ביול פסיכיאטריה. 2008 דצמבר 1; 64 (11): 941-50. Epub 2008 יולי 26.
Teegarden SL, Nestler EJ, Bale TL.
מָקוֹר
המחלקה לביולוגיה של בעלי חיים, אוניברסיטת פנסילבניה, פילדלפיה, הרשות הפלסטינית 19104-6046, ארה"ב.
תַקצִיר
רקע:
רגישות לתגמל היה מעורב כגורם נטייה להתנהגויות הקשורות לשימוש בסמים, כמו גם אכילת יתר. עם זאת, המנגנונים הבסיסיים התורמים לתגמול הרגישות אינם ידועים. שיערנו כי דיסרגולציה בסימפטום דופאמין עשויה להיות הסיבה הבסיסית להגברת רגישות הגמול, לפיה גירויים מתגמלים יכולים לפעול לנורמליזציה של המערכת.
שיטות:
השתמשנו במודל עכבר גנטי של רגישות גמול מוגברת, עכבר דלתא-overexpressing עכבר, כדי לבחון שינויים מסלול הפרס בתגובה לתזונה גבוהה שומן טעים. סמנים של אותות גמול בעכברים אלה נבדקו הן בסיסיים והן לאחר 6 שבועות של חשיפה לדיאטה טעים. עכברים נבדקו במבחן ההתנהגות בעקבות נסיגה דיאטה גבוהה שומן להעריך את הפגיעות של מודל זה כדי להסיר גירויים מתגמלת.
תוצאות:
התוצאות שלנו מדגימות שינוי מסלול מסלול גמול לאורך הגרעין accumbens- היפותלמי, הגחון אזור המעגל tegmental כתוצאה overexpression של דלתא FosB ב nucleus accumbens ואזורים striatal. רמות של חלבון אדנוזין monosphosphate phosphorylated (cAMP) תגובה רכיב חלבון מחייב (pCREB), נוירוטרופי (BDNF), ו דופמין ו מחזורית אדנוזין monophosphate מוסדר phosphoprotein עם המסה המולקולארית של 32 kDa (DARPP-32) בגרעין accumbens הופחתו עכברים דלתא FosB, מרמז על הפחתת דופמין איתות. שישה שבועות של חשיפה לדיאטה בשומן גבוה שיפרו לחלוטין את ההבדלים הללו, וחשפו את היכולת הגמישה של תזונה בריאה. עכברי דלתא FosB הראו גם עלייה משמעותית בפעילות של תנועה ותגובות הקשורות לחרדה 24 שעות לאחר נסיגה בשומן גבוה.
מסקנות:
תוצאות אלו קובעות רגישות בסיסית לשינויים בתגמול הקשור לדיסרגולציה של איתות דלתא FosB ודופאמין, שניתן לנרמל עם דיאטות טעימות, ועשויים להיות פנוטיפ נטייה בכמה צורות של השמנת יתר.
מבוא
למרות הידע הגובר שלנו על מערכות העצבים השולטים תיאבון ושובע, שיעורי ההשמנה ממשיכים לעלות בארצות הברית. טיפולים תרופתיים הנוכחי יש יעילות מוגבלת, ושינויים התנהגותיים סובלים תאימות לטווח ארוך מינימלי (1). הצריכה של מזונות צפופים וטעימים בקלוריות נקשרה לשינויים בלחץ ובמסלולי תגמול במוח, דבר המצביע על כך שהתכונות המתגמלות של מזונות אלו עשויות לעקוף את אותות מאזן האנרגיה (2-4). מזונות עשירים בשומן פועלים כתגמולים טבעיים, הפעלת מרכזי תגמול במוח באופן דומה לסמים של התעללות, וככאלה שימשו בפרדיגמות של ניהול עצמי (5-8). לכן, סביר להניח כי התנהגויות ומוטיבציה עבור אכילת יתר ושימוש בסמים חולקים מנגנונים בסיסיים הבסיסית, פוטנציאל לפתוח דרכים חדשות לטיפול בשני התנאים.
בחקר הקשר בין מזונות טעים לבין מסלולים המסדירים גמול ומתח במוח, זיהינו בעבר סמנים מולקולריים וביוכימיים של גמול מופחת ועלייה בלחץ בעקבות נסיגה מתזונה עשירה בשומן (HF). בדומה לתרופות של התעללות, חשיפה לתזונה חביבה במחקר שלנו גרמה לרמות גבוהות יותר של גורם השעתוק ΔFosB בגרעין Accumbens (NAc), מבנה מרכזי לתגמולי מוח (9, 10). עכברים כי Overexpress overexpress ΔFBB הצג הגדילה אינסטרומנטלי להגיב על תגמול מזון (11), מה שהופך אותם כלי חשוב לבחינת התפקיד של רגישות גמול dysregulation לטווח ארוך של מערכת תגמול תגובות מולקולריות וביוכימיות לתזונה טעים.
במחקר הנוכחי, ניצלנו את העכברים ΔFosB-overexpressing כדי לבחון שינויים ארוכי טווח בסמנים של תגמול ב neg-hypothalamus- הגחון אזור אזורית (VTA) neurocircuitry בתגובה לדיאטה HF טעים. בהתבסס על מחקרים קודמים בעכברים רגישים לתגמול, הנחנו ששינויים ב- ΔFB-Induced ברגישות הגמול כרוכים בהפרעה באיתות דופאמין הנובע ממשוב NAC ל- VTA. יתר על כן, שיערנו כי חשיפה לתגמול טבעי של דיאטה HF צפופה באנרגיה תאפשר לנרמל את המערכת הדופאמינרגית בעכברים אלה, וכתוצאה מכך תגובה מוגזמת ללחץ הנסיגה מתזונה זו של HF. ההיבט הייחודי של ניצול דיאטה טעים כחומר מתגמל מאפשר לנו לכלול את התשומות ההיפותלמיות לתגמל מעגלים בפנוטיפ שעשוי להיות מנבא של האוכלוסייה נטייה להשמנה עמיד להשמנה. כדי לבחון את ההשערה הזאת, למדנו סמנים של נוירוטרנסמינציה של דופאמין, כולל pCREB, BDNF ו- DARPP-32 ב- NAc ו- hydroxylase tyrosin ו- transporter dopamine ב- VTA, בעקבות חשיפה ל- HF. כמו כן בדקנו סמנים ספציפיים של מאזן האנרגיה הידוע להשפיע על הפלט דופמין, כולל קולטן לפטין ו אורקסין בביטוי VTA ו orexin בתוך ההיפותלמוס לרוחב.
חומרים ושיטות
בעלי חיים
עכברים bitransgenic זכר כי overexpress overexpress ΔFosB ב נוירונים חיובי dynorphin ב NAC ו הסטריאטום הגבי (Kelz et al., 1999) נוצרו על רקע מעורב (ICR: C57Bl6 / SJL) במרכז הרפואי של אוניברסיטת טקסס דרום מערב ומתוחזק ו נבדק באוניברסיטת פנסילבניה. כל העכברים נשמרו על דוקסיציקלין (100 מיקרוגרם / מ"ל במי השתייה) עד הגעתו לאוניברסיטת פנסילבניה. כדי לגרום overexpression, doxcycline הוסר (n = 23) (12). עכברי בקרה (n = 26) המשיכו לקבל את התרופה. עכברים הוקצו קבוצות דיאטה שמונה שבועות לאחר הסרת doxycyline שבו הביטוי זמן הוכח להגיע לרמות מקסימלי (13). עכברים נשמרו על 12: 12 אור כהה מחזור (אורות על 0700) עם מזון ומים זמין libitum המודעה. כל המחקרים בוצעו על פי פרוטוקולים ניסיוניים שאושרו על ידי אוניברסיטת פנסילבניה טיפול בבעלי חיים מוסדיים ועדת שימוש, וכל הנהלים נערכו בהתאם להנחיות המוסדיים.
חשיפה לדיאטה
עכברים נשמרו על chow הבית (n = 16) או להציב על HF (n = 16-17) במשך שישה שבועות. Chow הבית (דיאנה מעבדה Purina, סנט לואיס, MO) הכיל 4.00 קק"ל / גרם, המורכב 28% חלבון, 12% שומן, ו 60 פחמימות%. HF דיאטה (דיאטות מחקר, ניו ברונסוויק, ניו ג 'רזי) הכיל 4.73 קק"ל / גרם, המורכב 20% חלבון, 45% שומן, ו 35% פחמימות.
ביוכימיה וביטוי גנים
עכברים נותחו לאחר שישה שבועות של חשיפה לדיאטה. המוח הוסרו מן הגולגולת או הקפוא כולו על קרח יבש או NAC גזור (כ 0.5 - 1.75 מ"מ מ ברגמה, בעומק של 3.5 - 5.5 מ"מ) וקפא בחנקן נוזלי. רקמות נשמרו ב - 80 ° C עד assayed.
ניתוח ביוכימי
שיטות לכתמים מערביים מתוארות בחומרים משלימים. הנוגדנים בשימוש היו: Cdk5, CREB ו- BDNF (1: 500, סנטה קרוז ביוטכנולוגיה, סנטה קרוז, CA) ו phospho-CREB (pCREB) (133) (1: 500, תא איתות טכנולוגיה, Danvers, MA).
אוטורדיוגרפיה של קולטן
שיטות מפורטות עבור autoradiography מתוארים בחומרים משלימים. Ligands בשימוש היו 2 nM H3 - SCH 23390 ו 5 nM H3 - ספיפרן (PerkinElmer, בוסטון, MA).
הכלאה באתרו
עיבוד רקמות והכלאה בוצעו כמתואר לעיל (14). בדיקה DARPP-32 סופק בחביבות על ידי פ גרינגארד (אוניברסיטת רוקפלר), ואת בדיקה אוקססין על ידי ג 'יי Elmquist (אוניברסיטת טקסס Southwestern מרכז רפואי). Slides assayed עבור DARPP-32 היו apposed לקולנוע עבור 3 ימים, וכן שקופיות assayed עבור orexin היו apposed עבור הסרט עבור 4 ימים. כימות של תמונות הסרט נערך כפי שתואר לעיל (10).
QRT-PCR
RNA היה מבודד VTA ואת הביטוי של גנים בודדים מוערך באמצעות TaqMan מבחני ביטוי גנים (יישומית Biosystems, פוסטר סיטי, קליפורניה). שיטות מפורטות וניתוחים סטטיסטיים ניתן למצוא חומרים משלימים.
ניתוח התנהגותי
על מנת לבחון את ההשפעות של רגישות גמול על שינויים התנהגותיים המושרה על ידי דיאטה, קבוצת משנה של עכברים הוסרה מ HF לאחר ארבעה שבועות של חשיפה וחזרה לבית chow (n = 9 שליטה, n = 8 Δ FosB). עשרים וארבע שעות לאחר הנסיגה, העכברים נחשפו למבחן השדה הפתוח בהתאם לפרדיגמת הנסיגה שלנו שפורסמה בעבר (10). בקצרה, העכבר הונח במרכז מנגנון השדה הפתוח וניטור במשך חמש דקות. סך הכל חוצה קו, צואה, צואה במרכז, וצלבים למרכז נמדדו.
סטָטִיסטִיקָה
כל הנתונים למעט כתמים מערביים נותחו באמצעות ANOVA דו כיווני ואחריו בדיקת PLSD של פישר עם טיפול בדוקסיציקלין (ביטוי ΔFosB) ומצב התזונה כמשתנים הבלתי תלויים. לצורך ניתוחי RT-PCR, נעשה שימוש בהפחתת ערך P כדי לתקן השוואה מרובה בקבוצות של גנים קשורים (ראה חומרים משלימים). כתמים מערביים נותחו באמצעות מבחן t של סטודנט עם טיפול בדוקסיציקלין כמשתנה העצמאי, והשווה צפיפויות אופטית באותו כתם. כל הנתונים מוצגים כממוצע ± SEM.
תוצאות
הבדלים ביוכימיים ביזליים
כדי להבהיר את המסלולים המולקולריים העומדים בבסיס רגישות הגמול המשופרת בעכברים המבטאים יתר על המידה ב- ΔFosB, נבדקו רמות של כמה מולקולות איתות מפתח ב- NAc. הייתה מגמה של רמות מוגברות של Cdk5 ב- NAc של עכברי ΔFosB בהשוואה לבעלי שליטה על חברי המלטה המוחזקים על דוקסיציקלין (F = 5.1, P = 0.08; איור 1 א). עכברי ΔFosB ביטאו רמות מופחתות משמעותית של pCREB (F = 7.4, P <0.05; איור 1B) כמו גם רמות כולל של CREB (F = 5.4, P = 0.05; איור 1C). ירידה משמעותית ב- BDNF נצפתה גם ב- NAc של עכברי ΔFosB (F = 10.6, P <0.05; איור 1D).
איור 1
עכברים overexpressing ΔFosB הציג סמנים ביוכימיים של דופמין מופחתת איתות ב NAC
צריכת מזון ומשקל הגוף על דיאטה עשירה בשומן
לאחר מכן בחנו את ההשפעות של דיאטת HF מתגמלת באופן טבעי על שינויים במולקולות איתות בעכברים המבטאים יתר על המידה ΔFosB. לא היו הבדלים בין עכברי ΔFosB לבין בקרת צריכת המזון בבית או ב- HF. עם זאת, חלה ירידה כוללת בצריכת הקלוריות מנורמלת למשקל הגוף כאשר נחשפה ל- HF שהייתה ספציפית לעכברי ΔFosB (F = 11.2, P <0.01; איור 2 א '). בתום שישה שבועות של חשיפה לדיאטה, עכברים שקיבלו HF שקלו באופן משמעותי יותר מאלו שנמצאו בדיאטת Chow (F = 17.2, P <0.001) ועכברי ΔFosB שקלו פחות באופן כללי מהבקרות (F = 5.6, P <0.05; איור 2 א). 0.05B). השפעה זו הייתה ספציפית להבדלים בין הקבוצות בדיאטת Chow (P <XNUMX).
איור 2
ΔFosB overexpressing עכברים לא הראו הבדלים בצריכה מזון על דיאטה או צ 'יף גבוה (HF) שומן
הבדלים ביוכימיים על דיאטה עשירה בשומן
כדי לקבוע כיצד ההבדלים הבסיסיים של איתות NAC עשויים להשתנות על ידי דיאטה HF, אותם חלבונים איתות למדנו בתחילת המחקר נבדקו אצל בעלי חיים שקיבלו שישה שבועות של HF. לא נמצאו הבדלים משמעותיים ברמות Cdk5 (איור 3A). רמות pCREB ו CREB הכולל כבר לא היו שונים לאחר שישה שבועות של HF (איור 3B, C). רמות ה- BDNF היו גבוהות משמעותית בעכברי FOSB לאחר שישה שבועות של חשיפה ל- HF (F = 6.5, P = 0.05, איור 3D).
איור 3
High שומן (HF) דיאטה ameliorated איתות הבדלים שנצפו NAC של ΔFosB overexpressing עכברים
קולטני דופאמין אוטורדיוגרפיה
השתמשנו באוטורודיוגרפיה של קולטן כדי להעריך אם השינויים הנגרמים על ידי ΔFosB באיתות הדופמין ב- NAc קשורים לשינויים בביטוי קולטן הדופמין (איור 4 א). נראה כי תזונה עשירה בשומן מגדילה מעט את צפיפות קשירת הקולטן לדופמין D1 (P = 0.14), והבדל זה היה גדול יותר בעכברי ΔFosB (איור 4 ב). הייתה גם מגמה של עלייה באזור הכריכה של D1 בעקבות HF (P = 0.06), ובדיקות פוסט הוק הראו כי זה היה משמעותי בעכברי ΔFosB (P <0.05; איור 4C). בניגוד לקולטנים D1, אין שינויים בצפיפות מחייב הקולטן D2 (בקרת chow = 97.6 ± 6.9, בקרת HF = 101.1 ± 8.2, ΔFosB chow = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) או אזור מחייב (chow control = 47.3 ± 3.4, בקרת HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) ב- NAc נצפו.
איור 4
דיאטה עשירה בשומן (HF) הובילה לשינויים ב- D1 דופאמין מחייב קולטן ביטוי DARPP-32 בגרעין accumbens (NAC) של ΔFosB overexpressing עכברים
ביטוי DARPP-32 ב NAC
הכלאה באתרו שימשה לקביעת רמות הביטוי של DARPP-32 ב- NAc (איור 4D). תזונה עשירה בשומן הגדילה משמעותית את הביטוי של DARPP-32 באזור מוח זה (F = 5.1, P <0.05), והיה אינטראקציה משמעותית בין דיאטה לביטוי ΔFosB (F = 8.9, P <0.05), כאשר עכברי ΔFosB הראו כי שינוי המושרה על ידי דיאטה (איור 4 ה). הבדל בסיסי בביטוי DARPP-32 בין שליטה לעכברי ΔFosB נחשף על ידי בדיקות פוסט הוק (P <0.01), כמו גם עלייה משמעותית בביטוי DARPP-32 בעכברי ΔFosB ב- HF (P <0.01).
ביטוי גנים ב- VTA
QRT-PCR שימש להערכת שינויים בביטוי גנים ב- VTA, ומתמקד בכמה גנים מרכזיים שהיו מעורבים בעבר בהסדרת התגמול. כל הדגימות נורמלו ל- β- אקטין. כדי להבטיח שביטוי β- אקטין לא שונה על ידי הטיפול, נערך בדיקה נפרדת להשוואת β-actin לבקרה פנימית שנייה, GAPDH. לא היו הבדלים משמעותיים בביטוי β- אקטין (ערכי ΔCT, β-actin - GAPDH: chow בקרה = 2.29 ± 0.21, בקרת HF = 2.01 ± 0.04, ΔFosB chow = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
נצפתה מגמה לאינטראקציה בין ביטוי ΔFosB וטיפול בדיאטה לביטוי של טירוזין הידרוקסילאז (F = 3.6, P <0.06; איור 5 א). נראה כי שישה שבועות של חשיפה ל- HF הפחיתו את הביטוי של טירוזין הידרוקסילאז בעכברי בקרה והגבירו את הביטוי בעכברי ΔFosB. אינטראקציה משמעותית בין ביטוי ΔFosB לבין חשיפה לתזונה נצפתה לביטוי טרנספורטר הדופמין (F = 6.7, P <0.03; איור 5B). בדומה לטירוזין הידרוקסילאז, חשיפה ל- HF הפחיתה את הביטוי של טרנספורטר הדופמין בעכברי בקרה והגדילה את הביטוי באופן משמעותי בעכברי ΔFosB (P <0.05). ההבדל הבסיסי בביטוי טרנספורטר דופמין בין שליטה לעכברי ΔFosB לא הגיע למשמעות (P = 0.16), אך לאחר 6 שבועות של HF, עכברי ΔFosB ביטאו רמות גבוהות משמעותית של טרנספורטר דופמין בהשוואה לבקרות (P <0.05).
איור 5
High דיאטה דיאטה (HF) החשיפה הביטוי ΔFosB הוביל לשינויים בביטוי של מספר מולקולות מפתח VTA
הייתה מגמה המצביעה על השפעה של ביטוי ΔFosB מוגבר להפחתת רמות TrkB ב- VTA (F = 5.7, P <0.04; איור 5C). למרות שלא היו השפעות עיקריות על ביטוי קולטן κ-אופיואידים, הייתה מגמה של ביטוי מופחת בעכברי ΔFosB (P = 0.08; איור 5D). ביטוי של קולטן הלפטין נקבע גם ב- VTA. נמצאה השפעה משמעותית של חשיפה לדיאטה (F = 6.1, P <0.03), כאשר HF הפחית משמעותית את רמות הקולטן ללפטין ב- VTA בשני עכברי ה- ΔFosB וגם בקבוצת הביקורת (איור 5E). נבדק גם ביטוי של קולטן אורקסין 1 ב- VTA. הייתה השפעה משמעותית של תזונה על ביטוי קולטן האורקסין (F = 9.0, P <0.02), כאשר עכברים שנחשפו ל- HF מביעים רמות גבוהות יותר ב- VTA (איור 5F). הייתה גם מגמה של עכברי ΔFosB לבטא רמות גבוהות יותר של קולטן אורקסין 1 באזור מוח זה (P <0.05).
ביטוי Orexin בהיפותלמוס לרוחב
מדדנו רמות של אורקסין בהיפותלמוס לרוחב, מקור העצבנות האורקסינרגית של ה- VTA, על ידי הכלאה באתרו (איור 6 א). הייתה אינטראקציה משמעותית בין ביטוי ΔFosB לבין חשיפה לדיאטה על ביטוי אורקסין (F = 9.1, P <0.01), כאשר HF הגדיל משמעותית את רמות האורקסין בעכברי בקרה (P <0.05) וירידה בביטוי בעכברי ΔFosB (איור 6 ב). למרות שלא היו הבדלים משמעותיים בביטוי האורקסין במצב הבסיסי, בעקבות 6 שבועות של HF, עכברי ΔFosB הביעו רמות מופחתות משמעותית של אורקסין בהשוואה לבקרות (P <0.05).
איור 6
לתזונה עשירה בשומן (HF) היו השפעות דיפרנציאליות על הביטוי אוקסין בשליטה (Ctrl) ו- ΔFosB overexpressing עכברים
Beניתוחי
כדי להעריך שינויים מעוררים ורגשיים עקב שינוי תזונתי, עכברים נחשפו למבחן השדה הפתוח 24 שעות לאחר נסיגת הדיאטה HF (10). סה"כ צלבי קו, אשר נקלעו כמדד לעוררות, הושפעו משמעותית מהביטוי ΔFosB (F = 6.6, P <0.05) ודיאטה (F = 4.6, P <0.05; איור 7 א). עכברי ΔFosB היו פעילים יותר בסביבה החדשה מאשר התכווצויות, ובדיקות פוסט הוק הראו כי עכברים שנסוגו מ- HF היו פעילים יותר באופן משמעותי מאלו שנחשפו ל- chow (P <0.05). בולי צואה נספרו כמדד להתנהגות כמו חרדה (10). הייתה השפעה עיקרית של ביטוי ΔFosB (F = 10.2, P <0.01), כאשר עכברים המבטאים יתר על המידה ΔFosB מייצרים יותר בולים צואתיים בסביבה החדשה, במיוחד בקבוצות הנסיגה של בית Chow ו- HF (איור 7 ב). עכברי ΔFosB שנשמרו בדיאטת HF ייצרו פחות בולי צואה מאלה שנשמרו ב- chow ואלה שנסוגו 24 שעות לפני הבדיקה. נראה כי עכברי בקרה לא הושפעו מהתזונה. לא היו השפעות משמעותיות של ביטוי ΔFosB או דיאטה על זמן השהייה במרכז השדה הפתוח (בקרת צ'או = 14.5 ± 3.1 שניות, בקרת HF = 18.0 ± 3.2 שניות, בקרת W / D = 15.4 ± 1.9 שניות, ΔFosB צ'או = 16.9 ± 2.4 שניות, ΔFosB HF = 13.1 ± 3.9 שניות, ΔFosB W / D = 19.8 ± 2.6 שניות).
איור 7
עכברים מעל הביטוי ΔFosB היו רגישים יותר להשפעות של דיאטה שומן גבוה (HF) הנסיגה
דיון
בטיפול בהשמנת יתר, יש צורך קריטי לזיהוי גורמים המשפיעים על הרגישות לאכילת יתר ולעליה במשקל. מסלולי תגמול המוח ממלאים תפקיד חשוב במוטיבציה לתגובות מזון ושינויים תזונתיים (6, 10, 15, 16). כאשר אותות אורקסגניים ואנורקסגניים יכולים להשפיע באופן ישיר על תגמול גמול באמצעות מעגל היפותלמוס- VTA-NAc, הבהרת גנים המגיבים לתזונה עשירה בטעמי אנרגיה במרכזי גמול עשויה לספק מטרות טיפוליות חדשות בטיפול בהשמנת יתר (17, 18). לכן, בדקנו סמנים ביוכימיים ומולקולריים של תגמול ואיזון אנרגיה, לאורך מעגל ההיפותלמוס-VTA-NAc בתגובה לתזונה של HF ב- ΔFosB - overexpressing עכברים כמודל של רגישות משופרת לשינויים בתגמול (13, 19, 20) , ואת הרגישות ההתנהגותית הבאה דיאטה הנסיגה. שיערנו כי דיסרגולציה בסיסית של איתות דופאמין בעכברי ΔFosB תתבצע מנורמל על ידי ההשפעות המתגמלות של דיאטה HF, המקיפה את הצטלבות של סימני איזון האנרגיה ומערכת הדופמין.
כדי לבחון סמנים המעידים על dysregulation ב דופאמין איתות ב NAC, בחנו רמות קולטן D1 ומפעילי downstream. למרות שלא היו הבדלים משמעותיים מחייב קולטן D1, הייתה מגמה של חשיפת HF להגדלת האזור המחייב בעכברי ΔFosB. זה מעניין כמו אינדוקציה של ΔFosB על ידי סמים ותגמולים טבעי נראה להשתלט על תת סוג חיובי dynorphin של נוירונים קוצני בינוני המבטאים בעיקר קולטני D1 (9, 21). רמות של dPamine במורד הזרם איתות pCREB היו מופחת באופן משמעותי בעכברים ΔFosB, תומכת של הפעלת קולטן D1 מופחתת באזור זה במוח (22, 23). מעניין, אנו גם זיהה ירידה משמעותית ברמות CREB הכולל בעכברים ΔFosB, דבר המצביע על יכולת מופחתת נוספת עבור התמרת האות דופאמין כי עשוי להיות משני למשוב הנובע ירידה מתמשכת pCREB (24). הביטוי BDNF מוסדר על ידי pCREB, הוא מורם עם הפעלת D1, והוא מתווך חשוב של נוירופלסטיות הקשורות לתגמל ב NAC (25, 26). לפיכך, זיהינו ירידה משמעותית בחלבון BDNF ב NAC של עכברים ΔFosB.
כל נוירונים בינוני קוצני ב NAC Express DARPP-32 (27). המנגנונים הרבים שלה במורד הזרם הופכים אותה לשחקן חיוני במסלולי גמול (28), והיא מעורבת בהתמכרות לסמים ובהפרעות אחרות המעורבות במערכת הדופאמין, כולל הפרעות רגשיות וסכיזופרניה (27, 29). זיהינו ירידה בסיסית עמוקה בביטוי DARPP-32 ב NAC של עכברים ΔFosB. ביטוי DARPP-32 מוסדר על ידי BDNF, ולכן הביטוי המופחת עשוי להיות קשור ישירות להפחתות ברמות BDNF שזוהו בעכברי FOSB Δ (27, 29, 30). גם שינויים מתונים במצב זרחון של DARPP-32 יכול להוביל שינויים משמעותיים איתות תאיים בתוך NAC (27). מחקרים קודמים דיווחו על שום שינוי בחלבון DARPP-32 בעכברי ΔFosB בעקבות הסרת 12-wk מ- doxycycline כאשר בוצעה הערכה סטרלית רחבה יותר (31) מה שמרמז כי ההשפעות של ΔFosB על DARPP-32 עשוי להיות זמן אזור ספציפי.
הנחנו כי הירידות הדרמטיות במדדי דופאמין איתות ב NAC של עכברים ΔFosB כנראה מעורב שינויים נוירונים הקרנה VTA דופאמין, למרות ΔFOSB אינו overexpressed בתוך נוירונים אלה. לכן, בדקנו ביטוי של גנים הקשורים דופמין ב VTA, כולל tyrosine hydroxylase ו טרנספורטר דופמין. רמות של tyrosine hydroxylase ו דופמין טרנספורטר הם בקורלציה חיובית עם הפלט דופמין. היתה מגמה של עכברים ΔFosB להפגין מופחת טירוזין hydroxylase ו ירידה משמעותית של טרנספורטר דופמין, בהתאם dysregulation של דופאמין איתות ב NAC. כמו אלה הפחתות בסיסיות גנים הקשורים דופמין ב VTA של עכברים ΔFosB משוער משקפים משופרת מן NAC במהלך ביטוי לטווח ארוך ΔFosB overexpression, בחנו את הביטוי של קולטן BDNF, TrkB, כמנגנון אפשרי של משוב NAC ל- VTA (32). בדומה tyrosine hydroxylase ו טרנספורטר דופמין, ביטוי TrkB הראה גם מגמה להיות מופחת בעצם בעכברים ΔFosB כי לא הגיעו משמעות כאשר תוקנו עבור השוואות מרובות. מתחם BDNF-TrkB יכול להיות מועבר retrogradely ולפעול בתוך VTA כדי להשפיע על ביטוי הגן המקומי ולקדם צמיחה התא ותחזוקה (33). יתר על כן, הפעלת BDNF של trkb presynaptic בתוך NAC יכול ישירות לעורר דופאמין נוירוטרנסמינציה (32), התומכת בירידה הבסיסית של איתות דופמין בעכברים אלה.
הפעלת Dynorphin של קולטנים κ-opioid מווסתים דופאמין איתות הוא מנגנון נוסף שבו NAC מספק משוב VTA (34). מצאנו כי ביטוי קולטן κ-opioid ב VTA הראה מגמה להיות מופחת עכברים ΔFosB. כפי overexpression ΔFosB הוצגה כדי להקטין את הביטוי dynorphin ב NAC (20), עכברים ΔFosB כנראה יש הפחתות עמוקות נטו VTA κ-הפעלת אופיואידים. למרות שאיתות דינורפין מפעיל בדרך כלל השפעה מעכבת על נוירונים של דופמין (35), חולדות המראות על ניהול עצמי משופר של סמים של התעללות מציגות רמות מופחתות של דינורפין ב- NAc, ומצביעות על תפקיד של איתות דינורפין מופחת באופן בסיסי בשיפור רגישות התגמול (36). , 37). ויסות דיסיסטרי של מערכת הדינורפין - κ-אופיואידים נקשר לרכישה ולהתמדה של שימוש בסמים, ותומך באיזון קריטי של איתות אופיואידים בנורמליזציה של מסלולי הדופמין. (38).
בהתבסס על היכולת הגמישה של דיאטה HF צפופה באנרגיה, שיערנו כי דיסרגולציה בדופאמין ובאופיויד גמול איתות בעכברי ΔFosB יניע את העכברים האלה לתגמול תגמול משופר לתזונה שכזו, ובכך לנרמל את מערכת התגמולים באמצעות הפעלת ההיפותלמוס מעגל VTA-NAC. במהלך החשיפה לשבוע של שישה שבועות, לא נצפו הבדלים בצריכת המזון בין ΔFOSB לבין עכברי בקרה, דבר המצביע על כך שהשינויים שנמצאו בסמנים ביוכימיים ומולקולריים של אותות תגמול בעכברי Δ FOSB לא נבחנו מהבדלים בקלוריות הנצרכות. כצפוי, הבדלים ביזליים שהתגלו ב- pCREB, CREB הכולל, BDNF, DARPP-32 ורמות קולטנים מסוג κ-opioid בין ΔFOSB ועכברי בקרה נחלשו, ככל הנראה עקב עלייה בפלט הדופמין בעכברי FOSB ב- HF (29, 39-41) .
בחינה של שני hydroxylase טירוזין ו טרנספורטר דופמין ב VTA חשף תגובות מנוגדות מפתיע של ΔFOSB ועכברים שליטה בעקבות HF. עכברים שליטה הראו ירידה בטירוזין הידרוקסילאז ביטוי דופאמין טרנספורטר, בעוד עכברים ΔFosB הראה ביטוי מוגבר של שני גנים אלה הקשורים דופמין. מעניין, ביטוי tyrosine hydroxylase הוא השתנה VTA ידי קוקאין כרונית או מתאמפטמין הממשל (42-44), מה שמרמז כי עכברים ΔFosB עשוי למצוא את הפרס הטבעי של HF בולט יותר עכברים שליטה.
על מנת לבחון כיצד קלט היפותלמי פוטנציאלי ל - VTA עשוי להיות ממסר אותות המשקפים את מאזן האנרגיה, נבדקו גם ביטוי לקולטן לפטין וקולטן אורקסין - 1. רמות הלפטין במחזור מוגברות על ידי HF, ולפטין יכול בתורו לפעול ב- VTA כדי לשנות את אותות הדופאמין (18, 45). ביטוי VPT לקטין קולטן ירד באופן דומה על ידי HF הן ΔFOSB ועכברים שליטה, בהתאם לעלייה במשקל דומה צריכת הדיאטה בעוד על HF. שומן גבוה גם ביטוי מוגבר של קולטן אורקסין- 1 ב VTA של שניהם ΔFOSB ועכברים שליטה. Orexin מפעיל נוירונים דופאמין VTA, מקדם פלסטיות VTA, ומגביר את רמות הדופמין ב NAC (46-48). דיאטה עשירה בשומן הוכח להגברת ביטוי האורקסין בעכברים, בהתאם לתצפיות שלנו (49, 50). לפיכך, ביטוי מוגבר של קולטן אורקסין, כמו גם שינויים LPTIN איתות VTA יכול לקדם את הפרס לתגמל הן ΔFOSB ועכברים שליטה, תמיכה דיסוציאציה בין מסלולים ממסר אותות מאזן האנרגיה לבין אלה קשורה ישירות לתגמל.
כדי לבחון את ההשפעות המעוררות מתח של משיכת תגמול, נבדקו עכברים במבחן שדה פתוח 24 שעות לאחר הסרת HF. העכברים ΔFosB היו רגישים יותר להשפעות החריפות של דיאטת הדיאטה המועדפת, אשר הראו פעילות עוררות מוגברת ופולי בצואה בזק הפתוח בהשוואה לכל קבוצות הביקורת והדיאטה האחרות. עכברי ΔFosB הראו גם דפוס התנהגותי מעניין במבחן זה המרמז על תגמול ורגישות של מתח, עם דיאטה HF בתחילה הפחתת ייצור boli צואה יחסית chow, ואת הנסיגה שוב להגדיל את התגובה קשורה חרדה. עלייה זו נצפתה בפעילות בשטח הפתוח לא התאימה לשינויים בביטוי אורקסין, דבר המצביע על קשר עם עוררות המושרה על ידי מתח, שאינה רק השפעה של שינויים באיתות של אורקסין בתיווך. בסך הכל, נתונים אלה תומכים ההשערה שלנו כי עכברי ΔFosB יהיה רגיש יותר את ההשפעות החריפות של הנסיגה דיאטה המועדפת בשל רגישות תגמול מוגברת שלהם.
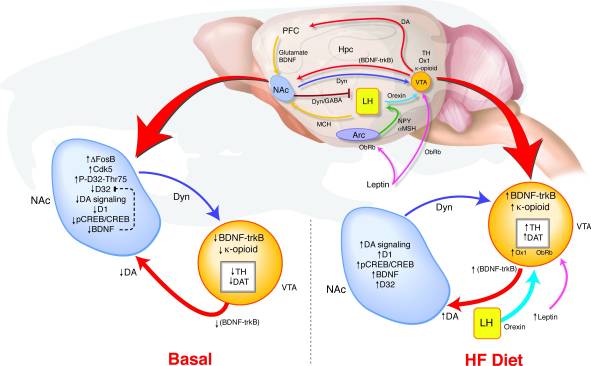
כיצד overexpression לטווח ארוך של ΔFosB ב NAC להוביל לשינויים כאלה התנהגות איתות גמול? אנו מציעים מודל של זיהוי VTA במקרה שבו משוב שונה מ NAC ו hypothalamus ממסר אותות לגבי המדינה לתגמל לקבוע את הרגולציה של מערכת דופמין שעשויים לתמוך קישור בין דיסרגולציה מסלול תגמול נטייה ההשמנה (איור 8). במהלך חשיפה HF, תשומות מרובות המשקף הן מאזן האנרגיה ואת המדינה לתגמל להתכנס על VTA. עליה בסימפטומים של לפטין ואורקסין, כמו גם שינוי משוב מה- NAC להיפותלמוס לרוחב, עשויה להשפיע על האופן שבו אותות אלה אוקסגניים מגיבים ל- HF בעכברי ΔFosB (17, 18, 45, 47, 51-53). העלאת רמות גבוהות של דיאטה בדם ב- BDNF עשויה לספק משוב לתגמול ה- VTA, ובכך לקדם את השינויים בביטוי גנים הקשור לדופאמין.
איור 8
דיאטה עשירה בשומן (HF) מנרמל את איתות הגמול dysregulated בעכברי FOSB Δ
תוצאות אלה מסמנות סמנים מולקולאריים של רגישות גמול ומראים כי dysregulation לטווח ארוך של מערכת דופמין עשוי להשפיע על הפרט להתמכרות והשמנת יתר. יתר על כן, נתונים אלה מספקים צעד חשוב לקראת איתור מטרות טיפוליות פוטנציאליות חדשות בטיפול ומניעה של השמנת יתר והפרעות אחרות שעלולות להתרכז במערכת התגמול. בעתיד, חשוב לחקור כיצד מערכת זו מגיבה להסרת דיאטה HF, כמו גם לחקור כל ההבדלים בין המינים ברגישות לתגמל וחשיפה גבוהה שומן דיאטה.
חומר משלים
Supp. שיטות
לחץ כאן לצפייה (61K, doc)
תודות
המחברים מבקשים להודות לקאתי סטפן על סיוע עם גידול בעלי חיים והעברתם. עבודה זו נתמכה על ידי מענק מאוניברסיטת פנסילבניה סוכרת מרכז (DK019525) ועל ידי מענקים מהמכון הלאומי לבריאות הנפש (R01 MH51399 ו P50 MH66172) ואת המכון הלאומי על שימוש בסמים (R01 DA07359).
הערות שוליים
גילוי כלכלי: כל המחברים מצהירים כי אין להם אינטרסים פיננסיים ביו-רפואיים או ניגודי אינטרסים פוטנציאליים.
הפניות
1. Wadden ת"א, ברקוביץ RI, Womble LG, Sarwer DB, Phelan S, קאטו RK, Hesson LA, Osei SY, קפלן R, Stankard AJ. ניסוי אקראי של שינוי באורח החיים ופרמקותרפיה להשמנה. N אנגל J מד. 2005; 353 (20): 2111-20. [PubMed]
2. Blendery JA, Strasser A, וולטרס CL, פרקינס KA, Patterson F, Berkowitz R, לרמן ג מופחת גמול ניקוטין בהשמנת יתר: השוואה הדדית האדם ועכבר. פסיכופארמקולוגיה (ברל) 2005
3. הפרעות IH, Muris P ההבדלים האישיים ברגישות הגמול קשורים למאוות מזון ומשקל גוף יחסי אצל נשים בריאות. תיאבון. 2005; 45 (2): 198-201. [PubMed]
4. Kelley AE, ברידג 'KC. מדעי המוח של תגמולים טבעיים: רלוונטיות לתרופות ממכרות. J Neurosci. 2002; 22 (9): 3306-11. [PubMed]
5. Cagniard B, Balsam PD, ברונר D, Zhuang X. עכברים עם עלייה כרונית מוגברת דופמין התעצמות מוטיבציה מוגברת, אבל לא למידה, עבור פרס מזון. Neuropsychopharmacology. 2006; 31 (7): 1362-70. [PubMed]
6. ליאנג NC, Hajnal A, Norgren ר ש 'האכלה שמן תירס מגביר דוממין accumbens בעכברוש. Am J פיסיול רגול אינטגר קומפי Physiol. 2006; 291 (5): R1236-9. [PubMed]
7. מנדוזה J, אנגלס קסטלאנוס M, Escobar C. Entrainment על ידי ארוחה טעים גורם לפעילות הצפויה למזון c-Fos הביטוי באזורים הקשורים לגמול של המוח. מדעי המוח. 2005; 133 (1): 293-303. [PubMed]
8. שרדר בי, בינזאק JM, קלי AE. פרופיל נפוץ של הפעלת קליפת המוח הפריפרונטלית לאחר החשיפה לרמזים הקשריים בניקוטין או שוקולד. מדעי המוח. 2001; 105 (3): 535-45. [PubMed]
9. נסלר EJ, Barrot M, עצמי DW. DeltaFosB: מתג מולקולרית מתמשכת להתמכרות. Proc Natl Acad Sci US A. 2001, 98 (20): 11042-6. [מאמר חינם PMC] [PubMed]
10. Teegarden SL, Bale TL. ירידה בהעדפות תזונתיים מייצרת רגישות מוגברת וסיכון לנסיגה בתזונה. ביול פסיכיאטריה. 2007; 61 (9): 1021-9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, טיילור JR. dFosB ב Nucleus Accumbens מסדיר מזון מחוזק התנהגות אינסטרומנטלית מוטיבציה. כתב העת של מדעי המוח. 2006; 26 (36): 9196-9204. [PubMed]
12. חן J, קלץ MB, זנג G, Sakai N, סטפן C, שוקט PE, Picciotto MR, Duman RS, Nestler EJ. חיות טרנסגניות עם ביטוי גנטי, ממוקד, במוח. מול פרמקול. 1998; 54 (3): 495-503. [PubMed]
13. קלוז, ג 'רדן L, Beckmann AM, סטפן C, ג' אנג YJ, Marotti L, עצמי DW, Tkatch T, Baranauskas G, Surmeier DJ, נווה RL, Duman RS, Picciotto MR, נסלר EJ. ביטוי של גורם שעתוק deltaFosB במוח שולט רגישות קוקאין. הטבע. 1999; 401 (6750): 272-6. [PubMed]
14. בייל TL, Dorsa DM. הבדלי מין והשפעות של אסטרוגן על קולטן אוקסיטוצין שליח הביטוי חומצה ריבונוקלאית ההיפותלמוס ventromedial. אנדוקרינולוגיה. 1995; 136 (1): 27-32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. עכברים תלויים בסוכר מראים תגובה משופרת לסוכר לאחר התנזרות: עדות להשפעות של מניעת סוכר. פיסיול בהב. 2005; 84 (3): 359-62. [PubMed]
16. האם MJ, Franzblau EB, קלי AE. גרעין אקומבנס מו-אופיואידים מווסת את צריכת הדיאטה דלת שומן באמצעות הפעלת רשת מוח מבוזרת. J Neurosci. 2003; 23 (7): 2882-8. [PubMed]
17. ג 'נג H, פטרסון LM, Berthoud HR. Orexin איתות באזור הטגנטלית הגחון נדרש לתיאבון בשומן גבוה המושרה על ידי גירוי אופיואידים של הגרעין accumbens. J Neurosci. 2007; 27 (41): 11075-82. [PubMed]
18. Hommel JD, Trinko R, סירס רם, Georgescu D, ליו ZW, גאו XB, Thurmon JJ, מרינלי M, DiLeone RJ. קולטן לפטין איתות נוירונים דופאמין המוח התיכון מווסת האכלה. נוירון. 2006; 51 (6): 801-10. [PubMed]
19. קולבי קר, ויסלר ק', סטפן סי, נסלר אי-ג'יי, עצמי ד"ו. סוג תא ספציפי של Overexion של DeltaFosB משפר תמריץ לקוקאין. J Neurosci. 2003; 23 (6): 2488-93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, קאסידי MP, קלץ MB, שו- Lutchman T, ברטון O, סים סלי LJ, Dileone RJ, קומאר א, Nestler EJ. תפקיד חיוני DeltaFosB בגרעין accumbens בפעולה מורפיום. Nat Neurosci. 2006; 9 (2): 205-11. [PubMed]
21. לי KW, קים Y, קים AM, הלמין K, Nairn AC, גרינגרד P. קוקאין המושרה השדרה הדנדריטים הדגימה D1 ו D2 דופאמין דופאמין המכילים נוירונים קוצניים בינוניים בגרעין accumbens. Proc Natl Acad Sci US A. 2006, 103 (9): 3399-404. [מאמר חינם PMC] [PubMed]
22. Blendy JA, Maldonado R. ניתוח גנטי של התמכרות לסמים: תפקידו של רכיב התגובה cAMP חלבון מחייב. J מולד. 1998; 76 (2): 104-10. [PubMed]
23. נסלר EJ. מנגנונים מולקולריים של התמכרות לסמים. נוירופרמקולוגיה. 2004; 47 1: 24-32. [PubMed]
24. Tanis KQ, Duman RS, ניוטון אס. CREB עיקול ופעילות במוח: ספציפיות אזורית והשראה ע"י התקף אלקטרו-פולסיבי. ביול פסיכיאטריה. 2007
25. קומאר א, Choi KH, רנטאל W, Tsankova NM, Theobald DE, Truong HT, רוסו SJ, Laplant Q, סאסאקי TS, ויסלר KN, נווה RL, עצמי DW, Nestler EJ. שיפוץ הכרומטין הוא מנגנון מפתח בבסיס הפלסטיות המושרה בקוקאין בסטריאטום. נוירון. 2005; 48 (2): 303-14. [PubMed]
26. גרהם DL, אדוארדס S, Bachtell RK, Dileone RJ, Rios M, עצמי DW. פעילות BDNF דינמית בגרעין אקומבנס עם שימוש בקוקאין מגבירה את הניהול העצמי וההישנות. Nat Neurosci. 2007; 10 (8): 1029-37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 מתווך את הפעולות של סמים מרובים של התעללות. APS J. 2005; 7 (2): E353-60. [מאמר חינם PMC] [PubMed]
28. פאלמר AA, Verbitsky M, Suresh R, Kamens HM, ריד CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, פיליפס TJ. הבדל ביטוי גנים בעכברים שנבחרו נבדק עבור רגישות מתאמפטמין. הגנום. 2005; 16 (5): 291-305. [PubMed]
29. בוגוש A, פדריני S, Pelta-Heller J, Chan T, יאנג ש, מאו Z, Sluzas E, Gieringer T, Ehrlich ME. AKT ו CDK5 / p35 מתווך המוח נגזר גורם neurotrophic אינדוקציה של DARPP-32 נוירונים קוצניים בגודל בינוני במבחנה. J ביול כימית. 2007; 282 (10): 7352-9. [PubMed]
30. Benavides DR, ביב JA. תפקידה של Cdk5 בשימוש בסמים ובפלסטיות. אן אן אקאד Sci. 2004; 1025: 335-44. [PubMed]
31. ביב ג 'א, צ' ין J, Svenningsson P, Nishi A, Snyder GL, יאן Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard פ ההשפעות של חשיפה כרונית קוקאין מוסדר על ידי חלבון נוירונים Cdk5. הטבע. 2001; 410 (6826): 376-80. [PubMed]
32. Blochl A, Sirrenberg C. Neurotrophins לעורר את שחרורו של דופמין מן העכברים mesencephalic נוירונים באמצעות Trk ו קולטנים p75Lntr. J ביול כימית. 1996; 271 (35): 21100-7. [PubMed]
33. ברטון O, McClung CA, Dileone RJ, קרישנן V, R Rento W, Russo SJ, גרהם D, Tsankova NM, Bolanos CA, ריוס M, מונטגיה LM, עצמי DW, Nestler EJ. תפקיד חיוני של BDNF במסלול dopamine mesolimbic הלחץ החברתי להביס. מדע. 2006; 311 (5762): 864-8. [PubMed]
34. נסלר EJ, קרלזון WA., ג 'וניור מעגל גמול דופמין mesolimbic בדיכאון. ביול פסיכיאטריה. 2006; 59 (12): 1151-9. [PubMed]
35. פורד CP, Beckstead MJ, וויליאמס JT. קאפה אופיואיד עיכוב של דופאמין somatodendritic מעכב זרמים postsynaptic. J Neurophysiol. 2007; 97 (1): 883-91. [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Brain dynorphin ו- enkephalin במעכבי פישר ולואיס: השפעות של סובלנות ומורפיום של מורפיום. מוח מוחי. 1995; 683 (1): 25-35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. ההבדלים בין אלכוהול מועדף (AA) ו-הימנעות אלכוהול (ANA) חולדות במערכות prodynorphin ו proenkealin. אלכוהול Clin Exp. 1994; 18 (5): 1272-9. [PubMed]
38. קריק MJ. קוקאין, דופמין ומערכת האופיואידים האנדוגניים. מכור J. 1996; 15 (4): 73-96. [PubMed]
39. Carlezon WA, ג 'וניור, Duman RS, Nestler EJ. הפנים הרבים של CREB. מגמות Neurosci. 2005; 28 (8): 436-45. [PubMed]
40. Dudman JT, איטון ME, ראג 'ייקשה A, Macias W, טאהר M, Barczak A, Kameyama K, Huganir R, Konradi C. Dopamine קולטנים D1 לתווך זרחון CREB באמצעות זרחון של קולטן NMDA ב Ser897-NR1. ניורוצ'ם. 2003; 87 (4): 922-34. [PubMed]
41. DW עצמי. תקנה של נטילת סמים ו - Seeking התנהגויות על ידי neuroadaptations במערכת mopolimbic dopamine. נוירופרמקולוגיה. 2004; 47 1: 242-55. [PubMed]
42. ביתר ג 'ונסון D, נסלר EJ. מורפין וקוקאין מפעילים פעולות כרוניות שכיחות על טירוזין הידרוקסילאז באזורי הגמילה של המוח הדופאמינרגי. ניורוצ'ם. 1991; 57 (1): 344-7. [PubMed]
43. Lu L, ג 'ים ג' יי, שחם Y, התקווה BT. מולקולרית neuroadaptations ב accumbens ואת אזור tegmental הגחון במהלך XNXX הימים הראשונים של התנזרות מאולצת של קוקאין עצמי הממשל בחולדות. ניורוצ'ם. 90; 2003 (85): 6-1604. [PubMed]
44. Shepard JD, Chuang DT, Shaham Y, Morales M. השפעת methamphetamine על הממשל העצמי של tyrosine hydroxylase ורמות טרנספורטר דופאמין במסופולימי ובכדורי דופאמין nigrostriatal של החולדה. Psychopharmacology (ברל) 2006, 185 (4): 505-13. [PubMed]
45. Fulton S, Pissios P, מנצ'ון RP, Stiles L, פרנק L, Pothos EN, Maratos-Flier E, פלייר JS. Leptin תקנה של מסלול דופמין mesoaccumbens. נוירון. 2006; 51 (6): 811-22. [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, סוזוקי T. השלכות של חלבון קינאז C ב oexin- המושרה מעלה של רמות דופמין תאיים ואת ההשפעה שלה מתגמל. Eur J Neurosci. 2007; 25 (5): 1537-45. [PubMed]
47. Narita M, Nagumo Y, Hhotimo S, Khotib J, Miyatake M, סאקאראי T, Yanagisawa M, Nakamachi T, Shioda S, סוזוקי T. מעורבות ישירה של מערכות אוקססינרגיות בהפעלת מסלול דופמין mesolimbic והתנהגויות הקשורות המושרה על ידי מורפין. J Neurosci. 2006; 26 (2): 398-405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, שדות HL, Bonci א Orexin ב VTA הוא קריטי עבור אינדוקציה של גמישות סינפטי רגישות התנהגותית קוקאין. נוירון. 2006; 49 (4): 589-601. [PubMed]
49. פארק ES, יי SJ, קים JS, לי HS, Lee IS, סונג JK, ג 'ין HK, Yoon YS. שינויים ב- orexin-A ו- neuropeptide Y ביטוי בהיפותלמוס של דיאטה בצום ובדיאטה עשירה. J Vet Sci. 2004; 5 (4): 295-302. [PubMed]
50. וורטלי KE, צ 'אנג GQ, Davydova Z, ליבוביץ SF. פפטידים המווסתים את צריכת המזון: ביטוי גנים של אורקסין גדל במהלך מצבים של hypertriglyceridemia. Am J פיסיול רגול אינטגר קומפי Physiol. 2003; 284 (6): R1454-65. [PubMed]
51. ג 'נג H, Corkern M, Stoyanova אני, Patterson LM, טיאן R, ברת'וד HR. פפטידים המווסתים את צריכת המזון: תיאבון המפעיל מניפולציה של אקומבנס מפעיל נוירונים אוקססין ההיפותלמיים ומעכב נוירונים של POMC. Am J פיסיול רגול אינטגר קומפי Physiol. 2003; 284 (6): R1436-44. [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, דניאל RA, Landry CF, KEley AE. הפעלת תת-עיכול של נוירונים היפותלמיים המכילים אורקסין / hypocretin, על ידי עיכוב של קולטן GABAA בתיווך של פגז הגרעין, אך לא על ידי חשיפה לסביבה חדשה. Eur J Neurosci. 2004; 19 (2): 376-86. [PubMed]
53. האריס GC, Wimmer M, אסטון ג 'ונס ג' תפקיד של נוירונים hypexalamic ההיפותלמיים בחיפוש גמול. הטבע. 2005; 437 (7058): 556-9. [PubMed]