Diante. Comportamento. Neurosci., 23 marzo 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Instituto de Investigación Traslacional e Instituto para a Saúde e Innovación Biomédica, Queensland University of Technology, Brisbane, QLD, Australia
- 2Escola de Ciencias Biomédicas, Universidade de Queensland, Brisbane, QLD, Australia
A dieta moderna endulzouse moito, obtendo niveis sen precedentes de consumo de azucre, particularmente entre os adolescentes. Aínda que se sabe que a inxestión crónica de azucre a longo prazo contribúe ao desenvolvemento de trastornos metabólicos, incluída a obesidade e a diabetes tipo II, pouco se sabe sobre as consecuencias directas do consumo de azucre a longo prazo e semellante ao azar no cerebro. BO azucre ecause pode provocar a liberación de dopamina no núcleo accumbens (NAc) de xeito similar aos fármacos de abuso, investigamos cambios na morfoloxía das neuronas desta rexión cerebral despois das curtas (4 semanas) e a longo prazo (12 semanas) binge- como o consumo de sacarosa usando un paradigma de elección intermitente de dúas botellas. Utilizamos a coloración de Golgi-Cox para impregnar neuronas de espiña media (MSNs) do núcleo NAc e cuncha de ratas consumidoras de sacarosa a curto e longo prazo e comparamos estas cos controis de auga correspondentes á idade. Demostramos que o consumo prolongado de sacarosa como binge diminuíu significativamente a lonxitude dendrítica total dos MSN shell de NAc en comparación cos ratos de control correspondentes á idade. Tamén descubrimos que a reestruturación destas neuronas resultou principalmente da complexidade dendrítica distal reducida. Pola contra, observamos un aumento da densidade de columna vertebral nas ordes distales de MSN shell de NAc de ratas que consumen sacarosa a longo prazo. Combinados, estes resultados poñen de relevo os efectos neuronais da inxestión prolongada de sacarosa tipo binge na morfoloxía MSN de cuncha de NAc.
introdución
Nos últimos anos 40, houbo un aumento documentado do consumo de bebidas edulcoradas con azucre e alimentos que conteñan azucres (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), con informes estimando que ata o 75% de todos os alimentos e bebidas conteñen altas cantidades de azucres engadidos (Ford e Dietz, 2013; Bray e Popkin, 2014). Durante este período, tamén houbo un aumento concorrente na prevalencia de obesidade e diabetes tipo II, especialmente nos adolescentes (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Estudos recentes demostraron que os nenos con exceso de peso e obesidade adoitan consumir altas cantidades de azucre engadido, non obstante a contribución de dietas con alto contido de azucre á aumento da incidencia de nenos con sobrepeso e obesos segue sendo controvertida (Hu, 2013; Bray e Popkin, 2014; Bucher Della Torre et al., 2015).
Aínda que un maior número de evidencias indica que o consumo de dietas ricas en azucre pode contribuír, en parte, ao aumento de peso entre nenos e adolescentes (Malik et al., 2010; Te Morenga et al., 2013; Bray e Popkin, 2014), prestouse menos atención ás consecuencias adversas non metabólicas derivadas da inxestión excesiva de azucre. Curiosamente, algúns patróns de comportamento e psicolóxicos comúns a miúdo xorden entre un grupo de persoas que consumen e manteñen dietas altas en azucres. O máis destacable é o desenvolvemento de trastornos alimentarios, incluído o comido por conxuntura, combinado co inicio simultáneo de síntomas psicolóxicos, incluída a falta de motivación e depresión (revisado en Sheehan e Herman, 2015). Ademais, debido a que os individuos que consumen desagradación adoitan presentar unha perda de control e unha incapacidade de autolimitarse a inxestión de azucre, é probable que estes comportamentos xurdan como consecuencia de adaptacións neurolóxicas en rexións cerebrais que avalían o valor hedonico dos alimentos altamente agradables. (Saper et al., 2002; Lutter e Nestler, 2009; Kenny, 2011). Esta fundamentación tamén se apoia en probas en humanos que demostran que o azucre e a dozura poden provocar ansias que son similares ás inducidas por drogas adictivas como o alcol e a nicotina (Volkow et al., 2012).
Aínda que as propiedades adictivas do azucre seguen sendo especulativas, estas observacións combináronse con estudos que demostran a contribución da inxestión excesiva de azucre aos cambios nos circuítos de recompensa e ao desenvolvemento de condutas e estados emocionais de tipo adictivo nos modelos animais. (Avena e col., 2008; Benton, 2010; Ventura et al., 2014), garante a necesidade de máis investigación. Estudos previos en roedores demostraron que o acceso intermitente á sacarosa altera a actividade de varios neurotransmisores dentro do sistema mesolimbico incluíndo dopamina, opioides e acetilcolina (revisado en Avena e col., 2008). Comprobouse que o consumo de sacarosa tipo binge facilita a liberación de dopamina no núcleo accumbens (NAc), de xeito similar ás drogas de abuso (Avena e col., 2008). Ademais, demostramos que o consumo a longo prazo de sacarosa empregando un paradigma de elección de dúas botellas de acceso intermitente 24 h (Simms et al., 2008) modula a expresión do receptor nicetínico da acetilcolina (nAChR) no NAc (Shariff et al., En prensa). Curiosamente, tamén observamos que os compostos de nAChR coñecidos por modular a actividade da dopamina e acetilcolina no NAc, teñen diferentes efectos sobre o consumo de sacarosa despois da inxestión a curto e a longo prazo (Shariff et al., En prensa).
Aínda que estes estudos demostraron semellanzas nos cambios de comportamento e neuroquímicos causados polo acceso intermitente ao azucre e as drogas de abuso, non se sabe se estes efectos facilitan cambios na morfoloxía neuronal no NAc. TO seu é en contraste con substancias de abuso incluíndo cocaína, anfetamina e nicotina que producen cambios ben caracterizados na morfoloxía de neuronas de espiña media (MSNs) na NAc, incluíndo o aumento da densidade da columna vertebral e a complexidade dendrítica alterada. (Robinson e Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Porque anteriormente demostramos que a exposición a longo prazo (semana 12) ao alcol e sacarosa usando o paradigma de elección intermitente de dúas botellas produce unha resposta diferencial ás intervencións farmacoterapéuticas en comparación coa inxestión a curto prazo (semanas 4; Steensland et al., 2007; Shariff et al., En prensa), evaluamos os efectos do consumo de sacarosa a curto e longo prazo sobre a morfoloxía de MSN no NAc. Permitimos que as ratas adolescentes consumisen sacarosa dun xeito seminario durante 4 (a curto prazo) ou 12 (a longo prazo) semanas e logo analizamos a morfoloxía de MSN NAc a partir de ratas consumidoras de sacarosa a curto e longo prazo e comparamos isto con controis adaptados á idade aos que só se lles daba acceso á auga. Os nosos resultados mostran que os MSNs do shell NAc están alterados despois do consumo de sacarosa a longo prazo, pero non a curto prazo, reducindo a lonxitude dendrítica, pero aumentaron a densidade dendrítica de columna vertebral. Ademais, atopamos a morfoloxía de MSN do núcleo NAc mantida relativamente intacta despois do consumo de sacarosa a curto e longo prazo. Estes resultados poñen de relevo unha consecuencia neurolóxica directa do consumo a longo prazo de sacarosa de xeito xenial. Ademais, estes datos demostran a necesidade de máis estudos encamiñados a dilucidar os cambios moleculares e neuroquímicos que acompañan a reestruturación morfolóxica dos MSN shell de NAc inducidos por unha inxestión prolongada de tipo sacarosa.
Materiais e Métodos
Declaración de ética
Todos os procedementos experimentais realizáronse de acordo co Código australiano para o coidado e uso de animais con fins científicos, 8th Edition (National Health and Medical Research Council, 2013). Os protocolos foron aprobados polo Comité de Ética Animal da Universidade de Queensland e o Comité de Ética Animal da Universidade de Queensland.
Animais e vivenda
As ratas Wistar masculinas de cinco semanas (adolescentes) (control: 176.4 ± 4.8 g; sacarosa: 178.3 ± 5.0 g) (ARC, WA, Australia), aloxáronse individualmente nun Plexiglas de dobre nivel ventilado.® gaiolas. As ratas aclimatáronse ás condicións individuais da vivenda, ao manexo e ao ciclo de luz inversa 5 días antes do inicio dos experimentos. Todas as ratas estaban aloxadas nun cuarto de luz invertida e ciclo escuro de 12-hr (luz apagada en 9 am) e con auga chow de rata estándar e auga dispoñible ad libitum.
Paradigma de bebida con elección de dúas botellas de acceso intermitente
O acceso intermitente 5% sacarosa elección de dúas botellas paradigma de bebida (Simms et al., 2008) foi adaptado a Sabio (1973). Todos os fluídos presentáronse en botellas de plástico graduado en 300 ml con botes de bebida de aceiro inoxidable inseridos a través de dúas pinzas na parte dianteira da gaiola despois do inicio do ciclo de luz escura. Os pes de cada botella rexistráronse antes da presentación da botella. Presentáronse simultaneamente dúas botellas: unha botella que contiña auga; a segunda botella que contén sacarosa 5% (p / v). A colocación da botella de sacarosa 5% (p / v) alternouse con cada exposición para controlar as preferencias laterais. Pesáronse as botellas de 24 h despois de que se presentaron os fluídos e as medidas se fixeron ao 0.1 g máis próximo. Tamén se mediu o peso de cada rata para calcular os gramos de inxestión de sacarosa por quilo de peso corporal. O día 1 do período de beber, ratas (n = 6-9) acceso a unha botella de sacarosa 5% (p / v) e a unha botella de auga. Despois de 24 h, a botella de sacarosa substituíuse por unha segunda botella de auga dispoñible para a próxima 24 h. Este patrón repetiuse o mércores e o venres. As ratas tiñan acceso ilimitado á auga os demais días. O consumo de sacarosa como o desembarco resultou nunha escalada na inxestión total de sacarosa (ml) ao longo do tempo (Figura complementaria 1) e estivo acompañado de niveis de bebida base base estables en función do peso corporal [20 ± 5 g / kg do 5% (p / v)] durante a curto prazo [~ 4 semanas (sesións de bebida 13)] e a longo prazo [ ~ 12 semanas (sesións de bebida 37)] períodos de bebida. Un grupo separado de ratas de control (n = 6-9) déronse acceso á auga en ambas botellas (é dicir, sen sacarosa) nas mesmas condicións descritas anteriormente. O peso corporal medio das ratas consumidoras de control e sacarosa ao final da exposición a curto prazo foi 405.7 ± 40.8 g e 426.4 ± 31.2 g respectivamente. Ao final da exposición a longo prazo, o peso corporal medio para os grupos control e sacarosa foi 578.8 ± 53.4 g e 600.2 ± 45.2 g.
Mancha de Golgi-Cox
Despois da última sesión de beber, os ratos foron trasladados da instalación animal para permitir o procesamento de mostras cerebrais na instalación de histoloxía da Escola de Ciencias Biomédicas da Universidade de Queensland (Santa Lucía, Australia). Tomáronse todas as medidas aprobadas para reducir o estrés durante o transporte, tras as que se permitiu que as ratas se recuperaran durante a noite. Ao día seguinte, as ratas sacrificáronse por sobredose pentobarbital sódica (60 – 80 mg / kg, ip Vetcare, Brisbane, Australia) e intracardialmente perfusionáronse con fluído cerebro-espinal artificial ~ 300 ml que contiña, (en mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2e 10 D-glicosa. A continuación, cada animal foi decapitado e o cerebro eliminado e incubado na escuridade na solución Golgi-Cox que contiña 5% dicromato de potasio, 5% cromato potásico e 5% cloruro mercurico (todos os produtos químicos de Sigma-Aldrich) que se fixo 3 días frescos. antes do sacrificio como se describiu anteriormente (Rutledge et al., 1969). Modificáronse os métodos de incubación de manchas de Golgi-Cox e post-procesamento Ranjan e Mallick (2010). Os cerebros de animais consumidores de sacarosa a curto prazo incubáronse durante 6 días a 37 ° C, mentres que os cerebros de animais consumidores de sacarosa a longo prazo incubáronse durante 10 días, con un cambio á solución Golgi-Cox fresca tras 4 días de incubación.
Despois da incubación, cortáronse seccións coronais de 300 μm usando un microtomo vibratorio Zeiss Hyrax V50 (Carl Zeiss, Alemaña). A continuación, as rodajas colocáronse secuencialmente en placas de pozo 24 cheas de sacarosa 30% (p / v) en solución salina tamponada con fosfato 0.1 M e procesáronse segundo o descrito en (Ranjan e Mallick, 2010). Brevemente, as seccións foron deshidratadas en 50% etanol durante 5 min, logo colocáronse en 0.1 M NH4Solución OH para 30 min, lavada dúas veces con auga destilada durante 5 min e colocada no fixador de películas de Fujihunt (Fujifilm, Singapur) para 30 min na escuridade. A continuación, as lavadas foron lavadas dúas veces en auga destilada durante 2 min cada unha e deshidratadas en 70, 90, 95 e 100% etanol dúas veces durante 5 min cada unha. As seccións foron despexadas en solución CXA (1: 1: 1 cloroformo: xileno: alcohol) para 10 min e montáronse en DPX (Sigma-Aldrich) en diapositivas Superfrost Plus (Menzel-Glaser, Lomb Scientific, Australia) e cubertas. (Menzel-Glaser, Alemaña). As diapositivas deixáronse á escuridade para secar a temperatura ambiente durante a noite.
Selección e rastrexo neuronal dentro do núcleo accumbens
Análise de franxas coronais entre bregma + 2.8 e + 1.7 para MSNs no núcleo e cuncha do NAc, empregando o ventrículo lateral e a comisión anterior como fitos coa axuda dun atlas cerebral de rata (Paxinos e Watson, 2007) (Figura 1). A función de contorno en Neurolucida 7 (MBF Bioscience, VT, USA) usouse para demarcar o núcleo de NAc e o shell de NAc en cada porción (Figura 2). Entre as neuronas 2 e 9 por rexión por animal rastrexáronse para parámetros de lonxitude dendrítica usando un obxectivo 63x ou para densidades de columna vertebral (indicadas como espinas por 100 μm) usando un obxectivo 100x nun Zeiss Axioskop II (Carl Zeiss, Alemaña) mediante un automatizado xyz escenario dirixido por Neurolucida® Software 7 (MBF Biosciences, VT, EUA). Todo o rastrexo realizouse de forma cegada con respecto ao tratamento. Os parámetros morfolóxicos das neuronas impregnadas de Golgi-Cox analizáronse dun xeito similar aos informes anteriores (Klenowski et al., 2015).
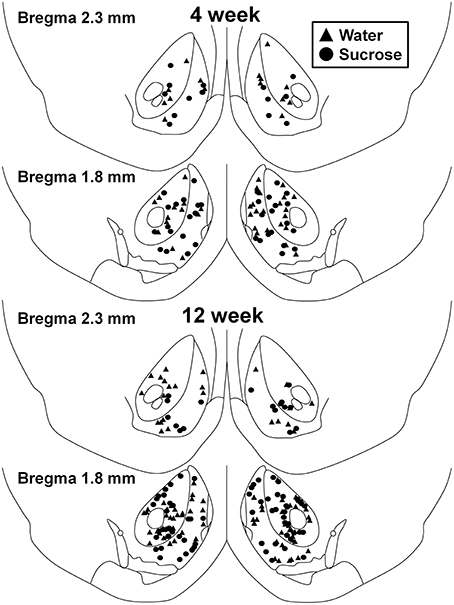
Figura 1. Mapa que mostra as localizacións de neuronas de espiña media mostradas do núcleo accumbens e a cuncha de ratas consumidoras de sacarosa 4 e 12 semana e controis correspondentes á idade. Os dous paneis principais mostran localización de neuronas mostradas do núcleo accumbens e núcleo de animais de control de semana 4 (triángulos) e sacarosa (círculos) animais. Os dous paneis inferiores mostran a posición das neuronas mostradas de animais de control de semana 12 (triángulos) e sacarosa (círculos).
Análise Estatística
Calculouse a media e o erro estándar da media (SEM) para cada conxunto de datos co animal como n, usando os datos de morfometría media de todos os MSNs NAc núcleo ou shell (n = 7 para o shell de NAc e n = 6 para o núcleo de NAc 4-semana, n = 9 para grupos de 12 semanas). Cando se indique, estudantes de dúas colas sen par trealizáronse probas ou ANOVA bidireccional con post-probas de Bonferroni para todas as análises que implican a comparación de medios de grupo, usando a versión 6.02 de GraphPad Prism (GraphPad Software, San Diego, CA). Aceptouse a importancia estatística en P <0.05. Todos os datos da sección de resultados preséntanse como medios ± SEM. Os cambios porcentuais calcúlanse en relación ao valor de control.
Resultados
As neuronas de espiña media do shell Nucleus Accumbens diminuíron a lonxitude dendrítica, diminuíron a complexidade dendrítica pero aumentaron a densidade media de columna vertebral nas ordes do ramo distal tras un consumo de sacarosa a longo prazo, pero non a curto prazo.
Despois do consumo de sacarosa a curto prazo (4 semanas), non houbo diferenzas significativas nos parámetros morfométricos de MSN do shell de NAc 1). Tampouco houbo diferenzas significativas entre o consumo de sacarosa a curto prazo e os controladores de auga de MSN de cuncha de NAc nas análises relacionadas coa orde de ramas centrífugas. É dicir, segmentos dendríticos por orde de sucursal (P = 0.4111), lonxitude dendrítica media por orde de sucursal (P = 0.5581) e a densidade media da columna vertebral por orde de sucursal (P = 0.2977, ANOVA bidireccionais) non foron significativamente diferentes entre grupos. Na Figura móstrase un mapa de localización que mostra as posicións aproximadas das neuronas mostradas 1.
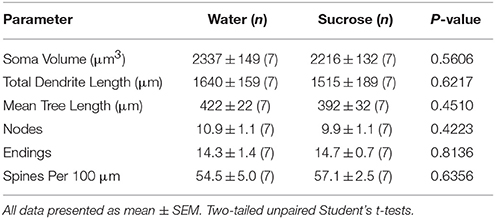
Táboa 1. Parámetros morfolóxicos xerais das neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a curto prazo e controis de auga correspondentes á idade.
Despois do consumo de sacarosa a longo prazo (12 semanas), a lonxitude total do árbo dendrítico dos MSN de cuncha de NAc diminuíu un 21% en comparación cos controis de consumo de auga (auga: 1827 ± 148 μm, n = 9; Sacarosa 1449 ± 78 μm, n = 9, *P = 0.0384, estudante sen par de dúas colas t-test, figura 2, Táboa 2). A comparación do número medio de bifurcacións dendríticas (nodos) e terminacións dendríticas entre os grupos de auga e sacarosa revelou un nivel reducido (aínda que non significativo) de complexidade dendrítica en MSNs cuncha de NAc (nodos: Auga 12.9 ± 1.4 n = 9, sacarosa 10.1 ± 0.8 n = 9, P = 0.0879; terminacións: Auga 17.9 ± 1.4 n = 9, sacarosa 14.8 ± 0.7 n = 9, P = 0.0657, estudante sen par de dúas colas t-test, Táboa 2). Non houbo cambio no volume de soma (P = 0.9400), lonxitude media da árbore dendrítica (P = 0.1646) ou densidade total de columna vertebral (P = 0.3662) en MSN shell de NAc procedentes de ratas que consumen sacarosa a longo prazo en comparación cos controis de auga. Estes parámetros morfométricos están detallados na táboa 2.
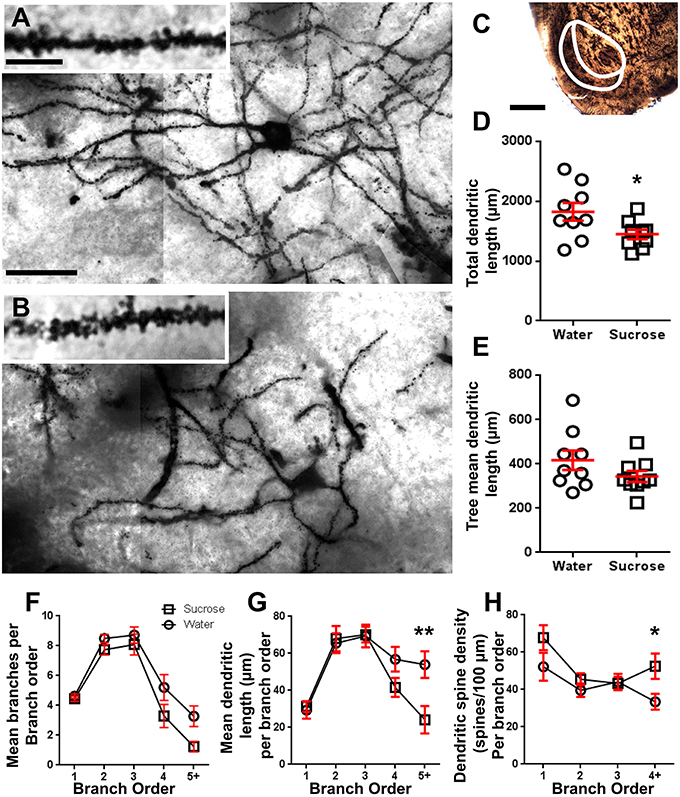 Figura 2. Diminución da lonxitude da árbore dendrítica e aumento da densidade da columna dendrítica distal das neuronas de espiña media (MSNs) do núcleo accumbens (NAc) de cuncha de ratas tratadas con sacarosa a longo prazo en comparación coas ratas control. (A, B) Amosar representacións de campo luminoso tratado con sacarosa (inferior) e control a longo prazo (12 semana) z-Mosaicos de MSN impregnados con Golgi do shell NAc (ampliación 63x). Inserto de (A, B) amosa imaxes de campo brillante tratadas con control e a longo prazo tratadas con sacarosa de dendritas de MSN impregnadas con Golgi e espinas dendríticas da cuncha de NAc (ampliación de 100x). (C) mostra as rexións anatómicas das que se probaron as MSN neste estudo. (D) mostra unha parcela de dispersión do arbor dendrítico total de MSN total (media ± SEM) do shell de NAc en animais de sacarosa a longo prazo (cadrados) en comparación con controles (círculos), estudantes sen parella t-test, *P <0.05, n = 9; control e n = 9; Xarcosidade semana 12. (E) mostra unha parcela de dispersión de lonxitude media de árbore dendrítica de MSN (media ± SEM) da cuncha de NAc en animais de sacarosa de longa duración (cadrados) en comparación con controles (círculos), estudantes sen parella t-test, P > 0.05, n = 9; control e n = 9; Xarcosidade semana 12. Análise de sucursais (media ± SEM) do número de segmento dendrítico por orde de sucursal (F), lonxitude media dendrítica por orde de sucursal (G) e densidade de columna dendrítica por orde de rama (H). O consumo de sacarosa a longo prazo diminuíu a lonxitude dendrítica en ordes de rama distal (5 +) e aumentou a densidade dendrítica de columna vertebral nas ordes de rama distal (4 +) en comparación cos controis (G, H), ANOVA bidireccional con post-probas de Bonferroni, *P <0.05, **P <0.01, n = 9; control e n = 9; sacarosa a longo prazo. Barras de escalas: (A, B) = 20 μm; inserción de (A, B) = 10 μm; (C) = 1 mm.
Figura 2. Diminución da lonxitude da árbore dendrítica e aumento da densidade da columna dendrítica distal das neuronas de espiña media (MSNs) do núcleo accumbens (NAc) de cuncha de ratas tratadas con sacarosa a longo prazo en comparación coas ratas control. (A, B) Amosar representacións de campo luminoso tratado con sacarosa (inferior) e control a longo prazo (12 semana) z-Mosaicos de MSN impregnados con Golgi do shell NAc (ampliación 63x). Inserto de (A, B) amosa imaxes de campo brillante tratadas con control e a longo prazo tratadas con sacarosa de dendritas de MSN impregnadas con Golgi e espinas dendríticas da cuncha de NAc (ampliación de 100x). (C) mostra as rexións anatómicas das que se probaron as MSN neste estudo. (D) mostra unha parcela de dispersión do arbor dendrítico total de MSN total (media ± SEM) do shell de NAc en animais de sacarosa a longo prazo (cadrados) en comparación con controles (círculos), estudantes sen parella t-test, *P <0.05, n = 9; control e n = 9; Xarcosidade semana 12. (E) mostra unha parcela de dispersión de lonxitude media de árbore dendrítica de MSN (media ± SEM) da cuncha de NAc en animais de sacarosa de longa duración (cadrados) en comparación con controles (círculos), estudantes sen parella t-test, P > 0.05, n = 9; control e n = 9; Xarcosidade semana 12. Análise de sucursais (media ± SEM) do número de segmento dendrítico por orde de sucursal (F), lonxitude media dendrítica por orde de sucursal (G) e densidade de columna dendrítica por orde de rama (H). O consumo de sacarosa a longo prazo diminuíu a lonxitude dendrítica en ordes de rama distal (5 +) e aumentou a densidade dendrítica de columna vertebral nas ordes de rama distal (4 +) en comparación cos controis (G, H), ANOVA bidireccional con post-probas de Bonferroni, *P <0.05, **P <0.01, n = 9; control e n = 9; sacarosa a longo prazo. Barras de escalas: (A, B) = 20 μm; inserción de (A, B) = 10 μm; (C) = 1 mm.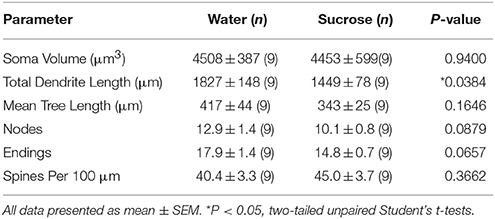
Táboa 2. Parámetros morfolóxicos xerais das neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a longo prazo e controis de auga correspondentes á idade.
Seguindo a caracterización da morfoloxía dendrítica xeral de MSN de shell de NAc que consumen sacarosa a longo prazo, analizamos as arborizacións dendríticas e as densidades da columna vertebral en canto ás súas características de orde rama. A nosa avaliación completa das árbores dendríticas cuantificou o número de segmentos dendríticos por orde de rama, a lonxitude media dos segmentos dendríticos por orde de rama e a densidade media de columna vertebral por orde de rama de MSN de cuncha de NAc de control de auga e ratas que consumen sacarosa a longo prazo. Na táboa móstrase un resumo dos datos e análises da orde de sucursal 3.
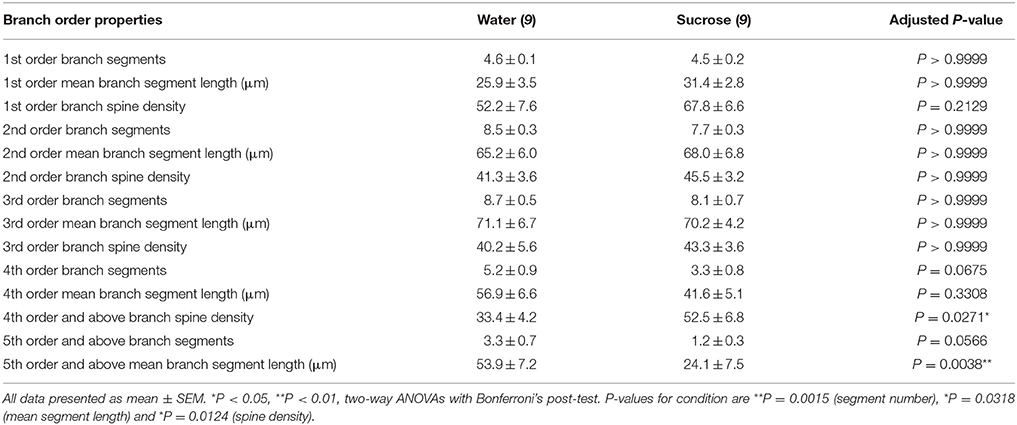 Táboa 3. Características de rama das neuronas de espiña media procedentes de sacarosa e ratas que beben auga.
Táboa 3. Características de rama das neuronas de espiña media procedentes de sacarosa e ratas que beben auga.O número de segmento dendrítico medio por rama por MSNs de cuncha de NAc reduciuse significativamente en ratas que consumen sacarosa a longo prazo en comparación cos controis de auga (**P = 0.0015, ANOVA bidireccional). As probas posteriores a Bonferroni revelaron unha tendencia cara a un número reducido de segmentos en 4th (Auga: 5.2 ± 0.9, n = 9; Sacarosa 3.3 ± 0.8, n = 9, P = 0.0675, Figura 2F, Táboa 3) e 5a orde e por encima das sucursais (Auga: 3.3 ± 0.7, n = 9; Sacarosa 1.2 ± 0.3, n = 9, P = 0.0566, Figura 2F, Táboa 3). A lonxitude media do segmento dendrítico por orde de rama de MSNs de cuncha de NAc tamén se reduciu significativamente en ratas que consumen sacarosa a longo prazo en comparación cos controis de auga (*P = 0.0444, ANOVA bidireccional). As probas posteriores a Bonferroni mostraron unha redución de 55% nas sucursais de orde 5 e máis alá (Auga: 53.9 ± 7.2 μm, n = 9; Sacarosa 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figura 2G, Táboa 3).
A análise das ordes das sucursais mostrou un aumento significativo na densidade da columna vertebral dendrítica dos MSN do shell de NAc de ratas que consumen sacarosa a longo prazo en comparación cos controis (*P = 0.0124, ANOVA bidireccional). As probas posteriores a Bonferroni mostraron un aumento da densidade da columna vertebral do 57% nas ramas distantes de orde 4 e máis alá (Auga: 33.4 ± 4.2, n = 9; Sacarosa 52.5 ± 6.8, n = 9, P = 0.0271 *, inserción de figuras 2A, B, H, Táboa 3). Nas imaxes representativas da arquitectura MSN en xeral e da densidade de columna vertebral distal (inserida) móstranse nas figuras 2A, B.
En conxunto, estes resultados indican que o consumo de sacarosa a curto prazo ten poucos efectos nos parámetros morfolóxicos dos MSN dentro do shell NAc. Non obstante, tras un consumo prolongado, hai unha diminución significativa na lonxitude e complexidade do arbor neuronal, particularmente nas ramas dendríticas distais. Tamén aumentan a densidade de columna distal concomitante nos MSN de cuncha de NAc de ratas que consumen sacarosa a longo prazo.
As neuronas de espiña media procedentes do núcleo do núcleo de Accumbens reducen a complexidade de ramificación despois do consumo de sacarosa a longo prazo, pero non a curto prazo.
Tras o curto prazo do consumo de sacarosa, non houbo diferenzas significativas nos parámetros morfométricos de MSN núcleo de NAc (Táboa 4). Tampouco houbo diferenzas significativas entre o consumo de sacarosa de 4 semana e os MSN núcleos de control de auga nas análises relacionadas coa orde de ramas centrífugas. É dicir, segmentos dendríticos por orde de sucursal (P = 0.7717), lonxitude dendrítica media por orde de sucursal (P = 0.2096), e a densidade media da columna vertebral por orde de sucursal (P = 0.3521, ANOVA bidireccionais) non eran diferentes entre grupos.
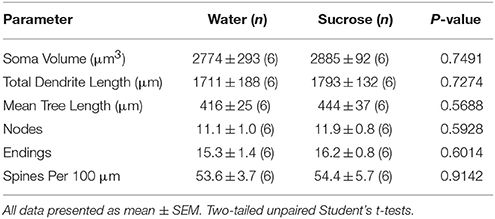 Táboa 4. Parámetros morfolóxicos xerais das neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a curto prazo e controis de auga correspondentes á idade.
Táboa 4. Parámetros morfolóxicos xerais das neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a curto prazo e controis de auga correspondentes á idade.O consumo prolongado de sacarosa tampouco tivo significativos nos parámetros morfométricos do núcleo de NAc (Táboa) 5). O número medio de segmento dendrítico por orde de rama de MSN núcleos de NAc reduciuse significativamente en ratas que consumen sacarosa a longo prazo en comparación cos controis de auga (*P = 0.0416, ANOVA bidireccional), non obstante non houbo diferenzas significativas na lonxitude dendrítica media por orde de sucursal (P = 0.0995) e a densidade media da columna vertebral por orde de sucursal (P = 0.4888, ANOVA de dúas vías) entre os MSN no núcleo de NAc de ratas que consumen sacarosa a longo prazo en comparación cos controis de auga. En conxunto, os nosos datos mostran que o núcleo NAc non responde ao consumo de sacarosa a longo prazo en comparación cos MSN da rexión de cuncha de NAc.
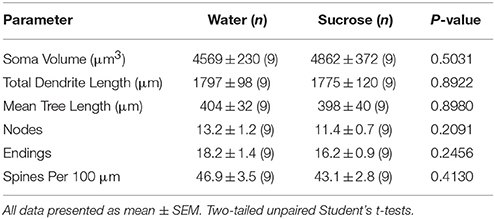 Táboa 5. Parámetros morfolóxicos xerais de neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a longo prazo e controis de auga correspondentes á idade.
Táboa 5. Parámetros morfolóxicos xerais de neuronas de espiña media do núcleo accumbens de ratas consumidoras de sacarosa a longo prazo e controis de auga correspondentes á idade.Conversa
A maior dispoñibilidade de alimentos altamente edulcorados na dieta occidental non só contribuíu ao aumento da prevalencia e carga económica da obesidade e a diabetes tipo II, senón que tamén provocou o inicio de trastornos alimentarios como a inxección alimentaria (Swanson et al., 2011; Kessler et al., 2013; Davis, 2015). Aínda que as propiedades adictivas dos azucres, incluída a fructosa e a sacarosa, seguen sendo especulativas, hai unha rechamante similitude nos correlacionais condutuais e neuronais que se manifestan como consecuencia do consumo excesivo e o consumo prolongado de drogas. (Avena e col., 2008, 2011). Ademais, o azucre activa os circuítos de recompensa do cerebro dun xeito similar ás drogas de abuso (Volkow et al., 2012) e os resultados de estudos humanos suxiren que o azucre e a dozura poden inducir ansias comparables en magnitude ás inducidas por drogas adictivas como o alcol e a nicotina (Volkow et al., 2012). Por iso, utilizamos un modelo de consumo de binge-sacarosa en ratas para determinar os efectos do consumo de sacarosa a curto prazo (4 semanas) e a longo prazo (12 semanas) na morfoloxía neuronal de MSNs na NAc, un compoñente clave do circuíto de recompensa superposto. que está modulada polo azucre e as drogas adictivas. Demostramos que os MSN do cáscara de NAc de ratas que consumen sacarosa crónica a longo prazo diminuíron significativamente a lonxitude e complexidade dendrítica, pero aumentaron a densidade distrinal da columna dendrítica. O consumo a longo prazo de sacarosa non tivo ningún efecto sobre a morfoloxía de MSN do núcleo NAc, mentres que o consumo de sacarosa a curto prazo tampouco tivo ningún efecto significativo na morfoloxía de MSN a partir do núcleo ou da cuncha de NAc. Estes resultados non só demostran un efecto directo da inxestión prolongada de sacarosa tipo binge na morfoloxía neuronal dos MSN shell de NAc, senón que tamén resaltan as consecuencias potencialmente nocivas do consumo prolongado de dietas con alto contido de azucre.
O NAc, que forma parte do estriat ventral, está formado principalmente por MSNs, que se caracterizan morfológicamente como neuronas de tamaño medio con extensas arborisaciones dendríticas e alta densidade de columna vertebral (Kemp e Powell, 1971; Graveland e DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). As neuronas glutamaterxicas e dopaminérxicas son as dúas entradas primarias aferentes ao NAc, que primordialmente entran en contacto cos eixes dendríticos e as espiñas dos MSN. (Groves, 1980; Kaiya e Namba, 1981; Groves et al., 1994). En concreto, a cuncha e núcleo de NAc reciben entrada glutamaterxica de áreas corticais funcionalmente distintas (Brog et al., 1993). A cuncha NAc tamén está innervada por aferentes excitadores de rexións subcorticais como o hipocampo, o tálamo e a amígdala basolateral (Brog et al., 1993; Wright e Groenewegen, 1995). Estudos previos demostraron que estas entradas glutamaterxicas xogan un papel fundamental na motivación e comportamentos dirixidos a obxectivos como a busca de alimentos e recompensas. (Maldonado-Irizarry et al., 1995; Kelley e Swanson, 1997; Reynolds e Berridge, 2003; Richard e Berridge, 2011). A outra entrada predominante nos MSNs NAc provén de aferentes dopaminérxicos que se proxectan desde a área tegmental ventral (Lindvall e Björklund, 1978; Veening et al., 1980; Kalivas e Miller, 1984). Curiosamente, estudos previos empregando modelos semellantes de acceso intermitente ao azucre demostraron que o consumo de tipo binge resultante produce un aumento da dopamina extracelular no NAc de xeito similar (aínda que en menor medida) aos fármacos de abuso. (Rada et al., 2005; Avena e col., 2006) e pode modular a expresión do receptor da dopamina (Colantuoni et al., 2001, 2002) no núcleo e shell de NAc. Curiosamente, o consumo de sacarosa tipo binge provoca unha escalada na inxestión ao longo do tempo de xeito similar á autoadministración de drogas de abuso como a cocaína e os heroesn (Ahmed e Koob, 1998; Ahmed et al., 2000, 2003), que está asociado ao desenvolvemento dun estado "adictivo como".
A nosa análise da morfometría das ordes ramais demostra que a redución global da lonxitude dendrítica dos MSN do shell de NAc causada pola inxestión de sacarosa a longo prazo, resulta principalmente da redución da complexidade das ordes ramas distais. Observamos redución de ramificacións distais (orde 4th e 5th e por encima das ordes ramais) e redución significativa da lonxitude media na orde 5th e por riba das dendritas, combinada co aumento da densidade da columna vertebral nestas ordes. Un factor común susceptible de influír neste tipo de reestruturación dendrítica inclúe cambios na conectividade e / ou na función sinápticas (Russo et al., 2010). Estudos anteriores demostraron que as sinapses glutamaterxicas nas MSN están formadas principalmente en columna vertebral, particularmente en dendritas distais (Groenewegen et al., 1999). Adicionalmente, co-localización de aportes de dopamina e glutamaterxicos da córtex prefrontal (Sesack e Pickel, 1992), hipocampo (Totterdell e Smith, 1989; Sesack e Pickel, 1990), e amígdala (Johnson et al., 1994) observáronse en espiñas dendríticas de MSN. Estas observacións combinadas co aumento da densidade de columna vertebral despois do consumo de sacarosa a longo prazo visto no noso estudo, apoian a formación de maiores entradas excitadoras. Polo tanto, xorde a posibilidade de que os efectos persistentes provocados por unha inxestión prolongada de sacarosa como binge podería facilitar o aumento da actividade sináptica excitadora nas dendritas distales de MSNs no shell de NAc. En consecuencia, pode producirse unha redución e / ou retracción de dendritas distales mediante un mecanismo homeostático sináptico (Reissner e Kalivas, 2010) Non obstante, isto está por determinar.
É interesante notar que Crombag e colegas demostraron que non houbo un aumento da densidade de columna na cáscara de NAc tras o consumo de sacarosa de 4 semana a través do paradigma de autoadministración nasa-poke, a pesar dunha adquisición máis robusta e unha maior taxa de resposta á sacarosa. con anfetamina (Crombag et al., 2005). A súa observación dunha ausencia de cambio na densidade da columna vertebral nas semanas 4 reflicte os nosos resultados. Por contraste, non obstante, o noso estudo demostra que despois da exposición a longo prazo (semana 12) ao consumo de sacarosa crónica, hai un aumento significativo da densidade de columna distal nos MSN dos ratos con experiencia en sacarosa. Ademais, o noso laboratorio demostrou anteriormente que o consumo de sacarosa a longo prazo (semana 12) facilita unha resposta farmacolóxica diferencial a farmacoterapéutica que demostrou que modulan as respostas de dopamina e acetilcolina ao nivel do NAc (Shariff et al., En prensa). En conxunto, isto suxire que a exposición a sacarosa a longo prazo (12 semanas e máis alá), que reflicte máis precisamente os escenarios do mundo real, ten como consecuencia adaptacións de morfoloxía a nivel do NAc.
En termos de drogas de abuso, a exposición repetida a varios medicamentos produce cambios de longa duración na estrutura de dendritas e espiñas dendríticas. Por exemplo, as anfetaminas e a cocaína aumentan a densidade da columna vertebral no NAc tanto na cuncha como no núcleo (Robinson e Kolb, 2004). Tamén se demostrou que a exposición á nicotina aumenta a densidade da columna vertebral na cuncha de NAc. Pola contra, a exposición á morfina conduce a unha diminución da densidade da columna vertebral e da complexidade da rama dendrítica (Robinson e Kolb, 2004). En termos de consumo de sacarosa a longo prazo, observamos un aumento da densidade da columna vertebral similar á anfetamina, a cocaína e a nicotina e oposto ao efecto da morfina. Non obstante, a diferenza da anfetamina e a cocaína, pero semellante á nicotina, o aumento da densidade da columna vertebral na exposición a longo prazo á sacarosa está limitado á cuncha de NAc. Tamén é interesante que cambien as ramificacións dendríticas (Robinson e Kolb, 1999) e densidade de columna vertebral (Li et al., 2003) producidos por anfetamina ou cocaína confínanse a dendritas distais de MSNs no NAc, o que reflicte os resultados do noso estudo. Ademais, e corroborando os cambios descritos anteriormente, o consumo de sacarosa tamén se demostrou anteriormente que aumenta a forza sináptica excitante sobre as neuronas de dopamina acumulada (Stuber et al., 2008b) así como outros compoñentes da vía de recompensa mesolímbica (Stuber et al., 2008a; Chen et al., 2010). En conxunto, isto posúe a sacarosa como un potente modulador da morfoloxía de neuronas despois dun uso pesado prolongado, similar aos efectos observados por drogas de abuso.
Aínda que se precisan novas investigacións para descubrir os mecanismos celulares e sinápticos que contribúen aos cambios morfolóxicos vistos neste estudo, os nosos resultados demostran importantes efectos neuronais provocados polo consumo a longo prazo de sacarosa. En particular, unha consideración non examinada no noso estudo é se os efectos morfolóxicos observados na sacarosa tamén poden orixinarse con edulcorantes non calóricos como a sacarina. A este respecto é importante destacar que Lenoir e compañeiros demostraron que a dulzura intensa supera a recompensa de cocaína, xa sexa xerada por sacarina ou sacarosa (Lenoir et al., 2007). Ademais, un estudo recente publicado polo noso laboratorio (Shariff et al., En prensa) demostra que a varenicline, un agonista parcial do receptor nicotínico da acetilcolina reduciu a inxestión de sacarina e sacarina en roedores seguindo o mesmo réxime de acceso intermitente a longo prazo usado no presente estudo. Curiosamente, estudos anteriores demostraron semellanzas nos efectos agudos de edulcorantes non calóricos como a sacarina e a sacarosa a nivel do NAc (Scheggi et al., 2013; Tukey et al., 2013; Carelli e West, 2014). Non obstante, son necesarios outros estudos para determinar se os edulcorantes non calóricos poden inducir efectos a longo prazo similares aos cambios na morfoloxía dos MSN shell de NAc provocados polo consumo de sacarosa a longo prazo.
A falta de efecto sobre a morfoloxía de MSN NAc tras o consumo de sacarosa a curto prazo, pon de manifesto a importancia de implementar estudos a longo prazo para avaliar o impacto dun abuso prolongado de drogas ou recompensas naturais como a sacarosa. En termos de dependencia, non só son ciclos repetidos de inxestión de binge e abstinencia compoñentes clave do ciclo de adicción, un crecente corpo de evidencias revelou que a transición á dependencia é un proceso progresivo que adoita producirse durante un período prolongado de tempo. Aínda que as propiedades adictivas dos azucres seguen sendo incertas, a plausibilidade da adicción a outros beneficios non farmacéuticos como o sexo, o xogo e a comida estase investigando cada vez máis. Os resultados deste estudo engaden un mérito á hipótese de que azucres como a sacarosa posiblemente teñan propiedades adictivas tras un consumo a longo prazo e semellante a binge. Os nosos resultados tamén teñen implicacións para o número crecente de nenos e adolescentes que manteñen hábitos alimentarios non saudables (alto consumo de azucre e consumo de azucre) na idade adulta. En consonancia co aumento do risco de desenvolver efectos metabólicos, tamén é posible que estas consecuencias neurolóxicas e psiquiátricas que afecten o estado de ánimo e a motivación poidan resultar destes comportamentos.
Contribucións do autor
Participou no deseño da investigación: PK, SB. Experimentos realizados: PK, MS, AB, MF, EM. Análise de datos: PK, MF, MS. Interpretou os datos e contribuíu á escritura do manuscrito: PK, MS, MF, EM, MB, SB. Todos os autores leron e aprobaron o manuscrito final para a súa presentación.
Declaración de conflitos de intereses
Os autores declaran que a investigación foi realizada en ausencia de relacións comerciais ou financeiras que puidesen interpretarse como un potencial conflito de intereses.
Os revisores SC, SA e Management Editor declararon a súa afiliación compartida e o Editor de manipulación afirma que o proceso cumpría as normas dunha revisión xusta e obxectiva.
Grazas
Este traballo contou co financiamento de axudas do Consello de Investigacións australianas (FT1110884) a SB e ao Consello Nacional de Saúde e Investigación Médica (1061979) a SB e MB.
Material complementario
O material suplementario deste artigo pódese atopar en liña en: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Figura complementaria 1. A inxestión de sacarosa e a preferencia das ratas consumidoras de sacarosa 4 e 12 semana. (A, B) amosar escalada na inxestión total de sacarosa (ml) durante 4 e 12 semanas de exposición. (C, D) mostran gran preferencia pola sacarosa sobre a auga durante os períodos de presentación de sacarosa.
References
Ahmed, SH e Koob, GF (1998). Transición dun consumo de drogas moderado a excesivo: cambio no punto de ajuste hedonico. ciencia 282, 298 – 300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF e Parsons, LH (2003). A escala da autoadministración da cocaína non depende dos núcleos inducidos por cocaína alterados nos niveis de dopamina. J. Neurochem. 86, 102 – 113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR e Koob, GF (2000). Aumento persistente da motivación de tomar heroína en ratas con antecedentes de escalada de drogas. Neuropsicopharmacoloxía 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Diabetes tipo 2 en nenos: aspectos clínicos e factores de risco. Res Horm 57 (sux. 1), 19 – 28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG, e Gold, MS (2011). Superposicións na nosoloxía do abuso de substancias e alimentación excesiva: as implicacións tradutivas da "adicción aos alimentos". Curr. Abuso de drogas Rev. 4, 133 – 139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. e Hoebel, BG (2008). Evidencia da adicción ao azucre: efectos comportamentais e neurocúmicos da intermitente e excesiva inxestión de azucre. Neurosci. Biobehav. Rev. 32, 20 – 39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N., e Hoebel, BG (2006). A trituración de sacarosa que se alimenta nun calendario de binge libera varias veces a dopamina e elimina a resposta á saciedade da acetilcolina. Neurociencia 139, 813 – 820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). A plausibilidade da adicción ao azucre e o seu papel na obesidade e nos trastornos alimentarios. Clin. Nutr. 29, 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA, e Popkin, BM (2014). Azucre dietético e peso corporal: chegamos a unha crise da epidemia de obesidade e diabetes ?: a saúde está condenada! Despeje o azucre. Coidado coa diabetes 37, 950 – 956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY e Zahm, DS (1993). Os patróns de inervación aferente do núcleo e a cuncha na parte "accumbens" do estriat ventral de rata: detección inmunohistoquímica de ouro de fluoro transportado retrógradamente. J. Comp. Neurol. 338, 255 – 278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., e Kruseman, M. (2015). Bebidas edulcoradas con azucre e risco de obesidade en nenos e adolescentes: unha análise sistemática sobre como a calidade metodolóxica pode influír nas conclusións. J. Acad. Nutr. Dieta. [Epub antes de imprimir]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM e West, EA (2014). Cando un bo sabor de boca vólvese mal: mecanismos neuronais que subxacen á aparición de afectación negativa e desvalorización da recompensa natural asociada pola cocaína. Neurofarmacoloxía 76 (P B), 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW e Bonci, A. (2010). Plasticidade sináptica no sistema mesolíbico: implicacións terapéuticas para o abuso de substancias. Ann. NY Acad. Sci. 1187, 129 – 139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002) A evidencia de que a inxestión excesiva de azucre intermitente provoca dependencia de opioides endóxena. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001) A inxestión excesiva de azucre altera a unión aos receptores de dopamina e mu-opioides no cerebro. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B., e Robinson, TE (2005). Efectos opostos da experiencia de autoadministración da anfetamina sobre columna dendrítica na córtex prefrontal medial e orbital. Cereb. Córtex 15, 341 – 348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014) Prevalencia da diabetes tipo 1 e tipo 2 entre nenos e adolescentes de 2001 a 2009. Jama 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). A epidemioloxía e a xenética do trastorno por alimentación coxestiva (BED). CNS Spectr. 20, 522 – 529. doi: 10.1017 / s1092852915000462
Ford, ES e Dietz, WH (2013). Tendencias na inxestión de enerxía entre os adultos nos Estados Unidos: resultados de NHANES. Am. J. Clin. Nutr. 97, 848 – 853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD, e Ogden, CL (2014). Prevalencia do sobrepeso, obesidade e obesidade extrema entre adultos: Estados Unidos, 1960 – 1962 a través de 2011 – 2012. Atlanta, GA: CDC.
Graveland, GA, e DiFiglia, M. (1985). A frecuencia e distribución de neuronas de tamaño medio con núcleos indentados no primate e o roedor neostriatum. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV, e Voorn, P. (1999). Converxencia e segregación de entradas e saídas ventrais estriais. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Terminacións sinápticas e os seus obxectivos postsinápticos en neostriatum: especializacións sinápticas reveladas a partir da análise de seccións en serie. Proc. Natl. Acad. Sci. Estados Unidos 77, 6926 – 6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC, e Young, SJ (1994). Axóns dopaminérxicos marcados con hidroxidopamina: reconstrucións tridimensionais de axóns, sinapses e dianas postsinápticas no neostriatum de rata. Neurociencia 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Resolto: hai suficientes evidencias científicas de que a diminución do consumo de bebidas edulcoradas con azucre reducirá a prevalencia de enfermidades relacionadas coa obesidade e a obesidade. Obes. Rev. 14, 606 – 619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z. e Totterdell, S. (1994). Entrada da amígdala ao núcleo de rata acumbens: a súa relación coa inmunoreactividade da tirosina hidroxilase e as neuronas identificadas. Neurociencia 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H., e Namba, M. (1981). Dous tipos de terminais nerviosos dopaminérxicos no neostriatum de rata. Un estudo ultraestrutural. Neurosci. Letra. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW e Miller, JS (1984). As neuronas de neurotensina na área tegmental ventral proxectan o núcleo medial accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ e Emson, PC (1990). Subtipos de proxección de células de matriz neostriatal de rata reveladas por inxección intracelular de biocitina. J. Neurosci. 10, 3421-3438.
Kelley, AE e Swanson, CJ (1997). A alimentación inducida polo bloqueo dos receptores AMPA e kainato dentro do estriat ventral: un estudo de mapeo de microinfusión. Behav. Res. Cerebral. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM e Powell, TP (1971). A organización sináptica do núcleo caudato. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403 – 412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mecanismos de recompensa na obesidade: novas ideas e direccións futuras. Neurona 69, 664 – 679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013) A prevalencia e as correlacións do trastorno alimentario por encargo nas enquisas de saúde mental da Organización Mundial da Saúde. Biol. Psiquiatría 73, 904 – 914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, e Bartlett, SE (2015). Caracterización estrutural e funcional de árbores dendríticos e entradas sinápticas GABAergic en interneuróns e células principais da amígdala basolateral de rata. J. Neurophysiol. 114, 942 – 957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., e Ahmed, SH (2007). A dozura intensa supera a recompensa de cocaína. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B., e Robinson, TE (2003). A localización de cambios persistentes inducidos pola anfetamina na densidade de espiñas dendríticas en neuronas de espiña media no núcleo accumbens e caudato-putamen. Neuropsicopharmacoloxía 28, 1082 – 1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O., e Björklund, A. (1978). Anatomía dos sistemas neuronais dopaminérxicos no cerebro de rata. Adv. Bioquímica. Psicofarmacol. 19, 1-23.
Lutter, M. e Nestler, EJ (2009). Os sinais homeostáticos e hedonicos interactúan na regulación da inxesta de alimentos. J. Nutr. 139, 629 – 632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ e Kelley, AE (1995). Os receptores do glutamato no núcleo accumbens controlan o comportamento da alimentación a través do hipotálamo lateral. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Tras, JP e Hu, FB (2010). Bebidas edulcoradas con azucre, obesidade, diabetes mellitus tipo 2 e risco de enfermidade cardiovascular. Circulación 121, 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM e Popkin, BM (2012). Uso de edulcorantes calóricos e non calóricos en alimentos envasados para consumidores estadounidenses, 2005-2009. J. Acad. Nutr. Dieta. 112, 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM e Popkin, BM (2002). Tendencias na inxestión de enerxía en Estados Unidos entre 1977 e 1996: cambios similares observados entre grupos de idade. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
Paxinos, G. e Watson, C. (2007). O cerebro de ratos en coordenadas estereotóxicas. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Que pasa co enfoque estadounidense da obesidade? Mentor virtual 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM e Hoebel, BG (2005). A inxección diaria de azucre libera repetidamente dopamina na cuncha de acumbens. Neurociencia 134, 737 – 744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW e McNeill, TH (1989). Estudo de Golgi do estriato do rato: cambios dendríticos relacionados coa idade en distintas poboacións neuronais. J. Comp. Neurol. 279, 212 – 227. doi: 10.1002 / cne.902790205
Ranjan, A. e Mallick, BN (2010). Un método modificado para a tinción consistente e fiable de Golgi-cox en tempo significativamente reducido. Diante. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Diabetes mellitus tipo 2 en nenos e adolescentes. Mundo J. Diabetes 4, 270 – 281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ, e Kalivas, PW (2010). Empregar a homeostase con glutamato como obxectivo para tratar trastornos adictivos. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM e Berridge, KC (2003). Conxuntos motivacionais do glutamato no núcleo accumbens: gradientes rostrocaudais de cuncha e medo. EUR. J. Neurosci. 17, 2187 – 2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM e Berridge, KC (2011). Nucleus accumbens dopamina / glutamate cambia a modificación para xerar desexo fronte ao temor: D (1) só para comer apetitoso pero D (1) e D (2) xuntos por medo. J. Neurosci. 31, 12866 – 12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE e Kolb, B. (1999). Alteracións na morfoloxía de dendritas e espiñas dendríticas do núcleo accumbens e cortiza prefrontal despois do tratamento repetido con anfetamina ou cocaína. EUR. J. Neurosci. 11, 1598 – 1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE e Kolb, B. (2004). Plasticidade estrutural asociada á exposición a drogas de abuso. Neurofarmacoloxía 47 (sux. 1), 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC e Nestler, EJ (2010). A sinapsis adicta: mecanismos de plasticidade sináptica e estrutural no núcleo accumbens. Tendencias Neurosci. 33, 267 – 276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J., e Beatty, N. (1969). Un estudo sobre os colaterais axonais das células piramidales en córtex cerebral adulto intacto e parcialmente illado. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC e Elmquist, JK (2002). A necesidade de alimentarse: o control homeostático e hedónico da alimentación. Neurona 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, e Gambarana, C. (2013). Influencia da palatabilidade na motivación para operar para alimentos non calóricos e non calóricos en ratas non privadas de alimentos e privadas de alimentos. Neurociencia 236, 320 – 331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR, e Pickel, VM (1990). No núcleo medial de rata acumbens, os terminais hipocampes e catecolaminérxicos converxen en neuronas espiñentas e están en disposición de estar entre si. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR, e Pickel, VM (1992). Efectos corticales prefrontal na sinapse de rata nos obxectivos neuronais non marcados de terminais de catecolamina no núcleo accumbens septi e sobre as neuronas de dopamina na zona tegmental ventral. J. Comp. Neurol. 320, 145 – 160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (en prensa). Os moduladores neurotínicos do receptor da acetilcolina reducen a inxestión de azucre. PLoS ONE.
Sheehan, DV, e Herman, BK (2015). Os factores psicolóxicos e médicos asociados ao trastorno da alimentación por trato non tratado. Prim. Atención ao coidado Disorder do sistema nervioso central. 17 doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008) O acceso intermitente ao 20% etanol induce un alto consumo de etanol en ratas Long-Evans e Wistar. Alcohol. Clin. Exp. Res. 32, 1816 – 1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, e Bartlett, SE (2007). Varenicline, un agonista parcial do receptor nicotínico da acetilcolina nicotínica alfa4beta2, reduce de xeito selectivo o consumo de etanol. Proc. Natl. Acad. Sci. Estados Unidos 104, 12518 – 12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., e Bonci, A. (2008a). A inxestión voluntaria de etanol potencia a forza sináptica excitadora na área tegmental ventral. Alcohol. Clin. Exp. Res. 32, 1714 – 1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Os indicios de recompensa aumentan a forza sináptica excitativa sobre as neuronas da dopamina cerebral. ciencia 321, 1690 – 1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J., e Merikangas, KR (2011). Prevalencia e correlacións dos trastornos alimentarios en adolescentes. Resultados do complemento nacional de replicación da enquisa sobre comorbididade. Arco. Xen. Psiquiatría 68, 714 – 723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., e Mann, J. (2013). Azucres dietéticos e peso corporal: revisión sistemática e metaanálises de ensaios controlados aleatorios e estudos de cohorte. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S., e Smith, AD (1989). Converxencia de hipocampal e dopaminérxica de entrada ás neuronas identificadas no núcleo accumbens da rata. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., et al. (2013). A inxestión de sacarosa induce un rápido tráfico de receptores AMPA. J. Neurosci. 33, 6123 – 6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM e Lieven, PA (1980). A organización tópica dos aferentes ao caudatoputamen da rata. Un estudo sobre a peroxidasa de raíz. Neurociencia 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R., e Contreras, AM (2014). Base neurobiolóxica da ansia de hidratos de carbono. Nutrición 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., e Baler, R. (2012). Recompensa de alimentos e drogas: circuítos superpostos en obesidade e adicción humana. Curr. Arriba. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Sabio, RA (1973). A inxestión voluntaria de etanol en ratas despois da exposición ao etanol en varios horarios. Psicopharmacoloxía 29, 203 – 210. doi: 10.1007 / BF00414034
Wright, CI, e Groenewegen, HJ (1995). Patróns de converxencia e segregación no núcleo mediano acumbens da rata: relacións de aferentes cortexos prefrontal, talamínico e amigdaloide basal. J. Comp. Neurol. 361, 383 – 403. doi: 10.1002 / cne.903610304
Palabras clave: consumo tipo binge, neurona de espiña media a longo prazo, núcleo accumbens, sacarosa
Citación: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC e Bartlett SE (2016) O consumo prolongado de sacarosa nun manexo de Binge-Like, altera a morfoloxía de neuronas espinas medias no núcleo de Accumbens. Diante. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Recibido: 03 2015 de decembro; Aceptado: 07 March 2016;
Publicado: 23 marzo 2016.
Editado por:
Nora Abrous, Institut des Neurosciences de Bordeaux, Francia
Avaliado por:
Serge H. Ahmed, Centro Nacional da Investigación Científica, Francia
Stéphanie Caille, Centro Nacional da Investigación Científica, Francia
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham e Bartlett. Este é un artigo de acceso aberto distribuído baixo os termos do documento Licenza de atribución de Creative Commons (CC BY). Está permitido o uso, distribución ou reprodución noutros foros sempre que se acredite o autor ou licenciador orixinal e cítase a publicación orixinal nesta revista, de acordo coa práctica académica aceptada. Non se permite ningún uso, distribución ou reprodución que non cumpra estes termos.
* Correspondencia: Selena E. Bartlett, [protexido por correo electrónico]