KOMEN YBOP: Kesan Coolidge berada di sebalik kehebatan pornografi Internet. Kesan Coolidge adalah fenomena yang dilihat pada spesies mamalia di mana lelaki (dan pada tahap yang lebih rendah wanita) menunjukkan minat seksual yang diperbaharui jika diperkenalkan kepada pasangan seksual yang baru, walaupun setelah menolak hubungan seks dari pasangan seksual sebelumnya tetapi masih ada. Kebaruan seksual mengatasi kebiasaan ini dengan kegembiraan yang baru disebabkan oleh dopamin yang lebih tinggi. Aliran kebaruan yang berterusan adalah yang menjadikan pornografi Internet sangat berbeza dengan pornografi masa lalu.
- Dennis F. Fiorino,
- Ariane Coury, dan
- Anthony G. Phillips
- Jurnal Neurosains, 15 Jun 1997, 17 (12): 4849-4855;
Abstrak
Kesan Coolidge menerangkan pengekalan tingkah laku seksual dalam haiwan yang "seksual memuaskan" sebagai tindak balas kepada pasangan baru yang diterima. Memandangkan peranan sistem mesoprak dopamin (DA) dalam inisiasi dan penyelenggaraan tingkah laku yang bermotivasi, mikrodialisis digunakan untuk memantau transmisi nukleus accumbens (NAC) semasa pengekalan, ketegangan seksual, dan pemulihan tingkah laku seksual. Dalam persetujuan dengan laporan terdahulu, pembentangan wanita estrous di sebalik skrin dan perkongsian dikaitkan dengan peningkatan ketara dalam NAC DA efflux. Pemulangan kepekatan NAC DA kepada nilai asas bertepatan dengan tempoh ketegangan seksual, walaupun kepekatan metabolit DA, asid dihydroxyphenylacetic dan asid homovanillic, kekal tinggi. Pembentangan seorang wanita yang menerima novel yang baru di belakang skrin menghasilkan sedikit peningkatan dalam NAC DA, yang ditambah dengan ketara semasa pembaharuan yang diperbaharui dengan novel wanita. Data yang ada sekarang menunjukkan bahawa sifat rangsangan dari seorang wanita yang baru diterima boleh meningkatkan transmisi NAC DA dalam tikus jantan yang seksual, dan ini pula berkaitan dengan pengekalan kelakuan seksual.
Pengenalan
Tikus jantan yang telah berpisah hingga kenyang dapat diinduksi menjadi pasangan lagi jika wanita awal digantikan dengan wanita baru yang menerima. Ini telah dikenali sebagai kesan Coolidge dan telah diperhatikan dalam beberapa spesies mamalia (Wilson et al., 1963). Faktor-faktor umum seperti keletihan atau kemurungan motorik tidak mencukupi untuk menjelaskan keadaan seksual yang jelas, kerana rangsangan dari seorang wanita novel masih dapat mendorong penyertaan. Ketegangan seksual juga boleh "dibalikkan" secara farmakologi, ke tahap yang signifikan, oleh pentadbiran pelbagai ubat yang boleh bertindak pada sistem neurotransmitter yang berbeza. Ubat-ubatan ini termasuk yohimbine, 8-OH-DPAT (Rodriguez-Manzo dan Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus dan Gorzalka, 1987; Rodriguez-Manzo dan Fernandez-Guasti, 1995a,b), dan apomorfin (Mas et al., 1995c). Walaupun tindakan periferi ubat-ubatan ini tidak dapat dikurangkan (contohnya, kesan adrenergik pada fungsi ereksi), kesan pada mekanisme pusat yang mendasari ketegangan seksual telah dicadangkan berdasarkan eksperimen lesi sentral noradrenergik terpilih (Rodriguez-Manzo dan Fernandez-Guasti, 1995a) dan eksperimen mikrodialysis yang memantau metabolisme dopaminergik di kawasan preoptic medial (Mas et al., 1995a,b).
Memandangkan mekanisme pusat dapat mengetengahkan ciri-ciri kelakuan seksual kesan Coolidge, calon yang mungkin adalah sistem dopamine mesolimbi (DA), yang mengunjurkan dari kawasan tegegalal ventral ke NAC. Mesolimbic DA seolah-olah bertindak sebagai modulator utama dalam proses integratif yang kompleks yang melibatkan penilaian rangsangan alam sekitar, seperti isyarat dari wanita yang menerima seksual, dan organisasi tingkah laku yang diarahkan dengan matlamat, termasuk persamaan (Fibiger dan Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Walaupun neuron DA tengah bertindak balas terhadap ganjaran utama dan petunjuk ramalan ganjaran, novel atau rangsangan alam sekitar yang tidak dapat diramalkan mendorong pengaktifan neuron yang paling mantap selama sesi latihan berulang (Fabre et al., 1983; Schultz, 1992; Mirenowicz dan Schultz, 1994). Terdapat banyak bukti yang menyokong peranan penting bagi DA mesolimbi dalam permulaan dan penyelenggaraan perilaku seks tikus (Pfaus dan Everitt, 1995), dan beberapa laporan kajian mikrodialisis meningkat dalam NAC DA efflux semasa faset selera dan perilaku seksual laki-laki (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Walau bagaimanapun, terdapat sedikit data mengenai korelasi saraf neurokimia dan pengekalan tingkah laku seksual. Pemakaian dalam vivo microdialysis untuk memantau gangguan neurotransmitter mesolimbic semasa kesan Coolidge memberikan peluang yang unik untuk mengkaji peranan NAC DA dalam perkongsian, ketegangan seksual, dan pengekalan penyertaan.
Percubaan microdialysis dilakukan untuk menentukan perkara berikut: (1) sama ada permulaan keseronokan seksual disertai dengan pemulangan kepekatan DA ekstrasel dalam NAC kepada nilai-nilai preopulation atau di bawah, dan (2) sama ada pengembalian semula kelakuan copulatory dalam " seksual "tikus lelaki dengan wanita yang baru menerima kaitan dengan kenaikan NAC DA efflux.
BAHAN DAN KAEDAH
Subjek. Tikus Sprague Dawley lelaki, yang diperolehi dari Pusat Penjagaan Haiwan (di Universiti British Columbia), dan tikus Long-Evans wanita, yang diperolehi dari Charles River Canada (St Constant, Quebec, Kanada), ditempatkan dalam kandang sangkar kawat (18 × 25 × 65 cm; lima setiap sangkar) di bilik koloni berasingan. Bilik-bilik koloni dikekalkan pada suhu ~ 20 ° C pada kitaran cahaya / siklus gelap 12 terbalik. Tikus mempunyai akses tanpa had kepada makanan (Purina Rat Chow) dan air.
Pembedahan dan ujian tingkah laku sebelum microdialysis otak.Tikus betina dilahirkan secara ovariectomized bilateral di bawah anestesia gas halothane (Fluothane, Ayerst Laboratories) sekurang-kurangnya 4 minggu sebelum ujian. Kesanggapan seksual dalam wanita rangsangan adalah disebabkan oleh suntikan subkutaneus estradiol benzoat (10 μg) dan progesteron (500 μg), 48 dan 4 jam, masing-masing, sebelum setiap sesi ujian. Tikus jantan telah disaring untuk tingkah laku seksual pada dua kesempatan, 4 d terpisah, di ruang Plexiglas (35 × 35 × 40 cm) dengan lantai mesh wire. Hanya tikus jantan yang mencapai kriteria prestasi, yang termasuk intromisi dalam min 5 penyampaian wanita dan ejakulasi dalam min 15 dari intromisi pertama, semasa ujian dua ujian ditanamkan dengan panduan probe microdialysis cannulae.
Tikus lelaki (n = 5) dibiakkan dengan ketamine hydrochloride (100 mg / kg, ip) dan xylazine (10 mg / kg, ip) sebelum pembedahan stereotaxic. Pemeriksaan probe mikrodialysis cannulae (tolok 19) ditanam secara bilateral ke atas NAC (koordinat dari bregma: anterior, + 1.7 mm; medial, ± 1.1 mm; ventral, -1.0 mm; tengkorak datar) dan dijamin ke tengkorak dengan akrilik gigi dan skru tukang jahit. Panduan dua hala implan cannulae digunakan untuk memaksimumkan peluang untuk percubaan microdialysis yang berjaya. Mujurlah, dalam eksperimen ini, hanya satu kanula diperlukan untuk setiap tikus. Tikus lelaki ditempatkan secara individu dalam sangkar plastik besar dengan tempat tidur corncob untuk selebihnya eksperimen. Satu minggu selepas pembedahan, tikus diuji untuk tingkah laku seksual. Dalam bahagian latihan ini, ruang ujian dilengkapi dengan skrin Plexiglas gelongsor yang membahagikan ruang ke dalam ruang besar dan kecil. Tikus lelaki diperkenalkan ke dalam petak besar dan min 15 kemudian, seorang wanita diletakkan di belakang layar. Selepas tempoh persediaan min 15, skrin telah dialih keluar, dan tikus-tikus dibenarkan untuk mengganti kotor untuk min 30. Tiga sesi latihan dijalankan, satu setiap 4 d. Semua tikus mencapai kriteria prestasi semasa setiap sesi.
Coolidge eksperimen kesan. Tikus telah ditanam secara unilateral dengan probe microdialysis 12-18 jam sebelum eksperimen kesan Coolidge dan diletakkan di ruang besar bilik uji dengan akses percuma ke makanan dan air. Pada pagi percubaan, sampel mikrodialisis dikumpulkan setiap min 15. Eksperimen ini terdiri daripada tujuh fasa berikut berturut-turut: (1) baseline (min min 60); (2) perempuan 1 di belakang skrin (min 15); (3) dengan wanita 1 sehingga tempoh min 30 diluluskan tanpa gunung; (4) pengenalan semula wanita 1 di belakang skrin (min 15); (5) akses kepada wanita 1 untuk tempoh min 15 yang disediakan tidak ada pemasangan (jika pemasangan telah berlaku, fasa ini dianggap sebagai fasa 3); (6) pengenalan wanita 2 di belakang skrin (min 15); 7) perkongsian dengan wanita 2 untuk min 60.
Kelakuan difilemkan dengan pencahayaan rendah menggunakan sistem video JVC dan diperhatikan pada monitor video yang terletak di luar bilik ujian. Ukuran standard tingkah laku seksual dicatat menggunakan komputer dan perisian yang sesuai (Holmes et al., 1987).
Selepas eksperimen mikrodialisis, haiwan diberi overdosis hidrat chloral dan perfusi dengan intraselin dengan garam dan formalin (4%). Otak dihiris dan dibekukan, dan, kemudian, bahagian coronal diwarnai dengan cresyl violet untuk menentukan penempatan probe microdialysis. Hanya tikus dengan penempatan probe dalam NAC digunakan untuk analisis tingkah laku dan neurokimia.
Mikrodialisis dan pengesanan elektrokimia HPLC. Probe mikrodialisis adalah reka bentuk sepusat dengan membran serat berongga semipermeable (membran 2 mm terdedah, diameter luar 340 μm, potongan berat molekul 65000, Filtral 12, Hospal) pada hujung distal. Probe telah diencerkan pada 1.0 μl / min dengan larutan Ringer yang diubahsuai (buffer 0.01 m sodium fosfat, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) menggunakan alat pemburu jarum (Hamilton, Reno, NV) dan pam jarring (model 22, Harvard Apparatus, South Natick, MA). Kolar panduan probe microdialysis digunakan untuk mengamankan probe microdialysis di dalam kannula panduan. Sebuah gegelung keluli, yang dipasang pada swivel cecair (Instech 375s) yang dipasang di atas ruang ujian, digunakan untuk melindungi tiub penyiasatan (Fiorino et al., 1993).
Analisis mikrodialisat, yang termasuk DA dan metabolitnya dihydroxhenylacetic acid (DOPAC) dan asid homovanillic (HVA), dipisahkan oleh kromatografi fasa terbalik (Kolom Ultrasphere; Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, diameter dalaman ) menggunakan penampan sodium acetate 0.083m, pH 3.5 (5% methanol). Penganalisis analisis dikira oleh pengesanan elektrokimia (EC). Alat ini terdiri daripada pam Bio-Rad (Richmond, CA), penyuntik dua posisi EC (EC10W) Valco Instruments (Houston, TX), pengesan Coulochem II EC ESA (Bedford, MA) dan perakam carta dua saluran (Kipp dan Zonen, Bohemia, NY). Parameter pengesan elektrokimia adalah seperti berikut: elektrod 1, + 450 mV; elektrod 2, -300 mV; dan sel penjaga, -450 mV. Pemeriksaan probe biasa, dilakukan vitro dan pada suhu bilik, adalah 22% untuk DA, 18% untuk DOPAC, dan 18% untuk HVA.
KEPUTUSAN
Kelakuan
Langkah-langkah kelakuan dari eksperimen kesan Coolidge dibentangkan dalam Jadual 1. Latihan untuk melekakan, mengikat, dan melancarkan ejakulasi, serta selang postakaculatori selepas ejakulasi pertama adalah serupa dengan sesi latihan sebelumnya (data tidak ditunjukkan). Ini menunjukkan bahawa prosedur microdialysis tidak mengubah tingkah laku seksual yang normal. Pengembangan keseronokan seksual, seperti yang ditunjukkan oleh bilangan purata ejakulasi sebelum kriteria dipenuhi (7.8 ± 0.5), penurunan progresif dalam bilangan intromisi sebelum setiap ejakulasi, dan peningkatan progresif dalam selang postejaculatory (data tidak ditunjukkan) , adalah serupa dengan yang dilaporkan dalam kajian terdahulu (Pantai dan Jordan, 1956; Fowler dan Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo dan Fernandez-Guasti, 1994; Mas et al., 1995d). Variabiliti individu diperhatikan sehubungan dengan bilangan ejakulasi yang dicapai dengan wanita 1, masa yang digunakan untuk bersalin dengan wanita 1, dan bilangan persembahan wanita 1 diperlukan untuk mencapai kriteria pemasahan (Jadual 1, bawah). Sesetengah tikus memerlukan banyak pengenalan semula 1 wanita sehingga fasa 5 selesai (n = 3). Tindakan meletakkan wanita 1 di belakang skrin dan penyingkiran partition itu mungkin berfungsi sebagai isyarat utama yang membawa kepada penyertaan. Ia juga perlu diperhatikan bahawa kriteria pemadatan min 30 tanpa gunung, walaupun digunakan sebelum ini (Pantai dan Jordan, 1965; Mas et al., 1995b), adalah sewenang-wenang dan tidak menjamin bahawa tikus tidak akan dipasang diberikan lebih banyak masa. Walau bagaimanapun, kelewatan atau penyingkiran dan prosedur penggantian tidak dapat dipercaya dalam perkongsian yang diperbaharui dengan 1 perempuan (contohnya, fasa 4 dan 5).

Kelakuan semasa eksperimen kesan Coolidge
Semua tikus mempamerkan kesan Coolidge. Aktiviti yang berkaitan dengan penempatan 2 perempuan di belakang skrin dan, khususnya, penyingkiran partition mungkin telah menyumbang kepada hasil ini, tetapi, sekali lagi, peristiwa-peristiwa ini tidak mencukupi untuk memperbaharui penyertaan sebelum eksperimen. Perbandingan antara langkah-langkah tingkah laku seksual dengan perempuan 1 dan wanita 2 dibuat menggunakan t ujian dengan pembetulan Bonferroni. Walaupun latensi gunung dan intromisi sebagai tindak balas terhadap 2 wanita tidak berbeza dengan ketara daripada mereka yang mengalami pertarungan positif dengan wanita 1, secara amnya, tingkah laku seksual dengan 2 wanita kurang kuat, seperti ditunjukkan oleh ejakulasi yang jauh lebih sedikit (min, 0.6 vs 4.2; F = 49.86;p <0.01) dan kemasukan (min = 11.2 vs 37.0;F = 20.17; p <0.05) pada jam pertama. Bilangan kenaikan pada jam pertama dengan wanita 1 dan 2 tidak berbeza secara signifikan.
Adalah penting untuk diperhatikan bahawa wanita yang digunakan semasa bahagian percubaan eksperimen (iaitu, wanita 1) masih mempamerkan tingkah laku yang kronik (iaitu, melompat dan melenggang) yang kuat dan tingkah laku (iaitu, lordosis) yang kuat untuk tempoh lengkap hubungan mereka dengan lelaki .
Neurokimia
Kepekatan nanomolar asas DA dan metabolitnya dalam microdialysates, dibentangkan sebagai min ± SEM, tiga sampel asas pertama adalah: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; dan HVA, 234.2 ± 49.0 (tidak dikesan untuk pemulihan probe;n = 5). Nilai ini mewakili skor asas bas 100.
Titik data yang ditakrifkan, bersesuaian dengan setiap fasa percubaan dan biasa kepada setiap tikus, digunakan untuk analisis neurokimia. Ini termasuk tujuh contoh berikut: (1) tujuh sampel selepas pengenalan pertama perempuan 1, (2) empat sampel yang mengiringi ketiadaan perilaku copulatory dengan wanita 1, dan (3) lima sampel selepas pembentangan wanita 2. Rajah 1 menggambarkan perubahan kepekatan DA (graf garis, tengah) dan metabolit DA (graf garis, bahagian) tingkah laku pengalihan paralel (graf bar, bawah) semasa ujian untuk kesan Coolidge.
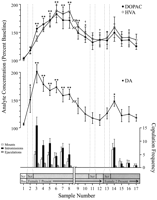
Lihat versi yang lebih besar:
Rajah. 1.
Nukleus mengakui hubungan neurokimia tingkah laku seksual semasa kesan Coolidge. Lapan sampel pertama mewakili titik data berterusan secara kronologi dari fasa 1 hingga 3. Sampel 1 adalah contoh baseline precopulation keempat dan terakhir (Bas). Contoh 2 mewakili pengenalan wanita 1 di belakang skrin (Scr). Selepas 15 min, skrin telah dialih keluar, dan tikus dibenarkan untuk mengawan (sampel 3-8). Thememecahkan pada x-axis sepadan dengan pengecualian data daripada tiga tikus yang disalin untuk tempoh yang panjang dengan wanita awal. Sembilan sampel terakhir juga berterusan secara kronologi. Sampel 9 dan 10 sepadan dengan tempoh pemasaan fasa 3 (iaitu, 30 min tanpa mount). 1 perempuan kemudian dimasukkan semula di belakang skrin (sampel 11) dan, 15 min kemudian, skrin telah dikeluarkan (sampel 12). Selepas 15 tanpa penyertaan, 2 perempuan diletakkan di belakang skrin (contoh 13). Sampel 14-17 sesuai dengan persamaan dengan 2 perempuan. Bilangan pemasangan, intromisi, atau ejakulasi yang berkaitan dengan setiap sampel microdialysis min 15 ditunjukkan di dalam graf bar bawah. Data neurokimia dinyatakan dari segi peratusan kepekatan baseline. Perubahan dalam NAC DA (petak tertutup), DOPAC (lingkaran tertutup), dan HVA (bulatan terbukaefflux dibentangkan sebagai graf garisan. Perbandingan berikut dibuat: sampel asas 1 berbanding sampel 2-10; sampel asas 10 berbanding sampel 11 dan 12; sampel asas baru 12 berbanding sampel 13-17 (*p <0.05; ** p <0.01). Bebas t Ujian dibuat antara nilai asas (sampel 1, 10, dan 12). Untuk perbezaan yang signifikan dari garis asas pertama (sampel 1), †p <0.05.
Mengasingkan satu arah, langkah-langkah ANOVA berulang dilakukan pada data neurokimia yang berkaitan dengan 1 wanita (sampel 1-12) dan 2 wanita (sampel 12-17). A priori perbandingan dibuat menggunakan ujian perbandingan pelbagai Dunn (Bonferroni t). Tiga perbandingan berikut telah dibuat: (1) baseline awal (sampel 1) berbanding sampel 2-10 (pendedahan pertama kepada wanita 1), (2) baseline kedua (sampel 10) berbanding sampel 11 dan 12 (reexposure kepada 1 perempuan) , dan (3) baseline ketiga (contoh 12) berbanding sampel 13-17 (pendedahan kepada 2 wanita).
Terdapat perubahan besar dalam DA efflux sebagai tindak balas kepada wanita 1 [F (11,44) = 8.48; p <0.001] dan wanita 2 [F (5,20) = 2.83;p <0.05]. Peningkatan ketara dalam eflux DA didapati ketika wanita 1 hadir di belakang skrin (+ 44%,p <0.05; contoh 2). Semasa kopulasi, kepekatan DA meningkat lebih jauh, mencapai nilai maksimum (+ 95%;p <0.01) semasa pertarungan kopulatif pertama (sampel 3). DA tetap meningkat sepanjang kopulasi dan hanya kembali ke kepekatan awal dalam tempoh 30 min di mana tidak ada pemasangan berlaku (sampel 9 dan 10). Baik pengenalan semula wanita 1 di belakang skrin (sampel 11) atau peluang untuk berinteraksi secara fizikal, tetapi tanpa pemasangan (sampel 12), peningkatan kepekatan DA relatif terhadap nilai asas kedua (sampel 10). Kehadiran wanita 2 di belakang skrin (sampel 13) menghasilkan peningkatan kecil dalam eflux DA (12%) dari nilai awal ketiga (sampel 12) yang tidak mencapai kepentingan statistik. Perbandingan yang diperbaharui dengan wanita 2 menghasilkan peningkatan (34%) yang ketara (p <0.05) dalam aliran masuk DA semasa sampel kopulasi pertama (sampel 14). Walaupun tingkah laku penyusutan yang lemah berlanjutan selama tiga sampel berikutnya, kepekatan DA menurun menjadi nilai awal (sampel 15-17). Bebas t Ujian yang dijalankan di antara sampel "baseline" (iaitu, 1, 10, dan 12) menunjukkan bahawa nilai-nilai ini tidak jauh berbeza.
Dalam ketiga-tiga tikus yang menyambung semula perkongsian apabila 1 wanita diperkenalkan semula, konsentrasi NAC DA meningkat apabila 1 wanita berada di belakang skrin (julat, 25-47%) dan semasa copulasi (julat, 13-37%), berbanding sampel sahaja sebelum pengenalan semula wanita. Walau bagaimanapun, kenaikan ini hanya berlaku apabila tingkah laku seksual bertambah kuat dan menyebabkan ejakulasi.
Perubahan keseluruhan yang ketara dalam DOPAC [F (11,44) = 9.57; p <0.001] dan HVA [F (11,44) = 12.47; p <0] kepekatan didapati sebagai tindak balas terhadap wanita 001, tetapi bukan wanita 1. Kepekatan metabolit meningkat sedikit (+ 2% dalam kedua-dua kes) semasa persembahan wanita 15 di belakang skrin (sampel 1), tetapi ini tidak signifikan secara statistik. Terdapat, bagaimanapun, peningkatan yang signifikan dalam kepekatan DOPAC dan HVA semasa kopulasi (sampel 2-3), mencapai nilai maksimum (+8 dan + 80%, masing-masing; p <0.01) selepas 60 min (sampel 6 dalam kedua kes). Walaupun kepekatan metabolit menurun dalam tempoh ketidakaktifan seksual pada akhir hubungan dengan wanita 1 (sampel 9 dan 10), kepekatan masih tetap meningkat sehubungan dengan tahap awal (p <0.05 dalam kedua-dua kes). Pengenalan semula wanita 1 di belakang skrin (sampel 11), akses ke wanita 1 setelah penyingkiran skrin (sampel 12), dan pengenalan wanita 2 (sampel 13) tidak menyebabkan perubahan dalam kepekatan metabolit. Sedikit, tetapi tidak signifikan secara statistik, peningkatan kepekatan DOPAC dan HVA (+ 23% dalam kedua-dua kes) berbanding dengan garis dasar (sampel 12) sesuai dengan pertarungan pertama dengan wanita 2 (sampel 14). Peningkatan ini hanya berumur pendek, dan menurun menjadi nilai awal untuk tiga sampel yang tinggal (15-17). Bebas tUjian yang dijalankan di antara sampel "baseline" (iaitu, 1, 10, dan 12) menunjukkan bahawa nilai baseline kedua dan ketiga (sampel 10 dan 12), walaupun tidak berbeza antara satu sama lain, kekal tinggi berbanding dengan sampel asas pertama untuk DOPAC dan HVA (p <0.05 dalam kedua-dua kes).
Histologi
Probe mikrodialisis terletak di NAC (Gamb.2) dalam pelbagai memperluas + 1.20 hingga + 1.70 mm dari bregma (tengkorak datar). Terdapat kebolehubahan juga dalam pesawat mediolateral; data mencerminkan pensampelan dari subkelen shell dan inti NAC.
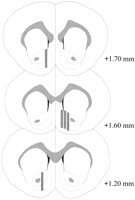
Lihat versi yang lebih besar:
Rajah. 2.
Lokasi pemeriksaan microdialysis dalam NAC tikus lelaki yang digunakan dalam eksperimen kesan Coolidge. Segi empat berbayang sesuai dengan kawasan membran terdedah daripada probe microdialysis. Bahagian otak coronal telah direkod semula dariPaxinos dan Watson (1986).
PERBINCANGAN
Dalam persetujuan dengan laporan terdahulu, keputusan sekarang menunjukkan peningkatan mesolimbic DA yang dikaitkan dengan komponen appetitive dan consummatory dari kelakuan seks tikus lelaki seperti yang dinilai olehdalam vivo microdialysis (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Di samping itu, keputusan ini memberikan kaitan neurokimia untuk pembesaran seksual dan reaksi pemulihan berikutnya sebagai tindak balas terhadap wanita baru yang menerima (kesan Coolidge). Data-data ini menunjukkan bahawa sifat-sifat rangsangan dari wanita baru yang diterima boleh meningkatkan transmisi NAC DA dalam tikus jantan yang seksual yang mungkin pula berkaitan dengan pemulihan tingkah laku seksual. Ini pertama kali dilihat dalam sedikit peningkatan dalam NAC DA semasa pembentangan seorang wanita novel di belakang skrin dan berlaku paling meyakinkan sebagai peningkatan yang lebih ketara semasa pembaharuan diperbaharui dengan wanita 2 (Gamb.1).
Kehadiran wanita yang menerima awal di belakang skrin menghasilkan kenaikan appetitive yang kuat dalam NAC DA efflux (44% dari garis dasar) sama besarnya dengan apa yang dilaporkan dalam eksperimen terdahulu menggunakan reka bentuk yang sama (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992). Juga sesuai dengan kajian-kajian ini ialah pemerhatian bahawa eflux NAC DA ditingkatkan lebih jauh semasa penyusunan (hingga> 95% di atas garis dasar dalam eksperimen ini). Walaupun kita dapat melihat tingkah laku penuh sebagai berkaitan dengan pelepasan NAC DA yang dipertingkatkan (Wenkstern et al., 1993; Wilson et al., 1995), adalah penting untuk mengkaji istilah "appetitive" dan "consummatory" dalam konteks tingkah laku seksual. Manakala fasa di mana wanita yang hadir di belakang skrin adalah semata-mata appetitive atau persediaan, tingkah laku semasa fasa kopulasi tidak boleh dipertimbangkan semata-mata. Kerana "selera" boleh digunakan untuk menggambarkan semua tingkah laku yang membawa kepada penyempurnaan kelakuan bermotivasi (copulasi), tingkah laku utama lelaki pameran semasa aktif dalam fasa "tahap" adalah digambarkan sebagai selera; lelaki menghabiskan sebahagian besar waktunya dan usaha mengejar wanita itu untuk menggigit. Dalam hal ini, kita dapat mengaitkan penghantaran NAC DA maksimal dengan penyesuaian serta komponen appetitive yang sengit dari kelakuan seks tikus lelaki.
Akses kepada wanita yang kedua, menghasilkan penyertaan yang diperbaharui dalam setiap mata pelajaran. Kajian terdahulu telah menunjukkan bahawa majoriti tikus yang dibenarkan untuk menggigit dengan kenyang, menggunakan protokol tingkah laku yang sama dengan yang digunakan dalam eksperimen ini, tidak meneruskan perkahwinan apabila diuji 24 hr kemudian (Pantai dan Jordan, 1956). Ia mungkin bahawa kehadiran sifat rangsangan novel wanita 2, yang mungkin termasuk penciuman serta isyarat visual dan auditori, mengakibatkan pembaharuan yang diperbaharui. Persoalan yang menarik, yang masih harus dijawab, adalah dengan mekanisme mana tikus lelaki membezakan seorang wanita novel dari seorang wanita yang dia telah kawin baru-baru ini. Sebuah tapak untuk mekanisme itu boleh terletak pada sistem pencium utama. Telah dilaporkan bahawa integriti sistem ini adalah penting untuk kesan Coolidge dalam hamster (Johnston dan Rasmussen, 1984). Sistem pengawal aksesori vomeronasal, bagaimanapun, di mana proses memori pheromonal diterangkan baru-baru ini dalam tikus (Kaba et al., 1994), juga merupakan calon utama. Dalam hal ini, perlu diperhatikan bahawa peningkatan dalam penghantaran NAC DA diukur menggunakan dalam vivo voltammetry dalam tikus lelaki yang dibentangkan dengan katil yang terdedah kepada tikus betina dalam estrus (Louillot et al., 1991; Mitchell dan Gratton, 1992). Selain itu, permohonan K+ terus ke lapisan saraf vomeronasal mentol olfaktori aksesori, serta mentol olfaktik aksesori itu sendiri, mencukupi untuk meningkatkan penghantaran NAC DA (Mitchell dan Gratton, 1992).
Pertembungan min 15 yang pertama dengan wanita 2 dikaitkan dengan kenaikan ketara dalam NAC DA. Berbeza dengan 1 perempuan, interaksi dengan wanita 2 tidak menghasilkan kenaikan NAC DA pada magnitud yang sama semasa fasa (12%) atau fungsinya (34%). Ini peningkatan kecil dalam NAC DA, bagaimanapun, berkorelasi baik dengan tahap pengurangan tingkah laku seksual yang dipaparkan dengan 2 perempuan dibandingkan dengan wanita 1. Kepekatan metabolit kekal tinggi semasa fasa kenyang, menghasilkan kepekatan baseline baru (sampel 10 dan 12) yang dinaikkan dengan ketara dari nilai asas awal (contoh 1).
Sikap temporal dalam peningkatan konsentrasi DOPAC dan HVA semasa persandingan adalah selaras dengan pembentukan mereka sebagai metabolit senyawa induk, DA. Telah dicadangkan bahawa kepekatan metabolit mikrodialisis, sekurang-kurangnya semasa tingkah laku semulajadi yang tidak digerakkan secara farmakologi, menyediakan indeks aktiviti neural yang bergunaDamsma et al., 1992; Fumero et al., 1994). Fakta bahawa kepekatan metabolit kekal tinggi walaupun pada masa ketidakaktifan seksual dalam eksperimen ini, apabila konsentrasi DA telah kembali ke nilai asas pretest, membuat keraguan atas cadangan ini.
Ketinggian kepekatan metabolit DA yang dilihat dalam eksperimen ini mencerminkan medan preoptic area (mPOA) profil metabolit DA yang diperhatikan pada tikus pada hari pertama setelah mereka berpisah dengan pemakanan (Mas et al., 1995a,b). Ketinggian konsentrasi DOPAC dan HVA yang berterusan di NAC atau mPOA tidak selalu dipatuhi apabila tempoh mengawan itu adalah tempoh tetap, jauh lebih pendek daripada masa yang diperlukan untuk mencapai tahap. Sebagai contoh, banyak kajian telah menunjukkan bahawa kepekatan DOPAC meningkat dan kekal tinggi semasa persamaan tetapi menolak kepada nilai-nilai asas tidak lama selepas wanita itu dikeluarkan (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Dalam kajian oleh Mas et al. (1995b), kepekatan extracellular basal DOPAC dan HVA dalam mPOA kekal dinaikkan pada hari-hari berturut-turut 4 yang bersamaan dengan tempoh ketidakaktifan seksual. Pada hari keempat, sebelum haiwan kembali menyambung, kepekatan metabolit dasar adalah dekat dengan nilai presatiasi. Penulis menyamakan corak perubahan neurokimia kepada mereka yang dilihat selepas pentadbiran penghalang reseptor DA (Zetterström et al., 1984; Imperato dan DiChiara, 1985) dan telah mencadangkan bahawa keadaan ketidakaktifan seksual boleh diantarkan melalui pembebasan prolaktin, yang boleh bertindak sebagai "neuroleptik endogen" (Mas et al., 1995a,b,d). Adalah jelas bahawa pentadbiran neuroleptik disertai dengan peningkatan kepekatan metabolit ekstraselular dan DA efflux (Zetterström et al., 1984; Imperato dan DiChiara, 1985). Malangnya, Mas et al. (1995a,b) tidak dapat mengesan kepekatan mPOA DA. Dalam kajian ini, kepekatan DA di NAC kembali kepada nilai awal, sedangkan kepekatan DOPAC dan HVA kekal tinggi. Corak ini tidak konsisten dengan peranan untuk bertindak neuroleptik endogen di NAC untuk mendorong ketegangan seksual.
Memandangkan penglibatan neuron DA mesolimbi dalam tingkah laku yang bermotivasi (Fibiger dan Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) dan kepekaan mereka terhadap rangsangan alam sekitar baru (Fabre et al., 1983; Schultz, 1992; Mirenowicz dan Schultz, 1994), kenaikan yang diperhatikan dalam kepekatan ekstraselular NAC DA sebagai tindak balas terhadap novel wanita konsisten dengan hipotesis bahawa aktiviti dalam sistem DA ini adalah penting untuk menghidupkan kembali tingkah laku seksual. Di samping itu, laporan mengenai kenaikan selera makan dan penghantaran dalam penghantaran DA (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) dan aktiviti neuron (Shimura et al., 1994) dalam mPOA tikus lelaki semasa tingkah laku seksual menunjukkan bahawa struktur ini juga boleh menyumbang kepada ciri-ciri copensi yang diperbaharui kesan Coolidge.
Selaras dengan peranan umum sistem DA mesolimbi dalam tingkah laku yang bermotivasi, adalah jelas bahawa kepekatan ekstraselular DA juga dinaikkan sebelum, semasa, dan serta-merta selepas makan, dengan pulangan kepada nilai asas ~ 30 min kemudian (Wilson et al., 1995). Adalah diketahui bahawa ketenangan yang disebabkan oleh makanan dipengaruhi oleh sifat deria. Manusia dan haiwan menolak makanan di mana mereka diberi makan untuk kenyang dan menelan makanan lain yang belum dimakan (Rolls, 1986). Ini menimbulkan persoalan sama ada DA efflux ekstraselular di NAC akan meningkat secara selektif dengan penyampaian jenis makanan baru, tetapi bukan oleh makanan yang dimakan baru-baru ini untuk kenyang dengan cara yang sama dengan yang dilaporkan dalam kajian ini dalam konteks motivasi seksual. Sekiranya disahkan, hubungan umum ini antara sifat deria ganjaran semula jadi, rasa kenyang, dan penghantaran mesolimbic DA akan menjadi peranan penting bagi sistem saraf ini dalam pengawalan proses motivasi, gangguan yang boleh mengakibatkan gangguan makan dan fungsi seksual yang serius .
RUJUKAN
- ↵
- Pantai FA,
- Jordan L
(1956) Keletihan dan pemulihan seksual pada tikus lelaki. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Ciri-ciri temporal kesan Coolidge dalam tingkah laku perilaku tikus lelaki. J Comp Physiol Psikiatri 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Fungsi dopamine dalam tingkah laku selera dan pertahanan. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Tingkah laku seksual meningkatkan penghantaran dopamin dalam nukleus accumbens dan striatum tikus lelaki: perbandingan dengan kebaruan dan lokomosi. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Kegiatan neuron di wilayah tegegal ventral monyet berkelakuan. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Ganjaran, motivasi, kognisi: psikobiologi sistem dopamine mesotelencephalic. dalam Buku Panduan fisiologi: sistem saraf IV, edisi Bloom FE, Geiger SD (American Physiology Society, Bethesda, MD), pp 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Rangsangan elektrik bagi laman-laman ganjaran di kawasan tegegalal ventral meningkatkan penghantaran dopamin dalam nukleus accumbens tikus. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Kesan variasi rangsangan terhadap pemakanan seksual di tikus lelaki. J Comp Physiol Psikiatri 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Variasi dalam rangsangan insentif dan tingkah laku seksual di tikus lelaki. J Comp Physiol Psikiatri 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Perubahan dalam perolehan monoamine di kawasan forebrain yang dikaitkan dengan tingkah laku seksual lelaki: kajian microdialysis. Otak Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) Sistem pungutan data berasaskan IBM-PC untuk merakam tingkah laku seksual tikus dan untuk rakaman acara umum. Physiol Behav 44: 825-828.
- ↵
- Hull EM,
- Eaton RC,
- Musa J,
- Lorrain DS
(1993) Penipuan meningkatkan aktiviti dopamin di kawasan preoptic medial tikus lelaki. Sains hidup 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Dopamine ekstraselular dalam medan preoptik medial: implikasi untuk motivasi seksual dan kawalan hormon terhadap perkongsian. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Pembebasan dan metabolisme dopamine dalam tikus berjaga-jaga selepas neuroleptik sistemik seperti yang dikaji oleh dialisis trans-striatal. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Pengiktirafan individu hamster wanita oleh lelaki: peranan isyarat kimia dan sistem penciuman dan vomeronasal. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Memindah ingatan penciuman dengan pengaktifan reseptor glutamat metabotropik. Sains/Ilmu 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Hooks MS
(1993) Farmakologi dan litar saraf pemekaan kepada psikostimulus. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Mesoporticolimbic neuron dopaminergik. Peranan fungsian dan peraturan. dalam Psychopharmacology: generasi keempat kemajuan, ed edisi Bloom FE, Kupfer DJ (Raven, New York), pp 283-294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Rangsangan penenang yang berhubungan dengan seks menyebabkan peningkatan selektif dalam pembebasan dopamin dalam nukleus akut tikus lelaki. Otak Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) Peningkatan pembebasan dopamine dalam nukleus akumulasi penyebaran tikus jantan seperti yang terbukti dalam vivo voltammetry. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Kaitan neurokimia keletihan seksual dan pemulihan seperti yang dinilai oleh mikrodialisis vivo. Otak Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Pemantauan voltammetrik dan microdialysis mengenai pembebasan neurotransmitter monoamine otak semasa interaksi sosioseksual. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Induksi perilaku mengawan oleh apomorphine dalam tikus yang bersenggama secara seksual. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
(1995d) Neurokimia kegemukan seksual. Model eksperimen yang menghalang keinginan. dalam Farmakologi fungsi seksual dan disfungsi, ed Bancroft J (Raven, New York), pp 115-126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Kepentingan ketidakpastian untuk membalas ganjaran dalam neuron dopaminergik primate. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Pelepasan dopamin Mesolimbic yang ditimbulkan oleh pengaktifan sistem pengawal aksesori: satu kajian chronoamperometrik berkelajuan tinggi. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) Otak tikus dalam koordinat stereotaxic (2nd ed). (Akademik, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Tingkah laku seksual meningkatkan penghantaran dopamin pusat di tikus lelaki. Otak Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Psikofarmakologi tingkah laku seksual. dalam Psychopharmacology: generasi keempat kemajuan, ed edisi Bloom FE, Kupfer DJ (Raven, New York), pp 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioid dan tingkah laku seksual. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Hubungan neurobiologi keadaan emosi positif: dopamin, jangkaan dan ganjaran. dalam kajian semula Antarabangsa tentang emosi, ed Strongman (Wiley, New York), pp 31-50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Korelasi pelepasan dopamin dalam nukleus akrab dengan tingkah laku seksual maskulin dalam tikus. Otak Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Pembalikan keletihan seksual oleh ejen serotonergik dan noradrenergik. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Penyertaan sistem noradrenergik pusat dalam reestablishment tingkah laku copulatori tikus yang haus seksual oleh yohimbine, naloxone, dan 8-OH-DPAT. Bruce Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Antagonis opioid dan fenomena pemadaman seksual. Psychopharmacol 122: 131-136.
- ↵
- Rolls BJ
(1986) Ketegangan khusus sensori. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) Neurokimia perilaku motivasi: isu metodologi dan konseptual dalam kajian aktiviti dinamik nukleus akui dopamin. Kaedah Neurosci 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Pembebasan dopamine di kawasan preoptik medial semasa tingkah laku penopang lelaki pada tikus. Otak Res 692: 66-70.
- ↵
- Schultz W
(1992) Aktiviti neuron dopamine dalam primata berkelakuan. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) Kawasan preoptik medial terlibat dalam kedua-dua gairah seksual dan prestasi dalam tikus lelaki: penilaian semula aktiviti neuron dalam haiwan bergerak bebas. Otak Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Peningkatan penghantaran dopamine dalam nukleus akut tikus lelaki semasa pendedahan pertama mereka terhadap tikus betina yang berisiko seksual. Otak Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Hubungan dopaminergik terhadap tingkah laku motivasi: pentingnya memandu. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Pantai FA
(1963) Pengubahsuaian dalam tingkah laku seksual tikus lelaki yang dihasilkan dengan menukar wanita rangsangan. J Comp Physiol Psikiatri 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Kesan ubat neuroleptik pada pembebasan dopamine dan metabolisme striatal dalam tikus terjaga yang dikaji oleh dialisis intracerebral. Eur J Pharmacol 106: 27-37.
Artikel yang memetik artikel ini
- Neuroplasticity yang disebabkan oleh Opioid endogenous Neuron Dopaminergik di Kawasan Tegmental Ventral Pengaruh Ganjaran Alam dan Opiate Jurnal Neurosains, 25 Jun 2014, 34 (26): 8825-8836
- Ketidakseimbangan dalam kepekaan terhadap pelbagai jenis ganjaran dalam perjudian patologi Otak, 1 Ogos 2013, 136 (8): 2527-2538
- Akta Ganjaran Semulajadi dan Ubat Mengenai Mekanisme Plastik Neural Bersama dengan {Delta} FosB sebagai Pengantara Utama Jurnal Neurosains, 20 Februari 2013, 33 (8): 3434-3442
- Tindak balas Neuronal dalam Nucleus Accumbens Shell semasa Kelakuan Seksual dalam Tikus Lelaki Jurnal Neurosains, 1 Februari 2012, 32 (5): 1672-1686
- Cinta romantis: sistem otak mamalia untuk pilihan pasangan Transaksi filosofi Royal Society B: Sains Biologi, 29 Disember 2006, 361 (1476): 2173-2186
- Hedonic Hot Spots dalam Otak Ahli sains Neuroses, 1 Disember 2006, 12 (6): 500-511
- Modulasi oleh Central and Basolateral Amygdalar Nuclei Dopaminergic Correlates of Feeding to Satiety dalam Tikus Nucleus Accumbens and Medial Prefrontal Cortex Jurnal Neurosains, 15 Disember 2002, 22 (24): 10958-10965
- Kekerapan Transfusi Kepekatan Dopamine Meningkatkan Strok Pinggang Punggung dan Ventral Tikus Muda semasa Pengenalan Kepentingan Jurnal Neurosains, 1 Disember 2002, 22 (23): 10477-10486
- Kajian Buku: Dinamik Dopamine Ekstraselular dalam Tindakan Akut dan Kronik Cocaine Ahli sains Neuroses, 1 Ogos 2002, 8 (4): 315-322
- Penglibatan Berbeza NMDA, AMPA / Kainate, dan Dopamin Receptors di Inti Nukleus Accumbens dalam Perolehan dan Pencapaian Kelakuan Pendekatan Pavlovian Jurnal Neurosains, 1 Disember 2001, 21 (23): 9471-9477
- Kawalan tingkah laku kokain yang dicari oleh rangsangan ubat-ubatan yang berkaitan dengan tikus: Kesan pemulihan paras-paras dopamin yang bertindak balas dan bertindak balas pada amigdala dan nukleus accumbens PNAS, 11 April 2000, 97 (8): 4321-4326
- Fasilitasi Perilaku Seksual dan Enhanced Dopamine Efflux di Nukleus Accumbens Tikus Laki-laki setelah Sensitisasi Perilaku D-Amphetamine-Induced Jurnal Neurosains, 1 Januari 1999, 19 (1): 456-463
- NEUROSCIENCE: Mendapatkan Perhatian Otak Sains, 3 Oktober 1997, 278 (5335): 35-37
- Korelasi Dopaminergik Ketegasan Khusus Spesifik di Kortex Prefrontal Medial dan Nucleus Accumbens of the Rat Jurnal Neurosains, 1 Oktober 1999, 19 (19): RC29