PASTABOS: Vienas iš nedaugelio tyrimų, apimančių abstinencijos poveikį dopamino receptorių lygiui primatuose.
- D2 receptoriai gana greitai atšoka - mažiau nei mėnesį
- D1 receptoriai per mėnesį yra per dideli, tačiau per 90 dienas sugrįžta.
- Dideli arba maži D1 receptoriai gali būti ūminio pasitraukimo ir potraukio raktai
Thomas JR Beveridge'as1, Hilary R Smith1, Michaelas A Naderis1 ir Linda J Porrino1
1Fiziologijos ir farmakologijos katedra, Narkotikų piktnaudžiavimo neurobiologinio tyrimo centras, „Wake Forest“ universiteto medicinos mokykla, Winston-Salem, NC, JAV
Korespondencija: dr. LJ Porrino, Fiziologijos ir farmakologijos katedra, Narkotikų piktnaudžiavimo neurobiologinio tyrimo centras, Wake Forest universiteto Medicinos mokykla, Medicinos centras bulvaras, Winston-Salem, NC 27157-1083, JAV. Tel: + 1 336 716 8575; Faksas: + 1 336 716 8501; El. Paštas: [apsaugotas el. paštu]
Gauta 29 m. Balandžio 2008 d .; Patikslinta 25 m. Liepos 2008 d .; Priimta 30 m. Liepos 2008 d .; Paskelbta internete 3 m. Rugsėjo 2008 d.
Abstraktus
Nors dopamino (DA) sistemos reguliavimas yra būdingas lėtinio kokaino poveikio bruožas, klausimas, ar šie pokyčiai išlieka abstinencijoje, iš esmės neatsakytas. Nežmoginiai primatai yra idealus modelis vertinant abstinencijos poveikį DA sistemai po lėtinio kokaino poveikio. Šiame tyrime vyrų reeso beždžionės patys vartojo kokainą (0.3 mg/kg injekcijos, 30 stiprintuvai per sesiją) pagal fiksuotą intervalą 3 min100 dienų po 30 arba 90 dienų abstinencijos. Tai parodė, kad ši kokaino savęs vartojimo trukmė mažina DA D2 tipo receptorių tankį ir padidina D1 tipo receptorių ir DA transporterių (DAT) kiekį.. Kontrolinių beždžionių atsakas buvo palaikomas pateikiant maistą pagal identišką protokolą ir tuos pačius abstinencijos laikotarpius. [3H]SCH 23390 privalomas DA D1 receptoriai po 30 dienų po abstinencijos buvo gerokai didesni visose striatumo dalyse, palyginti su kontroliniais gyvūnais, tuo tarpu \ t [3H]raclopido prisijungimas prie DA D2 receptorių grupėse nesiskyrė. [3H]WIN 35 428 prijungimas prie DAT taip pat buvo žymiai didesnis visoje beveik visose nugaros ir ventralinės stiatos dalyse po 30 abstinencijos dienų. Tačiau po 90 dienų abstinencijos metu DA D1 receptorių ir DAT koncentracija nesiskyrė nuo kontrolinių verčių. Nors šie rezultatai rodo, kad galimas atskirų DA sistemos elementų atsigavimas, jie taip pat išryškina šių komponentų dinaminį pobūdį pradiniuose abstinencijos etapuose nuo lėtinio kokaino savarankiško vartojimo.
Raktiniai žodžiai:
kokainas, dopaminas, autoradiografija, abstinencija, striatumas
ĮVADAS
Lėtinis kokaino vartojimas tarp žmonių, priklausančių nuo narkomanų, buvo susijęs su neuroadaptacija dopamino (DA) sistemoje (Malisonas et al, 1998; Volkow et al, 1993, 1997). Tai apima DA transporterių tankio padidėjimą (DAT) ir DA D2 tipo receptorių koncentracijų sumažėjimą (Mažai et al, 1999; Misa et al, 2002; Volkow et al, 1993). Be to, pastebėta ir DA išskyrimo pokyčių. Pavyzdžiui, tyrėjai, naudodami pozronų emisijos tomografijos (PET) tyrimus [11C]raclopidas ir metilfenidatas parodė mažėjančią DA lėtinio lėtinio kokaino vartotojų stresą (Volkow et al, 1997; Wong et al, 2006). Tačiau viena problema yra ta, kad dažnai sunku išskirti kitų veiksnių, pvz., Kitų nelegalių ir teisėtų narkotikų, ankstesnių narkotikų vartojimo ir vartojimo modelių skirtumus bei gyvenimo būdo skirtumus. Šie skirtumai, taip pat ir sąlygos, galinčios prieš narkotikų vartojimą, gali apriboti tyrimus žmogaus pacientams.
Nežmoginiai primatai, kuriuose kintamieji gali būti sistemingai manipuliuojami, yra alternatyvus metodas, kaip tirti lėtinio kokaino savęs administravimo ir vėlesnio abstinencijos pasekmes. Ankstesni tyrimai parodė, kad lėtinis kokaino poveikis yra susijęs su reikšmingu DA D2 receptorių koncentracijos sumažėjimu, taip pat D1 receptorių koncentracijos padidėjimu ir DAT tankumu.Letchworthas et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Šie poveikiai atspindi tuos, kurie pasireiškia žmonėms, ir tokiu būdu patvirtina šių vaistų poveikio modelių naudingumą.
Nors yra nemažai įrodymų apie DA sistemų reguliavimą, pasirodė esąs sunkiau įvertinti, ar yra požymių, kad po naudojimo nutraukus narkotikų vartojimą atsirastų įrodymų (Malisonas et al, 1998; Jacobsenas et al, 2000; Volkow et al, 1993) arba ar šie pokyčiai išlieka ilgiau nei tęsiamas kokaino poveikis. Vėlgi, nežmoniški primatai gali pateikti įžvalgų apie šį priklausomybės etapą. Farfel et al (1992) pranešta apie sumažėjusias DAT ir D1 tipo receptorių koncentracijas beždžionių striatume po susilaikymo nuo lėtinio nekontroliuojamo kokaino poveikio. Tačiau specifinį abstinencijos vaidmenį buvo sunku nustatyti dėl to, kad grupėje, kurioje nėra abstinencijos periodo, nebuvo matavimų. Panašiai, Melega et al (2008) pranešta, kad po 3 savaitės susilaikymo nuo didėjančio metamfetamino režimo pastebimai sumažėjo DAT koncentracija vervetinių beždžionių striatume. Tačiau stimuliantų vartojimas abiejuose tyrimuose buvo nesusijęs. Vartojimo būdas (kontingentas) vs nesusijęs), smegenų diferencialiai paveikė abiejų DA išskyrų atžvilgiu (Hemby et al, 1997) ir gliukozės metabolizmą (\ tGraham ir Porrino, 1995; Porrino et al, 2002). Taigi, savęs administravimo naudojimas šiame tyrime apeina šią problemą. Be to, ilgalaikio kokaino savarankiško vartojimo poveikis smegenų DA sistemoms buvo plačiai ištirtas naudojant šį savireguliacijos modelį reeso beždžionėse, tokiu būdu suteikiant pagrindą, kuriuo remiantis būtų galima įvertinti neuroadaptacijas, kurios atsiranda abstinencijos metu.
Todėl šių tyrimų tikslas buvo nustatyti, ar DAT ir DA D1 ir D2 receptorių koncentracijos pokyčiai, kurie anksčiau buvo įrodyti gyvūnams, patyrusiems kokainą savarankiškai.Letchworthas et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002) būtų pakeista po ilgų abstinencijos laikotarpių. Remiantis atliktais tyrimais su žmonių narkotikų vartotojais (plg Volkow et al, 1993), mes manėme, kad šie DA sistemos pokyčiai išliks net ir po 3 mėnesio abstinencijos. Tuo tikslu beždžionės savarankiškai vartojo kokainą 100 sesijoms, o bendras 900 kiekis mg/kg, po to po 30 arba 90 dienų abstinencijos nuo narkotikų. DA D1 ir D2 receptoriai, taip pat DAT buvo matuojami kiekybiškai in vitro receptorių autoradiografija.
METODAI
Tematika
Iš viso 17 eksperimentiškai neveikusių vyrų reeso beždžionių (Macaca mulatta) svėrimas tarp 7.7 ir 13 kg (vidurkis ± SD, 10.2 ± 1.32) tyrimo pradžioje buvo tiriamieji. Visos procedūros buvo atliktos pagal nusistovėjusią praktiką, kaip aprašyta Nacionaliniai sveikatos priežiūros institutai laboratorinių gyvūnų priežiūrai ir naudojimui. Be to, visas procedūras peržiūrėjo ir patvirtino Wake Forest universiteto Gyvūnų priežiūros ir naudojimo komitetas. Beždžionės buvo atskirai laikomos nerūdijančio plieno narveliuose su vandeniu ad libitum; gyvūnai buvo vienas su kitu fiziškai ir vizualiai. Jų kūno svoris buvo palaikomas maždaug 90 – 95% bandomųjų sesijų metu uždirbtų bananų skonio granulių ir papildomo „Lab Diet Monkey Chow“ maitinimo svorio, pateiktų ne anksčiau kaip „30“ min. po sesijos. Be to, ne mažiau kaip tris kartus per savaitę jie buvo švieži vaisiai ar žemės riešutai. Kiekvienas beždžionė buvo sveriama kartą per savaitę, o prireikus jų mityba buvo pakoreguota, kad būtų išlaikyti stabilūs svoriai.
Elgesio aparatai
Eksperimentiniai užsiėmimai buvo atliekami ventiliuojamose ir garso slopinimo operacinėse kamerose (1.5 × 0.74 × 0.76 m; „Med Associates Inc.“, „East Fairfield“, VT), skirta primatinės kėdės (R001; Primate Products, Redwood City, CA). Kameroje buvo žvalgybos skydas (48 × 69 cm), kurį sudarė dvi įtraukiamos svirties (5) cm pločio) ir trys stimuliatoriai. Svirties padėtis buvo lengvai pasiekiama iš beždžionės, sėdinčios primatų kėdėje. Bananų skonio maisto granulės (1 g; „Bio-Serv“, „Frenchtown“, NJ) buvo pristatytos iš padavimo kameros, esančios kameros viršuje. Vaistinių preparatų injekcijoms vartoti maždaug 7531 buvo naudojamas peristaltinis infuzinis siurblys (10-1; Cole-Parmer Co., Čikaga, IL) ml / 10 s tiems gyvūnams, kurie savarankiškai vartoja kokainą. Kamerų eksploatavimas ir duomenų kaupimas buvo atliktas naudojant „Power Macintosh“ kompiuterinę sistemą su sąsaja („Med Associates Inc.“).
Chirurginės procedūros
Visi beždžionės, įskaitant kontrolinius, buvo chirurgiškai paruošti steriliomis sąlygomis su intraveniniais kateteriais ir kraujagyslių patekimo prievadais (GPV modelis; Access Technologies, Skokie, IL). Beždžionės anestezuotos ketamino deriniu (15 mg/kg, im) ir butorfanolio (0.03 mg/kg, im) ir pjūvis buvo padarytas netoli šlaunikaulio. Po nelygios skilimo ir venų izoliacijos į veną buvo įterptas artimiausias kateterio galas, kad būtų apskaičiuotas atstumas, kuris baigsis žemesnėje vena cava. Diskalinis kateterio galas buvo supjaustytas poodiniu būdu į pjūvį, šiek tiek nutolusį nuo nugaros vidurio linijos. Kraujagyslių patekimo prievadas buvo patalpintas į kišenę, susidariusią nelyginiu skaidymu, šalia šio pjūvio. Beždžionėms buvo suteiktas 24 – 48 h atkūrimo laikas prieš grįžtant prie sustiprinto maisto. Maždaug prieš 5 dienas prieš terminalią procedūrą kiekvienas beždžionė buvo implantuota lėtiniu patekusiu kateteriu į gretimą šlaunies arteriją, kad būtų galima surinkti laikinus arterinius kraujo mėginius. Chirurginės procedūros buvo identiškos aprašytoms venų kateteriams. Paskutinės sesijos dieną buvo atliktas galutinis smegenų gliukozės metabolizmo tyrimas, kuriame beždžionės buvo švirkščiamos 2-[14C](2-DG) apytiksliai 2 min. po sesijos pabaigos ir kraujo mėginiai buvo gauti per arterinį kateterį per 45 min Beveridge et al, 2006 išsamiau). Šių tyrimų metabolizmo duomenys čia nepateikiami.
Savivaldos procedūros
Iš pradžių beždžionės buvo apmokytos reaguoti į vieną iš dviejų svertų, sustiprinant kiekvieną atsaką į tinkamą svirtį su maisto granulėmis. Per maždaug 3 savaitės laikotarpį tarp maisto granulių pasiekimo intervalas palaipsniui didėjo, kol pasiekiamas 3 min. Intervalas (ty fiksuoto intervalo 3 min. Stiprinimo tvarkaraštis; FI 3-min). Galutinėmis grafiko sąlygomis pirmas atsakymas į svirtį po 3 min. buvo gauta maisto granulių; sesijos baigėsi po 30 maisto pristatymo. Kiekvienos sesijos pabaigoje buvo ištrauktos atsakymo svirtys, užgęsta namų apšvietimas ir stimuliuojančios šviesos, o gyvūnai liko tamsioje kameroje maždaug 30 min. prieš juos grįžus į savo narvų narvus. Visi beždžionės atsakė pagal FI 3-min maisto pristatymo tvarkaraštį mažiausiai 20 sesijoms ir tol, kol buvo gautas stabilus veikimas (± 20% trijų iš eilės sesijų vidurkio, be atsakymų rodiklių tendencijų). Kai atsakas į maistą buvo stabilus, maitinimo šaltinis buvo atjungtas, o išnykimo poveikis atsakymui buvo ištirtas penkias iš eilės einančias sesijas, po kurių atsakas buvo atkurtas ir palaikomas pateikiant maistą.
Nustačius pradinę būklę, visi beždžionės buvo chirurgiškai paruoštos venų kateteriais, kaip aprašyta aukščiau, ir atsitiktinai priskiriamos vienai iš trijų grupių. Viena beždžionių grupė tarnavo kaip kontrolė ir toliau atsakė pagal FI 3-min maisto pristatymo tvarkaraštį iš viso 100 sesijų metu.N=6). Likusieji 11 beždžionės buvo priskirtos prie kokaino savarankiško vartojimo grupių (0.3 mg/kg). Kadangi 0.3 mg/1 kg kokaino vienam injekcijai buvo laikoma didele doze anksčiau kokaino nevartojusiems beždžionėms, daugumai gyvūnų ši dozė buvo pasiekta per dvi sesijas, iš pradžių leisdama beždžionėms patiems vartoti 0.1 mg/kg kokaino. Po operacijos (maždaug 4 – 6 dienos), kol prasidėjo kokaino savarankiško vartojimo sesijos, buvo užtikrinta, kad maistas palaikytų stabilumą. Prieš kiekvieną eksperimentinę sesiją gyvūno nugarėlė buvo išvalyta 95% etanolio ir betadino šveitimas ir 22 gabaritas Huber Point adata (modelis PG20-125) buvo įterpti į veną kateterį, jungiantį infuzinį siurblį su kokaino tirpalu į kateterį. Prieš pradedant sesiją, siurblys buvo naudojamas maždaug 3 s, užpildant uostą su kokaino doze, kuri buvo prieinama eksperimentinės sesijos metu. Sesijos baigėsi po 30 injekcijų; kaip kontrolinėmis sąlygomis beždžionės liko tamsioje kameroje maždaug 30 min. Kiekvienos sesijos pabaigoje uostas buvo užpildytas heparinizuotu fiziologiniu tirpalu (100 U/ml), kad būtų išvengta krešėjimo.
Eksperimentiniai užsiėmimai buvo atliekami maždaug tuo pačiu metu kiekvieną dieną ir tęsiami visoms 100 sesijoms. Baigus 100 sesijas, buvo įvestas 30 arba 90 dienų abstinencijos laikotarpis, kurio metu kateteriai kasdien buvo praplaunami su heparinizuotu fiziologiniu tirpalu, bet kokaino ar maisto savarankiško seanso nebuvo. Kontrolinės grupės atveju 30 dienų abstinencijos periodai buvo nustatyti keturiems gyvūnams ir 90 dienoms dviem kitiems. Kokaino grupei 30 dienų abstinencijos laikotarpiai buvo nustatyti aštuoniems gyvūnams ir 90 dienoms trims gyvūnams. Pasibaigus abstinencijos laikotarpiui, buvo atlikta viena galutinė savęs administravimo sesija (maisto kontrolė arba kokainas), o po sesijos pradėta procedūra „2-DG“. Dviejuose kontroliniuose tyrimuose ir keturiose kokaino savarankiškai vartojantys gyvūnai 30 dienos abstinencijos grupėje paskutiniame posėdyje kokaino nebuvo. Gyvūnai buvo humaniškai nužudyti pentobarbitalio perdozavimu (100 mg/kg, iv) 45 pabaigoje min žymeklio įsisavinimo laikas.
Audinių apdorojimas
Po nužudymo smegenys buvo nedelsiant pašalintos, užblokuotos ir užšaldytos izopentane –35 į –55 ° C ir po to laikomos –80 ° C temperatūroje. Tada audinių blokai, kuriuose yra striatumo, buvo nukirpti kriostatu -20 ° C temperatūroje koronalioje plokštumoje į 20 μm sekcijos, surenkamos į elektrostatiniu būdu įkrautus stiklelius, džiovinamos vakuume per naktį, esant 4 ° C temperatūrai, po to laikomos -80 ° C temperatūroje, kol apdorojamos autoradiografijai. Smegenų sekcijos buvo surenkamos iš caudato branduolio, putameno ir branduolių accumbens dalių, esančių rostrale iki priekinės komisijos. Šis regionas vadinamas išankstiniu tarpsniu. Be to, rostraliniai ir caudaliniai pirmtakinio striatumo lygiai buvo nurodyti pagal branduolį accumbens. Rostralinis išankstinis spiatumas yra regionas, kuriame branduolys accumbens nėra diferencijuojamas į atskiras apvalkalo ir pagrindinio subkomponento dalis. Caudalinis išankstinis spiatumas yra regionas, sutampantis su branduolio accumbens lukšto ir šerdies išvaizda, kuri yra užkandžių tuberkulio atsiradimui. Kiekviename iš jungiamųjų tyrimų buvo paimtos dvi gretimos sekcijos kiekviename iš penkių lygių (du rostriniai ir trys caudaliniai) per išankstinį spermą, skirtą visiems 10 skyriams vienam gyvūnui.
D1 Receptor Binding
Nustatyti DA D1 receptorių surišimo vietos tankiai [3H]SCH 23390 (specifinė veikla - 85 Ci/mmol; PerkinElmer, Boston, MA) kiekybiškai in vitro receptorių autoradiografija pagal procedūras, pritaikytas nuo Lidow et al (1991) ir Nader et al (2002). Sekcijos buvo inkubuotos 20 min. buferyje (50 mM Tris, 120 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, pH 7.4, 25 ° C), siekiant pašalinti endogeninę DA, kokainą ir [14C] iš 2-DG procedūros. Po to sekcijos buvo inkubuojamos 30 min. tame pačiame buferyje, pH 7.4, 25 ° C, kuriame yra 1 mM askorbo rūgšties, 40 nM ketanserino ir 1 nM [3H]SCH 23390. Po inkubacijos 20 du kartus buvo skalaujami s buferyje, kuriame yra 1 mM askorbo rūgšties, esant pH 7.4, 4 ° C, tada panardintas į distiliuotą vandenį 4 ° C temperatūroje ir džiovinamas vėsaus oro sraute. Nespecifinis surišimas buvo nustatytas inkubaciniame tirpale esančių gretimų sekcijų inkubacija, dalyvaujant 5 μM (+) -butaklamolis. Skyriai kartu su kalibravimu [3H] autoradiografiniai standartai (Amersham, Piscataway, NJ) buvo įtraukti į Kodak Biomax MR filmą (Fisher Scientific, Pittsburgh, PA) 6 savaitėms.
D2 Receptor Binding
Nustatyta DA D2 receptorių surišimo vietų tankis ir pasiskirstymas [3H]raclopidas (specifinis aktyvumas, 87 Ci/mmol; PerkinElmer) pagal procedūras, pritaikytas nuo Lidow et al (1991) ir Nader et al (2002). Sekcijos buvo inkubuotos 20 min. buferyje (50 mM Tris, 120 mM NaCl, 5 mM KCl, pH 7.4, 25 ° C), siekiant pašalinti endogeninę DA, kokainą ir [14C] iš 2-DG procedūros. Tada skaidrės buvo inkubuojamos 30 min. tame pačiame buferyje, kuriame yra 5 mM askorbo rūgšties ir 2 nM [3H]raclopride. Skyriai buvo nuplauti 3 × 2 min. buferyje, esant pH 7.4, 4 ° C, tada panardintas į distiliuotą vandenį 4 ° C temperatūroje ir džiovinamas vėsaus oro sraute. Nespecifinis surišimas buvo nustatytas inkubaciniame tirpale esančių gretimų sekcijų inkubacija, dalyvaujant 1 μM (+) –Butaklamolis. Skyriai kartu su kalibravimu [3H] autoradiografiniai standartai buvo įtraukti į „Kodak Biomax MR“ filmą 8 savaitėms.
Dopamino transporterio rišimas
DAT surišimo vietų tankis buvo nustatytas naudojant [3H]WIN 35,428 (specifinė veikla, 87 Ci/mmol; PerkinElmer) autoradiografija pagal procedūras, pritaikytas iš Canfield et al (1990) ir Letchworthas et al (2001). Audinių sekcijos buvo inkubuojamos buferyje (50 mM Tris, 100 mN NaCl, pH 7.4, 4 ° C) 20 min, kad pašalintumėte likusius DA, kokainus ir [14C] iš 2-DG procedūros. Po to sekcijos buvo inkubuojamos 2 h 4 ° C tame pačiame buferyje, kuriame yra 5 nM [3H]WIN 35 428. Skyriai buvo nuplauti iš viso 2 min. buferyje, esant 4 ° C temperatūrai, tada panardinamas distiliuotame vandenyje, esant 4 ° C temperatūrai, ir džiovinamas šalto oro sraute. Nespecifinis surišimas buvo nustatytas inkubaciniame tirpale esančių gretimų sekcijų inkubacija, dalyvaujant 30 μM kokainas. Skyriai kartu su kalibravimu [3H] autoradiografiniai standartai buvo įtraukti į „Kodak Biomax MR“ filmą 6 savaitėms.
Densitometrija ir duomenų analizė
Filmai buvo sukurti su „Kodak GBX“ kūrėju, „stopbath“ ir „Rapid Fixer“ (VWR, „West Chester“, PA), o paskui nuplauti. Autoradiogramų analizė atlikta kiekybiniu densitometrijos būdu su kompiuterizuota vaizdo apdorojimo sistema (MCID, Imaging Research; InterFocus Imaging Ltd, Kembridžas, Jungtinė Karalystė). Optinio tankio vertės buvo konvertuojamos į fmol/mg (drėgno audinio) pagal kalibruotą [3H] standartus. Specifinis surišimas buvo nustatytas skaitmeniniu būdu atimant nespecifinio rišimo vaizdus iš viršutinių gretimų vaizdų, esančių visiškame rišamajame. Konstrukcijos buvo identifikuojamos pagal Nissl dažymo sekcijas, esančias greta tų, kurie buvo analizuojami receptorių surišti. Kiekvieno tyrimo duomenys buvo analizuojami nepriklausomai, taikant vienpusį dispersijos analizę, po kurios sekė mažiausiai kvadratiniai skirtumai post-hoc daugkartinių palyginimų bandymai. Kiekvienas regionas sudarė atskirą analizę. Kadangi 30 ir 90 dienų kontrolinius gyvūnus sukaupę privalomi duomenys nesiskiria vienas nuo kito, panašūs į ankstesnius tyrimus (Nader et al, 2002), duomenys iš kontrolinių grupių buvo sujungti. Be to, duomenų apie tuos gyvūnus, kurie vartojo kokainą, ir tų, kurie nebuvo galutinėje sesijoje, duomenys reikšmingai nesiskyrė, todėl šių grupių duomenys taip pat buvo sujungti.
REZULTATAI
Lėtinio kokaino savęs administravimo abstinencijos poveikis 30 dienoms
Koncentracijos [3H]SCH 23390 surišimas į DA D1 receptorius priešakūniniame striatume parodytas Lentelė 1. Specifinis [3H]SCH 23390 sudarė daugiau nei 90% iš viso. Atitinka ankstesnes ataskaitas (Moore et al, 1998a; Nader et al, 2002), [3H]SCH 23390 prie D1 receptorių, vartojamų ne narkotikų veikiamuose kontroliniuose gyvūnuose, buvo nevienalytė, turinti pastebimus skirtumus tarp striatumo subregionų. Žymėjimas buvo tankesnis visose rostralinėse ir medialinėse porcijose per visą striatum.
![1 lentelė - 30 arba 90 abstinencijos dienų įtaka po lėtinio kokaino savarankiško vartojimo [Xsum] 3H [rsqb] SCH 23390 pririšimas prie D1 receptorių Rešus beždžionių priešgaisrinėje stadijoje - Deja, negalime pateikti prieinamo alternatyvaus teksto tai. Jei reikia pagalbos norėdami pasiekti šį vaizdą, prašome susisiekti su help@nature.com arba autoriu](https://yourbrainonporn.com/wp-content/uploads/2020/01/table_thumb.gif)
Po 30 dienų nuo susilaikymo nuo kokaino poveikio, prisijungimas prie D1 receptorių pasižymėjo plačiu mastu per visą rostrali-caudalinį ankstyvosios stiatos mastą, lyginant su nesusijusių kontrolinių gyvūnų (žr.Lentelė 1; 1 pav). Rostraliaus striatume koncentracija buvo žymiai didesnė caudatuose, įskaitant dorsolaterinę (+27%), centrinis (+27%), dorsomedial (+27%) ir ventromedial (+23%) porcijas, taip pat ir nugaros (+17%), centrinis (+22%) ir ventraliniai (+23%) putameno porcijos, lyginant su tankiu neužsikrėtusiuose kontroliniuose gyvūnuose. Reikšmingas padidėjimas taip pat buvo pastebėtas branduolių accumbens (+23%) šiuo lygiu. Stiatumo lygiu, kur branduolio accumbens šerdis ir lukštai yra labiausiai diferencijuoti, D1 tipo receptorių tankis taip pat buvo žymiai didesnis visoje dorsolateriniame (+31%), centrinis (+29%), dorsomedial (+30%) ir ventromedial (+18%) caudate branduolys, taip pat nugaros (+23%), centrinis (+29%) ir ventraliniai (+28%) putamen, lyginant su nekontroliuojamų kontrolinių medžiagų tankumu. Ventralinio striatumo viduje, šiuo lygiu, D koncentracija1 receptorių surišimo vietos buvo didesnės branduolio accumbens šerdyje (+45%) ir apvalkalas (+20%), taip pat uoslės tuberkle (+26%), lyginant su kontrolinių sistemų tankiais.
1 pav.
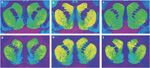 Reprezentatyvios autoradiogramos [3H] SCH 23390 jungiasi prie D1 receptorių (viršutinė plokštė) ir [3H]WIN 35428 prisijungimas prie dopamino transporterių (apatinis skydas) resuso beždžionių striatum koroninėse dalyse. (a, d) Kontroliuokite gyvūnų reagavimą į maisto stiprinimą. (b, e) Kokaino savarankiškai vartojamas gyvūnas su 30 dienų abstinencija. (c, f) Kokaino savarankiškai vartojamas gyvūnas, turintis 90 dienų abstinenciją.
Reprezentatyvios autoradiogramos [3H] SCH 23390 jungiasi prie D1 receptorių (viršutinė plokštė) ir [3H]WIN 35428 prisijungimas prie dopamino transporterių (apatinis skydas) resuso beždžionių striatum koroninėse dalyse. (a, d) Kontroliuokite gyvūnų reagavimą į maisto stiprinimą. (b, e) Kokaino savarankiškai vartojamas gyvūnas su 30 dienų abstinencija. (c, f) Kokaino savarankiškai vartojamas gyvūnas, turintis 90 dienų abstinenciją.
Koncentracijos [3H]Rasloprido surišimas į DA D2 receptorius prieškūniniame stiatume yra parodytas Lentelė 2. Konkretus susiejimas su [3H]raclopride buvo didesnis nei 90% iš viso. Pasiskirstymas [3H]Rasloprido prisijungimas prie D2 receptorių taip pat buvo nevienalytė visuose nugaros ir ventralinės striatos regionuose, kaip ir ankstesnėse ataskaitose (Moore et al, 1998b; Nader et al, 2002). Narkotikų ekspozicijos kontrolėje didesnė D2 surišimo vietų koncentracija buvo nugaros dalyje, palyginti su ventraliniu striatumu. Be to, buvo įrodyta, kad medialinis arba šoninis gradientas su didesne koncentracijų koncentracija yra daugiau šoninių striatumo dalių.
Po 30 dienų abstinencijos, D2 receptorių jungimosi su kokainu veikiančiais ir sustiprintais gyvūnais koncentracija daugelyje striatumo regionų nebuvo labai skirtinga. Didesnės jungimosi vietų koncentracijos buvo pastebėtos ventralinio putameno (+10%) ir priekiniai branduoliai accumbens (+12%) iš kokaino eksponuojamų beždžionių audiniuose, palyginti su kontroliniais preparatais. Kitų reikšmingų skirtumų nepastebėta.
Koncentracijos [3H]WIN 35428 surišimas į DAT išankstiniame spindulyje yra parodytas Lentelė 3. Atitinka ankstesnes ataskaitas (Letchworthas et al, 2001), nesusijusių su gyvūnais, kurie buvo veikiami ne DAT, Dors vietose buvo didesnė, palyginti su ventraliniu striatumu. Atliekant branduolį accumbens, šerdyje buvo pastebėtas didesnis tankis, palyginti su lukštų dalijimais. Galiausiai, nespecifinis įpareigojimas sudarė mažiau nei 10% iš viso.
Po 30 dienų po susilaikymo nuo kokaino poveikio, daugumai striatumo regionų, kurie buvo priskirti prie DAT, buvo žymiai didesnis jungimasis, palyginti su pririšimu prie nepageidaujamų vaistinių preparatų (žr.1 pav). Konkrečiai, DAT surišimo vietų koncentracija rostriniu lygiu buvo žymiai didesnė centrinėje (+22%), dorsomedial (+25%) ir ventromedial (+28%) caudatinis branduolys ir nugaros (+16%) ir centrinis (+23%) putamen, palyginti su sustiprintomis maisto kontrolėmis. Be to, taip pat buvo žymiai didesnė jungtis prie DAT priekiniame branduolyje (+37%) kokaino, palyginti su sustiprintais beždžionėmis. DAT surišimo vietų tankis buvo žymiai didesnis centrinėje caudate branduolyje (Caudal).+21%) ir putamen, centrinis (+20%) ir ventralinis (+19%; 1 pav). Ventralinio striatumo šitame lygyje prisijungimas prie DAT buvo žymiai didesnis branduolio accumbens šerdyje (+20%) ir uoslės tubercle (+24%) iš kokaino audinių \ t vs sustiprinti beždžionės.
Lėtinio kokaino savęs administravimo abstinencijos poveikis 90 dienoms
Priešingai nei plačiai paplitę D1 receptorių surišimo vietų tankio skirtumai, pastebėti vartojant su kokainu veikiančius gyvūnus po 30 abstinencijos dienų, po 90 dienų po abstinencijos nebuvo jokių reikšmingų skirtumų, lyginant su sustiprintu maistu kontroliuojamomis dalimis bet kokioje priešmokyklinio audinio dalyje. striatumas (Lentelė 1; 1 pav). Panašiai ir koncentracijos [3H]Rasloprido surišimas į DA D2 receptorius prieškomponentinėje striatume po 90 abstinencijos dienų taip pat reikšmingai nesiskyrė nuo tų, kurie buvo gydomi su vaistais nesusijusiems gyvūnams (Lentelė 2).
Koncentracijos [3H]WIN 35428 surišimas su DAT parodė panašų modelį, kaip ir vartojant D1 ir D2 receptorius. Po 90 abstinencijos dienų nebuvo pastebėta didelių DAT tankio skirtumų tarp kokaino eksponuojamų beždžionių, palyginti su nevartojamų vaistinių preparatų kiekiu (Lentelė 3; 1 pav), nors reikia pažymėti, kad buvo pastebėta tendencija, kad į priekinį branduolį susilieja aukštesnis jungimosi lygis.
DISKUSIJA
Ankstesni mūsų grupės tyrimai parodė, kad lėtinis kokaino savarankiško vartojimo poveikis kartu su reikšmingu nežmoginių primatų DA sistemos reguliavimu (Letchworthas et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Šio tyrimo rezultatai rodo, kad šis disreguliavimas lieka akivaizdus po kokaino vartojimo nutraukimo. Po 30 dienų abstinencijos, DA D1 receptorių ir DAT koncentracijos buvo žymiai padidintos visoje beždžionių striatume, turinčios lėtinio kokaino savęs vartojimo istoriją, lyginant su sustiprintais maisto produktais. Tačiau šiame tyrime taip pat pateikiami aiškūs įrodymai, kad DA sistemoje atsigavo po ilgesnių abstinencijos laikotarpių (90 dienų), o tai rodo, kad nėra didelių skirtumų tarp kokaino ekspozicijos ir kontroliuoja gyvūnai šiuo metu. Šie duomenys rodo, kad kokaino ekspozicija gali sukelti nuolatinį DA sistemos pakitimą, tačiau gali atsigauti po ilgalaikio susilaikymo nuo narkotikų vartojimo.
DAT koncentracijos reguliavimas, parodytas čia po abstinencijos, atitinka ankstesnius pranešimus apie nežmoginius primatus (Letchworthas et al, 2001), kuris parodė reikšmingą DAT surišimo vietų tankio padidėjimą tiek ventralinėje, tiek dorsalinėje stiatumoje. Nors tai nėra aiškiai išbandyta, atrodo, kad nutraukus vaisto ekspoziciją, DAT surišimo vietos tankio padidėjimas yra ne mažesnis kaip tokio dydžio ir plačiau paplitęs visuose striatum regionuose nei tie, kurie buvo nurodyti be pašalinimo laikotarpio (Letchworthas et al, 2001). Panašiai padidėjusios D1 receptorių surišimo vietų koncentracijos, pastebėtos po 30 abstinencijos dienų, taip pat atitinka ankstesnius tyrimus, kurie parodė, kad padidėjęs D1 receptorių surišimo tankis nežmoginių primatų striatume, veikiančiame identišką kokaino savarankiško vartojimo režimą (Nader et al, 2002). Priešingai, nebuvo pastebėta jokių reikšmingų skirtumų tarp D2 tipo receptorių surišimo tankio kokaino ekspozicijos ir kontrolinių gyvūnų striatume. Šis disreguliacijos trūkumas buvo pastebėtas, nepaisant didelių D2 receptorių koncentracijos sumažėjimo, apie kurį pranešta abiejose žmonių priklausomybėse (Volkow et al, 1993) ir kokaino savęs administravimo modeliai (Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Šie duomenys rodo, kad, palyginti su D1 panašiais receptoriais ir DAT, normalizuojama, kad būtų kontroliuojami lygiai šioje sistemoje. Apskritai, DA receptorių ir DAT pokyčiai aiškiai rodo, kad laikotarpis iš karto po kokaino savarankiško vartojimo nutraukimo yra labai geras, o vykstant DA sistemos reguliavimui vyksta žymūs pokyčiai, tačiau po to vėl keičiasi sistema, artėjanti labiau normaliam DA receptorių ir DAT pasiskirstymui po ilgesnio abstinencijos.
D1 receptorių pakeitimai
Plačiai paplitęs D1 receptorių tankis, pastebėtas nutraukus kokaino vartojimą, atitinka ataskaitas, rodančias padidėjusį D1 receptorių jautrumą nutraukimo metu. D1 jautrumo matavimas buvo atliktas atlikus tyrimą Henry ir White (1991), kuriame buvo nustatyta, kad vienoje branduolio accumbens neuronų vienetų įrašai yra jautresni D1 receptoriaus agonistui SKF 38393 po lėtinių kasdieninių kokaino injekcijų, lyginant su kontroliuojamais fiziologiniais tirpalais. Šis poveikis buvo patvarus, nes padidėjęs jautrumas išnyko iki vieno mėnesio. Autoriai teigė, kad D1 receptorių jautrinimas atsirado dėl D2 autoreceptoriaus jautrumo somatodendritinio A10 srityje, taip sumažinant slopinamąjį impulsų srautą per visą mezoaccumbens DA sistemą (Henry ir White, 1991). Šio tyrimo duomenys rodo, kad padidėjęs D1 receptorių susiejimo vieta yra abstinencijoje, o tai gali paaiškinti padidėjusį dopaminerginių neuronų jautrumą tiesioginio veikimo D1 receptorių agonistui. Be to, padidėjęs SKF 38393 poveikis branduolių accumbens neuronams nebuvo akivaizdus praėjus dviem mėnesiams po nutraukimo, o tai rodo, kad buvo nustatytas D1 receptorių jautrumas (Henry ir White, 1991); rezultatas, atitinkantis D1 receptorių tankumą, pastebėtą šiame tyrime po 90 abstinencijos dienų. Kitos ataskaitos taip pat palaiko svarbų D1 receptorių vaidmenį recidyvo metu. Tiesioginis D1 receptorių stimuliavimas branduolio accumbens lukštuose gali atkurti kokaino vartojimą abstinentiniuose graužikuose (Schmidt et al, 2006). Tačiau literatūroje šiek tiek prieštaraujama tuo, kad tiek D1 agonistai, tiek antagonistai gali susilpninti medikamentų, kuriuos sukelia kokaino prime arba kokaino sukeliami stimulai (Alleweireldt et al, 2002; De Vries et al, 1999; Khroyan et al, 2000; Savęs et al, 1996; Baltas et al, 2001). Neseniai Khroyan et al (2003) pranešė, kad D1 agonistai ir antagonistai mažina nepageidaujamą kokaino primimo modelį. Šie autoriai teigė, kad gali būti kritinis D1 receptorių aktyvumo diapazonas, reikalingas kokaino paieškai, ir kad abu antagonistai ir agonistai gali perkelti aktyvumą iš šio lango. Padidėjusios D1 receptorių koncentracijos, kurios lydi abstinenciją, gali pakeisti šį diapazoną, dėl kurio gali pasikeisti šios sistemos jautrumas. Kitas aspektas yra tai, kad D1 aktyvumas gali keisti aktyvumą D2 receptoriuose (Nolan et al, 2007; Ruskinas et al, 1999; Walters et al, 1987). Šie duomenys rodo, kad D1 ir D2 receptorių santykis abstinencijos metu keičiasi ir gali pakeisti šio moduliavimo veiksmingumą.
Nors, palyginti su dabartiniais duomenimis, buvo pranešta apie sumažėjusius D1 receptorių kiekius po ilgos kokaino savarankiško vartojimo.Moore et al, 1998a), tarp šių tyrimų yra didelių skirtumų, pavyzdžiui, kokaino dozė ir poveikio trukmė, bendras suvartojimas ir lyginamosios kontrolinės grupės. Todėl kartu suvokiantys įrodymai aiškiai rodo, kad pasitraukus iš lėtinio kokaino vartojimo, D1 sistema yra didelė.
Dopamino transporterio pakeitimai
DAT koncentracijos padidėjimas per kokaino poveikį patyrusių gyvūnų striatą po to, kai nutraukiamas narkotikų vartojimas, praplečia mūsų ankstesnių tyrimų rezultatus, rodančius, kad DAT surišimo vietų skaičius, susijęs su kokaino savarankišku vartojimu, yra nežmoginių primatų. Šie duomenys rodo, kad šis reguliavimas išlieka pradiniuose abstinencijos etapuose. Be to, jie rodo, kad atsigavimas iki kontrolinio lygio seka palyginti ilgą laiką (iki 90 dienų šioje studijoje). Ankstesniuose tyrimuose parodėme, kad, nors iš pradžių buvo apribota daugiausia ventralinė striatalo sritis, DAT surišimo vietų tankio pokyčiai išsiplėtė, kad apimtų daugiau dorsalinių ir rostrinių striatumo dalių su ilgesniais kokaino poveikio laikotarpiais (Letchworthas et al, 2001; Porrino et al, 2004). Šiame tyrime grįžtama prie kontrolinių DAT koncentracijų abstinencijos metu buvo didesnė ir greitesnė nugaros striatume nei ventralinėje stiatumoje, todėl atrodė, kad ji veikia atvirkštine anatomine trajektorija dėl lėtinio kokaino poveikio. .
Šie duomenys taip pat atitinka ataskaitas apie žmogaus kokaino vartotojus (Mažai et al, 1999; Malisonas et al, 1998; Misa et al, 2002; Staley et al, 1994), kurie, palyginti su kontroliniais preparatais, parodė, kad striatume yra daugiau jungimosi prie DAT vietų, o didžiausias padidėjimas yra ventralinėje stiatumoje. Pastaruoju metu šie padidėjimai parodė, kad kartu su 2 (VMAT2) suleidžiama daugybė vezikulinių monoamino transporterių (Mažai et al, 2003), rodo, kad DA neuronai yra realiai prarasti. Autoriai padarė išvadą, kad padidėjęs DAT greičiausiai atsirado dėl kokaino kompensacinio atsako į farmakologinį blokadą, o VMAT2 sumažėjimas labiau tikėtina, kad atspindėjo bendrus DA metabolizmo pokyčius, o tai sąlygojo hipodopaminerginę funkciją.
Nustatyta, kad žmogaus kokaino narkomanai sumažino DA koncentraciją ventralinėje striatoje, matuojant PET, reaguojant į metilfenidato poveikį, palyginti su sveikais kontroliniais preparatais (Volkow et al, 1997). Neseniai Martinez et al (2007) pranešė, kad kokaino vartotojas neatsakė į amfetamino poveikį ventraliniam striatui ir putamenui. Be to, šis amfetamino sukelto DA atpalaidavimo sumažėjimas buvo koreliuojamas su kokaino pasirinkimu atskirose savireguliavimo sesijose, kad vartotojai, turintys mažiausią DA išsiskyrimo laipsnį reaguojant į amfetaminą, greičiausiai pasirinko kokainą per alternatyvų gydytoją (Martinez et al, 2007). Neseniai atlikti tyrimai dėl kokaino vartojimo graužikų modelių taip pat palaiko šią idėją. Mateo et al (2005)Pavyzdžiui, pranešta, kad lėtinio kokaino savarankiško vartojimo poveikis yra susijęs su DAT funkcijos pokyčiais. Šie tyrėjai parodė, kad pradinis DA įsisavinimas buvo padidėjęs, dėl to atsirado greitesnis sinaptinės DA klirensas, todėl sumažėjo bazinis ekstraląstelinio DA kiekis arba hipodopaminerginė būsena. Taigi, tikėtina, kad padidėjusios DAT koncentracijos po to, kai nutraukiamas lėtinis kokaino savarankiškas vartojimas, pastebėtas šiame tyrime, yra kompensacinis atsakas, dėl kurio sumažėja ekstraląstelinio DA lygis.
D2 receptorių pakeitimai
Vienas iš šio tyrimo rezultatų buvo toks, kad D2 receptorių koncentracija vėl padidėjo po 30 abstinencijos dienų, lyginant su reikšmingu sumažėjimu gyvūnams, kuriems nenustatyta nutraukimo trukmė (Nader et al, 2002). Priešingai nei šiuo tyrimu, žmogaus vaizdavimo tyrimuose nustatyta, kad D2 receptorių kiekis yra mažesnis nei kontrolinių preparatų, vartojusių ilgalaikį susilaikymą nuo lėtinio kokaino poveikio (žr.Martinez et al, 2004; Volkow et al, 1993). Galimas šių žmogaus tyrimų ir dabartinio nežmoginio primato tyrimo skirtumų paaiškinimas apima kokaino vartojimo modelio ir trukmės skirtumus, taip pat galimybę iš anksto nustatyti mažesnius D2 receptorių kiekius žmonių narkomanuose.
Atsižvelgiant į pastarąją idėją, yra įrodymų, kad mažesni baziniai D2 receptorių kiekiai sveikiems žmonėms prognozuoja padidėjusį stimuliatorių, pvz. metilfenidatasirVolkow et al, 1999) ir panašiai beždžionėms, pradiniai D2 receptorių kiekiai prognozavo polinkį savarankiškai vartoti kokainą (\ tMorgan et al, 2002; Nader et al, 2006). Atsižvelgiant į tai, kad šie rezultatai yra lygiagrečiai būdingi kiekvienai rūšiai,Volkow et al, 1993) ir nežmoniškas primatas (Nader et al, 2006) vaizdavimo tyrimai parodė mažesnį D2 receptorių prieinamumo lygį po susilaikymo nuo kokaino poveikio. Pažymėtina, kad kokaino savarankiško vartojimo grafikas paskutiniame eksperimente (Nader et al, 2006) buvo panašus į šiame tyrime naudojamą sustiprinimo grafiką. Taigi, mažai tikėtina, kad šių dviejų tyrimų rezultatai yra susiję su metodologiniais skirtumais, pvz., Sustiprinimo ar suvartojimo per kokainą savanoriško vartojimo metu.
Labiau tikėtinas paaiškinimas susijęs su DA sistemos funkcine dinamika. D2 receptorių prieinamumo su PET priemonės apibūdinamos kaip „funkcinės“, nes signalas yra susijęs su baltymų kiekiu (šiuo atveju D2 receptorių tankiu) ir cirkuliuojančio neurotransmiterio kiekiu. (Žr. Laruelle, 2000; Nader ir Czoty, 2008 tolesnėms diskusijoms). Ipriešingai, receptorių autoradiografija yra neužteršta cirkuliuojančiais DA lygiais. Taigi, šiame tyrime, kartu su ankstesniu darbu, galima teigti, kad D2 receptorių tankis sumažėja dėl kokaino savarankiško vartojimo, tačiau receptorių lygis atsigauna abstinencijos metu. Panašiai atliktame PET tyrime (Nader et al, 2006), atsigavimas buvo pastebėtas trims iš penkių beždžionių. Dabartiniai duomenys rodo, kad šie beždžionės D2 receptorių tankiuose tikriausiai nesiskyrė, tačiau galbūt DA sistemos reakcija (ty cirkuliuojančios DA koncentracija abstinencijos metu) diferencijuojama tarp „susigrąžintų“ ir „negrąžintų“ asmenų.
Trūkumai
Svarbus dabartinių tyrimų apribojimas yra tas, kad mūsų tyrimuose negalima aptarti D1 ir D2 receptorių arba DAT funkcionalumo. Mes tikrinome tik receptorių baltymų tankio pokyčius. Nors rezultatai turi įtakos galimai šių sistemų vaidmenims, reikės atlikti tolesnius tyrimus, kad sužinotumėte čia pateiktų pakeitimų elgesio pasekmes. Kitas šio tyrimo apribojimas yra tas, kad autoradiografiniai ligandai dažnai nesiskiria tarp jų ląstelių ir citoplazminių vietų, nes radioaktyviai pažymėti antagonistai dažnai yra membraniniai. Mažai et al (2002) parodė, kad lėtinis kokaino poveikis lemia reikšmingą DAT reguliavimą membranos paviršiuje, tuo pačiu metu sumažėjusį ląstelių DAT koncentraciją transfekuotose ląstelėse. Neseniai Samuvel et al (2008) panašių tyrimų su žiurkių striatų sinaptosominiais preparatais. Šie rezultatai rodo, kad šiame tyrime pastebėti DAT pasiskirstymo pokyčiai gali rodyti pokyčius membranos paviršiuje, o ne ląstelėse.
Galiausiai, reikia laikytis atsargumo priemonių dėl ilgalaikio (90 dienų) abstinencijos tyrimų, nes šie duomenys buvo pagrįsti santykinai maža gyvūnų grupe (N=3). Nepaisant nedidelio tiriamųjų skaičiaus, iš šios grupės gauti duomenys buvo gana nuoseklūs, kaip matyti iš sklaidos diagramos, parodyta 2 pav. D1 receptorių surišimo vietų koncentracija per striatumą parodė, kad grupės viduje kinta nedaug, o tai rodo šių tyrimų patikimumą. Panašus nuoseklumas taip pat buvo akivaizdus D2 receptorių ir DAT surišimo tyrimų duomenyse. Nors reikia laikytis atsargumo priemonių, šie duomenys rodo, kad DAT ir DA receptorių koncentracijų atkūrimas striatume gali pasireikšti ilgai susilaikius.
2 pav.
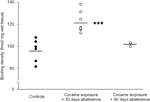 D1 receptorių rišamosios tankiai atskiriems gyvūnams vidutiniškai apskaičiuoti per visą striatumą po to, kai buvo sustiprintas maisto stiprinimas (kontrolė), arba 30 arba 90 dienų abstinencija nuo lėtinio kokaino savarankiško vartojimo. Grupėms skirtos priemonės vaizduojamos juodomis juostomis, ***p<0.001, palyginti su maisto sustiprintomis kontrolėmis.
D1 receptorių rišamosios tankiai atskiriems gyvūnams vidutiniškai apskaičiuoti per visą striatumą po to, kai buvo sustiprintas maisto stiprinimas (kontrolė), arba 30 arba 90 dienų abstinencija nuo lėtinio kokaino savarankiško vartojimo. Grupėms skirtos priemonės vaizduojamos juodomis juostomis, ***p<0.001, palyginti su maisto sustiprintomis kontrolėmis.
IŠVADOS
Apibendrinant galima pasakyti, kad kokaino savarankiško vartojimo poveikis sukėlė reikšmingų DA sistemų reguliavimo pokyčių, kurie išliko ankstyvosiose abstinencijos fazėse (pirmosiose 30 dienose). Tai labiausiai matyti D1 receptorių ir DAT koncentracijos reguliavime, atsižvelgiant į jų pokyčių dydį ir topografinį mastą. Priešingai, buvo įrodyta, kad normalizavosi ilgiau trukusio susilaikymo nuo kokaino vartojimo trukmė, nes DAT, D1 ir D2 receptorių koncentracijos po 90 abstinencijos dienų nesiskyrė nuo nekontroliuojamų kontrolinių preparatų. Ttačiau šios sistemos nebūtinai atitinka tą pačią laiko atkūrimo eigą, o tai rodo, kad yra tikėtinas DA lygių reguliavimo nestabilumas, ypač anksti susilaikius.. Šis dopaminerginis disreguliavimas gali turėti įtakos bet kokių galimų farmakologinių vaistų, vartojamų priklausomiems nuo kokaino vaistams, veiksmingumui, ypač jei vaisto veikimo mechanizmas priklauso nuo DA sistemos.
pastabos
ATSKLEIDIMAS/INTERESŲ KONFLIKTAS
Autoriai neturi interesų konflikto atskleisti.
Nuorodos
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). D1 dopamino receptorių blokada arba stimuliavimas slopina žiurkių elgesį su išnykusiu kokainu. Psichofarmakologija (Berl) 159: 284–293. | Straipsnis | PubMed | ChemPort |
- Beveridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Lėtinis kokaino savarankiškas vartojimas yra susijęs su kintančių funkcijų aktyvumu nežmoginių primatų laikinose skiltyse. Eur J Neurosci 23: 3109–3118. | Straipsnis | PubMed |
- Canfield DR, Spealman RD, Kaufman MJ, Madras BK (1990). Kokaino surišimo vietų autoradiografinis lokalizavimas [3H] CFT ([3H] WIN 35428) beždžionių smegenyse. Synapse 6: 189–195. | Straipsnis | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Dopaminerginiai mechanizmai, skatinantys ieškoti kokaino ir heroino po ilgalaikio IV vaisto savarankiško vartojimo nutraukimo. Psichofarmakologija (Berl) 143: 254–260. | Straipsnis | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Kartotinių kokaino injekcijų poveikis katecholamino receptorių surišimo vietoms, dopamino transporterio prisijungimo vietoms ir elgesiui reeso beždžionėje. Smegenų raiška 578: 235–243. | Straipsnis | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Kokaino savivaldos neuroanatominiai pagrindai. CRC: Boca Raton, FL.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Ekstraceliulinės dopamino koncentracijos skirtumai branduolio accumbens reakcijoje ir atsako nepriklausomame kokaino vartojime žiurkėms. Psichofarmakologija (Berl) 133: 7–16. | Straipsnis | PubMed | ChemPort |
- Henry DJ, White FJ (1991). Pakartotinis kokaino vartojimas sukelia nuolatinį D1 dopamino receptorių jautrumo padidėjimą žiurkių branduolyje. J Pharmacol Exp Ther 258: 882–890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR et al (2000). Padidėjęs centrinio serotonino transporterio prisijungimo prie akutinai priklausomų pacientų, priklausančių nuo kokaino, prieinamumas. Aš esu psichiatrija 157: 1134–1140. | Straipsnis | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Dopamino D1 ir D2 tipo receptorių mechanizmai recidyvuojant kokainą ieškančiam elgesiui: selektyvių antagonistų ir agonistų poveikis. J Pharmacol Exp Ther 294: 680–687. | PubMed | ISI | ChemPort |
- Khroyan TV, Platt DM, Rowlett JK, Spealman RD (2003). Dokamino D1 receptorių agonistų ir antagonistų, vartojamų nežmoginių primatų, recidyvo sumažėjimas. Psichofarmakologija (Berl) 168: 124–131. | Straipsnis | PubMed | ChemPort |
- Laruelle M (2000). Vaizdo sinchroninis neurotransmisija su in vivo privalomos konkurencijos metodai: kritinė apžvalga. J Cereb kraujo srauto metab 20: 423–451. | Straipsnis | PubMed | ISI | ChemPort |
- Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ (2001). Dopamino transporterio pokyčių pokyčiai, atsiradę dėl kokaino savarankiško vartojimo reeso beždžionėse. J Neuroscience 21: 2799–2807. | PubMed | ISI | ChemPort |
- Lidow MS, Goldman-Rakic PS, Gallager DW, Rakic P (1991). Dopaminerginių receptorių pasiskirstymas primityviame smegenų žieve: kiekybinė autoradiografinė analizė naudojant [3H] raclopride, [3H] spiperoną ir [3H] SCH 23390. Neurologijos 40: 657–671. | Straipsnis | PubMed | ChemPort |
- Little KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Kokaino dopamino transporterio prekyba žmonėmis į plazmos membraną. Mol Pharmacol 61: 436–445. | Straipsnis | PubMed | ISI | ChemPort |
- Little KY, Krolewski DM, Zhang L, Cassin BJ (2003). Žmogaus kokaino vartotojas praranda strialinį vezikulinį monoamino transporterio baltymą (VMAT2). Aš esu psichiatrija 160: 47–55. | Straipsnis | PubMed |
- Little KY, Zhang L, Desmond T, Frey KA, Dalack GW, Cassin BJ (1999). Žmogaus kokaino vartojantiems pacientams pasireiškė stuburo dopaminerginiai sutrikimai. Aš esu psichiatrija 156: 238–245. | PubMed | ChemPort |
- Malison RT, Best SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M et al (1998). Padidėjęs striatalo dopamino transporteris ūminio kokaino susilaikymo metu, išmatuotas pagal [123I] beta-CIT SPECT. Aš esu psichiatrija 155: 832–834. | PubMed | ChemPort |
- Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al (2004). Kokaino priklausomybė ir d2 receptorių prieinamumas funkciniuose striatumo padaliniuose: ryšys su elgesiu su kokainu. Neuropsychopharmacology 29: 1190–1202. | Straipsnis | PubMed | ISI | ChemPort |
- Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A et al (2007). Amfetamino sukeltas dopamino išsiskyrimas: akivaizdžiai sumažėjo priklausomybė nuo kokaino ir prognozuojama, kad pasirenkama savarankiškai vartoti kokainą. Aš esu psichiatrija 164: 622–629. | Straipsnis | PubMed | ISI |
- Mash DC, Pablo J, Ouyang Q, Hearn WL, Izenwasser S (2002). Kokaino vartotojai padidina dopamino transportavimo funkciją. J Ncurochcm 81: 292–300. | Straipsnis | PubMed | ChemPort |
- Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005). Sumažėjusi dopamino galinė funkcija ir nejautrumas kokainui po savarankiško savarankiško vartojimo ir netekimo. Neuropsychopharmacology 30: 1455–1463. | Straipsnis | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G et al (2008). Ilgalaikis metamfetamino vartojimas beždžionių modelių aspektuose, susijusiuose su žmogaus poveikiu: smegenų neurotoksiškumas ir elgesio profiliai. Neuropsychopharmacology 33: 1441–1452. | Straipsnis | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998a). Kokaino savarankiško vartojimo poveikis striatų dopamino D1 receptoriams reeso beždžionėse. Synapse 28: 1–9. | Straipsnis | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998b). Kokaino savarankiško vartojimo poveikis dopamino D2 receptoriams reeso beždžionėse. Synapse 30: 88–96. | Straipsnis | PubMed | ChemPort |
- Morgan D, Grant KA, Gage HD, Mach RH, Kaplanas JR, Prioleau O et al (2002). Socialinis dominavimas beždžionėse: dopamino D2 receptoriai ir kokaino savęs administravimas. Nat Neurosci 5: 169–174. | Straipsnis | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Smegenų vaizdavimas nežmoginiuose primatuose: narkomanijos įžvalgos. ILAR J 49: 89–102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR et al (2002). Kokaino savęs vartojimo poveikis striatų dopamino sistemoms reeso beždžionėse: pradinis ir lėtinis poveikis. Neuropsychopharmacology 27: 35–46. | Straipsnis | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al (2006). Dopamino D2 receptorių PET vaizdavimas per chronišką kokaino savarankišką vartojimą beždžionėms. Nat Neurosci 9: 1050–1056. | Straipsnis | PubMed | ChemPort |
- Nolan EB, Harrison LM, Lahoste GJ, Ruskin DN (2007). Elgesio sinergizmas tarp D (1) ir D (2) dopamino receptorių pelėse nepriklauso nuo tarpelių jungčių. Synapse 61: 279–287. | Straipsnis | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB et al (2002). Metabolinis kokaino poveikio žemėlapis pradiniuose savireguliacijos etapuose nežmogiškame primate. J Neuroscience 22: 7687–7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Kokaino savarankiškas vartojimas suteikia laipsnišką limbinės, asociacijos ir sensorių striatūros domenų dalyvavimą. J Neuroscience 24: 3554–3562. | Straipsnis | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Multisekundiniai svyravimai degimo spartoje globus pallidus: sinerginis moduliavimas pagal D1 ir D2 dopamino receptorius. J Pharmacol Exp Ther 290: 1493–1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, žr. RE, Ramamoorthy S (2008). Dopamino transporterio kontrabanda ir funkcijos po susilaikymo nuo kokaino savarankiško patekimo į žiurkes: diferencinio reguliavimo caudate putamen ir nucleus accumbens įrodymai. J Pharmacol Exp Ther 325: 293–301. | Straipsnis | PubMed | ChemPort |
- Schmidt HD, Anderson SM, Pierce RC (2006). D1 tipo arba D2 dopamino receptorių stimuliavimas branduolio accumbens lukštuose, bet ne šerdyje, atkuria kokainą ieškantį elgesį žiurkėse. Eur J Neurosci 23: 219–228. | Straipsnis | PubMed |
- Savęs DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). Priešais D1 ir D2 panašių dopamino receptorių agonistų kokaino paieškos elgesį. Mokslas 271: 1586–1589. | Straipsnis | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Didelės afiniškumo kokaino atpažinimo vietos, esančios dopamino transporteryje, yra padidėjusios dėl mirtinų kokaino perdozavimo aukų. J Pharmacol Exp Ther 271: 1678–1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al (1993). Sumažėjęs dopamino D2 receptorių prieinamumas yra susijęs su sumažėjusiu priekinio metabolizmo poveikiu kokaino vartojantiems asmenims. Synapse 14: 169–177. | Straipsnis | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A et al (1999). Prognozuojama, kad žmogaus smegenų dopamino D2 receptorių koncentracija sustiprins psichostimuliatorių reakciją. Aš esu psichiatrija 156: 1440–1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R et al (1997). Sumažėjęs striatalo dopaminerginis jautrumas nuo detoksikuotų kokaino priklausomų asmenų. Gamta 386: 830–833. | Straipsnis | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). D1 dopamino receptorių aktyvacija, reikalinga D2 agonistų poveikio postinaptinei ekspresijai. Mokslas 236: 719–722. | Straipsnis | PubMed | ChemPort |
- Weiss F, Martin-Fardon R, Ciccocioppo R, Kerr TM, Smith DL, Ben-Shahar O (2001). Ilgalaikis atsparumas su narkotikais susijusių užuominų sukeltam kokaino paieškos elgesiui. Neuropsychopharmacology 25: 361–372. | Straipsnis | PubMed | ISI | ChemPort |
- Wong DF, Kuwabara H, Schretlen DJ, Bonson KR, Zhou Y, Nandi A et al (2006). Padidėjęs dopamino receptorių įsisavinimas žmogaus striatume, kai pasireiškia kokaino troškimas. Neuropsychopharmacology 31: 2716–2727. | Straipsnis | PubMed | ChemPort |
