KOMENTĀRI: Lielisks pārskats par gandrīz visiem attiecīgajiem pētījumiem par androgēnu receptoriem, dopamīnu un seksuālo funkciju. Fantastiski zīmē hipotalāmu un atlīdzību ķēžu savstarpējās attiecības.
Horm Behav. 2008 maijs; 53(5): 647-658.
Publicēts tiešsaistē 2008 February 13. doi: 10.1016 / j.yhbeh.2008.01.010
Anotācija
Pusaudža vecums ir saistīts ar baudas meklētāju uzvedības palielināšanos, ko, savukārt, ietekmē hipotalāma-hipofīzes-gonādas ass pubertālā aktivizēšana. Dzīvnieku modeļos, kur uzvedība ir dabiski izdevīga, piemēram, dzimumam, sēklinieku androgēni veicina tēviņu uzvedības veidošanos un izpausmi. Lai panāktu uzvedības nobriešanu, pusaudža gados smadzenēs notiek nozīmīga pārveidošanās, un daudzas izmaiņas ir arī jutīgas pret androgēniem, domājams, ka tās darbojas caur androgēnu receptoriem (AR). Ņemot vērā dzimumdziedzeru hormonu un smadzeņu attīstības delikāto mijiedarbību, nav pārsteigums, ka hormonu līmeņa traucējumi šajā jutīgajā periodā ievērojami maina pusaudžu un pieaugušo uzvedību. Vīriešu kāmjiem normāla pieaugušā seksuālās izturēšanās izpausme ir pakļauta testosterona iedarbībai pusaudža gados. Tēviņiem, kuriem pubertātes laikā atņemti androgēni, ir novērojams ilgstošs pārošanās deficīts. Un otrādi, ar androgēniem vien nepietiek, lai izraisītu pārošanos vīriešiem pirmspubertālē, pat ja smadzeņu AR ir pirms pubertātes. Šajā kontekstā nopietnas bažas rada plaša anabolisko-androgēno steroīdu (AAS) lietošana pusaudža gados. AAS ļaunprātīga izmantošana var mainīt gan laiku, gan androgēnu līmeni pusaudžu vīriešiem. Kāmjiem pusaudžu AAS iedarbība palielina agresiju un izraisa ilgstošas izmaiņas neirotransmiteru sistēmās. Turklāt AAS pašas par sevi pastiprina, kā liecina testosterona un citu AAS pašpārvalde. Tomēr jaunākie pierādījumi liecina, ka androgēnu pastiprinošajai iedarbībai var nebūt nepieciešams klasiskais AR. Tāpēc, lai labāk izprastu AAS ļaunprātīgu izmantošanu, nepieciešama papildu izpēte par mijiedarbību starp androgēniem un atlīdzinošu izturēšanos pusaudža smadzenēs.
Pārskats
Pusaudža gados pamodina smadzenes gan priekam, gan riskam. Pusaudžiem tas bieži notiek eksperimentos ar narkotikām un seksu. Amerikas Savienotajās Valstīs vīriešu pirmā dzimumakta vidējais vecums ir 16.4 gadi, un 65% dzimumakta bija bijuši 12th pakāpe (Ķeizara ģimenes fonds, 2005). Tāpat šajā populācijā ir visaugstākais nelegālo narkotiku lietošanas rādītājs Amerikas Savienotajās Valstīs. Saskaņā ar 2004 nacionālo aptauju par narkotiku lietošanu un veselību, 38% vīriešu vecumā no 18 – 25 pēdējā gada laikā ir lietojuši nelikumīgas narkotikas (SAMHSA / OAS, 2005). Turklāt 31% pusaudžu zēnu pēdējās seksuālās saskarsmes laikā lietoja narkotikas vai alkoholu (Ķeizara ģimenes fonds, 2005). Turklāt pusaudža vecums ir nozīmīgs laiks noteiktu psihopatoloģiju etioloģijā, piemēram, depresija, nemiers, nesakārtota ēšana un uzvedības traucējumi. Mēs uzskatām, ka dzimumdziedzeru hormonu izdalīšana no pubertātes, smadzeņu steroīdu receptoru aktivizēšana un hormonu mijiedarbība ar pieredzi pusaudžu smadzeņu attīstībā veicina pusaudža gados novērotās uzvedības izmaiņas.
Mūsu mērķis šeit ir pārskatīt pierādījumus tam, ka dzimumdziedzeru androgēni meditē motivētas izturēšanās pusaudža nobriešanu un pieaugušo sniegumu, kā arī šīs izturēšanās labvēlīgās īpašības. Mēs arī iesniedzam pierādījumus, ka testosterons pats par sevi atalgo, kas, iespējams, veicina motivētas uzvedības nobriešanas izmaiņas pusaudža gados, kad testosterona līmenis strauji paaugstinās. Šī darba uzmanības centrā ir mūsu nervu ķēžu pētījumi, kas ir vīriešu seksuālās izturēšanās pamatā, īpaši Sīrijas kāmī, īpaši uzsverot mijiedarbību starp testosteronu un dopamīnu (DA). Mēs ierosinām, ka pubertāles androgēniem ir gan īslaicīga, gan ilgtermiņa ietekme uz atalgojuma shēmām un motivētu uzvedību. Mēs arī izvirzām hipotēzi, ka papildināšana ar eksogēniem androgēniem anabolisko androgēnu steroīdu (AAS) veidā palielina normālu pubertāles androgēnu iedarbību, tādējādi nelabvēlīgi ietekmējot pusaudža smadzeņu attīstību un uzvedību.
Pusaudža vecums kā jutīgs smadzeņu attīstības periods
Galu galā smadzenes ir gan ierosinātājs, gan mērķis androgēnu darbībai pusaudža gados. Jauniem zēniem (<12 gadi) un jauniem kāmjiem (<28 dienu vecumam) cirkulējošie androgēni un gonadotropīni ir bazālā līmenī. Tā kā luteinizējošā hormona sekrēcija no hipofīzes priekšējās daļas palielinās, reaģējot uz hipotalāma gonadotropīnu atbrīvojošo hormonu, cirkulējošā testosterona koncentrācija ievērojami palielinās. Tas notiek Tanner II / III stadijā (14 gadi) zēniem un līdz 28 dienu vecumam kāmjiem. Brīdī, kad zēni sasniedz Tannera IV / V stadiju (apmēram 16 gadu vecumā) vai kad kāmji ir 50–60 dienu veci, endogēnais testosterons ir pieaugušo vīriešu diapazonā. Pubertālā hormona sekrēcija sakrīt ar pusaudža periodu, kas cilvēkiem notiek apmēram no 12 līdz 20 gadu vecumam. Pubertātes hormoni ietekmē ne tikai perifēros audus, izraisot sekundāras dzimuma pazīmes, kas ir acīmredzamas pubertātes pazīmes, bet arī centrāli, lai ietekmētu gan pusaudžu smadzeņu pārveidošanu, gan uzvedības nobriešanu. Turklāt pubertātes hormonu izraisītās fizioloģiskās un neiroloģiskās izmaiņas izraisa būtiskas izmaiņas indivīda pieredzē, kas pats par sevi var būtiski mainīt smadzeņu attīstības gaitu. Tādējādi dzimumsteroīdu hormonu pubertātes pieaugums, ko nosaka reproduktīvās neiroendokrīnās ass attīstības nobriešana, savukārt pusaudžu uzvedības attīstību ietekmē gan tiešā, gan netiešajā nervu sistēmas ietekmē.
Cilvēka pusaudža vecums tagad tiek atzīts par galveno un dinamisko neironu attīstības periodu, kura laikā tiek pārveidotas un pilnveidotas uzvedības shēmas. Kaut arī 5 gadu veca bērna smadzenes jau ir 90% no tā pieaugušā lieluma (Dekaban, 1978), nozīmīga rekonstrukcija vēl ir jāveic. Šis jēdziens tika izveidots ar pētījumiem gan cilvēkiem, gan dzīvniekiem, dokumentējot, ka daudzi no attīstības pamata procesiem, kas notiek smadzeņu perinatālās attīstības laikā, tiek atkārtoti apkopoti pusaudža gados. Šajos procesos ietilpst neiroģenēze (Eckenhoff un Rakic, 1988; Viņš un apkalpes, 2007; Pinos, Collado, Rodriguez-Zafra, Rodriguez, Segovia un Guillamon, 2001; Rankin, Partlow, McCurdy, Giles un Fisher, 2003), ieprogrammēta šūnu nāve (Nunez, Lauschke un Juraska, 2001; Nunez, Sodhi un Juraska, 2002), dendrītisko arborāciju un sinapsu izstrāde un atzarošana (Andersens, Rutšteins, Benzo, Hosteters un Teišers, 1997; Huttenlocher un Dabholkar, 1997; Lenroot un Giedd, 2006; Sovels, Tompsons, Leonards, Welcome, Kan un Toga, 2004), mielinēšana (Benes, Turtle, Khan un Farol, 1994; Pauss, Kolinss, Evanss, Leonards, Pikē un Ziddenboss, 2001; Sowell, Thompson, Tessner un Toga, 2001) un seksuālo diferenciāciju (Chung, De Vries un Swaab, 2002; Deiviss, Šrīne un Gorski, 1996; Nunez et al., 2001). Tādējādi pēcdzemdību smadzeņu attīstības trajektorija nav lineāra, bet to drīzāk raksturo pusaudža strauju pārmaiņu pārrāvums un ietver gan progresīvus, gan regresīvus notikumus. Kā zina jebkurš attīstības biologs, strauju attīstības pārmaiņu periodi signalizē par paaugstinātu jutīgumu un neaizsargātību gan pret no pieredzes atkarīgām izmaiņām, gan pret pārmērīgas satraukuma un apvainojuma nelabvēlīgajām sekām, un nav pamata domāt, ka cilvēka pusaudža smadzeņu attīstība ir izņēmums (Andersen, 2003; Spear, 2000). Tādējādi tiek prognozēts, ka pubertālas hormonu ietekmes uz pusaudža smadzenēm laikā radītajām perturbācijām būs ilgstošas sekas pieaugušo uzvedībai.
Androgēni un neironu shēmas motivētai uzvedībai
Tā kā pusaudža vecums ir pārejoša un dinamiska attīstības fāze, pusaudža smadzenes un uzvedību būtu grūti novērtēt izolēti. Tā vietā, lai novērtētu pusaudža unikālo raksturu, ir noderīgi to pretstatīt nobriedušu pieaugušo cilvēku smadzenēm un uzvedībai. Tādējādi, pievēršoties šajā dokumentā vīriešu seksuālajai uzvedībai un atlīdzībai, šeit ir svarīgi ieviest pieaugušo vīriešu neironu ķēdes kopulācijai un seksuālajai motivācijai, ieskaitot gonādu steroīdu hormonu lomu uzvedības aktivizēšanā un androgēnu receptoru sadalījumā. (AR) un estrogēni (ER).
AR ir sastopamas šūnu grupās, kas veido neironu ķēdes, kas nodrošina atalgojošu sociālo uzvedību, piemēram, seksu. Turklāt smadzeņu AR tiek izteikti pirms pubertātes kāmīšos, un androgēni tos paaugstina gan nepilngadīgiem, gan pieaugušiem vīriešiem (Kashon, Hayes, Shek un Sisk, 1995; Lēnprātība, Romeo, Novaks un Sisk, 1997). Grauzēju smadzenēs AR un ER izplatība ievērojami pārklājas (Wood un Newman, 1995) un aromatāze (Celotti, Negri-Cesi un Poletti, 1997), ieskaitot estrogēna receptoru α un β formas (Shughrue, Lane un Merchenthaler, 1997). Saistoties ar ligandu, “klasiskais” AR un ER darbojas kā transkripcijas faktori, lai ierosinātu jaunu olbaltumvielu transkripciju un sintēzi. Nav pārsteidzoši, ka šie efekti notiek salīdzinoši lēnā laika posmā ar novēlotu darbības sākšanos. Vīriešu kāmju seksuālās izturēšanās ar steroīdiem stimulēšana (Noble un Alsum, 1975) ir saskanīgs ar darbībām, kas tiek veiktas ar klasisko genomu darbību palīdzību. Piemēram, 2 nedēļu laikā jāveic steroīdu iedarbība, lai atjaunotu pārošanos ilgtermiņa kastrātos. Jaunāki pētījumi ar žurkām ir parādījuši arī androgēnu ātru iedarbību uz šūnām smadzeņu reģionos, kuros ir maz klasisko receptoru (Mermelšteins, Bekers un Surmeiers, 1996). Tiek uzskatīts, ka šīs steroīdu darbības ir saistītas ar negenomiskiem receptoriem. Tā kā klasiskā AR un ER izplatība kāmja smadzenēs ir salīdzinoši ierobežota (Koks un Swann, 1999), potenciālie smadzeņu mērķi negenomiskas androgēnu darbībai ir daudz plašāki.
Mediālajai preoptiskajai zonai (MPOA) ir galvenā loma kopulācijā vīriešiem no zelta zivtiņas līdz cilvēkam (pārskatīts Korpuss, Vuds un Makkenna, 2006). Turklāt kāmja MPOA ar bagātīgu AR un ER starpniecību pārraida gonādu steroīdu hormonus, un MPOA testosterona implanti ir pietiekami, lai atjaunotu seksuālo aktivitāti ilgtermiņa kastrātos (Koks un Swann, 1999). Žurku tēviņiem dzimumdziedzeru steroīdi darbojas MPOA, lai regulētu bazālo DA izdalīšanos (Putnam, Sato un Hull, 2003) un stimulē pārošanos (Hull, Du, Lorrain un Matuszewich, 1995). Sākotnēji novērojams neliels DA pieaugums, kad aiz ekrāna tiek parādīta sieviete. Kopulācijas laikā MPOA DA palielinās vēl vairāk (+ 50% no sākotnējā līmeņa), un šim efektam nepieciešami androgēni (Hull et al., 1995; Putnam et al., 2003). Nav pārsteidzoši, ka kastrētiem tēviņiem, kuri nepārajas, MPOA DA nepalielinās (Hull et al., 1995). Šo rezultātu ir nedaudz grūti interpretēt, jo DA izdalīšanās trūkumu ierobežo seksuālās aktivitātes neesamība. Tomēr DA izdalīšanās MPOA korelē ar pārošanās zaudēšanu īstermiņa kastrātos (Hull et al., 1995) un ar testosterona izraisītu seksuālās aktivitātes atjaunošanu ilgstošos kastrātos (Du, Lorrain un Hull, 1998; Putnam, Du, Sato un Hull, 2001).
Grauzēju MPOA testosterona androgēniem un estrogēniem metabolītiem ir īpaša loma pārošanās regulēšanā (Putnam et al., 2003; Putnam, Sato, Riolo un Hull, 2005). Viens no seksuālās motivācijas rādītājiem ir latentums, lai sāktu kopulāciju (piestiprināšanu vai iejaukšanos). Seksuālās aktivitātes latentums ir jutīgs pret estrogēniem, uzturot MPOA slāpekļa oksīda sintāzi, kas, savukārt, uztur bazālo DA līmeni. Ar estrogēniem apstrādāti kastrāti uzrāda augstu bazālo DA līmeni, kas cieši korelē ar spēju uzsākt kopulāciju. Tomēr viņiem neizdodas parādīt sieviešu un kopulācijas izraisītu DA izdalīšanās pieaugumu, kas cieši korelē ar seksuālo sniegumu. Līdz ar to viņu seksuālā veiktspēja ir zemāka par neskarto līmeni. No otras puses, kastrātiem, kas apstrādāti tikai ar aromatizējamiem androgēniem, neuzrāda paaugstinātu bazālo DA līmeni, un tie nespēj uzsākt kopulāciju. Tāpēc normālai seksuālai sniegšanai ir nepieciešami gan estrogēni, gan androgēni. Seksuālo sniegumu parasti izsaka kā stiprinājumu, iejaukšanās un ejakulācijas biežuma mērus. Tikai tad, kad tiek aizstāti gan estrogēni, gan androgēni, kastrētiem tēviņiem ir paaugstināts DA līmenis (un īsāki latences rādītāji), un sieviešu un kopulācijas izraisītais DA pieaugums (un palielināts biežuma rādītājs). Tādā veidā MPOA estrogēni veicina seksuālo motivāciju, bet gan estrogēni, gan androgēni - seksuālo sniegumu.
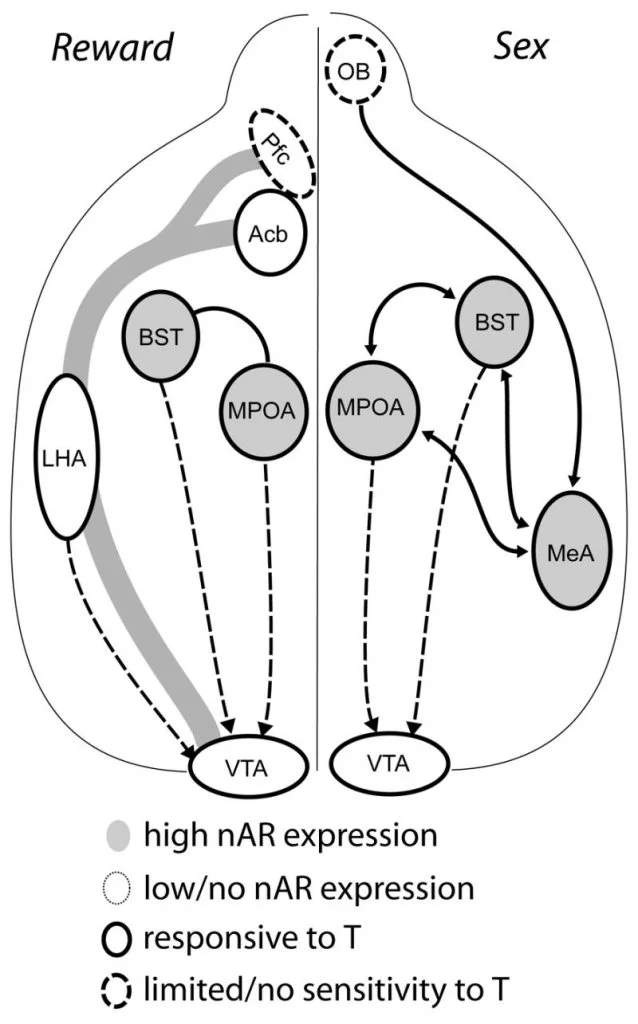
Lai gan testosterons ir nepieciešams MPOA DA atbrīvošanai no vīriešu kopulatīvās uzvedības un pašas pārošanās laikā, ne testosterons, ne pārošanās atsevišķi nevar izraisīt DA MPOA. Tā vietā DA izdalīšanai MPOA ir vajadzīgas arī konkrētu mātīšu ķīmiski jutīgās norādes. Grauzējiem vīriešu seksuālās izturēšanās ierosināšanai primārā sensoro īpašība ir kemosensorie stimuli (Fig. 1). Ķīmiski jutīgās norādes no ožas sīpoliem uz MPOA pārraida caur mediālo amygdaloid kodolu un stria terminalis gultnes kodolu, struktūrām ar bagātīgu AR un ER (Koks un Swann, 1999). Lai noteiktu ķīmiski jutīgo norāžu lomu pārošanās izraisītā DA, mēs izmērījām MPOA DA pārošanās laikā ar dzimumdziedzeriem neskartiem vīriešu kāmjiem ar vienpusēju ožas bulbektomiju (UBx, Triemstra, Nagatani un Wood, 2005). Kaut arī divpusēja ožas sīpolu noņemšana novērš seksuālo aktivitāti un MPOA DA izdalīšanos, vienpusēja bulbektomija netraucē pārošanos. Šajā pētījumā kopulācija izraisīja MPOA DA izdalīšanos, ja to mēra pretēji bojātajai ožas spuldzei, bet ne ipsilaterālā puslodē (Fig. 2). Līdzīgi rezultāti tika novēroti žurku tēviņiem ar vidējās amigdala bojājumiem (Dominguez, Riolo, Xu un Hull, 2001). Saistītā pētījumā mediālas amigdala ķīmiska stimulēšana žurkām izraisīja MPOA DA izdalīšanos, kas līdzvērtīga tai, kas notiek kopulācijas laikā (Dominguez un Hull, 2001). Kopumā šie dati liecina, ka testosterons rada pieļaujamu vidi, kas ļauj ārējiem maņu stimuliem sasniegt MPOA un izraisīt DA izdalīšanos kopulācijas laikā.

Fig. 2
Galu galā seksuālā izturēšanās un citas dabiskas atlīdzības aktivizē neironu atalgojuma ceļus. Mezokortikolimbiskā DA ķēde sastāv no ventrālā pamatvirsmas (VTA), kodolu akumulātiem (Acb) un prefrontālā garozas (Pfc). Dopamīna šūnu ķermeņi, kas dzīvo VTA projektā, virzoties uz ACB un Pfc (Koob un Nestler, 1997). Žurkām DA izdalās Acb seksa laikā (Pfaus, Damsma, Nomikos, Wenkstern, Blaha, Phillips un Fibiger, 1990). Daudzas ļaunprātīgas narkotikas darbojas arī mezolimbiskajā DA sistēmā, lai palielinātu DA izdalīšanos (amfetamīni) vai kavē DA atkārtotu uzņemšanu (kokaīns, Di Chiara un Imperato, 1988), tādējādi pastiprinot to atkarību izraisošās īpašības. Tādā veidā testosterons var ietekmēt DA izdalīšanos Acb gan ar seksuālās uzvedības uzlabošanu, gan ar narkotikām, kuras lieto ļaunprātīgi (skatīt zemāk).
Pašreizējie pierādījumi liecina, ka mezokortikolimbiskā DA sistēma nogatavojas pusaudža gados. Acb DA šķiedru blīvums pusaudža gados krasi palielinās, kas liecina, ka pusaudža periodā notiek ievērojams VTA dopamīnerģisko izvirzījumu nobriešana uz Acb (Lesting, Neddens un Teuchert-Noodt, 2005). Turklāt dopamīnerģisko ievadi GABA (γ-aminosviestskābes) -erģiskajās šūnās žurku mediālajā prefrontālajā garozā bagātina un modulē serotonerģiskās sistēmas pubertātes attīstības laikā (Benes, Taylor un Cunningham, 2000), un manipulācijas ar androgēniem pieaugušām žurkām izraisa dopamīnerģiskā aksona blīvuma izmaiņas prefrontālajā garozā (Kritzer, 2003). Pfc, Acb un VTA ir maz AR vai ER, kaut arī ERβ ir VTA (Shughrue et al., 1997). Tāpēc šķiet iespējams, ka androgēni ietekmē mezokortikolimfisko DA sistēmu, izmantojot androgēniem jutīgus afferentus vai caur ERβ VTA, kā hipotalāmā (Handa et al., Šis jautājums). Mūsu dati rāda, ka androgēnu jutīgās šūnas kāmju tēviņos nonāk VTA no struktūrām, kas saistītas ar uz steroīdiem jutīgu izturēšanos. Piemēram, gan MPOA, gan stria terminalis (BST) gultnes kodols satur lielu skaitu AR pozitīvu šūnu, kas izvirzījas uz VTA (Sato un koks, 2006). Ventrālais pallidums, galvenais Acb efektīvais mērķis (Zahm un Heimer, 1990), satur arī daudzas AR pozitīvas šūnas, kas projicējas uz VTA. Šīs projekcijas dod iespēju androgēniem mainīt mezokortikolimfiskās DA sistēmas aktivitāti.
No steroīdiem atkarīga uzvedības organizācija pusaudža gados
Tradicionālais viedoklis par hormonu darbību pusaudžu uzvedībā balstās uz steroīdu hormonu aktivizācijas efektiem, kas attiecas uz steroīdu spēju atvieglot uzvedību īpašos sociālos apstākļos, veicot darbību mērķa šūnās neironu ķēdēs, kas ir uzvedības pamatā. Aktivizācijas efekti ir īslaicīgi tādā nozīmē, ka tie rodas un iet kopā ar hormona klātbūtni un trūkumu, un parasti tie ir saistīti ar pieaugušo uzvedības izpausmēm. Turpretī organizatoriskā ietekme attiecas uz steroīdu spēju veidot nervu sistēmas struktūru attīstības laikā. Strukturālā organizācija ir pastāvīga, saglabājas arī pēc hormonu iedarbības perioda un nosaka neironu un uzvedības reakcijas uz steroīdiem pieaugušā vecumā. Mūsu izpratne par attīstības saistību starp steroīdu hormonu organizatorisko un aktivizācijas iedarbību ir attīstījusies pēdējos 50 gados. Fēnikss un kolēģi vispirms ierosināja, ka pieaugušo uzvedības (aktivizācijas) reakcijas uz steroīdiem hormoniem ieprogrammē (organizē) steroīdi hormoni maksimāli jutīgā perinatālās attīstības periodā (Fēnikss, Goijs, Geralls un Jauns, 1959). Vēlāk Skots un kolēģi izveidoja teorētisko pamatu vairāku jutīgu periodu pastāvēšanai nervu sistēmas progresējošai organizācijai un atzīmēja, ka jutīgi periodi, visticamāk, notiek strauju attīstības izmaiņu periodos (1974). Pēc tam Arnolds un Bredlovs norādīja, ka smadzeņu organizācija, kas atkarīga no steroīdiem, var notikt ārpus jutīgiem attīstības periodiem (Arnolds un Bredlovs, 1985). Pēdējo 15 gadu laikā veiktie pētījumi, izmantojot dažādus dzīvnieku modeļus un uzvedības sistēmas, skaidri parāda, ka pusaudža smadzenes ir jutīgas gan pret dzimumdziedzeru steroīdu aktivizēšanas, gan organizācijas iedarbību (pārskatīts Sisk un Zehr, 2005). Un, tāpat kā citi strauju attīstības pārmaiņu periodi, pusaudža vecums ir noteikts iespēju loks smadzeņu pārveidošanai, kas atkarīga no steroīdiem.
Mūsu darbs, izmantojot kāmīti kā dzīvnieku modeli, sniedz pierādījumus tam, ka pusaudža gados steroīdi maina vīriešu sociālo uzvedību (Šulcs, Menards, Smits, Albers un Sisks, 2006; Schulz un Sisk, 2006). Pirms pubertātes ārstēšanas ar testosteronu nevar aktivizēt kāmju seksuālo izturēšanos, kas liek domāt, ka vēl nav notikuši nobriešanas procesi, kas padara neironu ķēdes par jutīgām pret aktivizēšanu vai organizēšanu ar steroīdu hormonu palīdzību (Meek et al., 1997; Romeo, Ričardsons un Sisk, 2002a). Un otrādi, lai gan atklātai vīriešu reproduktīvās uzvedības izpausmei pieaugušā vecumā nav absolūti nepieciešama dzimumdziedzeru steroīdu klātbūtne pusaudža gados, maksimālajai uzvedības izpausmei tas ir nepieciešams. Salīdzinot vīrišķo reproduktīvo uzvedību vīriešiem, kas kastrēti pirms pubertātes (NoT@P) vai pēcpubertātes (T@P) un pēc tam pieaugušā vecumā ārstēti ar testosteronu, pirmspubertātes NoT@P kastrātiem ir vismaz 50% vīrišķās uzvedības deficīts, salīdzinot ar vīriešiem, kas kastrēti pēc pusaudža vecuma. (Fig. 3, Schulz, Richardson, Zehr, Osetek, Menard un Sisk, 2004). Turklāt reproduktīvās uzvedības deficīts ir ilgstošs, un to nevar novērst ne ilgstoša testosterona terapija, ne seksuāla pieredze pieaugušā vecumā (Schulz et al., 2004). Līdzīgi, pēc ārstēšanas ar estrogēnu un progesteronu, NoT@P vīriešiem ir īsāks lordozes latentums un ilgāks lordozes ilgums nekā vīriešiem, kas kastrēti kā pieaugušie (Schulz et al., 2004), kas liek domāt, ka pirmspubertāli kastrāti ir mazāk defeminēti nekā vīrieši, kas pakļauti pubertāles testosterona iedarbībai.

Fig. 3
Iespējams, ka NoT@P vīrieši cieš no pazeminātas seksuālās motivācijas. Viens no veidiem, kā risināt šo jautājumu, ir salīdzināt latentuma laiku, lai iesaistītos gan ano-ģenitāliju izmeklēšanā (AGI), gan piestiprināšanā starp vīriešiem, kuriem gonadektomija ir izoperēta pirms (NoT@P) un pēc pubertātes (T@P). Ja seksuālā motivācija ir atkarīga no dzimumdziedzeru hormonu iedarbības pusaudža gados, mēs prognozējam ilgāku latentumu, lai iesaistītos seksuālajā uzvedībā NoT@P vīriešiem. Patiešām, atkārtoti saskaroties ar estrus mātītēm, NoT@P tēviņiem ir nepieciešams ilgāks laiks, lai sāktu AGI un augtu, salīdzinot ar T@P tēviņiem (Fig. 4). Tādējādi, papildus seksuālās veiktspējas aspektu organizēšanai, šķiet, ka pubertātes hormoni organizē arī seksuālās uzvedības atalgojošos aspektus. Lai atbalstītu šo iespēju, DA agonista apomorfīna centrālā ievadīšana pieaugušā vecumā atjauno NoT@P tēviņu uzvedības pieaugumu līdz pieaugušajiem raksturīgajam līmenim, kas liecina, ka testosterons pusaudža gados parasti organizē dopamīnerģiskās nervu ķēdes (Salas-Ramirez, Montalto un Sisk, 2006. ). Tomēr joprojām ir daudz interesantu jautājumu. Vai NoT@P tēviņš nospiestu mātīti ar estrus vai arī izvēlētos nosacītu vietu pārošanās vietai? Turpmākajos pētījumos tiks pētīta pubertātes hormonu loma seksuālās motivācijas un seksuālās veiktspējas organizēšanā.

Fig. 4
Anogenitālās izmeklēšanas (AGI) latentums un ilgums kāmju tēviņiem, kam veikta gonadektomija pirms pubertātes (NoT@P) vai pēc pubertātes (T@P). Visi tēviņi tika sagatavoti ar testosteronu pieaugušā vecumā 7 nedēļas pēc gonadektomijas un vienu nedēļu pirms pirmā uzvedības testa. A. T@P tēviņiem uzrādīja līdzīgu AGI latentumu trijos testos ar estrus mātīti, turpretim NoT@P tēviņiem palielināja AGI latentumu trešā testa laikā ar estrus mātīti. B. T@P tēviņiem trijos uzvedības testos ar estrusu mātīti tika samazināts mount latentums, turpretim noT@P tēviņiem trijos uzvedības testos netika novērotas nekādas izmaiņas mount latentumā. Šie dati liecina, ka pubertātes dzimumdziedzeru hormoniem ir ilgstoša, veicinoša ietekme uz pieaugušo vīriešu motivāciju iesaistīties seksuālā uzvedībā ar sievieti. (Nepublicēti dati no dzīvniekiem Schulz, KM, Richardson, HN, Zehr, JL, Osetek, AJ, Menard, TA un Sisk, CL, 2004).
Prepubertālas uzvedības reakcija uz steroīdiem
Viena no ilgstošajām pusaudžu uzvedības attīstības mīklām ir iemesls, kāpēc reproduktīvās uzvedības aktivizēšana, reaģējot uz steroīdu iedarbību, tiek vājināta pirmspubertālajos vīriešu kāmjos. Ja zems androgēnu līmenis pirms pubertātes ierobežo vīriešu seksuālās izturēšanās izpausmes vīriešiem pirmspubertālē, tad, papildinot endogēnos androgēnus vīriešiem pirmspubertālē, vajadzētu radīt pārošanos. Izrādās, ka tas tā nav (Meek et al., 1997; Romeo, Cook-Wiens, Richardson un Sisk, 2001; Romeo, Vāgners, Jansens, Didrihs un Sisks, 2002b), neskatoties uz to, ka AR un ER skaits un sadalījums visā pārošanās ķēdē ir līdzīgi ar hormoniem ārstētiem pirmspubertāles un pieaugušajiem kastrātiem (Meek et al., 1997; Romeo, Diedrich un Sisk, 1999; Romeo et al., 2002a). Tāpēc šķiet, ka androgēni un AR ir nepieciešami, bet nav pietiekami, lai izteiktu vīriešu seksuālo uzvedību.
Līdz šim centieni identificēt faktorus, kas ierobežo seksuālo aktivitāti pirms pubertātes, ir bijuši dažādi. Fos reakcija uz ķīmijensorām, kas iegūtas no graujošām mātītēm, ir līdzīga prepubertālā un pieaugušo kāmju tēviņiem (Romeo, Parfitt, Richardson un Sisk, 1998). Šie dati parāda, ka maņu transdukcijas mehānismi ir nobrieduši pirms pubertātes. Tādējādi nepilngadīgi tēviņi spēj noteikt mātīšu kemosensorās norādes; atšķirībā no pieaugušajiem tas, kā viņi reaģē uz šīm norādēm. Viens no iespējamiem izskaidrojumiem ir tas, ka vīrieši pirmspubertālē nav motivēti iesaistīties seksuālā uzvedībā. Mēs esam noskaidrojuši, ka prepubertālajiem vīriešu kāmjiem MPOA nav izteiktas dopamīnerģiskas atbildes reakcijas uz sieviešu feromoniem, turpretim seksuāli naivu pieaugušiem vīriešiem ir spēcīgas MPOA dopamīnerģiskas reakcijas uz tiem pašiem stimuliem (Fig. 5, Schulz, Richardson, Romeo, Morris, Lookingland un Sisk, 2003). Tāpat pirmspubertāli vīrieši neuzrāda pieaugušajiem raksturīgo cirkulējošā testosterona pieaugumu pēc sieviešu feromonu iedarbības (Parfitt, Thompson, Richardson, Romeo un Sisk, 1999). Tādējādi sieviešu feromoni šķiet beznosacījuma stimuls neiroķīmiskajām un neiroendokrīnajām reakcijām pieaugušiem, bet ne pirmspubertāliem vīriešiem, kas liek domāt, ka šo sabiedriski nozīmīgo maņu stimulu izpausme mainās pubertātes attīstības laikā, iespējams, saistīta ar atalgojošo īpašību iegūšanu un seksuālo motivāciju. Turklāt, kaut arī testosterons atvieglo sievietes AGI sievietēm pirmspubertāles vīriešiem, šī iedarbība ir atkarīga no tā, vai vīrietis iepriekš ir bijis vai nav ticis pakļauts estrālā mātītei. Varbūt pārsteidzoši, ka ārstēšana ar testosteronu samazina latentumu un palielina AGI ilgumu tikai seksuāli naiviem pirmspubertāles vīriešiem (Fig. 6). Turklāt pirmspubertāles vīriešiem, kuriem ir bijusi viena iepriekšēja pieredze ar mātēm, ir daudz garāka AGI latence un īsāks AGI ilgums nekā vīriešiem, kuri pirmo reizi mijiedarbojas ar mātītēm, kuras ir uzņēmīgas (Fig. 6). Šie dati liek domāt, ka mijiedarbība ar sievišķīgu sievieti ir nelabvēlīga, nevis atlīdzinoša pirms pubertātes, tādējādi novēršot jebkādu testosterona veicinošo iedarbību uz AGI turpmākās mijiedarbības laikā ar mātīti. Būtu interesanti uzzināt, vai spēcīgas sievietes agrīnas iedarbības negatīvās uzvedības sekas saglabājas pusaudža un pieauguša cilvēka vecumā, īpaši ņemot vērā, ka atkārtota iedarbība uz sievietēm, kuras ir skaudras laikā pusaudža vecums parasti veicina vīriešu reproduktīvās uzvedības izpausmes (Molenda-Figueira, Salas-Ramirez, Schulz, Zehr, Montalto un Sisk, 2007).

Fig. 5
Prepubertāla un pieauguša vīrieša mediālās preoptiskās zonas (MPOA) dopamīnerģiskās reakcijas uz sieviešu feromoniem, kas atrodas maksts sekrēcijās. Pieaugušiem vīriešiem novēro MPOA dopamīnerģiskās aktivitātes palielināšanos, pakļaujot sieviešu maksts sekrēcijām, turpretim vīriešiem, kas atrodas pirmspubertāli, nav izteiktas MPOA dopamīnerģiskās reakcijas uz sieviešu feromoniem. (Atkārtoti no Šulcs, KM, Ričardsons, HN, Romeo, RD, Moriss, JA, Lookingland, KJ un Sisk, CL, 2003).

Lai arī ārstēšana pirmspubertālā androgēnā nevar izraisīt kopulāciju, jaunākais mūsu laboratorijas darbs liecina, ka kāmja nervu sistēma ir jutīga pret testosterona organizējošajām darbībām uz reproduktīvo uzvedību pirms pusaudža (Schulz, Zehr, Salas-Ramirez un Sisk, 2007). Kastrācija plus 19 dienu testosterona iedarbība pirms pusaudža vai tā laikā, bet ne pēc pusaudža vecuma atviegloja izturēšanos, kad testosterons tika aizstāts pieaugušā vecumā. Arī vīriešiem, kas pakļauti testosterona iedarbībai pirmspubertāli, pieaugušā vecumā tika novērota lielāka iejaukšanās nekā vīriešiem, kuri tika pakļauti testosterona iedarbībai pubertātes laikā vai pēc tās (Schulz et al., 2007). Šie dati liecina, ka testosterona spēja organizēt uzvedības neironu ķēdes samazinās līdz ar vecumu un pusaudža vecums iezīmē ilgstoša pēcdzemdību jutīga perioda beigas testosterona iedarbībai.
Farmakoloģiskie androgēni
Iepriekšējie dati liecina, ka endogēni dzimumdziedzeru steroīdi veicina motivētu uzvedību pusaudža gados. Kas notiek, ja kāds pats administrē androgēnus līmenī līdz 100x normālai fizioloģiskai koncentrācijai? Tā ir anaboliski androgēno steroīdu (AAS) ļaunprātīgas izmantošanas problēma (pārskatīta 2006. Gadā Brower, 2002; Klarks un Hendersons, 2003). Šeit ir piemērota īsa novirze: visi AAS ir testosterona atvasinājumi, visiem AAS ir oglekļa skelets ar 4 kausētiem gredzeniem, lielākajā daļā ir 19 ogleklis. AAS galvenokārt tiek izmantotas to anabolisko (muskuļu stiprināšanas) efektu dēļ. Tomēr, kā norāda viņu nosaukums, AAS ir arī androgēnas īpašības. Testosterons ir loģiska izvēle pētījumos ar dzīvniekiem, lai izpētītu androgēnu atlīdzības pamatmehānismus. Tā joprojām ir populāra izvēle arī lietotājiem, visbiežāk ilgstošas darbības testosterona esteru, piemēram, testosterona propionāta, veidā. 2006 testosterons bija visizplatītākā aizliegtā viela, kas tika konstatēta urīna analīzēs WADA akreditētās laboratorijās (WADA, 2006). Testosterons veidoja lielāko daļu (34%) no AAS pozitīvajiem urīna testiem 2000 Sidnejas olimpiskajās spēlēs (Van Eenoo un Delbeke, 2003). Tāpat AAS lietotāju urīna testos 41% testosterona (Pārlūks, Catlin, Blow, Eliopulos, Beresford, 1991). Lietojot lielas devas, AAS rada būtiskas izmaiņas uzvedībā. Jo īpaši, ņemot vērā to ciešo saistību ar testosteronu, AAS lietošana pusaudžu gados traucē normāli attīstīt pusaudžu nervu sistēmas steroīdu vidi, ieskaitot steroīdu iedarbības daudzumu, laiku un veidu.
Tāpat kā citu nelikumīgu narkotiku lietošana, AAS ļaunprātīga izmantošana cilvēkam ir pusaudžu problēma. Saskaņā ar 1994 nacionālo mājsaimniecību apsekojumu par narkotiku lietošanu (SAMHSA / OAS, 1996), steroīdu lietošanas maksimums vēlīnā pusaudža vecumā 18 gadu vecumā. Turklāt aptaujā Monitor the Future (Džonstons, O'Mallijs un Bahmans, 2003), steroīdu lietošanas biežums vidusskolas vecuma cilvēkiem (2.7%) mūža garumā bija salīdzināms ar kreka kokaīna (3.5%) vai heroīna (1.4%) rādītājiem. Steroīdu lietošana arvien biežāk tiek novērota arī jaunākā vecumā: 2.5% 8th klases skolēnu (13 – 14 gadi) ir lietojuši steroīdus, līdzīgi kā plaisu biežums (2.5%) un heroīna lietošana (1.6%). Šī tendence attiecībā uz AAS lietošanu agrīnā pusaudža vecumā ir īpaši satraucoša, ņemot vērā bažas par 1), ka pusaudži var būt īpaši jutīgi pret AAS ļaunprātīgu izmantošanu, un 2), ka pusaudžu AAS iedarbība farmakoloģiskā līmenī var būtiski mainīt normālu smadzeņu un uzvedība, lai izraisītu pārspīlētas morfoloģiskās un uzvedības reakcijas, akūti un hroniski.
Neatbilstoša agresija ir uzvedības reakcija, kas visbiežāk tiek saistīta ar cilvēku AAS ļaunprātīgu izmantošanu. Publicētajos gadījumu ziņojumos steroīdu lietošana ir saistīta ar vairākām vardarbīgām slepkavībām (Conacher un Workman, 1989; Pāvests un Katz, 1990; Pāvests, Kouri, Pauels, Kempbels un Katzs, 1996; Schulte, Hall un Boyer, 1993). Pašreizējo AAS lietotāju aptaujās paaugstināta agresija un aizkaitināmība bija AAS lietošanas biežākās uzvedības blakusparādības (Bonds, Choi un pāvests, 1995; Galligani, Renck un Hansen, 1996; Midlijs, Heather un Davies, 2001; Parrott, Choi un Davies, 1994; Perijs, Kučers, Lunds, Yates, Holmans un Demers, 2003). Tomēr, ņemot vērā androgēnu iedarbības diapazonu, psihisko simptomu daudzveidību un iepriekš pastāvošo psihisko disfunkciju potenciālu, ir grūti noteikt precīzu AAS lomu šajos cilvēku agresijas gadījumos. Potenciālo pētījumu rezultāti par brīvprātīgajiem cilvēkiem, kuri saņem AAS injekcijas, ir dažādi: Trikera et al (1996) un O'Connor et al (2004) nav ziņots par dusmīgas izturēšanās palielināšanos, kamēr citos pētījumos novērota pastiprināta agresija (Dalijs, Su, Šmits, Pikars, Mērfijs un Rubinovs, 2001; Hannans, Frīdls, Zolds, Ketlers un Plimāts, 1991; Kouri, Lukas, pāvests un Oliva, 1995; Pāvests un Katz, 1994; Su, Pagliaro, Schmidt, Pickar, Wolkowitz un Rubinow, 1993). Neskatoties uz to, ir svarīgi paturēt prātā, ka brīvprātīgajiem cilvēkiem ievadītās devas ir daudz mazākas nekā ķermeņa veidošanas vietnēs ieteiktās devas, un ārstēšanas ilgums parasti ir īss. Tādējādi kopumā šķiet taisnīgi secināt, ka AAS var uzlabot agonistisko izturēšanos vismaz jutīgos indivīdos. Pāvests un citi (1994) atklāja, ka AAS izraisa psihiskus simptomus neaizsargātām personām.
Pētījumi ar dzīvniekiem ir snieguši arī pārliecinošus pierādījumus par AAS izraisītu agresiju. Pusaudžu kāmju vīriešu kāmjiem, kurus hroniski ārstē ar lielām steroīdu devām, ir īsāks uzbrukuma latentums un lielāks uzbrukumu un kodumu skaits pret iebrucējiem vīriešiem, salīdzinot ar neārstētiem vīriešiem (Harisons, Konnors, Nowak, Nash un Melloni, 2000; Melloni, Connor, Hang, Harrison un Ferris, 1997). Līdzīgi, viegla provokācija (astes šķipsnu rašanās) rada pastāvīgu agresijas pieaugumu pusaudžu tēviņu tēviņiem, ieskaitot agresiju pret mātītēm (Kinthems un Makginniss, 2006). Vēl lielākas bažas rada tas, ka pusaudžu AAS iedarbība kāmjos izraisa ilgstošu agonistiskās izturēšanās palielināšanos, kas saglabājas pēc steroīdu lietošanas pārtraukšanas (Grimes un Melloni, 2006). Šīs uzvedības izmaiņas pavada ilgstoša nervu shēmas pārveidošana priekšējā hipotalāmā. Proti, pusaudžu AAS iedarbība kāmjos pastiprina arginīna vazopresīnu (AVP, Grimes un Melloni, 2006) un noregulē serotonīnu un serotonīnerģiskos 5HT1A un 5HT1B receptorus (Ricci, Rasakham, Grimes un Melloni, 2006). Nav pārsteigums, ka AAS maina arī AR smadzeņu līmeni. Testosterona vai nandrolona hroniska iedarbība palielina šūnu kodola AR žurku tēviņiem (Menards un Harlans, 1993; Wesson un McGinnis, 2006). Tādējādi AAS ir iespējams uzlabot no androgēniem atkarīgu uzvedību, gan papildinot endogēnos androgēnus, gan palielinot androgēno reakciju, palielinot AR ekspresiju.
Salīdzinot ar agonistisko uzvedību, AAS ir mazāk izteikta ietekme uz pārošanās izturēšanos vīriešu kārtas grauzējiem, un reakcija ir atkarīga no izmantotā steroīda (pārskatīts Klarks un Hendersons, 2003). Vīriešu kāmjiem, kas testosteronu patērē iekšķīgi lietojamos šķīdumos, ejakulācijas palielinājās atkarībā no devas (Wood, 2002). Tomēr ne testosterons, ne nandrolons neuzlaboja pārošanos pusaudžu žurku tēviņiem. Stanozolols, relatīvi mazāk spēcīgs AAS ar minimālu androgēnu aktivitāti, faktiski kavēja gan pārošanos, gan agresiju (Farrell un McGinnis, 2003), domājams, samazinot endogēno androgēnu līmeni.
Īpaši svarīgi atzīmēt, ka pusaudžu un pieaugušo kāmji var parādīt atšķirīgu uzvedības reakciju uz AAS iedarbību. Kaut arī AAS ievērojami uzlaboja agonistisko izturēšanos pusaudžu vīriešiem, tā pati attieksme pieaugušā vecumā izraisīja tikai nelielu agresīvas uzvedības pieaugumu un ievērojami samazināja seksuālo izturēšanos (Salas-Ramirez, Montaldo and Sisk, 2008). Tas saskan ar pusaudža vecuma jēdzienu kā jutīgu periodu androgēnu darbībai. Turklāt, tāpat kā pieaugušie kāmji vīrieši iegūst toleranci pret eksogēnu testosteronu (Peters un Wood, 2005), mēs uzskatām, ka jaunattīstības vīrieši iegūst toleranci pret testosteronu nobriešanas laikā. Tādējādi AAS ietekme mainās visā pusaudža attīstībā, un pusaudža AAS iedarbība var izraisīt pārmērīgu agresīvu un seksuālu uzvedību, kas var saglabāties arī pieaugušā vecumā.
Androgēnu pastiprinošā iedarbība
Pārošanās un cīņa ir gandarījums (vismaz, ja jūs uzvarējat cīņā). Žurku tēviņi atkārtoti nospiedīs sviru, lai kopētu ar mātīti (Everits un Stacey, 1987). Tāpat peles un vīriešu kāmji veidos kondicionētu vietas izvēli (CPP) vietām, kur viņi iepriekš ir uzvarējuši cīņās (Martinez, Guillen-Salazar, Salvador un Simon, 1995; Meisel un Joppa, 1994). Ja AAS var uzlabot sociālās izturēšanās atalgojumu virs līmeņa, kāds parasti tiek novērots vīriešiem, kuri ir neskarti ar dzimumdziedzeriem, ir loģiski sagaidīt, ka pats testosterons varētu būt izdevīgs. Tas tika pārbaudīts, izmantojot divus labi zināmus dzīvnieku atlīdzības un stiprināšanas modeļus: CPP un pašpārvaldi. Šo pētījumu rezultāti parāda, ka testosterons pastiprinās eksperimentālā kontekstā, kur anaboliskajai iedarbībai un sportiskajam sniegumam nav nozīmes. Izmantojot CPP, testējamā viela atkārtoti tiek savienota pārī ar unikālu vidi (piemēram, ar noteiktu kameru testēšanas aparātā). Tiklīdz dzīvnieks saistīs testējošo pastiprinošo vielu ar šo vidi, viņš meklēs vidi pat bez atlīdzības. Pirmajos ziņojumos par androgēnu atlīdzību laboratorijas dzīvniekiem tika izmantotas sistemātiskas testosterona injekcijas, lai izraisītu CPP peļu tēviņiem (Arnedo, Salvadora, Martinez-Sanchis un Gonzalez-Bono, 2000; Arnedo, Salvadora, Martinez-Sanchis un Pellicer, 2002) un žurkām (Aleksandrs, Packards un Hiness, 1994; de Beuns, Jansens, Slangens un Van de Polls, 1992). Pēc tam mūsu laboratorija izmantoja testosterona pašpārvaldi, lai pierādītu androgēnu pastiprināšanos (Johnson un Wood, 2001). Mēs noskaidrojām, ka vīriešu kāmji brīvprātīgi patērēs testosterona šķīdumus iekšķīgai lietošanai, izmantojot gan 2 pudeles izvēles testus, gan ēdiena izraisītu dzeršanu. Vēlākos pētījumos mēs pierādījām iv pašinjekciju žurku un kāmju tēviņiem (Vuds, Džonsons, Ču, Šads un Selfs, 2004). Intravenoza ievadīšana novērš garšas vai zarnu piepildījuma iespējamo jaukto ietekmi uz androgēnu uzņemšanu.
AAS ļaunprātīgas izmantošanas kontekstā ir svarīgi nošķirt androgēnu centrālo un perifēro iedarbību. Tā kā testosteronam ir plaši izplatīta iedarbība visā ķermenī, varētu apgalvot, ka atlīdzība un stiprināšana ar sistēmiskām testosterona injekcijām ir sekundāra salīdzinājumā ar testosterona sistēmiskajām anaboliskajām un androgēnām darbībām. Citiem vārdiem sakot, varbūt testosterons samazina muskuļu nogurumu un uzlabo locītavu darbību, lai dzīvnieki vienkārši justies labāk. Patiešām, šis skaidrojums ir izmantots klīniskajā literatūrā (kaut arī bez eksperimentāliem pierādījumiem), lai argumentētu pret AAS atkarības un atkarības iespējamību (DiPasquale, 1998). Tomēr Packard et al (Packards, Kornels un Aleksandrs, 1997) parādīja, ka testosterona injekcijas tieši žurku smadzenēs var izraisīt CPP. Tāpat mūsu laboratorija ir pierādījusi intracerebroventrikulāru (icv) testosterona pašpārvaldi kāmjiem vīriešiem (Wood et al., 2004). Intracerebrālā CPP un icv pašpārvalde ar testosteronu apgalvo, ka galvenie mērķi ir mediēt androgēnu stiprināšanu.
Jāatzīmē, ka testosterona stiprināšana ne vienmēr notiek saskaņā ar tiem pašiem mehānismiem, kas iepriekš tika noteikti steroīdu iedarbībai uz seksuālo uzvedību. Kā jau iepriekš tika runāts, MPOA ir galvenā vietne vīriešu grauzēju seksuālās uzvedības organizēšanai (Korpuss, Meisel un Sachs, 2002). Kāmjiem MPOA ir bagātīgi steroīdu receptori, un testosterona implanti MPOA atjauno seksuālo aktivitāti ilgstošos kastrātos (Koks un Swann, 1999). Šo steroīdu iedarbības laiks ir lēns: izturēšanās izturēšanās saglabājas vairākas nedēļas pēc orhidektomijas, un, lai atjaunotu pārošanos ilgtermiņa kastrātos, ir nepieciešama pastiprināta steroīdu iedarbība (Noble un Alsum, 1975). Tomēr testosterona injekcijas žurku tēviņu MPOA neizraisa CPP (Kings, Packards un Aleksandrs, 1999). Tas liek domāt, ka citi smadzeņu reģioni ir svarīgi androgēnu stiprināšanai.
Turpretī žurku tēviņi veidos CPP testosterona injekcijām Acb (Packard et al., 1997). Tāpat kā citu narkotiku lietošana, DA, iespējams, ir galvenais testosterona pastiprināšanas neiromediators: sistēmiskas testosterona injekcijas izraisītu CPP bloķē D1 un D2 dopamīna receptoru antagonisti (Schroeder un Packard, 2000). Tomēr atšķirībā no citām narkotikām, kas saistītas ar ļaunprātīgu izmantošanu, mūsu pētījumi ar kāmjiem liecina, ka testosterons neizraisa Acb DA izdalīšanos (Triemstra, Sato un Wood, presē). Tāpat žurku tēviņu pētījumi rāda, ka androgēni neietekmē DA bazālo līmeni vai amfetamīna stimulētu DA izdalīšanos (Birgners, Kindlundh-Hogberg, Nyberg un Bergstrom, 2006; bet arī redzēt Klarks, Lindenfelds un Gibbons, 1996). Turklāt testosterons salīdzinoši maz ietekmē Acb DA audu līmeni (Thiblin, Finn, Ross un Stenfors, 1999). Šie dati kopā liecina, ka, kaut arī testosterona stiprināšana galu galā var mainīt DA aktivitāti Acb, šie mehānismi var atšķirties no kokaīna vai citu stimulantu mehānismiem. Šajā sakarā jaunākie dati liecina, ka hroniska AAS iedarbība var mainīt jutīgumu pret DA, mainot DA metabolismu (Kurling, Kankaanpaa, Ellermaa, Karila un Seppala, 2005), DA receptoru līmenis (Kindlundh, Lindblom, Bergstrom, Wikberg un Nyberg, 2001; Kindlundh, Lindblom un Nyberg, 2003) vai DA transportētāju (Kindlundh, Bergstrom, Monazzam, Hallberg, Blomqvist, Langstrom un Nyberg, 2002).
Pašlaik īpašie steroīdu signāli, receptori un smadzeņu darbības vietas testosterona pastiprināšanai nav zināmi. Balstoties uz neseno mūsu laboratorijas pētījumu par kāmjiem, šķiet, ka testosterona pastiprinošo iedarbību ietekmē gan androgēni, gan estrogēni (DiMeo un Wood, 2006). Bieži ļaunprātīgi izmantotā AAS ietver gan aromatizējamus, gan aromātiskus androgēnus (Gallaway, 1997; WADA, 2006). Tas nozīmē, ka gan AR, gan ER var atlīdzināt par steroīdu stimuliem. Pastāv papildu iespēja, ka testosterona stiprināšanu var izraisīt klasisko un negenomisko receptoru kombinācija.
Vairāki pierādījumi norāda uz genogēnu receptoru darbību AAS pastiprinošajā iedarbībā. Papildus retajam AR sadalījumam Acb un VTA, androgēnu pastiprināšanas laika kurss ir ātrs (<30 min), un signāla apstrāde, izmantojot klasisko AR, var nebūt pietiekami ātra pastiprināšanai. Attiecīgi, lai pārbaudītu neģenomas AR nozīmi AAS pastiprināšanā, mēs izmantojām divas papildu metodes (Fig. 7). Vienā eksperimentā (Sato, Johansen, Jordānija un Wood, 2006), mēs ļāvām žurkām ar sēklinieku feminizācijas mutāciju (Tfm, skatīt šo jautājumu) pašiem ievadīt dihidrotestosteronu (DHT), kas nav aromātisks androgēns. Tfm mutācija ievērojami samazina ligandu saistīšanos AR. Neskatoties uz to, Tfm žurkas un viņu savvaļas tipa vīriešu kārtas māsas un brāļi un māsas, kuras pašiem ievada aptuveni vienādu daudzumu DHT. Tas norāda uz DHT negenomisko iedarbību. Turpmākajā pētījumā mēs noskaidrojām, vai vīriešu kāmji pats administrēs DHT, kas konjugēts ar liellopu seruma albumīnu (BSA, Fig. 8, Sato un koks, 2007). DHT-BSA konjugāti ir membrāncaurlaidīgi; tādējādi to iedarbība aprobežojas ar šūnas virsmu. Kāmji, ko pats ievada DHT, kā tika pierādīts iepriekš (DiMeo un Wood, 2006). Viņi parādīja līdzīgu priekšroku DHT-BSA konjugātiem, bet nespēja pašiem ievadīt BSA vien.

Fig. 7

Fig. 8
Šie dati norāda uz šūnu virsmas AR centrālo lomu androgēnu stiprināšanā. Pašlaik precīzs šādu receptoru raksturs nav zināms. Ir ierosināts, ka androgēni var darboties uz šūnu virsmas, saistoties ar tam paredzēto membrānu AR (Tomass, Dressing, Pangs, Bergs, Tubbs, Benninghoff un Doughty, 2006, skatiet arī šo jautājumu). Tas var būt klasiskā ekstrakodolenda formā, kā ziņots hipokampā (šajā jautājumā Sarkey et al.). Alternatīvi, iepriekšējie pētījumi ir aprakstījuši arī steroīdu saistošās vietas citās neirotransmiteru sistēmās. Konkrēti, dažādi steroīdu hormoni, ieskaitot AAS, var allosteriski modulēt GABA-A receptoru (Hendersons, 2007; Lamberts, Belelli, Pedens, Vardijs un Peters, 2003). Tāpat sulfāti neirosteroīdi var mainīt aktivitāti N-metil-D-partelatīvo receptoru apakštipi (Malajevs, Gibbs un Farbs, 2002) receptori. Šī ir svarīga joma turpmākiem pētījumiem.
Kāpēc vajadzētu būt membrānas AR? Kā jau iepriekš tika runāts, pastāv cieša saistība starp androgēnu sekrēciju un sociālās izturēšanās atalgošanu. Mēs varam spekulēt, ka testosterona sekrēcijas palielināšanās, kas seko pārošanai vai cīņai, pastiprina uzvedību. Ja tā, ir nepieciešams ātri stimulēt (uzvedība) un atlīdzība (testosterons). To vislabāk var panākt, saistoties ar membrānu AR. Šajā sakarā būtu interesanti noteikt, vai androgēnu sekrēcijas nostiprināšana pārošanās laikā mazina seksuālās uzvedības labvēlīgo ietekmi.
Kopsavilkums
Šeit mēs apskatām pierādījumus tam, ka androgēni ir spēcīgi mediatori pieaugušo motivētai uzvedībai, un tālāk, ka androgēnu iedarbības laiks attīstības programmu laikā no androgēnu atkarīgas motivētas uzvedības pieaugušā vecumā. Anaboliskie steroīdi ātri kļūst par iecienītākajām narkotikām, ko pusaudži ASV ļaunprātīgi izmanto. Kaut arī AAS var nebūt atkarīga no kokaīna vai heroīna atkarības, mēs tikai sākam izprast androgēnu stiprināšanas un atkarības potenciālu. Jo īpaši, tā kā jauniešu sports kļūst konkurētspējīgāks, pieaug spiediens uz jaunattīstības sportistiem lietot steroīdus, sākot no jaunākiem gadiem. Šī tendence ir satraucoša, ņemot vērā jaunus pierādījumus par steroīdiem jutīgu nervu nobriešanu pusaudžiem.
Neskatoties uz to, ka gan sabiedrība, gan zinātniskā sabiedrība ir palielinājusi izpratni par dziļajām neironu izmaiņām, kas pavada pusaudža gados, pubertātes attīstības neirobioloģijas eksperimentālie pētījumi ir ierobežoti. Dzīvnieku pusaudžu attīstības modeļi ir nepieciešami, lai izpētītu, kā hormonu iedarbības laiks attīstības laikā palielina indivīda psihopatoloģijas un narkotiku lietošanas risku un kāda veida pieredze mazina vai pastiprina pubertāles laika noviržu uzvedības ietekmi. Piemēram, sociālie faktori, piemēram, vienaudžu ietekme, saasina pubertātes laika ietekmi uz vielu un alkohola lietošanu (Bīls, Natsuaki un Ge, 2007; Patton, Novy, Lee un Hickok, 2004; Simons-Mortons un Haynie, 2003; Vištroms un Pedersens, 2001). Dzīvnieku pubertātes laika modeļi arī ietekmēs cilvēku pētījumu centienus, un tie, iespējams, novedīs pie efektīvākas terapeitiskās iejaukšanās pusaudža gados.
Pateicības
Mēs pateicamies Eleni Antzoulatos, Cortney Ballard, Lucy Chu, Kelly Peters, Jennifer Triemstra, Jane Venier, Lisa Rogers un Pamela Montalto par palīdzību šajos pētījumos. Šo darbu atbalsta NIH dotācijas (DA12843 RIW, MH68764 CLS un MH070125 KMS).
Zemsvītras piezīmes
Izdevēja atruna: Šis ir PDF fails, kurā nav publicēta manuskripta, kas ir pieņemts publicēšanai. Kā pakalpojums mūsu klientiem sniedzam šo rokraksta agrīno versiju. Manuskripts tiks pakļauts kopēšanu, apkopošanu un iegūto pierādījumu pārskatīšanu, pirms tas tiek publicēts tā galīgajā citējamajā formā. Lūdzu, ņemiet vērā, ka ražošanas procesa laikā var rasties kļūdas, kas var ietekmēt saturu, un attiecas uz visiem žurnālam piemērojamiem juridiskajiem atrunas.
Atsauces
- Alexander GM, Packard MG, Hines M. Testosterons apbalvo emocionālas īpašības vīriešu kārtas žurkām: sekas seksuālās motivācijas bioloģiskajam pamatam. Uzvedības neirozinātne. 1994;108: 424-8. [PubMed]
- Andersen SL. Smadzeņu attīstības trajektorijas: neaizsargātība vai iespēju logs? Neirozinātnes un bioloģiskās uzvedības atsauksmes. 2003;27: 3-18. [PubMed]
- Andersen SL, Rutstein M, Benzo JM, Hostetter JC, Teicher MH. Seksu atšķirības dopamīna receptoru pārprodukcijā un eliminācijā. Neuroreport. 1997;8: 1495-8. [PubMed]
- Arnedo MT, Salvador A, Martinez-Sanchis S, Gonzalez-Bono E. Testosterona īpašības veseliem vīriešu pelēm: izmēģinājuma pētījums. Farmakoloģija, bioķīmija un uzvedība. 2000;65: 327-32.
- Arnedo MT, Salvador A, Martinez-Sanchis S, Pellicer O. Līdzīga testosterona atalgojošā iedarbība pelēm, kas novērtētas kā īstermiņa un ilgstoša uzbrukuma latentuma indivīdi. Atkarības bioloģija. 2002;7: 373-9. [PubMed]
- Arnolds AP, Breedlove SM. Seksa steroīdu organizatoriskā un aktivizējošā ietekme uz smadzenēm un uzvedību: reanalīze. Hormoni un izturēšanās. 1985;19: 469-98. [PubMed]
- Benes FM, Taylor JB, Cunningham MC. Monoaminergisko sistēmu konverģence un plastiskums vidējā prefronta garozā pēcdzemdību periodā: sekas psihopatoloģijas attīstībai. Smadzeņu garoza. 2000;10: 1014-27. [PubMed]
- Benes FM, Turtle M, Khan Y, Farol P. Hipokampas veidošanās galvenās releju zonas mielinēšana notiek cilvēka smadzenēs bērnības, pusaudža un pieaugušo vecuma laikā. Vispārējās psihiatrijas arhīvs. 1994;51: 477-84. [PubMed]
- Biehl MC, Natsuaki MN, Ge XJ. Pubertātes laika ietekme uz alkohola lietošanu un smagas alkohola lietošanas trajektorijām. Jauniešu un pusaudžu žurnāls. 2007;36: 153-167.
- Birgner C, Kindlundh-Hogberg AM, Nyberg F, Bergstrom L. Neirozinātņu vēstules. 2006. Mainīti ārpusšūnu DOPAC un HVA līmeņi žurku kodola akumulācijas apvalkā, reaģējot uz subhronisku nandrolona ievadīšanu un sekojošu amfetamīna izaicinājumu.
- Bonds AJ, Choi PY, pāvests HG., Jr. Anabolisko-androgēno steroīdu lietotāju un nelietotāju uzmanības novirzes un noskaņojuma novērtēšana. Atkarība no narkotikām un alkohola. 1995;37: 241-5. [PubMed]
- Brower KJ. Anaboliskie steroīdu ļaunprātīga izmantošana un atkarība. Pašreizējie psihiatrijas ziņojumi. 2002;4: 377-87. [PubMed]
- Pārlūks KJ, Catlin DH, Blow FC, Eliopulos GA, Beresford TP. Anabolisko-androgēno steroīdu ļaunprātīgas izmantošanas un atkarības klīniskā pārbaude un urīna pārbaude. American Journal of Drug & Alcohol Abuse. 1991;17: 161-171. [PubMed]
- Celotti F, Negri-Cesi P, Poletti A. Steroīdu metabolisms zīdītāju smadzenēs: 5alpha redukcija un aromatizācija. Prāta pētījumu biļetens. 1997;44: 365-75. [PubMed]
- Chung WC, De Vries GJ, Swaab DF. Stria terminalis gultnes kodola seksuālā diferenciācija cilvēkiem var izvērsties pieaugušā vecumā. Neiroloģijas žurnāls. 2002;22: 1027-33. [PubMed]
- Clark AS, Henderson LP. Uzvedības un fizioloģiskās reakcijas uz anaboliskiem-androgēniem steroīdiem. Neirozinātnes un bioloģiskās uzvedības atsauksmes. 2003;27: 413-36. [PubMed]
- Clark AS, Lindenfeld RC, Gibbons CH. Anaboliski androgēnie steroīdi un smadzeņu atalgojums. Farmakoloģija, bioķīmija un uzvedība. 1996;53: 741-5.
- Conacher GN, Workman ģenerāldirektorāts. Vardarbīgs noziegums, iespējams, saistīts ar anabolisko steroīdu lietošanu. American Journal of Psychiatry. 1989;146: 679. [PubMed]
- Cunningham RL, McGinnis MANS. Fabrikāņu pubertāles anabolisko androgēno steroīdu iedarbībā esošās žurku tēviņi izraisa agresiju pret mātītēm. Hormoni un izturēšanās. 2006;50: 410-6. [PubMed]
- Daly RC, Su TP, Schmidt PJ, Pickar D, Murphy DL, Rubinow DR. Cerebrospinālais šķidrums un uzvedības izmaiņas pēc metiltestosterona ievadīšanas: provizoriski atklājumi. Vispārējās psihiatrijas arhīvs. 2001;58: 172-7. [PubMed]
- Deiviss EC, Shryne JE, Gorski RA. Žurku hipotalāma anteroventralajā periventrikulārajā kodolā esošie strukturālie seksuālie dimorfismi ir jutīgi pret peronāli dzimumdziedzeru steroīdiem, bet attīstās perubertāli. Neuroendokrinoloģija. 1996;63: 142-8. [PubMed]
- de Beun R, Jansen E, Slangen JL, Van de Poll NE. Testosterons kā apetītes un diskriminējošs stimuls žurkām: no dzimuma un devas atkarīga ietekme. Fizioloģija un uzvedība. 1992;52: 629-34. [PubMed]
- AS Dekaban. Smadzeņu svara izmaiņas cilvēka dzīves laikā: smadzeņu svara attiecība pret ķermeņa augstumu un ķermeņa svaru. Neiroloģijas Annals. 1978;4: 345-56. [PubMed]
- Di Chiara G, Imperato A. Cilvēki, kurus ļaunprātīgi lietojuši cilvēki, galvenokārt palielina sinaptisko dopamīna koncentrāciju brīvi kustīgu žurku mesolimbiskajā sistēmā. Amerikas Savienoto Valstu Zinātņu akadēmijas darbi. 1988;85: 5274-8. [PMC bezmaksas raksts] [PubMed]
- DiMeo AN, koksnes RI. Estrogēna un dihidrotestosterona pašregulācija tēviņiem. Hormoni un izturēšanās. 2006;49: 519-26. [PubMed]
- DiPasquale M. Anaboliskie steroīdi. In: Tarter RE, Ammerman RT, Ott PJ, redaktori. Rokasgrāmata par vielu ļaunprātīgu izmantošanu. Plēnuma prese; NY: 1998. 547 – 565 lpp.
- Dominguez J, Riolo JV, Xu Z, Hull EM. To regulē kopulācijas mediālā amigdala un mediālā preoptiskā dopamīna izdalīšanās. Neiroloģijas žurnāls. 2001;21: 349-355. [PubMed]
- Dominguez JM, Hull EM. Mediālās amigdala stimulēšana pastiprina mediālo preoptisko dopamīna izdalīšanos: tas ietekmē vīriešu žurku seksuālo izturēšanos. Smadzeņu izpēte. 2001;917: 225-229. [PubMed]
- Du J, Lorrain DS, Hull EM. Kastrācija samazina ārpusšūnu, bet palielina intracelulāro, dopamīna līmeni žurku tēviņu mediālajā preoptiskajā zonā. Smadzeņu izpēte. 1998;782: 11-17. [PubMed]
- Eckenhoff MF, Rakic P. Hipokampu protezējošo šūnu raksturs un liktenis Hemokampā dentet Gyrus laikā rēzus pērtiķa dzīves laikā. Neiroloģijas žurnāls. 1988;8: 2729-47. [PubMed]
- Everitt BJ, Stacey P. Pētījumi par instrumentālo uzvedību ar seksuālu pastiprināšanu vīriešu kārtas žurkām (Rattus norvegicus): II. Preoptiskās zonas bojājumu, kastrācijas un testosterona ietekme. Salīdzinošās psiholoģijas žurnāls. 1987;101: 407-19. [PubMed]
- Gallaway S. Steroīdu Bībele. Belle International Press; Sakramento, Kalifornijā: 1997.
- Galligani N, Renck A, Hansen S. Vīriešu, kas lieto anaboliskos androgēnos steroīdus, personības raksturojums. Hormoni un izturēšanās. 1996;30: 170-5. [PubMed]
- Grimes JM, Melloni RH., Jr Ilgstošas serotonīna neironu sistēmas izmaiņas pēc pusaudžu anabolisko-androgēno steroīdu iedarbības pārtraukšanas kāmjiem (Mesocricetus auratus) Uzvedības neirozinātne. 2006;120: 1242-51. [PubMed]
- Hannan CJ, Jr, Friedl KE, Zold A, Kettler TM, Plymate SR. Psiholoģiskas un seruma homovanilskābes izmaiņas vīriešiem, kuriem ievadīti androgēni steroīdi. Psihoneiroendokrinoloģija. 1991;16: 335-43. [PubMed]
- Harrison RJ, Connor DF, Nowak C, Nash K, Melloni RH., Jr Hroniska anaboliska-androgēna steroīdu terapija pusaudža gados palielina priekšējā hipotalāma vazopresīnu un agresiju neskartos kāmjos. Psihoneiroendokrinoloģija. 2000;25: 317-38. [PubMed]
- Viņš J, apkalpes FT. Neiroģenēze samazinās smadzeņu nobriešanas laikā no pusaudža vecuma līdz pieauguša cilvēka vecumam. Farmakoloģija, bioķīmija un uzvedība. 2007;86: 327-33.
- Hendersons LP. GABAA receptoru starpniecības steroīdu modulācija hipotalāmā: ietekme uz reproduktīvo funkciju. Neirofarmakoloģija. 2007;52: 1439-53. [PMC bezmaksas raksts] [PubMed]
- Korpuss EM, Du J, Lorrain DS, Matuszewich L. Āršūnu dopamīns mediālajā preoptiskajā zonā: ietekme uz seksuālo motivāciju un kopulācijas hormonālo kontroli. Neiroloģijas žurnāls. 1995;15: 7465-7471. [PubMed]
- Korpusa EM, Meisel RL, Sachs BD. Vīriešu seksuālā izturēšanās. In: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, editors. Hormoni, smadzenes un uzvedība. Akadēmiskā prese; Ņujorka: 2002. 3 – 137 lpp.
- Hull EM, Wood RI, McKenna KE. Vīriešu seksuālās uzvedības neirobioloģija. In: Neill JD, redaktors. Reprodukcijas fizioloģija. Vol. 1. Elsevier Press; Ņujorka: 2006. 1729 – 1824 lpp.
- Huttenlocher PR, Dabholkar AS. Sinagoģenēzes reģionālās atšķirības cilvēka smadzeņu garozā. Salīdzinošās neiroloģijas žurnāls. 1997;387: 167-78. [PubMed]
- Johnson LR, Wood RI. Iekšķīgi testosterona pašpārvalde vīriešu kāmjiem. Neuroendokrinoloģija. 2001;73: 285-92. [PubMed]
- Johnston LD, O'Malley PM, Bachman JG. Vidusskolnieki (NIH publikācijas Nr. 03 – 5375) I. Bethesda, MD: Nacionālais narkomānijas novēršanas institūts; 2003. Nākotnes nacionālā apsekojuma rezultātu uzraudzība par narkotiku lietošanu, 1975 – 2002.
- Ķeizara ģimenes fonds. ASV pusaudžu seksuālā aktivitāte. 2005. lpp. # 3040 – 02.
- Kashon ML, Hayes MJ, Shek PP, Sisk CL. Smadzeņu androgēnu receptoru imūnreaktivitātes regulēšana ar androgēniem prepubertālos vīriešu seskiem. Reprodukcijas bioloģija. 1995;52: 1198-205. [PubMed]
- Kindlundh AM, Bergstrom M, Monazzam A, Hallberg M, Blomqvist G, Langstrom B, Nyberg F. Dopamīnerģiskie efekti pēc hroniskas ārstēšanas ar nandrolonu, kas vizualizēta žurku smadzenēs ar pozitronu emisijas tomogrāfiju. NeiroPsihofarmakoloģijas un bioloģiskās psihiatrijas progress. 2002;26: 1303-8.
- Kindlundh AM, Lindblom J, Bergstrom L, Wikberg JE, Nyberg F. Anaboliski androgēnais steroīds nandrolona dekanoāts ietekmē dopamīna receptoru blīvumu žurku vīriešu smadzenēs. European Journal of Neuroscience. 2001;13: 291-6. [PubMed]
- Kindlundh AM, Lindblom J, Nyberg F. Hroniska ievadīšana ar nandrolona dekanoātu izraisa izmaiņas dopamīna D (1) un D (2) receptoru gēnu transkripcijas saturā žurku smadzenēs. Smadzeņu izpēte. 2003;979: 37-42. [PubMed]
- Karalis BE, Packard MG, Aleksandrs GM. Testosterona intramediālā preoptiskā apgabala injekciju iedarbība uz žurku tēviņiem. Neirozinātņu vēstules. 1999;269: 149-52. [PubMed]
- Koob GF, Nestler EJ. Narkotiku atkarības neirobioloģija. Neiropsihiatrijas un klīnisko neirozinātņu žurnāls. 1997;9: 482-97. [PubMed]
- Kouri EM, Lukas SE, pāvests HG, Jr, Oliva PS. Pastiprināta agresīva reakcija vīriešiem brīvprātīgajiem pēc pakāpeniski pieaugošu testosterona cypionate devu ievadīšanas. Atkarība no narkotikām un alkohola. 1995;40: 73-9. [PubMed]
- Kritzer MF. Ilgstoša gonadektomija ietekmē pieaugušo vīriešu žurku mediālās prefrontālās korekcijas tirozīna hidroksilāzes, bet ne dopamīna-beta-hidroksilāzes, holīna acetiltransferāzes vai serotonīna-imunoreaktīvo axonu blīvumu. Smadzeņu garoza. 2003;13: 282-296. [PubMed]
- Kurling S, Kankaanpaa A, Ellermaa S, Karila T, Seppala T. Subhroniskas nandrolona dekanoāta apstrādes ietekme uz dopamīnerģisko un serotonerģisko neironu sistēmu žurku smadzenēs. Smadzeņu izpēte. 2005;1044: 67-75. [PubMed]
- Lambert JJ, Belelli D, Peden DR, Vardy AW, Peters JA. GABAA receptoru neirosteroīdu modulācija. Neirobioloģijas attīstība. 2003;71: 67-80. [PubMed]
- Lenroot RK, Giedd JN. Smadzeņu attīstība bērniem un pusaudžiem: ieskats no anatomiskās magnētiskās rezonanses attēlveidošanas. Neirozinātnes un bioloģiskās izturēšanās pārskats. 2006;30: 718-29.
- Lesting J, Neddens J, Teuchert-Noodt G. Dopamīna inervācijas ontogēnija gerbilu kodolos. Smadzeņu izpēte. 2005;1066: 16-23. [PubMed]
- Malajevs, Gibbs TT, Farb DH. NMDA atbildes reakcijas inhibēšana, lietojot pregnenolona sulfātu, atklāj NMDA receptoru selektīvo modulāciju ar sulfātu steroīdu palīdzību. Lielbritānijas žurnāls par farmakoloģiju. 2002;135: 901-9. [PMC bezmaksas raksts] [PubMed]
- Martinez M, Guillen-Salazar F, Salvador A, Simon VM. Veiksmīga starpmaņu agresija un nosacīta vietas izvēle pelēm. Fizioloģija un uzvedība. 1995;58: 323-8. [PubMed]
- Lēnām LR, Romeo RD, Novak CM, Sisk CL. Testosterona darbības prepubertālā un postpubertālajā vīriešu kāmīšos: ietekmes uz reproduktīvo uzvedību un smadzeņu androgēnu receptoru imūnreaktivitātes disociācija. Hormoni un uzvedība. 1997;31: 75-88.
- Meisel RL, Joppa MA. Pēc agresīvas vai seksuālas tikšanās sievietes kāmjiem priekšroka tiek dota. Fizioloģija un uzvedība. 1994;56: 1115-8. [PubMed]
- Melloni RH, Jr, Connor DF, Hang PT, Harrison RJ, Ferris CF. Anaboliski androgēno steroīdu iedarbība pusaudža gados un agresīva izturēšanās zelta kāmjiem. Fizioloģija un uzvedība. 1997;61: 359-64. [PubMed]
- Menards C, Harlans R. Androgēnu receptoru imūnreaktivitātes regulēšana žurku smadzenēs ar androgēniem-anaboliskiem steroīdiem. Smadzeņu izpēte. 1993;622: 226-236. [PubMed]
- Mermelstein PG, Becker JB, Surmeier DJ. Estradiols samazina kalcija plūsmas žurku neostriatāla neironos caur membrānas receptoriem. Neiroloģijas žurnāls. 1996;16: 595-604. [PubMed]
- Midgley SJ, Heather N, Davies JB. Agresijas līmeņi anabolisko-androgēno steroīdu lietotāju grupā. Medicīna, zinātne un likumi. 2001;41: 309-14.
- Molenda-Figueira HN, Salas-Ramirez KY, Schulz KM, Zehr JL, Montalto PR, Sisk CL. Pusaudžu sociālā pieredze atjauno pieaugušo ejakulācijas uzvedību Sīrijas kāmju vīriešiem, kuriem trūkst pubertāles testosterona. Uzvedības neiroendokrinoloģijas biedrība; Klusā okeāna birža, Kalifornija: 2007.
- Noble RG, Alsum PB. No hormoniem atkarīgi dzimuma dimorfismi zeltainā kāmī (Mesocricetus auratus) Fizioloģija un uzvedība. 1975;14: 567-74. [PubMed]
- Nunez JL, Lauschke DM, Juraska JM. Šūnu nāve žurku tēviņu un mātīšu aizmugurējā garozas attīstībā. Salīdzinošās neiroloģijas žurnāls. 2001;436: 32-41. [PubMed]
- Nunez JL, Sodhi J, Juraska JM. Olnīcu hormoni pēcdzemdību dienā 20 samazina neironu skaitu žurku primārajā redzes garozā. Neirobioloģijas žurnāls. 2002;52: 312-21. [PubMed]
- O'Connor DB, Archer J, Wu FC. Testosterona ietekme uz garastāvokli, agresiju un seksuālu izturēšanos jaunos vīriešos: dubultaklā, placebo kontrolētā, savstarpēji salīdzinošā pētījumā. Klīniskās endokrinoloģijas un metabolisma žurnāls. 2004;89: 2837-45. [PubMed]
- Packard MG, Kornels AH, Aleksandrs GM. Apsverot testosterona injekcijas introduktīvās iekšējās asinsspiediena pazīmes. Uzvedības neirozinātne. 1997;111: 219-24. [PubMed]
- Parfitt DB, Thompson RC, Richardson HN, Romeo RD, Sisk CL. GnRH mRNS palielinās līdz ar pubertāti vīriešu Sīrijas kāmju smadzenēs. Neuroendokrinoloģijas žurnāls. 1999;11: 621-7. [PubMed]
- Parrott AC, Choi PY, Davies M. Anabolisko steroīdu lietošana sportistiem amatieriem: ietekme uz psiholoģisko noskaņojumu. Sporta medicīnas un fiziskās sagatavotības žurnāls. 1994;34: 292-8. [PubMed]
- Patton PE, Novy MJ, Lee DM, Hickok LR. Diagnoze un reproduktīvais iznākums pēc pilnīgas dzemdes, dubulta dzemdes kakla un maksts starpsienas ķirurģiskas ārstēšanas. Amerikas Dzemdību un ginekoloģijas žurnāls. 2004;190: 1669 – 75. 1675 – 8. [PubMed]
- Paus T, Collins DL, Evans AC, Leonard G, Pike B, Zijdenbos A. Baltās vielas nogatavināšana cilvēka smadzenēs: magnētiskās rezonanses pētījumu pārskats. Prāta pētījumu biļetens. 2001;54: 255-66. [PubMed]
- Perijs PJ, Kutscher EC, Lunds BC, Yates WR, Holman TL, Demers L. Agresijas un garastāvokļa izmaiņu mēri vīriešu svarcēlājos ar androgēnu anabolisko steroīdu lietošanu un bez tās. Tiesu zinātņu žurnāls. 2003;48: 646-51. [PubMed]
- Peters KD, Wood RI. Androgēnu atkarība no kāmjiem: pārdozēšana, tolerance un iespējamie opioidergiskie mehānismi. Neirozinātne. 2005;130: 971-81. [PubMed]
- Pfaus JG, Damsma G, Nomikos GG, Wenkstern DG, Blaha CD, Phillips AG, Fibiger HC. Seksuālā uzvedība uzlabo centrālo dopamīna pārnešanu žurku tēviņiem. Smadzeņu izpēte. 1990;530: 345-348. [PubMed]
- Fīniksa CH, Goy RW, Gerall AA, Young WC. Pirmsdzemdību laikā ievadītā testosterona propionāta darbības organizēšana uz audiem, kas veicina jūrascūciņu pārošanās izturēšanos. Endokrinoloģija. 1959;65: 369-82. [PubMed]
- Pinos H, Collado P, Rodriguez-Zafra M, Rodriguez C, Segovia S, Guillamon A. Dzimumu atšķirību attīstība žurkas lokusā coeruleus. Prāta pētījumu biļetens. 2001;56: 73-8. [PubMed]
- Pāvests HG, Jr, Katz DL. Slepkavības un gandrīz slepkavības, ko izdarījuši anabolisko steroīdu lietotāji. [Skatīt komentāru] Klīniskās psihiatrijas žurnāls. 1990;51: 28-31. [PubMed]
- Pope HG, Jr, Katz DL. Anabolisko-androgēnu steroīdu lietošanas psihiskie un medicīniskie efekti. Kontrolēts 160 sportistu pētījums. Vispārējās psihiatrijas arhīvs. 1994;51: 375-82. [PubMed]
- Pāvests HG, Jr, Kouri EM, Powell KF, Campbell C, Katz DL. Anabolisko androgēno steroīdu lietošana ieslodzīto vidū 133. Visaptveroša psihiatrija. 1996;37: 322-7. [PubMed]
- Putnam SK, Du J, Sato S, Hull EM. Testosterona kopulatīvās uzvedības atjaunošana korelē ar vidēju preoptisku dopamīna izdalīšanos kastrētu žurku tēviņiem. Hormoni un izturēšanās. 2001;39: 216-224. [PubMed]
- Putnam SK, Sato S, Hull EM. Testosterona metabolītu ietekme uz kopulāciju un vidējo preoptisko dopamīna izdalīšanos kastrētu žurku tēviņos. Hormoni un izturēšanās. 2003;44: 419-26. [PubMed]
- Putnam SK, Sato S, Riolo JV, Hull EM. Testosterona metabolītu ietekme uz kopulāciju, mediālo preoptisko dopamīnu un NOS imūnreaktivitāti kastrētu tēviņu tēviņiem. Hormoni un izturēšanās. 2005;47: 513-522. [PubMed]
- Rankin SL, Partlow GD, McCurdy RD, Giles ED, Fisher KR. Pēcdzemdību neiroģenēze cūku hipotalāmu vazopresīnu un oksitocīnu saturošā kodolā. Smadzeņu izpēte. 2003;971: 189-96. [PubMed]
- Ricci LA, Rasakham K, Grimes JM, Melloni RH., Jr Serotonīna-1A receptoru aktivitāte un ekspresija modulē pusaudžu anabolisko / androgēno steroīdu izraisīto agresiju kāmjos. Farmakoloģija, bioķīmija un uzvedība. 2006;85: 1-11.
- Romeo RD, Cook-Wiens E, Richardson HN, Sisk CL. Dihidrotestosterons aktivizē seksuālo uzvedību pieaugušiem kāmju vīriešiem, bet ne mazuļiem. Fizioloģija un uzvedība. 2001;73: 579-84. [PubMed]
- Romeo RD, Diedrich SL, Sisk CL. Estrogēna receptoru imūnreaktivitāte prepubertāli un pieaugušiem Sīrijas kāmju vīriešiem. Neirozinātņu vēstules. 1999;265: 167-70. [PubMed]
- Romeo RD, Parfitt DB, Richardson HN, Sisk CL. Feromoni izraisa līdzvērtīgu Fos imūnreaktivitātes līmeni prepubertāli un pieaugušiem Sīrijas kāmjiem. Hormoni un izturēšanās. 1998;34: 48-55. [PubMed]
- Romeo RD, Richardson HN, Sisk CL. Pubertāte un vīriešu smadzeņu nobriešana un seksuālā uzvedība: uzvedības potenciāla pārstrādāšana. Neirozinātnes un bioloģiskās uzvedības atsauksmes. 2002a;26: 381-91. [PubMed]
- Romeo RD, Wagner CK, Jansen HT, Diedrich SL, Sisk CL. Estradiols inducē hipotalāma progesterona receptorus, bet pirms pubertātes neaktivizē pārošanās izturēšanos kāmju tēviņos (Mesocricetus auratus). Uzvedības neirozinātne. 2002b;116: 198-205. [PubMed]
- Salas-Ramirez KY, Montalto PR, Sisk CL. Anaboliskie androgēnie steroīdi (AAS) atšķirīgi ietekmē sociālo uzvedību pusaudžu un pieaugušu Sīrijas kāmju vīriešu vidū. Hormoni un izturēšanās. 2008 presē.
- SAMHSA / OAS. DHHS publikācijas Nr. (SMA) 1996. 1994 Nacionālais mājsaimniecību apsekojums par narkotiku lietošanu, galvenie secinājumi 1994; 96 – 3085 lpp.
- SAMHSA / OAS. Rezultāti no 2004 nacionālā apsekojuma par narkotiku lietošanu un veselību: nacionālie atzinumi. 2005. NSDUH sērija H-28, DHHS publikācijas Nr. SMA 05 – 4062.
- Sato SM, Johansen J, Jordan CL, Wood RI. Androgēnu pašpārvalde Tfm žurkām. Uzvedības neiroendokrinoloģijas biedrības gadskārtējā sanāksme.10.
- Sato SM, Wood RI. Membrānnecaurlaidīgu anabolisko-androgēno steroīdu (aas) pašievadīšana Sīrijas kāmjos. Uzvedības neiroendokrinoloģijas biedrības 11 gadskārtējā sanāksme.2007.
- Schroeder JP, Packard MG. Dopamīna receptoru apakštipu loma testosterona nosacītas vietas izvēles iegūšanā žurkām. Neirozinātņu vēstules. 2000;282: 17-20. [PubMed]
- Schulte HM, MJ zāle, Boyer M. Vardarbība ģimenē, kas saistīta ar anabolisko steroīdu ļaunprātīgu izmantošanu. American Journal of Psychiatry. 1993;150: 348. [PubMed]
- Šulcs KM, Menards TA, Smits DA, Albers HE, Sisk CL. Sēklinieku hormonu iedarbība pusaudža gados organizē sānu marķēšanu un vazopresīna receptoru saistīšanos sānu starpsienā. Hormoni un izturēšanās. 2006;50: 477-83. [PubMed]
- Schulz KM, Richardson HN, Romeo RD, Morris JA, Lookingland KJ, Sisk CL. Mediālas preoptiskas zonas dopamīnerģiskas reakcijas uz sieviešu feromoniem attīstās Sīrijas kāmja vīrieša pubertātes laikā. Smadzeņu izpēte. 2003;988: 139-45. [PubMed]
- Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk CL. Dzimumdziedzeru hormoni maskulinizē un mazina reproduktīvo uzvedību pubertātes laikā Sīrijas kāmja tēviņiem. Hormoni un izturēšanās. 2004;45: 242-9. [PubMed]
- Schulz KM, Sisk CL. Pubertālie hormoni, pusaudžu smadzenes un sociālās uzvedības nobriešana: Sīrijas kāmja mācības. Molekulārā un šūnu endokrinoloģija. 2006:254–255. 120–6.
- Schulz KM, Zehr JL, Salas-Ramirez KY, Sisk CL. Neirozinātņu sanāksmju plānotājs. Sandiego, CA: Neirozinātnes biedrība; 2007. Vai pusaudža vecums ir otrs jutīgais periods testosterona organizējošajai ietekmei uz pieaugušu vīriešu reproduktīvo uzvedību? 2007 tiešsaistē.
- Skots JP, Stjuarts JM, De Gets VJ. Kritiskie periodi sistēmu organizācijā. Attīstības psihobioloģija. 1974;7: 489-513. [PubMed]
- Shughrue PJ, Lane MV, Merchenthaler I. Estrogēna receptoru alfa un beta beta mRNS salīdzinošais sadalījums žurku centrālajā nervu sistēmā. Salīdzinošās neiroloģijas žurnāls. 1997;388: 507-25. [PubMed]
- Simons-Mortons BG, Haynie DL. Sesto pakāpju smēķēšanas pakāpes psihosociālie prognozētāji. American Journal of Health Behavior. 2003;27: 592-602. [PubMed]
- Sisk CL, Zehr JL. Pubertālie hormoni organizē pusaudžu smadzenes un uzvedību. Neuroendokrinoloģijas robežas. 2005;26: 163-74. [PubMed]
- Sowell ER, Thompson PM, Leonard CM, Welcome SE, Kan E, Toga AW. Galvas smadzeņu garozas biezuma un smadzeņu augšanas kartēšana normāliem bērniem. Neiroloģijas žurnāls. 2004;24: 8223-31. [PubMed]
- Sowell ER, Thompson PM, Tessner KD, Toga AW. Turpināt smadzeņu augšanu un pelēkās vielas blīvuma samazināšanos dorālā frontālā garozā: inversijas attiecības pēcdzemdību smadzeņu nobriešanas laikā. Neiroloģijas žurnāls. 2001;21: 8819-29. [PubMed]
- Spear LP. Pusaudžu smadzeņu un ar vecumu saistītās uzvedības izpausmes. Neirozinātnes un bioloģiskās izturēšanās pārskats. 2000;24: 417-63.
- Su TP, Pagliaro M, Schmidt PJ, Pickar D, Wolkowitz O, Rubinow DR. Anabolisko steroīdu neiropsihiatriskā iedarbība normāliem vīriešiem vīriešiem. JAMA. 1993;269: 2760-4. [PubMed]
- Thiblin I, Finn A, Ross SB, Stenfors C. Palielināta dopamīnerģiskā un 5-hidroksitriptaminerģiskā aktivitāte žurku tēviņu smadzenēs pēc ilgstošas ārstēšanas ar anaboliskiem androgēniem steroīdiem. Lielbritānijas žurnāls par farmakoloģiju. 1999;126: 1301-6. [PMC bezmaksas raksts] [PubMed]
- Thomas P, mērci G, Pang Y, Berg H, Tubbs C, Benninghoff A, Doughty K. Progestin, estrogēnu un androgēnu G-proteīnu saistītie receptori zivju sugās. Steroīdi. 2006;71: 310-6. [PubMed]
- Tricker R, Casaburi R, Storer TW, Clevenger B, Berman N, Shirazi A, Bhasin S. Testosterona suprafizioloģisko devu ietekme uz dusmīgu uzvedību veseliem eugonadal vīriešiem - klīnisko pētījumu centra pētījums. Klīniskās endokrinoloģijas un metabolisma žurnāls. 1996;81: 3754-8. [PubMed]
- Triemstra JL, Nagatani S, Wood RI. Ķīmiski jutīgās norādes ir būtiskas, lai pārošanās izraisītu dopamīna izdalīšanos Sīrijas kāmju tēviņu MPOA. Neuropsychopharmacology. 2005;30: 1436-42. [PubMed]
- Triemstra JL, Sato SM, Wood RI. Testosterons un kodols accumbens dopamīnu vīriešu Sīrijas kāmja. Psihoneiroendokrinoloģija (presē)
- Van Eenoo P, Delbeke FT. Dopinga izplatība Flandrijā salīdzinājumā ar dopinga izplatību starptautiskajos sporta veidos. Starptautiskais sporta medicīnas žurnāls. 2003;24: 565-570. [PubMed]
- WADA. Akreditēto laboratoriju ziņojumi par nelabvēlīgiem analīzes rezultātiem. 2006. http://www.wada-ama.org.
- Wesson DW, McGinnis MANS. Anabolisko androgēno steroīdu (AAS) sakraušana pubertātes laikā žurkām: neiroendokrīna un uzvedības novērtējums. Farmakoloģija, bioķīmija un uzvedība. 2006;83: 410-9.
- Wichstrom L, Pedersen W. Anabolisko-androgēno steroīdu lietošana pusaudža gados: uzvarēt, izskatīties labi vai būt slikti? Alkohola pētījumu žurnāls. 2001;62: 5-13. [PubMed]
- Koksnes RI, Džonsons LR, Ču L, Šads C, Self DW. Testosterona pastiprināšana: intravenoza un intracerebroventrikulāra pašregulācija žurku tēviņiem un kāmjiem. Psihofarmakoloģija. 2004;171: 298-305. [PubMed]
- Koka RI, Newman SW. Androgēnu un estrogēnu receptori vienlaikus pastāv atsevišķos neironos Sīrijas kāmja smadzenēs. Neuroendokrinoloģija. 1995;62: 487-97. [PubMed]
- Koka RI, Swann JM. Ķīmiski jutīgo un hormonālo signālu neironu integrācija, kas kontrolē vīriešu seksuālo izturēšanos. In: Wallen K, Schneider JS, redaktori. Reproducēšana kontekstā. MIT prese; Kembridža: 1999. 423 – 444 lpp.
- Zahm DS, Heimer L. Divi transpalīdzīgi ceļi, kuru izcelsme ir žurkas kodolā uzkrāšanās. Salīdzinošās neiroloģijas žurnāls. 1990;302: 437-46. [PubMed]