 Martinas Zackas
Martinas Zackas- 1Kognityvinės psichofarmakologijos laboratorija, Neurologijos departamentas, Priklausomybės ir psichinės sveikatos centras, Torontas, ON, Kanada
- 2Transliacinė neurologijos programa, Pensilvanijos universiteto Medicinos mokyklos psichiatrijos skyrius, Filadelfija, PA, JAV
- 3Biopsichologijos skyrius, Neurologijos skyrius, Priklausomybės ir psichikos sveikatos centras, Torontas, ON, Kanada
Priklausomybė laikoma smegenų liga, kurią sukelia lėtinis narkotikų poveikis. Smegenų dopamino (DA) sistemų jautrinimas iš dalies sukelia šį poveikį. Patologinis lošimas (PG) laikomas elgesio priklausomybe. Todėl PG gali atsirasti dėl lėtinio lošimo poveikio. Šią galimybę patvirtintų azartinių lošimų sukelto DA sistemų jautrinimo nustatymas. Azartinių lošimų apdovanojimai primena DA išleidimą. Vienas lošimo automatų žaidimo epizodas perkelia DA atsaką nuo atlygio įteikimo iki užuominų (sukančių būgnų) už atlygį pradžią, laikantis laiko skirtumų mokymosi principų. Taigi sąlyginiai dirgikliai (CS) vaidina pagrindinį vaidmenį DA atsakuose į lošimą. Primatų DA atsakas į CS yra stipriausias, kai atlygio tikimybė yra 50%. Pagal šį tvarkaraštį CS tikimasi atlygio, tačiau nepateikia informacijos apie tai, ar jis įvyks tam tikro bandymo metu. Lošimo metu 50 % tvarkaraštis turėtų sukelti maksimalų DA išleidimą. Tai labai atitinka atlygio dažnumą (46 %) komerciniuose lošimo automatuose. DA išsiskyrimas gali prisidėti prie jautrinimo, ypač amfetamino. Lėtinis CS poveikis, kuris numato atlygį 50 % laiko, gali imituoti šį poveikį. Šią hipotezę išbandėme trijuose tyrimuose su žiurkėmis. Gyvūnai gavo 15 × 45 minučių ekspoziciją CS, kuri numatė atlygį su 0, 25, 50, 75 arba 100% tikimybe. CS buvo lengvas; atlygis buvo 10% sacharozės tirpalas. Po treniruotės žiurkės gavo jautrinamąjį režimą – penkias atskiras d-amfetamino dozes (1 mg/kg). Galiausiai prieš 0.5 minučių trukmės lokomotorinio aktyvumo testą jie gavo 1 arba 90 mg/kg amfetamino. Visuose trijuose tyrimuose 50 % grupė pasižymėjo didesniu aktyvumu nei kitos grupės reaguojant į abi provokacines dozes. Poveikio dydžiai buvo nedideli, bet nuoseklūs, kaip rodo reikšminga grupės × rango asociacija (ϕ = 0.986, p = 0.025). Lėtinis į azartinius lošimus panašus atlygio nuspėjamųjų dirgiklių grafiko poveikis gali paskatinti jautrumą amfetaminui, panašiai kaip paties amfetamino poveikis.
Įvadas
Priklausomybė buvo apibūdinama kaip smegenų liga, kurią sukelia lėtinis piktnaudžiavimo narkotikais poveikis (Leshner, 1997). Manoma, kad neuroplastiškumas tarpininkauja tokio poveikio padariniams („Nestler“, „2001“). Smegenų dopamino (DA) sistemų jautrinimas yra neuroplastiškumo forma, susijusi su hiperreaktyvumu į sąlyginius vaistų dirgiklius (CS) ir kompulsyviu narkotikų ieškojimu.Robinson ir Berridge, 2001). Jautrinimas buvo apibrėžiamas kaip padidėjęs DA išsiskyrimas reaguojant į CS už atlygį ir padidėjęs lokomotorinis atsakas į farmakologinį DA iššūkį (Robinson ir Berridge, 1993; Pierce ir Kalivas, 1997; Vanderschuren ir Kalivas, 2000). Nors jautrinimas yra tik vienas iš daugelio smegenų pokyčių, susijusių su priklausomybe (plg. Robbins ir Everitt, 1999; Koob ir Le Moal, 2008), buvo pasiūlyta, kad presinapsinio dopamino išsiskyrimo pokyčiai atspindi įprastas neuroadaptacijas, susijusias su narkomanija, susijusia su priklausomybe (pvz., atkryčiu), kai vaistai sukelia lokomotorinį jautrumą opiatams (pvz., morfijui) arba stimuliuojančių medžiagų poveikiui (pvz., amfetaminui). , taip pat atkuria užgesusias operacines reakcijas į heroino ar kokaino savarankišką vartojimą – tai gyvūno atkryčio modelis (Vanderschuren ir kt., 1999). Įrodymai, kad skatinamasis jautrinimas (padidėjusi atlygio už narkotikus vertė) yra ryškiausias po pradinio poveikio priklausomybę sukeliantiems narkotikams, taip pat rodo, kad jautrinimas gali būti susijęs ir su ankstyvomis priklausomybės stadijomis (Vanderschuren ir Pierce, 2010).
Patologinis lošimas (PG) buvo apibūdintas kaip elgesio priklausomybė ir neseniai 5-ajame Psichikos sutrikimų diagnostikos ir statistikos vadovo leidime perklasifikuotas į tą pačią kategoriją kaip priklausomybės nuo narkotikų sutrikimai.Frascella ir kt., 2010; APA, 2013 m). Tai reiškia, kad PG gali atsirasti dėl lėtinio į azartinius lošimus panašios veiklos, kad bendri mechanizmai gali tarpininkauti lošimo ir narkotikų poveikio (Zack ir Poulos, 2009; Leeman ir Potenza, 2012); ir tai, kad smegenų DA trajektorijų jautrinimas gali būti svarbus šio proceso elementas.
Klinikiniai įrodymai netiesiogiai patvirtina šią galimybę: Naudodami pozitronų emisijos tomografiją (PET) Boileau ir jo kolegos nustatė, kad PG tiriamųjų vyrų striatalinis DA išsiskyrimas yra žymiai didesnis, reaguodamas į amfetaminą (0.4 mg/kg), nei sveikų kontrolinių vyrų (Boileau ir kt., 2013). Bendri grupių skirtumai buvo reikšmingi asociatyviniame ir somatosensoriniame striatum. Limbiniame striatum, kuriame yra nucleus accumbens, grupės nesiskyrė. Tačiau PG tiriamiesiems DA išsiskyrimas limbiniame striate tiesiogiai koreliavo su PG simptomų sunkumu. Šie radiniai atitinka smegenų DA takų jautrinimą PG, bet taip pat rodo keletą svarbių skirtumų su nuo žmogaus medžiagų priklausomais asmenimis ir klasikiniu gyvūnų jautrinimo amfetaminu modeliu. Skirtingai nei PG tiriamieji ir gyvūnai, veikiami mažomis amfetamino dozėmis (plg. Robinson ir kt., 1982), žmonėms, turintiems priklausomybę nuo medžiagų, nuolat sumažėja DA išsiskyrimas į stimuliatorių (Volkow ir kt., 1997; Martinez ir kt., 2007), o gyvūnų gauti duomenys rodo, kad tai gali atspindėti DA funkcijos trūkumą pradinėse abstinencijos stadijose po besaikio piktnaudžiavimo medžiagomis (Mateo ir kt., 2005). Atliekant tyrimus, kuriuose buvo įrodytas jautrinimas stimuliatoriams su gyvūnais, padidėjęs DA išsiskyrimas dažniausiai stebimas limbiniame striatum, o ne nugariniame (asociaciniame, somatosensoriniame) striatum.Vezina, 2004). Tačiau užuominų sukeltas (ty sąlyginis) narkotikų ieškojimas gyvūnams, pakartotinai veikiamiems kokaino, buvo susijęs su padidėjusiu DA išsiskyrimu nugaros juostoje – manoma, kad rezultatas rodo įprastesnę motyvuoto elgesio formą.Ito ir kt., 2002). Taigi bendras DA išsiskyrimo padidėjimas PG tiriamųjų nugaros regionuose gali būti susijęs su įpročiu pagrįstu (nelanksčiu, įprastu) atlygio siekimu, apimančiu „progresavimą nuo ventralinės iki nugarinės striatum srities“ (Everitt ir Robbins, 2005, p. 1481), kadangi nuo šiol tiriamųjų asmenų, priklausančių nuo DA, sunkumas priklauso nuo stimuliacinio jautrumo, kaip paprastai modeliuojama gyvūnams. PET rezultatai neatskleidžia, ar DA hipergeaktyvumas buvo anksčiau buvęs šių PG subjektų požymis, galintis sukelti azartinių lošimų poveikį, ar visiškai kito proceso rezultatas. Norint išspręsti šį klausimą, būtina įrodyti, kad chroniškų lošimų poveikis yra jautrus žmonėms, kurie yra normalūs prieš poveikį. Dėl to kyla klausimų dėl to, kokie lošimo požymiai gali sukelti jautrumą.
Skinneris pažymėjo, kad kintamas stiprinimo grafikas buvo esminis azartinių lošimų patrauklumo (ar bent jau jo išlikimo) veiksnys.Skinner, 1953). Lažybų elgesys lošimo automatų žaidime gerai atitinka pagrindinius instrumentinio kondicionavimo principus, kuriuos atspindi numatoma koreliacija tarp piniginės išmokos ir statymo dydžio už nuoseklius sukimus (Tremblay ir kt., 2011). Taigi, kintamasis koeficientas operantas reaguoja, atrodo, suteikia išoriškai galiojantį lošimo automatų lošimo modelį.
Naujausi tyrimai su gyvūnais suteikia tvirtą pradinį pagrindą priežastiniam lošimo poveikiui jautrinimui. Dainininkas ir jo kolegos ištyrė 55 1 val. kasdienių fiksuoto (FR20) arba kintamo (VR20) sacharino sustiprinimo seansų operantinio svirties paspaudimo paradigmoje poveikį tolesniam lokomotoriniam atsakui į mažą amfetamino dozę (0.5 mg/kg) sveikiems vyrams. Sprague Dawley) žiurkės (Singer ir kt., 2012). Jie teigė, kad jei azartiniai lošimai sukels jautrumą, žiurkėms, veikiančioms kintamąjį grafiką, kuris imituoja azartinius lošimus, turėtų būti didesnis atsakas į amfetaminą, nei žiurkėms, veikiančioms nustatytą grafiką. Kaip prognozuota, VR20 grupė rodė 50% didesnį lokomotorinį atsaką į amfetaminą nei FR20 grupė. Priešingai, grupėms pasireiškė lygiavertis judėjimas po fiziologinio tirpalo injekcijos. Šie duomenys patvirtina, kad lėtinis kintamojo sutvirtinimo poveikis yra pakankamas, kad sukeltų hiperreaktyvumą DA poveikiui sveikiems gyvūnams, atsitiktinai suskirstytiems į atitinkamus grafikus.
Iš šio rezultato kyla daug klausimų: pirma, kokiu mastu suvokiamas atsitiktinumas – ar jo nebuvimas – tarp operantinio atsako ir jo rezultato tarpininkauja šiems poveikiams? Kalbant apie mokymosi terminus, ar šis poveikis apima „tikėtiną atsako rezultato tikimybę“, ar panašus poveikis gali būti matomas nesant operantinio atsako, ty „stimuliatoriaus ir rezultato tikėtinos“ pagal Pavlovo paradigmą (plg. Bolles, 1972)? Antra, ar ankstesnio įvykio (atsako ar stimulo) ir jo rezultato kontingento laipsnis turi įtakos jautrumo laipsniui?
Antrasis klausimas yra susijęs su neapibrėžtumo svarba. Pavyzdžiui, ar žaidimai, kurių rezultatas yra tikrai atsitiktinis - visiškai nenuspėjami, turi didesnį potencialą sukelti jautrumą nei žaidimai, kuriuose laimėjimo šansai yra aiškiai apibrėžti, bet ne atsitiktiniai, net jei absoliutus atlygio lygis yra mažas? Šiame tyrime nagrinėjami šie klausimai.
Eksperimento planas buvo pagrįstas pradiniu tyrimu dėl atlygio tikėjimo ir DA neuronų atsako beždžionėms (Fiorillo ir kt., 2003). Šio tyrimo metu gauti gyvūnai gavo atlygį už sultis (US) pagal 0, 25, 50, 75 arba 100% kintamųjų santykių grafikus. Grafikai buvo pažymėti 1 iš 4 skirtingų CS (piktogramos). 0% grafikas pristatė atlygį taip dažnai, kaip ir 100% grafikas, bet praleido CS. DA neuronų degimo greitis per laikotarpį tarp CS pradžios ir JAV pristatymo ar neveikimo buvo pagrindinė priklausoma priemonė. Tyrimas parodė, kad DA atsakas padidėjo kaip atlygio pristatymo netikrumo funkcija. Taigi pagal 100% schemą CS sukėlė nedidelį aktyvumą, pagal 25 ir 75% grafikus, CS sukėlė vidutinį ir panašų aktyvumo lygį, o pagal 50% schemą CS sukėlė maksimalų aktyvumą. Kiekvienu atveju šaudymo greitis padidėjo per CS-US intervalą, ty, kaip tikėtina, kad tikėtina, kad tikėtasi.
Šie faktai rodo, kad DA veikla ne tik kinta, ar atlygis yra tikras (fiksuotasis santykis), ar nežinomas (kintamasis santykis), bet taip pat kinta atvirkščiai proporcingai informacijos apie atlygį pristatymui, kurį teikia CS. Esant 100% sąlygoms, CS sukelia tikėtiną atlygį ir puikiai prognozuoja jo pristatymą. Esant 25 ir 75% sąlygoms, CS sukelia tikimybę ir prognozuoja atlygio pristatymą tris kartus iš keturių kartų. Esant 50% sąlygai, CS sukelia tikėtiną laiką, bet nesuteikia informacijos apie atlygį tik po tikimybės. Remdamiesi savo duomenimis, Fiorillo ir kt. padarė išvadą: „Šis neapibrėžtumas padidėjęs dopamino kiekis gali prisidėti prie naudingų lošimų savybių“ (p. 1901).
50 % kintamo atlygio per vieną seanso poveikis neturėtų keistis per kelis seansus, nes kiekvieno bandymo metu atlygio tikimybė išlieka visiškai nenuspėjama. Taigi, svarstant sąlygas, kurios maksimaliai padidintų lėtinį DA neuronų aktyvavimą pasikartojančių lošimo epizodų metu, 50 % tvarkaraštis turėtų sukelti patvariausią ir stipriausią poveikį. Tai pažymėtina, nes ilgalaikis atlygio koeficientas (išmokėjimas > 0), stebimas tūkstančius sukimų komerciniame lošimo automate, buvo 45.8 % (Tremblay ir kt., 2011). Taigi, 50% kintamasis atlygis tiksliai atspindi mokėjimų grafiką, kurį valdo tikrieji lošimo įrenginiai.
Šiame tyrime buvo naudojami tie patys kondicionavimo grafikai kaip ir Fiorillo ir kt. lėtinio poveikio atveju tarp grupių su žiurkėmis. Gyvūnams buvo atliekamos ~ 3 savaičių kasdieninės kondicionavimo sesijos, kur CS (šviesa) buvo sujungta su JAV (nedidelis sacharozės kiekis). Po dresūros fazės gyvūnai pailsėjo prieš įvertindami jautrumą, kurį indeksavo judėjimo reakcija į amfetaminą. Remiantis literatūra, buvo prognozuota, kad žiurkėms, kurioms taikomas skirtingas atlygio grafikas, nesiskirtų jų judėjimo be narkotikų elgesys, tačiau po amfetamino atsirastų žymiai skirtingi judesio lygiai, o 50% grupei būdavo didesnis judėjimo reakcija į vaistą, kitoms grupėms dozių metu, kurio galima tikėtis, jei 50% gyvūnų anksčiau buvo paveiktos papildomos amfetamino dozės (ty kryžminis jautrinimas).
Eksperimentuokite 1
Medžiagos ir metodai
Tematika
Keturios grupės (n = 8 / grupė) suaugusių (300 – 350 g) žiurkių Sprague-Dawley žiurkių (Charles River, St. Constant, Kvebekas, Kanada) buvo atskirai laikomos atvirose polikarbonato dėžutėse (20 × 43 × 22 cm), esant atvirkštiniam 12: 12 šviesos ir tamsos ciklas. Jie gavo ad libitum galimybė gauti maisto ir vandens, o eksperimentuotojas 2 savaites prieš tyrimą kasdien tvarko. Kiekviena grupė buvo kondicionuojama pagal vieną iš keturių kintamų atlygio grafikų: 0, 25, 50 arba 100%. 75% grupė šiame pradiniame tyrime buvo praleista, kaip Fiorillo ir kt. (2003 m.) nustatė lygiavertį CS DA išleidimą pagal 25 ir 75% atlygio grafikus, todėl abi sąlygos lėmė didesnį DA išleidimą nei 100% CS-US sąlyga, bet mažiau nei 50% sąlyga.
Aparatūra
Prieiga prie sacharozės pateikimo ir CS buvo teikiama atskirai operantų kondicionavimo dėžėse (33 × 31 × 29 cm). Kiekvienoje dėžutėje buvo įrengtas žurnalas, esantis ant priekinės sienelės. Žurnalas žurnalo viršuje tarnavo kaip CS. Į žurnalo grindis gali būti pakeltas motorinis, solenoidinis valdomas skystis. Rinkinyje esančius įvykius kontroliavo „Med Associates“ įranga ir programinė įranga, naudojant vidinę programą, parašytą MED-PC. Lokomotoriniai bandymai buvo atlikti individualiai plexiglas narveliuose (27 × 48 × 20 cm). Kiekviename narve buvo sumontuota stebėjimo sistema, sudaryta iš šešių foto spindulių ląstelių, kad būtų galima nustatyti horizontalų judėjimą.
Procedūra
mokymas. Tyrimas buvo atliktas laikantis Kanados gyvūnų priežiūros tarybos nustatytų etinių gairių. Tyrimo metu žiurkės buvo maistui ribotos iki 90% savo kūno svorio ir laikomos atskirai. Kiekvienas žiurkė gavo 15 dienų sacharozės atlyginimų mokymą (10% vandens tirpalas 0.06 ml už atlygį): 5 iš eilės einančios dienos × 3 savaitės su savaitgaliais. Prieš ir po treniruočių fazės gyvūnai buvo laikomi standartiniu būdu. sacharozės ekspozicija apsiribojo penkiolika 40 min. treniruočių. Kiekvieną dieną sesija susideda iš 15 stimulų pateikimo (šviesos, CS), kurių kiekviena buvo atskirta tarpininkaujant 120 s. Šviesa buvo laikoma viršutiniame žurnalo skydelyje ir liko 25 s, su sacharoze per paskutinius 5 s. 0 grupės atveju sacharozės nuleidimo priemonė buvo pakelta kas 140 s (5 s), tačiau stimuliatoriaus šviesa nebuvo apšviesta. Tai prilygsta intervalui tarp nuleidimo 0 grupėje ir kitų grupių (120 + 25 s). Kiekviena gydymo sesija truko ~ 40 min. Vidutiniškai 25 grupė kartą per keturis CS pristatymus gavo sacharozę; 50 grupė gavo vieną sacharozę kiekvienam dviem CS pristatymams, o 100 grupė po kiekvieno CS pateikimo gavo sacharozę.
Testavimas. Praėjus dviem savaitėms po paskutinės sacharozės prieigos (arba „kondicionavimo“) sesijos, buvo įvertintas lokomotorinis atsakas į d-amfetaminą (AMPH; ip). Žiurkėms buvo suteiktos trys 2-h sesijos, norint priprasti prie lokomotyvų dėžių, po kurių sekė šešios AMPH bandymų sesijos. AMPH bandymo dienos įvyko 1-wk intervalais. Bandymo dienomis žiurkėms buvo duodama 30 min, kad būtų galima priprasti prie dėžių, tada gavo vieną 0.5 mg / kg AMPH dozę, o po atskirų savaitės sesijų - penkiomis 1.0 mg / kg dozėmis (viena dozė per dieną) tiriamosiomis dienomis 1 per 5 . Kiekvienos sesijos metu po AMPH lokomotyvo buvo vertinama 90 min.
Duomenų analizės metodas
Statistinės analizės buvo atliktos su SPSS (v. 16 ir v. 21; SPSS Inc., Chicago IL). Skubus elgesio atsakas į CS buvo įvertintas pagal nosį, kai sacharozė buvo išduota. Po to vidutinis nosies pokesų skaičius per šį intervalą (5 s per bandymą) buvo lyginamas su vidutiniu nosies pokes skaičiumi tuo pačiu laikotarpiu (5 s), vidutiniškai per visą CS buvimo laiką. Grupė × Sėdynės ANOVAs, kuriose yra CS ir yra buvusios ir nėra stebimos, yra diskriminacinio atsako į cue ir neidentiškus nosies kiškio atsakymus įsigijimas pagal skirtingus grafikus 15 sacharozės treniruočių metu.
Gydymo poveikis lokomotoriniam atsakui buvo įvertintas grupėje × Seanso ANOVAs, kai pasireiškė be ligos atsiradimo fazė (trys sesijos), priešinsibilizacija 0.5 mg / kg AMPH (vienas seansas) ir penkių sesijų metu 1 mg / kg AMPH jautrinimo režimas, kai tikimasi, kad grupės skirsis, reaguodamos į kartotines AMPH dozes. Grupė × Sesijos ANOVA taip pat įvertino be vaistų lokomotorinį atsaką 30 min. Planuojami palyginimai įvertino 50 grupės ir 0 grupės (vidutinės trukmės kontrolė) ir 100 grupės (be neapibrėžties kontrolės) vidutinio efektyvumo skirtumą. t-testai (Sveiki, 1992 m), naudojant MS klaidos ir df klaidų terminus atitinkamam efektui (ty grupės arba grupės × seanso sąveikai) iš ANOVA (Vynininkas, 1971 m). Polinomų tendencijų analizė išbandė pokyčių profilį sesijų metu.
Norint nustatyti, ar 15 sacharozės treniruočių metu pasireiškiantys atsakymai į CSN buvimą ir nebuvimą prisidėjo prie lokomotorinio atsako į AMPH kitimą, arba dėl AMPH atsako tarpininkaujančių grupių skirtumų, AMPH buvo atlikta tolesnė kovariacijos (ANCOVA) analizė. judesio duomenys, įskaitant bendrą nosies pokesą (sumą 15 sesijoms), kai CS nebuvo kaip kovaria. Reikšmingas kovariato poveikis rodo, kad be narkotikų metodo atsakas sumažino (įtakojo) grupės ar sesijos poveikį. Grupės ar sesijos poveikio reikšmingumo sumažėjimas, esant reikšmingam kovariatui, parodytų, kad požiūris į reagavimą, kuris yra tarpininko (sesijos) poveikis, yra tarpinis. Grupės ar sesijos pasekmių sumažėjimas, nesant reikšmingo bendrų efektų, paprasčiausiai atspindėtų statistinės galios praradimą dėl df perskirstymo iš klaidos termino į kovariaciją ir neturėtų įtakos interpretavimui. grupės ar sesijos poveikis.
rezultatai
Nosis pokes per sacharozės kondicionavimo sesijas
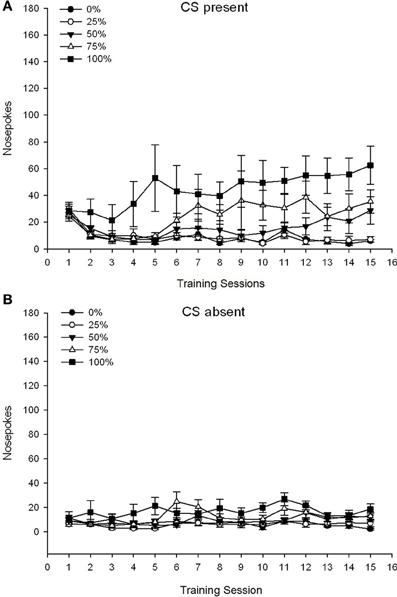
CS yra. Pav 1A 25, 50 ir 100 grupių vidutinis nosies pokštas, kai CS buvo 15 sacharozės kondicionavimo sesijose (nosies pokes nebuvo užregistruotos 0 grupei, kuri negavo CS). 3 grupė × 15 sesija ANOVA davė reikšmingą pagrindinį grupės poveikį, F(2, 21) = 5.63, p = 0.011 ir sesija, F(14, 294) = 14.00, p <0.001 kartu su reikšminga grupės × seanso sąveika, F(28, 294) = 2.93, p <0.001. Pav 1A nurodo, kad pagrindinis sesijos poveikis atspindėjo nosies pokesų padidėjimą per visas tris grupes, o pagrindinis grupės poveikis 100 grupėje apskritai buvo didesnis, o 25 grupėje - 50. Svarbi grupė × sesijos sąveika kubinei tendencijai, F(2, 21) = 4.42, p = 0.030, nurodė greitą pakilimą, kritimą ir išlyginimą nosies pokse per 100 grupės sesijas, lyginant su linijiniu 50 grupės sesijų padidėjimu ir mažesniu linijiniu padidėjimu per sesijas 25 grupėje.
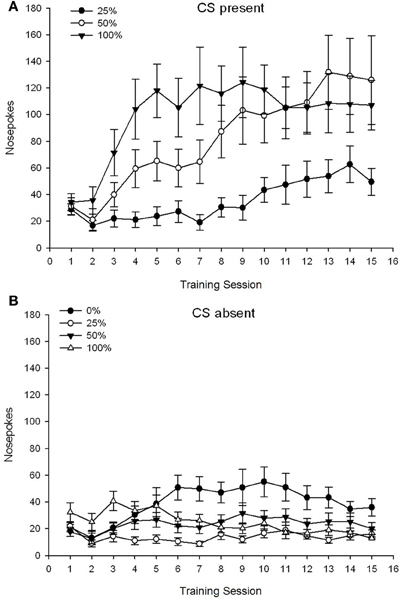
1 pav. Vidutinis (SE) priartėjimo atsakas (nosies kišimas) per 15 sacharozės treniruočių Sprague Dawley žiurkių grupėse (n = 8 / grupė), veikiami sacharozės atlygiu (10% tirpalas), pristatomas pagal kintamąjį 0, 25, 50 arba 100%. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. () Balai, kai buvo CS (5 s × 15 bandymų). (B) Balai, kai CS nebuvo (vidutiniškai 5 × 15 s, kai šviesa buvo išjungta).
CS nėra. Pav 1B rodo vidutinę nosies pokesę visoms keturioms grupėms lygiavertės trukmės laikotarpiu (5 s × 15 tyrimai), vidutiniškai per visą CS buvimo laiką. 4 grupė × 15 sesija ANOVA davė reikšmingą pagrindinį grupės poveikį, F(3, 28) = 7.06, p = 0.001 ir sesija F(14, 392) = 2.84, p <0.001 kartu su reikšminga grupės × seanso sąveika, F(42, 392) = 3.93, p <0.001. Reikšminga grupės × seanso sąveika pagal kvadratinę tendenciją, F(3, 28) = 3.91, p = 0.019, be kubinės tendencijos sąveikos, F(3, 28) <0.93, p > 0.44, atspindėjo „invertuoto U“ profilį, kai nosis verčiasi per 0 grupės sesijas, palyginti su paprastai stabiliu profiliu per kitų grupių sesijas.
Gyvenimo vietos judėjimo kamerose
4 grupė × 3 sesija ANOVA davė pagrindinį sesijos poveikį, F(2, 56) = 5.67, p = 0.006 ir jokio kito reikšmingo poveikio, F(3, 28) <1.60, p > 0.21. Vidutinis (SE) pluošto lūžis per 2 valandas judėjimo variklio dėžėse buvo 1681 (123) pirmojo seanso metu, 1 (1525) per 140 seansą ir 2 (1269) per 96 seansą. Planuoti palyginimai nerado reikšmingų skirtumų tarp 3 grupės ir grupės 50 arba 0 grupė pirmojoje arba paskutinėje pripratimo sesijoje, t(84) <1.69, p > 0.05. Taigi, nesant AMPH, pakartotinis bandymų laukelių poveikis buvo susijęs su nuosekliu savaiminio judėjimo aktyvumo mažėjimu keturiose grupėse (ty seanso efektas) ir jokio skirtingo atsako, priklausančio nuo sacharozės treniruočių grafiko (be sąveikos). .
Bandymų sesijos
Išankstinio jautrinimo poveikis 0.5 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 4 grupės vienpusis judėjimo atsako ANOVA 30-min prieš injekcijos etapą nepadarė reikšmingo poveikio, F(3, 28) <1.05, p > 0.38. Suplanuoti palyginimai nerado reikšmingo skirtumo tarp 50 grupės ir 0 grupės ar 100 grupės, t(32) <0.87, p > 0.40. Todėl pradiniai judėjimo prieš injekciją skirtumai neatsižvelgė į grupių judėjimo reakcijos į AMPH skirtumus. Vidutinis (SE) mėginio pluošto lūžis buvo 559 (77).
Po injekcijos atliktas judėjimas prieš galutinį vaistą be vaistų. 4 grupės × 2 sesija ANOVA palygino grupių judėjimo reakcijas per paskutinę pripratimo sesiją ir iškart po išankstinio sensibilizavimo 0.5 mg / kg AMPH. Įpratimo sesijos (120 min.) Balai buvo keičiami taip, kad atitiktų AMPH testo sesijos trukmę (90 min.) (Neapdoroto pripratimo balas × 90/120). Analizė davė reikšmingą pagrindinį „Session“ poveikį, F(1, 28) = 34.16, p <0.001 ir jokio kito reikšmingo poveikio nėra, F(3, 28) <2.26, p > 0.10. „Session“ poveikis atspindėjo vidutinio (SE) pluošto lūžių padidėjimą, atsižvelgiant į dozę, nuo 952 (72) iki 1859 (151). Suplanuoti palyginimai nerado reikšmingų skirtumų tarp 50 ir 0 grupių arba 100 grupės, atsižvelgiant į dozę, t(56) <1.72, p > 0.10. Tačiau pluošto lūžių balų (M; SE) rangų tvarka sutapo su hipoteze: 50 grupė (2205; 264)> 0 grupė (2025; 203)> 100 grupė (1909; 407)> 25 grupė (1296; 299) .
1 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 4 grupė × 5 seanso ANOVA, kai 30 min. Prieš injekciją pasireiškė 1 mg / kg AMPH bandymų sesijų metu, buvo atliktas pagrindinis Sesijos poveikis, F(4, 112) = 43.64, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(3, 28) <0.97, p > 0.42. Suplanuoti palyginimai nerado reikšmingo skirtumo tarp 50 grupės ir 0 grupės ar 100 grupės per pirmąjį arba paskutinį bandymo seansą, t(140) <0.84, p > 0.30. Todėl pradiniai judėjimo skirtumai neatsižvelgė į grupių judėjimo reakcijos į AMPH skirtumus. Vidutiniai (SE) spindulių lūžio balai prieš dozę pripratimo fazėje 1–5 sesijose buvo: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
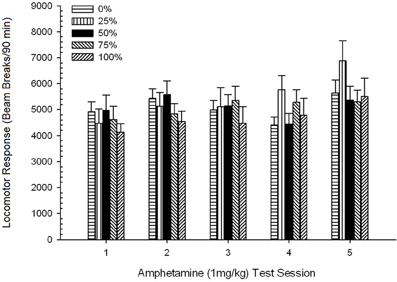
Po injekcijos sukeltas judėjimas. Pav 2 rodo keturių 1 mg / kg AMPH (vieną kartą per savaitę) injekcijų poveikį keturių grupių lokomotorinio aktyvumo rodikliams. 4 grupė × 5 sesija ANOVA davė pagrindinį sesijos poveikį, F(4, 112) = 8.21, p <0.001, ribinis pagrindinis grupės poveikis, F(2, 45) = 3.28, p = 0.085 ir nėra reikšmingos sąveikos, F(12, 122) <0.77, p > 0.68.
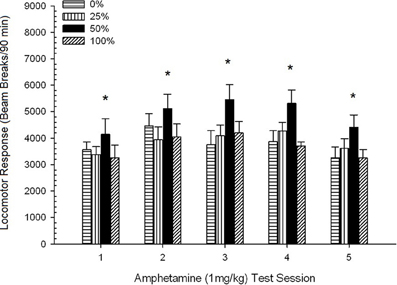
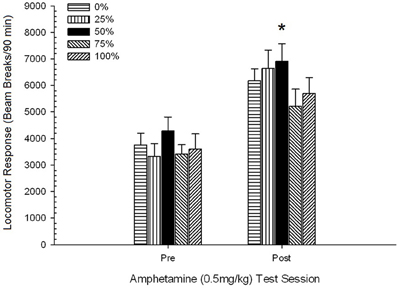
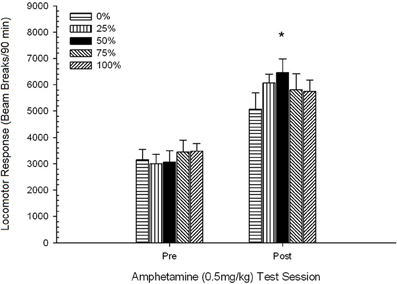
2 paveikslas. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) iki 1 mg/kg d-amfetamino (ip) per 5 savaitinius seansus Sprague Dawley žiurkių grupėse (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. *p < 0.05 vidutiniam skirtumui tarp 50 ir 0 grupės bei 100 grupės, remiantis planuojamais palyginimais.
Planuojami palyginimai parodė, kad 50 grupės rezultatai labai skyrėsi nuo 0 grupės, t(14) = 2.19, p = 0.037 ir grupė 100, t(14) = 2.36, p = 0.025 [ir šiek tiek skyrėsi nuo 25 grupės, t(14) = 2.03, p = 0.051]. Taigi 50 grupėje lokomotorinis atsakas į 1 mg / kg AMPH patikimai viršijo kitų trijų grupių poveikį visose penkiose bandymų sesijose. Polinomo tendencijos analizė nustatė didelę kvadratinę tendenciją per sesijas, F(1, 28) = 32.47, p <0.0001 ir jokių kitų reikšmingų tendencijų nėra, F(1, 28) <1.78, p > 0.19. Pav 2 rodo, kad šis rezultatas atspindi „apverstą U“ modelį per sesijas.
Kontrolė, susijusi su nosies kūgio pokyčiais, atsakant į sacharozės mokymą
Tolesni ANCOVA po lokalizacijos reakcijų į 1 mg / kg AMPH, kai nosies žarnos (CS), kaip kovariacija, trijose grupėse, kurios gavo CS, davė ribinį pagrindinį grupės poveikį, F(2, 20) = 3.07, p = 0.069, ir nėra reikšmingų kovariacijų, F(4, 80) <0.05, p > 0.85. Taigi pasirinktas metodas, atsakantis į treniruotes, nepaaiškino reikšmingų judėjimo reakcijos į 1 mg / kg AMPH skirtumų 25, 50 ar 100 grupėse.
Tolesnis ANCOVA dėl lokomotorinio atsako į 1 mg / kg AMPH, kai nosies užpakalis (CS nėra) kaip kovariacija, davė reikšmingą kovariato poveikį, F(1, 27) = 6.17, p = 0.020, reikšmingas pagrindinis grupės poveikis F(3, 27) = 4.13, p = 0.016, ribinė sesija × Kovariatyvi sąveika, p = 0.080 ir jokio kito reikšmingo poveikio, F(4, 108) <1.48, p > 0.21. Taigi, nenurodytas (neatskiriamas) metodas, reaguojantis treniruotės metu, paaiškino reikšmingą judėjimo reakcijos į 1 mg / kg AMPH skirtumus. Tačiau šis variantas nesutapo su grupe susijusio dispersijos, nes kovariato įtraukimas į analizę padidino, o ne sumažino grupės efekto reikšmingumą.
Diskusija
Duomenys apie nosį, kai buvo buvę CS, rodo, kad grupės įgijo ryšį tarp CS ir sacharozės tiekimo, kaip rodo padidėjęs atsakas į mokymus per mokymus. Atsakymo į sesijas metu, kai buvo buvęs CS, profilis parodė, kad 100 ir 50% CS-US tvarkaraščiai buvo vienodai veiksmingi skatinant požiūrį, o 25% grafikas paskatino kuklesnį didėjimo koeficientą. Duomenys apie nosį, kai CS nebuvo, rodo, kad grupės, gavusios bet kurį iš trijų CS-sacharozės treniruočių tvarkaraščių (grupė 25, 50, 100), greitai išmoko mažinti savo nosį, kai nėra CS, o 0 grupės gyvūnai , kurie negavo CS, išmoko tik ribotai sumažinti savo požiūrį į elgesį po išsamių mokymų.
Pritaikymo duomenys rodo, kad grupės nesiskyrė prieš AMPH ir kad pakartotinis ekspozicija su bandymų dėžėmis buvo susijęs su sumažėjusiu vaistu nesuderinamu lokomotoriniu atsaku. Todėl skirtumai tarp grupių ir padidėjęs atsakas į pakartotines AMPH dozes negali būti siejami su ankstesniais lokomotorinio elgesio skirtumais.
Išankstinio jautrinimo 0.5 mg / kg AMPH tyrimo rezultatai patvirtino, kad vaistas padidino lokomotorinį aktyvumą, palyginti su galutiniu vaistu nesuteikiantį pripratimo dieną. Atsižvelgiant į hipotezę, 50 grupė buvo didesnė už 0 arba 100 (taip pat 25 grupę), atsižvelgiant į vidutinį atsaką į dozę, nors vidutiniai skirtumai tarp grupių nebuvo reikšmingi.
Sensibilizacijos seansų metu planuoti palyginimai tarp grupių parodė, kad ankstesnis 50% sąlyginės sacharozės atlygio poveikis reikšmingai padidino judėjimo reakciją į 1.0 mg / kg amfetamino dozę, palyginti su kitomis trimis schemomis. Šis poveikis buvo akivaizdus nuo pirmosios dozės ir, vartojant kartotines dozes, pastebimai nepakito. Tendencijų analizė parodė dvifazį (viso mėginio) atsaką į pakartotines AMPH dozes, didėjant iki trečios dozės ir vėliau mažėjant. ANKVA stebėjimo su nosies išspaudimais (CS nėra), nes kovariatas, rezultatai patvirtino, kad keturių grupių judėjimo reakcijos į 1 mg / kg AMPH skirtumai nebuvo tarpininkaujami be atsako, atsakant per sacharozės treniruotes.
Grupės poveikis jautrinimo sesijų metu atitinka mūsų hipotezę. Dviejų fazių seanso efektas neatitinka numatomo tolesnio lokomotorinio atsako didėjimo su kartotinėmis AMPH dozėmis. Tai gali būti susiję su dozavimo intervalu. Siekiant išspręsti šią problemą, reikia taikyti procedūrą (pakaitinę paros dozę), kuri parodė, kad sukelia pastovią lokomotorinio atsako didėjimą 1.0 mg / kg AMPH dozėms (ty elgsenos jautrinimas). AMPH jautrinimo režimo poveikis tolesnei reakcijai į antrąjį 0.5 mg / kg poveikį dar labiau sustiprintų šio poveikio bendrumą. Įterpiant fiziologinį tirpalą prieš AMPH, būtų nustatytas tikėtinas ar su injekcijomis susijęs (pvz., Streso) poveikis lokomotoriniam atsakui į AMPH. 75% sąlygojamos sacharozės grupės įtraukimas padėtų išsiaiškinti atlygio neapibrėžtumo ir atlygio retumo įtaką 50 ir 25 grupių atsakymų modeliui. Be to, norint, kad ANCOVA įvertintų vaistų be vaistų metodo reakciją į judėjimą pagal AMPH (naudojant nosies pokes su CS, kaip kovariato), nosies pokesas taip pat buvo koduotas 0 grupei per intervalą, kai CS buvo buvusi kitose keturiose grupėse (ty taip, kad iš visų penkių grupių - įskaitant 0 grupę, kuri negavo jokio CS) - gali būti įtraukta į kovariacijos su CS, kaip kovariacijos, analizę. Šie patobulinimai buvo įtraukti į eksperimentą 2.
Eksperimentuokite 2
Medžiagos ir metodai
2 eksperimento metodika buvo panaši į 1 eksperimento metodiką, tačiau buvo peržiūrėta, kad geriau atitiktų režimą, kuris patikimai sukelia AMPH jautrinimą (Fletcher ir kt., 2005). Pakeitimai buvo tokie: a) 75% CS-sacharozės grupė (n = 8); (b) Sacharozės mokymo metu žiurkės (išskyrus 0 grupę) gavo 20 CS (šviesos) prezentacijas (priešingai nei 15 eksperimente 1); (c) kiekvienas CS pateikimas buvo atskirtas vidutiniu 90 s intervalo tarpiniu intervalu; diapazonas: 30 – 180 s (palyginti su 120 s eksperimente 1), kuris kompensuoja mokymų bandymų padidėjimą, kad kiekvienos treniruotės trukmė atitiktų eksperimento 1 trukmę; (d) kiekvienos iš trijų bendrinimo sesijų trukmė buvo sumažinta nuo 120 iki 90 min, kad atitiktų testavimo sesijų trukmę; (e) Buvo pridėta fiziologinio tirpalo (ip, 1 ml / kg) (90 min.) (po sacharozės treniruočių dienos 8), siekiant įvertinti injekcijos poveikio judesiui poveikį. Rep (pvz., lūkesčiai, stresas); f) 1 mg / kg jautrinimo sesijos vyko pakaitinėmis darbo dienomis (po 12 – 21 treniruočių dienos), o ne kas savaitę, kaip ir eksperimente 1; (g) Kartu su 0.5 mg / kg AMPH poveikiu (po 9 treniruotės) buvo įtrauktas antrasis jautrinimas 0.5 mg / kg AMPH (po sacharozės treniruočių dieną 28), kad būtų galima išbandyti bendrą gydymo 0 dozę. jautrinimo poveikis dozėmis; (h) visose grupėse (įskaitant 5 grupę) buvo koduojami nosies užsikimšimai, kai CS buvo; (i) nosies pokes, kai CS nebuvo, buvo įrašytos specialiai iš XNUMX intervalo prieš pat CS pradžią, kad būtų galima indeksuoti priešlaikinį požiūrį.
rezultatai
Nosis pokes per sacharozės kondicionavimo sesijas
5 grupė × 15 sesija × 2 fazė (CS, CS nėra) ANOVA nosies pokes davė reikšmingą pagrindinį grupės poveikį. F(4, 19) = 2.89, p = 0.050, sesija F(14, 266) = 2.28, p = 0.006 ir fazė, F(1, 19) = 14.72, p = 0.001, taip pat didelė trijų krypčių sąveika, F(56, 266) = 1.38, p = 0.050. Skydai (A, B) pav 3 Nubraižykite grupių vidutinius nosies smūgio balus atitinkamai CS esamų ir CS nesančių fazių atžvilgiu. Palyginus dvi plokštes, paaiškėjo, kad pagrindinis fazės poveikis atspindėjo daugiau bendro atsako į nosį, kai CS buvo, o jo nebuvo. Todėl atsakymai į atsakymus pasitaikydavo žymiai dažniau nei priešlaikiniai atsakymai. Pagrindiniai grupės ir sesijos padariniai nebuvo lengvai interpretuojami dėl aukštesnės eilės sąveikos. Pastarasis rezultatas atspindėjo penkių grupių balų konvergenciją santykinai stabiliu žemu lygiu per sesijas, kai nebuvo CS (pav.) 3B), kartu su balų skirtumu į aukštą (75 grupė, 100 grupė), tarpinį (50 grupė) ir mažą (0 grupės, 25 grupės) lygį, kai atsakas per sesijas, kai CS buvo (žr. 3A). Iš žemesnės eilės polinomų tendencijų (tiesinė, kvadratinė, kubinė) tik reikšminga linijinės tendencijos trijų krypčių sąveika, F(4, 19) = 2.32, p = 0.094, atspindintis apskritai monotonišką nosies pokesų padidėjimą per 75 grupės sesijas ir santykinai spartesnę stabilizaciją esant aukštai, vidutinei ir žemai reaguojant kitose grupėse, kai CS buvo.
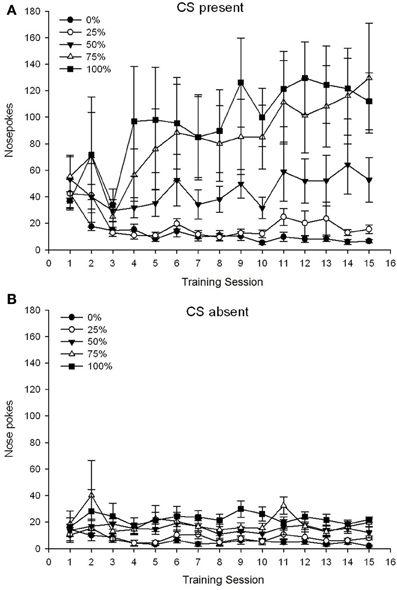
3 pav. Vidutinis (SE) priartėjimo atsakas (nosies kišimas) per 15 sacharozės treniruočių Sprague Dawley žiurkių grupėse (n 8 / 10, 0, 25 arba 50% kintamųjų tvarkaraščių, veikiamų sacharozės atlyginimu (75% tirpalu).. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. () Balai, kai buvo CS (5 s × 20 bandymų). (B) Balai, kai CS nebuvo (vidutiniškai 5 × 20 s, kai šviesa buvo išjungta).
Vietos judėjimo dėžės
5 grupė × 3 sesija ANOVA be vaistų lokomotorinio atsako davė reikšmingą pagrindinį Sesijos poveikį. F(2, 70) = 60.01, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <0.70, p > 0.60. Planuoti 50 grupės palyginimai su 0 grupe ir 100 grupėmis pirmojo ir paskutinio įpratimo sesijose reikšmingo poveikio nedavė, tyra <0.84, p > 0.40. Todėl prieš bandymą vidutinis lokomotorinis atsakas be narkotikų pagrindinėse grupėse nesiskyrė. Vidutinis (SE) pluošto pertraukų skaičius per 90 min. Buvo 2162 (118) 1 sesijoje, 1470 (116) 2 sesijoje ir 1250 (98) 3 sesijoje.
Bandymų sesijos
Druskingas. 5 grupė × 2 sesija ANOVA palygino lokomotorinį atsaką į galutinį įpročio seansą ir fiziologinio tirpalo užduotį. ANOVA davė pagrindinį Sesijos poveikį, F(1, 35) = 62.46, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <0.65, p > 0.64. Pav 4 parodo grupės priemones ir parodo, kad sesijos efektas atspindi bendrą judesio atsako sumažėjimą nuo galutinio vaistų be pripratimo sesijos iki fiziologinio tirpalo sesijos, kuri nesiskyrė pagal grupę. Taigi, tęstinis lokomotorinio atsako sumažėjimas per tris įpročius buvo tęsiamas ketvirtoje be narkotikų poveikio bandymų dėžėse.
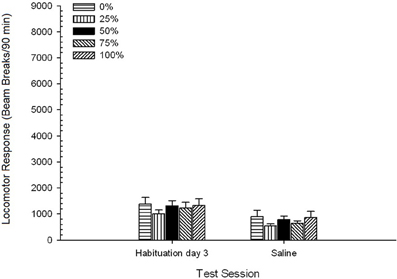
4 pav. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) per paskutinius 3 pripratimo be vaistų seansus ir kitą seansą po fiziologinio tirpalo injekcijos (ip, 1 ml/kg) grupėse Sprague Dawley žiurkių (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų.
0.5 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 5 grupė × 2 sekcijos ANOVA prieš injekciją (30-min) prieš ir po jautrinimo 0.5 mg / kg AMPH bandymo dienos davė reikšmingą pagrindinį Sesijos poveikį, F(1, 35) = 13.39, p = 0.001 ir jokio kito reikšmingo poveikio, F(4, 35) <1.79, p > 0.15. Suplanuoti palyginimai nerado reikšmingų skirtumų tarp 50 grupės ir 0 grupės ar 100 grupės pirmojo seanso metu, t(70) <1.00, p > 0.30. Tačiau antrojo (po sensibilizavimo) seanso metu 50 grupėje (1203; 121) buvo žymiai daugiau pluoštų pertraukų prieš injekciją (M; SE) nei 100 grupėje (756; 103), t(70) = 5.11, p <0.001, bet nesiskyrė nuo 0 grupės (1126; 211), t(7) <0.88, p > 0.40. Todėl pradiniai judesio skirtumai neatsižvelgė į judėjimo reakcijos į pirmąją 0.5 mg / kg AMPH dozę grupių skirtumus, tačiau galėjo prisidėti prie 50 ir 100 grupės judėjimo atsako į antrąją 0.5 mg / kg AMPH dozę skirtumų. . Vidutinis (SE) pluošto lūžis prieš injekciją fazės metu per pirmąjį ir antrąjį 0.5 mg / kg AMPH testo seansus buvo 757 (41) ir 974 (59).
Po injekcijos sukeltas judėjimas. 5 grupė × 2 seanso ANOVA su 0.5 mg / kg AMPH lokomotoriniu atsaku prieš ir po 5 dozės jautrinančio režimo davė pagrindinį Sesijos poveikį, F(1, 35) = 76.05, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <1.10, p > 0.37. Pav 5 rodo kiekvienos grupės ir sesijos vidurkius.
5 pav. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) iki 0.5 mg/kg d-amfetamino atskiruose seansuose prieš ir po 5 seansų jautrinimo d-amfetamino (1.0 mg/ kg; ip per seansą) Sprague Dawley žiurkių grupėse (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. *p < 0.05 vidutiniam skirtumui tarp 50 ir 0 grupės bei 100 grupės, remiantis planuojamais palyginimais.
Paveiksle matyti, kad Sesijos efektas reikšmingai padidino vidutines vidutines (SE) spindulių pertraukas 90 min. 0.5 mg / kg 1, 3674 (216) dozės 0.5, 2 (6123). Sąveikos ar grupės poveikio nebuvimas parodė, kad jautrinimas AMPH nesiskiria patikimai skirtingose grupėse. Nepaisant to, kad ANOVA neturėjo reikšmingo grupinio poveikio, skaičiavimas parodo, kad 275 grupė rodė didžiausią atsaką tiek į pirmąją, tiek į antrąją 50 mg / kg dozę. Numatytas atsako į pirmąją 0.5 mg / kg dozę palyginimas neatskleidė reikšmingo skirtumo tarp 0.5 grupės ir 50 grupės arba 0 grupės, t's(35) <0.48, p > 0.50. Tačiau, reaguojant į antrąją (po sensibilizavimo) 0.5 mg / kg dozę, 50 grupės judėjimas buvo žymiai didesnis nei 0 grupės, t(35) = 2.00, p <0.05, taip pat 100 grupė, t(35) = 3.29, p <0.01.
Atsižvelgiant į reikšmingą grupių skirtumą prieš pradedant injekciją antroje 0.5 mg / kg AMPH sesijoje, apie kurią buvo pranešta, buvo atlikta tolesnė 5 grupė × 2 sesija ANCOVA, kai lokomotorinis atsakas į 0.5 mg / kg AMPH buvo kontroliuojamas. antrojoje sesijoje. Ši analizė davė didelį poveikį kovariacijai, F(1, 34) = 8.65, p = 0.006, pagrindinis Sesijos poveikis F(1, 34) = 10.83, p = 0.002 ir jokio kito reikšmingo poveikio, F(4, 34) <0.85, p > 0.50. Svarbu tai, kad planuoti palyginimai, pagrįsti MS paklaida ir ANCOVA df paklaida, patvirtino, kad vidutinis judėjimo atsakas į antrąją 0.5 mg / kg AMPH dozę 50 grupėje išliko žymiai didesnis nei 100, t(34) = 3.09, p <0.01 ir 0 grupė, t(34) = 1.88, p <0.05 (viena uodega), kai prieš injekciją buvo kontroliuojami 2 seanso pokyčiai. Taigi 50 grupėje pastebimas žymiai didesnis lokomotorinis atsakas po sensibilizacijos į 0.5 mg / kg AMPH, nei 100 ar 0 grupėje, ir šių grupių skirtumai nebuvo tarpininkaujant judesiui prieš injekciją bandymo dienomis.
1.0 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 5 grupė × 5 30-min. 1 min. Sekos ANNOVA XNUMX mg / kg AMPH jautrinimo sesijose davė pagrindinį Sesijos poveikį, F(4, 140) = 16.70, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <0.94, p > 0.45. Suplanuoti palyginimai nerado reikšmingo skirtumo tarp injekcijos prieš injekciją tarp 50 grupės ir 0 grupės arba 100 grupės pirmojo seanso metu, t(175) <1.66, p > 0.10. Tačiau paskutinėje sesijoje 50 grupėje (1167; 140) buvo žymiai daugiau pluoštų pertraukų (M; SE) nei 100 grupėje (1000; 99), t(175) = 2.35, p <0.05, bet nesiskyrė nuo 0 grupės (1085, 120), t(175) <1.16, p > 0.20. Todėl judėjimo prieš injekciją skirtumai prisidėjo prie 50 ir 100 grupių skirtumų, susijusių su judėjimo reakcija į galutinę 1 mg / kg AMPH dozę. Vidutinis (SE) mėginio pluošto lūžis prieš injekciją 1–5 sesijų metu buvo: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
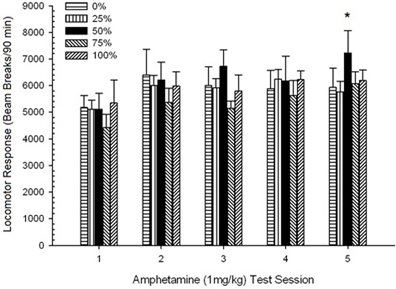
Po injekcijos sukeltas judėjimas. 5 grupė × 5 sesijos ANOVA atsakas į 1 mg / kg AMPH davė reikšmingą pagrindinį Sesijos poveikį, F(4, 140) = 6.72, p <0.001, ribinė grupės × seanso sąveika, F(16, 140) = 1.57, p = 0.085, ir pagrindinis grupės poveikis, F(4, 35) <0.44, p > 0.77. Daugianario tendencijų analizė atskleidė reikšmingą tiesinę tendenciją, F(1, 35) = 9.19, p = 0.005 ir kubinė tendencija, F(1, 35) = 21.63, p <0.001, per 1–5 seansus 6 rodo kiekvienos grupės ir sesijos vidutinius judesio balus.
6 paveikslas. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) iki 1 mg/kg d-amfetamino (ip) per 5 savaitinius seansus Sprague Dawley žiurkių grupėse (n = 8/grupei), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų.
Paveiksle matyti, kad sesijos efektas parodė, kad bendras 1, 4624 (213) ir 5, 5736 (272) seansų bendrasis vidutinis (SE) spindulių pertraukos padidėjo, o tai patvirtina jautrumo AMPH atsiradimą. Kubinė tendencija žymėjo santykines maksimumas sesijose 1, 3, ir 5, su 2 ir 4 sesijomis, ypač 0 ir 50 grupėms. Paveiksle taip pat atskleidžiama, kad, nepaisant didelės sąveikos stokos, 25 grupė palaipsniui parodė didesnį judesio atsaką per sesijas ir labai skyrėsi nuo kitų grupių 4 ir 5 sesijose (9 ir 22% didesnis nei kitoje aukščiausioje grupėje). Planuojami palyginimai parodė, kad 50 grupė žymiai nesiskyrė nuo 0 arba 100 grupių, t(175) <0.89, p > 0.40 per pirmąjį arba paskutinį 1 mg / kg AMPH testo seansą.
Kontrolė, susijusi su nosies kūgio pokyčiais, atsakant į sacharozės mokymą
Dviejų 5 grupių × 2 sesija ANCOVA, kai lokomotorinis atsakas į 0.5 mg / kg AMPH prieš ir po jautrinimo režimo, įskaitant bendrą nosį, sacharozės treniruotės metu dalyvaujant CS, ir CS nėra atskirų kovariacijų, nerado reikšmingo poveikio nei kovariatui, nei F(1, 18) <1.03, p > 0.31. Todėl požiūris, atsakant į treniruotes, netiesiogiai sąlygojo grupių skirtumus atsakant į 0.5 mg / kg AMPH.
Dviejų 5 grupių × 5 sesija ANCOVA, kai lokomotorinis atsakas į 1 mg / kg per jautrinimo sesijas su visais nosies pokaisiais (CS buvimas, CS nėra), nes atskiri koviniai junginiai nesukėlė reikšmingo kovariato poveikio, kai CS buvo, F(4, 104) <1.04, p > 0.38 ir ribinis pagrindinis kovariato poveikis, kol nebuvo CS, F(1, 18) = 3.32, p = 0.085.
Diskusija
Šio tyrimo rezultatai neatitiko hipotezės, kad 50 grupė parodys didesnį lokomotorinį atsaką per sesijas, palyginti su kitomis grupėmis. 1 mg / kg AMPH duomenys patvirtino jautrumo atsiradimą vartojant kitą paros dozę. Grupių modeliai parodė didesnio jautrumo tendenciją pastarųjų sesijų metu 25 grupėje, be tokių įrodymų 50 grupei. Priešingai, 0.5 mg / kg dozės rezultatai parodė, kad 50 grupėje pastebėta didesnio jautrumo tendencija, tuo pačiu metu patvirtinant reikšmingą bendrą lokomotorinio atsako padidėjimą grupėse iki antrosios ir pirmosios 0.5 mg / kg AMPH dozės. Nulinis fiziologinio tirpalo injekcijos poveikis patvirtino, kad tikėtinas ar su injekcijomis susijęs stresas neprisidėjo prie AMPH poveikio.
Nosies kišimo duomenys dar kartą atskleidė, kad per treniruotes, kai buvo KS, bendras požiūris į reagavimą padidėjo, o kai KS nebuvo, atitinkamo padidėjimo nebuvo. Todėl atrodė, kad gyvūnai įgyja ryšį tarp CS ir sacharozės atlygio. Grupės skirtumai tarp nosies dūrių dažnio, kai buvo CS, maždaug atitiko atlygio įteikimo dažnį pagal atitinkamus grafikus: 75 ir 100 grupėse buvo daugiausiai nosies dūrių, 50 grupėje – tarpinis nosies dūrių skaičius, o 0 ir 25 grupėse. 50 rodo mažiausiai nosies dūrių. Šie rezultatai rodo, kad CS pradėjo kontroliuoti požiūrį, reaguodamas taip, kad atitiktų bendrą atlygio tikimybę. Nors ir spekuliatyvus, vienas iš galimų paaiškinimų dėl mažesnio nosies kišimo dažnio, kai KS yra 2 grupėje 1 eksperimente, palyginti su 1 eksperimentu, gali būti intervalo tarp bandymų sutrumpinimas, nes atrodo, kad ilgesni tarp bandymų intervalai (XNUMX eksperimentas) skatina impulsyvumą. tendencijos ir tai yra susijusi su padidėjusia DA apykaita priekinėje cingulinėje, priešlimbinėje ir infralimbinėje žievėje (Dalley ir kt., 2002). Todėl 30% sumažėjimas tarp bandymų 2 (ir 3) tyrimų gali būti pakeistas kortikos DA lygiais ir skatinamas selektyvesnis (ty, atsižvelgiant į santykinį atlyginimų dažnį) ir impulsyvus (nesukeliantis atlygio dažnio) metodas 50 grupėje, reaguodama 2 eksperimento metu, palyginti su eksperimentu 1.
Nėra reikšmingo su kovu susijusio poveikio, susijusio su nosies užsikimšimu CS dabartinėje būsenoje, ANCOVA rodo, kad metodas, reaguojant per sacharozės mokymą, nepadarė skirtingų CS-sacharozės grafikų poveikio atsakams į AMPH. Nežymiai reikšmingas kovariato poveikis CS nesant būklei ANCOVA lokomotorinio atsako į 1 mg / kg AMPH atveju rodo, kad tendencija prieš ankstyvą vaisto nesukeliantį atsaką paaiškino kai kuriuos AMPH lokomotorinio poveikio pokyčius jautrinimo sesijų metu.
Kartu įrodymai rodo, kad gydymo istorijoje poveikis gali būti labiau pastebimas 0.5 AMPH, nei vartojant 1 mg / kg AMPH, ir kad protokolas, kuris sukelia jautrumą, jei nėra jokio kito manipuliavimo, gali užtemdyti arba padaryti nereikalingą poveikį. jautrumo skatinimo elgesio manipuliacija (ty lėtinis kintamasis atlygis).
Elgesio jautrinimas AMPH yra tvirtas poveikis laboratorijoje. Tačiau už laboratorijos ribų tik mažuma chroniškai lošiančių asmenų pakyla iki patologinio lygio. Nors įjautrinimo rizika yra susijusi su priklausomybės (arba narkotikų vartojimo) rizika, ypač psichostimuliatoriams (Vezina, 2004; Flagel et al., 2008), daugelis veiksnių, be įjautrinimo rizikos, gali sukelti priklausomybę (pvz., Verdejo-Garcia ir kt., 2008; Conversano ir kt., 2012 m; Volkow ir kt., 2012). Nepaisant to, bruožų veiksniai, suteikiantys jautrumą jautrinimui, gali sąveikauti su kondicionavimo istorija, siekiant pabrėžti nenuspėjamo atlygio (ty 50% CS-US tvarkaraščio) poveikį DA sistemos reaktyvumui. Norėdami ištirti šią galimybę, eksperimentas 3 taikė tą pačią procedūrą kaip ir eksperimentas 2, bet naudojo Lewis padermę vietoj Sprague Dawley padermės žiurkių.
Sprague Dawley žiurkėms būdingas vidutinis DA transporterių lygis, mažesnis nei Wistar padermės žiurkėms (Zamudio ir kt., 2005 m), bet didesnis nei Wistar-Kyoto žiurkių (į „depresiją“ panašus padermės) lygis nucleus accumbens, migdoliniame kūne, ventralinėje krūtinės srityje ir juodojoje juodojoje medžiagoje.Jiao ir kt., 2003 m). Dėl šio profilio Sprague Dawley žiurkės gali būti tik vidutiniškai jautrios aplinkos ar farmakologinėms DA funkcijos manipuliacijoms. Priešingai, Lewis žiurkėms būdingas mažas DA transporterių, taip pat D2 ir D3 DA receptorių kiekis branduolyje ir nugaros juostoje, palyginti su kitomis padermėmis (pvz., F344).Flores ir kt., 1998). Šie morfologiniai skirtumai gali prisidėti prie skirtingo Lewiso žiurkių atsako į DA manipuliacijas. Lewiso žiurkės taip pat pasižymi įvairiais paryškintais atsakais į eksperimentines manipuliacijas vaistais, palyginti su kitomis padermėmis (pvz., F344). Svarbiausia, kad Lewiso žiurkės yra labiau jautrios metamfetaminui, kurioms būdingas mažas atsakas į pradines dozes, bet didesnis atsakas į vėlesnes dozes.Camp ir kt., 1994 m). Lewis žiurkės taip pat pasižymi didesniu lokomotoriniu jautrumu įvairioms kokaino dozėms.Kosten ir kt., 1994; Haile ir kt., 2001). Remiantis šiuo poveikiu, mes manėme, kad Lewis žiurkės leis mums ištirti, ar jautrumas jautrumui sustiprina kondicionavimo grafiko poveikį tolesniam atsakui į AMPH.
Eksperimentuokite 3
Medžiagos ir metodai
Metodika buvo tokia pati, kaip ir eksperimente 2, be Lewis žiurkių naudojimo (200 – 225 g atvykus, Charles River, Quebec, Kanada).
rezultatai
Nosis pokes per sacharozės kondicionavimo sesijas
5 grupė × 15 sesija × 2 fazė (CS, CS nėra) ANOVA nosies pokes davė reikšmingą pagrindinį grupės poveikį. F(4, 34) = 6.12, p = 0.001, sesija, F(14, 476) = 3.42, p <0.001 ir fazė, F(1, 34) = 20.83, p <0.001, taip pat reikšminga trijų krypčių sąveika, F(56, 476) = 1.56, p = 0.008. Skydai (A, B) pav 7 Nubraižykite grupių vidutinius nosies smūgio balus atitinkamai CS esamų ir CS nesančių fazių atžvilgiu. Palyginus dvi plokštes, paaiškėjo, kad pagrindinis fazės poveikis atspindėjo daugiau bendro atsako į nosį, kai CS buvo, o jo nebuvo. Todėl atsakymai užfiksuoti žymiai dažniau nei anksčiau subrendę atsakymai. Pagrindiniai grupės ir sesijos padariniai nebuvo lengvai interpretuojami dėl aukštesnės eilės sąveikos. Trijų krypčių sąveika atspindėjo penkių grupių balų konvergenciją santykinai stabiliu žemu lygiu per sesijas, kai nebuvo CS (B)], kartu su balų skirtumu, kai CS buvo įtrauktas į santykinai diskretiškus profilius kiekvienai grupei, lygiagrečiai jų reitingų eilės tvarka: nuo aukščiausio (100 grupė) iki mažiausios (grupė 25) [skydas ()]. Tik tiesinė sąveikos tendencija buvo reikšminga, F(4, 34) = 4.03, p = 0.009, atspindintis bendrą nuoseklų nosies pokesų padidėjimą per 100 grupės sesijas, kai CS dalyvavo, palyginti su santykinai nenuosekliu nosies pokesio profiliu kitose grupėse per šį etapą.
7 pav. Vidutinis (SE) priartėjimo atsakas (nosies kišimas) per 15 sacharozės treniruočių Lewiso žiurkių grupėse (n 8 / 10, 0, 25 arba 50% kintamųjų tvarkaraščių, veikiamų sacharozės atlyginimu (75% tirpalu).. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. () Balai, kai buvo CS (5 s × 20 bandymų). (B) Balai, kai CS nebuvo (vidutiniškai 5 × 20 s, kai šviesa buvo išjungta).
Vietos judėjimo dėžės
5 grupė × 3 sesija ANOVA davė pagrindinį sesijos poveikį, F(2, 70) = 23.07, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(8, 70) <1.47, p > 0.18. Kreivinis vidutinio (SE) judesio judesio balų modelis paaiškėjo nuo 1 sesijos 1076 (74) iki 2 sesijos, 644 (48), iki 3 sesijos, 762 (59). Suplanuoti 50 grupės palyginimai su 0 ir 100 grupėmis pirmojo ir paskutinio pripratimo sesijų metu atskleidė žymiai mažiau spindulių pertraukų 50 grupėje (M = 911; SE = 109) prieš grupę 0 (M = 1103; SE = 176) įpratimo sesijoje 1, t(105) = 2.02, p <0.05, bet nėra skirtumo tarp 50 ir 100 grupių (M = 1066; SE = 150), t(105) <1.20, p > 0.20, šioje sesijoje. 50 grupė paskutinėje pripratimo sesijoje reikšmingai nesiskyrė nei nuo 0, nei nuo 100 grupės, t(105) <0.93, p > 0.30. Todėl vidutinis lokomotorinis atsakas be narkotikų pagrindinėse grupėse prieš bandymą nuosekliai nesiskyrė.
Bandymų sesijos
Druskingas. 5 grupė × 2 seanso ANOVA galutinio įpročio sesijos metu ir fiziologinio tirpalo sesija davė reikšmingą pagrindinį Sesijos poveikį. F(1, 35) = 50.12, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <0.57, p > 0.68. Pav 8 parodo abiejų seansų grupės vidurkius ir nurodo, kad sesijos efektas atspindi reikšmingą sumažėjimą nuo pripratimo prie fiziologinio tirpalo. Taigi, injekcijos gavimas Rep (pvz., tikimybė, stresas) nepadidino lokomotorinio atsako.
8 pav. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) per paskutinius 3 pripratimo be vaistų seansus ir kitą seansą po fiziologinio tirpalo injekcijos (ip, 1 ml/kg) grupėse Lewiso žiurkių (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų.
0.5 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 5 grupė × 2 seanso anovacija ANOVA davė reikšmingą pagrindinį Sesijos poveikį. F(1, 35) = 15.04, p <0.001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <1.19, p > 0.33. Suplanuoti palyginimai nenustatė reikšmingo skirtumo tarp 50 ir 0 grupių arba 100 grupės per bet kurį bandymo seansą, t(70) <0.99, p > 0.30. Todėl pradiniai judėjimo prieš injekciją skirtumai neatsižvelgė į grupių judėjimo reakcijos į 0.5 mg / kg AMPH skirtumus. Pirmojo ir antrojo (po sensibilizavimo) 0.5 mg / kg seansų vidutinis (SE) pluošto lūžis prieš injekciją buvo 325 (25) ir 473 (36).
Po injekcijos sukeltas judėjimas. 5 grupė × 2 sesija ANOVA, kai lokomotorinis atsakas į 0.5 mg / kg dozes, tiekiamas prieš ir po lėtinio 1 mg / kg AMPH, davė pagrindinį Session poveikį. F(1, 34) = 87.44, p <0.0001 ir jokio kito reikšmingo poveikio nėra, F(4, 34) <0.94, p > 0.45. Pav 9 parodo kiekvienos grupės ir sesijos vidutinius judesio balus ir parodo, kad sesijos efektas atspindi padidėjusį bendrą atsaką į antrąją 0.5 mg / kg dozę, atitinkančią jautrumą. Paveiksle taip pat matyti, kad grupės 1 sesijos buvo labai panašios, tačiau 50 grupė rodė daugiau lokomotorinio aktyvumo nei kitos grupės 2 sesijoje. Numatyta, kad, palyginus su pirmąja 0.5 mg / kg doze, nebuvo nustatyta reikšmingų skirtumų tarp 50 ir 0 grupės arba 100 grupės, t(35) <1.28, p > 0.20. Tačiau 50 grupės judėjimo reakcija į antrąją 0.5 mg / kg dozę buvo žymiai didesnė nei 0 grupėje, t(35) = 4.32, p <0.001 arba 100 grupė, t(35) = 2.24, p <0.05.
9 pav. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) iki 0.5 mg/kg d-amfetamino atskiruose seansuose prieš ir po 5 seansų jautrinimo d-amfetamino (1.0 mg/ kg; ip per seansą) Lewiso žiurkių grupėse (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. *p < 0.05 vidutiniam skirtumui tarp 50 ir 0 grupės bei 100 grupės, remiantis planuojamais palyginimais.
1 mg / kg AMPH poveikis.
Judėjimas prieš injekciją. 5 grupė × 5 30 min min. Priešpurškimo sekų ANOVA seka jautrinimo seansams davė pagrindinį Sesijos poveikį, F(4, 140) = 4.10, p = 0.004 ir jokio kito reikšmingo poveikio, F(4, 35) = 1.25, p > 0.31. Suplanuoti palyginimai parodė, kad pluošto lūžiai prieš injekciją (M; SE) buvo žymiai mažesni 50 grupėje (395; 62) nei 100 grupėje (508; 62), t(175) = 2.58, p <0.01, bet ne 0 grupė, t(175) <1.83, p > 0.10, atliekant 1 mg / kg AMPH sesiją. Paskutinėje 1 mg / kg AMPH sesijoje planiniai palyginimai taip pat parodė, kad 1 grupės (50; 378) judėjimas prieš injekciją buvo žymiai mažesnis nei 60 grupės (100; 650) ), t(175) = 6.17, p <0.001, bet ne 0 grupėje, t(175) <1.84, p > 0.10. Kadangi šių grupių skirtumų kryptis (kontrolinė grupė = 50 grupė) buvo priešinga hipotezuotam modeliui, grupiniai judėjimo po injekcijos skirtumai, derinantys su hipoteze, negali būti priskiriami prie pradinių injekcijos skirtumų. Vidutinis (SE) bendras pluošto lūžis prieš injekciją 1–5 seansų metu buvo: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Po injekcijos sukeltas judėjimas. 5 grupė × 5 sesijos ANOVA atsako į 1 mg / kg dozes davė reikšmingą pagrindinį Sesijos poveikį, F(4, 140) = 6.15, p <0.001 ir jokio kito reikšmingo poveikio nėra, F(4, 35) <0.57, p > 0.68. Daugianario tendencijų analizė atskleidė reikšmingą tiesinę tendenciją, F(1, 35) = 9.34, p = 0.004 ir kubinė tendencija, F(1, 35) = 5.08, p = 0.031, pastarasis rezultatas reiškia santykines maksimalias seansų 3 ir 5 reikšmes. Pav 10 parodo šiuos balus ir parodo, kad, nepaisant reikšmingos sąveikos su ANOVA, 50 grupė, reaguodama į galutinę 1 mg / kg dozę, parodė žymiai didesnį judėjimą nei kitos keturios grupės. Atitinkamai, planuojami palyginimai parodė, kad 5 grupėje 50 grupėje buvo daug didesnių vidurkių nei visose kitose grupėse, t(35) > 3.68, p <0.001.
10 paveikslas. Vidutinis (SE) lokomotorinis atsakas (spindulio pertrūkių skaičius elektroninėje matricoje per 90 min.) iki 1 mg/kg d-amfetamino (ip) per 5 savaites Lewis žiurkių grupėse (n = 8 grupėje), kurie anksčiau buvo paveikti 15 kasdienių kondicionavimo seansų su sacharozės atlygio (10 % tirpalo) 0, 25, 50, 75 arba 100 % kintamu grafiku.. Sąlyginis dirgiklis buvo šviesa (120 s). 0 grupė gavo tiek pat apdovanojimų kaip ir 100 grupė, nesant sąlyginių stimulų. *p < 0.05 vidutiniam skirtumui tarp 50 ir 0 grupės bei 100 grupės, remiantis planuojamais palyginimais.
Kontrolė, susijusi su nosies kūgio pokyčiais, atsakant į sacharozės mokymą
Dviejų 5 grupių × 2 sesija ANCOVA, kai lokomotorinis atsakas į 0.5 mg / kg AMPH prieš ir po jautrinimo režimo, įskaitant bendrą nosį, sacharozės treniruotės metu dalyvaujant CS, ir CS nėra atskirų kovariacijų, nerado reikšmingo poveikio nei kovariatui, nei F(1, 32) <0.44 p > 0.51. Dvi 5 grupės × 5 seanso judėjimo reakcijos į 1 mg / kg AMPH judesio jautrumo sesijų metu su viso nosies išspaudimu (CS esama, CS nėra), nes atskiri kovariatai reikšmingo kovariato poveikio nesukėlė, kol CS buvo arba jo nebuvo, F(1, 33) <0.14, p > 0.71. Todėl taikant metodą be narkotikų nebuvo atsižvelgta į grupių judėjimo reakcijos skirtumus į bet kurią AMPH dozę.
Diskusija
Jautrinimas išsivystė pakartotinio 1.0 mg/kg amfetamino poveikiui. Pripratimo ir fiziologinio tirpalo duomenys patvirtina, kad šį poveikį lėmė ne anksčiau buvę skirtumai, lūkesčiai ar su stresu susijusios reakcijos į injekciją. ANCOVA su nosies kišimu patvirtina, kad toks poveikis atsirado ne dėl elgesio be narkotikų. Patys nosies kišimo duomenys parodė, kad grupės įgijo ryšį tarp CS ir sacharozės atlygio. Grupių atsako į nosies kišimą lygis treniruotės pabaigoje atitiko bendrą atlygio dažnį pagal skirtingus grafikus nuo didžiausio (100 grupė) iki mažiausio (0 grupė), kaip ir 2 eksperimente. Santykinai žemesnis bendras vidurkis Nosies kišimo lygis šiame eksperimente, palyginti su 1 ir 2 eksperimentais, gali atspindėti selektyvesnį požiūrį, reaguojant į užuominas už atlygį Lewis žiurkėms (Kosten ir kt., 2007).
0.5 mg / kg dozės duomenys parodė, kad Lewis žiurkių pradinis lokomotorinis atsakas į AMPH 9) buvo šiek tiek slopinta, palyginti su Sprague Dawley žiurkėmis (eksperimentas 2; 5), tačiau po 0.5 sesijos AMPH režimo atsakas į antrąją Lewis žiurkių dozę padidėjo daug (beveik dvigubai daugiau nei pirmoji 5 mg / kg dozė). Labiausiai, 50 grupėje pasireiškė didesnis lokomotorinis atsakas išskyrus visas grupes, išskyrus 25 grupę į antrą (ty po jautrinimo) 0.5 mg / kg AMPH dozę ir didesnį lokomotorinį atsaką nei visos kitos grupės, įskaitant 25 grupę, į galutinę 1 mg / kg AMPH dozę (galutinė jautrinimo sesija) .
Eksperimentų grupės reitingų suvestinė
Norint nustatyti jautrumo grupių skirtumų patikimumą, neparametrine analize buvo įvertintas atsitiktinumas tarp grupės ir vidutinio lokomotorinio atsako į antrąją (po lėtinio AMPH) 0.5 mg/kg dozę ir galutinę 1.0 mg/kg AMPH dozę. iš 3 eksperimentų. Analizė davė reikšmingą poveikį, φ = 0.986, p = 0.025, atspindintis faktą, kad 50 grupė pirmą kartą užėmė visus palyginimus. Geresnis 50 grupės lygis, palyginti su visomis kitomis grupėmis, reaguojant į antrąją (po chroniško AMPH) 0.5 mg / kg dozę, pavaizduotas Fig. 5 (eksperimentas 2) ir paveikslas 9 (eksperimentas 3). Geresnis 50 grupės lygis, palyginti su kitomis grupėmis, atsižvelgiant į galutinę 1.0 mg / kg dozę, pavaizduotas Fig. 2 (eksperimentas 1) ir paveikslas 10 (eksperimentas 3). Vienintelė šio modelio išimtis buvo atsakas į galutinę 1.0 mg / kg dozę Sprague-Dawley žiurkėms 2 eksperimento metu.
Bendra diskusija
Šioje eksperimentų serijoje buvo patikrinta hipotezė, kad nuolatinis į azartinius lošimus panašus atlygio grafikas gali įjautrinti smegenų DA kelius, panašiai kaip lėtinis piktnaudžiavimo narkotikais poveikis. Tokio poveikio įrodymai rodo, kad neuroplastiškumą, kuris, kaip manoma, prisideda prie priklausomybės nuo narkotikų, gali sukelti nuolatinis nenuspėjamas atlygio grafikas. Remiantis literatūra apie priklausomybę nuo narkotikų, lokomotorinis atsakas į 0.5 ir 1.0 mg/kg AMPH dozes indeksuoja DA sistemos reaktyvumą, o didesnis judėjimas reaguojant į vėlesnes dozes, kurios operatyviai apibrėžia jautrumą (plg. Robinson ir Berridge, 1993; Pierce ir Kalivas, 1997; Vanderschuren ir Kalivas, 2000).
Apskritai rezultatai atitinka mūsų hipotezę. Tačiau jie taip pat rodo, kad dėl procedūrinių veiksnių eksperimentinis poveikis labai skiriasi. Gydymo grafiko poveikis buvo nedidelis, tačiau nuoseklus, o 50 grupėje po penkių dozavimo režimų abiejų dozių atsakas buvo didesnis nei kitoms keturioms grupėms. Nors apskritai F- su grupe susijusio poveikio vertės dispersijos analizėse dažnai nebuvo reikšmingos, pagrindiniai grupių skirtumai buvo patvirtinti pagal suplanuotus palyginimus. Šiuo atžvilgiu reikėtų pažymėti, kad „dabartinis mąstymas yra tai, kad bendra reikšmė yra [ F ANOVA] nėra būtina. Visų pirma, bendrosios bandymo ir daugkartinio palyginimo testo išbandytos hipotezės yra gana skirtingos, turinčios gana skirtingus galios lygius. Pavyzdžiui, bendras F iš tikrųjų paskirsto grupių skirtumus tarp grupių laisvės laipsnių. Tai sumažina bendrą kiekį F situacijoje, kai kelios grupės priemonės yra lygios viena kitai, bet skiriasi nuo kito vidurkio“ (Sveiki, 1992 m, p. 338). Būtent tokia situacija buvo taikoma šiuose eksperimentuose, kur 50 grupė buvo skirtinga nuo 0 grupės ir 100 grupės kontrolinių grupių, tačiau 25 grupei ar 75 grupei tarp šių kontrolinių grupių skirtumų nenustatyta.
Nosies dūrio duomenys patvirtino, kad kiekviename eksperimente gyvūnai įgijo ryšį tarp CS ir sacharozės atlygio perspektyvos. Skirtumas tarp skirtingų grupių nosies dūrio dažnio ir bendras jų atlyginimo dažnumas pagal atitinkamus mokymo tvarkaraščius leidžia manyti, kad vidutinė sacharozės norma yra atsakas į gydymą be narkotikų. Vis dėlto, dėl to, kad ANCOVA'uose trūksta tarpinio poveikio, susijusio su grupėmis susijusiais lokomotoriniais atsakais į AMPH, paaiškėjo, kad šie du veiksmai yra atskiri procesai.
Kai kuriais atvejais atsako į pirmąją AMPH dozę pasireiškė kondicionavimo grafiko poveikis; kitais atvejais jis atsirado tik po kartotinių dozių. Grupės lokomotorinio atsako į pirmąją AMPH dozę skirtumai rodo, kad azartinių lošimų atlygio grafikų poveikis yra pakankamas sukelti jautrinimą. Grupiniai skirtumai, susiję su judėjimu po daugelio AMPH dozių, rodo subtilesnį poveikį, kuris gali būti apibūdinamas kaip „jautrumas“, kuris pasireiškia tik tuo atveju, kai jis derinamas su nuolatiniu pirminio jautrinančio poveikio poveikiu (ty amfetaminu).
Eksperimentų atsako modelio skirtumai rodo, kad ilgesnis intervalas tarp treniruotės ir pradinio AMPH iššūkio gali maksimaliai padidinti galimybę aptikti būdingą jautrinamąjį kondicionuojančio gydymo poveikį. Tai savo ruožtu rodo, kad sąlyginio atlygio poveikis laikui bėgant gali išlikti, o šis reiškinys taip pat pastebimas su stimuliuojančiu jautrumu (Grimm et al., 2006). Atsakas į dvi amfetamino dozes rodo, kad 0.5 mg / kg dozė gali būti efektyvesnė atskleidžiant gydymo istorijos poveikį. Tai savo ruožtu rodo, kad kondicionavimo poveikis pagal dabartinį mokymo protokolą yra šiek tiek subtilus ir gali būti apsuptas lubų poveikiu pagal AMPH dozes ir sąlygas, kurios sukelia de novo jautrinimas.
3 eksperimento metu dvifazis atsakas į 0.5 mg/kg dozes ir laipsniškas pranašumo atsiradimas 50 grupėje atitinka tikėtiną Lewis žiurkių atsaką į metamfetaminą (Camp ir kt., 1994 m). Tai palaiko dabartinių išvadų pagrįstumą ir siūlo sutapti veiksnius, kurie mažina jautrumą psichostimuliatoriaus jautrinimui ir lošimų tipo atlygio grafikus.
Visuose eksperimentuose 50 grupės lokomotorinis atsakas po jautrinimo paprastai viršijo kitų grupių, vartojančių skirtingas amfetamino dozes ir skirtingų padermių gyvūnų, atsaką. Tačiau didelis kintamumas grupės viduje ir nedidelis poveikio dydžiai tarp grupių rodo kitų veiksnių vaidmenį DA sistemos reaktyvumui su amfetaminu po skirtingų sąlyginės sacharozės atlygio grafikų. Nors DA neuronų atsakas į atlygio signalus gali sudaryti grubų lošimo modelį (Fiorillo ir kt., 2003), kaip ir visi modeliai, dėl parsimonijos prarandama informacija - ty parodyti pagrindinį procesą. Kaip rezultatas, poveikis CS-US sąlygomis originalioje Fiorillo et al. tyrimas ne visiškai apibendrina lokomotorinį atsaką į amfetaminą. Reikalingi tolesni modelio patobulinimai, kad būtų visiškai užfiksuoti lošimų aspektai, turintys įtakos DA sistemos funkcijai.
Apibendrinant, šios eksperimentų serijos rezultatai preliminariai patvirtina hipotezę, kad lėtinis į azartinius lošimus panašus atlygio grafikas padidina smegenų DA sistemos reaktyvumą į psichostimuliatorių. Taigi jie išplečia išvadas Singer ir kt. (2012 m.) kurie įrodė, kad, palyginti su fiksuotu grafiku, ankstesnis kintamo sustiprinimo grafiko poveikis operantinėje paradigmoje sustiprina vėlesnį lokomotorinį atsaką į amfetaminą. Konkrečiau, šios išvados rodo, kad atlygio teikimo neapibrėžtumas yra esminis veiksnys, lemiantis kintamo atlygio poveikį. Poveikių dydis operantinėje paradigmoje buvo daug didesnis nei poveikis, nustatytas dabartiniuose eksperimentuose. Tai gali reikšti didesnį lėtinį į azartinius lošimus panašios veiklos poveikį (55, palyginti su 15 dienų); tai gali atspindėti operantinio atsako reikalavimo, kad gautų atlygį (ty agentūros vaidmenį), o ne pasyvaus poveikio, kaip šiame tyrime, poveikį. Šiuos klausimus padėtų išspręsti mokymo trukmės padidinimas pagal dabartinę paradigmą.
Iš šių eksperimentų negalima suprasti kintamo atlygio ir sustiprinimo tvarkaraščių, kaip lošimo modelių, pagrįstumo. Šią problemą galėtų išspręsti būsimi tyrimai, kuriuose nagrinėjamas kondicionavimo istorijos poveikis rizikingam elgesiui, kai žaidžiama graužikai. Panašiai, atitikimas tarp čia nustatyto elgesio jautrinimo ir padidėjusio striato DA atsako į amfetaminą, neseniai aptiktą patologinių lošėjų tarpe, turi laukti tolesnio tyrimo.Boileau ir kt., 2013). Šis klausimas gali būti sprendžiamas mikro-dializėje, o prognozėmis, pagrįstomis žmogaus duomenimis, būtų galima teigti, kad didesnis DA išskyrimas 50 grupėje „azartinių lošimų fenotipas“ būtų akivaizdžiai matomas dorsalinio (sensorinio) striatume, o ne ventralinėje (limbinėje) striatume . 50% kintamojo CS + atlygio poveikio patvirtinimas šiose kitose paradigmose paremtų jo naudingumą kaip bona fide eksperimentinį PG modelį.
Nors kai kurios lošimo formos aiškiai reiškia instrumentinį atsaką (pvz., lošimo automatai), kitose lošimo formose (pvz., loterijoje) ryšys tarp veiksmo (bilieto įsigijimo, ty statymo), atlygio užuominų (ty , loterijos skaičiai), o pats atlygis (laimėjimo skaičius ir piniginis laimėjimas) yra daug labiau išsklaidytas. Nepaisant to, gali įvykti DA aktyvinimas CS-US intervalo metu. Tai gali paaiškinti, kodėl, paskelbus „laimėjimo skaičių“, dėmesys prikaustomas, nes kiekvienas atskiras loterijos kamuoliukas krenta iš eilės, kad sudarytų konkrečią laimėjimo skaičiaus skaitmenų seką. Nors konkretaus skaitmens atsiradimo tikimybė yra apibrėžta matematiškai, kiekvieno atskiro loterijos kamuoliuko rezultatas yra dvejetainis – pataikyti (atitinka žaidėjo numerį) arba praleisti (neatitinka žaidėjo skaičiaus), o bet kurio bandymo rezultatas nežinomas. Toks scenarijus gali geriau apibūdinti 50 grupės patirtį dabartiniuose eksperimentuose, kai atlygis buvo teikiamas ne atsitiktinai, bet ir nenuspėjamai, o CS tik nurodė atlygio potencialą, neatskleidžiant, ar jis įvyks tam tikro bandymo metu. Lošimo automatai yra labiau susiję su PG nei loterijos bilietai (Cox ir kt., 2000; Bakken ir kt., 2009), nurodant svarbų instrumentinių veiksnių (ir betarpiškumo) vaidmenį teikiant azartinių lošimų naudingumą šiai populiacijai (Loba ir kt., 2001 m). Nepaisant to, dabartiniuose eksperimentuose suprojektuotas Pavlovijos procesas (CS + neapibrėžtas atlygis) atrodo būtinas ar ne pakankamas lošimų patirties elementas.
Be to, kad nėra aiškių priemonių reikalavimo, keletas kitų dizaino ypatybių galėjo prisidėti prie palyginti kuklios ir kintančios eksperimentinio poveikio. Grupės skiriasi nuo bendros sacharozės ekspozicijos, taip pat nuo CS ir sacharozės atlygio. Nors tai galėjo prisidėti prie grupės narių kintamumo, ji negali lengvai paaiškinti, kodėl gyvūnai, kurių ekspozicija yra didžiausia (100 grupė), jautriau nei 50 grupėje. Be to, 0 grupėje nebuvo jokio stimulo prieš sacharozės ekspoziciją kiekviename tyrime. Nors tai užkerta kelią paskaičiuotam atlygio lūkesčiui, ji nekontroliavo stimulo buvimo prieš atlygio pristatymą, kuris egzistavo visose kitose grupėse. Siekiant išspręsti šią problemą, būsimi moksliniai tyrimai turėtų apimti sąlygą, kai gyvūnai gauna atlygį už kiekvieną bandymą, atsitiktinai paveikus neutralų stimulą (ty, kurio buvimas neparodo atlygio potencialo).
Kitas dizaino apribojimas yra galimas papildomas elgesys, galintis turėti įtakos treniruočių grafiko poveikiui. Neapibrėžtumo akivaizdoje gyvūnams gali išsivystyti prietaringas elgesys, skirtas sustiprinti suvokiamą kontrolę ir sumažinti netikrumo sukeltą DA aktyvavimą (plg. Harris ir kt., 2013). Todėl įmanoma, kad nekontroliuojami eksperimentinio projekto aspektai leido gyvūnams kompensuoti kondicionavimo grafiko poveikį. Toks poveikis galėtų prisidėti prie santykinai kuklios ir kintančios reakcijos į amfetaminą 50 grupėje po CS + sacharozės mokymo. Ateityje atliekami moksliniai tyrimai turėtų užfiksuoti spontanišką elgesį, išskyrus nosies pokes, treniruočių metu, siekiant išbandyti šią galimybę, ir ją statistiškai kontroliuoti, jei ji atsiras. Kadangi tikimasi, kad toks elgesys sušvelnins ar sumažins grafiko sukeltos netikrumo poveikį, 50 grupėje reikia sustiprinti lokomotorinį atsaką į amfetaminą, kai jis yra kontroliuojamas (procedūriniu ar statistiniu požiūriu). Todėl dabartinis (nekontroliuojamas) dizainas užtikrina konservatyvų 50% CS + atlygio poveikio amfetamino jautrumo tyrimą.
Kalbant apie išorinį pagrįstumą, žiurkių patinų naudojimas taip pat riboja rezultatų apibendrinimą. Aiškios „bausmės“ sąlygos nebuvimas taip pat skiriasi nuo azartinių lošimų, kur dažnai patiriami dideli piniginiai nuostoliai ir jie daro svarbų motyvacinį poveikį (Nieuwenhuis ir kt., 2005; Singhas ir Khanas, 2012 m). Galimybė kaupti atlygį taip pat nėra dabartinėje paradigmoje ir buvo nustatyta, kad sukaupti laimėjimai lošimo automatų žaidime sąveikauja su DA manipuliacijomis žmonėms (Tremblay ir kt., 2011; Smart et al., 2013). Panašiai ir galimybė laimėti jackpotą yra svarbus skirtumas tarp dabartinio modelio ir tikrojo lošimo.
Nepaisant šių apribojimų, dabartiniai rezultatai rodo, kad 50% kintamas CS + atlygis gali įtraukti DA kelius, susijusius su stiprėjančiu lošimo poveikiu (Fiorillo ir kt., 2003; „Anselme“, „2013“). Kryžminis atsako į AMPH jautrinimas pagal šį į lošimą panašų tvarkaraštį atitinka pagrindinį DA vaidmenį azartinių lošimų ir psichostimuliuojančių vaistų poveikiui.Zack ir Poulos, 2009) ir pratęsia ankstesnius tyrimus, susijusius su AMPH sukeliamu kryžmine motyvacija lošti patologiniais lošėjais (Zack ir Poulos, 2004). Dabartiniai rezultatai taip pat netiesiogiai rodo, kad nedidelės AMPH dozės, kurios nesukelia suprafiziologinio DA išsiskyrimo, gali geriau modeliuoti smegenų veiklą, reaguojant į nutrūkstančius atlygio signalus (ty lošimo metu), nei veikiant dideliam (ty, panašaus į persivalgymą) stimuliuojančių vaistų dozės (plg. Vanderschuren ir Pierce, 2010). Tiesioginė parama šiai korespondencijai gali būti nustatoma vertinant DA išsiskyrimą, reaguojant į kintamąjį CSN-US planą 50% ir skirtingas AMPH dozes, naudojant mikrodializę.
Eksperimentiniu požiūriu dabartinis Pavlovijos modelis ir ankstesnis kintamo stiprinimo operacinis modelis, atrodo, sukelia fenotipą, panašų į žmogaus patologinį žaidėją. Tokiu būdu jie yra vertingas papildymas graužikų lošimų užduotims, kurios modeliuoja lošimų elgesį (kaip priklausomą priemonę), tačiau iki šiol naudojo tik sveikus gyvūnus, lygiaverčius žmonių socialiniams lošėjams. Remiantis literatūra, gyvūnai, kurie nuolat patiria kintamą atlygį, šiose užduotyse gali labai skirtis, ypač atsakant į DA-ergologinius vaistus. Derinant žiurkių lošimų fenotipą su lošimo užduotimis, gali būti sistemingai kuriami vaistai, skirti gydyti PG, kurie gali būti visiškai nevykdomi vien su sveikais gyvūnais. Papildomi eksperimentinio projektavimo ir mokymo režimo patobulinimai, kaip aprašyta pirmiau, turėtų pagerinti šio paradigmoje apmokytų gyvūnų ir faktinių patologinių žaidėjų atitiktį.
Klinikiniu ir sociologiniu požiūriu išvada, kad 50 % kintamo CS + atlygis, kuris labai atitinka atlygio grafiką komerciniuose lošimo automatuose (Tremblay ir kt., 2011), keičia smegenų DA sistemą patikimais ir ilgalaikiais būdais, rodo, kad kai kuriais atvejais azartinių lošimų veikla, kaip ir piktnaudžiavimo vaistai, gali būti „patogenas“, galintis sukelti priklausomybę. Tačiau nedidelis poveikis ir didelis kintamumas, reaguojant į 50% CS + atlygį, rodo, kad, kaip ir piktnaudžiavimo narkotikais, azartinių lošimų skatinimo tvarkaraščių tendencija skatinti priklausomybę labai priklausys nuo ankstesnio lošėjo rizikos profilio. Vis dėlto, norint išvengti didelės rizikos asmenų, galinčių sukelti galimą neigiamą poveikį azartiniams lošimams, atrodo pagrįsta, kad piktnaudžiavimo narkotikais prevencijos ir žalos mažinimo politika taip pat galėtų būti taikoma ir lošimams.
Interesų konflikto pareiškimas
Autoriai teigia, kad tyrimas buvo atliktas nesant jokių komercinių ar finansinių santykių, kurie galėtų būti laikomi galimu interesų konfliktu.
Padėka
Šis tyrimas buvo finansuojamas Kanados gamtos mokslų ir inžinerijos mokslinių tyrimų tarybos dotacijomis Pauliui J. Fletcheriui. Nuoširdžiai dėkojame Djurdja Djordjevičiui už skaitinių parengimą.
Nuorodos
Anselme, P. (2013). Dopaminas, motyvacija ir evoliucinė azartinių lošimų elgsenos reikšmė. Behav. Brain Res. 256, 1 – 4. doi: 10.1016 / j.bbr.2013.07.039
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
APA (2013). Psichikos sutrikimų diagnostikos ir statistikos vadovas, 5th Edn. Arlingtonas, VA: Amerikos psichiatrijos leidyba.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG ir Oren, A. (2009). Azartinių lošimų elgesys ir lošimo problemos Norvegijoje 2007 m. Scand. J. Psychol. 50, 333 – 339. doi: 10.1111 / j.1467-9450.2009.00713.x
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Boileau, I., Payer, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA ir kt. (2013). In vivo Įrodymai dėl didesnio amfetamino sukelto dopamino išsiskyrimo sergant patologiniu lošimu: pozitronų emisijos tomografijos tyrimas su [C]-(+)-PHNO. Mol. Psichiatrija doi: 10.1038/mp.2013.163. [Epub prieš spausdinimą].
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Bolles, RC (1972). Sustiprinimas, laukimas ir mokymasis. Psychol. Rev. 79, 394 – 409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE ir Robinson, TE (1994). Metamfetamino ir kokaino poveikis motoriniam elgesiui ir ekstraląsteliniam dopaminui Lewiso ir Fischer 344 žiurkių ventraliniame striatum. Smegenų raiška. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G. ir Dell'Osso, L. (2012). Patologinis lošimas: sisteminga biocheminių, neurovaizdinių ir neuropsichologinių radinių apžvalga. Harv. kun. Psichiatrija 20, 130 – 148. doi: 10.3109 / 10673229.2012.694318
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Cox, BJ, Kwong, J., Michaud, V. ir Enns, MW (2000). Problema ir galimas patologinis lošimas: bendruomenės apklausos svarstymai. Gal. J. Psichiatrija 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. ir Robbins, TW (2002). Impulsų kontrolės trūkumai, susiję su toniškai padidėjusia serotoninergine funkcija žiurkės prefrontalinėje žievėje. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Everitt, BJ ir Robbins, TW (2005). Narkotikų stiprinimo neuronų sistemos: nuo veiksmų iki įpročių iki prievartos. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Fiorillo, CD, Tobler, PN ir Schultz, W. (2003). Dopamino neuronų neuronų atlyginimų tikimybės ir neapibrėžties diskretiškas kodavimas. Mokslas 299, 1898 – 1902. doi: 10.1126 / science.1077349
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Flagel, SB, Watson, SJ, Akil, H. ir Robinson, TE (2008). Individualūs paskatų priskyrimo su atlygiu susijusiam signalui skirtumai: įtaka kokainui. Behav. Brain Res. 186, 48 – 56. doi: 10.1016 / j.bbr.2007.07.022
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V. ir Kapur, S. (2005). Jautrinimas amfetaminui, bet ne PCP, pablogina dėmesio rinkinio pasikeitimą: D1 receptorių agonisto, suleidžiamo į medialinę prefrontalinę žievę, pakeitimą. Psichofarmakologija (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Flores, G., Wood, GK, Barbeau, D., Quirion, R. ir Srivastava, LK (1998). Lewis ir Fischer žiurkės: dopamino transporterio ir receptorių lygių palyginimas. Smegenų raiška. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Frascella, J., Potenza, MN, Brown, LL ir Childress, AR (2010). Bendras smegenų pažeidžiamumas atveria kelią priklausomybei nuo ne medžiagų: priklausomybė nuo raižinio prie naujo sąnario? Ann. NY Acad. Sci. 1187, 294 – 315. doi: 10.1111 / j.1749-6632.2009.05420.x
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A. ir Wells, B. (2006). Nuo laiko priklausomas kokaino dozės ir atsako poveikio disociacija sacharozės troškimui ir judėjimui. Behav. Pharmacol. 17, 143–149. doi: 10.1097/01.fbp.0000190686.23103.f8
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Haile, CN, Hiroi, N., Nestler, EJ ir Kosten, TA (2001). Skirtingos elgsenos reakcijos į kokainą yra susijusios su mezolimbinių dopamino baltymų dinamika Lewis ir Fischer 344 žiurkėse. Synapse 41, 179 – 190. doi: 10.1002 / syn.1073
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Harris, JA, Andrew, BJ ir Kwok, DW (2013). Žurnalo požiūris per signalą maistui priklauso nuo Pavloviško, o ne instrumentinio kondicionavimo. J. Exp. Psychol. Anim. Behav. Procesas. 39, 107 – 116. doi: 10.1037 / a0031315
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Howell, DC (1992). Statistiniai psichologijos metodai. Bostonas, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW ir Everitt, BJ (2002). Dopamino išsiskyrimas nugaros juostoje ieškant kokaino, kontroliuojant su narkotikais susijusią užuominą. J. Neurosci. 22, 6247 – 6253.
Jiao, X., Pare, WP ir Tejani-Butt, S. (2003). Įtempių skirtumai tarp dopamino transporterių vietų pasiskirstymo žiurkės smegenyse. Prog. Neuropsichofarmacol. Biol. Psichiatrija 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Koob, GF ir Le Moal, M. (2008). Apžvalga. Neurobiologiniai mechanizmai oponento motyvacijos procesams priklausomybėje. Filosas. Trans. R. Soc. Lond. B Biol. Sci. 363, 3113 – 3123. doi: 10.1098 / rstb.2008.0094
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Kosten, TA, Miserendino, MJ, Chi, S. ir Nestler, EJ (1994). Fischer ir Lewis žiurkių padermės rodo skirtingą kokaino poveikį sąlygiškai pasirenkant vietą ir elgsenos jautrinimą, bet ne dėl lokomotorinio aktyvumo ar sąlyginio skonio baimės. J. Pharmacol. Gal. Ther. 269, 137 – 144.
Kosten, TA, Zhang, XY ir Haile, CN (2007). Kokaino savarankiško vartojimo palaikymo skirtumai ir jų ryšys su naujoviškos veiklos atsakais. Behav. Neurosci. 121, 380 – 388. doi: 10.1037 / 0735-7044.121.2.380
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Leeman, RF ir Potenza, MN (2012). Panašumai ir skirtumai tarp patologinių azartinių lošimų ir medžiagų vartojimo sutrikimų: dėmesys impulsyvumui ir kompulsyvumui. Psichofarmakologija (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Leshner, AI (1997). Priklausomybė yra smegenų liga, ir tai svarbu. Mokslas 278, 45 – 47. doi: 10.1126 / science.278.5335.45
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Loba, P., Stewart, SH, Klein, RM ir Blackburn, JR (2001). Manipuliacijos standartinių vaizdo loterijų terminalo (VLT) žaidimų ypatybėmis: poveikis patologiniams ir nepatologiniams lošėjams. J. Gambl. Stud. 17, 297–320. doi: 10.1023 / A: 1013639729908
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A. ir kt. (2007). Amfetamino sukeltas dopamino išsiskyrimas: labai sumažinta priklausomybė nuo kokaino ir nuspėjamas pasirinkimas savarankiškai vartoti kokainą. Esu. J. Psichiatrija 164, 622 – 629. doi: 10.1176 / appi.ajp.164.4.622
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC ir Jones, SR (2005). Sumažėjusi dopamino galinė funkcija ir nejautrumas kokainui po kokaino vartojimo ir nepritekliaus. Neuropsychopharmacology 30, 1455 – 1463. doi: 10.1038 / sj.npp.1300687
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
„Nestler“, EJ (2001). Ilgalaikio priklausomybės plastikos molekuliniai pagrindai. Nat. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB ir Yeung, N. (2005). Aktyvumas žmogaus atlygiui jautriose smegenų srityse labai priklauso nuo konteksto. Neuroimage 25, 1302 – 1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Pierce, RC ir Kalivas, PW (1997). Elgesio jautrumo amfetaminams panašiems psichostimuliatoriams išraiškos schema. Brain Res. Brain Res. Rev. 25, 192. doi: 10.1016/S0165-0173(97)00021-0
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Robbins, TW ir Everitt, BJ (1999). Priklausomybė nuo narkotikų: blogi įpročiai didėja. Gamta 398, 567 – 570. doi: 10.1038 / 19208
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Robinson, TE, Becker, JB ir Presty, SK (1982). Ilgalaikis amfetamino sukeltos sukimosi elgsenos ir striatalinio dopamino išsiskyrimo, kurį sukelia vienkartinis amfetamino poveikis, palengvinimas: lyčių skirtumai. Smegenų raiška. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Robinson, TE ir Berridge, KC (1993). Narkotikų troškimo nervų pagrindas: skatinamojo jautrumo priklausomybės teorija. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Robinson, TE ir Berridge, KC (2001). Skatinamasis jautrinimas ir priklausomybė. Polinkis 96, 103 – 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Singer, BF, Scott-Railton, J. ir Vezina, P. (2012). Neprognozuojamas sacharino sutvirtinimas sustiprina lokomotorinį atsaką į amfetaminą. Behav. Brain Res. 226, 340 – 344. doi: 10.1016 / j.bbr.2011.09.003
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Singh, V. ir Khan, A. (2012). Sprendimų priėmimas dėl Ajovos lošimo užduoties apdovanojimo ir bausmės variantų: „numatymo“ ar „įrėminimo“ įrodymas? Priekyje. Neurosci. 6: 107. doi: 10.3389 / fnins.2012.00107
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Skinner, BF (1953). Mokslas ir žmogaus elgesys. Niujorkas, NY: Laisva spauda.
Smart, K., Desmond, RC, Poulos, CX ir Zack, M. (2013). Modafinilas padidina atlygį lošimo automatų žaidime esant mažo ir didelio impulsyvumo patologiniams lošėjams. Neurofarmakologija 73, 66 – 74. doi: 10.1016 / j.neuropharm.2013.05.015
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Tremblay, AM, Desmond, RC, Poulos, CX ir Zack, M. (2011). Haloperidolis modifikuoja lošimo automatų lošimo instrumentinius aspektus patologiniams lošėjų ir sveikų asmenų lošimams. Addict. Biol. 16, 467 – 484. doi: 10.1111 / j.1369-1600.2010.00208.x
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Vanderschuren, LJ ir Kalivas, PW (2000). Dopaminerginio ir glutamaterginio perdavimo pokyčiai sukeliant ir išreiškiant elgesio jautrinimą: kritinė ikiklinikinių tyrimų apžvalga. Psichofarmakologija (Berl.) 151, 99 – 120. doi: 10.1007 / s002130000493
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Vanderschuren, LJ ir Pierce, RC (2010). Jautrinimo procesai priklausomybėje nuo narkotikų. Curr. Į viršų. Behav. Neurosci. 3, 179–195. doi: 10.1007/7854_2009_21
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH ir De Vries, TJ (1999). Dopaminerginiai mechanizmai, tarpininkaujantys ilgalaikei lokomotorinio jautrumo išraiškai po išankstinio morfino ar amfetamino poveikio. Psichofarmakologija (Berl.) 143, 244 – 253. doi: 10.1007 / s002130050943
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Verdejo-Garcia, A., Lawrence, AJ ir Clark, L. (2008). Impulsyvumas kaip medžiagų vartojimo sutrikimų pažeidžiamumo žymuo: didelės rizikos tyrimų, probleminių lošėjų ir genetinių asociacijų tyrimų išvadų apžvalga. Neurosci. Biobehav. Rev. 32, 777 – 810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Vezina, P. (2004). Vidurinių smegenų dopamino neuronų reaktyvumo jautrinimas ir psichomotorinius stimuliuojančių vaistų savarankiškas vartojimas. Neurosci. Biobehav. Rev. 27, 827 – 839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R. ir kt. (1997). Sumažėjęs striato dopaminerginis atsakas detoksikuotiems nuo kokaino priklausomiems asmenims. Gamta 386, 830 – 833. doi: 10.1038 / 386830a0
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Volkow, ND, Wang, GJ, Fowler, JS ir Tomasi, D. (2012). Priklausomybės grandinė žmogaus smegenyse. Annu. Rev. Pharmacol. Toksikolis. 52, 321–336. doi: 10.1146/annurev-pharmtox-010611-134625
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Winer, B. (red.). (1971). Eksperimentinio projektavimo statistiniai principai. Niujorkas, NY: McGraw-Hill.
Zack, M. ir Poulos, CX (2004). Amfetaminas skatina lošti azartinius lošimus ir su azartiniais lošimais susijusius semantinius tinklus probleminiuose lošėjus. Neuropsychopharmacology 29, 195 – 207. doi: 10.1038 / sj.npp.1300333
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
M. Zackas ir Poulos, CX (2009). Paraleliniai dopamino vaidmenys patologinių lošimų ir psichostimuliatorių priklausomybėje. Curr. Piktnaudžiavimas narkotikais, red. 2, 11 – 25. doi: 10.2174 / 1874473710902010011
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F. ir Flores, G. (2005). Dopamino receptorių lygio skirtumai ir su dopaminu susijęs elgesys žiurkėms. Brain Res. Bull. 65, 339–347. doi: 10.1016/j.brainresbull.2005.01.009
Pubmed Santrauka | Išleistas visas tekstas | „CrossRef“ visas tekstas
Raktažodžiai: patologinis lošimas, jautrinimas, amfetaminas, dopaminas, netikrumas
Citata: Zack M, Featherstone RE, Mathewson S ir Fletcher PJ (2014) Lėtinis į azartinius lošimus panašus atlygio nuspėjamųjų dirgiklių grafiko poveikis gali paskatinti žiurkių jautrumą amfetaminui. Priekyje. Behav. Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Gauta: 01 lapkritis 2013; Priimta: 23 Sausis 2014;
Paskelbta internete: 11 m. vasario 2014 d.
Redagavo:
Bryan F. SingerMičigano universitetas, JAV
Peržiūrėjo:
Loukas Vanderschurenas, Utrechto universitetas, Nyderlandai
Ruudas Van Den Bosas, Radboud universitetas Nijmegen, Nyderlandai
Patrick AnselmeLiège universitetas, Belgija
Autorių teisės © 2014 Zack, Featherstone, Mathewson ir Fletcher. Tai yra atviros prieigos straipsnis, platinamas pagal sąlygas „Creative Commons“ priskyrimo licencija (CC BY). Leidimas naudoti, platinti ar atgaminti kituose forumuose yra leidžiamas, jei originalus autorius (-iai) arba licencijos išdavėjas yra įskaitytas (-i) ir kad originalus leidinys šiame žurnale yra cituojamas pagal priimtą akademinę praktiką. Negalima naudoti, platinti ar atgaminti, kuri neatitinka šių sąlygų.
* Korespondencija: Martinas Zackas, Kognityvinės psichofarmakologijos laboratorija, Neurologijos departamentas, Priklausomybės ir psichikos sveikatos centras, 33 Russell Street, Torontas, ON M5S 2S1, Kanada el. paštas: [apsaugotas el. paštu]
