הערות: ככל שמחקרים מאוחרים יותר יחשפו DeltaFosB הוא המתג המולקולרי הנפוץ הן לסמים והן להתמכרויות התנהגותיות. זהו גורם שעתוק שמשמעותו שהוא משפיע על הגנים שמפעילים או מכבים. כאמור במקומות אחרים, סמים ממכרים חוטפים רק מנגנונים נורמליים. לכן זה טיפשי להציע שהתמכרויות התנהגותיות לא יכולות להתקיים.
Proc Natl Acad Sci US א. 2001 ספטמבר 25; 98 (20): 11042-11046.
doi: 10.1073 / pnas.191352698.
אריק ג 'נסטלר *, מישל בארו, ודוד וו
המחלקה לפסיכיאטריה ומרכז לחקר המוח, המרכז הרפואי של אוניברסיטת טקסס, 5323 הארי היינס בולווארד, דאלאס, טקסס 75390-9070
תַקצִיר
תוחלת החיים של חלק מהפרעות ההתנהגותיות המאפיינות התמכרות לסמים הציעה כי הרגולציה של ביטוי גנים עצביים עשויה להיות מעורבת בתהליך שבו תרופות של התעללות גורמות למצב של התמכרות. אניראיות ncreasing עולה כי גורם שעתוק ΔFosB מייצג מנגנון אחד שבו תרופות של התעללות לייצר שינויים יציבים יחסית במוח אשר תורמים פנוטיפ התמכרות. ΔFosB, חבר במשפחת גורמי השעתוק, מצטבר בתוך תת-קבוצה של נוירונים של הגרעין המכהן והסטריאטום הגבי (אזורי מוח חשובים להתמכרות) לאחר ניהול חוזר של סוגים רבים של סמים. הצטברות דומה של ΔFosB מתרחשת לאחר ריצה כפייתית, דבר המצביע על כך ש- Δ FOSB עשוי לצבור בתגובה לסוגים רבים של התנהגויות כפייתיות. חשוב לציין, ΔFosB נמשכת נוירונים לתקופות זמן ארוכות יחסית בגלל יציבות יוצאת דופן שלה. לכן, ΔFOSB מייצג מנגנון מולקולרי שיכול ליזום ולאחר מכן לקיים שינויים ביטוי גנים כי נמשכים זמן רב לאחר החשיפה לסמים נפסקת. מחקרים בעכברים המהונדסים מהונדסים כי overexpress או ΔFosB או מעכב שלילי דומיננטי של החלבון לספק ראיות ישירות כי ΔFOSB גורם רגישות מוגברת ההשפעות ההתנהגותיות של סמים של התעללות, ואולי, גדל התנהגות סמים המבקשים. עבודה זו תומכת בתפיסה שפונקציות ΔFosB משמשות כסוג של "מתג מולקולרי" מתמשך הממיר בהדרגה תגובות סמים חריפות להתאמות יציבות יחסית התורמות לפלסטיות העצבית והתנהגותית שבבסיס ההתמכרות.
מחקר התמכרות מתמקד בהבנת הדרכים המורכבות שבהן תרופות של התעללות משנות את המוח כדי לגרום לחריגות התנהגותיות המאפיינות התמכרות. אחד האתגרים הקריטיים בתחום הוא לזהות שינויים יציבים יחסית בתרופות המופיעות במוח על מנת להסביר את הפרעות ההתנהגותיות האלה, שהן ארוכות במיוחד. לדוגמה, אדם מכור עלול להיות בסיכון מוגבר להישנות אפילו לאחר שנים של הינזרות.
היציבות של הפרעות התנהגותיות אלה הובילה להצעה כי הם עשויים להיות מתווכת, לפחות בחלקו, באמצעות שינויים ביטוי גנים (1-3). לפי תפיסה זו, החשיפה החוזרת ונשנית לתרופה של התעללות חוזרת ונשנית על התמסורת בסינפסות מסוימות במוח הרגישות לתרופה. הפרעות כאלה מסמנות בסופו של דבר באמצעות מפלרי שליח תאיים אל הגרעין, שבו הם יוזמים תחילה ולאחר מכן שומרים על שינויים בביטויים של גנים ספציפיים. מנגנון עיקרי שבאמצעותו מסלולי התמרת אות משפיעים על ביטוי גנים הוא ויסות גורמי שעתוק, חלבונים המחוברים לאזורים רגולטוריים של גנים ומשנים את התעתיק שלהם.
מטרה אחת של מחקר התמכרות, אם כן, היתה לזהות גורמי שעתוק אשר השתנו באזורי מוח מעורבים התמכרות לאחר ניהול כרוני של סמים של התעללות. מספר גורמי תעתיק כאלה זוהו בעשור האחרון (1-6). המוקד של סקירה זו הוא על גורם שעתוק מסוים בשם ΔFosB.
אינדוקציה של ΔFosB על ידי סמים של התעללות
ΔFosB, המקודד על ידי הגן fosB, הוא חבר במשפחת גורמי השעתוק של Fos, הכוללים גם C-Fos, FosB, Fra1 ו- Fra2 (7). אלה Fos משפחה חלבונים heterodimerize עם יוני חלבונים המשפחה (c-Jun, Junb, או JunD) כדי ליצור אקטיבי AP-1 (activator חלבון-1) גורמי שעתוק המחייבים AP-1 אתרים (רצף קונצנזוס: TGAC / GTCA) נוכח את היזמים של גנים מסוימים כדי להסדיר את שעתוקם.
אלה Fos המשפחה חלבונים הם המושרה במהירות transiently באזורים במוח מסוים לאחר ניהול חריף של סמים רבים של התעללות (איור 1) (8-11). אזורים בולטים הם גרעין ההרכב והסטריאטום הגבי, שהם מתווכים חשובים לתגובות התנהגותיות לתרופות, ובמיוחד לתגמולים המניעים את התנועה שלהם (12, 13). חלבונים אלו חוזרים לרמות הבסיסיות בתוך שעות של ניהול סמים.
איור 1
תכנית המציגה את ההצטברות ההדרגתית של ΔFOSB לעומת האינדוקציה המהירה והחולפת של חלבונים משפחתיים אחרים של פוס בתגובה לתרופות של התעללות. (א) Autoradiogram מדגים את אינדוקציה הדיפרנציאלי של אלה חלבונים שונים על ידי גירוי חריף (1-2 שעה לאחר חשיפה תרופה אחת) לעומת גירוי כרוני (1 יום לאחר חשיפה חוזרת ונשנית של סמים). (B) כמה גלים של חלבונים דמויי FOS [C-FOS (52- ל- 58-kDa isoformform), FosB (46- ל- 50-kDa isoformform), ΔFosB (33-kDa isoform) ו- Fra1 או Fra2 40 kDa)] הם המושרה גרעין accumbens ו נוירונים striatal הגבי ידי הממשל אקוטי של סמים של התעללות. המושרה גם הם איזופורמים שונה ביוכימית של ΔFosB (35-37 kDa); גם הם נגרמים (אם כי ברמות נמוכות) לאחר טיפול חריף בסמים, אך מתמידים במוח במשך תקופות ארוכות בשל יציבותם. (C) עם חזרה חוזרת (למשל, פעמיים ביום) התרופה, כל גירוי חריף גורם רמה נמוכה של isoforms ΔFosB יציבה, אשר מסומן על ידי קבוצה התחתונה של שורות חופפות המציינים ΔFosB המושרה על ידי כל גירוי חריף. התוצאה היא עלייה הדרגתית ברמות הכוללות של ΔFOSB עם גירויים חוזרים במהלך קורס של טיפול כרוני, אשר מסומן על ידי קו הגדילה הגובר בגרף.
תגובות שונות מאוד נראים לאחר ניהול כרוני של סמים של התעללות (איור 1). האיזופורמים המתוקנים ביוכימית של ΔFosB (המסה המולקולרית 35-37 kDa) מצטברים בתוך אזורי מוח שונים לאחר חשיפה חוזרת ונשנית של סמים, בעוד שכל בני משפחת פוס האחרים מראים סובלנות (כלומר, אינדוקציה מופחתת בהשוואה לחשיפה ראשונית לתרופות). הצטברות כזו של ΔFosB נצפתה עבור קוקאין, מורפין, אמפטמין, אלכוהול, ניקוטין, phencyclidine (11, 14-18). יש כמה ראיות כי אינדוקציה זו היא סלקטיבית עבור dynorphin / החומר המכיל משנה של נוירונים קוצני בינוני הממוקם באזורים אלה במוח (15, 17), אם כי יש צורך בעבודה נוספת כדי לבסס את זה בוודאות. 35- ל 37-kDa isoformform של ΔFosB dimerize בעיקר עם ג 'ונד כדי ליצור פעיל וארוך טווח AP-1 מורכבים בתוך אזורים אלה במוח (19, 20). אלה isoformform ΔFBB לצבור עם חשיפה כרונית לסמים בגלל חייהם מחצית חיים יוצא דופן (21), ולכן מתעקשים נוירונים לפחות כמה שבועות לאחר הפסקת ניהול התרופות. זה מעניין לציין כי אלה ΔFosB isoformform הם מוצרים יציבים מאוד של גן מוקדם מיידי (fosb). היציבות של האיזופורמים ΔFosB מספקת מנגנון מולקולרי חדשני שבאמצעותו שינויים המביאים לתרופות בביטוי גנים יכולים להתקיים למרות תקופות ארוכות יחסית של נסיגה של תרופות.
למרות שהגרעין הממלא תפקיד מרכזי בהשפעה המתגמלת של סמים של התעללות, הוא האמין כי הוא פועל כרגיל על ידי הסדרת תגובות לחוזרים טבעיים, כגון מזון, שתייה, סקס ואינטראקציות חברתיות (12, 13). כתוצאה מכך, יש עניין רב בתפקיד אפשרי של אזור מוח זה בהתנהגויות כפייתיות אחרות (למשל, אכילת יתר פתולוגית, הימורים, פעילות גופנית וכו '). מסיבה זו, בדקנו אם Δ FOSB מוסדר במודל חיה של ריצה כפייתית. ואכן, יציבה יציבה 35 ל- 37-kDa isoformforms של ΔFosB הם המושרה באופן סלקטיבי בתוך גרעין accumbens חולדות המציגות התנהגות ריצה כפייתי. †
זהות ביוכימית של איזופורמים יציבים ΔFosB
כפי שצוין לעיל, האיזופורמים ΔFosB המצטברים לאחר ניהול כרוני של סמים של התעללות או ריצה כפייתית מראים מסה מולקולארית של 35-37 kDa. הם יכולים להיות מובחנים מן האיזופור 33-kDa של ΔFosB כי הוא המושרה במהירות אבל transiently לאחר חשיפה תרופה אחת (איור 1) (14, 19, 22). עדויות עדכניות מראות כי האיזופור 33-kDa הוא הצורה הטבעית של החלבון, אשר משתנה כדי ליצור את 35- יציב יותר XNX-kDa מוצרים (37, 19). עם זאת, אופי השינוי הביוכימי הממיר את האיזופורם הבלתי יציב של 21-kDa ל- 33 היציב ל- 35-kDa isoformform נשאר מעורפל. זה כבר השערות כי זרחון עשוי להיות אחראי (37). לדוגמה, אינדוקציה של ΔFOSB הוא attenuated בעכברים חסרים DARPP-11, חלבון מועשר steratal (32, 23). בגלל DARPP-24 מסדיר את הפעילות הקטליטית של חלבון phosphatase- 32 ו חלבון קינאז A (1, 25), הדרישה לחלבון זה עבור הצטברות נורמלית של ΔFosB יציבה איזופורמים מציע תפקיד אפשרי זרחון בדור של מוצרים יציבים אלה.
תפקידו של ΔFosB בהתנהגות פלסטיות לסמים של התעללות
תובנה תפקידו של ΔFos ב התמכרות לסמים באה בעיקר מן המחקר של עכברים מהונדס שבו ΔFOSB יכול להיגרם באופן סלקטיבי בתוך הגרעין accumbens ושאר אזורים striatal של בעלי חיים בוגרים (27, 28). חשוב לציין, עכברים אלה overexpress ΔFosB סלקטיבי ב dynorphin / חומר P המכיל נוירונים קוצניים בינוניים, שבו סמים מאמינים כי לעורר את החלבון. הפנוטיפ ההתנהגותי של עכברי ΔFosB- overexpressing, אשר במובנים רבים דומה לבעלי חיים לאחר חשיפה סמים כרונית, מסוכם בטבלה 1. העכברים מראים תגובת תנועה מוגברת לקוקאין לאחר ניהול חריף וכרוני (28). בנוסף, הם מראים רגישות משופרת להשפעות הגמישות של קוקאין ומורפינים במבחני איזון (11, 28), והם ינהגו בעצמם מינונים נמוכים יותר של קוקאין מאשר מינרלים שאינם מפריזים יתר על המידה ΔFosB ‡ לעומת זאת, רגישות לקוקאין ולמידה מרחבית רגילה במבוך המים של מוריס (28). Tנתונים אלה מצביעים על כך ש- ΔFosB מגביר את רגישות החיה לקוקאין ואולי לסמים אחרים של התעללות ועשוי להוות מנגנון לרגישות ממושכת יחסית לתרופות..
 הסטריאטום
הסטריאטום
| פעילות מוגברת של תנועה בתגובה לאקוויטי וחוזר על עצמו. |
| התגברות התגובות מתגמל קוקאין מורפיום מבחני במקום. |
| הגברת הניהול העצמי של מינונים נמוכים של קוקאין. |
| מוטיבציה מוגברת קוקאין מבחני יחס פרוגרסיבי. |
| התגברות תגובות חרדה לאלכוהול. |
| התנהגות ריצה מוגברת כפייתית. |
מבוסס על נתונים בשופטים. 28 ו 29.† ‡ §¶
פלסטיות התנהגותית מתווכת על ידי ΔFOSB בסטריאטום גרעין אקומבנס-גבי
Iבנוסף, ישנן ראיות ראשוניות לכך שההשפעות של ΔFOSB עשויות להאריך מעבר לרגולציה של רגישות לסמים לתהליכים מורכבים יותר הקשורים לתהליך ההתמכרות. עכברים המבטאים ΔFosB עבודה קשה יותר לניהול עצמי קוקאין ב מתקדמת מבחנים עצמית עצמית יחס, suggesting כי ΔFosB עשוי להרגיש בעלי חיים לתמריץ המוטיבציה של קוקאין ובכך להוביל נטייה להישנות לאחר משיכת התרופהl. ‡ ΔFosB- להביע עכברים גם להראות השפעות חרדה משופרת של אלכוהול, § פנוטיפ כי כבר קשורה עם צריכת אלכוהול מוגברת בבני אדם. יחד, הממצאים המוקדמים הללו מצביעים על כך ש- Δ FOSB, בנוסף להגברת הרגישות לסמים של התעללות, מייצר שינויים איכותיים בהתנהגות המקדמים התנהגות של חיפוש תרופתי. לפיכך, ΔFOSB עשוי לתפקד כמתג מולקולרי מתמשך המסייע ליזום ולאחר מכן לשמור על היבטים מכריעים של המדינה המכורה. שאלה חשובה בחקירה הנוכחית היא האם הצטברות Δ FOSB במהלך החשיפה לתרופות מקדמת התנהגות של התמודדות עם סמים לאחר תקופות נסיגה ממושכות, גם לאחר שרמות ה- Δ FOSB מנורמל (ראה להלן).
מְבוּגָר עכברים כי overexpress ΔFosB באופן סלקטיבי בתוך הגרעין accumbens ו הסטריאטום הגבי גם להציג ריצה כפייתית גדולה יותר בהשוואה littermates שליטה. † תצפיות אלה להעלות את האפשרות המעניינת כי Δ צבירה FosB בתוך נוירונים אלה משמשת תפקיד כללי יותר ביצירת ותחזוקה של זיכרונות הרגל וכפייתית התנהגויות, אולי על ידי חיזוק היעילות של מעגלים עצביים שבהם נוירונים לתפקד.
ΔFosB מצטבר באזורים מסוימים במוח מחוץ לגרעין accumbens ו הסטריאטום הגבי לאחר חשיפה כרונית קוקאין. בולט בין אלה אזורים הם קליפת המוח האמיגדלה והמדיאלית הפריפרונטלית (15). המטרה העיקרית של המחקר הנוכחי היא להבין את התרומות של אינדוקציה ΔFosB באזורים אלה כדי פנוטיפ התמכרות.
עבודה קודמת על עכברי נוקאאוט של fosB העלתה כי בעלי חיים אלה אינם מצליחים לפתח רגישות להשפעות התנועתיות של קוקאין, העולה בקנה אחד עם ממצאי העכברים המבטאים יתר על המידה ב- ΔFosB שהוזכרו לעיל (22). עם זאת, המוטציות של fosB הראו רגישות משופרת להשפעות החריפות של קוקאין, שאינה עולה בקנה אחד עם ממצאים אחרים אלה. פרשנות הממצאים עם המוטציות fosB, אם כי, מסובכת מכך שלחיות אלה חסר לא רק ΔFosB, אלא גם FosB באורך מלא. יתר על כן, למוטציות חסרים שני חלבונים בכל המוח ומהשלבים הראשונים של ההתפתחות. ואכן, עבודה עדכנית יותר תומכת במסקנות מהעכברים המביעים יתר על המידה ΔFosB: ביטוי יתר המושרה של מוטציה קטומה של c-Jun, המשמש כנוגד שלילי דומיננטי של ΔFosB, באופן סלקטיבי בגרעינים ובסטריאטום הגבי מראה רגישות מופחתת להשפעות הגמול של קוקאין .¶ ממצאים אלה מדגישים את הזהירות שיש להשתמש בפירוש תוצאות של עכברים עם מוטציות מכוננות וממחישים את חשיבותם של עכברים עם מוטציות ספציפיות הנגרמות ותאי סוג במחקרי פלסטיות במוח הבוגר.
גני מטרה עבור ΔFosB
בגלל ΔFOSB הוא גורם שעתוק, כנראה החלבון גורם גמישות התנהגותית באמצעות שינויים בביטוי של גנים אחרים. ΔFosB נוצר על ידי שחבור אלטרנטיבי של הגן fosB וחסר חלק של התחום C טרנזקטיבציה C הקיימת באורך מלא FosB. כתוצאה מכך, הוצע במקור כי פונקציות ΔFosB כמו מדחס תעתיק (29). עם זאת, עבודה בתרבית תאים הוכיח בבירור כי ΔFosB יכול לגרום או להדחיק AP-1 בתיווך תמליל בהתאם AP-1 האתר בשימוש (21, 29-31). FosB באורך מלא מפעילה את אותו אפקט כמו ΔFOSB על שברי מקדם מסוימים, אך השפעות שונות על אחרים. עבודה נוספת יש צורך להבין את המנגנונים העומדים מאחורי פעולות מגוונות אלה של ΔFosB ו FosB.
הקבוצה שלנו השתמשה בשתי גישות לזיהוי גני יעד עבור ΔFosB. האחת היא גישת הגן המועמדת. בתחילה ראינו קולטני גלוטמט α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic (AMPA) כמו יעדים משוערים, בהתחשב בתפקיד החשוב של העברה גלוטמטרגית בגרעין. העבודה עד היום הצביעה על כך שיחידת משנה מסוימת של קולטן AMPA גלוטמט מסוים, GluR2, עשויה להיות מטרה בתום לב ל- ΔFosB (איור 2). ביטוי GluR2, אך לא ביטוי של יחידות משנה אחרות של קולטן AMPA, מוגבר בגרעין האקומבנס (אך לא בסטריאטום הגבי) לאחר ביטוי יתר של ΔFosB (28), וביטוי של מוטציה שלילית דומיננטית מחליש את יכולתו של קוקאין לגרום לחלבון. בנוסף, האמרגן של הגן GluR2 מכיל אתר קונצנזוס AP-1 הקושר את ΔFosB (28). ביטוי יתר של GluR2 בגרעין האקומבנס, באמצעות העברת גנים בתיווך נגיפי, מגביר את הרגישות של בעל החיים להשפעות המתגמלות של קוקאין, ובכך מחקה חלק מהפנוטיפ שנראה בעכברים המבטאים ΔFosB (28). אינדוקציה של GluR2 עשויה להסביר את הרגישות האלקטרופיזיולוגית המופחתת של נוירונים גרעיניים לאגוניסטים של קולטן AMPA לאחר מתן קוקאין כרוני [32], מכיוון שקולטני AMPA המכילים GluR2 מראים מוליכות כוללת מופחתת וחדירות Ca2 +. היענות מופחתת של נוירונים אלה לתשומות מעוררות עשויה לשפר את התגובות לתרופה של התעללות. עם זאת, הדרכים בהן האותות הדופמינרגיים והגלוטמטרגיים בגרעין האגן מווסתים את ההתנהגות הממכרת נותרו עלומים; זה ידרוש רמת הבנה במעגל העצבי, שעדיין אינה זמינה.
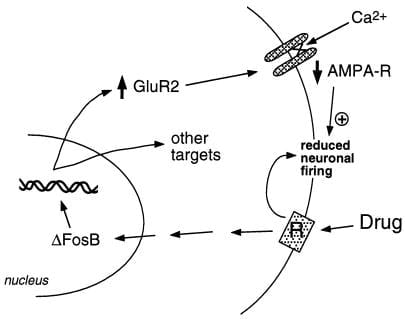
איור 2
יחידת המשנה לקולטני AMPA גלוטמט, GluR2, היא מטרה משוערת עבור ΔFosB. מוצג כיצד אינדוקציה ΔFosB בתיווך של GluR2 עשויה לשנות את ההיענות הפיזיולוגית של נוירונים גרעיניים, ולהוביל לתגובות רגישות לסמים של התעללות. על פי תוכנית זו, סמים של התעללות מייצרים את השפעותיהם המחזקות החריפות באמצעות עיכוב של נוירונים בגרעין. עם חשיפה חוזרת ונשנית, התרופות גורמות ל- ΔFosB, המסדיר גני יעד רבים, כולל GluR2. זה מגדיל את חלקם של קולטני AMPA (AMPA-R) בתאי עצב גרעין המכילים את יחידת המשנה GluR2, מה שגורם להפחתת זרם ה- AMPA הכולל ולהפחתת זרם ה- Ca2 +. רגישות מופחתת זו עשויה להפוך את הנוירונים לרגישים יותר להשפעות המעכבות החריפות של התרופות ובכך להשפעות המחזקות של התרופות..
מטרה משוערת נוספת עבור ΔFosB הוא גינון קידוד הגן. כאמור, dynorphin מתבטא בתת קבוצות של נוירונים נוצצים בינוניים וקוצניים המראים אינדוקציה של ΔFOSB. Dynorphin נראה לתפקד בלולאה משוב בין תאיים: השחרור שלו מעכב את הנוירונים דופאמינרגי כי innervate נוירונים קוצני בינוני, דרך קולטנים אופיואידים κ נוכח על מסופי עצבים דופאמינרגיים בגרעין accumbens וגם על גופי התא ודנדריטים באזור הטגנטלית הגחון (איור 3) (33-35). רעיון זה עולה בקנה אחד עם היכולת של אגוניסט קולטן κ, על ניהול לתוך אחד משני אזורי המוח האלה, כדי להפחית את התרופה rewarד (35).
Rעבודה ecent הצביע כי ΔFOSB מקטין את הביטוי של dynorphin, "" אשר יכול לתרום להגברת מנגנוני התגמול לראות עם אינדוקציה Δ FOSB. באופן מעניין, גורם נוסף שגורם לתרופות, CREB (מרכיב CAMP בתגובה לחלבון מחייב) (2, 3), מפעיל את האפקט ההפוך: הוא גורם לביטוי דינורפין בגרעין האקומבנס ומקטין את התכונות הגמלות של קוקאין ומורפיום (4). **
Bהפעלת ecucause של CREB מתפוגגת במהירות לאחר ניהול התרופה, רגולציה הדדית זו של דינורפין על-ידי CREB ו- ΔFOSB יכולה להסביר את השינויים ההתנהגותיים ההדדיים המתרחשים במהלך השלבים המוקדמים והמאוחרים של הנסיגה, עם תסמינים רגשיים שליליים ורגישות מופחתת של התרופה השולטת בשלבים המוקדמים של נסיגה, ורגישות לתגמולים המניעים והתמריצים של התרופות השולטות בנקודות זמן מאוחרות יותר.
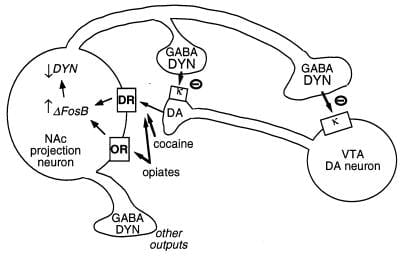
איור 3
Dynorphin הוא יעד putative עבור ΔFosB. המוצג הוא אזור הגחון tegmental (VTA) דופמין (DA) נוירון innervating בכיתה של נוקלאוס Accumbens (NAC) נוירון הקרנה GABAergic המבטא dynorphin (DYN). Dynorphin משמש מנגנון משוב במעגל זה: dynorphin, משוחררים מסופים של נוירונים NAC, פועל על קולטנים אופיואידים κ הממוקם על מסופי העצבים וגופים התא של נוירונים DA כדי לעכב את התפקוד שלהם. ΔFosB, על ידי עיכוב ביטוי dynorphin, עשוי למטה לווסת את הלולאה משוב זה ולשפר את המאפיינים מתגמלת של סמים של התעללות. לא מוצג הוא אפקט הדדי של CREB על מערכת זו: CREB משפר ביטוי dynorphin ובכך מחלישה את התכונות מתגמלת של סמים של התעללות (4). GABA, γ-aminobutyric חומצה; DR, קולטן דופמין; או, קולטן אופיואידים.
הגישה השנייה המשמשת לזיהוי גני יעד עבור ΔFosB כוללת ניתוח microarray DNA. ביטוי יתר של אינדוקציה של ΔFosB מגביר או מקטין את הביטוי של גנים רבים בגרעין האשכבה (36). למרות שנדרשת כעת עבודה לא מבוטלת כדי לאמת כל אחד מהגנים הללו כמטרות פיזיולוגיות של ΔFosB ולהבנת תרומתם לפנוטיפ ההתמכרות, נראה כי יעד חשוב אחד הוא Cdk5 (קינאז -5 תלוי-ציקלין). לפיכך, Cdk5 זוהה בתחילה כ- ΔFosB מוסדר על ידי שימוש במערכי מיקרו, ומאוחר יותר הוכח כי הוא נגרם בגרעין שריר ובסטריאטום הגבי לאחר מתן קוקאין כרוני (37). ΔFosB מפעיל את הגן cdk5 דרך אתר AP-1 הקיים בתוך היזם של הגן (36). יחד, נתונים אלה תומכים בתוכנית בה קוקאין גורם לביטוי Cdk5 באזורי מוח אלה באמצעות ΔFosB. נראה כי אינדוקציה של Cdk5 משנה את האיתות הדופמינרגי לפחות באופן חלקי באמצעות זרחון מוגבר של DARPP-32 (37), אשר מומר ממעכב חלבון פוספטאז -1 למעכב קינאז חלבון A עם זרחונו על ידי Cdk5 (26).
תפקידו של ΔFosB בתיווך פלסטיות "קבועה" לסמים של התעללות
למרות האות ΔFosB הוא ארוך יחסית, זה לא קבוע. ΔFOSB מתדרדר בהדרגה ואינו יכול עוד להתגלות במוח לאחר 1-2 חודשים של נסיגה של תרופות, למרות שחריגות התנהגותיות מסוימות נמשכות לפרקי זמן ארוכים יותר. לכן, Δ FOSB כשלעצמה לא היה מסוגל להיות מסוגל לתווך אלה abnormalities התנהגותית חצי למחצה. הקושי במציאת ההתאמות המולקולריות שביסוד השינויים ההתנהגותיים היציבים ביותר הקשורים בהתמכרות מקביל לאתגרים העומדים בפני תחום הלמידה והזיכרון. למרות שמדובר במודלים תאיים ומולקולריים אלגנטיים של למידה וזיכרון, לא ניתן היה עד כה לזהות התאמות מולקולריות וסלולריות שהן מספיקות זמן רב כדי להסביר זיכרונות התנהגות יציבים מאוד. ואכן, ΔFosB הוא ההתאמה הארוכה ביותר הידועה המתרחשת במוח הבוגר, לא רק בתגובה לסמים של התעללות, אלא גם לכל הפרעה אחרת (שאינה כוללת נגעים). שתי הצעות התפתחו, הן בתחום ההתמכרות והן בתחום הלמידה והזיכרון, כדי להסביר את הפער הזה.
אפשרות אחת היא ששינויים ארעיים יותר בביטוי גנים, כגון אלה המתווכים באמצעות ΔFOSB או גורמי שעתוק אחרים (למשל, CREB) עשויה לתווך שינויים ארוכי טווח יותר במורפולוגיה העצבית ובמבנה הסינפטי. לדוגמה, גידול בצפיפות של הקוצים הדנדריטים (בעיקר גידול שדרה דו-ראשי) מלווה יעילות מוגברת של סינפסות גלוטמטריות ב נוירונים פירמידליים בהיפוקמפוס במהלך potentiation לטווח ארוך (38-40), וקביל הרגישות ההתנהגותית משופרת קוקאין בתיווך ברמה של נוירונים קוצניים בינוניים של הגרעין accumbens (41). זה לא ידוע אם שינויים מבניים כאלה מספיק זמן רב כדי להסביר שינויים יציבים מאוד בהתנהגות, אם כי אלה נמשכים לפחות 1 חודש של נסיגה סמים. עדויות חדשות מעלות את האפשרות כי ΔFOSB, וזיהויו של Cdk5, הוא מתווך אחד של שינויים המביאים לתרופות במבנה הסינפטי בגרעין האקומבנס (איור 4). ‡‡ כך, עירוי של מעכב Cdk5 לתוך הגרעין המכהן מונע את היכולת של חוזרת קוקאין חוזרת להגדיל את הצפיפות עמוד השדרה הדנדריטים באזור זה. זה עולה בקנה אחד עם הדעה כי Cdk5, אשר מועשר במוח, מסדיר את המבנה העצבי ואת הצמיחה (ראה שופט 36 ו 37). זה אפשרי, אם כי בשום אופן לא הוכח, כי שינויים כאלה במורפולוגיה העצבית עשויה להאריך את האות ΔFosB עצמו.
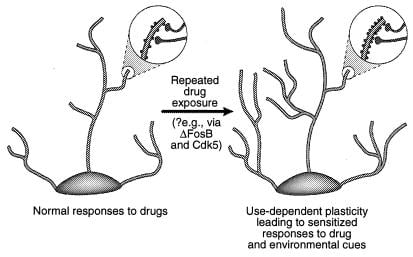
איור 4
ויסות המבנה הדנדריטי על ידי סמים של התעללות. מוצג הרחבת העץ הדנדריטי של נוירון לאחר חשיפה כרונית לתרופה של התעללות, כפי שנצפה עם קוקאין בגרעין העורקים ובקליפת המוח הקדם חזיתית (41). אזורי ההגדלה מראים עלייה בקוצים הדנדריטים, הנחשבת להתרחש בשילוב עם מסופי עצב מופעלים. עלייה זו בצפיפות עמוד השדרה הדנדריטית עשויה להיות מתווכת באמצעות ΔFosB וכתוצאה מכך אינדוקציה של Cdk5 (ראה טקסט). שינויים כאלה במבנה הדנדריטי, הדומים לאלה שנצפו בחלק ממודלי הלמידה (למשל, עוצמה ארוכת טווח), יכולים לתווך תגובות רגישות לאורך זמן לתרופות של התעללות או רמזים סביבתיים. [משוכפל באישור של נ.צ. 3 (זכויות יוצרים 2001, מגזינים מקמיליאן בע"מ)].
אפשרות נוספת היא כי אינדוקציה חולפת של גורם שעתוק (למשל, ΔFosB, CREB) מוביל לשינויים קבועים יותר ביטוי גנים באמצעות שינוי של chromatin. גורמים אלה ורבים אחרים שעתוק הם האמינו להפעיל או לדכא את שעתוק הגן היעד על ידי קידום acetylation או deacetylation, בהתאמה, של היסטונים באזור הגן (42). אף על פי שאסטילציה ודה-סטיילציה כאלה של היסטונים יכולים כנראה להתרחש במהירות רבה, ייתכן כי ΔFOSB או CREB עשויים לייצר הסתגלות ממושכת יותר במנגנון האנזימטי השולט על אצטילציית ההיסטון. ΔFOSB או CREB עשויים גם לקדם שינויים ארוכי טווח בביטוי גנים על ידי ויסות שינויים אחרים של הכרומטין (למשל, DNA או היסטון מתילציה), אשר היו מעורבים בשינויים הקבועים בתעתיק גנים המתרחשים במהלך הפיתוח (ראה ש '42 ו- 43) . אף על פי שאפשרויות אלה נותרות ספקולטיביות, הן יכולות לספק מנגנון שבו הסתגלות ארעית לתרופה של התעללות (או כל הפרעה אחרת) תוביל לתוצאות התנהגותיות משמעותיות לכל החיים.
הפניות
- ↵
- Nestler EJ,
- הופ BT,
- ווידנל ק
(1993) נוירון 11: 995-1006.
- ↵
- Berke JD,
- היימן SE
(2000) נוירון 25: 515-532.
- ↵
- נסלר EJ
(2001) Nat Rev Neurosci 2: 119-128.
- ↵
- Carlezon WA Jr,
- Thome J,
- אולסון VG,
- Lane-Ladd SB,
- Brodkin ES,
- הירוי N,
- Duman RS,
- נווה ר.ל.,
- נסלר EJ
(1998) מדע 282: 2272-2275.
- אודונובן ק.ג'יי,
- Tourtellotte WG,
- Millbrandt J,
- ברבן
(1999) מגמות Neurosci 22: 167-173.
- ↵
- Mackler SA,
- Korutla L,
- צ'ה XY,
- קובה,
- Fournier KM,
- באוארס MS,
- Kalivas PW
(2000) J Neurosci 20: 6210-6217.
- ↵
- Morgan JI,
- קאראן ט
(1995) מגמות Neurosci 18: 66-67.
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci ארה"ב 88: 1291-1295.
- גרייביל,
- מורתאללה ר,
- רוברטסון
(1990) Proc Natl Acad Sci ארה"ב 87: 6912-6916.
- תקווה B,
- קוסופסקי ב,
- היימן SE,
- נסלר EJ
(1992) Proc Natl Acad Sci ארה"ב 89: 5764-5768.
- ↵
- קלץ,
- נסלר EJ
(2000) Curr דעות Neurol 13: 715-720.
- ↵
- Koob GF,
- סאנה PP,
- בלום FE
(1998) נוירון 21: 467-476.
- ↵
- חכמים
(1998) אלכוהול התלות אלכוהול 51: 13-22.
- ↵
- הופ BT,
- Nye HE,
- קלץ,
- עצמי DW,
- Iadarola MJ,
- Nakabeppu Y,
- Duman RS,
- נסלר EJ
(1994) נוירון 13: 1235-1244.
- ↵
- Nye H,
- הופ BT,
- קלץ ז,
- Iadarola M,
- נסלר EJ
(1995) J Pharmacol Exp 275 1671: 1680-XNUMX.
- Nye HE,
- נסלר EJ
(1996) מול פרמקול 49: 636-645.
- ↵
- מורתאללה ר,
- אליבול B,
- Vallejo M,
- גרבאל
(1996) נוירון 17: 147-156.
- ↵
- Pich EM,
- Pagliusi SR,
- Tessari M,
- טלבוט-אייר ד,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) מדע 275: 83-86.
- ↵
- חן JS,
- Nye HE,
- קלץ,
- הירוי N,
- Nakabeppu Y,
- הופ BT,
- נסלר EJ
(1995) מול פרמקול 48: 880-889.
- ↵
- הירוי N,
- Brown J,
- Ye H,
- Saudou F,
- Vaidya VA,
- Duman RS,
- גרינברג,
- נסלר EJ
(1998) J Neurosci 18: 6952-6962.
- ↵
- חן J,
- קלץ,
- הופ BT,
- Nakabeppu Y,
- נסלר EJ
(1997) J Neurosci 17: 4933-4941.
- ↵
- הירוי N,
- Brown J,
- היילה C,
- Ye H,
- גרינברג,
- נסלר EJ
(1997) Proc Natl Acad Sci ארה"ב 94: 10397-10402.
- ↵
- פיינברג AA,
- הירוי N,
- מרמלשטיין,
- שיר WJ,
- סניידר GL,
- Nishi A,
- Cheramy A,
- O'Callaghan JP,
- מילר D,
- קול DG,
- et al.
(1998) מדע 281: 838-842.
- ↵
- הירוי N,
- פיינברג א,
- היילה C,
- Greengard P,
- נסלר EJ
(1999) Eur J Neurosci 11: 1114-1118.
- ↵
- Greengard P,
- אלן PB,
- Nairn AC
(1999) נוירון 23: 435-447.
- ↵
- BIB JA,
- סניידר GL,
- Nishi A,
- Yan Z,
- Meijer L,
- פיינברג AA,
- Tsai LH,
- Kwon YT,
- Girault JA,
- Czernik AJ,
- et al.
(1999) טבע (לונדון) 402: 669-671.
- ↵
- חן JS,
- קלץ,
- Zeng GQ,
- Sakai N,
- סטפן C,
- שוקט PE,
- Picciotto M,
- Duman RS,
- נסלר EJ
(1998) מול פרמקול 54: 495-503.
- ↵
- קלץ,
- חן JS,
- קרלזון WA,
- ויסלר ק,
- גילדן L,
- בקמן,
- סטפן C,
- ג 'אנג YJ,
- Marotti L,
- SW עצמי,
- et al.
(1999) טבע (לונדון) 401: 272-276.
- ↵
- Dobrazanski P,
- Noguchi T,
- Kovary K,
- Rizzo CA,
- לאזו PS,
- בראבו ר
(1991) Mol Cell Biol 11: 5470-5478.
- Nakabeppu Y,
- Nathans D
(1991) תא 64: 751-759.
- ↵
- ין J,
- חוכמה RM,
- Tratner I,
- Verma IM
(1991) Proc Natl Acad Sci ארה"ב 88: 5077-5081.
- ↵
- לבן FJ,
- הו XT,
- ג'אנג XF,
- וולף
(1995) J Pharmacol Exp 273 445: 454-XNUMX.
- ↵
- היימן SE
(1996) נוירון 16: 901-904.
- קריק MJ
(1997) Pharmacol Biochem Behav 57: 551-569.
- ↵
- שיפנברג TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449-455.
- ↵
- חן JS,
- ג 'אנג YJ,
- קלץ,
- סטפן C,
- Aneg,
- זנג L,
- נסלר EJ
(2000) J Neurosci 20: 8965-8971.
- ↵
- BIB JA,
- חן JS,
- טיילור ג '
- Svenningsson P,
- Nishi A,
- סניידר GL,
- Yan Z,
- Sagawa ZK,
- Nairn AC,
- Nestler EJ,
- et al.
(2001) טבע (לונדון) 410: 376-380.
- ↵
- Luscher C,
- ניקול ר,
- Malenka RC,
- מולר ד
(2000) Nat Neurosci 3: 545-550.
- מלינוב,
- Mainen ZF,
- הייאשי י
(2000) Curr דעות Neurobiol 10: 352-357.
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133-141.
רובינסון, TE & Kolb, B. (1999) (1997) Eur. ג 'יי Neurosci.11, 1598-1604.
- ↵
- קארי M,
- Smale ST
(2000) תקנה תעתיק ב Eukaryotes (Cold Spring Harbor Lab. Press, Plainview, NY).
- ↵
- ספנסר VA,
- דייווי ג 'ר
(1999) גן 240: 1-12.
 פייסבוק
פייסבוק- טויטר
- CiteULike
- טָעִים מְאוֹד
 Digg
Digg- Mendeley
HighWire Press-hosted מאמרים citing מאמר זה
- טבעי ותרופות Rewards חוק על מנגנוני פלסטיות עצביים נפוצים עם {דלתא} FosB כמגשר מפתח ג 'יי Neurosci. 2013 33 (8) 3434-3442
- סמים, פשע, ואת אפיגנטיקה של אלדוזיס הדוני כתב העת של המשפט הפלילי עכשווי 2012 28 (3) 314-328
- תַקצִיר
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- תַקצִיר
- טקסט מלא (HTML)
- טקסט מלא (PDF)
- מורפין מפעילה את E-XX עשרים שעתוק תמלול פקטור 1 / סרום תגובה פקטור נתיב דרך קינאזות מווסתים תאיים 1 / 2 בתאי F11 נגזר נוירונים שורש הגב שורש ג 'יי פרמקול. Exp. . 2012 342 (1) 41-52
- מנגנון מולקולרי עבור Gateway סמים: שינויים epigenetic יזום על ידי ניקוטין ראש ביטוי ג 'ין על ידי קוקאין 2011 3 (107) 107ra109
- משופרת Sucrose וקוקאין עצמי המינהל ואת קיו- Induced סמים מחפשים אחרי אובדן של VGLUT2 ב נוירונים דופאמין Midbrain בעכברים ג 'יי Neurosci. 2011 31 (35) 12593-12603
- היפוקסיה כרונית לסירוגין מגבירה את לחץ הדם ואת הביטוי של FosB / {Delta} FosB באזורים מרכזיים אוטונומיים Am ג 'יי פיסיול. רגול. אינטגר. Comp. פיסיול. 2011 301 (1) R131-R139
- היעדר קולטן GPR37 / PAEL פוגע סטריאטלית Act ו- ERK2 זרע, הביטוי דלתא הביטוי FosB, והעדפה מקום מותנה אמפטמין וקוקאין FASEB 2011 25 (6) 2071-2081
- הקשר בין משך חשיפה ראשונית לאלכוהול לבין ההתמדה של סובלנות מולקולרית הוא בעליל לא לינארי ג 'יי Neurosci. 2011 31 (7) 2436-2446
- ב Vivo פליטת אור הדמיה חושפת Redox- מוסדר Activator חלבון- 1 הפעלה גרעין Paraventricular של עכברים עם יתר לחץ דם ורידוסקולרית לחץ דם 2011 57 (2) 289-297
- ביטוי יתר של סטארטלי של {Delta} FosB מחזירה תנועה כרונית של לבודופה ג 'יי Neurosci. 2010 30 (21) 7335-7343
- גישור אפיגנטי של השפעות סביבתיות בהפרעות פסיכוטיות מרכזיות Schizophr בול 2009 35 (6) 1045-1056
- בדיקות מבוססות MRI מבוססות דנ"א לאיתור ספציפי של חשיפה כרונית לאמפתמין במוח החי ג 'יי Neurosci. 2009 29 (34) 10663-10670
- השתנות הפלסטית הדנדריטי של עמוד השדרה בחולצות קוקאין ג 'יי Neurosci. 2009 29 (9) 2876-2884
- Overexpression מסך תסיסנית מזהה תפקידים העצבית של GSK-3 {beta} / שאגי כמו רגולטור של AP-1- גנטיקה 2008 180 (4) 2057-2071
- תמליל MRI: תצוגה חדשה של המוח החי Neuroscientist 2008 14 (5) 503-520
- {דלתא} אינדוקציה FosB בקורטקס אורביטופרונטלי מתווך סובלנות לתפקוד קוגניטיבי ג 'יי Neurosci. 2007 27 (39) 10497-10507
- פגיעות מתמשכת כדי להחזיר את ההתנהגות של מתאמפטמין המבקש בקו גליה קו נגזר גורם neurotrophic עכברים מוטציה FASEB 2007 21 (9) 1994-2004
- (דלתא) FosB בגרעין Accumbens מסדיר מזון מחוזק התנהגות אינסטרומנטלית מוטיבציה ג 'יי Neurosci. 2006 26 (36) 9196-9204
- תקנה של {דלתא} יציבות FosB על ידי זרחון. ג 'יי Neurosci. 2006 26 (19) 5131-5142
- ביטוי של קולטנים NMDA מוטציה ב Dopamine D1 תאים המכילים קולטן מונע רגישות קוקאין ומפחית העדפת קוקאין ג 'יי Neurosci. 2005 25 (28) 6651-6657
- D1 דופמין קולטנים מווסתים {דלתא} אינדוקציה FosB בתאי עכברוש לאחר ניהול מורפיום לסירוגין ג 'יי פרמקול. Exp. . 2005 314 (1) 148-154
- נוירוביולוגיה של עכברים נבחרה עבור פעילות ריצה גבוהה וולונטרי אינטגר. Comp. ביול. 2005 45 (3) 438-455
- ההשפעות של מניעת מים והתייבשות ב- c-Fos ו FosB מכתים בגרעין עכברוש supraoptic ו lamina terminalis באזור Am ג 'יי פיסיול. רגול. אינטגר. Comp. פיסיול. 2005 288 (1) R311-R321
- אינדוקציה תעתיק של FosB / [דלתא} FosB גנים על ידי מתח מכני Osteoblasts J Biol כימית 2004 279 (48) 49795-49803
- אינדוקציה של {דלתא} FosB בתגובות מוח הקשורות למוח לאחר לחץ כרוני ג 'יי Neurosci. 2004 24 (47) 10594-10602
- מינון גנטי Sim1 מודולציה התגובה ההומיאוסטית האכלה שומן מוגבר בשומן בעכברים Am ג 'יי פיסיול. אנדוקרינול. מטאב. 2004 287 (1) E105-E113
- DNA ניתוח microarray של ביטוי גנים אנושיים בראש עצב astrocytes בתגובה ללחץ hydrostatic פיסיול. גנומיקה 2004 17 (2) 157-169
- Superoxide מעורב במערכת העצבים המרכזית הפעלה וסימפטוקסיסיטציה של אי ספיקת לב אוטם שריר הלב מעגל. Res. 2004 94 (3) 402-409
- קולטני אדנוזין A2A בהתאמה עצבית לגירוי דופמינרגי חוזר ונשנה: השלכות לטיפול בדיסקינזיות במחלת פרקינסון. נוירולוגיה 2003 61 (90116) S74-81
- Cytoplasmic מול לוקליזציה גרעינית של חלבונים הקשורים FOS ב צפרדע, רנה esculenta, Testis: ב ויוו ישיר במינון השפעת גונדוטרופין משחרר הורמון אגוניסט ביול. Reprod. 2003 68 (3) 954-960
- עכברים Periadolescent הצג משופרת דלתא FosB upregulation בתגובה קוקאין ואמפתמין ג 'יי Neurosci. 2002 22 (21) 9155-9159
- דלתא FosB מסדיר את גלגל ריצה ג 'יי Neurosci. 2002 22 (18) 8133-8138
- פעילות CREB בגרעין פגז גרעיני שולטת של תגובות התנהגותיות לגירויים רגשיים מעבד נטל. אקאד. Sci. ארה"ב 2002 99 (17) 11435-11440
- פסיכוגנומיקה: הזדמנויות להבנה התמכרות ג 'יי Neurosci. 2001 21 (21) 8324-8327