples. O complexo BDNF-TrkB pódese transportar de xeito retrogrado e actuar dentro do VTA para afectar a expresión xénica local e promover o crecemento e mantemento celular (33). Ademais, a activación do BDNF de TrkB presináptico dentro do NAc pode estimular directamente a neurotransmisión de dopamina (32), soportando unha diminución subxacente da sinalización de dopamina nestes ratos.
A activación da dinorfina dos receptores opioides κ regula a sinalización da dopamina e é outro mecanismo polo cal o NAc proporciona feedback ao VTA (34). Descubrimos que a expresión do receptor κ opiáceos no VTA mostrou unha tendencia a ser reducida nos ratones FosB. Como se demostrou que a sobreexpresión de FosB diminúe a expresión da dinorfina no NAc (20), os ratos ΔFosB probablemente teñen reducións profundas na VTA κ netasactivación de opiáceos. Aínda que a sinalización de dinorfina normalmente exerce un efecto inhibidor sobre as neuronas dopaminéticas (35), as ratas que mostran unha autoadministración mellorada de drogas de abuso presentan niveis reducidos de dinorfina no NAc, apuntando a un papel na sinalización de dinorfina basalmente reducida na mellora da sensibilidade á recompensa (36 , 37). A desregulación do sistema dinorfina-opioide estivo relacionada coa adquisición e persistencia do abuso de drogas, apoiando un balance crítico de sinalización de opioides na normalización das vías de dopamina. (38).
Baseado na gratificante capacidade dunha dieta de densidade enerxética densa de enerxía, pensamos que unha desregulación na sinal de recompensa de dopamina e opioides nos ratos BFosB predisporía a estes ratos para mellorar as respostas a tal dieta, normalizando así o sistema de recompensa mediante a activación do hipotálamo -VTA-NAc circuíto. Durante a exposición á dieta de seis semanas, non se observaron diferenzas na inxestión de alimentos entre wereFosB e ratos control, suxerindo que os cambios atopados nos marcadores bioquímicos e moleculares da sinalización de recompensa nos ratos ΔFosB non se debían a diferenzas nas calorías consumidas. Como se anticipou, as diferenzas basais detectadas nos niveis de pCREB, CREB total, BDNF, DARPP-32 e κ-opiáceos entre ΔFosB e rato control foron atenuadas, probablemente debido ao aumento da produción de dopamina en ratos ΔFosB en HF (29, 39-41) .
O exame de tirosina hidroxilase e do transportista de dopamina no VTA revelou sorprendentes respostas opostas de ΔFosB e rato control despois de HF. Os ratos de control mostraron unha diminución na expresión do transportista de tirosina hidroxilase e dopamina, mentres que os ratos BFosB mostraron unha expresión aumentada de ambos xenes relacionados coa dopamina. Curiosamente, a expresión da tirosina hidroxilase é alterada no VTA por administración crónica de cocaína ou metanfetamina (42-44), o que suxire que os ratos ΔFosB poden atopar a recompensa natural de HF máis saliente que os ratos control.
Co fin de examinar como se poden transmitir sinais de potencial hipotálamo para a VTA que reflicten o equilibrio enerxético, tamén se examinou a expresión do receptor de leptina e do receptor 1 de orexina. Os niveis de leptina circulante aumentan mediante HF e, á súa vez, a leptina pode actuar no VTA para alterar a sinalización da dopamina (18, 45). A expresión do receptor de leptina VTA diminuíu de xeito similar por HF en ratos ΔFosB e control, de acordo coa ganancia de peso similar e a inxestión de dieta mentres estaba en HF. A expresión 1 do receptor de orexina en graxa alta tamén aumentou no VTA de ambos os ratos ΔFosB e control. A Orexina activa as neuronas de dopamina na VTA, promove a plasticidade VTA e aumenta os niveis de dopamina no NAc (46-48). Demostrouse que a dieta rica en graxas aumenta a expresión da orexina en ratos, de acordo coas nosas observacións (49, 50). Así, o aumento da expresión do receptor de orexina, así como os cambios na sinalización da leptina no VTA, poderían promover a recompensa da dieta tanto nos ratos BFosB como nos controles, soportando unha disociación entre as vías que transmiten sinais de balanza enerxética e as relacionadas directamente coa recompensa.
Para examinar os efectos provocadores da retirada de recompensa, os ratos foron examinados nunha proba de campo aberto horas de 24 despois da eliminación de HF. Os ratones BFosB eran máis sensibles aos efectos agudos da retirada da dieta preferida, mostrando unha maior actividade de excitación e produción de boli fecal na novela aberta en comparación con todos os demais grupos de control e dieta. Os ratones BFosB tamén mostraron un interesante patrón de comportamento nesta proba que suxire unha sensibilidade de recompensa e estrés, coa dieta HF reducindo inicialmente a produción de boli fecal en relación ao chow, e a retirada aumentando de novo esta resposta relacionada coa ansiedade. Este aumento observado na actividade de campo aberto non se correlacionou cos cambios na expresión de orexina, o que suxire unha relación coa excitación inducida polo estrés que non é só un efecto dos cambios na sinalización mediada por orexina. En xeral, estes datos apoian a nosa hipótese de que os ratos BFosB serían máis sensibles aos efectos agudos da retirada da dieta preferida debido á súa sensibilidade máis recompensa..
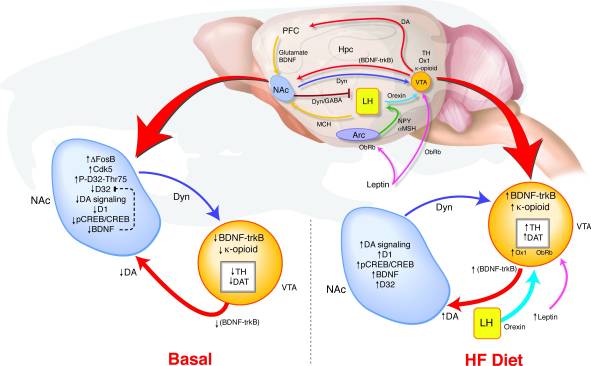
Como a sobreexpresión a longo prazo de ΔFosB no NAc leva a tales cambios no comportamento e na sinalización de recompensas? Propuxemos un modelo de detección coincidente VTA no que a retroalimentación do NAc e do hipotálamo transmite sinais en relación ao estado de recompensa para determinar a regulación do sistema de dopamina que pode soportar unha conexión entre a desregulación da vía de recompensa e unha predisposición á obesidade (fig. 8). Durante a exposición a HF, múltiples entradas que reflicten o estado de equilibrio enerxético e de recompensa converxen ao VTA. Os aumentos da sinalización de leptina e orexina, así como a retroalimentación da NAc ao hipotálamo lateral poden afectar a como estes sinais orexixénicos responden a HF nos ratos ΔFosB (17, 18, 45, 47, 51-53). As elevacións inducidas por unha dieta rica en graxas no BDNF poden proporcionar retroalimentación á VTA, promovendo aínda máis os cambios na expresión xénica relacionada coa dopamina.
figura 8
A dieta alta en graxa (HF) normaliza a sinalización de recompensas disreguladas nos ratos BFosB
Estes resultados delinean marcadores moleculares de sensibilidade de recompensa e indican que a desregulación a longo prazo do sistema de dopamina pode predispor un individuo á adicción e á obesidade. Ademais, estes datos proporcionan un paso importante para identificar novas dianas terapéuticas no tratamento e prevención da obesidade e outros trastornos que poden centrarse no sistema de recompensa. No futuro, será importante investigar como este sistema responde á eliminación da dieta HF, así como investigar as diferenzas sexuais na sensibilidade á recompensa e á exposición a unha dieta rica en graxas.
Material complementario
Supp. Métodos
Fai clic aquí para ver. (61K, doc)
Grazas
Os autores desexan agradecer a Cathy Steffen axuda para a mellora e transferencia de animais. Este traballo foi apoiado por unha bolsa do Centro de Diabetes da Universidade de Pensilvania (DK019525) e por subvencións do Instituto Nacional de Saúde Mental (R01 MH51399 e P50 MH66172) e Instituto Nacional de Abuso de Drogas (R01 DA07359).
Notas ao pé
Revelacións financeiras: Todos os autores declaran que non teñen intereses financeiros biomédicos ou posibles conflitos de interese.
References
1. Wadden TA, Berkowitz RI, Womble LG, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Kaplan R, Stunkard AJ. Proba aleatoria de modificación do estilo de vida e farmacoterapia para a obesidade. N Engl J Med. 2005; 353 (20): 2111 – 20. [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Diminución da recompensa de nicotina na obesidade: comparación cruzada en humanos e rato. Psicofarmacoloxía (Berl) 2005
3. Franken IH, Muris P. As diferenzas individuais na sensibilidade de recompensa están relacionadas co desexo dos alimentos e o peso relativo das mulleres sans. Apetito. 2005; 45 (2): 198 – 201. [PubMed]
4. Kelley AE, Berridge KC. A neurociencia das recompensas naturais: relevancia para as drogas adictivas. J Neurosci. 2002; 22 (9): 3306 – 11. [PubMed]
5. Cagniard B, PD Balsam, Brunner D, Zhuang X. Os ratos con dopamina crónica elevada mostran unha motivación mellorada, pero non aprendendo, para unha recompensa de alimentos. Neuropsicofarmacoloxía. 2006; 31 (7): 1362 – 70. [PubMed]
6. Liang NC, Hajnal A, Norgren R. Sham alimentando aceite de millo aumenta a dopamina accumbens na rata. Am J Physiol Regul Integr Comp Physiol. 2006; 291 (5): R1236 – 9. [PubMed]
7. Mendoza J, Angeles-Castellanos M, Escobar C. O arrastramento por unha comida apetecible induce a actividade anticipatoria de alimentos e a expresión c-Fos en áreas do cerebro relacionadas coa recompensa. Neurociencia. 2005; 133 (1): 293 – 303. [PubMed]
8. Schroeder BE, Binzak JM, Kelley AE. Un perfil común de activación cortical prefrontal tras a exposición a sinais contextuais asociadas á nicotina ou ao chocolate. Neurociencia. 2001; 105 (3): 535 – 45. [PubMed]
9. Nestler EJ, Barrot M, Self DW. DeltaFosB: un interruptor molecular sostido para a dependencia. Proc Natl Acad Sci US A. 2001; 98 (20): 11042 – 6. [Artigo libre de PMC] [PubMed]
10. Teegarden SL, Bale TL. As diminucións da preferencia na dieta producen unha maior emocionalidade e un risco de recaída na dieta. Psicoloxía biolóxica. 2007; 61 (9): 1021 – 9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR. dFosB no Nucleus Accumbens regula o comportamento e a motivación instrumental reforzado por alimentos. The Journal of Neuroscience. 2006; 26 (36): 9196 – 9204. [PubMed]
12. Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, PE Shockett, MR Picciotto, Duman RS, EJ Nestler. Animais transxénicos con expresión xénica inducible e dirixida no cerebro. Mol Pharmacol. 1998; 54 (3): 495 – 503. [PubMed]
13. Kelz MB, Chen J, WA Carlezon, Jr, K Whisler, Gilden L, Beckmann AM, Steffen C, Zhang YJ, Marotti L, DW Self, Tkatch T, G Baranauskas, DJ Surmeier, Neve RL, Duman RS, Picciotto MR, Nestler EJ. A expresión do factor de transcrición deltaFosB no cerebro controla a sensibilidade á cocaína. Natureza. 1999; 401 (6750): 272 – 6. [PubMed]
14. Bale TL, Dorsa DM. Diferenzas sexuais e efectos do estrógeno na expresión do ácido ribonucleico do receptor de oxitocina no hipotálamo ventromedial. Endocrinoloxía. 1995; 136 (1): 27 – 32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. Os ratos dependentes do azucre mostran unha resposta mellorada para o azucre despois da abstinencia: evidencia dun efecto de privación de azucre. Physiol Behav. 2005; 84 (3): 359 – 62. [PubMed]
16. Will MJ, Franzblau EB, Kelley AE. Os mu-opioides de Nucleus accumbens regulan a inxestión dunha dieta rica en graxas mediante a activación dunha rede de cerebro distribuída. J Neurosci. 2003; 23 (7): 2882 – 8. [PubMed]
17. Zheng H, Patterson LM, Berthoud HR. A sinalización da orxina na área tegmental ventral é necesaria para un apetito alto en graxas inducido pola estimulación dos opiáceos do núcleo accumbens. J Neurosci. 2007; 27 (41): 11075 – 82. [PubMed]
18. Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, Gao XB, Thurmon JJ, Marinelli M, DiLeone RJ. A sinalización do receptor de leptina nas neuronas de dopamina do cerebro medio regula a alimentación. Neurona. 2006; 51 (6): 801 – 10. [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. A sobreexpresión específica do tipo de célula estriatal en DeltaFosB aumenta o incentivo para a cocaína. J Neurosci. 2003; 23 (6): 2488 – 93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. Un papel esencial para DeltaFosB no núcleo accumbens na acción da morfina. Nat Neurosci. 2006; 9 (2): 205 – 11. [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. Formación dendrítica da columna vertebral inducida pola cocaína en neuronas espiñas medias que conteñen receptores de dopamina D1 e D2 no núcleo accumbens. Proc Natl Acad Sci US A. 2006; 103 (9): 3399 – 404. [Artigo libre de PMC] [PubMed]
22. Blendy JA, Maldonado R. Análise xenética da adicción ás drogas: o papel da proteína de unión ao elemento de resposta do cAMP. J Mol Med. 1998; 76 (2): 104 – 10. [PubMed]
23. Nestler EJ. Mecanismos moleculares da adicción ás drogas. Neurofarmacoloxía. 2004; 47 1: 24 – 32. [PubMed]
24. Tanis KQ, Duman RS, Newton SS. Encadernación e actividade do cerebro no cerebro: especificidade rexional e inducción por crise electroconvulsiva. Psicoloxía biolóxica. 2007
25. Kumar A, Choi KH, Renthal W, Tsankova NM, Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ. A remodelación da cromatina é un mecanismo fundamental na plasticidade inducida por cocaína no estriado. Neurona. 2005; 48 (2): 303 – 14. [PubMed]
26. Graham DL, Edwards S, Bachtell RK, Dileone RJ, Rios M, Self DW. A actividade dinámica do BDNF no núcleo accumbens co uso de cocaína aumenta a autoadministración e a recaída. Nat Neurosci. 2007; 10 (8): 1029 – 37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 media as accións de varias drogas de abuso. Aaps J. 2005; 7 (2): E353 – 60. [Artigo libre de PMC] [PubMed]
28. Palmer AA, Verbitsky M, Suresh R, Kamens HM, Reed CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, Phillips TJ. As diferenzas de expresión xénica nos ratos son diverxentemente seleccionadas para a sensibilidade á metanfetamina. Mamm Genome. 2005; 16 (5): 291 – 305. [PubMed]
29. Bogush A, Pedrini S, Pelta-Heller J, Chan T, Yang Q, Mao Z, Sluzas E, Gieringer T, Ehrlich ME. AKT e CDK5 / p35 median a inducción do factor neurotrófico derivado do cerebro de DARPP-32 en neuronas espiñas de tamaño medio in vitro. J Biol Chem. 2007; 282 (10): 7352 – 9. [PubMed]
30. Benavides DR, Bibb JA. Papel do Cdk5 no abuso de drogas e na plasticidade. Ann NY Acad Sci. 2004; 1025: 335 – 44. [PubMed]
31. Os Bibb JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Os efectos da exposición crónica á cocaína son regulados pola proteína neuronal Cdk5. Natureza. 2001; 410 (6826): 376 – 80. [PubMed]
32. Blochl A, Sirrenberg C. As neurotrofinas estimulan a liberación de dopamina a partir de neuronas mesencefálicas de rata a través dos receptores Trk e p75Lntr. J Biol Chem. 1996; 271 (35): 21100 – 7. [PubMed]
33. Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, CA Bolanos, Ríos M, Monteggia LM, Self DW, Nestler EJ. O papel esencial do BDNF na vía de dopamina mesolímbica no estrés da derrota social. Ciencia. 2006; 311 (5762): 864 – 8. [PubMed]
34. Nestler EJ, Carlezon WA., Jr O circuíto de recompensa de dopamina mesolímbica na depresión. Psicoloxía biolóxica. 2006; 59 (12): 1151 – 9. [PubMed]
35. Ford CP, Beckstead MJ, Williams JT. Inhibición de opioides de Kappa de correntes postsinápticas inhibidoras de dopamina somatodendríticas. J Neurophysiol. 2007; 97 (1): 883 – 91. [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Sistemas de dinorfina cerebral e encefalina en ratas Fischer e Lewis: efectos da tolerancia á morfina e da retirada. Res. Cerebral. 1995; 683 (1): 25 – 35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Diferenzas entre ratas que prefiren o alcohol (AA) e que evitan o alcohol (ANA) nos sistemas de prodinorfina e proenefalina. Alcohol Clin Exp Res. 1994; 18 (5): 1272 – 9. [PubMed]
38. Kreek MJ. Cocaína, dopamina e sistema opioide endóxeno. J Addict Dis. 1996; 15 (4): 73 – 96. [PubMed]
39. Carlezon WA, Jr, Duman RS, Nestler EJ. As moitas caras do CREB. Tendencias Neurosci. 2005; 28 (8): 436 – 45. [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Taher M, Barczak A, Kameyama K, Huganir R, Konradi C. Os receptores D1 de Dopamina median a fosforilación CREB a través da fosforilación do receptor NMDA en Ser897-NR1. J Neurochem. 2003; 87 (4): 922 – 34. [PubMed]
41. Auto DW. Regulación dos comportamentos de toma de drogas e de busca por neuroadaptacións no sistema de dopamina mesolímbica. Neurofarmacoloxía. 2004; 47 1: 242 – 55. [PubMed]
42. Beitner-Johnson D, Nestler EJ. A morfina ea cocaína exercen accións crónicas comúns sobre a tirosina hidroxilase nas rexións dopaminérxicas de recompensa cerebral. J Neurochem. 1991; 57 (1): 344 – 7. [PubMed]
43. Lu L, Grimm JW, Shaham Y, Hope BT. Neuroadaptacións moleculares na área accumbens e tegmental ventral durante os primeiros días 90 de abstinencia forzada da auto-administración de cocaína en ratas. J Neurochem. 2003; 85 (6): 1604 – 13. [PubMed]
44. Shepard JD, Chuang DT, Shaham Y, Morales M. Efecto da auto-administración de metanfetamina en niveis de transportadores de tirosina hidroxilase e dopamina en vías de dopamina mesolímbica e nigrostriatal da rata. Psicofarmacoloxía (Berl) 2006; 185 (4): 505 – 13. [PubMed]
45. Fulton S, Pissios P, Manch RP, Stiles L, Frank L, Pothos EN, Maratos-Flier E, Flier JS. Regulación da leptina da vía de dopamina de mesoaccumbens. Neurona. 2006; 51 (6): 811 – 22. [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Implicación da proteína quinasa C na elevación inducida por orexina dos niveis de dopamina extracelular e o seu efecto gratificante. Eur J Neurosci. 2007; 25 (5): 1537 – 45. [PubMed]
47. Narita M, Nagumo Y, Hashimoto S, Khotib J, Miyatake M, Sakurai T, Yanagisawa M, Nakamachi T, Shioda S, Suzuki T. Implicación directa de sistemas orexinérxicos na activación da vía de dopamina mesolímbica e comportamentos relacionados inducidos pola morfina. J Neurosci. 2006; 26 (2): 398 – 405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, Fields HL, Bonci A. A Orexin A no VTA é fundamental para a inducción de plasticidade sináptica e sensibilización do comportamento á cocaína. Neurona. 2006; 49 (4): 589 – 601. [PubMed]
49. Park ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS. Cambios na expresión de orexina-A e neuropéptido Y no hipotálamo das ratas alimentadas con dieta alta e gorda. J Vet Sci. 2004; 5 (4): 295 – 302. [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF. Os péptidos que regulan a inxestión de alimentos: a expresión xénica da orexina aumenta durante os estados de hipertrigliceridemia. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1454 – 65. [PubMed]
51. Zheng H, Corkern M, Stoyanova I, Patterson LM, Tian R, Berthoud HR. Os péptidos que regulan a inxestión de alimentos: a manipulación de accumbens que indican o apetito activa as neuronas de orexina hipotalámica e inhibe as neuronas POMC. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1436 – 44. [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, Daniel RA, Landry CF, Kelley AE. A activación dunha subpopulação de neuronas hipotálamas que conteñen orexina / hipocretina polo inhibidor mediado polos receptores GABAA do núcleo accumbens shell, pero non pola exposición a un novo ambiente. Eur J Neurosci. 2004; 19 (2): 376 – 86. [PubMed]
53. Harris GC, Wimmer M, Aston-Jones G. Un papel para as neuronas hipotálamas laterales de orexina en busca de recompensas. Natureza. 2005; 437 (7058): 556 – 9. [PubMed]