- 1Centro tarpdisciplininis de Valparaíso neuropiencias, Valparaíso Universidad, Valparaíso, Čilė
- 2Departamento de Neurociencia, Medicinos fakultetas, Čilės universitetas, Santjagas, Čilė
- 3Núcleo Milenio NUMIND Neuropsichiatrinių sutrikimų biologija, Universidad de Valparaíso, Valparaíso, Čilė
- 4Pontificia Universidad Católica de Chile, Santiago, Čilė, Biologinių mokslų fakulteto Ląstelinės ir molekulinės biologijos katedra
- 5Pontificia Universidad Católica de Chile, Santjagas, Čilė, Chemijos fakulteto Farmacijos katedra ir Tarpdisciplininis neuromokslų centras
Į tikslą orientuoto elgesio stiprumą reguliuoja vidurinių smegenų dopamino neuronai. Dopaminerginių grandinių funkcijos sutrikimai stebimi priklausomybės nuo narkotikų ir obsesinio-kompulsinio sutrikimo atvejais. Kompulsinis elgesys yra abiejų sutrikimų požymis, susijęs su padidėjusiu dopamino neurotransmisija. Vidurinių smegenų dopamino neuronų aktyvumą daugiausia reguliuoja homeostatinis dopamino poveikis per D2 receptorius (D2R), kurie sumažina neuronų sudegimą, taip pat dopamino sintezę ir išsiskyrimą. Dopamino perdavimą taip pat reguliuoja heterologinės neuromediatorių sistemos, tokios kaip kappa opioidų sistema. Didžioji dalis mūsų dabartinių žinių apie kappa opioidų sistemą ir jos įtaką dopamino perdavimui yra iš ikiklinikinių gyvūnų smegenų ligų modelių. 1988 m., Naudojant smegenų mikrodializę, buvo parodyta, kad ūmus Kappos opioidinių receptorių (KOR) aktyvinimas sumažina sinapsinį dopamino kiekį striatumoje. Šis slopinamasis KOR poveikis prieštarauja lengvinančiam narkotikų vartojimui dopamino išsiskyrimą, todėl KOR agonistai turi būti naudojami kaip farmakologinė terapija kompulsiniam vaistų vartojimui. Keista, bet po 30 metų KOR antagonistai yra siūlomi gydyti priklausomybę nuo narkotikų. Kas galėjo nutikti per tuos metus, paskatinusį šį drastišką paradigmos pasikeitimą? Surinkti įrodymai rodo, kad KOR poveikis sinapsinio dopamino lygiui yra sudėtingas, priklausomai nuo KOR aktyvavimo dažnio ir laiko, kai kiti stimuliuoja dopamino neuronai, taip pat nuo lyties ir rūšių skirtumų. Priešingai nei ūminis poveikis, atrodo, kad lėtinis KOR aktyvinimas palengvina dopamino neurotransmisiją ir dopamino sukeliamą elgesį. Priešingi veiksmai, atlikti dėl ūmaus ir lėtinio KOR aktyvavimo, buvo siejami su pradiniu neigiamu ir uždelstu atlygį sukeliančiu poveikiu narkotikų vartojimo metu. Pakartotinis D2R aktyvinimas sukelia kompulsinį elgesį taip pat sustiprina nuolatinį KOR aktyvavimą, kuris koreliuoja su sumažėjusiu sinapsiniu dopamino ir sensibilizuoto D2R lygiu. Taigi nuo laiko priklausomas KOR aktyvavimas daro tiesioginį poveikį dopamino lygiui ir daro įtaką motyvuoto elgesio derinimui. Šioje apžvalgoje analizuojamas kappa opioidų sistemos indėlis į kompulsinio elgesio dopaminerginius ryšius.
Įvadas
Dopaminerginė sistema esant kompulsiniam elgesiui
Prievarta - tai negalėjimas savarankiškai sustoti atlikti įprastą veiksmą, kurio rezultatas žinomas, nepaisant neigiamų padarinių (Robbins ir kt., 2012). Be kitų psichinių ligų, kompulsyvus elgesys yra obsesinio-kompulsinio sutrikimo (OKS) ir priklausomybės nuo narkotikų požymis. Elgesio tikrinimas labai dažnas obsesinio-kompulsinio spektro sutrikimų atvejais, kai būdingas nuolatinis tam tikros rutinos kartojimas stereotipiniu ar ritualiniu būdu (Williams ir kt., 2013). Įvairiausias įprastas elgesys (pvz., Tikrinimas, valymas, rankų plovimas ir tt) OKS sergantiems pacientams gali virsti kompulsyviu ir paprastai kyla dėl reaguojančių į obsesines ir kankinančias mintis, sukeliančias nerimą. Panašiai narkotikų ieškojimas ir vartojimas tampa priklausomi nuo narkomanų. Kaip ir sergant OKS, nerimas vaidina pagrindinį vaidmenį, sukeliantį kompulsyvų narkotikų vartojimą patyrusiems narkotikų vartotojams. Abiem atvejais pastebimi tie patys atlygio ir bausmės vykdymo sutrikimai (Figee ir kt., 2016 m), kuris paskatino kai kuriuos autorius aptarti OKS kaip priklausomybę nuo elgesio (Holden, 2001).
Vienas galimas kompulsyvaus elgesio mechanizmas yra įtrauktas į priklausomybės stimuliavimo ir sensibilizacijos teoriją, ty priklausomybės metu išsivysto sustiprinta narkotiko motyvacija („norėjimas“), nesusidarant sustiprintam maloniam („patinkančiam“) efektui (Berridge ir kt., 1989; Berridge ir Robinson, 2016). Nuolatinis atlygio (motyvacijos) įjautrinimas yra susijęs su skatinamuoju jautrinimu, susijusiu su narkotikų paieškomis. Atlygio / motyvacijos grandinę sudaro vidurinių smegenų dopamino neuronai substantia nigra (SN) ir ventralinė pagrindinė sritis (VTA), nukreiptos atitinkamai į nugaros ir vidurinę striatumo pakopas. Dopamino neuronai, kurie išsikiša į vidurinę striatinę ar branduolio akumuliatorius (NAc), tradiciškai buvo siejami su į tikslą orientuota elgsena, tuo tarpu dopamino neuronai, kurie išsikiša į nugaros striatumą, buvo siejami su įpročių įgijimu (Everitt ir Robbins, 2005; Išminčius, 2009; Yager ir kt., 2015 m; Volkow ir kt., 2017).
Graužikams stebimas atlygio (motyvacijos) grandinės jautrumas, nes palaipsniui padidėja lokomotorinis aktyvumas, kurį sukelia pakartotinis galimai priklausomybę sukeliančių vaistų fiksuotos dozės (Pierce ir Kalivas, 1997; Robinson ir Berridge, 2001). Lokomotorinis sensibilizavimas yra ilgalaikis reiškinys, nes jis pastebimas praėjus savaitėms, mėnesiams ir net metams po narkotikų vartojimo (Robinson ir Berridge, 1993). Anksti buvo pasiūlyta, kad jautrinant atlygio / motyvavimo schemą prisidedama prie priverstinio narkotikų paieškos (Robinson ir Berridge, 1993). Atitinkamai, dėl lokomotorinės sensibilizacijos palengvinamas savaiminis kokaino atstatymas (De Vries ir kt., 2002). Be to, žiurkėms, turinčioms daugiau galimybių savarankiškai vartoti kokainą, pasireiškia didesnis lokomotorinis atsakas į kokainą nei žiurkėms, kurių prieiga ribota (Ferrario ir kt., 2005). Be to, neurocheminiai pokyčiai, dėl kurių padidėja lokomotorinė sensibilizacija psichostimuliatoriams, taip pat stebimi kompulsyvių vaistų ieškant (Steketee ir Kalivas, 2011; Giuliano ir kt., 2019). Šie duomenys patvirtina ankstyvą siūlomą atitikimą tarp lokomotorinės sensibilizacijos ir kompulsinių vaistų, stebėtų žmonėms (Robinson ir Berridge, 1993; Vanderschuren ir Kalivas, 2000). Mechaniškai kartojamas narkotikų vartojimas sensibilizuoja mezolimbines dopamino grandines, didindamas dopaminerginį neurotransmisiją. Psichostimuliatoriai, tokie kaip kokainas ar amfetaminai, kurie blokuoja plazmos membranos dopamino pernešėją (DAT), sukelia didelį dopamino padidėjimą sinapsinėje erdvėje striatumoje ir NAc, tokiu būdu suaktyvindami judėjimą (Steketee ir Kalivas, 2011). Kaip ir priklausomybėje nuo narkotikų, jautrus dopamino apdovanojimų / motyvacijos ciklas prisideda prie kompulsinio elgesio, pastebimo OKS. Iš tikrųjų pakanka pakartotinio D2 dopamino receptorių (D2R) aktyvavimo, kad būtų galima sukelti lokomotorinį jautrinimą ir patikrinti elgesį tiek žiurkėms, tiek pelėms (Szechtman ir kt., 1998; Szechtman ir kt., 1999; Sun ir kt., 2019). Pakartotinis chinpirolio, D2R / D3R agonisto, vartojimas yra priimtas OKS modelis, nes jis pakartoja veido pagrįstumą, padidindamas kompulsyvų patikrinimą ir stereotipinį elgesį, prognozuojamą pagrįstumą, kaip matyti iš kompulsyvaus elgesio sumažėjimo po lėtinio gydymo serotoninu reabsorbcijos. inhibitoriai (SRI) ir konstrukto pagrįstumas, nes šiame modelyje dalyvaujančios smegenų struktūros yra bendros su patologijomis (Stuchlik ir kt., 2016; Szechtman ir kt., 2017). Apibendrinant galima pasakyti, kad pakartotinis dopamino perdavimo suaktyvinimas tiek prieš sinapsinius (dopamino atpalaidavimo), tiek po sinapsinius (D2R aktyvinimas) mechanizmus sukelia lokomotorinį jautrinimą ir kompulsyvų elgesį.
Kappa opioidų sistema yra viena iš labiausiai vyraujančių sistemų, kontroliuojančių dopamino pernešimą atlygio / motyvacijos grandinėje. Įrodymai rodo, kad kappa-opioidų perdavimas prieštarauja dopamino poveikiui; ūmus kappa opioidinių receptorių (KOR) aktyvinimas neutralizuoja psichostimuliatorių sukeltą lokomotorinį aktyvumą (Gray ir kt., 1999). Atvirkščiai, pakartotinis KOR aktyvinimas palaiko ir sustiprina kompulsyvų ir įprastą narkotikų vartojimą (Koob, 2013). Piktnaudžiavimo narkotikais vartojimas padidina homeostatinį kappa opioidų perdavimą, greičiausiai prisidedantį prie neigiamų emocinių disforijos būsenų (Koob, 2013) sukelia priverstinį narkotikų vartojimą (Chavkinas ir Koobas, 2016 m). Tiesą sakant, KOR blokada užkirto kelią nikotino atstatymui streso, bet ne narkotikų (Džeksonas ir kt., 2013), kokainas (Beardsley ir kt., 2005) ir etanolis (Sperling ir kt., 2010 m). Remiantis šiais duomenimis, KOR blokada panaikina amfetamino jautrinamų žiurkių dorsolateralinio striatumo dopaminerginius pokyčius, nekeisdama jų sustiprinto lokomotorinio atsako į vaistą (Azocar ir kt., 2019 m). Taigi, atrodo, kad KOR sistema sustiprina neigiamą pastiprinimą, padidindama narkotikų vertę. OKS metu neigiamą sustiprėjimą sukelia obsesijos, kurios sustiprina tam tikrą prievartą, kad būtų išvengta šios manijos. Nors neigiamas pastiprinimas nebuvo tiesiogiai patikrintas, jis gali turėti įtakos chinpirolio sensibilizacijai. Iš tiesų, D2R dalyvauja kuriant neigiamą armatūrą. Pvz., Pelėms, neturinčioms ilgos D2R izoformos, nebuvo išvengta vietos, kurioje būtų morfino atsitraukimo poros.Smith et al., 2002) ir pakartotinis gydymas chinpiroliu abstinencijos laikotarpiu atkuria kokainą ir heroiną, ieškant automatinio vartojimo paradigmos - poveikis, susijęs su sensibilizuotu chinpirolio lokalizavimu (De Vries ir kt., 2002), siūlydama bendrus psichostimuliatorių ir chinpirolio sukeltos sensibilizacijos mechanizmus. Be to, namų narvo, bet ne naujo narvo įvedimas į atviro lauko areną sumažina lokomotorinio jautrinimą ir kompulsinį tikrinimo elgesį (Szechtman ir kt., 2001), nurodant, kad saugos / pažįstami patarimai gali konkuruoti su neigiamais aplinkos ženklais, kurie skatina jautrumą. Panašiai kaip psichostimuliatorių sukeltas sensibilizavimas, pakartotinis KOR aktyvinimas palengvina lokomotorinę sensibilizaciją (Escobar ir kt., 2017 m) ir kompulsinis tikrinimo elgesys (Perreault ir kt., 2007) sukeltas pakartotinai naudojant chinpirolį. Reikia išsiaiškinti, ar šis stiprinantis poveikis yra padidėjusio neigiamo sustiprinimo pasekmė.
Neseniai atlikta išsami analizė rodo, kad kappa-opioidų sistemos poveikis dopaminerginiam perdavimui yra sudėtingas: jis priklauso nuo dopamino kelio (Margolis ir kt., 2006; Margolis ir kt., 2008) ir laiko tarpą tarp KOR receptoriaus aktyvacijos ir dopamino receptoriaus aktyvavimo (Chartoff ir kt., 2016). Atsižvelgiant į šį sudėtingumą, plačiai aptariamas galimas KOR ligandų terapinis panaudojimas. Buvo pasiūlyta, kad KOR agonistas gali būti kliniškai naudingas vaisto vartojimo metu, silpninantis vaisto sukeltą hiperdopaminergiją (Shippenberg ir kt., 2007). Kita vertus, KOR antagonistas gali būti naudingas gydant abstinencijos sindromą, kurį sukelia padidėjęs dinorfinų ekspresija po pakartotinio vaisto vartojimo (Wee ir Koob, 2010). Atitinkamai buvo pasiūlyta, kad KOR dalinis agonistas (Béguin ir kt., 2012 m) galėtų būti terapinė galimybė gydyti tiek kompulsyvų vaistų vartojimą, tiek abstinencijos simptomus priklausomiems asmenims (Chartoff ir kt., 2016; Callaghan ir kt., 2018). Šioje apžvalgoje mes analizuojame elgesio sensibilizacijos ir kompulsyvumo dopaminerginių koreliacijų priklausomybę nuo laiko / konteksto.
Anatominis ir funkcinis kappa opioidų ir dopaminerginių sistemų skerspjūvis striatos ir vidurinės smegenų srityse
Stuburo regionai
KOR yra su baltymais sujungti receptoriai, kurių baltymai yra stipriai išreikšti vidurinės smegenų dalies dopamino sistemoje (Mansour ir kt., 1996). Šie receptoriai priklauso opioidinių receptorių grupei, kurią sudaro mu (MOR), delta (DOR) ir kappa (KOR). Šių receptorių endogeniniai agonistai yra atitinkamai endorfinai, enkefalinas ir dinorfinas. Stuburyje dinorfiną sintetina dopamino D1 receptorių (D1R) turintys vidutinio dydžio neuronai (MSN), turintys pasikartojančius aksonus, aktyvinančius KOR iš tų pačių branduolių (Mansour ir kt., 1995). Žiurkės NAc elektroninės mikroskopijos vaizdai rodo, kad KOR daugiausia randama DAT turinčiose presinapsinėse struktūrose, o nedidelė KOR dalis yra dendrituose, skiriant DAT (Svingos ir kt., 2001; Kivell ir kt., 2014 m). Imunofluorescenciniai tyrimai, apibūdinantys presinapsinius-sinaptosominius preparatus iš NAc, rodo, kad KOR ir D2R pirmiausia yra kartu sinaptosomose, kuriose yra dopaminą sintetinančio fermento tirozino hidroksilazės (TH) (Escobar ir kt., 2017 m). Be to, KOR yra gausu NAc ir striatum ląstelių kūnuose ir yra kolokalas su D2Rs ląstelių subpopuliacijoje (Escobar ir kt., 2017 m). Remiantis genetinėmis ir molekulinėmis įžvalgomis, buvo pasiūlyta, kad 20% viso KOR jungimosi striatumoje būtų stebima DA terminaluose (Van't Veer ir kt., 2013 m). Be to, Tejeda ir kt. (2017 m.) parodė, kad tiek D1R, tiek D2R MSN išreiškia KOR, labiau pasirinkdami D1R turinčius MSN (Tejeda ir kt., 2017 m). Šie anatominiai duomenys rodo, kad KOR yra prieš ir po postsinapsinių, reguliuojančių dopamino neurotransmisiją atlygio / motyvacijos grandinėje.
Keli eksperimentiniai metodai rodo, kad KOR aktyvacija slopina dopamino išsiskyrimąe. Ūmus KOR aktyvinimas atliekant sisteminę injekciją arba vietinę agonistų infuziją sumažina tarpląstelinį dopamino kiekį NAc (Di Chiara ir Imperato, 1988; Spanagel ir kt., 1992; Fuentealba ir kt., 2006) ir nugaros striatum (Gehrke ir kt., 2008 m). Remiantis tonizuojančiu KOR slopinančiu dopamino neurotransmisijos poveikiu, tai ilgalaikio ir selektyvaus KOR antagonisto nor-binaltorphimino (nor-BNI) tiesioginė infuzija (Broadbear ir kt., 1994) padidina bazinį dopamino kiekį NAc (Spanagel ir kt., 1992) ir dopamino išsiskyrimas pakaušio dalyje (Azocar ir kt., 2019 m). Galutiniai KOR toninio dopamino slopinimo įrodymai buvo parodyti KOR išmušimo pelėms, kurioms nustatytas padidėjęs tarpląstelinis dopamino lygis striatumoje ir NAc (Chefer ir kt., 2005). Mechanizmai, atsakingi už dopamino išsiskyrimo KOR slopinimą, nėra visiškai išaiškinti. Tačiau gerai žinoma, kad aktyvavus KOR, padidėja K + ir sumažėja Ca2 + laidumas, taip sukeliant ląstelių hiperpoliarizaciją ir vezikulinių neuromediatorių išsiskyrimo blokadą (Bruchas ir Chavkinas, 2010 m; Margolis ir Karkhanis, 2019 m).
Be, in vitro ir in vivo Funkciniai duomenys rodo, kad KOR modifikuoja Dopamino tarpląstelinį lygį moduliuodamas DAT aktyvumą. Pavyzdžiui, dėl KOR aktyvavimo EM4 ląstelėse, kurios kartu išreiškia KOR ir DAT, padidėja dopamino įsisavinimas, matuojant voltamperimetru (Kivell ir kt., 2014 m). An ex vivo analizė, naudojant voltamperometriją ir neskaidytiems audiniams, parodė, kad sisteminė KOR agonisto U-69593 injekcija padidino dopamino įsisavinimą NAc (Thompson ir kt., 2000). Panašus naujausias straipsnis rodo, kad „Nor-BNI“ neleidžia didinti dopamino įsisavinimo vidurinėje ir nugaros dalyje, kurį sukelia ūmus sisteminis MP1104, mišraus Kappa / Delta opioidų receptorių agonisto, injekcija (Atigari ir kt., 2019 m). Nepaisant to, KOR aktyvacijos poveikis dopamino įsisavinimui dar nėra iki galo išaiškintas. Sistemingai skiriant KOR dalinį agonistą nalmefeną, priklausomai nuo striatos dopamino suvartojimo dozės sumažėjo, kiekybiškai įvertinant greitojo skenavimo ciklinę voltamometriją (FSCV) (Rose ir kt., 2016). Suaugusiems žiurkių patinams atlikus tiesioginę mikrodializę, KOR blokavimas padidėjo ekstrahavimo frakcija (Ed), o tai yra netiesioginis dopamino įsisavinimo matas (Chefer ir kt., 2006; Azocar ir kt., 2019 m), leidžia daryti prielaidą, kad toninis KOR aktyvinimas slopina DAT aktyvumą (dopamino įsisavinimas). Šie rezultatai pabrėžia sudėtingą endogeninio KOR aktyvumo įtaką dopamino įsisavinimui kontroliuojant dopamino tarpląstelinį lygį. Didesnės laikinės skiriamosios gebos metodai, tokie kaip FSCV, neįrodė KOR įtakos dopamino įsisavinimui (Ebner et al., 2010 m; Ehrichas ir kt., 2015 m; Hoffman ir kt., 2016), leidžiant manyti, kad KOR, stiprinančiam DAT aktyvumą strijaus regionuose, reikia inkubacinio periodo. linijos (Kivell ir kt., 2014 m).
Vidurinės smegenų regionai
Autoradiografiniai tyrimai, atlikti žiurkės vidurinėje smegenyse, rodo reikšmingą KOR ryšį su SN ir VTA rostrokoadaline ašimi (Speciale ir kt., 1993 m). Kita vertus, elektroninės mikroskopijos duomenys rodo, kad dinorfinų turintys terminalai sinapsuoja tiesiai ant TH teigiamų dendritų SN ir VTA (Sesack ir Pickel, 1992), teigdamas, kad KOR lokalizuojasi dopamino neuronų somatodendritiniuose skyriuose. Striatal D1R turintys MSN yra vienas iš dynorfinų įvadų į smegenų vidurinės smegenų dopamino neuronus. Įdomu tai, kad KOR blokavimas nekeičia slopinančio D1R-MSN poveikio VTA dopamino neuronams, nurodydamas, kad šį slopinimą skatina GABA (Edwards et al, 2017 m). KOR moduliuoja dopamino vidurinių smegenų neuronų somatodendritinius atsakus. Elektrofiziologiniai tyrimai rodo, kad KOR aktyvacija VTA hiperpolarizuoja ir sumažina dopamino neuronų spontaninį sudeginimo greitį (Margolis ir kt., 2003). Taigi KOR agonistų infuzija sumažina somatodendritinį dopamino ištekėjimą (Smith et al., 1992; Dalmanas ir O'Malley, 1999 m). Tačiau atrodo, kad šis slopinantis KOR poveikis dopamino neuronams priklauso nuo grandinės. Kapa-opioidų agonistų infuzija VTA sumažina dopamino išsiskyrimą medialinėje priešfrontalinėje žievėje (mPFC) (Margolis ir kt., 2006), bet ne NAc (Devine ir kt., 1993; Margolis ir kt., 2006 m). Be to, Margolis ir kt. (2006 m.) nustatė, kad KOR slopina VTA dopamino neuronus, kurie išsikiša į mPFC ir bazolateralinę amigdalą, bet ne tuos, kurie projektuoja NAc. Tais pačiais metais „Ford“ ir kt. (2006 m.) parodė, kad KOR agonistų pritaikymas pelių VTA skiltelėse sukėlė didesnę dopamino neuronų, einančių į NAc, išorinę srovę, palyginti su tais, kurie išsikiša į bazolateralinę amygdalą, parodydami, kad KOR labiau slopina dopamino neuronus, kurie skleidžia NAc, nei į amygdalą. Be to, KOR aktyvinimas sumažina sužadinimo amplitudę (Margolis ir kt., 2005) ir slopinantis (Ford ir kt., 2007) postsinapsinės srovės į smegenų vidurinės grandies dopamino neuronus. Skirtumai tarp rūšių ir kompleksinių efektinių VTA provokacijų į mPFC ir NAc (Van Bockstaele ir Pickel, 1995; Carr ir Sesack, 2000) sunku nustatyti, ar KOR selektyviai slopina kai kurias VTA neuronų dopamino populiacijas. Nepaisant to, čia apibendrinti duomenys rodo, kad KOR yra dopamino neuronų somoje ir terminaluose, taip pat juos reguliuojančiuose įvaduose, taigi, puikioje padėtyje kontroliuoti vidurinių smegenų dopamino neuronų sinapsinį aktyvumą.
KOR vaidmuo kontroliuojant dopamino neurotransmisiją psichostimuliatorių sukeltame sensibilizavime ir kompulsiniame elgesyje
Priklausomybė nuo narkotikų yra procesas, kuris iš pradžių apima impulsyvų narkotikų ieškojimą, siejamą su jų teigiamą poveikį stiprinančiu poveikiu. Kita vertus, kompulsyvumas yra asmenybės bruožas, pastebimas narkomanams. Buvo pasiūlyta keletas neuroadaptinių priemonių dopaminerginiais būdais siekiant atsižvelgti į kompulsyvų narkotikų paiešką ir vartojimą po pakartotinio narkotikų vartojimo (Everitt ir Robbins, 2005; Koob ir Volkow, 2016). Viena iš siūlomų hipotezių, lemiančių kompulsinį narkotikų vartojimą, yra jo neigiamą poveikį sukeliančio poveikio jautrinimas (Koob, 2013). Kapa opioidų sistemos, slopinančios dopamino išsiskyrimą, kontrolė galėtų prisidėti prie piktnaudžiavimo narkotikų neigiamą poveikį sukeliančių savybių. Tačiau KOR aktyvavimo pasekmės dopamino neurotransmisijai ir kompulsinių vaistų paieškai atrodo sudėtingos ir akivaizdžiai prieštaringos. Iš tiesų, amfetamino ir kokaino sukeltas dopamino išsiskyrimą mažina kartu skiriant KOR agonistus (Heidbrederis ir Shippenbergas, 1994 m; Maisonneuve ir kt., 1994 m; Thompson ir kt., 2000) ir netgi sumažina kokaino administravimą (Negus ir kt., 1997 m). Be to, KOR slopina mezolimbinio kelio dopamino atpalaidavimą, reaguodamas į nuolatinį post-sinapsinio D1R aktyvavimą, kuris vyksta pakartotinai veikiant psichostimuliatoriams (Cole ir kt., 1995; „Nestler“, „2001“). Paradoksalu, bet KOR aktyvinimas taip pat gali palengvinti dopamino išsiskyrimą atlygio / motyvacijos kelyje (Fuentealba ir kt., 2006; Fuentealba ir kt., 2007) ir psichostimuliatorių vartojimas (Wee ir kt., 2009). Fuentealba ir kt. (2007 m.) parodė, kad po keturių dienų, vartojusių U69593, KOR agonistą, padidėjo amfetamino sukeltas dopamino išsiskyrimas NAc. Neseniai buvo parodyta, kad KOR blokavimas panaikina dopamino išsiskyrimo ir įsisavinimo pokyčius nugaros juosmeninėje dalyje, vykstančius amfetamino sukeltos lokomotorinės sensibilizacijos metu (Azocar ir kt., 2019 m). Iš viso šie duomenys rodo, kad KOR aktyvinimas taip pat gali prisidėti prie teigiamų narkotikų savybių sustiprinimo (Chartoff ir kt., 2016).
Be to, atrodo, kad KOR aktyvinimas taip pat prisideda prie kompulsyvių narkotikų paieškų; KOR blokada mažina kokainą (Wee ir kt., 2009), heroinas (Schlosburg et al., 2013 m) ir metamfetaminas (Whitfield ir kt., 2015 m) suvartojimas žiurkėms su neribota prieiga prie vaisto (Wee ir kt., 2009 m). Šis poveikis taip pat pastebimas ieškant narkotikų dėl streso. Pvz., KOR išmušimo pelės nepasižymėjo kokaino pasirinkimo vieta po priverstinio maudymo streso (McLaughlin ir kt., 2006a). KOR blokavimas silpnina nikotino vietos pasirinkimą, kurį sukelia priverstinis plaukimo streso poveikis (Smith et al., 2012). Įdomu tai, kad KOR blokavimas susilpnina kokaino ir nikotino paieškas, kurias sukelia stresas, tačiau neturėjo įtakos paieškoms, kurias sukelia narkotikų iššūkis (Beardsley ir kt., 2005; Džeksonas ir kt., 2013). Atrodo, kad šį streso sukeltą palengvinamąjį KOR poveikį daro atlygio / motyvacijos grandinė (Shippenberg ir kt., 2007; Wee ir Koob, 2010). Elegantiškame tyrime, kurį atliko daktarė Kauer ir jos grupė, buvo parodyta, kad KOR blokavimas VTA anksčiau arba po ūmaus streso slopina kokaino ieškojimą iš naujo - poveikį, susijusį su ilgalaikio slopinamųjų sinapsių potencialo padidėjimas dopamino neuronuose (Graziane ir kt., 2013 m; Polteris ir kt., 2014 m).
Atrodo, kad KOR palengvina psichostimuliatorių vartojimą, atsižvelgiant į narkotikų poveikio laiką. KOR agonisto U50488 paskyrimas 1 val. Prieš kokaino ekspoziciją sustiprina tiek kokaino pasirinkimą, tiek santykinį dopamino išsiskyrimą, kurį sukelia kokainas NAc, tuo tarpu priešingas poveikis pastebimas skiriant 15 min. Prieš (McLaughlin ir kt., 2006a; Ehrichas ir kt., 2014 m). Naudojant intrakranijinę savęs stimuliaciją Chartoff ir kt. (2016 m.) pastebėjo, kad KOR agonistas Salvinorinas A turi pradinį neigiamą ir uždelstą atsilyginti poveikį, lydimą atitinkamai sumažėjusio ir padidėjusio stimuliuojamo dopamino išsiskyrimo NAc. Visi šie duomenys rodo nuo laiko priklausomą KOR aktyvacijos poveikį naudingosioms kokaino savybėms ir nurodo streso sukeltą KOR aktyvavimą kaip pagrindinį veiksnį, skatinant kompulsyvų narkotikų vartojimą.
Chinpirolio sukeltas lokomotorinis jautrinimas ir kompulsinis elgesys
Faktus, kad dopamino sistema dalyvauja sensibilizacijos ir kompulsyvumo susidaryme, sustiprina elgesys, stebimas graužikams, gydytiems D2R agonistu, chinpiroliu. Trumpai tariant, D2R yra receptoriai, sujungti su Gi, išreikšti atlygio / motyvacijos grandinėje; jie yra išreikšti somatodendritiškai ir dopamino neuronų aksonų terminaluose (Sesack ir kt., 1994), o jo aktyvinimas sumažina tarpląstelinio dopamino kiekį (Imperato ir Di Chiara, 1988). Stuburyje D2R taip pat yra postsinapsiniu būdu ant vidutinio smaigalio neuronų (Sesack ir kt., 1994), o jo aktyvacija slopina netiesioginį kelią, leidžiantį judėti.
Gydytojas Henry Szechtmanas pradėjo tyrinėti chinpirolio poveikį žiurkių elgesiui, pasibaigusiam 1980 m. Vartojant mažas dozes (0.03 mg / kg), jis mažina judėjimo aktyvumą, o vartojant didesnes (> 0.5 mg / kg) - didėja. (Eilamas ir Szechtmanas, 1989 m). Šis poveikis yra susijęs su didelio afiniškumo presinapsinių D2R ir mažo afiniškumo postsinapsinių D2R aktyvinimu (Usiello ir kt., 2000). Netikėtai pakartotinis (kas antrą dieną) chinpirolio vartojimas sukelia laipsnišką ir nuolatinį lokalizacijos padidėjimą, panašų į psichostimuliatorių sukeltą lokomotorinį jautrinimą (Szechtman ir kt., 1993; Szechtman ir kt., 1994). Nustatyta, kad lokomotorinis jautrinantis poveikis priklauso nuo D2R, nes pelėms, kurioms trūksta šio receptoriaus, lokomotorinis jautrumas chinpiroliui neatsiranda (Escobar ir kt., 2015 m).
Dešimtojo dešimtmečio pradžioje Szechtmanas ir Eilamas pranešė, kad žiurkės kartu su judėjimo jautrinimu jautrino stereotipinį elgesį, kuris sustiprėja kiekvieną kartą vartojant kvinpirolį (Eilamas ir Szechtmanas, 1989 m; Szechtman ir kt., 1993). Šiandien chinpirolio pakartotinis vartojimas yra patvirtintas OKS modelis (Szechtman ir kt., 1999; Szechtman ir kt., 2001; Eilamas ir Szechtmanas, 2005 m; Stuchlik ir kt., 2016; Szechtman ir kt., 2017), pagrįstą pastebėjimu, kad žiurkių elgesys tampa vis labiau struktūruotas ir nelankstus, primenantis ritualinį elgesį, būdingą kompulsyviam tikrinimo elgesiui (Szechtman ir kt., 1998; Szechtman ir kt., 2017). Naujausi tyrimai rodo, kad kartotinis chinpirolis taip pat sukelia priverstinį elgesį su pelėmis, pavyzdžiui, kompulsinį patikrinimą (Sun ir kt., 2019), elgesio nelankstumas ir kompulsinis kramtymas (Asaoka ir kt., 2019 m), pastarąjį atmetė D2Rs blokada striatumoje, dar labiau patvirtindama, kad norint sukelti kompulsyvų elgesį reikia pakartotinio D2Rs aktyvavimo. Duomenys rodo, kad D2R vaidmuo vidurinės smegenų dalies dopamino keliuose sukelia lokomotorinį jautrinimą ir kompulsyvumą. Pakartotinis chinpirolio vartojimas užkerta kelią kokaino sukeltam stereotipiniam elgesiui (Thompson ir kt., 2010) ir amfetamino lokomotorinis poveikis (Cope ir kt., 2010), sustiprindamas mintį, kad D2Rs aktyvinimas yra psichostimuliatorių sukeltas sensibilizacijos pagrindas, ir siūlydamas bendrą chinpirolio ir psichostimuliatorių sukeltos sensibilizacijos mechanizmą. Įdomu tai, kad pakartotinio D2R aktyvavimo jautrusis poveikis atrodo stipresnis nei tą sukelia psichostimuliatoriai, nes kiekviena žiurkė, gydoma chinpiroliu, patiria lokomotorinį jautrinimą (Escobar ir kt., 2015 m), o maždaug šešiasdešimt procentų žiurkių jautrina amfetaminui (Escobar ir kt., 2012 m; Casanova ir kt., 2013 m).
Pakartotinai suaktyvinus D2R, elgesio jautrinimą lydi adaptacija atlygio / motyvacijos grandinėje. Žiurkėms, jautrioms chinpirolio, NAc dopaminerginis tonusas yra mažesnis, stebimas kaip sumažėjęs bazinis (Koeltzow ir kt., 2003 m) ir skatinamas tonizuojantis ir fazinis dopamino išsiskyrimas (Escobar ir kt., 2015 m), nurodant sumažėjusį dopamino vidurio smegenų grandinės dopamino atpalaidavimo pajėgumą. Sinapsinio dopamino kiekį NAc kontroliuoja tiek DAT, tiek dopamino neuronų aktyvumas (Goto ir Grace, 2008), kuris in vivo susideda iš tonizuojančio ir sudegusio (Wilson ir kt., 1977; Grace ir Bunney, 1980). Ankstesnės ataskaitos rodo, kad žiurkėms, jautrioms nuo chinpirolio, tonizuojantis ir sprogstantis VTA rodo mažesnį dopamino neuronų skaičių (Sesia ir kt., 2013 m). Šie duomenys kartu rodo, kad dopamino išsiskyrimo sumažėjimas po sensibilizacijos su chinpiroliu yra sumažėjęs bendras dopamino neuronų aktyvumas. Dėl sumažėjusio NAc dopaminerginio tono D2R jautrumas gali būti kompulsinis elgesys ir jautrus lokomotorinis aktyvumas, kurį sukelia pakartotinis gydymas chinpiroliu. Iš tiesų žiurkėms, jautrioms chinpirolio, padidėja dopamino D2R (Culver ir kt., 2008 m) ir padidėjęs šių receptorių afinitetas (Perreault ir kt., 2007), palaikydamas šią hipotezę.
KOR-dopamino sąveika su chinpirolio sukeltu kompulsiniu elgesiu
Pradiniai tyrimai apie KOR vaidmenį D2R sukeltame kompulsiniame elgesyje taip pat buvo iš Szechtmano laboratorijos. Ši grupė ištyrė KOR agonisto U69593 kartu su kvinpirolu vartojimą dėl judėjimo aktyvumo. Konkrečiai, autoriai suleido žiurkėms poodines injekcijas su U69593 ir chinpirolio mišiniu, kol bus atlikta 8–10 injekcijų. Priešingai nei hipolokomotorinis vien U69593 poveikis, hiperlokomocija buvo pastebėta vartojant kartu su mažomis (presinapsinėmis) ir didelėmis (postsinapsinėmis) kvinpirolio dozėmis. U69593 pakeitė presinaptinės chinpirolio dozės hipopolomotorinį poveikį į hiperlokomociją ir sustiprino postinapsinės chinpirolio dozės hiperlokomotorinį poveikį (Perreault ir kt., 2006). Kartu suaktyvinant KOR, taip pat paspartėjo lokomotorinių jautrinimų indukcija ir sustiprėjo D2Rs aktyvacijos poveikis, nes dvigubo gydymo būdu pasiekta didžiausia lokomotyva dubliuoja vien chinpirolio sukeltą lokomotorinį poveikį (Perreault ir kt., 2006; Escobar ir kt., 2017 m). Bendras KOR aktyvinimas taip pat pagreitina kompulsinio tikrinimo elgesio įgijimą (Perreault ir kt., 2007). Šis potencialus KOR poveikis chinpirolio sukeltai elgsenai reikalauja pakartotinio KOR aktyvavimo. Tiesą sakant, ūminė KOR agonisto U69593 injekcija toliau nepakeitė žiurkių, jautrintų chinpiroliu, lokomotorinio aktyvumo (Escobar ir kt., 2017 m). KOR stiprinančio D2R sukeltos sensibilizacijos mechanizmas nežinomas. Viena galimybė yra tai, kad pati endogeninė kappa opioidų sistema tarpininkauja nuo D2R priklausomai sensibilizacijai. Tačiau ši galimybė buvo atmesta parodžius, kad iš anksto paskyręs norBNI nepakeitė lokomotorinio sensibilizacijos chinpiroliui, o tai rodo, kad dinorfinas neišleidžiamas pasroviui D2R aktyvinant (Escobar ir kt., 2017 m). Šie duomenys neatmeta galimybės, kad dynorfinas gali turėti įtakos sensibilizuojančiam kompulsiniam elgesiui, pavyzdžiui, stresas skatina dinorfino išsiskyrimą ir aktyvuoti KOR, o tai palengvina kompulsinį elgesį (McLaughlin ir kt., 2003; McLaughlin ir kt., 2006a; McLaughlin ir kt., 2006b).
Tarp D2R ir KOR esančių kryžminių elementų yra sudėtinga ir atrodo, kad tai priklauso nuo to, ar abiejų receptorių aktyvacija sutampa, ar laikinai atskirtos. Anatominiai duomenys rodo, kad D2Rs ir KOR skerspjūvis gali įvykti presinapsiniu būdu dopamino neuronų aksonuose ir somatose, taip pat postinaptiškai - striatumo MSN. Nors anatominiai duomenys neatmeta KOR, esančių kitų neurocheminių sistemų aksonuose, vaidmens, jie aiškiai rodo tiesioginį KOR, reguliuojančio D2R, vaidmenį. Ar ūmus, ar kartojamas, KOR aktyvinimas sumažina slopinančią D2Rs funkciją dopamino neuronuose. Elektrofiziologiniai tyrimai parodė, kad ūmus KOR aktyvavimas VTA ir SN dopamino neuronuose slopina D2R tarpininkaujamą slopinančią postsinapsinę srovę, o tai sukelia pre- ir postsinapsiniai mechanizmai, nes KOR mažina dopamino išsiskyrimą, o dinorfinas blokuoja vonioje slopinamą dopamino slopinamąjį poveikį (Ford ir kt., 2007). Neurocheminiai tyrimai parodė, kad pakartotinis KOR aktyvavimas blokuoja D2R sukeltą dopamino išsiskyrimą NAc (Fuentealba ir kt., 2006). Be to, dėl sutapto D2R ir KOR aktyvacijos sumažėja dopamino išsiskyrimo slopinimas NAc, palyginti su kiekvieno receptoriaus poveikiu atskirai (Escobar ir kt., 2017 m). Taigi presinapsiniai KOR neveikia papildomai arba sinergijoje su presinapsiniais D2R, atvirkščiai, KOR slopina arba slopina D2R slopinamąjį poveikį. Šis mechanizmas galėtų paaiškinti ūmios KOR agonistų dozės, susijusios su maža chinpirolio doze, lokomotorinį aktyvinamąjį poveikį (Perreault ir kt., 2006).
Neseniai atliktas tyrimas rodo, kad KOR aktyvacija VTA tarpininkauja kompulsiniam elgesiui, matuojamam kaip elgesio slopinimas ir marmuro užkasimas (Abraham et al., 2017 m), patvirtinančią mintį, kad KOR aktyvacija iš tikrųjų yra kompulsyvumo priežastis. Duomenis paskelbė Margolis ir kt. (2006 m.); 2008) rodo, kad KOR ir D2R sąveika turėtų vykti dopamino neuronuose, nukreipiančiuose į mPFC (Margolis ir kt., 2006; Margolis ir kt., 2008). Nepaisant to, „Ford“ ir kt. (2006 m.); 2007) nustatė, kad D2R tarpininkaujamo IPSC slopinimas KOR vyksta dopamino neuronuose, nukreipiančiuose į NAc (Ford ir kt., 2006; Ford ir kt., 2007). Šie duomenys kartu rodo, kad KOR sąveika su D2R dopamino neuronų somatodendritiniame skyriuje gali atsirasti dėl to paties dopamino neurono skerspjūvio. Ar tai atsitiks mezolimbinėse ar mezokortikinėse projekcijose, vis dar diskutuojama.
Pažymėtina, kad KOR buvo rastas NAc MSN (Escobar ir kt., 2017 m; Tejeda ir kt., 2017 m), taip nurodant, kad D2R sukeltas kompulsinis elgesys gali sustiprėti ir dėl tiesioginių veiksmų dopamino neuronų tikslinėms ląstelėms. Šiuo atžvilgiu verta paminėti, kad pakartotinis U69593 skyrimas padidina D2R kiekį didelio afiniteto būsenoje (Perreault ir kt., 2007). Neurocheminiai duomenys rodo, kad sumažėjęs tarpląstelinis dopamino kiekis yra susijęs su D2R jautrinimu. KOR koaktyvacija dar labiau nesumažina tarpląstelinio dopamino lygio NAc, jau sumažėjus kartojant D2R (Escobar ir kt., 2017 m), paneigdamas presinapsinių KOR vaidmenį, pagreitinantis ar sustiprinantis D2R jautrumą NAc, naudojant šį mechanizmą. Todėl KOR sužadina lėtai molekulinius mechanizmus, kurie dar labiau jautrina D2R neurocheminį ir elgesio poveikį, ir tai rodo, kad lokalizacijos sensibilizacijos padidėjimą gali lemti adaptyvus postsinapsinis, o ne presinapsinis poveikis. Šiuo atžvilgiu pakartotinis KOR aktyvinimas gali paskatinti D2R netiesioginio striatos kelio slopinimą, keičiant D1R / D2R balansą į D1R, sukeliantį kompulsyvumą (1 pav).
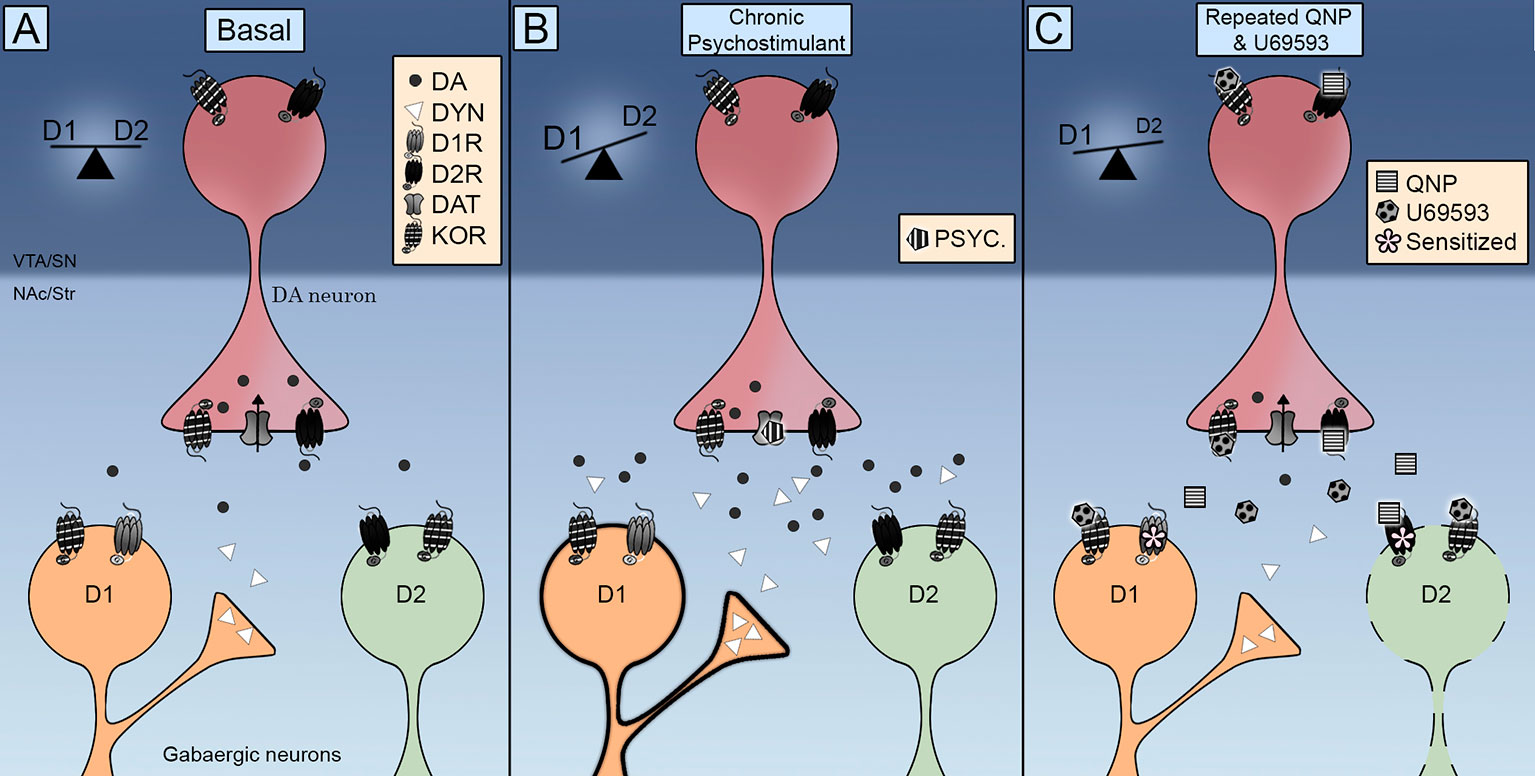 1 pav Integruota Kappa opioidinių receptorių (KOR) kontrolės schema tiesia (D1R) ir netiesiogine (D2R) striatos srityje. () KOR yra prieš sinaptiką dopamino terminaluose ir po sinaptikos - vidutinio dydžio neuronuose (MSN). Jo aktyvacija kontroliuoja tarpląstelinį dopamino lygį, o jo lokalizacija skatina sąveiką su dopamino pernešėjo (DAT) ir dopamino D2 receptoriais. (B) Kartotinį psichostimuliatoriaus poveikį lydi padidėjęs tarpląstelinio dopamino ir dinorfinų kiekis. Suaktyvinus D1 ir D2 receptorius, pusiausvyra keičiama į D1R tiesioginį kelią, skatinant lokomotorinį jautrinimą. (C) Kartu vartojant chinpirolį ir U69593, sumažėja tarpląstelinis dopamino kiekis. Kartu suaktyvinant KOR ir D2 receptorius, D2 netiesioginis kelias sukelia kompulsinį elgesį.
1 pav Integruota Kappa opioidinių receptorių (KOR) kontrolės schema tiesia (D1R) ir netiesiogine (D2R) striatos srityje. () KOR yra prieš sinaptiką dopamino terminaluose ir po sinaptikos - vidutinio dydžio neuronuose (MSN). Jo aktyvacija kontroliuoja tarpląstelinį dopamino lygį, o jo lokalizacija skatina sąveiką su dopamino pernešėjo (DAT) ir dopamino D2 receptoriais. (B) Kartotinį psichostimuliatoriaus poveikį lydi padidėjęs tarpląstelinio dopamino ir dinorfinų kiekis. Suaktyvinus D1 ir D2 receptorius, pusiausvyra keičiama į D1R tiesioginį kelią, skatinant lokomotorinį jautrinimą. (C) Kartu vartojant chinpirolį ir U69593, sumažėja tarpląstelinis dopamino kiekis. Kartu suaktyvinant KOR ir D2 receptorius, D2 netiesioginis kelias sukelia kompulsinį elgesį.
KOR-dopamino sąveikos lyties skirtumai kompulsiniame elgesyje
Klinikiniai tyrimai parodė lytinius skirtumus, susijusius su kompulsyviu elgesiu, įskaitant kompulsyvų narkotikų vartojimą. Anksčiau OCD simptomai pasireiškė vyrams, palyginti su moterimis (Mathis ir kt., 2011 m), kai moterys dažniau pasireiškia užterštumu ir valymo simptomais (Labad ir kt., 2008 m). Kalbant apie lyčių skirtumus priklausomybėje nuo narkotikų, klinikiniai duomenys rodo, kad nors narkotikai yra labiau paplitę vyrams, moterys labiau linkę į vyrus linkti į narkotikų vartojimą nei vyrai (Hernandez-Avila ir kt., 2004 m; Fattore ir Melis, 2016 m).
Pastaruoju metu ikiklinikiniai įrodymai labai išryškino klinikinių tyrimų metu pastebėtus neurobiologinius pagrindus, kuriais grindžiami lytiniai narkotikų vartojimo skirtumai (Beckeris ir Chartoffas, 2019 m). Ankstyvieji stebėjimai, naudojant nefluksinę srauto mikrodializę, parodė, kad tarpląstelinės dopamino koncentracija liemens striatumoje estrozo ciklo metu kinta, o proestrus ir estrus padidėja, palyginti su diestrus. Be to, nors kiaušidžių kaklelio metu sumažėja striatralinio dopamino tarpląstelinė koncentracija žiurkių patelėse, žiurkių patinų kastracija nekeičia dopamino striatralinės tarpląstelinės koncentracijos („Xiao and Becker“, 1994 m), nurodant svarbų kiaušidžių hormonų vaidmenį dopamino aktyvumui. Be to, moteriški hormonai reguliuoja atsaką į psichostimuliatorius. Ankstyvas in vitro Eksperimentai parodė, kad estradiolis kartu su progesteronu atstato amfetamino sukeltą dopamino išsiskyrimą iš striatos audinio, gauto iš kiaušidžių išbrėžtų žiurkių patelių (Becker ir Ramirez, 1981). Neseniai atlikti greitos skenavimo ciklinės voltamperometrijos tyrimai parodė, kad moterys, palyginti su vyrais, išskiria ir absorbuoja elektroparodą labiau (Walker ir kt., 2000). Šie dopamino neurotransmisijos lytiniai skirtumai gali sąlygoti didesnį kokaino ir amfetamino ieškojimą tarp moterų. (Roberts ir kt., 1989; Cox ir kt., 2013).
KOR reguliavimas tarpląsteliniame dopamino lygyje taip pat rodo lyčių skirtumus (Chartoffas ir Mavrikaki, 2015 m). Naudojant intrakranijinę savęs stimuliaciją ir ciklinę voltamperometriją, Conway ir kt. (2019 m.) parodė, kad mažesnis jautrumas KOR agonisto ūminiam anhedoniniam poveikiui, pastebėtam žiurkių patelėms, palyginti su patinėliais, lydimas silpnesnio stimuliuojamo dopamino išsiskyrimo NAc (Conway ir kt., 2019). Buvo teigiama, kad estradiolis prisideda prie neryškaus dopamino išsiskyrimo slopinimo žiurkių patelėms po KOR aktyvavimo (Abraham et al., 2018 m). Nors KOR ir dopamino signalizacijos šaltinis buvo tiriamas vyrams („Tejeda“ ir „Bonci“, 2019 m), šios sąveikos ir jos įtakos moterų priklausomybės procesui tyrimų nėra (Chartoffas ir Mavrikaki, 2015 m). Žiurkių patelėms ūmus KOR agonisto U69593 paskyrimas susilpnino kokaino sukeltą hiperkomokiją tiek kontrolinėms, tiek kiaušidžių kiaušintakmenėms turinčioms žiurkėms. Įdomu tai, kad pakartotinis U69593 vartojimas susilpnino kokaino sukeltą hiperlokomociją priklausomai nuo estradiolio (Puig-Ramos ir kt., 2008 m). Šie duomenys rodo, kad estradiolis daro KOR poveikį žiurkių patelėms, o tai gali būti susiję su lyties skirtumais reaguojant į stresą (Puig-Ramos ir kt., 2008 m). Neatsakytas klausimas, ar pakartotinis KOR aktyvinimas patelių žiurkėms palengvina stropinės dopamino išsiskyrimą, kaip pastebėta patinams.
Nors patelėms, palyginti su patinėliais, pastebimas palengvinamas psichostimuliatorių sukeltas dopamino išsiskyrimas, amfetamino lokomotorinės sensibilizacijos dopamino mechanizmų lyčių skirtumai nėra visiškai išaiškinti (Becker, 1999). Pakartotinis amfetamino poveikis sukelia didesnį lokomotorinį aktyvumą tiek paaugliams („Mathews“ ir „McCormick“, „2007“) ir suaugusios žiurkių patelės (Milesi-Hallé ir kt., 2007 m), kai žiurkių patelės paauglės, pakartotinai veikdamos amfetaminu, jautresnės lokomotorinės reakcijos. Naujagimių D2 receptorių aktyvinimas suintensyvino amfetamino sukeltą sensibilizaciją tik žiurkių patelėms (Brown et al., 2011). Kaip minėta anksčiau, žiurkių patinėliams buvo pastebėtas kartotinis D2 agonisto poveikis sukelia lokomotorinį jautrinimą ir kompulsinį elgesį (Dvorkin ir kt., 2006 m). Be to, kartu veikiantis KOR sustiprina lokomotorinį jautrinimą, kurį sukelia pakartotinis chinpirolio poveikis, palengvindamas D2 receptorių slopinamą kontrolę DA išsiskyrimo metu NAc (Escobar ir kt., 2017 m). Lyčių skirtumai, tokie kaip pastebėtas mažesnis jautrumas KOR slopinančiam poveikiui moterims dopamino išsiskyrimui (Conway ir kt., 2019) gali lemti skirtingą KOR indėlį ieškant kompulsyvių narkotikų.
Išvados
Kaip KOR moduliuoja dopamino signalizaciją motyvuotam elgesiui formuoti ir kada tai sukelia jautrų kompulsinį elgesį? Anatominiai duomenys rodo, kad KOR yra puikioje padėtyje kontroliuoti vidurinių smegenų dopamino neuronų sinapsinį aktyvumą. Funkciniai duomenys rodo, kad KOR kontroliuoja DAT ir D2R funkcionavimą, taip pat dopamino neuronų šaudymo greitį. Pirminiai įrodymai, kad dėl ūmaus KOR aktyvavimo sumažėja piktnaudžiavimo narkotikais sukeliamas dopamino išsiskyrimas, buvo papildyti duomenimis, rodančiais, kad pakartotinis KOR aktyvinimas palengvina dopamino išsiskyrimą ir kompulsyvų narkotikų ieškojimą. Dopamino signalizacijos pusiausvyra tiesiogiai ir netiesiogiai išstumia kelius iš striatos sritiesPav. 1A). Arba lėtinė stimuliacija psichostimuliatoriais, kurie padidina dopamino išsiskyrimą, aktyvindami ir D1R, ir D2R (Pav. 1B) arba chinpirolis, kurie aktyvina tik D2R (Pav. 1C) sukelia lokomotorinį sensibilizavimą ir kompulsyvų elgesį debilizuotu D2R netiesioginiu keliu, taigi balansas keičiamas į D1R tiesioginį kelią. KOR perdavimas pagerėja vartojant lėtinį psichostimuliatorių, padidėjant dinorfinui striatos D1 neuronuose (Pav. 1B). Padidėjęs KOR perdavimas yra imituojamas OKS farmakologiniame modelyje, skiriant U69593. Šis kartu vykdomas KOR aktyvumas dar labiau atitolina D2 netiesioginį kelią (Pav. 1C). Ateities tyrimai turėtų būti atlikti siekiant išsiaiškinti KOR aktyvacijos padarinius DAT aktyvumui, suprasti endogeninės KOR sistemos vaidmenį chinpirolio sukeltame kompulsyvume ir nustatyti KOR sistemos indėlį į lyčių skirtumus, stebimus kompulsyvaus elgesio metu.
Autoriaus įnašai
AE, MA ir JF prisidėjo prie rankraščio koncepcijos sudarymo. AE ir JF parašė pirmąjį rankraščio projektą su MA indėliu. MA ir JC prisidėjo prie kritinio rankraščio peržiūros ir redagavimo. Visi autoriai patvirtino jį leidybai.
Finansavimas
Šioje apžvalgoje cituojamų autorių darbai paremti FONDECYT stipendijų numeriais: 1110352 ir 1150200 MA; 1141088 į JF; DIPOG dotacija JF 391340281; FONDECYT doktorantas 3170497 JC ir 3190843 AE.
Interesų konfliktas
Autoriai teigia, kad tyrimas buvo atliktas nesant jokių komercinių ar finansinių santykių, kurie galėtų būti laikomi galimu interesų konfliktu.
Tvarkymo redaktorius šiuo metu rengia tyrimų temą su vienu iš autorių JF ir patvirtina, kad jokio kito bendradarbiavimo nėra.
Nuorodos
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL ir kt. (2017 m.). Kappos opioidinio receptoriaus aktyvacija dopamino neuronuose sutrikdo elgesio slopinimą. Neuropsychopharmacology 43 (2), 362–372. doi: 10.1038 / npp.2017.133
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ ir kt. (2018 m.). Dėl GRK2 estrogeno reguliavimo inaktyvinamas kappa opioidinių receptorių signalas, sukeliantis analgeziją, bet ne baimė. J. Neurosci. 38 (37), 8031–8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019 m.). Adenozino A2A receptorių antagonistas pagerina kartotinių chinpirolio sukeltų psichozių simptomus. eNeuro 6 (1), 1–16. ENEURO.0366 m. doi: 18.2019 / ENEURO.10.1523-0366
„Atigari“, DV, „Uprety“, R., Pasternak, G. G., Majumdar, S., Kivell, BM (2019). Mišrus kappa-delta opioidų receptorių agonistas MP1104 pasižymi antikokaino savybėmis ir turi mažesnį šalutinį poveikį žiurkėms. Neurofarmakologija 150, 217 – 228. doi: 10.1016 / j.neuropharm.2019.02.010
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, ME, Fuentealba, JA (2019). Kapa-opioidinio receptoriaus blokavimas panaikina dorsolateralinio striatum dopamino dinamikos pokyčius amfetamino sensibilizacijos metu. J. Neurochem. 148, 348–358. doi: 10.1111 / jnc.14612
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE ir kt. (2012). 12-epi-salvinorino A ir jo analogų diferencialinės signalizacijos savybės kappa opioidų receptoriuose. Bioorg. Med. Chem. Lett. 15; 22 (2), 1023–1026. doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Diferencinis naujojo kappa opioidų receptorių antagonisto JDTic poveikis atstatant kokaino paieškas, sukeltas pėdsakų stresorių, palyginti su kokaino PRIM, ir jo antidepresantų panašų poveikį žiurkėms. Psichofarmakolis. (Berlas) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Beckeris, JB, Chartoffas, E. (2019). Lyties skirtumai neuroniniuose mechanizmuose, tarpinančiuose atlygį ir priklausomybę. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Beckeris, JB, Ramirezas, VD (1981). Amfetamino lyties skirtumai paskatino katecholaminų išsiskyrimą iš žiurkės striatos audinio in vitro. Brain Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Beckeris, JB (1999). Lyties skirtumai dopaminerginėje funkcijoje striatumoje ir branduolio akumuliatoriuose. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Berridge, KC, Robinson, TE (2016). Mėgstamumas, norėjimas ir skatinamojo jautrumo priklausomybės teorija. Esu. Psychol. 71, 670–679. doi: 10.1037 / amp0000059
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Berridge, KC, Venier, IL, Robinson, TE (1989). 6-hidroksidopamino sukeltos afagijos skonio reaktyvumo analizė: poveikis dopamino funkcijos žadinimo ir anhedonijos hipotezėms. Behav. Neurosci. 103, 36 – 45. doi: 10.1037 / 0735-7044.103.1.36
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Skirtingas sistemiškai vartojamo norbanaltorphimino (nor-BNI) poveikis kappa-opioidų agonistams pelių riešo nustatymo tyrime. Psichofarmakologija 115, 311 – 319. doi: 10.1007 / BF02245071
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Brownas, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Jautrumas amfetamino lokomotoriuose ir sąlygota vieta paaugliams žiurkėms patinams ir patelėms, naujagimiams gydytiems chinpiroliu. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Bruchas, MR, Chavkin, C. (2010). Kinazės kaskados ir į ligandą nukreiptas signalas apie kappa opioidų receptorius. Psichofarmakolis. (Berlas) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Galimi opioidų receptorių vaidmenys motyvacijos ir didžiosios depresijos sutrikimų atveju. Prog. Brain Res. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Carr, DB, Sesack, SR (2000). GABA turintys neuronai žiurkės ventraliniame pagrindiniame rajone išsikiša į prefrontalinę žievę. Synapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Jautrinant amfetamino lokomotorius, padidėja didelis K + stimuliuoto dopamino išsiskyrimas žiurkės medialinėje prefrontalinėje žievėje. Behav. Brain Res. 237, 313 – 317. doi: 10.1016 / j.bbr.2012.09.052
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Chartoff, EH, Mavrikaki, M. (2015). Kapo opioidinių receptorių funkcijos lytiniai skirtumai ir jų galimas poveikis priklausomybei. Priekyje. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME, et al. (2016). Santykinis laikas tarp kappa opioidinių receptorių aktyvacijos ir kokaino lemia poveikį atlygiui ir dopamino išsiskyrimui. Neuropsychopharmacology 41, 989 – 1002. doi: 10.1038 / npp.2015.226
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Chavkinas, C., Koobas, GF (2016). Dinorfinas, disforija ir priklausomybė: priklausomybės stresas. Neuropsychopharmacology 41, 373 – 374. doi: 10.1038 / npp.2015.258
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Endogeninės kapa-opioidinių receptorių sistemos reguliuoja mezoakumbalinio dopamino dinamiką ir pažeidžiamumą kokainui. J. Neurosci. 25, 5029 – 5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Kiekybinė mikrodializė neto srauto metu leidžia nustatyti dopamino įsisavinimo padidėjimą ir sumažėjimą pelės branduolio akumuliatoriuose. J. Neurosci. Metodai 155, 187 – 193. doi: 10.1016 / j.jneumeth.2005.12.018
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Cole, R. L., Konradi, C., Douglass, J., Hyman, SE (1995). Neuronų adaptacija prie amfetamino ir dopamino: molekuliniai prodynorphin geno reguliavimo mechanizmai žiurkių striatumoje. Neuronas 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Conway, SM, Puttickas, D., Russellas, S., Poteris, D., Roitmanas, MF, Chartoffas, E. H. (2019). Moterys yra mažiau jautrios nei vyrai dėl kapos opioidinių receptorių aktyvacijos motyvacinį ir dopaminą slopinančio poveikio. Neurofarmakologija 146, 231 – 241. doi: 10.1016 / j.neuropharm.2018.12.002
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Cope“, ZA, „Huggins“, KN, „Sheppard“, AB, „Noel“, DM, Roane, DS, Brown, RW (2010). Gydymas naujagimiams chinpiroliu padidina lokomotorinę aktyvaciją ir dopamino išsiskyrimą branduolio branduolyje, reaguojant į amfetamino gydymą suaugus. Synapse 64, 289 – 300. doi: 10.1002 / syn.20729
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Cox, BM, Young, AB, See, RE, Reichel, CM (2013). Lyties skirtumai metamfetamino paieškoje žiurkėms: oksitocino poveikis. Psichoneuroendocrinologija 38, 2343 – 2353. doi: 10.1016 / j.psyneuen.2013.05.005
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Culveris, KE, Szechtmanas, H., Levantas, B. (2008). Pasikeitęs dopamino D2 receptorių jungimasis žiurkėms, kurių elgesys sensibilizuoja chinpirolį: išankstinio gydymo Ro 41-1049 poveikis. Euras. J. Pharmacol. 592, 67 – 72. doi: 10.1016 / j.ejphar.2008.06.101
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Dalmanas, FC, O'Malley, KL (1999). kappa-opioidų tolerancija ir priklausomybė dopaminerginių vidurinių smegenų neuronų kultūrose. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
De Vries, T. J., Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Dopamino D2 receptorių sąlygotas elgesys su kokainu ir heroinu, kurio ieškoma, priklauso nuo laiko ir yra susijęs su elgesio sensibilizacija. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Diferencinis ventrinių tegmentinių mu, delta ir kappa opioidinių receptorių dalyvavimas modifikuojant bazinį mezolimbinio dopamino išsiskyrimą: in vivo mikrodializės tyrimai. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Priešingas mu ir kappa opiatų agonistų poveikis dopamino atpalaidavimui laisvai judančių žiurkių branduolyje ir pakaušio kaudate. J. Pharmacol. Exp. Ther. 244, 1067 – 1080. doi: 10.1073 / pnas.85.14.5274
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Dvorkinas, A., „Perreault“, L. L., Szechtmanas, H. (2006). Pakartotinių dopamino agonisto chinpirolio injekcijų sukeltas kompulsinio tikrinimo vystymas ir laikinas organizavimas pagal obsesinio-kompulsinio sutrikimo gyvūnų modelį. Behav. Brain Res. 169, 303 – 311. doi: 10.1016 / j.bbr.2006.01.024
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Ebner“, SR, „Roitman“, MF, „Potter“, DN, „Rachlin“, AB, „Chartoff“, EH (2010). Kappa opioidų receptorių agonisto salvinorino A panašus į depresiją poveikis yra susijęs su sumažėjusiu fazinio dopamino išsiskyrimo akumuliatorių branduolyje. Psichofarmakolis. (Berlas) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., et al. (2017 m.). Grandinės specifiškumas VTA slopinančioje architektūroje reguliuoja kokaino sukeltą elgesį. Nat. Neurosci. 20 (3), 438–448. doi: 10.1038 / nn.4482
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Ehrichas, J. M., Phillipsas, PEM, Chavkinas, C. (2014). Kappos opioidinių receptorių aktyvinimas sustiprina kokaino sukeltą dopamino išsiskyrimo padidėjimą, in vivo užfiksuotą pelės branduolio akumuliatoriuose. Neuropsychopharmacology 39, 3036 – 3048. doi: 10.1038 / npp.2014.157
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, et al. (2015). Kappos opioidų receptorių sukeltam susirgimui reikia p38 MAPK aktyvacijos VTA dopamino neuronuose. J. Neurosci. 35, 12917 – 12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Eilam, D., Szechtman, H. (1989). Dviejų fazių D-2 agonisto chinpirolio poveikis judėjimui ir judesiams. Euras. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Eilam, D., Szechtman, H. (2005). Psichostimuliatorių sukeltas elgesys kaip obsesinio-kompulsinio sutrikimo gyvūnų modelis: etologinis požiūris į kompulsinių ritualų formą. CNS Spectr. 10, 191–202. doi: 10.1017 / S109285290001004X
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012). Pakartotinis gydymas kappa opioidinių receptorių agonistu U69593 panaikina padidintą K + sukeltą dopamino išsiskyrimą akumuliatorių branduolyje, tačiau nerodo lokomotorinio sensibilizacijos iš amfetamino jautrių žiurkių. Neurochem. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). Sumažėjęs dopamino ir gliutamato neuronų perdavimas žiurkių, kuriems jautrus chinpirolis, branduolio akumuliatoriuose, rodo uždaromąją D2 autoreceptorių funkciją. J. Neurochem. 134, 1081–1090. doi: 10.1111 / jnc.13209
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., et al. (2017 m.). Kapa opioidinio receptoriaus dopamino D2 receptorių funkcijos stiprinimo mechanizmai žiurkėms sukėlus chinpirolio sukeltą lokomotorinę jautrinimą. Vid. J. Neuropsychopharmacol. 20, 660–669. doi: 10.1093 / ijnp / pyx042
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Everitt, BJ, Robbins, TW (2005). Priklausomybės nuo narkotikų sustiprinimo neuroninės sistemos: nuo veiksmų iki įpročių iki priverstinio. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Fattore, L., Melis, M. (2016). Seksualiniai impulsyvaus ir kompulsyvaus elgesio skirtumai: dėmesys narkomanijai. Addict. Biol. 21 (5), 1043–1051. doi: 10.1111 / adb.12381
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Neuroninis ir elgesio plastiškumas, susijęs su perėjimu nuo kontroliuojamo prie padidėjusio kokaino vartojimo. Biol. Psy. 58 (9), 751–9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., et al. (2016). Priverstinis poveikis obsesinių-kompulsinių sutrikimų ir priklausomybių atvejais. Euras. Neuropsichofarmacol. 26, 856 – 868. doi: 10.1016 / j.euroneuro.2015.12.003
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Ford“, CP, „Mark“, GP, „Williams“, JT (2006). Mezolimbinių dopamino neuronų savybės ir opioidų slopinimas priklauso nuo tikslo vietos. J. Neurosci. 26, 2788 – 2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Ford“, CP, Beckstead, MJ, Williams, JT (2007). Kappa opioidas slopina somatodendritinį dopaminą slopinančias postsinapsines sroves. J. Neurophysiol. 97, 883 – 891. doi: 10.1152 / jn.00963.2006
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). Pakartotinis selektyvaus kappa-opioidinio receptoriaus agonisto U-69593 vartojimas padidina stimuliuojamą tarpląstelinį dopamino kiekį žiurkės branduolio akumuliatoriuose. J. Neurosci. Res. 84, 450 – 459. doi: 10.1002 / jnr.20890
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Padidėjęs lokomotorinis atsakas į amfetaminą, kurį sukelia pakartotinis selektyvaus kappa-opioidinio receptoriaus agonisto U-69593 vartojimas. Synapse 61, 771 – 777. doi: 10.1002 / syn.20424
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Ūmaus ir pakartotinio salvinorino A vartojimo poveikis dopamino funkcijai žiurkės nugaros juosmens srityje. Psichofarmakolis. (Berlas) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Kompulsinis alkoholio vartojimas atsiranda dėl nesugebėjimo atjungti dorsolateralinio striatos elgesio kontrolės. J. Neurosci. 39 (9), 1744–1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Goto, Y., Grace, AA (2008). Limbinės ir žievės informacijos apdorojimas akumuliatorių branduolyje. Tendencijos neurosci. 31, 552 – 558. doi: 10.1016 / j.tins.2008.08.002
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Grace, AA, Bunney, BS (1980). Nigraliniai dopamino neuronai: tarpląstelinis registravimas ir identifikavimas naudojant L-dopa injekciją ir histofluorescencija. Mokslas 210, 654 – 656. doi: 10.1126 / science.7433992
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Grey, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). Κ-opioidų agonistas U-69593 sumažina ūminį amfetamino sukeliamą elgesį ir dopamino bei glutamato dializuojamų dopamino ir glutamato koncentracijas vidurinėje žarnoje. J. Neurochem. 73, 1066 – 1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Kappos opioidiniai receptoriai reguliuoja streso sukeltą kokaino paiešką ir sinapsinį plastiškumą. Neuronas 77, 942 – 954. doi: 10.1016 / j.neuron.2012.12.034
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Heidbreder, CA, Shippenberg, TS (1994). U-69593 apsaugo nuo kokaino sensibilizacijos, normalizuodamas bazinius akumuliatorius dopaminą. Neuroreportas 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Nuo opioidų, kanapių ir alkoholio priklausomos moterys rodo spartesnį gydymą piktnaudžiavimu narkotinėmis medžiagomis. Priklauso nuo alkoholio. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Hoffman“, AF, „Spivak“, CE, „Lupica“, CR (2016). Padidėjęs dopamino išsiskyrimas dopamino transportavimo inhibitoriais, aprašytas ribotos difuzijos modeliu ir greitojo nuskaitymo cikliniu voltamperimetru. ACS Chem. Neurosci. 7, 700–709. doi: 10.1021 / acschemneuro.5b00277
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Holden, C. (2001). „Elgesio“ priklausomybės: ar jos egzistuoja? Mokslas 294, 980 – 982. doi: 10.1126 / science.294.5544.980
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Imperato, A., Di Chiara, G. (1988). Lokaliai taikomų D-1 ir D-2 receptorių agonistų ir antagonistų poveikis buvo tiriamas atliekant smegenų dializę. Euras. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Džeksonas, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Kapa opioidų receptorių antagonisto norbinaltorphimino poveikis stresui ir vaistų sukeltas nikotino sąlygotos vietos pirmenybėse pelėms. Psichofarmakolis. (Berlas) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., et al. (2014). Salvinorinas A reguliuoja dopamino pernešėjo funkciją per kappa opioidų receptorius ir nuo ERK1 / 2 priklausomą mechanizmą. Neurofarmakologija 86, 228 – 240. doi: 10.1016 / j.neuropharm.2014.07.016
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Elgesio jautrinimas chinpiroliui nėra susijęs su padidėjusiu branduolio akumuliatorių dopamino perpildymu. Neurofarmakologija 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Koob“, „GF“, „Volkow“, ND (2016). Priklausomybės neurobiologija: neurocirkuliacijos analizė. Lancet Psichiatrija 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Koobas, GF (2013). Priklausomybė yra atlygio deficitas ir streso perteklius. Priekyje. Psichiatrija 4, 72. doi: 10.3389 / fpsyt.2013.00072
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Labad, J. 1., Menchon, J. M., Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., et al. (2008). Obsesinių-kompulsinių simptomų lyčių skirtumai. Depresuoti nerimą. 25 (10), 832–838. doi: 10.1002 / da.20332
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, kappa opioidų receptorių agonistas, susilpnina kokaino sukeltą tarpląstelinio dopamino padidėjimą žiurkių branduolio akumuliatoriuose. Neurosci. Lett. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Opioidų receptorių mRNR raiška žiurkių CNS: anatominiai ir funkciniai padariniai. Tendencijos neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Mansour, A., Burke, S., Pavlic, R. J., Akil, H., Watson, SJ (1996). Imunohistocheminė klonuotų kappa 1 receptorių lokalizacija žiurkių CNS ir hipofizėje. Neurologijos 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Margolis, EB, Karkhanis, AN (2019). Dopaminerginis ląstelių ir grandinės indėlis į kappa opioidų receptorius sąlygoja baimę. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Kappa-opioidų agonistai tiesiogiai slopina vidurinių smegenų dopaminerginius neuronus. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Tiek Kappa, tiek Mu opioidiniai agonistai slopina glutamaterginį patekimą į veninius segmentinius neuronus. J. Neurophysiol. 93, 3086 – 3093. doi: 10.1152 / jn.00855.2004
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Kappos opioidai selektyviai kontroliuoja dopaminerginius neuronus, išsikišančius į priekinę priekinę žievę. Proc. Natl. Acad. Sci. JAV 103, 2938 – 2942. doi: 10.1073 / pnas.0511159103
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Vidurinės smegenų dopamino neuronai: projekcijos taikinys nustato veikimo potencialo trukmę ir dopamino D (2) receptorių slopinimą. J. Neurosci. 28, 8908 – 8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Mathews, IZ, McCormick, CM (2007). Žiurkių patelės ir patinai vėlyvoje paauglystėje nuo amfetamino sukeltos lokomotorinės veiklos skiriasi nuo suaugusiųjų, tačiau ne amfetamino, o sąlygotosios vietos atžvilgiu. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T. ir kt. (2011). Obsesinio-kompulsinio sutrikimo lyčių skirtumai: literatūros apžvalga. Brazas. J. Psichiatrija 33 (4), 390–399. doi: 10.1590 / S1516-44462011000400014
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Kappos opioidinių receptorių antagonizmas ir prodynorphin geno sutrikimas blokuoja streso sukeltą elgesio reakciją. J. Neurosci. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Ankstesnis kappa opioidinių receptorių aktyvinimas U50,488 imitavo pakartotinį priverstinį maudymo stresą, kad sustiprintų kokaino pasirinkimą. Neuropsychopharmacology 31, 787 – 794. doi: 10.1038 / sj.npp.1300860
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). Socialinio pralaimėjimo streso sukeltas elgesio reakcijas tarpininkauja endogeninė kappa opioidų sistema. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Lyties skirtumai tarp (+) - amfetamino ir (+) - metamfetamino sukeltų elgesio reakcijų žiurkių patinų ir patelių Sprague-Dawley. Pharmacol. Biochem. Behav. 86, 140 – 149. doi: 10.1016 / j.pbb.2006.12.018
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Kappa opioidų poveikis kokaino savarankiškam vartojimui. J. Pharmacol. Exp. Ther. 282, 44-55.
„Nestler“, EJ (2001). Ilgalaikio priklausomybės plastikos molekuliniai pagrindai. Nat. Rev. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Perreault, L. L., Grahamas, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Kappa-opioidinis agonistas U69593 stiprina lokomotorinę sensibilizaciją D2 / D3 agonisto chinpiroliui: pre- ir postsinapsiniai mechanizmai. Neuropsychopharmacology 31, 1967 – 1981. doi: 10.1038 / sj.npp.1300938
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Perreault“, L. L., Seemanas, P., Szechtmanas, H. (2007). Kappa-opioidinių receptorių stimuliacija paspartina kompulsinio tikrinimo patogenezę pagal obsesinio-kompulsinio sutrikimo (OKS) chinpirolio sensibilizacijos modelį. Behav. Neurosci. 121, 976 – 991. doi: 10.1037 / 0735-7044.121.5.976
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Pierce, RC, Kalivas, PW (1997). Elgesio jautrumo amfetaminą primenantiems psichostimuliatoriams schema. Brain Res. Brain Res. Rev. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Polteris, AM, vyskupas, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). „Kappa“ opioidinių receptorių blokas „Post Stress“ išgelbėjo ilgalaikę slopinamųjų sinapsių potenciją ir neleidžia atkurti kokaino. Biol. Psichiatrija 76, 785 – 793. doi: 10.1016 / j.biopsych.2014.04.019
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Puig-Ramos, A., Santjagas, GS, Segarra, AC (2008). U-69593, kappa opioidų receptorių agonistas, sumažina kokaino sukeliamą sensibilizaciją žiurkių patelėms. Behav. Neurosci. 122, 151 – 160. doi: 10.1037 / 0735-7044.122.1.151
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Neurokognityvūs impulsyvumo ir kompulsyvumo endofenotipai: dimensinės psichiatrijos link. Tendencijų pažinimas. Mokslas. 16, 81 – 91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Estrouzinis ciklas turi įtakos kokaino savarankiškam vartojimui pagal laipsnišką santykį su žiurkėmis. Psichofarmakolis. (Berlas) 98, 408 – 411. doi: 10.1007 / BF00451696
Robinsonas, TE, Berridge, KC (1993). Neuroninis narkotikų potraukio pagrindas: priklausomybės stimuliavimo-jautrinimo teorija. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Robinsonas, TE, Berridge, KC (2001). Skatinamasis jautrinimas ir priklausomybė. Polinkis 96, 103 – 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Skirtingas nalmefeno poveikis dopamino įsisavinimo greičiui ir kappa opioidinių receptorių aktyvumui branduolių akumuliatoriuose po lėtinio protarpinio etanolio poveikio. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, et al. (2013). Ilgalaikis κ opioidų receptorių antagonizmas neleidžia didėti heroino vartojimui ir didėja jo motyvacija. J. Neurosci. 33 (49), 19384–19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Sesack, SR, Pickel, V. M. (1992). Dviguba ultrainfrastruktūrinė enkefalino ir tirozino hidroksilazės imunoreaktyvumo lokalizacija žiurkės ventraliniame pagrindiniame rajone: daugybė substratų opiatų ir dopamino sąveikai. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Sesack, SR, Aoki, C., Pickel, V. M. (1994). D2 receptorių imunoreaktyvumo ultrainfrastruktūrinis lokalizavimas vidurinės smegenų dopamino neuronuose ir jų striatiniuose taikiniuose. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Sesia, T., Bizup, B., Grace, AA (2013). Obsesinio-kompulsinio sutrikimo gyvūnų modelių įvertinimas: koreliacija su fazinio dopamino neuronų aktyvumu. Vid. J. Neuropsychopharmacol. 16, 1295–1307. doi: 10.1017 / S146114571200154X
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Shippenberg“, TS, „Zapata“, A., „Chefer“, VI (2007). Dinorfinas ir narkomanijos patofiziologija. „Pharmacol“. Tem. 116, 306–321. doi: 10.1016 / j.pharmthera.2007.06.011
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Smithas, JA, Loughlin, SE, Leslie, FM (1992). kappa-opioidas slopina [3H] dopamino išsiskyrimą iš žiurkės ventralinių mezencefalinių disocijuotų ląstelių kultūrų. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Dopamino D2L receptorių išmušimo pelėms trūksta teigiamų ir neigiamų morfino stiprinamųjų savybių ir vengiama mokytis. Neurologijos 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Streso sukelta dinorfino / κ-opioidų receptorių sistemos aktyvacija amygdaloje sustiprina nikotino sąlygotos vietos pasirinkimą. J. Neurosci. 32, 1488 – 1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Spanagel, R., Herz, A., Shippenberg, TS (1992). Priešingos toniškai aktyvios endogeninės opioidų sistemos moduliuoja mezolimbinį dopaminerginį kelią. Proc. Natl. Acad. Sci. 89, 2046 – 2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., German, DC (1993). Opioidiniai receptoriai žiurkės vidurinės smegenų dopaminerginiuose regionuose. II. Kappos ir delta receptorių autoradiografija. J. Neural Transm. Gen. Sekta. 91, 53 – 66. doi: 10.1007 / BF01244918
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Endogeninis kappa-opioidinis tarpinis streso sukeltas potencialo stiprinimas etanolio sąlygojamoje vietoje ir savarankiškas vartojimas. Psichofarmakolis. (Berlas) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Narkotikų poreikis: sensibilizacija ir narkotikų ieškančio elgesio atkrytis, sunbley DR, red. Pharmacol. Rev. 63, 348–365. doi: 10.1124 / pr.109.001933
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., et al. (2016). Chinpirolio sensibilizacijos žiurkių OKS modelio galiojimas: susieti su gyvūnais atliktų ir klinikinių tyrimų duomenys. Priekyje. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Sun, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G. ir kt. (2019 m.). Bazolaterinis amigdalos įvedimas į medialinę prefrontalinę žievę kontroliuoja į obsesinius-kompulsinius sutrikimus panašų elgesį. Proc. Natl. Acad. Sci. JAV 116, 3799 – 3804. doi: 10.1073 / pnas.1814292116
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Didelis β-opioidinių receptorių ir dopamino pernešėjo ekspresija branduolio akumuliatorių aksoniniuose profiliuose. Synapse 42, 185 – 192. doi: 10.1002 / syn.10005
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Dopamino agonisto chinpirolio sukeltos sensibilizacijos aplinkos ir elgesio komponentai. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dopamino agonisto chinpirolio sukeltas elgesio jautrumo dinamika ir siūlomas centrinis energijos kontrolės mechanizmas. Psichofarmakolis. (Berlas) 115, 95 – 104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Chinpirolis sukelia kompulsinį tikrinimo elgesį su žiurkėmis: tai yra galimas obsesinio-kompulsinio sutrikimo (OKS) modelis gyvūnams. Behav. Neurosci. 112, 1475 – 1485. doi: 10.1037 / 0735-7044.112.6.1475
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Szechtman, H., Culver, K., Eilam, D. (1999). Dopamino sistemų vaidmuo obsesiniame-kompulsiniame sutrikime (OKS): naujojo psichostimuliatorių sukeltų gyvūnų modelio padariniai. Pol. J. Pharmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, Ca, JZ, M., et al. (2001). Kompulsinis žiurkėms, jautrioms chinpirolio, elgesio su obsesiniu-kompulsiniu sutrikimu (OKS) modelis: forma ir kontrolė. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Szechtman, H., Ahmari, SE, Beninger, R. J., Eilam, D., Harvey, BH, Edemann-Callesen, H., et al. (2017 m.). Obsesinis-kompulsinis sutrikimas: įžvalgos iš gyvūnų modelių. Neurosci. Biobehav. Rev. 76, 254 – 279. doi: 10.1016 / j.neubiorev.2016.04.019
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Tejeda, HA, Bonci, A. (2019). Dinorfino / kappa-opioidinių receptorių dopamino dinamikos kontrolė: neigiamos afektinės būsenos ir psichiniai sutrikimai. Brain Res. 1713, 91 – 101. doi: 10.1016 / j.brainres.2018.09.023
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J. ir kt. (2017). Sužadinimo ir slopinimo balanso kelio ir ląstelių specifinis kappa-opioidinių receptorių moduliavimas skirtingai vartoja d1 ir d2 akumuliuoja neuronų aktyvumą. Neuronas 93 (1), 147–163. doi: 10.1016 / j.neuron.2016.12.005
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Thompson, AC, Zapata, A., Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Kappa-opioidinių receptorių aktyvinimas keičia dopamino įsisavinimą akumuliatorių branduolyje ir prieštarauja kokaino poveikiui. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Thompson, D., Martini, L., Whistler, JL (2010). Pasikeitęs D1 ir D2 dopamino receptorių santykis pelių striatoje yra susijęs su elgesio jautrinimu kokainui. PloS One 5, e11038. doi: 10.1371 / journal.pone.0011038
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., et al. (2000). Skirtingos dviejų dopamino D2 receptorių izoformų funkcijos. Gamta 408, 199 – 203. doi: 10.1038 / 35041572
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Van Bockstaele, EJ, Pickel, V. M. (1995). GABA turintys neuronai ventralinėje pagrindinėje srityje išsikiša į žiurkės smegenyse esančius branduolius. Brain Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY ir kt. (2013). Kapa-opioidų receptorių pašalinimas iš smegenų dopamino neuronų turi panašų į anksiolitinį poveikį ir padidina kokaino sukeltą plastiškumą. Neuropsychopharmacology 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Vanderschuren, LJ, Kalivas, PW (2000). Dopaminerginio ir glutamaterginio perdavimo pokyčiai sukeliant ir išreiškiant elgesio sensibilizaciją: kritinė ikiklinikinių tyrimų apžvalga. Psichofarmakolis. (Berlas) 151 (2–3), 99–120. doi: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017). Dopamino motyvų sistema: priklausomybė nuo narkotikų ir maisto. Nat. Rev. Neurosci. 18, 741–752. doi: 10.1038 / nrn.2017.130
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Walkeris, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Dopamino išsiskyrimas ir įsisavinimas yra didesnis nei žiurkių patinų patinų striatumoje, matuojant greita cikline voltamperometrija. Neurologijos 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Wee, S., Koob, GF (2010). Dinorfino – κ opioidų sistemos vaidmuo stiprinant piktnaudžiavimo narkotikais poveikį. Psichofarmakolis. (Berlas) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Kapa opioidinių receptorių slopinimas sumažino padidėjusį kokaino vartojimą žiurkėms ir padidino kokaino prieinamumą. Psichofarmakolis. (Berlas) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr, Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., et al. (2015). κ Opioidų receptoriai branduolio akumuliatoriaus apvalkale padeda didinti metamfetamino suvartojimą. J. Neurosci. 35 (10), 4296–4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Williamsas, MT, Mugno, B., Franklinas, M., Faberis, S. (2013). Obsesinio-kompulsinio sutrikimo simptomų dimensijos: fenomenologija ir gydymo rezultatai naudojant ekspoziciją ir ritualinę prevenciją. Psichopatologija 46, 365 – 376. doi: 10.1159 / 000348582
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Wilsonas, CJ, Young, SJ, Groves, PM (1977). Statistinės neuronų smaigalių traukinių savybės pagrindinėje nigroje: ląstelių tipai ir jų sąveika. Brain Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Išminčius, RA (2009). Nigrostriatalio, o ne tik mezokortikolimbinio, dopamino vaidmuo atlygio ir priklausomybės srityse. Tendencijos neurosci. 32, 517 – 524. doi: 10.1016 / j.tins.2009.06.004
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Xiao, L., Becker, JB (1994). Tarpląstelinės striatos dopamino koncentracijos kiekybinis mikrodializės nustatymas žiurkių patinams ir patelėms: estrozinio ciklo ir lytinių liaukų pokyčiai. Neurosci. Lett. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
„Yager“, LM, „Garcia“, AF, Wunsch, AM, Ferguson, SM (2015). Stuburio pjūviai ir trūkumai: vaidmuo priklausomybėje nuo narkotikų. Neurologijos 301, 529 – 541. doi: 10.1016 / j.neuroscience.2015.06.033
PubMed Santrauka | „CrossRef“ visas tekstas | "Google Scholar"
Raktažodžiai: kappa opioidų receptoriai, dopaminas, kompulsyvumas, amfetaminas, chinpirolis, lokomotorinė sensibilizacija
Citacija: „Escobar AdP“, „Casanova JP“, „Andrés ME“ ir „Fuentealba JA“ (2020) „Kappa“ opioidų ir dopamino sistemų skerspjūvis kompulsyvaus elgesio metu. Priekis Pharmacol. 11: 57. doi: 10.3389 / fphar.2020.00057
Gauta: 16 m. Spalio 2019 d .; Priimta: 22 m. Sausio 2020 d .;
Paskelbta: 18 Vasaris 2020.
Redagavo:
Gonzalo E. Jevenesas, Koncepciono universitetas, Čilė
Peržiūrėjo:
Luisas Gerardo Aguayo, Koncepciono universitetas, Čilė
Hugo Tejeda, Nacionalinis piktnaudžiavimo narkotikais institutas (NIDA), JAV
Cecilija Skorza, Instituto de Investigaciones Biológicas Clemente Estable (IIBCE), Urugvajus
Autorinės teisės © 2020 „Escobar“, „Casanova“, „Andrés“ ir „Fuentealba“. Tai yra atviros prieigos straipsnis, platinamas pagal „Creative Commons“ priskyrimo licencija (CC BY). Leidžiama naudoti, platinti ar atgaminti kituose forumuose, jei įskaitomas originalus autorius (-iai) ir autorių teisių (-ių) savininkas (-iai) ir kad minėtas leidinys yra paskelbtas šiame žurnale, laikantis pripažintos akademinės praktikos. Negalima naudoti, platinti ar atgaminti, kuri neatitinka šių sąlygų.
* Korespondencija: José Antonio Fuentealba, [apsaugotas el. paštu]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675
