Front Behav Neurosci. 2015 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Abstracto
No acondicionamento clásico, un estímulo inicialmente neutro (estímulo condicionado, CS) asóciase cun suceso bioloxicamente salientable (estímulo incondicionado, EUA), que pode ser dor (condicionamento aversivo) ou comida (condicionamento apetitivo). Despois dunhas poucas asociacións, o CS é capaz de iniciar respostas defensivas ou consumativas, respectivamente. Ao contrario do condicionamento aversivo, o condicionamento apetitivo raramente se investiga en humanos, aínda que a súa importancia para os comportamentos normais e patolóxicos (por exemplo, obesidade, adicción) é innegable. O presente estudo ten a intención de traducir os descubrimentos dos animais sobre o condicionamento apetitivo aos humanos que usan alimentos como Estados Unidos. Trinta e tres participantes foron investigados entre as 8 e as 10 horas sen almorzar para asegurar que sentían fame. Durante dúas fases de adquisición, unha forma xeométrica (avCS +) prediu un aversivo EE.UU. (descarga eléctrica dolorosa), outra forma (appCS +) predixo un apetitivo EUA (chocolate ou pretzel salgado segundo a preferencia dos participantes) e unha terceira forma (CS- ) non previu nin EE. Nunha fase de extinción, estas tres formas máis unha forma novedosa (NOVO) presentáronse de novo sen entrega aos Estados Unidos. As valoracións de valencia e excitación, así como as respostas de sobrecarga e condutancia da pel (SCR) foron recollidas como índices de aprendizaxe. Atopamos condicionamentos aversivos e apetitosos exitosos. Por unha banda, o avCS + clasificouse como máis negativo e máis excitante que o CS e induciu o potencial de sobresalto e o SCR mellorado. Por outra banda, a appCS + clasificouse como máis positiva que a CS e provocou unha atenuación de sobresalto e un SCR maior. En resumo, confirmamos con éxito os achados dos animais en humanos (famentos) demostrando unha aprendizaxe apetitiva e unha aprendizaxe aversiva normal.
introdución
A predición de ameazas e alimentos é de crucial importancia para a supervivencia de calquera organismo. No acondicionamento clásico (Pavlov, 1927), un sinal precede a un evento aversivo, como un lixeiro choque eléctrico doloroso (estímulo incondicional aversivo, EE. UU.), ou un evento apetito, como un pellet de comida (EUA apetitivo), varias veces. Posteriormente, este estímulo só (agora denominado CS, CS +) é capaz de provocar respostas defensivas ou consumidoras, respectivamente. O primeiro tipo de aprendizaxe asociativa chámase condicionamento aversivo, mentres que o segundo chámase acondicionamento apetito.
Aínda que a previsión de eventos apetitivos é tan importante para a supervivencia como a previsión de eventos aversivos, o apetito acondicionado é bastante menos investigado nos animais (Bouton e Peck, 1989; Koch et al., 1996; McDannald et al., 2011, para ver unha revisión Martin-Soelch et al., 2007) así como en humanosKlucken et al., 2009, 2013; Austin e Duka, 2010; Delgado et al., 2011; Levy and Glimcher, 2011). Esta falta de investigación podería deberse á complexidade do paradigma apetito en comparación coa aversiva. Por exemplo, os alimentos como reforzo primario deben entregarse cando o organismo ten fame para ser gratificante (para unha revisión recente vexa Dickinson e Balleine, 1994; Clark et al., 2012). Na investigación humana, esta dificultade superouse empregando o diñeiro (Austin e Duka, 2010; Delgado et al., 2011; Levy and Glimcher, 2011) ou imaxes eróticas (Klucken et al., 2009, 2013). Non obstante, os estudos de neurotoxía sinalaron que os reforzadores primarios (é dicir, bocadillos ou bebidas) e secundarios (é dicir, diñeiro) activan algunhas rexións cerebrais comúns (por exemplo, o estriado) pero tamén conteñen patróns distintos de activación (Delgado et al., 2011; Levy and Glimcher, 2011).
Polo que sabemos, só algúns estudos de condicionamento humano examinaron os efectos dos reforzadores apetitivos primarios como o cheiro (Gottfried et al., 2002), auga (Kumar et al., 2008), ou comida (Prévost et al., 2012). No primeiro estudo, Gottfried et al. (2002) caras neutras asociadas (CSs) cun olor desagradable, agradable ou neutro. Curiosamente, atoparon unha maior activación no córtex orbitofrontal (OFC) e no estriado ventral en resposta ao CS + apetitivo contra o CS + aversivo. Concluíron que o OFC procesa o valor do olor e está implicado na transferencia do valor afectivo do sistema olfativo (EU) ao visual (CS). A activación (ventro) estriatal interpretouse como que reflicte o aperitivo CR provocado polo aperitivo CS +. No segundo estudo, Kumar et al. (2008) Invitou aos seus participantes (controis sans e pacientes con depresión maior) ao laboratorio cedo na mañá e pediulles que se abstivesen de beber durante a noite para asegurarse de ter sede antes do escaneo. As imaxes fractal eran CSs e 0.1 ml de auga en Estados Unidos. Curiosamente, entre outras activacións, participantes sans (pero non pacientes de depresión) mostraron unha maior activación no estriado ventral ao apetito CS +, o que suxire que este estímulo foi procesado como gratificante. No terceiro estudo, Prévost et al. (2012) tamén presentaron imaxes fractais como CS e lanches doces ou salgados como Estados Unidos segundo a preferencia dos participantes. O CS presentouse durante 6 segundos e durante o último segundo presentouse unha imaxe de comida. Cada vez que aparecía a foto da comida, o experimentador colocaba un anaco de comida nas mans dos participantes, aos que se lles permitía consumir a merenda de inmediato. Desafortunadamente, os autores non informaron da activación cerebral ao CS + apetitivo durante a fase de acondicionamento clásico; con todo, observaron unha menor desaceleración cardíaca ao CS + recompensado en comparación co CS non recompensado, o que indica efectos condicionantes diferenciais.
É sorprendente a falta de estudos que empreguen a resposta de sobresaltos como índice de condicionamento apetitivo, especialmente tendo en conta o seu amplo uso en condicionamentos aversivos. A resposta de Startle é unha resposta defensiva ancestral e automática ante eventos aversivos repentinos, inesperados e fortes (Koch, 1999). Esta resposta defensiva está mediada por unha vía neuronal relativamente sinxela que implica as neuronas raíces cocleares, o núcleo pontino caudal da formación reticular (PnC) e os motoneuróns espinais (Fendt e Fanselow, 1999; Koch, 1999). Os estudos en animais revelaron que a potenciación sobresaliente depende das proxeccións desde a amígdala ata o PnC (Fendt e Fanselow, 1999; Koch, 1999), mentres que a atenuación do arranque depende dun núcleo intacto accumbens (NAcc, Koch, 1999). Tal modulación da resposta do sobresalto é unha medida implícita útil para a valencia dos estímulos presentados en primeiro plano. Así, a potenciación indica valencia negativa, mentres que a atenuación indica valencia positiva, tanto sen ser moi influenciada polos procesos cognitivos (Hamm e Weike, 2005; Andreatta et al., 2010). Segundo o noso coñecemento, só un estudo animal investigou o condicionamento do apetito medindo as respostas de sobresaliente como medida dependente. Curiosamente, as ratas examinadas demostraron unha atenuación no CS + (ou sexa, lixeira) asociada á entrega dunha solución de sacarosa. En particular, tal atenuación foi afectada en animais con lesións NAcc, pero non en aquelas con lesións de amígdala, o que suxire que o NAcc xoga un papel específico na obtención de CR apetitosos e na atenuación das respostas iniciais (Koch et al., 1996).
O estudo actual é o que sabemos o primeiro que traduce este paradigma de condicionamento apetitoso aos humanos empregando reforzadores primarios como EU, é dicir, doce (Smarties de chocolate)®) ou comida salada (pretzel salgado pequeno) e modulación de sobresaltos como medida de CR. Esperabamos que a appCS + desencadease CRs apetitivos fortes, como se reflicte na atenuación do inicio, SCR mejorada e clasificación de valencia positiva en comparación cos outros estímulos, é dicir, o avCS + e o CS-.
Materiais e Métodos
os participantes
Corenta e dous voluntarios aceptaron participar no estudo e recibiron créditos do curso. Nove participantes foron excluídos da análise, dous por problemas técnicos, tres porque foron codificados como non respondedores (amplitude media de sobresalto <5 μV) e catro porque non presentaron respostas de sobresalto suficientes por condición (mínimo = 2; para detalles, ver Materiais e método). Ao final, consideráronse 33 participantes na análise (16 homes; idade media: 22.09 anos, SD: 2.84; rango: 18-29 anos). Catro participantes non eran alemáns nativos e seis foron zurdos. Un participante non ignoraba as asociacións CSs-US durante todo o experimento (ver Procedemento), pero decidimos non excluír a este participante porque as súas respostas eran normais e non afectaron aos resultados.
Obras
Estímulos non condicionados (Estados Unidos)
Utilizáronse dous tipos de EE. Como Estados Unidos aversivo, aplicamos unha descarga eléctrica levemente dolorosa no antebrazo non dominante dos participantes. A descarga eléctrica saldouse mediante dous electrodos con diámetro 9 mm e espacio 30 cm. O estímulo eléctrico consistiu nun estímulo de pulso cunha frecuencia de 50 Hz e duración de 200 ms, xerado por un estimulador de corrente (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Reino Unido, 400 V, máximo de 9.99 mA). A intensidade da descarga eléctrica determinouse individualmente mediante un procedemento de limiar descrito anteriormente (Andreatta et al., 2010). Brevemente, os participantes sufriron dúas series de intensidades ascendentes e descendentes en intervalos de 0.5 mA. Tiveron que valorar cada estímulo nunha escala visual que vai desde 0 (sen sentir nada) ata 10 (dor realmente intensa) con 4 como áncora para o limiar (só dor perceptible). A intensidade media do estímulo eléctrico foi 2.12 mA (SD = 0.56) e considerouse como doloroso (M = 6.45, SD = 1.73). Os EEUU apetitosos consistían nun chocolate (Smarties)®) ou pretzel salgado pequeno. A elección dos apetitosos Estados Unidos dependía da preferencia individual do participante segundo se informou durante a entrevista preliminar. A saber, os participantes tiveron que informar se normalmente comen salgados ou doces durante o almorzo. Tamén poderían escoller libremente se preferían o chocolate ou o pretzel salgado durante o experimento. Ao final, 22 participantes escolleron o chocolate e 11 o pretzel salgado pequeno.
Estímulos condicionados (CS)
As formas xeométricas (cadrado azul, círculo amarelo, triángulo verde, hexágono vermello) cunha diagonal de 8 cm presentáronse como CS. As formas presentáronse no medio dunha pantalla de ordenador negra para s 8. Unha forma (avCS +) estivo sempre asociada co US aversivo (choque eléctrica dolorosa), unha forma (appCS +) estivo sempre asociada cos EEUU apetitosos (chocolate ou pretzel salado), unha forma (CS-) nunca se asociou con ningún dos dous aversivos. ou os EEUU apetitosos, e a cuarta forma (NOVA) presentouse en fase de extinción pero non durante a fase de adquisición para asegurar a súa neutralidade.
Sonda Startle
Como sonda de sobresalto empregouse un ruído branco de 103 dB cunha duración de 50 ms. Os estímulos acústicos presentáronse binauralmente a través dos auriculares e producíronse ao azar 4-6 s despois do inicio da forma.
Cuestionarios
Antes e despois do experimento, os participantes tiveron que cubrir as versións alemás do Inventario Estatal de trazos de ansiedade (STAI, Laux et al., 1981) e o calendario de afectos negativos positivos (PANAS, Krohne et al., 1996). O STAI é un inventario para avaliar o trazo e / ou a ansiedade do estado dos participantes e consta de 20 elementos tanto para o trazo como para as versións do estado. Nivel de ansiedade dos participantes antes (M = 37.06, SD = 7.80) e despois (M = 39.33, SD = 9.16) o experimento non cambiou significativamente [t(32) = 1.61, p = 0.117]. As puntuacións de ansiedade do rasgo na mostra actual oscilaron entre 20 e 58 (M = 36.6, SD = 8.98), que é comparable ao intervalo normal de adultos publicado (Laux et al., 1981). As PANAS (Krohne et al., 1996) é un índice de humor positivo e negativo. Os individuos con puntuacións altas na escala de efectos positivos (PAS) son propensos a emocións como o entusiasmo, mentres que os individuos con puntuacións altas na escala de efectos negativos (NAS) son propensos a emocións como a angustia. Cada elemento consta dun adxectivo e os participantes indican nunha escala que vai de 1 (moi lixeiramente) a 5 (extremadamente) ata que punto o adxectivo reflicte os seus sentimentos nese momento concreto. Non se atoparon diferenzas significativas no efecto negativo dos participantes como resultado do experimento [comezar: M = 11.67, SD = 2.29; final: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Dalgún xeito, os participantes cambiaron significativamente o seu estado de ánimo positivo dende o principio (M = 26.72, SD = 4.70) ata o final (M = 23.66, SD = 7.02) do experimento [t(31) = 3.11, p = 0.004]. Esta diminución do estado de ánimo positivo dos participantes puido estar relacionada co desagradable paradigma (presentáronse descargas eléctricas dolorosas e tamén un ruído branco aversivo).
Procedemento
Ao chegar ao laboratorio, os participantes leron e asinaron un formulario de consentimento informado aprobado polo comité de ética do Departamento de Psicoloxía da Universidade de Würzburg. Non se lles informou sobre a continxencia entre os CS e os Estados Unidos. Despois de cubrir os cuestionarios, conectáronse os electrodos e realizouse o procedemento do limiar da dor como se describiu anteriormente.
Durante o fase de habituación, as catro formas xeométricas presentáronse dúas veces cun intervalo de proba (ITI) variando entre 18 e 25 s (media: 21.5 s). Non se entregaron probas estadounidenses nin de inicio durante esta fase.
Antes da fase de adquisición, entregáronse sete momentos de ruído branco cada 7-15 s para diminuír a reactividade inicial.
Os dous seguintes fases de adquisición eran idénticos (figura 1). Cada fase de adquisición consistiu en ensaios 24: ensaios 8 CS, ensaios 8 avCS + e ensaios 8 appCS +. A secuencia CS era pseudorandoma coa única restrición de que o mesmo estímulo non se podía presentar máis de dúas veces seguidas. Destacable, o avCS + foi presentado xunto cun raio como un símbolo para a descarga eléctrica e o doloroso EUA foi entregado ao offset. A appCS + presentouse en composto cunha imaxe de Smarties ou unha pretzel salada e o participante puido escoller un Smarties ou un pretzel dun tarro. O CS- presentouse xunto cun símbolo de prohibición e non se entregaron ningún EUA. Durante tres das presentacións de 8 CS de cada tipo, entregouse unha sonda de arranque entre 4 e 6 s despois do inicio do estímulo. Durante a ITIs presentáronse tres sondas de arranque adicionais para asegurar a súa imprevisibilidade e reducir a habituación do arranque. O ITI, consistente nunha pantalla negra, variou entre 18 e 25 s cunha media de 21.5 s.
FIGURA 1
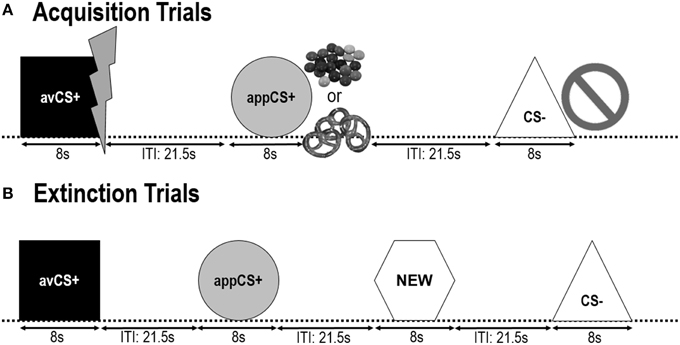
Figura 1. Ensaios durante as dúas fases de adquisición (A) e fase de extinción (B). Os participantes souberon que unha forma (avCS +) predicía unha choque eléctrica dolorosa leve, unha forma (appCS +) predicía unha peza de chocolate ou unha pretzel salada pequena (segundo a súa preferencia) e unha terceira forma (CS-) non predicía ningunha. evento bioloxicamente salientable. Cada forma presentábase en conxunto cunha imaxe que describe unha descarga eléctrica, smarties / pretzel salado ou nada dependendo da asociación con EE. Durante a fase de extinción, presentáronse de novo as tres formas xeométricas, pero non se entregaron os EE. Ademais, presentouse unha cuarta forma xeométrica (NOVA) como un control neutral.
Durante o fase de extinción, os participantes viron de novo as tres formas xeométricas (isto é, o avCS +, o appCS +, eo CS–) máis unha nova forma neutra (NOVO). Non se entregaron Estados Unidos e as formas non se presentaron en conxunto con imaxes de flashes, pretzeles salgados de chocolate ou prohibicións. Cada estímulo presentouse oito veces nun orde pseudoaleatorio (é dicir, o mesmo estímulo non se presentou máis de dúas veces seguidas), creando probas 32. Durante a presentación de estímulos 4 de cada tipo de CS presentáronse estímulos de sonda sobresalientes durante o 8. Ao igual que na fase de adquisición e de habituación, as IIT variaban entre 18 e 25 s, e as sondas de sobres adicional 4 entregáronse de xeito imprevisible durante as ITI.
Despois de cada fase, os participantes valoraron a valencia (agradable) e a excitación (excitatoria) dos CS con escalas analóxicas visuais (VAS) que van desde 1 a 9. A escala de valance varía de "1" indicando "moi desagradable" a "9" indicando "moi agradable" a escala de excitación variou de "1" indicando "calma" a "9" indicando "emocionante". avaliados despois das dúas fases de adquisición e da fase de extinción. Os participantes viron unha forma xeométrica para 1 s e despois tiñan que indicar se esta forma estaba asociada ao choque eléctrico, co chocolate / o salgado de pretzel, sen nada, ou se non eran capaces de facer ningunha asociación. En particular, todos os participantes (excepto un) tiveron coñecemento de continxencia despois da adquisición 2.
Redución de datos
As respostas fisiolóxicas rexistráronse cun amplificador V-Amp 16 e un software V-Amp Edition de Vision Recorder (versión 1.03.0004, BrainProducts Inc., Munich, Alemaña). Aplicáronse unha taxa de mostraxe de 1000 Hz e un filtro de entalhe 50 Hz. As análises fóra de liña realizáronse con Brain Vision Analyzer (versión 2.0; BrainProducts Inc., Múnic, Alemaña).
Resposta sobresaliente
A resposta de sobremia foi medida por electromiografía (EMG) á esquerda orbicularis oculi músculo con dous electrodos 5 mm Ag / AgCl. De acordo coas directrices (Blumenthal et al., 2005), colocouse un electrodo baixo a pupila eo segundo 1 cm lateralmente. Os electrodos de terra e de referencia colocáronse nos mastoides dereita e esquerda, respectivamente. Antes de conectar os electrodos, a pel foi rasgada e limpada con alcohol para manter a impedancia por baixo de 10 kΩ. A sinal electromiográfica foi filtrada sen conexión cun filtro de corte baixo 28 Hz e cun filtro de corte alto 500 Hz. Entón, o sinal EMG foi rectificado e unha media móbil de 50 ms foi aplicada. Usamos 50 ms antes da aparición da sonda de inicio como liña de base (Grillon et al., 2006). As respostas ás sondas sobresalientes foron puntuadas manualmente e os ensaios con cambios de liña de base excesivos (± 5 μV) ou artefactos de movemento foron excluídos da análise posterior. As respostas de sobresaltos inferiores a 5 μV foron codificadas como cero e consideradas para o cálculo da magnitude de sobrecarga (Blumenthal et al., 2005). En total, 10.4% dos ensaios foron rexeitados, e un mínimo de 2 fóra de 3 respostas de inicio en fases de adquisición e 4 fóra de 8 respostas de inicio en fase de extinción para cada condición foron obrigados a manter o participante no grupo de análise. Por esta razón, excluíronse catro participantes. A amplitude máxima definíase como o pico máximo relativo á liña de base durante a xanela de tempo 20-120 ms despois do inicio da sonda de sobresaltado. Os datos en bruto normalizáronse dentro dos suxeitos usando z- Puntuacións e logo puntuacións T para reducir a influencia da variabilidade individual e detectar mellor os procesos psicolóxicos. A puntuación T fixouse en promedio para cada condición (avCS +, appCS +, CS–, NEW e ITI). Para investigar a potenciación de sobresaltos ou a atenuación de sobresaltos, as puntuacións para as respostas de sobresalto ITI foron restadas das respostas de sobrecarga de cada condición.
Resposta á conduta da pel (SCR)
A resposta da condutancia da pel (SCR) rexistrouse usando dous electrodos 5 mm Ag / AgCl colocados na palma da man non dominante. A resposta galvánica foi filtrada sen conexión cun filtro de corte QNUMX Hz alto. O SCR definíase como a diferenza (en μS) entre o inicio da resposta (1-1 s despois do inicio do estímulo) eo pico de resposta (Tranel e Damasio, 1994; Delgado et al., 2011). As análises do SCR non consideraron as probas que conteñan sondas de sobresalto. As respostas por baixo de 0.02 μS foron codificadas como cero. Cinco participantes foron excluídos da análise de SCR porque tiñan un SCR medio menor que 0.02 μS. Os datos de condutividade da pel en bruto foron de raíz cadrada transformados para normalizar a distribución e as puntuacións foron promediadas para cada condición por separado para as dúas fases de adquisición (avCS +, appCS +, CS–) e a fase de extinción (avCS +, appCS +, CS– e NEW ).
Análise Estatística
Todos os datos analizáronse con SPSS para Windows (versión 20.0, SPSS Inc.). Para as respostas fisiolóxicas, calculáronse análises multivariantes de varianza (ANOVA) para as dúas fases de adquisición e a fase de extinción. O ANOVA para as fases de adquisición tiña estímulos (avCS +, appCS +, CS–) e fase (Adquisición 1, Adquisición 2) como factores dentro dos suxeitos. O ANOVA para a fase de extinción só tiña estímulos (avCS +, appCS +, CS–, NEW) como factor dentro dos suxeitos. Analizáronse os índices de valencia, excitación e continxencia con ANOVA separados que conteñen os estímulos dos factores dentro dos suxeitos (avCS +, appCS +, CS– e NEW) e fase. Este factor tiña catro niveis de avaliación de valencia e excitación (T1: despois da fase de habituación, T2: despois da primeira fase de adquisición, T3: despois da segunda fase de adquisición, T4: despois da fase de extinción), pero tres niveis para os índices de continxencia (T1: despois primeira fase de adquisición, T2: despois da segunda fase de adquisición, T3: despois da fase de extinción).
O nivel alfa (α) foi establecido en 0.05 para todas as análises. O tamaño do efecto divídese como parcial η2.
Resultados
A valoración de valencia e excitación de cada fase móstrase na figura 2; as respostas de sobresaltos e os SCR móstranse na figura 3.
FIGURA 2
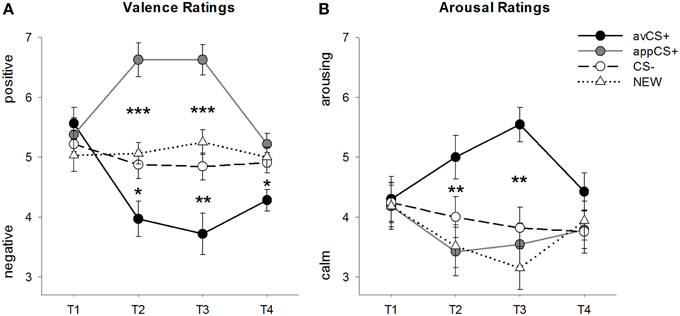
Figura 2. Valoración de valencia (A) e excitación (B). As liñas (con erros estándar) amosan as cualificacións tras a fase de habituación (T1), a adquisición 1 (T2), a adquisición 2 (T3) ea fase de extinción (T4). A aversiva CS + (liña sólida negra) adquiriu valencia negativa e alta excitación logo das dúas fases de adquisición en comparación co CS– (liña discontinua negra) e NOVA (liña punteada negra). É importante destacar que o apetito CS + (liña gris sólida) adquiriu valencia positiva en comparación co CS– e o NOVO. *p <0.05, **p > 0.01, ***p <0.001.
FIGURA 3
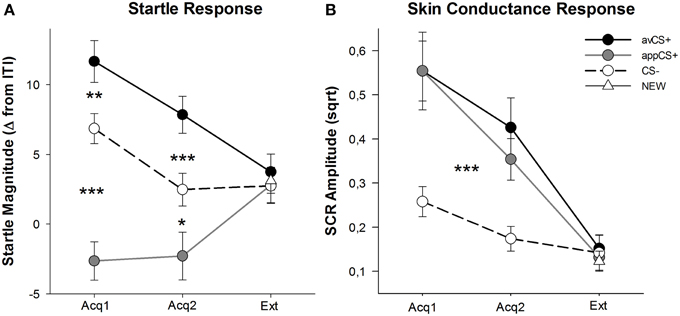
Figura 3. Respostas sobresalientes (A) e condutancia da pel (B) (con erros estándar) durante a primeira fase de adquisición (Acq1), a segunda fase de adquisición (Acq2) ea fase de extinción (Ext). As respostas de sobresaltos foron potenciadas significativamente co CS + aversivo (liña sólida negra) e significativamente atenuadas ao CS + (liña continua sólida) en comparación co CS– (liña discontinua negra) durante as fases de adquisición. O SCR foi significativamente maior ao avCS + e ao appCS + en comparación co CS–. Non se revelaron diferenzas durante a fase de extinción. *p <0.05, **p > 0.01, ***p <0.001.
Puntuacións
O ANOVA para o valoracións de valencia durante a adquisición revelaron efectos principais significativos do estímuloF(3, 93) = 17.26, GG-ε = 0.801, p <0.001, parcial η2 = 0.358] e fase [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, parcial η2 = 0.096], así como unha interacción significativa entre o estímulo ea fase.F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, parcial η2 = 0.235]. Seguimento t-as probas indican que as valencias das catro formas xeométricas no inicio do experimento eran idénticas (ps> 0.19), mentres que despois da adquisición 1 e da adquisición 2, avCS + clasificouse como especialmente negativa e a appCS + como especialmente positiva. En concreto, o avCS + clasificouse como máis negativamente valorado en comparación co CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], o NOVO [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] e a aplicación CS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. A appCS + clasificouse como significativamente máis positiva que a CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] e o NOVO [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. As diferenzas entre o CS e o NOVO nunca foron significativas (ps> 0.18).
É importante destacar que non se atoparon diferenzas significativas ao comparar os índices de valencia do appCS + asociado ao pretzel de chocolate versus salado [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Despois da fase de extinción, o avCS + seguía sendo máis negativo que o CS– [t(31) = 2.40, p = 0.023], o NOVO [t(31) = 3.40, p = 0.002], e os appCS + [t(31) = 3.35, p = 0.002]. Pola contra, a valencia do appCS + non se diferenciaba máis do CS– [t(31) = 1.77, p = 0.086] ou o NOVO [t(31) = 1.07, p = 0.293].
O ANOVA para o avaliacións de excitación durante a adquisición revelou un efecto principal significativo do estímuloF(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, parcial η2 = 0.181], pero non de fase [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, parcial η2 = 0.038], e unha interacción significativa entre o estímulo ea fase.F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, parcial η2 = 0.124]. Seguimento tas probas non indicaron diferenzas significativas entre os estímulos en relación á súa excitación inicialps> 0.74). Non obstante, despois das dúas fases de adquisición, o avCS + foi máis estimulante que o NOVO [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] e a aplicación CS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001], e marxinalmente máis excitante que o CS– despois da primeira fase de adquisición [t(32) = 1.96, p = 0.058], pero moito máis espertar logo da segunda fase de adquisición [t(32) = 3.65, p = 0.001]. A diferenza das clasificacións de valencia, a excitación do appCS + non difire da excitación do CS– e do NOVO (ps > 0.13) despois das fases de adquisición.
Do mesmo xeito que nas clasificacións de valencia, CS– e NEW non se diferencian na súa excitación.ps> 0.07).
Como foi o caso da valencia, non se revelaron diferenzas para o appCS + asociado co chocolate eo appCS + asociado co pretzel salado [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Despois da fase de extinción, non se revelaron diferenzas significativas nas clasificacións de excitación.ps> 0.08).
Para adicional t post-hoc-testes que comparan as clasificacións despois da fase de habituación, a primeira e segunda fases de adquisición e a fase de extinción, vexa Materiais suplementarios.
Resposta sobresaliente
O ANOVA para as fases de adquisición devolveu un efecto principal do estímulo [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, parcial η2 = 0.609], pero non fase [F(1, 32) = 3.16, p = 0.085, parcial η2 = 0.090], e unha interacción significativa Fase ×F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, parcial η2 = 0.095]. Seguimento t-as probas revelaron unha potenciación significativa da sobrecarga do avCS + en comparación co CS– durante o primeirot(32) = 3.27, p = 0.003] eo segundo [t(32) = 4.00, p <0.001] fases de adquisición. As respostas sobresalientes ao avCS + tamén se potenciaron significativamente en comparación coas do appCS +, de novo despois de ambas a adquisición 1 [t(32) = 8.20, p <0.001] e adquisición 2 [t(32) = 5.74, p <0.001]. É importante destacar que a magnitude de sobresalto da appCS + atenuouse significativamente en comparación coa CS– durante a adquisición 1 [t(32) = 6.34, p <0.001] e adquisición 2 [t(32) = 2.91, p = 0.007]. De novo e en consonancia coas clasificacións, non se revelaron respostas de sobresa diferencial para o appCS + cando se asocia co chocolate ou co pretzel salado [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Durante a fase de extinción, non se atopou ningún efecto significativo.F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, parcial η2 = 0.008].
Resposta á conduta da pel (SCR)
Dos ANOVAs para o SCR durante as dúas fases de adquisición, o estímulo de efectos principais [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, parcial η2 = 0.401] e fase [F(1, 27) = 20.91, p <0.001, parcial η2 = 0.436] resultou ser significativo, pero non a súa interacción [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, parcial η2 = 0.024]. T post-hoc-as probas indicaron un SCR significativamente maior ao avCS + [t(27) = 6.46, p <0.001] e á aplicación CS + [t(27) = 4.84, p <0.001] en comparación co CS–, mentres que os participantes mostraron SCR comparable ao avCS + e á appCS + [t(27) = 0.64, p = 0.527]. En particular, non se atoparon diferenzas nos SCR para os appCS + de chocolate e para os appCS + de pretzel salado [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Como era certo para as clasificacións e a resposta de sobres, non se atoparon efectos significativos para a fase de extinción [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, parcial η2 = 0.010].
Conversa
O obxectivo deste estudo foi traducir os descubrimentos dos animais aos humanos utilizando un paradigma de acondicionamento apetitivo clásico cun reforzo primario como estímulo incondicional (EEUU). Para iso, os participantes chegaron ao laboratorio cedo pola mañá sen almorzar para asegurarse de que tiñan fame, e segundo a súa preferencia, os anacos de chocolate ou salero de pretzels eran usados como apetitosos dos Estados Unidos. Durante a fase de adquisición, unha forma xeométrica (avCS +) asociouse cun choque eléctrico levemente doloroso (aversivo dos Estados Unidos), outra forma (appCS +) co apetito dos EUA e unha terceira forma (CS–) sen o EUA apetitivo nin co EUA aversivo. Os resultados indican un condicionamento aversivo e apetitivo exitoso no nivel verbal explícito (é dicir, valoracións), sobre o nivel de comportamento implícito (é dicir, a resposta de sobresalto) e ao nivel fisiolóxico (é dicir, SCR). Especificamente, o avCS + en comparación co CS- xerou clasificacións de valencia máis negativas, maiores puntuacións de excitación, potenciación de sobrecarga e maior SCR. Máis importante, o appCS + en comparación co CS - desencadeou valoracións de valencia máis positivas, atenuación de sobresaltos e maior SCR. Os nosos descubrimentos sobre o condicionamento aversivo estaban en consonancia coas expectativas, xa que estudos anteriores descubriron que unha estimación de prevención da ameaza (avCS +) é aversiva, provoca maiores respostas ao medo e aumenta a excitación fisiolóxica (Fendt e Fanselow, 1999; Hamm e Weike, 2005; Andreatta et al., 2010, 2013). Os nosos resultados no condicionamento apetito tamén estiveron en liña cos estudos anteriores en humanos e animais que amosaban que unha estimación de recompensa (appCS +) é valorada como positiva, inhibe as respostas ao medo e aumenta a excitación fisiolóxica (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin e Duka, 2010; Prévost et al., 2012). Segundo nós, este é o primeiro estudo que demostra unha atenuación de sobresaltos condicionada en humanos a un estímulo que predice unha recompensa primaria. É importante destacar que fomos capaces de transferir e confirmar os resultados dun estudo con animais (Koch et al., 1996). Este estudo en animais demostrou que a atenuación de sobrecarga nos ratos depende das proxeccións do NAcc (parte do estriado ventral) ao PnC. Polo tanto, a atenuación de sobresaltos do noso estudo podería implicar a actividade de NAcc, que tamén está en consonancia cos descubrimentos de fMRI (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy and Glimcher, 2011). De aí, concluímos que o noso paradigma de acondicionamento apetitivo foi exitoso, xa que tanto a valencia positiva explícita (avaliación) como a validez implícita (sobrescritura) indicáronse.
Ademais deste novo resultado, pero bastante predecible, merecen mención outros dous resultados interesantes. As primeiras respostas de excitación verbal e fisiolóxica ao appCS + disociáronse. En segundo lugar, atopamos a extinción máis rápida dos CR apetitivos en comparación cos CR aversivos.
O SCR indica a activación simpática, que se incrementa tanto no estímulo asociado á recompensa (appCS +) como no estímulo asociado á ameaza (avCS +). Este resultado está en liña cun estudo de condicionamento anterior, no que as imaxes eróticas foron usadas como EUA apetitiva (Klucken et al., 2013), e un estudo que revela un SCR comparable a un sinal de predicción de diñeiro e a unha suxestión prevendo un ruído aversivoAustin e Duka, 2010). En particular, o SCR é unha resposta orientadora relacionada coa activación do sistema simpático. Esta resposta suxeriuse para reflectir a preparación dunha reacción de comportamento a eventos salientais de motivación (Bradley, 2009). En base a isto, pensamos que tanto os estímulos asociados á ameaza como os asociados á recompensa provocaron unha resposta preparatoria para respostas comportamentais sucesivas. Noutras palabras, os sinais de comida e dor iniciaron a preparación para o comportamento de aproximación e evitación, respectivamente. En contraste coa alta excitación fisiolóxica, as respostas verbais indicaron unha baixa excitación no estímulo asociado á recompensa. É posible que a excitación verbal poida estar máis influenciada pola natureza esperta dos Estados Unidos que pola activación fisiolóxica per se. De feito, pensamos que o appCS + vs avCS + foi clasificado como pobre en contra e moi espertador porque estaban vinculados a Estados Unidos altamente e altamente excitantes, respectivamente. Desafortunadamente, non recompilamos os índices de excitación para os Estados Unidos e, polo tanto, dependerá de estudos futuros probar esta hipótese de xeito explícito.
Durante a fase de extinción, non se entregaron Estados Unidos. Isto pode inducir unha nova aprendizaxe inhibitoria chamada extinción (para ver unha revisión) Milad e Quirk, 2012), tendo en consecuencia tanto os CR aversivos coma os apetitosos. Evidentemente, nos niveis de comportamento (é dicir, a resposta sobrecogedora) e fisiolóxica (é dicir, SCR), xa non se detectaron reaccións diferenciais ao avCS +, ao appCS + e ao CS–. Do mesmo xeito, a excitación explícita dos estímulos condicionados evaluada despois da fase de extinción igualouse a un nivel baixo, o que suxire unha aprendizaxe de extinción exitosa. Non obstante, o avCS + seguía sendo considerablemente máis negativo que o CS–, mentres que a valencia appCS + xa non difería da valencia CS–. A máis lenta extinción da resposta explícita aversiva pode deberse a un conservadorismo evolutivo, o que significa que os sinais de ameaza son especialmente difíciles de esquecer porque a falta de resposta a un sinal de ameaza podería ser unha ameaza á vida.
Aínda queda unha pregunta: por que entón a resposta de sobresalto (é dicir, a valencia implícita) extinguíase completamente durante a fase de extinción? En primeiro lugar, estas respostas foron calculadas durante todo o curso da fase. Polo tanto, é concebible que os CR discriminativos aínda sexan detectables durante os primeiros ensaios da fase de extinción. De xeito exploratorio, seguimos esta hipótese e analizamos as respostas de sobresalto e SCR durante a fase de extinción (ver Material suplementario). Aínda que non atopamos diferenzas significativas, observamos unha magnitude de sobrecarga lixeiramente superior para o avCS + en comparación co CS– e o novo estímulo de control. Tamén observamos un pouco máis de atenuación de sobresalto ao appCS + en comparación co CS– e o novo estímulo de control ao comezo da fase de extinción, que, con todo, desapareceu nuns poucos ensaios. O SCR ao appCS + caeu xa despois do ensaio de extinción 2nd, mentres que o SCR ao avCS + permaneceu máis alto para case toda a extinción en comparación co SCR co NEW. Aínda que debemos interpretar estes resultados con moita cautela, as respostas de sobresalto parecían paralelas ás valoracións de valencia. Ademais, ambas respostas sobresalientes e SCR apoian aínda máis a idea dun conservadurismo evolutivo cara aos estímulos da ameaza.
Finalmente, temos que recoñecer algunhas limitacións deste estudo. En primeiro lugar, debido a un problema técnico, non se puideron informar as calificacións de agradable (e excitación) para o chocolate eo pretzel salado. Non obstante, os CR apetitivos suxiren que os participantes experimentaron realmente os dous Estados Unidos como apetitivos. En segundo lugar, a duración da aversión e do apetitivo dos Estados Unidos difería moito. Así, o choque eléctrico doloroso foi entregado ao compensar o avCS + e durou exactamente 200 ms, mentres que o chocolate eo pretzel salgado presentáronse aos participantes preto de 2 s despois do inicio do appCS +, ea duración era indefinible porque dependía da rapidez con o individuo comeu. A elección para entregar o apetito dos EUA deste xeito baseouse nun estudo previo en humanos (Prévost et al., 2012). Non obstante, sería metodolóxicamente máis elegante entregar un apetito estadounidense máis comparable ao aversivo dos Estados Unidos, como un sorbete de zume ou xenxibre na compensación appCS +. Para compensar esta gran diferenza perceptiva, presentamos formas xeométricas xunto cun estímulo visual que simboliza os Estados Unidos. En terceiro lugar, non podemos excluír definitivamente a posibilidade de que as respostas fisiolóxicas rapidamente extinguidas na fase de extinción sexan debidas a un aspecto metodolóxico. De feito, os estímulos visuais nesta fase non se presentaron en conxunto co símbolo dos Estados Unidos como foi o caso nas fases de adquisición. É posible que só a presentación dos CSs influíse nos CR e, por iso, non se viu ningunha diferenza significativa durante os primeiros ensaios de extinción. Non obstante, as nosas respostas rápidas extinguidas están en consonancia coas respostas extinguidas nun estudo anterior no que o CS + (cara) presentouse en composto con EEUU (grito) durante a adquisición pero non durante a extinción (Lissek et al., 2008).
En conclusión, atopamos respostas condicionadas e aversivas exitosas a un estímulo asociado á ameaza e a un estímulo asociado á recompensa, respectivamente. Curiosamente, o nivel de respostas explícito (avaliacións) e implícito (reflexo sobresaliente) funcionaba de xeito sinérgico, xa que o avCS + foi denunciado como negativo e inducido por potenciación de sobrecarga e o apCS + reportouse como atenuación de sobresalto positiva e inducida. Ademais, a excitación explícita (valoración) e fisiolóxica (SCR) do appCS + disociouse, reflectindo dous procesos distintos.
Declaración de conflitos de intereses
Os autores declaran que a investigación foi realizada en ausencia de relacións comerciais ou financeiras que puidesen interpretarse como un potencial conflito de intereses.
Grazas
Este traballo foi apoiado polo Centro de Investigación Colaborativa “Trastornos de medo, ansiedade e ansiedade”, SFB-TRR 58, proxecto B1.
Material complementario
O material suplementario deste artigo pódese atopar en liña en: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
References
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. e Pauli, P. (2013). A previsibilidade da dor inverte as valoracións de valencia dun estímulo asociado ao relevo. Diante. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B. e Pauli, P. (2010). Unha brecha entre a valencia condicionada implícita e explícita tras a aprendizaxe do alivio da dor en humanos. Proc. Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Austin, AJ e Duka, T. (2010). Mecanismos de atención para resultados apetitosos e aversivos no condicionamento Pavloviano. Behav. Res. Cerebral. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV e van Boxtel, A. (2005). Informe do comité: directrices para estudos electromiográficos de luces oculares. Psicofisioloxía 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Bouton, ME e Peck, CA (1989). Efectos do contexto no condicionamento, extinción e reintegración nunha preparación de acondicionamento apetitiva. Anim. Aprende. Behav. 17, 188 – 198. doi: 10.3758 / BF03207634
Texto completo de CrossRef | Google Scholar
Bradley, MM (2009). Atención selectiva natural: orientación e emoción. Psicofisioloxía 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Clark, JJ, Hollon, NG e Phillips, PEM (2012). Sistemas de valoración pavloviana na aprendizaxe e na toma de decisións. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Delgado, MR, Jou, RL e Phelps, EA (2011). Sistemas neuronais subxacentes ao condicionamento aversivo en humanos con reforzos primarios e secundarios. Diante. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Dickinson, A. e Balleine, B. (1994). Control motivacional da acción dirixida por obxectivos. Anim. Aprende. Behav. 22, 1 – 18. doi: 10.3758 / BF03199951
Texto completo de CrossRef | Google Scholar
Fendt, M. e Fanselow, MS (1999). A base neuroanatómica e neuroquímica do medo condicionado. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Gottfried, JA, O'Doherty, J. e Dolan, RJ (2002). A aprendizaxe olfativa apetitiva e aversiva en humanos estudou mediante resonancia magnética funcional relacionada con eventos. J. Neurosci. 22, 10829 – 10837. Dispoñible en liña en: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. e Johnson, L. (2006). Condicionamento do contexto e evitación do comportamento nun ambiente de realidade virtual: efecto da previsibilidade. Biol. Psiquiatría 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Hamm, AO e Weike, AI (2005). A neuropsicología da aprendizaxe do medo e da regulación do medo. Int. J. Psychophysiol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Activacións neuronais da adquisición da excitación sexual condicionada: efectos do coñecemento de continxencia e do sexo. J. Sexo. Med. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). O polimorfismo 5-HTTLPR está asociado con respostas hemodinámicas alteradas durante o acondicionamento apetito. Hum. Mapa do cerebro. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Koch, M. (1999). A neurobioloxía do asombro. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Koch, M., Schmid, A. e Schnitzler, H.-U. (1996). A atenuación do pracer do asunto é interrompida por lesións do núcleo accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W. e Tausch, A. (1996). Untersuchungen mit einer deutschen version der "Programa de afectación positiva e negativa" (PANAS). Diagnostica 42, 139-156.
Kumar, P., Garçom, G., Ahearn, T., Milders, M., Reid, I. e Steele, JD (2008). Signos de aprendizaxe de recompensa por diferenzas temporais anormales na depresión maior. Cerebro 131, 2084 – 2093. doi: 10.1093 / brain / awn136
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P. e Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: Beltz Test.
Levy, DJ e Glimcher, PW (2011). Comparación de mazás e laranxas: utilizando unha representación de valor subxectivo específico de recompensa e recompensa no cerebro. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Condición elevada do medo a estímulos socialmente incondicionales relevantes no trastorno de ansiedade social. Am. J. Psiquiatría 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Martin-Soelch, C., Linthicum, J. e Ernst, M. (2007). Condicionamento apetitivo: bases neurais e implicacións para a psicopatoloxía. Neurosci. Biobehav. Rev. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Resumo | Texto completo de CrossRef | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. e Schoenbaum, G. (2011). O estriado ventral e o córtex orbitofrontal son necesarios para a aprendizaxe de reforzo baseada no modelo, pero non modelo. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Milad, MR e Quirk, G. (2012). Extinción de medo como modelo de neurociencia translacional: dez anos de progreso. Annu. Rev. Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Pavlov, IP (1927). Reflexos condicionados: unha investigación da actividade fisiolóxica da cortiza cerebral. Londres: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM e O'Doherty, JP (2012). Correlados neuronais de transferencia pavloviana a instrumental específica e xeral dentro de subrexións amigdalares humanas: un estudo de resonancia magnética magnética. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Tranel, D. e Damasio, H. (1994). Correlados neuroanatómicos das respostas da condutancia da pel electrodérmica. Psicofisioloxía 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Resumo | Texto completo de CrossRef | Google Scholar
Palabras clave: condicionamento clásico, recompensa, castigo, resposta de sobresalto, resposta da condutancia da pel
Citación: Andreatta M e Pauli P (2015) Condicionamento apetitivo contra a aversión en humanos. Diante. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Recibido: 11 febreiro 2015; Aceptado: 05 Maio 2015;
Publicado: 19 Maio 2015.
Editado por:
Niels Birbaumer, Universidade de Tuebingen, Alemaña
Avaliado por:
Tim Klucken, Justus Liebig University Giessen, Alemania
Alessandro Angrilli, Universidade de Padua, Italia
Copyright © 2015 Andreatta e Pauli. Este é un artigo de acceso aberto distribuído baixo os termos da Licenza de atribución de Creative Commons (CC BY). Está permitido o uso, distribución ou reprodución noutros foros sempre que se acredite o autor ou licenciador orixinal e cítase a publicación orixinal nesta revista, de acordo coa práctica académica aceptada. Non se permite ningún uso, distribución ou reprodución que non cumpra estes termos.
* Correspondencia: Marta Andreatta, Departamento de Psicoloxía (Psicoloxía Biolóxica, Psicoloxía Clínica e Psicoterapia), Universidade de Würzburg, Marcusstraße 9-11, D-97070 Würzburg, Alemania, [protexido por correo electrónico]