ការពិភាក្សាគ្លីនិក Neurosci ។ 2013 Dec;15(4):431-43.
អរូបី
ទោះបីជាមានសារៈសំខាន់នៃកត្តាចិត្តសាស្ត្រជាច្រើនក៏ដោយវានៅតែជាកត្តាញៀនគ្រឿងញៀនពាក់ព័ន្ធនឹងដំណើរការជីវសាស្រ្តៈសមត្ថភាពនៃការប៉ះពាល់ម្តងហើយម្តងទៀតចំពោះគ្រឿងញៀននៃការរំលោភបំពានដើម្បីនាំឱ្យមានការផ្លាស់ប្តូរនៅក្នុងខួរក្បាលងាយរងគ្រោះដែលជំរុញឱ្យមានការស្វែងរកនិងការប្រើថ្នាំនិងការបាត់បង់ការគ្រប់គ្រង។ លើការប្រើប្រាស់គ្រឿងញៀនដែលកំណត់ស្ថានភាពញៀន។ នៅទីនេះយើងពិនិត្យឡើងវិញនូវប្រភេទនៃការបន្សាំនៃម៉ូលេគុលនិងកោសិកាដែលកើតឡើងនៅក្នុងតំបន់ខួរក្បាលជាក់លាក់ដើម្បីសម្របសម្រួលភាពមិនប្រក្រតីនៃអាកប្បកិរិយាដែលទាក់ទងនឹងការញៀន។ ទាំងនេះរួមបញ្ចូលទាំងការផ្លាស់ប្តូរនៅក្នុងហ្សែនហ្សែនដែលទទួលបានមួយផ្នែកតាមរយៈយន្ដការខាងហ្សែនហ្សែនប្លាស្ទិកក្នុងមុខងារប្រព័ន្ធប្រសាទប្រសាទនៃប្រសាទនិងប្រសាទនិងប្លាស្ទិចដែលជាប់ទាក់ទងក្នុងប្រសាទប្រសាទនិងប្រសាទត្រូវបានសម្របសម្រួលមួយផ្នែកដោយការផ្លាស់ប្តូរកត្តាសរសៃប្រសាទ។ ប្រភេទនីមួយៗនៃការកែប្រែគ្រឿងញៀនអាចត្រូវបានគេមើលឃើញថាជាទម្រង់នៃ“ ការចងចាំកោសិកាឬម៉ូលេគុល” ។ លើសពីនេះទៅទៀតវាគួរឱ្យកត់សម្គាល់ថាទម្រង់ប្លាស្ទិកដែលទាក់ទងនឹងការញៀនភាគច្រើនស្រដៀងនឹងប្រភេទនៃប្លាស្ទិកដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងទម្រង់បុរាណនៃ“ ការចងចាំអាកប្បកិរិយា” ប្រហែលជាឆ្លុះបញ្ចាំងពីផលប៉ះពាល់តិចតួចនៃយន្តការបន្សាំដែលមានចំពោះណឺរ៉ូននៅពេលប្រឈមនឹងបរិស្ថាន។ បញ្ហាប្រឈម។ ទីបំផុតការសម្របខ្លួនម៉ូលេគុលនិងកោសិកាទាក់ទងនឹងការញៀនពាក់ព័ន្ធនឹងតំបន់ខួរក្បាលភាគច្រើនដែលសម្របសម្រួលការចងចាំបែបបុរាណកាន់តែច្រើនស្របនឹងទស្សនៈថាការចងចាំមិនធម្មតាគឺជាកត្តាជំរុញសំខាន់នៃរោគសញ្ញាញៀន។ គោលដៅនៃការសិក្សាទាំងនេះដែលមានគោលបំណងដើម្បីពង្រីកមូលដ្ឋានម៉ូលេគុលនិងកោសិកានៃការញៀនគ្រឿងញៀនគឺដើម្បីអភិវឌ្ឍការធ្វើរោគវិនិច្ឆ័យផ្អែកលើជីវសាស្ត្រក៏ដូចជាការព្យាបាលដែលមានប្រសិទ្ធភាពជាងមុនសម្រាប់ជំងឺញៀន។
សេចក្តីផ្តើម
ការញៀនគ្រឿងញៀនដែលអាចត្រូវបានកំណត់ថាជាការស្វែងរកនិងប្រើប្រាស់គ្រឿងញៀនទោះបីជាមានផលវិបាកគួរឱ្យរន្ធត់ឬបាត់បង់ការគ្រប់គ្រងលើការប្រើប្រាស់គ្រឿងញៀនក៏ដោយគឺបណ្តាលមកពីការផ្លាស់ប្តូរថ្នាំដែលមានរយៈពេលយូរដែលកើតឡើងនៅក្នុងតំបន់ខួរក្បាលជាក់លាក់។1 ទោះយ៉ាងណាមានតែបុគ្គលខ្លះទេដែលញៀននឹងការប្រឈមមុខនឹងការប្រើថ្នាំម្តងហើយម្តងទៀតខណៈដែលអ្នកផ្សេងទៀតមានសមត្ថភាពប្រើថ្នាំធម្មតានិងគេចផុតពីរោគសញ្ញាញៀន។ កត្តាហ្សែនមានចំនួនប្រហែល 50% នៃភាពខុសគ្នានៃភាពខុសគ្នានៃភាពងាយរងគ្រោះនៃការញៀននេះហើយកំរិតនៃភាពចាស់ជរានេះមានភាពស្មោះត្រង់ចំពោះក្រុមថ្នាំញៀនសំខាន់ៗទាំងអស់រួមមានថ្នាំរំញោចអាភៀនអាល់កុលនីកូទីននិងថ្នាំ cannabinoids ។.2 វាមិនទាន់អាចកំណត់អត្តសញ្ញាណហ្សែនភាគច្រើនដែលមានហានិភ័យហ្សែននេះទេដោយសារតែការចូលរួមនៃការប្រែប្រួលហ្សែនប្រហែលជារាប់រយដែលត្រូវបានបូកបញ្ចូលក្នុងបុគ្គលតែមួយដើម្បីបង្ហាញពីភាពងាយរងគ្រោះនៃការញៀន (ឬចំពោះបុគ្គលដទៃទៀតការតស៊ូ) ។
ហានិភ័យ ៥០ ភាគរយទៀតនៃការញៀនគឺមកពីកត្តាបរិស្ថានដែលកើតឡើងពេញមួយជីវិតដែលមានទំនាក់ទំនងជាមួយសមាសធាតុហ្សែនរបស់មនុស្សម្នាក់ៗដែលអាចធ្វើឱ្យគាត់ឬនាងងាយនឹងញៀនក្នុងកម្រិតធំជាងឬតិចជាងនេះ។ កត្តាបរិស្ថានជាច្រើនប្រភេទត្រូវបានបង្កប់ក្នុងការញៀនរាប់បញ្ចូលទាំងស្ត្រេសផ្នែកចិត្តសាស្ត្រប៉ុន្តែកត្តាដែលមានឥទ្ធិពលបំផុតគឺការប៉ះពាល់ទៅនឹងគ្រឿងញៀននៃការរំលោភបំពានខ្លួនឯង។ ថ្នាំ“ ក្លោងទ្វារ” មួយចំនួនជាពិសេសជាតិនីកូទីនត្រូវបានបង្ហាញថាបង្កើនភាពងាយរងគ្រោះរបស់មនុស្សម្នាក់ចំពោះការញៀននឹងថ្នាំមួយទៀត។3 លើសពីនេះទៅទៀតមានភ័ស្តុតាងកើនឡើងថាទោះបីជាមានហានិភ័យហ្សែនជាច្រើនសម្រាប់ការញៀននៅទូទាំងប្រជាជនក៏ដោយការប៉ះពាល់នឹងកម្រិតខ្ពស់នៃថ្នាំអស់រយៈពេលជាយូរមកហើយអាចបំប្លែងអ្នកដែលមានផ្ទុកហ្សែនទាបទៅជាអ្នកញៀន។.4
វឌ្ឍនភាពដ៏អស្ចារ្យត្រូវបានធ្វើឡើងក្នុងរយៈពេលពីរទសវត្សកន្លងមកនេះក្នុងការកំណត់តំបន់ដាច់ស្រយាលនៃខួរក្បាលដែលមានសារៈសំខាន់ក្នុងការសំរបសំរួលរោគសញ្ញានៃការញៀនក៏ដូចជាប្រភេទនៃការផ្លាស់ប្តូរនៅកំរិតម៉ូលេគុលនិងកោសិកាដែលគ្រឿងញៀននាំអោយមាននៅក្នុងតំបន់ទាំងនេះដើម្បីបង្ហាញពីទិដ្ឋភាពសំខាន់ៗ។ ញៀន។1,5 សៀគ្វីដែលទទួលបានការចាប់អារម្មណ៍បំផុតត្រូវបានគេហៅថាប្រព័ន្ធដូប៉ូមីលីកដែលទាក់ទងនឹងណឺរ៉ូនឌីផូមីញ៉ូមនៅក្នុងតំបន់ដែលមានខ្យល់ដង្ហើម (VTA) នៃណឺរ៉ូនប្រសាទមធ្យមដែលមានពន្លឺព្រះអាទិត្យនៅក្នុងនុយក្លីដិនដាប់ប៊ែល (ណៃស៊ីដែលជាផ្នែកមួយនៃកោសិកាឈាមរត់) ។ ណឺរ៉ូន VTA ទាំងនេះក៏ធ្វើឱ្យមានផ្នែកខាងមុខនៃតំបន់ជាច្រើនទៀតរួមមានហ៊ីបភីផាំមូសអឹមឌីដាឡានិងផ្នែកខាងកណ្តាលផ្នែកខាងមុខ (PFC) ។
វាសមហេតុផលក្នុងការពិចារណាពីយន្ដការញៀនដែលបង្កឡើងដោយគ្រឿងញៀនទាំងនេះក្នុងបរិមាណនេះលើការចងចាំសម្រាប់ហេតុផលបីជាន់គ្នា។6
- ទីមួយរាល់ការបន្សាំទៅនឹងថ្នាំអាចត្រូវបានគេមើលឃើញថាជាប្រភេទនៃ“ ការចងចាំម៉ូលេគុលឬកោសិកា៖ កោសិកាប្រសាទដែលមានការផ្លាស់ប្តូរបែបនេះគឺខុសគ្នាដោយសារលទ្ធផលនៃការប្រើថ្នាំហើយដូច្នេះមានប្រតិកម្មខុសគ្នាពីថ្នាំដដែល។ ជាលទ្ធផលថ្នាំដទៃទៀតឬចំពោះសកម្មភាពរំញោចផ្សេងៗជាលទ្ធផល។.
- ទីពីរវាគួរឱ្យចាប់អារម្មណ៍ដែលភាគច្រើននៃប្រភេទនៃការផ្លាស់ប្តូរដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងស្ថានភាពនៃការញៀន។ (ឧទាហរណ៏ការប្តូរហ្សែនដែលផ្លាស់ប្តូរហ្សែនហ្សែនហ្សែនហ្សែនប្លាស្ទិកកោសិកានិងកោសិកាទាំងមូលនិងលក្ខណៈរូបវិទ្យានិងប្រព័ន្ធប្រសាទប្រសាទ) ត្រូវបានផ្សារភ្ជាប់ផងដែរនៅក្នុងទម្រង់ប្រពៃណីនៃ "ការចងចាំអាកប្បកិរិយា" ដូចជាការចងចាំទំហំការភ័យខ្លាចនិងលក្ខខណ្ឌប្រតិបត្តិការក្នុងចំណោមឧបករណ៍ផ្សេងទៀត។
- ទីបីក្នុងចំណោមតំបន់ខួរក្បាលដែលរងផលប៉ះពាល់ដោយគ្រឿងញៀននៃការរំលោភបំពានគឺជាតំបន់ដែលមានមុខងារទាក់ទងនឹងសរសៃប្រសាទសម្រាប់ការចងចាំអាកប្បកិរិយារួមទាំង hippocampus, amygdala និង PFC ។ នេះស្របពេលជាមួយនឹងការយល់ដឹងកាន់តែច្បាស់ថាលក្ខណៈពិសេសសំខាន់បំផុតនៃការញៀនដែលមើលឃើញតាមគ្លីនិក (ឧទាហរណ៍ការឃ្លានគ្រឿងញៀននិងការថយចុះវិញ) ឆ្លុះបញ្ចាំងពីភាពមិនប្រក្រតីនៅក្នុងសៀគ្វីចងចាំជាប្រពៃណីជាមួយនឹងការចងចាំរយៈពេលវែងនៃបទពិសោធថ្នាំដែលជាកត្តាជំរុញដ៏ខ្លាំងក្លានៃរោគញៀន។4,7,8 ផ្ទុយទៅវិញតំបន់រង្វាន់នៃខួរក្បាល (ឧទាហរណ៍ VTA និង NAc) ជះឥទ្ធិពលយ៉ាងខ្លាំងដល់ការចងចាំអាកប្បកិរិយា។
អត្ថបទនេះផ្តល់នូវទិដ្ឋភាពទូទៅនៃប្រភេទសំខាន់ៗនៃការផ្លាស់ប្តូរម៉ូលេគុលនិងកោសិកាដែលកើតឡើងនៅក្នុងតំបន់ខួរក្បាលជាច្រើននៅក្នុងគំរូនៃការញៀនសត្វដោយផ្តោតលើនុយក្លេអ៊ែរដែលព័ត៌មានភាគច្រើនមាននាពេលបច្ចុប្បន្ន។ សំខាន់វាមានលទ្ធភាពកាន់តែខ្លាំងឡើងក្នុងការធ្វើឱ្យមានការផ្លាស់ប្តូរខ្លះនៃការញៀនរបស់មនុស្សដោយផ្អែកលើការសិក្សាអំពីខួរក្បាលក្រោយខួរក្បាល។ ទោះបីជាការពិតដែលថាគ្រឿងញៀននៃការរំលោភបំពានមានរចនាសម្ព័ន្ធគីមីខុសគ្នានិងធ្វើសកម្មភាពលើគោលដៅប្រូតេអ៊ីនខុសគ្នាក៏ដោយវាគួរអោយចាប់អារម្មណ៍ថាការបន្សាំទាក់ទងនឹងការញៀនដែលលេចធ្លោគឺមានជាទូទៅចំពោះមនុស្សជាច្រើនហើយក្នុងករណីខ្លះគ្រឿងញៀននៃការរំលោភបំពាននិងទំនងជារួមចំណែកដល់លក្ខណៈពិសេសនៃឯកសារ រោគសញ្ញាញៀន។.4,9 ផ្ទុយទៅវិញការបន្សាំគ្រឿងញៀនជាច្រើនផ្សេងទៀតមានលក្ខណៈជាក់លាក់ចំពោះថ្នាំដែលបានផ្តល់ហើយអាចសំរបសំរួលទិដ្ឋភាពប្លែក ៗ បន្ថែមទៀតនៃការញៀន។ យើងផ្តោតសំខាន់លើថ្នាំរំញោចនិងប្រើគ្រឿងញៀននៃការរំលោភបំពានដែលបង្កើតផលប៉ះពាល់គួរឱ្យកត់សម្គាល់នៅក្នុងគំរូសត្វបើប្រៀបធៀបជាមួយថ្នាំដទៃទៀត។ យើងក៏គូសបញ្ជាក់ផ្នែកសំខាន់ៗសម្រាប់ការស្រាវជ្រាវនាពេលអនាគតដែលនឹងបង្កើនចំណេះដឹងបន្ថែមទៀតនៃរោគសញ្ញាញៀននិងបកប្រែការជឿនលឿនទាំងនេះទៅជាការធ្វើតេស្តនិងការព្យាបាលដែលមានភាពប្រសើរឡើង។
យន្ដការចម្លងនិងអេពីកំណើត។
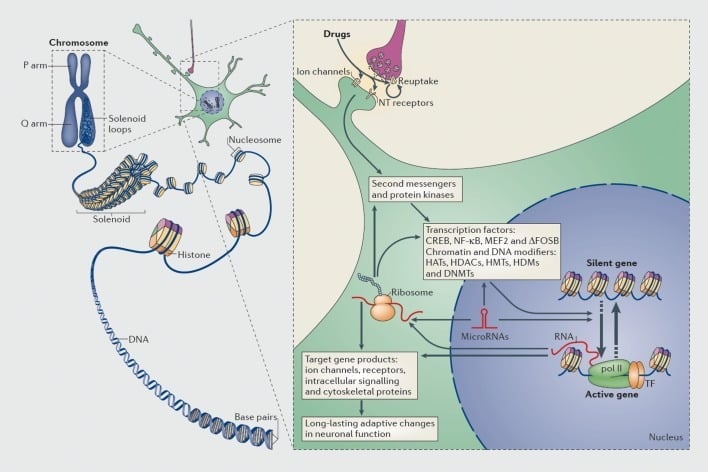
ចំណេះដឹងដែលថាអ្នកញៀនអាចនៅតែមានហានិភ័យខ្ពស់ក្នុងការធូរស្បើយវិញទោះបីជាការចៀសវាងច្រើនឆ្នាំមានន័យថាការញៀនពាក់ព័ន្ធនឹងការផ្លាស់ប្តូរថ្នាំដែលបង្កឡើងដោយខួរក្បាលដែលអាចមានស្ថេរភាព។ នេះបាននាំឱ្យក្រុមជាច្រើនពិចារណាការផ្លាស់ប្តូរនៅក្នុងការបញ្ចេញហ្សែនដែលជាសមាសធាតុសំខាន់នៃដំណើរការញៀន។ (រូបភាព 1) ។ ដូច្នោះហើយការសិក្សាអំពីហ្សែនរបស់បេក្ខជនឬការស៊ើបអង្កេតពន្ធុពាក់ព័ន្ធនឹងអតិសុខុមទស្សន៍ឌីអិនអេនិងថ្មីៗនេះ RNA-seq (លំដាប់ខ្ពស់នៃ RNA ដែលបានបង្ហាញ) បានកំណត់ហ្សែនជាច្រើនដែលការបញ្ចេញមតិត្រូវបានផ្លាស់ប្តូរនៅក្នុងតំបន់ខួរក្បាលដែលបានផ្តល់ឱ្យជាគំរូនៃសត្វកករនិងសត្វនៃការញៀននិងក្នុងការញៀនមនុស្ស។ (ឧទាហរណ៍ការបដិសេធ 10-17) ។ ឧទាហរណ៍នៃហ្សែនបែបនេះត្រូវបានពិភាក្សានៅក្នុងផ្នែកជាបន្តបន្ទាប់នៃការពិនិត្យឡើងវិញនេះ។
យន្តការនៃបទបញ្ញត្តិចម្លងនិងអេពីកំណើតដោយគ្រឿងញៀននៃការរំលោភបំពាន។ នៅក្នុងកោសិកា eukaryotic ឌីអិនអេត្រូវបានរៀបចំឡើងដោយរុំព័ទ្ធជុំវិញអ៊ីស្ត្រូសែនដើម្បីបង្កើតនុយក្លេអ៊ែរដែលបន្ទាប់មកត្រូវបានរៀបចំនិងបង្រួមដើម្បីបង្កើតជាក្រូម៉ូសូម (ផ្នែកខាងឆ្វេង) ។ មានតែការបង្រួបបង្រួម chromatin បង្រួមជាបណ្ដោះអាសន្នប៉ុណ្ណោះដែលឌីអិនអេនៃហ្សែនជាក់លាក់មួយអាចចូលដំណើរការបានដោយប្រើគ្រឿងម៉ាស៊ីនចម្លង។ គ្រឿងញៀននៃការរំលោភបំពានធ្វើសកម្មភាពតាមរយៈគោលដៅដែលអាចធ្វើសមកាលកម្មបានដូចជាយន្តការចាប់យកបណ្តាញអ៊ីយ៉ុងនិងប្រព័ន្ធទទួលសរសៃប្រសាទបញ្ជូនសរសៃប្រសាទ (NT) ទៅនឹងការផ្លាស់ប្តូររណសិរ្សបញ្ជូនសញ្ញាដែលមានអន្តរកម្ម (ផ្នែកខាងស្តាំ) ។ នេះនាំឱ្យមានការធ្វើឱ្យសកម្មឬការហាមឃាត់កត្តាចម្លង (TFs) និងគោលដៅនុយក្លេអ៊ែរជាច្រើនទៀតរួមទាំងប្រូតេអ៊ីនដែលមានបទប្បញ្ញត្តិ chromatin (បង្ហាញដោយព្រួញក្រាស់); យន្ដការលម្អិតដែលពាក់ព័ន្ធនឹងបទបញ្ញត្តិស៊ីចង្វាក់គ្នានៃប្រូតេអ៊ីន - នីកូទីលនៅតែមិនទាន់យល់ច្បាស់នៅឡើយ។ ដំណើរការទាំងនេះនៅទីបំផុតនាំឱ្យមានការបង្កើតឬការបង្ក្រាបនៃហ្សែនជាក់លាក់រួមទាំងដំណើរការ RNA ដែលមិនមែនជាកូដដូចជា microRNA ។ ការផ្លាស់ប្តូរការបង្ហាញនៃហ្សែនទាំងនេះខ្លះអាចផ្លាស់ប្តូរការផ្លាស់ប្តូរហ្សែន។ វាត្រូវបានគេស្នើសុំថាការផ្លាស់ប្តូរដែលបង្កឡើងដោយគ្រឿងញៀនមួយចំនួននៅកម្រិតក្រូម៉ូសូមមានស្ថេរភាពខ្លាំងហើយដោយហេតុនេះបានបង្ហាញអាកប្បកិរិយាយូរអង្វែងដែលកំណត់ការញៀន។ CREB, ប្រូតេអ៊ីនភ្ជាប់ឆ្លើយតបទៅនឹង AMP ដែលទាក់ទងនឹងធាតុ; DNMTs, DNA methyltransferases; HATs, histone acetyltransferases; HDACs, histone deacetylases; HDMs, អ៊ីស្ត្រូស្យូមស៊ីអ៊ីលលីន; HMTs, histone methyltransferases; MEF2, កត្តាកែលម្អជាក់លាក់ myocyte 2; NF-kB, កត្តានុយក្លេអ៊ែរ-KB; pol II, RNA polymerase II ។ ផលិតឡើងវិញពី Ref 44: Robison AJ, Nestler EJ ។ យន្ដការចម្លងនិងអេពីកំណើតនៃការញៀន។ ណាត Rev Neurosci ។ 2011; 12: 623-637 ។
ដូចគ្នានេះដែរកត្តាចម្លងជាច្រើនប្រភេទ - ប្រូតេអ៊ីនដែលភ្ជាប់ទៅនឹងតំបន់បទប្បញ្ញត្តិនៃហ្សែននិងដោយហេតុនេះបង្កើនឬបន្ថយការចម្លងហ្សែនទាំងនោះ - ត្រូវបានបង្កប់ក្នុងការសំរបសំរួលផលប៉ះពាល់រយៈពេលវែងនៃគ្រឿងញៀននៃការរំលោភលើការបញ្ចេញហ្សែននៅក្នុងខួរក្បាល។។ ឧទាហរណ៍លេចធ្លោរួមមាន CREB (ប្រូតេអ៊ីនភ្ជាប់ធាតុឆ្លើយតបរបស់ cAMP), ΔFosB (កត្តាចម្លងគ្រួសារ Fos), NFkB (កត្តានុយក្លេអ៊ែ kB), MEF2 (កត្តាបង្កើន myocyte-2) និងអ្នកទទួលជាតិ glucocorticoid ក្នុងចំណោមអ្នកដទៃទៀត។5,10,18-22 គេអាចយល់កាន់តែច្បាស់អំពីផ្លូវបញ្ជូនសញ្ញាកោសិកាតាមរយៈការរំលោភបំពានគ្រឿងញៀនដែលធ្វើឱ្យកត្តាចម្លងក្នុងខួរក្បាលមានឥទ្ធិពលហើយភ្ជាប់ការធ្វើឱ្យសកម្មទៅនឹងហ្សែនគោលដៅរបស់កត្តាចម្លងនេះនិងទិដ្ឋភាពអាកប្បកិរិយាជាក់លាក់នៃការញៀន (មើល រូបភាពទី 1). វឌ្ឍនភាពនេះត្រូវបានបង្ហាញដោយការពិចារណារបស់ CREB និងΔFosBដែលជាកត្តាចម្លងការសិក្សាដែលល្អបំផុតនៅក្នុងគំរូនៃការញៀន។
ធាតុឆ្លើយតបរបស់អេមអេហ្វអេមអេហ្វអេស។
ថ្នាំរំញោចនិងថ្នាំអាភៀននៃការរំលោភធ្វើឱ្យសកម្ម។ CREB ។ នៅក្នុងតំបន់ខួរក្បាលមួយចំនួនដែលមានសារៈសំខាន់សម្រាប់ការញៀនរួមទាំងភាពលេចធ្លោនៅក្នុង NAc ។.23,24 CREB ត្រូវបានគេដឹងថាត្រូវបានធ្វើឱ្យសកម្មនៅក្នុងប្រព័ន្ធផ្សេងទៀតដោយស៊ីអេហ្វអេ, កា។2+និងមាគ៌ានៃកត្តាកំណើន25 ហើយគេមិនទាន់ដឹងថាតើថ្នាំណាមួយដែលសំរបសំរួលសកម្មភាពរបស់ខ្លួននៅក្នុង NAc ដោយគ្រឿងញៀននៃការរំលោភបំពាន។ សកម្មភាពគ្រឿងញៀនរបស់ CREB នៅ NAc ត្រូវបានបង្ហាញថាតំណាងឱ្យយន្តការផ្តល់យោបល់អវិជ្ជមានបែបបុរាណ។, ដោយហេតុនេះ CREB ជួយកាត់បន្ថយភាពប្រែប្រួលរបស់សត្វទៅនឹងផលចំណេញនៃថ្នាំទាំងនេះ (និងការអត់អោន) និងដើម្បីសំរួលអារម្មណ៍អវិជ្ជមានក្នុងពេលដកគ្រឿងញៀន (ការពឹងផ្អែក).18,26,27 ផលប៉ះពាល់ទាំងនេះត្រូវបានបង្ហាញនាពេលថ្មីៗនេះដើម្បីជំរុញការគ្រប់គ្រងថ្នាំដោយខ្លួនឯងនិងការកើនឡើងវិញជាពិសេសតាមរយៈដំណើរការនៃការពង្រឹងអវិជ្ជមាន។28 សកម្មភាពទាំងនេះរបស់ CREB ហាក់ដូចជាពាក់ព័ន្ធទៅនឹងអនុប្រភេទធំ ៗ នៃណឺត្រូនប្រសាទស្ពឺដែលជាសញ្ញាបង្ហាញដែលលើសលុប1 ទល់នឹងឃ។2 អ្នកទទួលថ្នាំដូប៉ាមីន។.24 Iគួរឱ្យកត់សម្គាល់ស្ថាប័នអក្សរសិល្ប៍ដ៏ធំមួយបានបង្ហាញថា CREB ដែលដើរតួជាហ៊ីបភីផាំបនិងអាមមីដាឡាគឺជាម៉ូលេគុលសំខាន់នៅក្នុងការចងចាំអាកប្បកិរិយា.29-31 តួនាទីដ៏ទូលំទូលាយនៅក្នុងការញៀននិងការចងចាំអាកប្បកិរិយាទំនងជាឆ្លុះបញ្ចាំងពីការពិតដែលថាណឺរ៉ូនត្រូវបានភ្ជាប់ជាមួយនឹងចំនួនម៉ូទ័រម៉ូលេគុលដែលមានកំណត់ដែលត្រូវសម្របខ្លួនទៅនឹងបរិយាកាសដែលមានការផ្លាស់ប្តូរឥតឈប់ឈរ។
ហ្សែនគោលដៅសម្រាប់ CREB ដែលសម្របសម្រួល phenotype អាកប្បកិរិយានេះត្រូវបានកំណត់អត្តសញ្ញាណតាមរយៈការអះអាងទូទៅនិងការខិតខំប្រឹងប្រែងដែលបានជ្រើសរើស។10,18,32 ឧទាហរណ៍មួយគឺ dynorphin peptide opioid៖ ការជម្រុញអោយមានការបញ្ចេញមតិ dynorphin នៅក្នុងណឺរ៉ូនណឺរ៉ូនដែលត្រូវបានសំរបសំរួលតាមរយៈ CREB បង្កើនការធ្វើឱ្យសកម្មរបស់ឌីផូលីនទទួលអ្នក K opioid នៅលើណឺត្រុងឌីអ៊ីប៉ូតាមីននិងអាចបញ្ចោញការបញ្ជូនដូប៉ាមីនទៅឌីអេនអេ។ និងរង្វាន់អន់ថយ។.18 គោលដៅរបស់ CREB ជាច្រើនត្រូវបានគេបង្ហាញថាមានសារៈសំខាន់សម្រាប់ប្លាស្ទិកប្លាស្ទិកដែលត្រូវបានបង្កឡើងដោយគ្រឿងញៀនដូចដែលបានពិភាក្សាខាងក្រោម។ ខណៈពេលដែល CREB ត្រូវបានធ្វើឱ្យសកម្មផងដែរនៅក្នុងតំបន់ខួរក្បាលជាច្រើនទៀតដោយភ្នាក់ងាររំញោចនិងអាភៀន។23,24 មិនសូវត្រូវបានគេដឹងអំពីផលវិបាកនៃឥរិយាបទនៃឥទ្ធិពលនេះនិងហ្សែនគោលដៅដែលវាកើតឡើង។ ដូចគ្នានេះដែរគេមិនសូវដឹងអំពីតួនាទីរបស់ CREB ក្នុងការសម្រុះសម្រួលសកម្មភាពថ្នាំដទៃទៀតនៃការរំលោភបំពាន។19
ΔFosB
ការប៉ះពាល់ទៅនឹងថ្នាំដែលរំលោភបំពានស្ទើរតែបង្កឱ្យមានកត្តាចម្លងគ្រួសារ Fos ទាំងអស់នៅក្នុង NAc និងតំបន់ខួរក្បាលជាច្រើនទៀត។ អាំងឌុចស្យុងនេះមានល្បឿនលឿនប៉ុន្តែក៏មានការផ្លាស់ប្តូរយ៉ាងលឿនផងដែរជាមួយនឹងកំរិតប្រូតេអ៊ីន Fos ត្រលប់ទៅធម្មតាវិញក្នុងរយៈពេល 8 ដល់ 12 ម៉ោង។ អ្វីដែលប្លែកក្នុងចំនោមប្រូតេអ៊ីនគ្រួសារ Fos ទាំងនេះគឺΔFosBដែលជាផលិតផលកាត់ចេញនៃហ្សែន FosB ។ ដោយគុណធម៌នៃស្ថេរភាពមិនធម្មតារបស់វាបន្តិចម្តង ៗ បណ្តើរ ៗ ឆ្លងកាត់វគ្គនៃការប៉ះពាល់គ្រឿងញៀនម្តងហើយម្តងទៀត។ ហើយក្លាយជាប្រូតេអីនកូសលេចធ្លោក្រោមល័ក្ខខ័ណ្ឌទាំងនេះ។22,33 លើសពីនេះទៅទៀតដោយសារតែស្ថេរភាពនេះកម្រិតនៃΔFosBនៅតែបន្តអស់ជាច្រើនសប្តាហ៍បន្ទាប់ពីការដកគ្រឿងញៀន។ អាំងវឺតទ័ររ៉ាំរ៉ៃបែបនេះត្រូវបានបង្ហាញសម្រាប់ការរំលោភបំពានស្ទើរតែទាំងអស់នៃគ្រឿងញៀន។34 ហើយសំរាប់ថ្នាំភាគច្រើនគឺជ្រើសរើសសំរាប់ណឺរ៉ូន Dl-type NAc ។34,35 វាក៏បាន បង្ហាញនៅក្នុងអ្នកញៀនមនុស្ស។35 តួអក្សរសិល្ប៍ធំមួយបានបង្ហាញថាអាំងឌុច B ហ្វ័រប៊ីនៅក្នុងឌី1-ណឺរ៉ូនប្រភេទណុនជួយបង្កើនភាពប្រែប្រួលរបស់សត្វចំពោះថ្នាំក៏ដូចជារង្វាន់ធម្មជាតិនិងលើកកម្ពស់ការគ្រប់គ្រងថ្នាំដោយខ្លួនឯងតាមរយៈដំណើរការនៃការពង្រឹងវិជ្ជមាន។ (សូមមើលឯកសារយោង 34 ដល់ 38). អ្វីដែលគួរឱ្យចាប់អារម្មណ៍នោះគឺថាការបញ្ចូលគ្រឿងញៀនΔFosBក្នុងអិនអេអេគឺមានភាពទាក់ទាញខ្លាំងចំពោះសត្វវ័យជំទង់ដែលជាពេលវេលាងាយរងការញៀនខ្លាំង។,39 ហើយការបង្កើតរបស់វាដោយជាតិនីកូទីនត្រូវបានបង្ហាញដល់ការធ្វើឱ្យប្រសើរឡើងនៃការទទួលបាននូវកូកាអ៊ីនដែលធ្វើឱ្យមានលក្ខណៈល្អប្រសើរឡើង.40
ចំពោះ CREB, ហ្សែនគោលដៅជាច្រើនសម្រាប់ΔFosBត្រូវបានកំណត់អត្តសញ្ញាណនៅក្នុង NAc ដោយការប្រើប្រាស់ហ្សែនបេក្ខជននិងវិធីសាស្រ្តធំទូលាយហ្សែន។10,32 ខណៈពេលដែល CREB បង្កើតឌីណាម៉ូនΔFosBបង្ក្រាបវាដែលរួមចំណែកដល់ផលចំណេញគាំទ្រΔFosB។.38 គោលដៅΔFosBមួយទៀតគឺ cFos៖ ដោយសារΔFosBកកកុញជាមួយនឹងការប៉ះពាល់គ្រឿងញៀនម្តងហើយម្តងទៀតវាបង្ក្រាប c-Fos និងរួមចំណែកដល់កុងតាក់ម៉ូលេគុលដែលΔFosBត្រូវបានជ្រើសរើសនៅក្នុងស្ថានភាពព្យាបាលដោយថ្នាំរ៉ាំរ៉ៃ។.41 គោលដៅΔFosBជាច្រើនទៀតត្រូវបានបង្ហាញដើម្បីសម្របសម្រួលសមត្ថភាពថ្នាំជាក់លាក់នៃការរំលោភបំពានក្នុងការធ្វើឱ្យប្លាស្ទិកស៊ីបស៊ីបនៅក្នុងអេនអេចនិងការផ្លាស់ប្តូរដែលទាក់ទងទៅនឹងការបង្កើតប្រព័ន្ធប្រសាទប្រសាទប្រសាទមធ្យមអាអេសស៊ីដូចដែលនឹងត្រូវបានពិភាក្សាខាងក្រោម។
ផលវិបាកមុខងាររបស់ΔFosB induction ក្នុងតំបន់ខួរក្បាលផ្សេងទៀតមិនសូវត្រូវបានគេដឹងនោះទេទោះបីជាការចាប់ផ្តើមរបស់វានៅក្នុងគន្លងអវកាស (OFC) ត្រូវបានសិក្សាលម្អិតក៏ដោយ។ នៅទីនេះΔFosBសម្របសម្រួលការអត់ធ្មត់ដែលកើតឡើងចំពោះផលប៉ះពាល់នៃការយល់ដឹងរបស់កូកាអ៊ីនក្នុងកំឡុងពេលនៃការប៉ះពាល់រ៉ាំរ៉ៃហើយការបន្សាំនេះត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើនការគ្រប់គ្រងដោយខ្លួនឯងរបស់កូកាអ៊ីន។.42,43
ការសន្និដ្ឋានទូទៅរបស់ហ្សែនបានស្នើហ្សែនគោលដៅសក្តានុពលជាច្រើនដែលសម្របសម្រួលផលប៉ះពាល់ទាំងនេះ។42 ទោះបីជា propertiesFosB មានលក្ខណៈសម្បត្តិខាងសាច់ឈាមតែមួយនិងចំណេះដឹងថាវាត្រូវបានបង្កើតឡើងនៅក្នុងសៀគ្វីនៃការចងចាំតាមបែបប្រពៃណី (ឧទាហរណ៍ហ៊ីបភីផាប់) ក៏ដោយក៏មិនទាន់មានការរុករកតួនាទីΔFosBក្នុងការចងចាំអាកប្បកិរិយាដែលជាប្រធានបទគួរឱ្យចាប់អារម្មណ៍សម្រាប់ការស្រាវជ្រាវនាពេលអនាគត។
យន្តការអេក្វេនិច
ក្នុងប៉ុន្មានឆ្នាំថ្មីៗនេះការសិក្សាអំពីការចម្លងត្រូវបានជំរុញមួយជំហានទៀតឆ្ពោះទៅរកអេពីដេមី។44 (សូមមើល រូបភាពទី 1)ដែលអាចត្រូវបានកំណត់យ៉ាងទូលំទូលាយថាជាការផ្លាស់ប្តូរកន្សោមហ្សែនដែលកើតឡើងក្នុងករណីដែលគ្មានការផ្លាស់ប្តូរលំដាប់ឌីអិនអេ។ យន្ដការពន្ធុសេនេទិចគ្រប់គ្រងការវេចខ្ចប់ឌីអិនអេនៅក្នុងស្នូលកោសិកាតាមរយៈអន្តរកម្មរបស់វាជាមួយអ៊ីស្ត្រូសែននិងប្រូតេអ៊ីននុយក្លេអ៊ែរជាច្រើនប្រភេទផ្សេងទៀតដែលរួមបញ្ចូលគ្នារវាងក្រូទីន។ ការបង្ហាញហ្សែនត្រូវបានគ្រប់គ្រងដោយស្ថានភាពនៃការវេចខ្ចប់នេះតាមរយៈការកែប្រែនៃអ៊ីស្ត្រូសែនប្រូតេអ៊ីនដទៃទៀតនិងឌីអិនអេខ្លួនឯង។ ដូចឧទាហរណ៍ខ្លះអ៊ីស្ត្រូលីននៃអ៊ីស្ត្រូសែនមាននិន្នាការជំរុញការធ្វើឱ្យហ្សែនសកម្មមេតាណុលនៃអ៊ីស្តូនអាចជម្រុញសកម្មភាពហ្សែនឬការបង្ក្រាបអាស្រ័យលើសំណល់លីសដែលឆ្លងកាត់ការកែប្រែនេះហើយមេតាណុលឌីអិនអេជាទូទៅទាក់ទងនឹងការបង្ក្រាបហ្សែនទោះបីជាទម្រង់មេតាណុលមានការប្រែប្រួលមួយចំនួនក៏ដោយ ( ឧទាហរណ៍ 5-hydroxymethylation) អាចត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងសកម្មភាពហ្សែន។
Epigenetics គឺជាយន្តការទាក់ទាញពីព្រោះនៅក្នុងប្រព័ន្ធដទៃទៀតឧទាហរណ៍ការវិវត្តនៃជីវវិទ្យាមហារីកនិងការផ្លាស់ប្តូរអេទីឡែនជាក់លាក់អាចមានជាអចិន្ត្រៃយ៍។ សម្រាប់ហេតុផលនេះ, epigenetics ត្រូវបានគេដេញតាមទាំងនៅក្នុងការរៀនសូត្រនិងគំរូនៃការចងចាំ (ឧទាហរណ៍ការបដិសេធ 45-48) ក៏ដូចជាការញៀន;44,49 នៅក្នុងប្រព័ន្ធទាំងពីរការផ្លាស់ប្តូរយ៉ាងជ្រាលជ្រៅត្រូវបានគេរាយការណ៍នៅក្នុងអ៊ីស្តូស្តេរ៉ូននិងមេតាណុលនិងនៅក្នុងមេទីលអិមអេទីល។ ដូចឧទាហរណ៍មួយអ៊ីស្ត្រូទីនមេទីលហ្វីហ្វហ្វីក G9a ត្រូវបានបង្កប់នៅក្នុងការចងចាំទាំងពីរ50 និងការញៀន។51,52 នៅក្នុងម៉ូដែលញៀន កន្សោម G9a ត្រូវបានកំណត់។ នៅក្នុង NAc ឆ្លើយតបទៅនឹងថ្នាំរំញោចឬថ្នាំអាភៀននៃការរំលោភបំពាននិង t ។គាត់ត្រូវបានបង្ហាញដើម្បីបង្កើនប្រសិទ្ធិភាពនៃថ្នាំទាំងនេះ។.51,52 គួរឱ្យចាប់អារម្មណ៍ការបង្ក្រាបកូកាអ៊ីន G9a ត្រូវបានសម្របសម្រួលដោយΔFosB។ G9a ជួយជំរុញភាពមិនស៊ីសង្វាក់នៃ Lys9 នៃអ៊ីស្ត្រូនអេសអេជអរអេស (H3K3me9) ដែលជាអ្នកសម្រុះសម្រួលដ៏សំខាន់នៃការបង្ក្រាបហ្សែន។ បន្ទះឈីបឈីបឬឈីប - ស៊ីក (ហ្សែនទីរ៉ូហ្សែលការពារមិនជាប់តាមរៀងៗខ្លួនដោយបន្ទះឈីបផ្សព្វផ្សាយឬលំដាប់ខ្ពស់) ត្រូវបានប្រើដើម្បីទទួលបានផែនទីហ្សែនដែលមានហ្សែននៅក្នុងអិនអេចដែលបង្ហាញការផ្លាស់ប្តូរ H2K3me9 បន្ទាប់ពីការរំញោចឬការប៉ះពាល់អាភៀន។32,52,53 តាមរយៈការត្រួតលើបញ្ជីហ្សែនទាំងនេះជាមួយនឹងបញ្ជីហ្សែននៃការផ្លាស់ប្តូរការបង្ហាញហ្សែននិងជាមួយផែនទីពន្ធុនៃទម្រង់ជាច្រើននៃការផ្លាស់ប្តូរហ្សែនហ្សែន (ឧទាហរណ៍ការចងΔFosB, ការចង CREB, ការកែប្រែអ៊ីស្តូន។ ល។ )32,53 វាគួរតែអាចកំណត់អត្តសញ្ញាណហ្សែនពេញលេញមួយដែលត្រូវបានកំណត់ដោយថ្នាំនៃការរំលោភបំពាននិងដើម្បីស្វែងយល់ពីយន្តការអេដ្យូមេទិកដែលពាក់ព័ន្ធ។
ទម្រង់មួយទៀតនៃបទប្បញ្ញត្តិអេហ្សែនទិកទាក់ទងនឹងការចងចាំនិងការញៀនគឺការបង្កើត microRNAs ។ RNAs ដែលមិនមែនជាខ្សែអក្សរតូចៗទាំងនេះភ្ជាប់ទៅនឹងតំបន់បន្ថែមនៃ MRNA ហើយដោយហេតុនេះអាចរារាំងការបកប្រែរបស់ពួកគេឬនាំឱ្យមានការរិចរិលរបស់ពួកគេ។ ការលុបអារីហ្គូតដែលជាប្រូតេអ៊ីនមានសារៈសំខាន់សម្រាប់ដំណើរការមីរ៉ាអិនកែប្រែការឆ្លើយតបអាកប្បកិរិយាចំពោះកូកាអ៊ីនដោយឥទ្ធិពលខុសគ្នាត្រូវបានគេសង្កេតឃើញចំពោះឌីអេសអេសអិច - ធៀបនឹងណឺរ៉ូនណឺរ៉ូនប្រភេទមធ្យម។54 miRNA ជាក់លាក់មួយចំនួនត្រូវបានបង្ហាញថាត្រូវបានកំណត់ដោយការប៉ះពាល់នឹងគ្រឿងញៀននិងដើម្បីធ្វើឱ្យមានឥទ្ធិពលលើការឆ្លើយតបអាកប្បកិរិយាចំពោះថ្នាំ (ឧទាហរណ៍ការបដិសេធ 55,56) ។ វានឹងគួរឱ្យរំភើបនៅក្នុងការសិក្សានាពេលអនាគតដើម្បីកំណត់គោលដៅ mRNA នៃ miRNA ទាំងនេះនិងបង្ហាញពីលក្ខណៈដែលវាប៉ះពាល់ដល់ដំណើរការញៀន។
ប្លាស្ទិក Synaptic ។
ប្រភេទទូទៅនៃការកែប្រែស៊ីចង្វាក់គ្នានៅចង្វាក់ភ្លេង glutamatergic synapses ដែលមានជាប់ទាក់ទងនឹងហ៊ីបភីផាំបនិងអាមីឌីដានៅក្នុងការចងចាំអាកប្បកិរិយា (សូមមើលអត្ថបទផ្សេងទៀតនៅក្នុងបញ្ហានេះ) ត្រូវបានបង្ហាញស្រដៀងគ្នានៅក្នុងតំបន់រង្វាន់ខួរក្បាលនៅក្នុងគំរូញៀននិងមានសារៈសំខាន់ក្នុងការសម្រុះសម្រួល។ ដំណើរការញៀន។.57,58 ប្លាស្ទិច synaptic ដែលបណ្តាលមកពីគ្រឿងញៀនត្រូវបានគេពិពណ៌នានៅក្នុងតំបន់ខួរក្បាលមួយចំនួនទោះជាយ៉ាងណាយើងផ្តោតអារម្មណ៍នៅទីនេះនៅលើ NAc ដែលការស្រាវជ្រាវភាគច្រើនផ្តោតលើកាលបរិច្ឆេទ។ (រូបភាព 2).
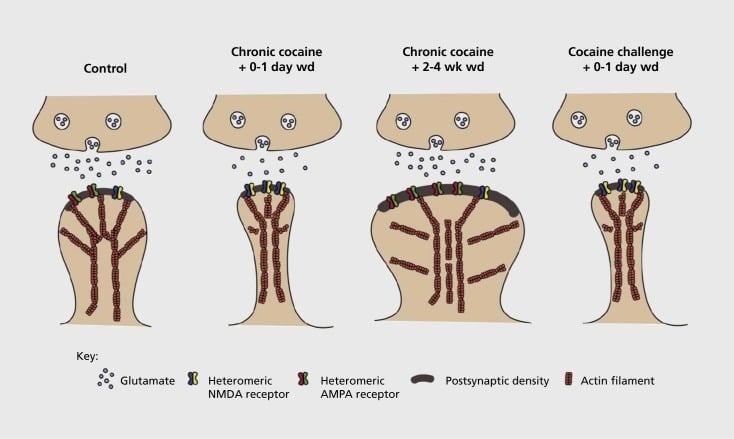
គំរូនៃប្លាស្ទិកប្លាស្ទិកដែលទាក់ទងនឹងការញៀននិងរចនាសម្ព័នដែលទាក់ទងនឹងការញៀននុយក្លេអ៊ែរ។ ការប៉ះពាល់នឹងកូកាអ៊ីនរ៉ាំរ៉ៃធ្វើឱ្យមានការរៀបចំឡើងវិញដោយពឹងផ្អែកលើពេលវេលានិងការផ្លាស់ប្តូរអាស៊ីដα-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) និងអាស៊ីត N-methyl-D-aspartic (NMDA) នៅ NAc ។ ប្រសាទណឺរ៉ូន (អេសអិនអិន) រួមបញ្ចូលគ្នាក៏ដូចជាការផ្លាស់ប្តូររចនាសម្ព័ន្ធនៅក្នុងក្បាលឆ្អឹងខ្នងរបស់អេអេអិនអេអេសអេសអិលដែលទាក់ទងនឹងទម្រង់ប្លាស្ទិកស៊ីបស៊ីប។ ឧទហរណ៍កូកាអ៊ីនរ៉ាំរ៉ៃបង្កឱ្យមានការបង្ហាញលើផ្ទៃរបស់អ្នកទទួល NMDA ការបង្កើតភាពស្ងាត់ស្ងៀមនិងការធ្លាក់ទឹកចិត្តរយៈពេលវែង (LTD) នៅចំណុចពេលវេលាដកដំបូង។ ក្នុងកំឡុងពេលដកយូរ (ការអូសបន្លាយ) ការផ្លាស់ប្តូរសមកាលកម្មទាំងនេះបញ្ច្រាស់ជាមួយនឹងលទ្ធផលដែលត្រូវបានបង្កើនការបង្ហាញនៃផ្ទៃ AMPA ដែលជាការច្របាច់បញ្ចូលគ្នាទៅក្នុងឆ្អឹងខ្នងដែលមានរាងដូចផ្សិតនិងសក្តានុពលរយៈពេលវែង (LTP) ។ ផលប៉ះពាល់ទាំងនេះត្រលប់មកវិញម្តងទៀតយ៉ាងឆាប់រហ័សនៅពេលដែលប៉ះពាល់នឹងកូកាអ៊ីនដែលនាំឱ្យមានរចនាសម្ព័ន្ធឆ្អឹងខ្នងទៅជាឆ្អឹងខ្នងស្ដើងនិងធ្លាក់ទឹកចិត្តនៃកម្លាំងសមកាលកម្ម។
ការពិសោធន៍ដំបូងបានបង្ហាញថាការប៉ះពាល់ម្តងហើយម្តងទៀតទៅនឹងថ្នាំរំញោចនៃការរំលោភបំពាននាំឱ្យមាន LTD (ការធ្លាក់ទឹកចិត្តរយៈពេលវែង) ដែលមានលក្ខណៈដូចនៅនឹងកន្លែងច្រើនលើសលុបនៅ NAc ។59 ទោះយ៉ាងណាការងារថ្មីៗកាន់តែច្រើនបានបង្ហាញពីភាពប្លាស្ទិកបែបនេះគឺពឹងផ្អែកទៅលើពេលវេលាខ្ពស់ដោយក្រុមហ៊ុនអិល។ ធី។ អិលបានកើតឡើងនៅដំណាក់កាលដំបូងបន្ទាប់ពីការប៉ះពាល់កូកាអ៊ីនចុងក្រោយកំពុងវិវឌ្ឍន៍ទៅជារដ្ឋ LTP (សក្តានុពលយូរអង្វែង) ដូចរដ្ឋបន្ទាប់ពីការដកពេលវេលាកាន់តែយូរ។60,61 ការងារនេះដែលរហូតមកដល់បច្ចុប្បន្ននេះត្រូវបានអនុវត្តជាចម្បងដោយប្រើអ្នកស៊ើបអង្កេតដែលបានគ្រប់គ្រងផ្ទុយពីថ្នាំដែលគ្រប់គ្រងដោយខ្លួនឯងបានកំណត់ពីតម្រូវការសម្រាប់ការស៊ើបអង្កេតជាប្រព័ន្ធបន្ថែមទៀតនៅក្នុងគំរូនៃការគ្រប់គ្រងរដ្ឋបាលខ្លួនឯងដែលតាមដានទម្រង់ប្លាស្ទិកដែលត្រូវគ្នាដែលកើតឡើងនៅដំណាលគ្នា។ NAc ក្នុងរយៈពេលវគ្គសិក្សាលម្អិតពីការទទួលបានការគ្រប់គ្រងរដ្ឋបាលខ្លួនឯងទៅនឹងការថែរក្សារបស់ខ្លួនតាមរយៈពេលវេលាផ្សេងៗគ្នានៃការដកនិងការផុតពូជហើយជាការឆ្លើយតបទៅនឹងការរំញោចនៃការវិលត្រឡប់។ ការងាររហូតមកដល់បច្ចុប្បន្ននេះបានកំណត់នូវយន្តការម៉ូលេគុលមួយចំនួនដែលរួមចំណែកដល់ប្លាស្ទិចស៊ីបស៊ីបដែលបណ្តាលមកពីគ្រឿងញៀនរួមទាំងការជួញដូរអេមភីអេសអេចអេសអេសដើម្បីវិវឌ្ឍន៍ប្រហែលជាត្រូវបានសំរបសំរួលមួយផ្នែកតាមរយៈ CaMKII (Ca2+/ កាល់ស្យូមពឹងផ្អែកលើប្រូតេអ៊ីន kinase II) phosphorylation នៃធាតុទទួល AMPA ជាក់លាក់ក៏ដូចជាការបញ្ចេញមតិដែលផ្លាស់ប្តូរនៃអនុផ្នែកទទួល AMPA (ឧទាហរណ៍ 60,62-65, តួលេខ 2 និង 3). តួនាទីរបស់ CREB និងΔFosBត្រូវបានជាប់ទាក់ទងនៅក្នុងបាតុភូតទាំងនេះក៏ដូចជាការផ្លាស់ប្តូរដែលទាក់ទងទៅនឹងលក្ខណៈនៃវណ្ណយុត្តិនៃពពួក glutamatergic (សូមមើលខាងក្រោម) ។ ឧទាហរណ៍ GluAl គឺជាគោលដៅសម្រាប់ CREB នៅ NAc ដែល GluA2 និង CaMKII គឺជាគោលដៅទាំងពីររបស់ΔFosBនៅក្នុងតំបន់ខួរក្បាលនេះ។ .35,36,66,67 ការឆ្ពោះទៅមុខវាជាការសំខាន់ដើម្បីភ្ជាប់ការសម្របខ្លួនជាក់លាក់ទៅនឹងការផ្លាស់ប្តូរដែលពឹងផ្អែកលើពេលវេលាក្នុងមុខងារស៊ីចង្វាក់គ្នានិងលក្ខណៈអាកប្បកិរិយានៃការញៀន។
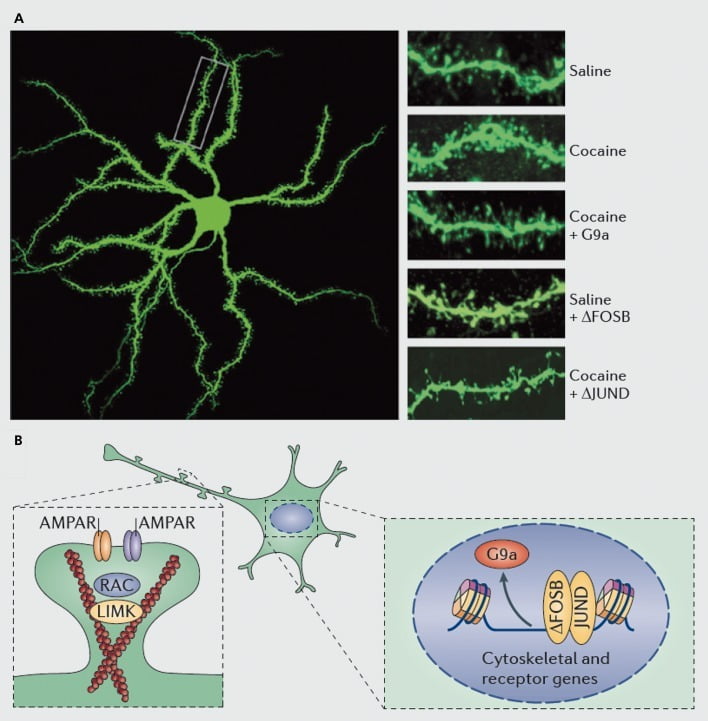
យន្ដការម៉ូលេគុលដែលស្ថិតនៅក្រោមកូកាអ៊ីននៃឆ្អឹងខ្នងដែលមានពន្លឺព្រះអាទិត្យនៅលើណឺត្រូនប្រសាទប្រសាទមធ្យម។ ក) បង្ហាញពីការកើនឡើងកូកាអ៊ីនក្នុងចំនួនឆ្អឹងខ្នងដែលអាចត្រូវបានរារាំងដោយការរីករាលដាលនៃវីរុស G9a ឬ JunD (អ្នកប្រឆាំងនឹងការចម្លង AP1 - ក្លែងបន្លំ) ឬត្រូវបានធ្វើត្រាប់តាមការរីករាលដាលនៃវីរុស FosB ។ ខ) បទបញ្ញត្តិនៃការជួញដូរ AMPA (AMPAR) និងការធ្វើ cytoskeleton (ខាងឆ្វេង) ក៏ដូចជាបទបញ្ញត្តិនៃការចម្លងនៃអ្នកទទួល glutamate និងប្រូតេអ៊ីននិយតកម្ម actin (ឧទាហរណ៍ដូចជាការសម្រុះសម្រួលតាមរយៈΔFosB, ស្តាំ) ត្រូវបានបង្ហាញថាដើរតួនាទីយ៉ាងសំខាន់ ក្នុងការសម្រុះសម្រួលបទបញ្ជារបស់កូកាអ៊ីននៃដង់ស៊ីតេឆ្អឹងខ្នងរបស់អេនអេអេស។ UMK, LIM domain kinase; RAC, ស្រទាប់ការពារជាតិពុល C3 ដែលទាក់ទងនឹង Ras ។
ឧបករណ៍ពិសោធន៍ថ្មីកំពុងធ្វើឱ្យវាមានលទ្ធភាពដំបូងដើម្បីកំណត់ជាមួយនឹងភាពជាក់លាក់កាន់តែច្បាស់ដែលសៀគ្វីជាក់លាក់ណាមួយបង្ហាញទម្រង់ប្លាស្ទិកស៊ីបស៊ីបនិងភាពមិនប្រក្រតីនៃអាកប្បកិរិយាដែលពួកគេសម្រុះសម្រួល។ ឧទាហរណ៍ t ។គាត់សែលនិងអនុតំបន់នៃ NAc បង្ហាញភាពខុសគ្នានៃប្លាស្ទិកស៊ីបស៊ីបដែលបណ្តាលមកពីគ្រឿងញៀនដូច D1- ធៀបនឹងណឺរ៉ូនអេឡិចត្រូនិចប្រភេទ D2 នៅក្នុងអនុតំបន់នីមួយៗ។60,63,64,67 ដូចគ្នានេះដែរការពិសោធន៍អុបទិកបានផ្តល់ការយល់ដឹងពីប្រលោមលោកទៅក្នុងការចូលរួមចំណែកនៃទម្រង់ប្លាស្ទិកដែលអាចធ្វើសមកាលកម្ម (ឧទាហរណ៍អិល។ អិល។ អិល។ អិល។ អិល។ ) នៅចំនួនប្រជាជនជាក់លាក់នៃខ្ទង់រាងពងក្រពើនៅអិនអេចឧទាហរណ៍លទ្ធផលដែលកើតឡើងពីផ។ ស។ ស។ ភ។ ធៀបនឹងអាមីដាប់ប៊ែលឡាទល់នឹងអនុផ្នែក ventral (ទិន្នផលសំខាន់) ។ នៃ hippocampus) ។68-70 នៅទីបំផុតវាចាំបាច់ក្នុងការត្រួតគ្នាទៅនឹងការបន្សាំនៃម៉ូលេគុលដែលបង្កដោយគ្រឿងញៀននៅក្នុងណឺត្រុងប្រសាទនីមួយៗជាមួយនឹងការបន្សាំទៅនឹងភាពជាក់លាក់ដែលកើតឡើងនៅក្នុងឌីផេរ៉ង់ស្យែលរបស់ពួកគេដើម្បីចងក្រងការយល់ដឹងពេញលេញអំពីវិធីដែលគ្រឿងញៀននៃការរំលោភបំពានកែប្រែសៀគ្វីខួរក្បាលដើម្បីជំរុញផ្នែកជាក់លាក់នៃ រដ្ឋញៀន។ ការខិតខំនេះនឹងតម្រូវឱ្យមានការកោតសរសើរកាន់តែខ្លាំងឡើងចំពោះប្លាស្ទិចដែលបណ្តាលមកពីគ្រឿងញៀននៅតាមកន្លែងរារាំងនៅក្នុងតំបន់ខួរក្បាលដូចគ្នាដែលជាតំបន់មួយដែលទទួលបានការយកចិត្តទុកដាក់តិចតួចបំផុតរហូតមកដល់បច្ចុប្បន្ននេះ។65
ប្លាស្ទិកកោសិកាទាំងមូល។
ខណៈពេលដែលការស្រាវជ្រាវភាគច្រើនពាក់ព័ន្ធនឹងការផ្លាស់ប្តូរប្រព័ន្ធប្រសាទសាស្រ្តនៃណឺរ៉ូននៅក្នុងបាតុភូតរំលោភបំពានគ្រឿងញៀនដូចជានៅក្នុងការសិក្សានិងបាតុភូតនៃការចងចាំបានផ្តោតលើប្លាស្ទិចដែលមានភាពស៊ីចង្វាក់គ្នាមានភ័ស្តុតាងកើនឡើងសម្រាប់សារៈសំខាន់នៃប្លាស្ទិកកោសិកាទាំងមូល។ ប្លាស្ទិកកោសិកាទាំងមូលដែលត្រូវបានគេស្គាល់ផងដែរថាជាប្លាស្ទិកក្នុងស្បូន។71 ពាក់ព័ន្ធនឹងការផ្លាស់ប្តូរភាពរំភើបដែលអាចកើតមាននៃកោសិកាសរសៃប្រសាទទាំងមូលក្នុងលក្ខណៈមួយដែលវាមិនជាក់លាក់។ ដោយសារលក្ខណៈពិសេសមួយចំនួននៃការញៀនគ្រឿងញៀនពាក់ព័ន្ធនឹងការបង្កើនឬបន្ថយភាពប្រែប្រួលទៅនឹងថ្នាំវាធ្វើឱ្យយល់ថាការបង្កើនឬកាត់បន្ថយភាពរំជើបរំជួលអគ្គិសនីរបស់កោសិកាសរសៃប្រសាទជាក់លាក់រួមចំណែកដល់ការសម្របសម្រួលឥរិយាបថទាំងនេះ។.5
នេះ ឧទាហរណ៏ដែលបានបង្កើតឡើងល្អបំផុតនៃប្លាស្ទិកកោសិកាទាំងមូលទៅនឹងគ្រឿងញៀននៃការរំលោភបំពានគឺជាសមត្ថភាពនៃថ្នាំអាភៀនរ៉ាំរ៉ៃដើម្បីបង្កើនភាពរំជើបរំជួលនៃសរសៃប្រសាទណឺរ៉ូននៃណឺរ៉ូនសូលុយស្យុង។).72 ភាពរំភើបដែលកើនឡើងនេះត្រូវបានសំរបសំរួលតាមរយៈ CREB និងការបង្កើតអាំងស៊ុយលីននៃខ្យល់ព្យុះ adenylyl ដែលជំរុញការកើនឡើងនៃណឺរ៉ូនអេសអិលប្រហែលជាតាមរយៈការបង្កើតបណ្តាញណា + ។72-75 hyperexcitabilty នៃណឺរ៉ូនអេសអិលនេះតំណាងឱ្យយន្តការបុរាណនៃការអត់ធ្មត់និងការពឹងផ្អែកហើយជំរុញឱ្យមានរោគសញ្ញានិងរោគសញ្ញាមួយចំនួននៃការដកអាភៀន។ គួរឱ្យចាប់អារម្មណ៍, CREB សំរបសំរួលទម្រង់ស្រដៀងគ្នានៃប្លាស្ទិចកោសិកាទាំងមូលនៅក្នុងណឺរ៉ូនណឺរណឺរណឺរៀរ, ដែលត្រូវបានបង្ហាញយ៉ាងផុយស្រួយដោយការប៉ះពាល់រ៉ាំរ៉ៃទៅនឹងថ្នាំនៃការរំលោភបំពានតាមរយៈ CREB ។76 ដូច្នេះវានឹងមានសារៈសំខាន់ក្នុងការស៊ើបអង្កេតនាពេលអនាគតដើម្បីយល់ពីរបៀបដែលស៊ីអេសប៊ី - ប្លាស្ទិចធ្វើសមកាលកម្មនៃភាពស៊ីឡាំងស្កូបឺរនៅលើណឺត្រូនប្រសាទមធ្យម។65,66 សមូហភាពជាមួយនឹង CREB - សំរបសំរួលផ្ចាញ់ផ្ចាលនៃប្រសាទប្រសាទទាំងនេះ។76 ដើម្បីគ្រប់គ្រងលក្ខណៈពិសេសនៃអាកប្បកិរិយានៃការញៀន។
ឧទាហរណ៍មួយទៀតនៃប្លាស្ទិកកោសិកាទាំងមូលនៅក្នុងគំរូនៃការញៀនគឺភាពប្រថុយប្រថាននៃសរសៃប្រសាទណឺរដូវីតាមីនដែលកើតឡើងបន្ទាប់ពីការប៉ះពាល់រ៉ាំរ៉ៃទៅនឹងថ្នាំអាភៀននៃការរំលោភបំពាន។e (រូបភាព 4).77,78 ការសម្របខ្លួននេះដែលត្រូវបានផ្សារភ្ជាប់ទៅនឹងការផ្លាស់ប្តូរ morphological នៅក្នុងកោសិកាសរសៃប្រសាទទាំងនេះ (សូមមើលផ្នែកបន្ទាប់) មិនត្រូវបានសម្របសម្រួលដោយ CREB ទេប៉ុន្តែសម្រេចបានជំនួសតាមរយៈបទប្បញ្ញត្តិនៃ cascades បញ្ជូនសញ្ញាប្រព័ន្ធប្រសាទដូចបានរៀបរាប់ខាងក្រោម។

គំរូធ្វើការនៃអាដាប់ធ័រម៉ូណុលរ៉ាំរ៉ៃនៅក្នុងតំបន់ដែលរំញោចរំអិល (VTA) ណឺរ៉ូន។ ការថយចុះទំហំម៉ូណូរ៉ាំរ៉ៃ VTA dopamine (DA) មិនត្រឹមតែបង្កើនភាពរំជើបរំជួលនៃសរសៃប្រសាទទេខណៈពេលដែលការបញ្ជូនដូប៉ាមីនទៅកាន់សរសៃប្រសាទនុយក្លេអ៊ែរមានការថយចុះ។ ផលប៉ះពាល់សុទ្ធរបស់ម៉ូល្លីនគឺជាមាគ៌ារង្វាន់ដែលមិនសូវឆ្លើយតបទេពោលគឺភាពអត់ធ្មត់នៃរង្វាន់។ ការធ្លាក់ចុះនៃសញ្ញា IRS2-AKT នៅក្នុង VTA សម្របសម្រួលផលប៉ះពាល់នៃម៉ូលេគុលរ៉ាំរ៉ៃលើទំហំសូម៉ានិងភាពរំភើបអគ្គិសនី។ ឥទ្ធិពលលើភាពរំភើបត្រូវបានសម្របសម្រួលតាមរយៈការថយចុះអាស៊ីដγ-aminobutyric (GABA) ចរន្តនិងការបញ្ចោញកន្សោមឆានែល K។ ការធ្លាក់ចុះនៃម៉ូទ័រហ្វីននៃសកម្មភាព mTORC2 នៅក្នុង VTA គឺមានសារៈសំខាន់ណាស់សម្រាប់ការសម្របខ្លួនម៉ូទ័រនិងសរីរវិទ្យាដែលបង្កើតឡើងដោយអរម៉ូនទាំងនេះក៏ដូចជាសម្រាប់ការទទួលយករង្វាន់។ ផ្ទុយពី mT0RC2 ម៉ូទ័រម៉ូណូរ៉ាំរ៉ៃបង្កើនសកម្មភាព mTORCI ដែលមិនមានឥទ្ធិពលលើអាដាប់ធ័រដែលបង្កើតដោយម៉ូម៉ូន។ BDNF, កត្តាសរសៃប្រសាទដែលទទួលបានពីខួរក្បាល; សារធាតុ IRS, សារធាតុទទួលអាំងស៊ុយលីន; mTORC, ស្មុគស្មាញ mTOR; AKT, ប្រូតេអ៊ីន kinase B ផលិតឡើងវិញពី Ref 77 ។
ប្លាស្ទិកប្លាស្ទិកនិងយន្តការសរសៃប្រសាទ។
ភ័ស្តុតាងដែលកំពុងកើនឡើងភាគច្រើនបានមកពីការសិក្សាអំពីកោសិកាប្រសាទហ៊ីបភីផបលនិងខួរក្បាលផ្នែកខួរក្បាលបានបង្ហាញថាការផ្លាស់ប្តូរប្លាស្ទិកស៊ីបស៊ីដត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការផ្លាស់ប្តូរលក្ខណៈរូបវិទ្យានៅ synapses ។ ឧទាហរណ៍អិល។ អិល។ ឌី។ អេសនិងជំនាន់នៃភាពស្ងាត់ស្ងៀមត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើតឆ្អឹងខ្នងដែលមានរាងដូចផ្សិតហើយចំណែកឯអិលភីធីត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងឆ្អឹងខ្នងដែលមានរាងដូចផ្សិត។79,80 ដូច្នេះវាគួរឱ្យចាប់អារម្មណ៍ណាស់ដែលថាកន្លែងប្រើគ្រឿងញៀនបានផ្តោតលើការផ្លាស់ប្តូរដែលបង្កឡើងដោយគ្រឿងញៀននៅក្នុងឆ្អឹងខ្នងដែលមិនមានភាពវឹកវរអស់រយៈពេល ១៥ ឆ្នាំ។ ការប៉ះពាល់រ៉ាំរ៉ៃទៅ។ ថ្នាំរំញោចនៃការរំលោភបំពានបង្កើនដង់ស៊ីតេឆ្អឹងខ្នងផ្នែកខាងសរសៃប្រសាទនៃអេណុកដែលជាការផ្លាស់ប្តូរដែលលេចធ្លោចំពោះណឺរ៉ូនប្រភេទ Dl ។.67,81,82 ការលួងលោមឆ្អឹងខ្នងត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងផ្នែកភាគច្រើនជាមួយនឹងការឆ្លើយតបអាកប្បកិរិយាចំពោះថ្នាំទាំងនេះទោះបីជាភ័ស្តុតាងខ្លះមានទំនាស់នឹងទស្សនៈនេះក៏ដោយ។
ដូចគ្នានឹងការសិក្សាអំពីប្លាស្ទិកធ្វើសមកាលកម្មដែរការងារជាច្រើនត្រូវបានគេត្រូវការជាចាំបាច់ដើម្បីកំណត់ជាលក្ខណៈប្រព័ន្ធនូវការផ្លាស់ប្តូរឆ្អឹងខ្នងដែលកំពុងកើតមានក្នុងកំឡុងពេលនៃការគ្រប់គ្រងខ្លួនឯងការដកប្រាក់និងការកើតឡើងវិញ។ Sការជាប់ទាក់ទងរហូតមកដល់បច្ចុប្បន្នដែលពាក់ព័ន្ធនឹងអ្នកស៊ើបអង្កេត - និងថ្នាំគ្រប់គ្រងខ្លួនឯងបានបង្ហាញថាការផ្លាស់ប្តូរឆ្អឹងខ្នងខុសគ្នាខ្លាំងដែលកើតឡើងនៅចំណុចពេលវេលាដកប្រាក់ខុសគ្នានិងនៅក្នុងសែល NAc ធៀបនឹងអនុតំបន់ស្នូល។.83-86 វាក៏សំខាន់ផងដែរក្នុងការកំណត់យន្តការម៉ូលេគុលច្បាស់លាស់ដែលកូកាអ៊ីនឬសារធាតុរំញោចមួយផ្សេងទៀតបង្កើតផលប៉ះពាល់ជាក់លាក់អាស្រ័យលើពេលវេលានិងកោសិកា។ ΔFosBត្រូវបានបង្ហាញថាមានភាពចាំបាច់និងគ្រប់គ្រាន់សម្រាប់ការបញ្ចូលឆ្អឹងខ្នងដែលមិនទាន់ពេញវ័យនៅលើកោសិកាសរសៃប្រសាទប្រភេទ Dl-NAc ។.35,51,67 បទប្បញ្ញត្តិបែបនេះកើតឡើងនៅក្នុងការប្រគុំតន្រ្តីជាមួយកូកាអ៊ីននិងបទបញ្ញត្តិΔFosBនៃប្រូតេអ៊ីនជាច្រើនដែលត្រូវបានគេស្គាល់ថាគ្រប់គ្រងការរៀបចំឡើងវិញនៃអរម៉ូនស៊ីធីកូសសុន។ ដូចឧទាហរណ៍មួយបទបញ្ញត្តិប្រតិចារិកនៃកត្តាផ្លាស់ប្តូរ nucleotide ហ្គីណេនជាច្រើននិងប្រូតេអ៊ីនដែលធ្វើឱ្យសកម្មរបស់ជីភីធីស្យូសជីភីអេសភីអេសជីអេសអេសសម្រាប់ការថយចុះនៃសកម្មភាពថយចុះក្នុងសកម្មភាពឆ្លើយតបនឹងការប៉ះពាល់កូកាអ៊ីននីមួយៗហើយការថយចុះថេរនៃសកម្មភាព Rac1 ត្រូវបានបង្ហាញដោយប្រើការត្រួតពិនិត្យអេកូទិក។ នៃ Rac1 ដើម្បីសំរបសំរួលការបញ្ចូលឆ្អឹងខ្នងដែលមិនទាន់ពេញវ័យ។87 ផលប៉ះពាល់ទាំងនេះរបស់រ៉ាល់សន្មតកើតឡើងតាមរយៈការគ្រប់គ្រងរបស់វាលើកូហ្វីលីននិងប្រូតេអីនប្រូតេអ៊ីនដទៃទៀតដែលត្រូវបានបង្ហាញផងដែរដើម្បីសម្របសម្រួលបទប្បញ្ញត្តិកូកាអ៊ីននៃការលូតលាស់ឆ្អឹងខ្នង។87,88 ទោះយ៉ាងណាក៏ដោយវាចាំបាច់ត្រូវបញ្ជាក់ថានេះគ្រាន់តែជាផ្លូវមួយដែលពាក់ព័ន្ធនឹងបទប្បញ្ញត្តិរបស់កូកាអ៊ីននៃឆ្អឹងខ្នងដែលមិនទាន់ពេញវ័យចាប់តាំងពីប្រូតេអ៊ីនដទៃទៀតត្រូវបានបង្ហាញថាដើរតួយ៉ាងសំខាន់ផងដែររួមមានស៊ីឌីខេ ៥ (ស៊ីនស៊ីន - គីនស៊ីស -៥), CaMKII, NFkB , MEF5, CREB, G5a, និង DNMT2 (ឌីអិនអេមេទីលត្រាហ្វលុបបំបាត់ ៣ ក) ដើម្បីដាក់ឈ្មោះមួយចំនួន។20,21,35,51,67,89,90 អ្វីដែលគួរឱ្យចាប់អារម្មណ៍នោះគឺបទប្បញ្ញត្តិកូកាអ៊ីននៃហ្សែនមួយចំនួនក្នុងចំណោមនោះរួមមានការបង្កើតស៊ីខេខេអេសអេចស៊ីខេអេអេខេនិងអាយអិនខេប៊ីនិងការបង្ក្រាបហ្គិចស៊ីចក៏ត្រូវបានសម្របសម្រួលតាមរយៈΔFosB។20,35,51,91
គួរឱ្យភ្ញាក់ផ្អើល, ថ្នាំអាភៀននៃការរំលោភបំពានមានឥទ្ធិពលផ្ទុយនិងកាត់បន្ថយដង់ស៊ីតេឆ្អឹងខ្នង dendritic នៃណឺរ៉ូនប្រសាទស្ពឺមធ្យម.81 គេដឹងតិចតួចអំពីផលវិបាកនៃអាកប្បកិរិយានៃការសម្របខ្លួននេះនិងយន្តការម៉ូលេគុលដែលពាក់ព័ន្ធ។ បាតុភូតនេះគឺ ទោះយ៉ាងណាគួរឱ្យភ្ញាក់ផ្អើលដែលថា CREB និងΔFosBត្រូវបានបង្កឡើងដោយភ្នាក់ងាររំញោចនិងអាភៀនហើយទាំងពីរនេះត្រូវបានបង្កប់ដោយអាំងតង់ស៊ីតេឆ្អឹងខ្នងដែលរំញោចដោយដង់ស៊ីតេឆ្អឹងខ្នង NAc ។. នេះធ្វើឱ្យមានសំណួរថាតើថ្នាំអាភៀនបង្ក្រាបដង់ស៊ីតេឆ្អឹងខ្នងរបស់អេអាអេកទោះបីជាពួកគេត្រូវបានបង្កើតកត្តាទាំងនេះក៏ដោយ។
ទម្រង់សំខាន់នៃប្លាស្ទិករូបវិទ្យាដទៃទៀតដែលត្រូវបានគេឃើញក្នុងគំរូនៃការប្រើប្រាស់គ្រឿងញៀនគឺការកាត់បន្ថយទំហំរាងកាយរបស់សូម៉ាកោសិកាណឺត្រូនដូផូអ៊ីតដែលបង្កើតឡើងដោយរដ្ឋបាលអាភៀនរ៉ាំរ៉ៃ។.77,92,93 ការបន្សាំស្រដៀងគ្នាកើតឡើងក្នុងការឆ្លើយតបទៅនឹង cannabinoids ។94 ការរួញតូចនៃណឺរ៉ូនឌីប៉ូតាមីនដែលកើតឡើងជាមួយនឹងការគ្រប់គ្រងអាភៀន។93 ហើយត្រូវបានចងក្រងជាឯកសារក្នុងការញៀនថ្នាំហេរ៉ូអ៊ីនរបស់មនុស្ស77 ហាក់ដូចជាសំរបសំរួលការអត់ឱនរង្វាន់ហើយត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការកាត់បន្ថយការបញ្ចេញនូវសារធាតុដូប៉ាមីននៅក្នុងអេអឹមខេ។. ភស្តុតាងដែលអាចពិចារណាបានឥឡូវនេះបង្ហាញថាការកាត់បន្ថយទំហំសូម៉ាកោសិកាត្រូវបានសម្របសម្រួលដោយការបង្ក្រាបអាភៀននៃការបញ្ចេញកោសិកាប្រសាទ (BDNF) ដែលទាក់ទងនឹងកោសិកាប្រសាទទាំងនេះ។ យើងបានភ្ជាប់ដោយផ្ទាល់នូវការដកចេញនូវអាភៀននៃការគាំទ្រ BDNF និងការបង្រួម VTA ណឺរ៉ូនដើម្បីកាត់បន្ថយសកម្មភាពរបស់ cascades សញ្ញា BDNF ចុះក្រោមនៅក្នុងណឺរ៉ូនឌីដូថុនដែលកាត់បន្ថយសកម្មភាពពិសេសរបស់ IRS2 (អាំងស៊ុយលីនអេស្ត្រូស្យូមអេស - អេសអិន), AKT (សឺរ - threonine) ។ kinase) និង TORC2 (គោលដៅនៃ rapamycin-2 ដែលមិនងាយនឹង rapamycin) ។77,93 យើងក៏បានផ្សារភ្ជាប់ការធ្លាក់ចុះនៃ BDNF ដែលផ្តល់សញ្ញាដោយផ្ទាល់ទៅនឹងការកើនឡើងគួរឱ្យរំភើបដែលម៉ូត្រុនបង្កើតនៅក្នុងណឺរ៉ូនទាំងនេះដូចដែលបានកត់សម្គាល់មុន។77,78 ជាការពិតការថយចុះទំហំសូម៉ាកោសិកានិងការបង្កើនភាពរំភើបត្រូវបានភ្ជាប់យ៉ាងតឹងរ៉ឹងដូចជាការបញ្ចូលមួយនាំទៅរកការផ្ទុយនិងមួយទៀត។ ការគ្រប់គ្រងនេះលើភាពរំជើបរំជួលរបស់កោសិកាពាក់ព័ន្ធនឹងការបង្ក្រាបរបស់ខេ។+ ឆានែលនិងរបស់ GABA ។A ចរន្តនៅក្នុងណឺរ៉ូនទាំងនេះ។
តួនាទីនេះសម្រាប់ប៊ី។ អេ។ អិល។ អេ។ ក្នុងការគ្រប់គ្រងការឆ្លើយតបម៉ូលេគុលនៅកម្រិត VTA ផ្ទុយពីការចូលរួមខុសគ្នាខ្លាំងរបស់វានៅក្នុងសកម្មភាពរបស់កូកាអ៊ីននិងសារធាតុរំញោចផ្សេងៗ។ សារធាតុរំញោចជំរុញឱ្យមានសញ្ញា BDNF ដល់ NAc ដែលជាផលប៉ះពាល់ដោយសារតែការកើនឡើងនៃការសំយោគមូលដ្ឋាននៃ BDNF ក៏ដូចជាការកើនឡើងនូវការចេញផ្សាយពីតំបន់ដែលទាក់ទងគ្នាជាច្រើន។95 លើសពីនេះទៅទៀតការកើនឡើងនៃសញ្ញា BDNF នៅក្នុង NAc ប៉ុន្តែមិនមាននៅក្នុង VTA ត្រូវបានបង្ហាញដើម្បីលើកកម្ពស់ផលប៉ះពាល់អាកប្បកិរិយានៃថ្នាំទាំងនេះរួមទាំងការគ្រប់គ្រងខ្លួនឯង។95,96 បទបញ្ញត្តិផ្ទុយគ្នានៃសញ្ញា BDNF នៅក្នុងផ្លូវ VTA-NAc ដោយ opiates និងថ្នាំរំញោចបង្កើនលទ្ធភាពដែលថាភាពខុសគ្នាបែបនេះសម្របសម្រួលបទប្បញ្ញត្តិផ្ទុយគ្នានៃថ្នាំនៃឆ្អឹងខ្នង dendritic NAc ដែលជាលទ្ធភាពដែលឥឡូវនេះកំពុងស្ថិតក្រោមការស៊ើបអង្កេត។
ទិសដៅអនាគត
និទានកថាខាងលើគូសបញ្ជាក់អំពីការជឿនលឿនយ៉ាងខ្លាំងដែលបានបង្កើតឡើងក្នុងការស្វែងយល់អំពីការបន្សាំនៃម៉ូលេគុលនិងកោសិកាដែលកើតឡើងនៅក្នុងតំបន់រង្វាន់ខួរក្បាលដើម្បីឆ្លើយតបទៅនឹងការប៉ះពាល់គ្រឿងញៀនម្តងហើយម្តងទៀតនិងទាក់ទងទៅនឹងការសម្របខ្លួនរបស់បុគ្គលទៅនឹងលក្ខណៈពិសេសនៃអាកប្បកិរិយានៃការញៀននៅក្នុងគំរូសត្វ។ ។ ទោះបីមានការជឿនលឿនទាំងនេះក៏ដោយក៏សំណួរចំបងនៅតែមាន។ ចំណេះដឹងដែលមានស្រាប់របស់យើងភាគច្រើនផ្តោតលើ VTA និង NAc ដែលមានព័ត៌មានតិចជាងមុនអំពីតំបន់ខួរក្បាលដែលមានអវយវៈសំខាន់ៗដែលមានសារៈសំខាន់សម្រាប់ការញៀនគ្រឿងញៀនផងដែរ។ លើសពីនេះទៀតរាល់ការសាកល្បងពិសោធន៍ទាំងអស់នៃតួនាទីមូលហេតុនៃការបន្សាំនឹងម៉ូលេគុល - កោសិកាក្នុងឥរិយាបថទាក់ទងនឹងគ្រឿងញៀនបានរៀបចំការបន្សាំរបស់បុគ្គលម្នាក់ៗក្នុងពេលតែមួយ។ ដើម្បីសម្របខ្លួនទៅនឹងការបន្សាំជាច្រើនក្នុងពេលតែមួយពិតជាពិបាកជាងប៉ុន្តែវាក៏ចាំបាច់ដែរព្រោះយើងដឹងថាគ្រឿងញៀនផលិតនូវប្រភេទនៃការផ្លាស់ប្តូរខុសៗគ្នាជាច្រើនសូម្បីតែនៅក្នុងកោសិកាសរសៃប្រសាទនីមួយៗដែលទំនងជាប្រមូលផ្តុំដោយវិធីស្មុគស្មាញដើម្បីជះឥទ្ធិពលដល់ឥរិយាបថ។ វិធីសាស្រ្តជីវវិទ្យាប្រព័ន្ធមួយនេះមានសារៈសំខាន់ណាស់ក្នុងការបង្ក្រាបការជម្រុញជីវសាស្រ្ត។ នៅចុងបញ្ចប់ការខិតខំស្វែងយល់ពីយន្តការម៉ូលេគុល - កោសិកានៃការចងចាំទាក់ទងនឹងការញៀនរកឃើញខ្លួនឯងនៅចំណុចដែលកិច្ចខិតខំប្រឹងប្រែងផ្សេងទៀតដើម្បីយល់ពីមូលដ្ឋានជីវសាស្ត្រនៃការចងចាំអាកប្បកិរិយាកំពុងតែតស៊ូ: សមត្ថភាពរបស់យើងទាក់ទងបាតុភូតជីវសាស្ត្រទៅនឹងការចងចាំអាកប្បកិរិយាស្មុគស្មាញនៅតែពិបាកខ្លាំង។ ការយកឈ្នះលើការបែងចែកនេះប្រហែលជាបញ្ហាប្រឈមធំបំផុតក្នុងវិស័យវិទ្យាសាស្ត្រប្រសាទ។
អក្សរកាត់និងអក្សរកាត់ដែលបានជ្រើសរើស។
- ន៉ាក
- nucleus accumbens
- CREB ។
- ធាតុឆ្លើយតបនៃ cAMP ភ្ជាប់ប្រូតេអ៊ីន។
- ΔFosB
- កត្តាប្រតិចារិកគ្រួសារ Fos ។
- VTA
- តំបន់ប្រដាប់បន្តោង
- AMPA
- អាស៊ីតα-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
- អិលធីឌី
- ការធ្លាក់ទឹកចិត្តរយៈពេលវែង។
- LTP
- protentiation រយៈពេលវែង។
- BDNF
- កត្តា neurotrophic កើតចេញពីខួរក្បាល
- NKkB ។
- កត្តានុយក្លេអ៊ែរខេប៊ី។
សេចក្តីយោង