Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
Anotācija
Pusaudža vecums ir attīstības periods, ko raksturo nepietiekami optimāli lēmumi un darbības, kas palielina nejaušas traumas un vardarbību, alkohola un narkotiku lietošanu, neparedzētu grūtniecību un seksuāli transmisīvās slimības. Tradicionālie neirobioloģiskie un kognitīvie pusaudžu uzvedības izskaidrojumi nav devuši iespēju ņemt vērā nelineārās izmaiņas uzvedībā, kas novērotas pusaudža gados, salīdzinot ar bērnību un pieaugušo vecumu. Šis pārskats sniedz bioloģiski ticamu konceptuālo neironu mehānismu koncepciju, kas balstās uz šīm nelineārajām uzvedības izmaiņām, kā paaugstināta reakcija uz stimuliem, bet impulsa kontrole šajā periodā joprojām ir relatīvi nenobriedusi. Nesenie cilvēka attēlveidošanas un dzīvnieku pētījumi sniedz bioloģisku pamatu šim viedoklim, kas liek domāt par limbisko atalgojuma sistēmu atšķirīgu attīstību attiecībā uz augšupvērstām kontroles sistēmām pusaudža gados salīdzinājumā ar bērnību un pieaugušo vecumu. Šis attīstības modelis var saasināties pusaudžiem, kam ir nosliece uz riska uzņemšanos, palielinot sliktu rezultātu risku.
Saskaņā ar Nacionālā veselības statistikas centra datiem katru gadu Amerikas Savienotajās Valstīs ir vairāk nekā 13,000 pusaudžu nāves gadījumu. Aptuveni 70% no šiem nāves gadījumiem rodas automobiļu avāriju, nejaušu ievainojumu, slepkavību un pašnāvību rezultātā (Eaton et al., 2006). 2005 Nacionālā jaunatnes riska uzvedības apsekojuma (YRBS) rezultāti rāda, ka pusaudži iesaistās uzvedībā, kas palielina viņu nāves vai slimības iespējamību, braucot transportlīdzekli pēc dzeršanas vai bez drošības jostas, nēsājot ieročus, izmantojot nelegālas vielas un iesaistoties neaizsargātā dzimuma dzīvē izraisot nejaušu grūtniecību un STS, tostarp HIV infekciju (\ tEaton et al., 2006). Šī statistika uzsver, cik svarīga ir izpratne par riskantu izvēli un rīcību pusaudžiem.
Ir izteiktas vairākas kognitīvās un neirobioloģiskās hipotēzes, kāpēc pusaudži iesaistās suboptimālajā izvēlē. Nesenā literatūras pārskatā par cilvēka pusaudžu smadzeņu attīstību, \ t Yurgelun-Todd (2007) norāda, ka kognitīvā attīstība caur pusaudžu gadiem ir saistīta ar kognitīvās kontroles kapacitātes pakāpenisku palielināšanu. Šī efektivitāte ir raksturota kā atkarīga no prefronta garozas nogatavināšanas, ko apliecina pastiprināta aktivitāte fokusa prefrontajos reģionos (Rubia et al., 2000; Tamm, Menons un Reiss, 2002. gads) un samazināta aktivitāte neatbilstīgos smadzeņu reģionos (Brown et al., 2005; Durston et al., 2006).
Šis vispārējais modelis, uzlabota kognitīvā kontrole ar prefronta garozas nogatavināšanu, liecina par lineāru pieaugumu no bērnības līdz pilngadībai. Tomēr pusaudža laikā novērotās suboptimālās izvēles un darbības ir nelineāra uzvedības izmaiņas, ko var atšķirt no bērnības un pieauguša cilvēka vecuma, kā to apliecina Nacionālais veselības statistikas centrs par pusaudžu uzvedību un mirstību. Ja kognitīvā kontrole un nenobriedis prefrontālais garozas pamats bija suboptimāla izvēle, tad bērniem, ņemot vērā to mazāk attīstīto prefronālo garozu un kognitīvās spējas, vajadzētu izskatīties ļoti līdzīgi vai pat sliktāk nekā pusaudži. Tādējādi tikai nenobrieduša prefronta funkcija nevar ņemt vērā pusaudžu uzvedību.
Precīza kognitīvo un neirobioloģisko izmaiņu konceptualizācija pusaudža gados ir jāapskata pusaudža vecumam kā pārejas periodam (Spear, 2000), nevis vienu momentuzņēmumu (Keisija, Totenhema, Listons un Durstons, 2005. gads). Citiem vārdiem sakot, lai saprastu šo attīstības periodu, pārejas uz pusaudžu vecumu un no tās ir nepieciešamas, lai atšķirtu šīs attīstības stadijas atšķirīgās īpašības. Kognitīvo un neironu procesu attīstības trajektoriju noteikšana ir būtiska, lai raksturotu šīs pārejas un ierobežotu interpretācijas par izmaiņām uzvedībā šajā periodā. Kognitīvā vai uzvedības līmenī pusaudži tiek raksturoti kā impulsīvi (ti, trūkst kognitīvās kontroles), un riska uzņemšanās ar šīm konstrukcijām tiek izmantota sinonīmi un nenovērtējot katras atsevišķās attīstības trajektorijas. Neirobioloģiskā līmenī cilvēka attēlveidošanas un dzīvnieku pētījumi liecina par atšķirīgām neirobioloģiskām bāzēm un attīstības trajektorijām nervu sistēmām, kas ir šo atsevišķo impulsu kontroles un riska lēmumu pamatā.
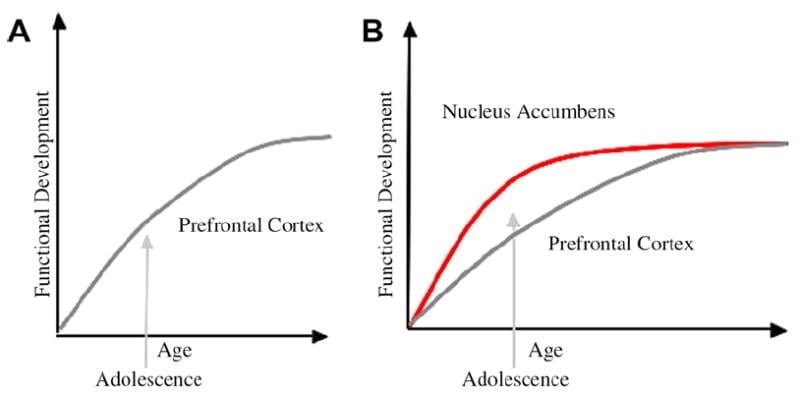
Šajā sistēmā esam izstrādājuši pusaudžu attīstības neirobioloģisko modeli, kas balstās uz grauzēju modeļiem (Laviola, Adriani, Terranova un Gerra, 1999. gads; Spear, 2000) un jaunākie pusaudžu attēlveidošanas pētījumi (Ernst et al., 2005; Galvans, Zaķis, Voss, Glovers un Keisijs, 2007. gads; Galvan et al., 2006). Fig. 1 zemāk attēlots šis modelis. Kreisajā pusē ir tradicionāls pusaudža raksturojums, kas gandrīz pilnībā saistīts ar prefrontālās garozas nenobriedumu. Labajā pusē ir mūsu ierosinātā neirobioloģiskā modeļa paraugs, kas ilustrē, kā limbiskie subortikālie un prefrontālie augšupvērsti kontroles reģioni ir jāaplūko kopā. Karikatūra ilustrē dažādas šo sistēmu attīstības trajektorijas, un limbiskās sistēmas attīstās agrāk nekā prefrontālās kontroles reģioni. Saskaņā ar šo modeli indivīds ir vairāk nobriedis no funkcionāli nobriedušiem limbiskajiem reģioniem pusaudža vecumā (ti, limbiskā nelīdzsvarotība attiecībā pret prefrontālo kontroli), salīdzinot ar bērniem, kuriem šīs sistēmas (ti, limbiskā un prefrontālā) joprojām attīstās; salīdzinot ar pieaugušajiem, kuriem šīs sistēmas ir pilnībā nobriedušas. Šī perspektīva nodrošina pamatu nelineārām izmaiņām uzvedībā pāri attīstībai, jo šī limbiskā brieduma agrākā nobriešana ir saistīta ar mazāk nobriedušu augšupvērstu prefronta kontroles reģionu. Ar attīstību un pieredzi, funkcionālais savienojums starp šiem reģioniem nodrošina mehānismu šo reģionu augšupējai kontrolei (Zaķis, Voss, Glovers un Keisijs, 2007a). Turklāt modelis saskaņo pretrunu ar veselības statistiku par riskantu uzvedību pusaudža vecumā ar izsmalcinātu novērojumu, ko veic Reyna un Farley (2006) ka pusaudži spēj saprast un saprast uzvedības riskus, kuros viņi iesaistās. Saskaņā ar mūsu modeli emocionāli svarīgās situācijās limbiskā sistēma uzvarēs pār kontroles sistēmām, ņemot vērā tās briedumu attiecībā pret prefrontālās vadības sistēmu. Pierādījumi no uzvedības un cilvēka attēlveidošanas pētījumiem, lai atbalstītu šo modeli, tiek sniegti saistībā ar darbībām, kas ir atalgojošas un emocionālas (Galvan et al., 2006, 2007; Zaķis, Voss, Glovers un Keisijs, 2007b; Hare et al., 2007a). Turklāt mēs spekulējam, kāpēc smadzenes var attīstīties šādā veidā un kāpēc dažiem pusaudžiem var būt lielāks risks pieņemt optimālus lēmumus, kas noved pie sliktākiem ilgtermiņa rezultātiem (Galvan et al., 2007; Hare et al., 2007b).
Tiek ieteikts, ka tradicionālais pusaudžu uzvedības skaidrojums ir saistīts ar prefrontālās garozas (A) ilgstošo attīstību. Mūsu modelis ņem vērā prefrontālās garozas attīstību kopā ar subortikālajiem limbiskajiem reģioniem (piem., Kodolu accumbens), kas ir iesaistīti riskantās izvēles un darbībās (B).
Mērķtiecīgas uzvedības attīstība
Kognitīvās attīstības stūrakmens ir spēja nomākt nepiemērotas domas un darbības par labu mērķtiecīgām, jo īpaši pārliecinošu stimulu (Keisija, Galvans un Zaķis, 2005. gads; Casey et al., 2000b; Keisijs, Tomass, Deivids-dēls, Kuncs un Franzens, 2002.a.; Keisija, Totenhema un Foselja, 2002.b.). Vairāki klasiski attīstības pētījumi liecina, ka šī spēja attīstās bērnībā un pusaudža gados (Korpuss, 1972; Flavell, Feach un Chinsky, 1966. gads; Kītings un Bobbits, 1978. gads; Pascual-Leone, 1970). Vairāki teorētiķi apgalvoja, ka kognitīvā attīstība ir saistīta ar apstrādes ātruma un efektivitātes pieaugumu, nevis garīgās spējas pieauguma dēļ (piemēram, Bjorkland, 1985; Bjorkland, 1987; Korpuss, 1985). Citi teorētiķi ir iekļāvuši “inhibējošo” procesu konstruēšanu savā kognitīvās attīstības kontā (Harnishfeger & Bjorkland, 1993. gads). Saskaņā ar šo kontu nenobriedušo izziņu raksturo uzņēmību pret konkurējošu avotu iejaukšanos, kas ir jānovērš (piemēram, Brainerd & Reyna, 1993. gads; Keisijs, Tomass, Deividsons, Kuncs un Franzens, 2002.a.; Dempster, 1993; Dimants, 1985; Munakata & Yerys, 2001. gads). Tādējādi mērķtiecīgai uzvedībai ir nepieciešama impulsu kontrole vai apmierinātības kavēšanās, lai optimizētu rezultātus, un šī spēja izzūd pāri bērnībai un pusaudža vecumam.
Pusaudžu uzvedība ir raksturota kā impulsīva un riskanta, gandrīz sinonīmi, tomēr šīs konstrukcijas balstās uz dažādiem kognitīviem un neironu procesiem, kas liecina par atšķirīgām konstrukcijām ar atšķirīgām attīstības trajektorijām. Konkrētāk, literatūras apskats liecina, ka impulsivitāte ar vecumu samazinās bērnībā un pusaudža gados (Casey et al., 2002a; Casey, Galvan et al., 2005; Galvan et al., 2007) un ir saistīta ar ilgstošu prefronta garozas attīstību (Casey, Galvan et al., 2005), lai gan pastāv atšķirības atkarībā no tā, cik lielā mērā konkrēts indivīds ir impulsīvs vai nē, neatkarīgi no vecuma.
Pretstatā tam, impulsa / kognitīvās kontroles gadījumā, šķiet, ka riska uzņemšanās pusaudža vecumā palielinās salīdzinājumā ar bērnību un pieaugušo vecumu, un tā ir saistīta ar subkortu sistēmām, par kurām zināms, ka tās piedalās atalgojuma novērtēšanā. Cilvēka attēlveidošanas pētījumi, kas tiks pārskatīti, liecina, ka, veicot riskantus lēmumus, palielinās subortikālā aktivācija (piemēram, accumbens).Kuhnen & Knutson, 2005. gads; Matthews u.c., 2004. gads; Montāga un Berns, 2002. gads), kas ir pārspīlēts pusaudžiem, salīdzinot ar bērniem un pieaugušajiem (Ernst et al., 2005; Galvan et al., 2006). Šie atklājumi liecina par dažādām trajektorijas tendencēm, kas balstītas uz atalgojuma vai stimulējošu uzvedību, iepriekš izstrādājot šīs sistēmas attiecībā uz kontroles sistēmām, kurās parādās ilgstošs un lineārs attīstības virziens, proti, nepiemērotu nepareizu izvēli un rīcību par labu mērķtiecīgām.
Pierādījumi, kas iegūti, pētot cilvēka smadzeņu attīstību
Jaunākie pētījumi par pusaudžu smadzeņu attīstību ir balstīti uz progresu neiromodēšanas metodoloģijās, ko var viegli izmantot ar jaunattīstības cilvēku populācijām. Šīs metodes balstās uz magnētiskās rezonanses (MRI) metodēm (skatīt. \ T Fig. 2) un ietver: strukturālo MRI, ko izmanto, lai izmērītu struktūru lielumu un formu; funkcionāls MRI, ko izmanto smadzeņu darbības modeļu mērīšanai; un difūzijas tenzora attēlveidošana (DTI), ko izmanto, lai indeksētu savienojumu ar balto vielu šķiedrām. Pierādījumus par mūsu konkurences attīstību starp kortikālajiem un subortikālajiem reģioniem apstiprina nenobriedis strukturālais un funkcionālais savienojums, ko mēra attiecīgi ar DTI un fMRI.
MRI pētījumi par cilvēka smadzeņu attīstību
Vairāki pētījumi ir izmantojuši strukturālo MRI, lai kartētu normālās smadzeņu attīstības anatomisko gaitu (sk Durston et al., 2001). Lai gan kopējais smadzeņu lielums ir aptuveni 90% no pieaugušo lieluma, seši gadi, smadzeņu pelēkās un baltās vielas sastāvdaļas turpina dinamiski mainīties pusaudža vecumā. Pēdējo garenisko MRI pētījumu dati liecina, ka pelēkās vielas tilpumam ir apvērsts U-veida modelis, ar lielāku reģionālo variāciju nekā baltajai vielai (Giedd, 2004; Gogtay et al., 2004; Sowell et al., 2003; Sowell, Thompson un Toga, 2004). Kopumā reģioni, kas izmanto primārās funkcijas, piemēram, motora un sensorās sistēmas, ir nobrieduši agrāk; augstākas kārtības asociācijas apgabali, kas integrē šīs primārās funkcijas, ir nobrieduši vēlāk (Gogtay et al., 2004; Sowell, Thompson un Toga, 2004). Piemēram, pētījumi, kuros izmanto MRI balstītus pasākumus, liecina, ka kortikālo pelēkās vielas zudums notiek ātrāk primārajās sensorimotoru zonās un vēlākais dorsolaterālā prefrontālā un sānu laikmeta dzīslās (Gogtay et al., 2004). Šis modelis atbilst cilvēka primāta un cilvēka pēcdzemdību pētījumiem, kas liecina, ka prefronta garoza ir viens no pēdējiem smadzeņu reģioniem, kas ir nobrieduši (Buržujs, Goldmens-Rakičs un Rakičs, 1994. gads; Huttenlocher, 1979). Atšķirībā no pelēkās vielas, balto vielu tilpums palielinās aptuveni lineārā veidā, pieaugot visā attīstībā līdz pieaugušo vecumam (Gogtay et al., 2004). Šīs pārmaiņas, iespējams, atspoguļo notiekošo aksonu mielinēšanu ar oligodendrocītiem, kas veicina neironu vadīšanu un komunikāciju.
Lai gan, analizējot strukturālās pārmaiņas, mazāka uzmanība tiek pievērsta subkortāliem reģioniem, tomēr šajos reģionos, īpaši bazālajos ganglijos, ir vērojamas dažas lielākās pārmaiņas smadzenēs.Sowell et al., 1999Skatīt Fig. 3) un jo īpaši vīriešiem (Giedd et al., 1996). Attīstības izmaiņas strukturālajā tilpumā bazālajos ganglijos un prefrontālajos reģionos ir interesantas, ņemot vērā zināmos attīstības procesus (piemēram, dendrīta arborizāciju, šūnu nāvi, sinaptisko atzarošanu, mielinēšanu), kas notiek bērnībā un pusaudža gados. Šie procesi ļauj precīzi pielāgot un nostiprināt savienojumus starp prefrontālajiem un subortikālajiem reģioniem ar attīstību un mācīšanos, kas var sakrist ar lielāku kognitīvo kontroli. Kā šīs strukturālās izmaiņas attiecas uz izziņas izmaiņām? Vairāki pētījumi ir saistīti ar frontālās daivas strukturālo nobriešanu un kognitīvo funkciju, izmantojot neiropsiholoģiskos un kognitīvos pasākumus (piemēram, Sowell et al., 2003). Konkrētāk, ir ziņots par saistībām starp MRI balstītiem prefrontālo kortikālo un bazālo gangliju reģionālajiem apjomiem un kognitīvās kontroles mērījumiem (ti, spēja ignorēt neatbilstošu reakciju par labu citam vai novērst uzmanību uz neatbilstošu stimulu atribūtu par labu attiecīgajam stimulējošajam atribūtam (Casey, Trainor et al., 1997). Šie atklājumi liecina, ka kognitīvās pārmaiņas atspoguļojas strukturālās smadzeņu pārmaiņās un uzsver subortikālo (bazālo gangliju), kā arī kortikālo (piem., Prefrontālās garozas) attīstību.

DTI pētījumi par cilvēka smadzeņu attīstību
Pārskatītie MRI balstītie morfometrijas pētījumi liecina, ka kortikālie savienojumi tiek pielāgoti, novēršot sinapšu pārpilnību un stiprinot attiecīgos sakarus ar attīstību un pieredzi. Nesenie sasniegumi MRI tehnoloģijā, piemēram, DTI, ir potenciāls līdzeklis, lai detalizētāk pārbaudītu konkrētu balto vielu klātbūtni smadzeņu un uzvedības attīstībā. Atbilstoši šim ziņojumam ir pētījumi par neirofotografēšanu, kas saistījuši šķiedru traktu attīstību ar kognitīvo spēju uzlabojumiem. Konkrētāk, ir parādītas saiknes starp DTI balstītajiem prefrontālās baltās vielas attīstības un kognitīvās kontroles pasākumiem bērniem. Vienā pētījumā šīs jaudas attīstība pozitīvi korelēja ar prefrontālo-parietālo šķiedru traktiem (Nagijs, Vesterbergs un Klingbergs, 2004. gads), kas atbilst funkcionāliem neirofotogrāfijas pētījumiem, kas liecina par šo reģionu atšķirīgu pieņemšanu darbā, salīdzinot ar pieaugušajiem.
Izmantojot līdzīgu pieeju, Liston et al. (2005) ir parādījuši, ka baltās vielas trakti starp prefrontālo-bazālo gangliju un -posta šķiedru traktiem turpina attīstīties bērnībā līdz pieauguša cilvēka vecumam, bet tikai tie tronti starp prefrontālo garozu un bazālo gangliju ir korelēti ar impulsu kontroli, ko mēra ar veiktspēju uz go / nogo uzdevums. Prefrontālās šķiedras trases tika definētas fMRI pētījumā identificēto interešu reģionos, izmantojot to pašu uzdevumu. Gan attīstības DTI pētījumos šķiedru trakta pasākumi tika korelēti ar attīstību, bet konkrētu šķiedru traktu specifika ar kognitīvo veiktspēju tika parādīta, atdalot konkrēto traktu (Liston et al., 2005) vai kognitīvās spējas (Nagy et al., 2004). Šie atklājumi uzsver, ka ir svarīgi pārbaudīt ne tikai reģionālās, bet arī ar shēmu saistītās izmaiņas, kad tiek izteiktas prasības par vecuma atkarīgām kognitīvās attīstības nervu substrātu izmaiņām.
Funkcionālie MRI pētījumi par uzvedības un smadzeņu attīstību
Lai gan strukturālās izmaiņas, kas izmērītas ar MRI un DTI, ir saistītas ar uzvedības izmaiņām attīstības laikā, tiešāka pieeja struktūras un funkcijas asociācijas pārbaudei ir vienlaicīgi izmērīt smadzeņu un uzvedības izmaiņas, tāpat kā fMRI. Spēja izmērīt funkcionālās pārmaiņas jaunattīstības smadzenēs ar MRI ir nozīmīgs attīstības zinātnes potenciāls. Pašreizējā dokumenta kontekstā fMRI nodrošina līdzekli, lai ierobežotu pusaudžu uzvedības interpretācijas. Kā jau minēts iepriekš, tiek uzskatīts, ka prefronta garozas attīstībai ir svarīga nozīme augstāku kognitīvo spēju, piemēram, lēmumu pieņemšanas un kognitīvās kontroles, nobriešanā (Keisija, Totenhema un Fosella 2002b; Casey, Trainor et al., 1997). Lai novērtētu šo spēju neirobioloģisko pamatu, tostarp flanker, Stroop un go / nogo uzdevumus, kopā ar fMRI ir izmantotas daudzas paradigmas.Casey, Castellanos et al., 1997; Keisijs, Džids un Tomass, 2000a; Durston et al., 2003). Kopumā šie pētījumi rāda, ka bērni, veicot šos uzdevumus, pieņem darbā atšķirīgus, bet bieži vien lielākus, difūzākus prefrontālus reģionus, nekā pieaugušajiem. Darbības modelis smadzeņu reģionos, kas ir centrāli uzdevumu veikšanai (ti, kas korelē ar kognitīvo veiktspēju), kļūst vairāk fokusēts vai pielāgots vecumam, bet reģioni, kas nav saistīti ar uzdevuma izpildi, samazinās aktivitātē ar vecumu. Šis modelis ir novērots abos šķērsgriezumos (Brown et al., 2005) un garengriezuma pētījumi (Durston et al., 2006) un dažādās paradigmās. Kaut arī neiromogrāfijas pētījumi nevar pilnībā raksturot šādu attīstības izmaiņu mehānismu (piemēram, dendrīta arborizāciju, sinaptisko atzarošanu), konstatējumi atspoguļo attīstību un uzlabošanu projekcijās uz un no aktivētiem smadzeņu reģioniem ar nobriešanu un liek domāt, ka šīs izmaiņas notiek ilgstoši laika periods (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya un Gabrieli, 2002. gads; Casey, Trainor et al., 1997; Casey et al., 2002a; Crone, Donohue, Honomichl, Wendelken & Bunge, 2006. gads; Luna et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro un Eden, 2003).
Kā šī metodika mums var sniegt informāciju par to, vai pusaudžiem patiešām trūkst pietiekamas kognitīvās kontroles (impulsīva) vai ir riskanti savu izvēli un rīcību? Impulsa kontrole, ko mēra ar kognitīvās kontroles uzdevumiem, piemēram, uzdevuma veikšanas / nogo uzdevums, parāda lineāru attīstības modeli bērnībā un pusaudžā, kā aprakstīts iepriekš. Tomēr nesenie neirofotogrāfijas pētījumi ir sākuši izskatīt ar atalgojumu saistītu apstrādi, kas raksturīga riska uzņemšanai pusaudžiem (Bjork et al., 2004; Ernst et al., 2005; May et al., 2004). Šie pētījumi galvenokārt ir vērsti uz accumbens reģionu, daļu no bazālo gangliju, kas iesaistīti atalgojuma prognozēšanā, nevis šīs reģiona attīstības raksturojumu saistībā ar augšupvērstajiem kontroles reģioniem (prefrontālā garoza). Lai gan nesenais ziņojums par mazāk vēdera priekšdziedzera aktivitāti pusaudžiem salīdzinājumā ar pieaugušajiem monetāro lēmumu pieņemšanas uzdevumā par riska uzņemšanos ir pierādīts (Ešels, Nelsons, Blērs, Pine un Ernsts, 2007. gads).
Kopumā daži pētījumi ir pētījuši, kā mainās apbalvošanas shēmu attīstība subortikālajos reģionos (piemēram, accumbens) saistībā ar kortikālo prefrontālo reģionu attīstību. Turklāt tas, kā šīs nervu pārmaiņas sakrīt ar atalgojuma meklēšanu, impulsivitāti un riska uzņemšanos, joprojām ir relatīvi nezināms. Mūsu neirobioloģiskais modelis liek domāt, ka paaugstinātas reakcijas uz atlīdzību un nenobriedumu kombinācija uzvedības kontroles jomās var novirzīt pusaudžus, lai meklētu tūlītēju, nevis ilgtermiņa ieguvumu, iespējams, izskaidrojot to pieaugumu riskantā lēmumu pieņemšanā un impulsīvā uzvedībā. Subkortikālo (piemēram, accumbens) un kortikālo (piemēram, prefrontālo) lēmumu izstrādes sekošana bērnībā ar pieaugušo vecumu nodrošina papildu ierobežojumus tam, vai pusaudža gados ziņotās izmaiņas ir specifiskas šim attīstības posmam, vai atspoguļo nobriešanu, kas pastāvīgi parādās nedaudz lineārs modelis no bērnības līdz pilngadībai.
Nesen veiktā fMRI pētījuma empīriskie pierādījumi palīdz atbalstīt mūsu neirobioloģisko modeli un ņem pārejas pieeju pusaudža izpratnei, pārbaudot izmaiņas pirms un pēc pusaudža vecuma. Šajā pētījumā (Galvan et al., 2006), mēs pārbaudījām uzvedības un neironu atbildes reakciju uz atalgojuma manipulācijām visā attīstībā, koncentrējoties uz smadzeņu reģioniem, kas saistīti ar atalgojumu saistītu mācīšanos un uzvedību dzīvniekiem (Hikosaka & Watanabe, 2000. gads; Pecina, Kagniard, Berridge, Aldridge un Zhuang, 2003; Schultz, 2006) un pieaugušo attēlveidošanas pētījumi (piemēram, Knutsons, Adamss, Fongs un Hommers, 2001. gads; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) un atkarības pētījumos (Himans un Malenka, 2001. gads; Volkow & Li, 2004. gads). Pamatojoties uz grauzēju modeļiem (Laviola et al., 1999; Spear, 2000) un iepriekšējais attēlveidošanas darbs (Ernst et al., 2005), mēs pieņēmām, ka attiecībā pret bērniem un pieaugušajiem pusaudži uzrāda pārspīlētu akumbēnu aktivizāciju, vienlaicīgi ar mazāk nobriedušu augšupejošu prefrontālās kontroles reģionu pieņemšanu. Nesenais darbs, kas liecina par aizkavētu funkcionālo savienojamību starp šiem prefrontālajiem un limbiskajiem subkortikālajiem reģioniem pusaudža gados salīdzinājumā ar pieaugušajiem, nodrošina mehānismu šo reģionu augšupējas kontroles trūkumam (Hare et al., 2007a).
Mūsu rezultāti atbilst grauzēju modeļiem (Laviola, Makri, Morlijs-Flečers un Adriani, 2003) un iepriekšējie attēlveidošanas pētījumi (Ernst et al., 2005), kas liecina par pastiprinātu akumbēnu darbību, lai gūtu atalgojumu pusaudža laikā. Patiešām, attiecībā pret bērniem un pieaugušajiem pusaudži uzrādīja pārspīlētu atbildes reakciju, gaidot atalgojumu. Tomēr gan bērni, gan pusaudži bija mazāk nobrieduši prefrontālās kontroles reģionos nekā pieaugušie. Šie atklājumi liecina, ka šo reģionu attīstības trajektorijas var būt par pamatu akumbēnu aktivitātes uzlabošanai attiecībā pret bērniem vai pieaugušajiem, kas savukārt var attiekties uz paaugstinātu impulsu un riskantu uzvedību, kas novērota šajā attīstības periodā (skatīt Fig. 4).

Dažos attīstības fMRI pētījumos ir ziņots par prefrontālo un subortikālo reģionu diferenciālu pieņemšanu darbā.Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Parasti šie secinājumi ir interpretēti kā nenobrieduši prefrontālie reģioni, nevis nelīdzsvarotība starp prefrontālo un subkortālo reģionālo attīstību. Ņemot vērā prefrontālo reģionu pierādījumus, lai vadītu piemērotus pasākumus dažādos kontekstos (Millers un Koens, 2001. gads) nenobriedis prefrontālais darbs var kavēt nākotnes rezultātu pienācīgu novērtēšanu un riskantu izvēļu novērtēšanu, un tādējādi tas varētu būt mazāk ietekmīgs attiecībā uz atalgojuma novērtēšanu, nekā to nosaka. Šis modelis atbilst iepriekšējiem pētījumiem, kas liecina par paaugstinātu subkortikālo iedarbību, salīdzinot ar kortikālo aktivitāti, kad lēmumi ir neobjektīvi tūlītēji ilgtermiņā (McClure, Laibson, Loewenstein un Cohen, 2004). Turklāt fumbrija aktivitāte ir pierādīta ar pozitīvu korelāciju ar turpmākiem riska uzņemšanās paradumiem (Kuhnen & Knutson, 2005. gads). Pusaudža vecumā, salīdzinot ar bērnību vai pieaugušo vecumu, nenobriedis ventrālā prefrontālā garoza var nenodrošināt pietiekamu lejupvērstu vadāmu spēcīgi aktivizētu atalgojuma apstrādes reģionu (piemēram, accumbens) kontroli, kā rezultātā prefrontālās sistēmas (orbitofrontālās garozas) mazāk ietekmē attiecībā pret atalgojumu novērtējums.
Kāpēc smadzenes būtu jāplāno tā, lai tās attīstītos?
Pusaudža vecums ir pārejas periods starp bērnību un pieaugušo vecumu, kas bieži vien notiek ar pubertāti. Pubertāte norāda uz seksuālās nobriešanas sākumu (Greibers un Brūkss-Ganns, 1998. gads) un to var definēt ar bioloģiskiem marķieriem. Pusaudža vecumu var raksturot kā pakāpenisku pāreju uz pieaugušo vecumu ar miglainu ontogenētisko laika gaitu (Spear, 2000). Evolucionāri runājot, pusaudža vecums ir periods, kurā tiek iegūtas neatkarības prasmes, lai palielinātu panākumus, atdaloties no ģimenes aizsardzības, lai gan palielinās izredzes uz kaitīgiem apstākļiem (piemēram, traumas, depresija, trauksme, narkotiku lietošana un atkarība).Kellija, Šošeta un Landrija, 2004. gads). Neatkarības meklējumi ir izplatīti dažādās sugās, piemēram, vienaudžu vērstas sociālās mijiedarbības palielināšanās un jaunrades un riska uzņemšanās paradumu pastiprināšanās. Psihosociālie faktori ietekmē pusaudžu tieksmi uz riskantu uzvedību. Tomēr riskanta uzvedība ir bioloģiski virzītas nelīdzsvarotības rezultāts starp paaugstinātu novitāti un sajūtu meklēšanu saistībā ar nenobriedušu “pašregulācijas kompetenci” (Steinberg, 2004). Mūsu neirobioloģiskie dati liecina, ka tas notiek, diferencējot šīs divas sistēmas (limbisko un kontroles).
Spekulācija liek domāt, ka šis attīstības modelis ir evolūcijas iezīme. Jums ir jāiesaistās augsta riska uzvedībā, lai atstātu savu ģimeni un ciemu, lai atrastu mate un riska uzņemšanos, tajā pašā laikā, kad hormoni vada pusaudžus, lai meklētu seksuālos partnerus. Mūsdienu sabiedrībā, kad pusaudža vecums var ilgt neierobežoti, ar bērniem, kas dzīvo kopā ar vecākiem un kuriem ir finansiāla atkarība un kas vēlāk izvēlas biedrus, šī attīstība var tikt uzskatīta par nepiemērotu.
Pastāv pierādījumi, kas liecina, ka pusaudžu gados paaugstināta novatorisma un riska uzņemšanās. Tāda paša vecuma vienaudžu meklējumi un cīņa ar vecākiem, kas palīdz pusaudžiem izkļūt no mītnes teritorijas pārošanai, ir redzama citās sugās, tostarp grauzējiem, primātiem, kas nav cilvēkveidīgie primāti, un dažiem putniem (Spear, 2000). Salīdzinot ar pieaugušajiem, periadolescentās žurkas uzrāda lielāku novitāti vērstu uzvedību brīvas izvēles jaunumu paradigmā (Laviola et al., 1999). Neiroķīmiskie pierādījumi liecina, ka pusaudžu smadzeņu līdzsvars starp kortikālo un subortikālo dopamīna sistēmu sāk pāriet uz lielāku kortikālo dopamīna līmeni pusaudža gados (Spear, 2000). Līdzīga ilgstoša dopamīnerģiskā enervācija ar pusaudžu vecumu līdz pieaugušo vecumam ir parādīta arī cilvēka primāta prefrontālā garozā (Rozenbergs un Lūiss, 1995. gads). Līdz ar to, šķiet, ka šī paaugstinātā šķietamā riska uzņemšanās ir dažāda veida un tiem ir svarīgi adaptīvie mērķi.
Bioloģiskās nosliece, attīstība un risks
Individuālas atšķirības impulsu kontrolē un risku uzņemšanā psiholoģijā jau kādu laiku ir atzītas (Benthins, slovāks un Seversons, 1993. gads). Iespējams, viens no klasiskajiem individuālo atšķirību piemēriem, kas šajās prasmēs atspoguļotas sociālajā, kognitīvajā un attīstības psiholoģijas literatūrā, ir apmierinātības kavēšanās (Mischel, Shoda un Rodriguez, 1989). Atalgojuma aizkavēšanās parasti tiek vērtēta 3-4 gadus veciem mazuļiem. Bērnam jautā, vai viņi dod priekšroku nelielam atalgojumam (vienam sīkfailam) vai lielam atalgojumam (diviem sīkfailiem). Tad bērnam tiek paziņots, ka eksperimentētājs atstās istabu, lai sagatavotos gaidāmajām aktivitātēm, un paskaidro bērnam, ka, ja viņa paliks savā sēdeklī un nepēdīs sīkdatni, viņa saņems lielu atalgojumu. Ja bērns nedara vai nevar gaidīt, viņai jāzvana zvans, lai izsauktu eksperimentētāju un tādējādi saņemtu mazāku atalgojumu. Pēc tam, kad bērns saprot šo uzdevumu, viņa sēž pie galda ar divām balvām un zvanu. Trauksme telpā ir samazināta līdz minimumam, bez rotaļlietām, grāmatām vai attēliem. Eksperts atgriežas pēc 15 min vai pēc tam, kad bērns ir nociris zvanu, ēdis atlīdzību vai parādījis jebkādas briesmu pazīmes. Mischel parādīja, ka bērni parasti uzvedas vienā no diviem veidiem: (1) vai nu gandrīz tūlīt zvana zvans, lai saņemtu sīkfailu, kas nozīmē, ka viņi saņem tikai vienu; vai (2) viņi gaida un optimizē savus ieguvumus un saņem abus sīkfailus. Šis novērojums liek domāt, ka daži indivīdi ir labāki nekā citi, kas spēj kontrolēt impulsus, ņemot vērā ļoti nozīmīgus stimulus, un šo aizspriedumu var atklāt agrīnā bērnībā (Mischel et al., 1989) un tie, šķiet, paliek pāri pusaudža vecumam un jauniešiem (Eigsti et al., 2006).
Kas varētu izskaidrot individuālās atšķirības optimālā lēmumu pieņemšanā un uzvedībā? Daži teorētiķi ir apgalvojuši, ka dopamīnerģiskā mezolimbiskā shēma, kas saistīta ar atalgojuma apstrādi, ir pamatā riskantai uzvedībai. Atsevišķas atšķirības šajā shēmā, piemēram, aleliskie varianti ar dopamīnu saistītos gēnos, kas izraisa pārāk maz vai pārāk daudz dopamīna subkortikālos reģionos, var attiekties uz tendenci iesaistīties riskantā uzvedībā (O'Doherty, 2004). Ir pierādīts, ka kodols accumbens palielina aktivitāti tieši pirms riska riska paradigmu izvēles.Kuhnen & Knutson, 2005. gads; Matthews et al., 2004; Montāga un Berns, 2002. gads) un kā aprakstīts iepriekš, pusaudžiem ir pārspīlēta akumbens aktivitāte, lai atalgotu rezultātus attiecībā uz bērniem vai pieaugušajiem (Ernst et al., 2005; Galvan et al., 2006). Kopumā šie dati liecina, ka pusaudži var būt vairāk pakļauti riskantām izvēlei kā grupai (Dārznieks un Šteinbergs, 2005. gads), bet daži pusaudži būs vairāk pakļauti riskantajai uzvedībai nekā citi, liekot tiem potenciāli lielāku risku negatīviem rezultātiem. Tādēļ ir svarīgi ņemt vērā individuālo mainīgumu, pārbaudot sarežģītas smadzeņu uzvedības attiecības, kas saistītas ar riska uzņemšanos un atalgojuma apstrādi attīstības populācijās.
Lai izpētītu individuālas atšķirības riska uzņemšanās uzvedībā, Galvan et al. (2007) nesen pārbaudīja saistību starp darbību, kas saistīta ar atalgojuma neironu ķēdēm, gaidot lielu naudas atalgojumu ar personības iezīmēm, kas saistītas ar riska uzņemšanos un impulsivitāti pusaudža gados. 7 un 29 gadu vecumā indivīdiem tika iegūtas funkcionālas magnētiskās rezonanses attēlveidošanas un anonīmas pašvērtējuma skalas riskantai uzvedībai, riska uztverei un impulsivitātei. Bija pozitīva saistība starp accumbens darbību un iespējamību iesaistīties riskantā uzvedībā visā attīstībā. Šī aktivitāte mainījās atkarībā no indivīdu vērtējuma par šādas rīcības paredzamajām pozitīvajām vai negatīvajām sekām. Tie cilvēki, kuri uztvēra riskantu uzvedību, izraisot drausmīgas sekas, aktivizēja akumbenus mazāk, lai atalgotu. Šo asociāciju lielā mērā noteica bērni, un pieaugušie novērtēja šādas rīcības sekas pēc iespējas. Impulsiju vērtējumi nebija saistīti ar akumbēnu darbību, bet gan ar vecumu. Šie rezultāti liecina, ka pusaudža gados daži cilvēki var būt vairāk pakļauti riskantai uzvedībai attīstības izmaiņu dēļ, ņemot vērā mainīgo indivīda noslieci iesaistīties riskantā uzvedībā, nevis vienkārši mainīt impulsivitāti (skat. Fig. 5).

Pusaudžu uzvedība ir vairākkārt raksturota kā impulsīva un riskanta (Steinberg, 2004, 2007), tomēr šī attēlveidošanas literatūras pārskatīšana liecina par dažādiem neirobioloģiskiem substrātiem un dažādām attīstības tendencēm šīm uzvedībām. Konkrēti, impulsivitāte ir saistīta ar nenobriedušu vēdera prefrontālo attīstību un pakāpeniski samazinās no bērnības līdz pieauguša cilvēka vecumam (Casey, Galvan et al., 2005). Negatīvā korelācija starp impulsivitātes reitingiem un vecumu pētījumā Galvan et al. (2007) turklāt atbalsta šo jēdzienu. Turpretim riska uzņemšanās ir saistīta ar akumbēnu darbības pieaugumu (Kuhnen & Knutson, 2005. gads; Matthews et al., 2004; Montāga un Berns, 2002. gads), kas pusaudžiem ir pārspīlēts attiecībā pret bērniem un pieaugušajiem (\ tErnst et al., 2005; Galvan et al., 2006). Līdz ar to pusaudžu izvēli un uzvedību nevar izskaidrot tikai ar prefrontālās garozas impulsivitāti vai ilgstošu attīstību, jo pēc tam tiek prognozēts, ka bērni būs lielāki. Rezultāti sniedz neironu pamatu tam, kāpēc dažiem pusaudžiem ir lielāks risks nekā citiem, bet sniedz pamatu tam, kā pusaudžu uzvedība atšķiras no bērniem un pieaugušajiem riska uzņemšanā.
Kopumā šie dati liecina, ka, lai gan pusaudži kā grupa tiek uzskatīti par riska ņēmējiem (Dārznieks un Šteinbergs, 2005. gads), daži pusaudži būs vairāk pakļauti riskantajai uzvedībai nekā citi, liekot viņiem potenciāli lielāku risku negatīviem rezultātiem. Šie atklājumi uzsver, cik svarīgi ir ņemt vērā individuālo mainīgumu, pārbaudot sarežģītas smadzeņu uzvedības attiecības, kas saistītas ar riska uzņemšanos un atlīdzības apstrādi attīstības populācijās. Turklāt šīs individuālās un attīstības atšķirības var palīdzēt izskaidrot neaizsargātību dažiem indivīdiem attiecībā uz risku, kas saistīts ar vielas lietošanu, un galu galā, atkarību.
secinājumi
Cilvēka attēlveidošanas pētījumi rāda strukturālas un funkcionālas izmaiņas frontostriatālajos reģionos (Giedd et al., 1996, 1999; Jernigan et al., 1991; Sowell et al., 1999; pārskatīšanai, Casey, Galvan et al., 2005), kas, šķiet, paralēli palielina izziņas kontroli un pašregulāciju (\ tCasey, Trainor et al., 1997; Luna un Svīnijs, 2004. gads; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004; Skatīt arī Steinberg, 2008, šis jautājums). Šķiet, ka šīs izmaiņas parāda pāreju prefrontālo reģionu aktivizēšanā no difūzas uz vairāk fokusa rekrutēšanu laika gaitā (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor et al., 1997; Durston et al., 2006; Moses et al., 2002) un paaugstinātu subkortikālo reģionu pieņemšanu pusaudža gados (\ tCasey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Kaut arī neiromogrāfijas pētījumi nevar pilnībā raksturot šādu attīstības izmaiņu mehānismu, šīs apjoma un struktūras izmaiņas var atspoguļot attīstību un uzlabošanu projekcijās uz un no šiem smadzeņu reģioniem nogatavināšanas laikā, kas liecina par sistēmas precizēšanu ar attīstību.
Kopumā šeit iegūtie rezultāti liecina, ka pusaudža vecumā palielināta riska uzņemšanās uzvedība ir saistīta ar dažādām subortikālo izklaides un kortikālo kontroles reģionu attīstības trajektorijām. Šīs attīstības izmaiņas var saasināt individuālās atalgojuma sistēmu darbības atšķirības. Lai gan pusaudža vecums ir atšķirīgs kā periods, ko raksturo atalgojuma meklējumi un riska uzņemšanās (Dārznieks un Šteinbergs, 2005. gads; Spear, 2000) individuālās atšķirības neironu reakcijās uz atalgojumu, dod priekšroku dažiem pusaudžiem uzņemties vairāk risku nekā citi, liekot viņiem lielāku risku negatīviem rezultātiem. Šie atklājumi sniedz būtisku pamatu, sintezējot dažādus secinājumus, kas saistīti ar riska uzņemšanos pusaudža vecumā, kā arī izpratni par individuālajām atšķirībām un attīstības marķieriem, lai noteiktu negatīvas uzvedības spējas.
Pateicības
Šo darbu daļēji atbalstīja Zāļu ļaunprātīgas izmantošanas valsts institūts R01 DA18879 un Nacionālais garīgās veselības institūts 1P50 MH62196.
Atsauces
- Benthin A, Slovic P, Severson H. Psihometriskais pētījums par pusaudžu riska uztveri. Pusaudža žurnāls. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Stimulējoša smadzeņu aktivācija pusaudžiem: līdzības un atšķirības no jauniešiem. Neiroloģijas žurnāls. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Konceptuālo zināšanu loma organizācijas attīstībā bērnu atmiņā. In: Brainerd CJ, Pressley M, redaktori. Atmiņas attīstības pamatprocesi: progress kognitīvās attīstības pētījumos. Ņujorka: Springer-Verlag; 1985. lpp. 103 – 142.
- Bjorkland DF. Kā vecuma izmaiņas zināšanu bāzē veicina bērnu atmiņas attīstību: interpretācijas pārskats. Attīstības pārskats. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenesis rēzus pērtiķu prefrontālajā garozā. Smadzeņu garoza. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Atmiņas neatkarība un atmiņas traucējumi izziņas attīstībā. Psiholoģiskais pārskats. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Attīstības izmaiņas cilvēka smadzeņu funkcionālajā organizācijā vārda ģenerēšanai. Smadzeņu garoza. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Nenobriedušas frontālās daivas iemaksas kognitīvajā kontrolē bērniem: fMRI pierādījumi. Neirons. 2002;33: 301-311. [PubMed]
- R. gadījums. Neo-piatrijas kapacitātes konstrukcijas apstiprināšana. Bērnu psiholoģijas eksperiments. 1972;14: 287-302.
- Lieta R. Intelektuālā attīstība: dzimšana līdz pilngadībai. Ņujorka: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. un citi. Labās frontostriatālās shēmas ietekme uz reakcijas inhibīciju un uzmanības deficīta / hiperaktivitātes traucējumiem. Amerikas Bērnu un pusaudžu psihiatrijas akadēmijas žurnāls. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Izmaiņas smadzeņu funkcionālajā organizācijā kognitīvās attīstības laikā. Pašreizējais atzinums neirobioloģijā. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Strukturālā un funkcionālā smadzeņu attīstība un tās saistība ar izziņas attīstību. Bioloģiskā psiholoģija. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Striatīvas un hipokampusa funkcijas atdalīšana ar stimulēšanas reakcijas saderības uzdevumu. Neiroloģijas žurnāls. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Velsa TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Atbildes konflikta sadalīšana, uzmanības atlase un paredzamība ar funkcionālo magnētiskās rezonanses attēlu. Nacionālās Zinātņu akadēmijas darbi. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Klīniskās, attēlveidošanas, bojājumu un ģenētiskās pieejas uz kognitīvās kontroles modeli. Attīstības psihobioloģija. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Attīstošās smadzeņu attēlveidošana: Ko mēs esam uzzinājuši par kognitīvo attīstību? Kognitīvās zinātnes tendences. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Attīstošs funkcionāls MRI pētījums par prefronta aktivizāciju, veicot uzdevumu. Kognitīvās neiroloģijas žurnāls. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Brain reģioni, kas veicina elastīgu noteikumu izmantošanu attīstības laikā. Neiroloģijas žurnāls. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Izturība pret traucējumiem: attīstības izmaiņas pamata apstrādes mehānismā. In: Howe ML, Pasnak R, redaktori. Jaunās tēmas kognitīvajā attīstībā Sējums 1: pamati. Ņujorka: Springer; 1993. lpp. 3 – 27.
- Diamond A. Attīstīt spēju izmantot atsaukšanu, lai vadītu rīcību, kā norāda zīdaiņu sniegums AB. Bērna attīstība. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Konflikta un reaģēšanas konkurences parametru manipulācija, izmantojot ātru jauktu izmēģinājumu notikumu fMRI. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, et al. Pāreja no difūzas uz centrālo kortikālo darbību ar attīstību. Attīstības zinātne. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Attīstošā cilvēka smadzeņu anatomiskā MRI: ko mēs esam iemācījušies? ASV Bērnu pusaudžu psihiatrijas akadēmijas žurnāls. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. Jauniešu riska uzvedības uzraudzība - ASV, 2005, uzraudzības kopsavilkumi. Nedēļas ziņojums par saslimstību un mirstību. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, et al. Prognozējot kognitīvo kontroli no pirmsskolas līdz vēlai pusaudža vecumam un jauniešiem. Psiholoģiskā zinātne. 2006;17: 478-484. [PubMed]
- Ernsts M, Nelsons EE, Jazbecs S, McClure EB, Monk CS, Leibenlufts E, et al. Amygdala un nucleus accumbens reaģē uz guvumu saņemšanu un izlaišanu pieaugušajiem un pusaudžiem. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neironu substrātu izvēle pēc izvēles pieaugušajiem un pusaudžiem: Ventrolaterālo prefrontālo un priekšējo cingulējošo spuru veidošanās. Neuropsychologia. 2007;45: 1270-1279. [PMC bezmaksas raksts] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontāna verbāla mēģinājums atmiņas uzdevumā kā vecuma funkcija. Bērna attīstība. 1966;37: 283-299. [PubMed]
- Galvans A, Hare TA, Parra CE, Penss J, Voss H, Glover G, et al. Agrāka akumbēnu attīstība saistībā ar orbitofrontālo garozu varētu būt pamatā riska uzņemšanās paradumiem pusaudžiem. Neiroloģijas žurnāls. 2006;26: 6885-6892. [PubMed]
- Galvāns A, Harē T, Voss H, Glover G, Casey BJ. Riska uzņemšanās un pusaudžu smadzenes: Kas ir apdraudēts? Attīstības zinātne. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Peer ietekme uz riska uzņemšanos, riska izvēli un riskantu lēmumu pieņemšanu pusaudža un pieaugušo vecumā: eksperimentāls pētījums. Attīstības psiholoģija. 2005;41: 625-635. [PubMed]
- Giedd JN. Pusaudžu smadzeņu strukturālā magnētiskā rezonanse. Ņujorkas Zinātņu akadēmijas Annals. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Smadzeņu attīstība bērnībā un pusaudža gados: ilgstošs MRI pētījums. Nature Neuroscience. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Cilvēka smadzeņu attīstības kvantitatīvā magnētiskā rezonanse: Agn 4 – 18. Smadzeņu garoza. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Cilvēka garozas attīstības dinamiskā kartēšana bērnībā ar agru pieaugušo vecumu. Amerikas Savienoto Valstu Zinātņu akadēmijas darbi. 2004;101: 8174-8179. [PMC bezmaksas raksts] [PubMed]
- Graber JA, Brooks-Gunn J. Pubertāte. In: Blechman EA, Brownell KD, redaktori. Uzvedības medicīna un sievietes ir visaptveroša rokasgrāmata. Ņujorka, Ņujorka: Guilford Press; 1998. lpp. 51 – 58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Pusaudžu smadzenes un potenciālais trauksmes un depresijas risks. 2007a Iesniegts publicēšanai.
- Hare TA, Voss HU, Glover GH, Casey BJ. Konkurence starp prefrontālo un subortikālo limbisko sistēmu pamato emocionālo reaktivitāti pusaudža laikā. 2007b Iesniegts publicēšanai.
- Harnishfeger KK, Bjorkland F. Inhibīcijas mehānismu ontogēnums: atjaunota pieeja kognitīvai attīstībai. In: Howe ML, Pasnek R, redaktori. Kognitīvās attīstības tēmas. Vol. 1. Ņujorka: Springer-Verlag; 1993. lpp. 28 – 49.
- Hikosaka K, Watanabe M. Mērkaņa orbitālās un sānu prefronālās neironu aizkavēšanās aktivitāte, kas atšķiras ar dažādiem ieguvumiem. Smadzeņu garoza. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Synaptic blīvums cilvēka priekšējā garozā - attīstības izmaiņas un novecošanās sekas. Smadzeņu izpēte. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Atkarība un smadzenes: piespiedu neirobioloģija un tās noturība. Dabas apskats Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Magnētiskās rezonanses attēlveidošanas anomālijas lēcu kodolos un smadzeņu garozā šizofrēnijā. Vispārējās psihiatrijas arhīvs. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Garīgās spējas kognitīvās apstrādes komponentu individuālās un attīstības atšķirības. Bērna attīstība. 1978;49: 155-167.
- Kelley AE, Šocets T, Landijs C. Ņujorkas Zinātņu akadēmijas Annals. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Prognozēšana par pieaugošās monetārās atlīdzības saņemšanu selektīvi pieņems darbā kodolu. Neiroloģijas žurnāls. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Finanšu risku uzņemšanas neirālais pamats. Neirons. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psihobioloģiskie riska faktori, kas saistīti ar psihostimulantu neaizsargātību pret pusaudžiem un dzīvnieku modeļiem. Neirozinātnes un uzvedības pārskati. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstrakta riska uzvedība pusaudžu pelēm: psihobioloģiskie faktori un agrīna epigenetiskā ietekme. Neirozinātnes un uzvedības pārskati. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Frontostriatāla mikrostruktūra modulē efektīvu kognitīvās kontroles iegūšanu. Smadzeņu garoza. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Sadarbības smadzeņu funkcijas parādīšanās: FMRI pētījumi par reakcijas inhibīcijas attīstību. Ņujorkas Zinātņu akadēmijas Annals. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, et al. Plaši izplatītas smadzeņu funkcijas nogatavināšana pakļauj izziņas attīstību. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Selektīva kodola accumbens aktivizācija, pieņemot lēmumus par risku. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Maijs JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Ar notikumiem saistīta funkcionāla magnētiskās rezonanses attēlveidošana ar smadzenēm saistītajās smadzeņu shēmās bērniem un pusaudžiem. Bioloģiskā psihiatrija. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Atsevišķas neironu sistēmas novērtē tūlītēju naudas atlīdzību. Zinātne. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Integrālās teorija par prefronālo garozas funkciju. Neiroloģijas gada pārskats. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Bērnu apmierināšanas aizkavēšanās. Zinātne. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Pusaudžu nenobriedums saistībā ar smadzeņu piesaisti emocionālām sejas izteiksmēm. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neiroloģiskā ekonomika un vērtēšanas bioloģiskie substrāti. Neirons. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Bērnu globālās un vietējās apstrādes funkcionālais MRI. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Tagad visi kopā: kad izzūd disociācijas starp zināšanām un rīcību. Psiholoģiskā zinātne. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Balto vielu nogatavināšana ir saistīta ar kognitīvo funkciju attīstību bērnībā. Kognitīvās neiroloģijas žurnāls. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Kopsavilkums par atalgojumu un sodiem cilvēka orbitofrontālajā garozā. Nature Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Atlīdzības prezentācijas un ar atalgojumu saistītas mācības cilvēka smadzenēs: ieskats no neirofotogrāfijas. Pašreizējie viedokļi neirobioloģijā. 2004;14: 769-776.
- Pascual-Leone JA. Matemātiskais modelis pārejai Piaget attīstības stadijās. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hiperdopamīnerģiskās mutācijas pelēm ir augstāks „vēlas”, bet ne “patika”, lai gūtu labumu. Neiroloģijas žurnāls. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Pusaudžu lēmumu pieņemšanas risks un racionalitāte: ietekme uz teoriju, praksi un sabiedrisko kārtību. Psiholoģiskā zinātne sabiedrības interesēs. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Pērles pēc dzemdībām pērtiķu prefrontālās un motora dzeltenās dopamīnerģiskās inervācijas gadījumā: tirozīna hidroksilāzes imūnhistoķīmiskā analīze. Salīdzinošās neiroloģijas žurnāls. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Funkcionālā frontalizācija ar vecumu: neiroloģiskās attīstības trajektoriju kartēšana ar fMRI. Neirozinātnes un uzvedības pārskati. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funkcionālās neuroanatomiskās atšķirības starp pieaugušajiem un skolas vecuma bērniem atsevišķu vārdu apstrādē. Zinātne. 2002;296: 1476-1479. [PubMed]
- Schultz W. Uzvedības teorijas un atalgojuma neirofizioloģija. Gada pārskats par psiholoģiju. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Welcome SE, Henkenius AL, Toga AW. Kartikas izmaiņu kartēšana cilvēka dzīves laikā. Nature Neuroscience. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo pierādījumi par pēcdzemdību smadzeņu nobriešanu frontālajos un striatālajos reģionos. Nature Neuroscience. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Kartes izmaiņas cilvēka garozā visā dzīves laikā. Neirologs. 2004;10: 372-392. [PubMed]
- Spear LP. Pusaudžu smadzeņu un ar vecumu saistītās uzvedības izpausmes. Neirozinātnes un uzvedības pārskati. 2000;24: 417-463. [PubMed]
- Steinberg L. Riska uzņemšanās pusaudža gados: kādas izmaiņas un kāpēc? Ņujorkas Zinātņu akadēmijas Annals. 2004;1021: 51-58. [PubMed]
- Steinberg L. Riska uzņemšanās pusaudža gados: jaunas perspektīvas no smadzeņu un uzvedības zinātnes. Pašreizējie psiholoģijas zinātnes virzieni. 2007;16: 55-59.
- Steinberg L. Sociālā neirozinātnes perspektīva par pusaudžu riska uzņemšanos. Attīstības pārskats. 2008;28: 78-106. [PMC bezmaksas raksts] [PubMed]
- Tamm L, Menon V, Reiss AL. Smadzeņu darbības nobriešana, kas saistīta ar reakcijas inhibīciju. Amerikas Bērnu un pusaudžu psihiatrijas akadēmijas žurnāls. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Pierādījumi par attīstības atšķirībām implanta secībā: FMRI pētījums par bērniem un pieaugušajiem. Kognitīvās neiroloģijas žurnāls. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Ziedi DL, Zeffiro TA, Eden GF. Neironu mehānismu izstrāde lasīšanai. Nature Neuroscience. 2003;6: 767-773.
- Volkow ND, Li TK. Narkotiku atkarība: uzvedības neirobioloģija. Dabas apskats Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Emocionālās un kognitīvās izmaiņas pusaudža laikā. Pašreizējais atzinums neirobioloģijā. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Dopamīna pārnešana cilvēka striatumā naudas atalgojuma uzdevumu laikā. Neiroloģijas žurnāls. 2004;24: 4105-4112. [PubMed]