Biol Psikiatri. 2008 Dec 1; 64 (11): 941-50. Epub 2008 Jul 26.
Teegarden SL, Nestler EJ, Bale TL.
Source
Jabatan Biologi Haiwan, Universiti Pennsylvania, Philadelphia, PA 19104-6046, Amerika Syarikat.
Abstrak
LATAR BELAKANG:
Sensitiviti terhadap ganjaran telah dikaitkan sebagai faktor predisposisi untuk tingkah laku yang berkaitan dengan penyalahgunaan dadah serta makan berlebihan. Walau bagaimanapun, mekanisme yang menyumbang untuk memberi ganjaran sensitiviti tidak diketahui. Kami membuat hipotesis bahawa disyskulasi dalam isyarat dopamin mungkin menjadi penyebab utama kepekaan ganjaran yang tinggi di mana rangsangan yang bermanfaat dapat bertindak untuk menormalkan sistem.
KAEDAH:
Kami menggunakan model tetikus genetik kepekaan ganjaran yang meningkat, tetikus Delta FosB-overexpressing, untuk memeriksa perubahan jalur ganjaran sebagai tindak balas kepada diet tinggi lemak yang enak. Penanda ganjaran memberi isyarat pada tikus ini diperiksa kedua-duanya pada dasarnya dan mengikuti minggu 6 pendedahan diet yang sesuai. Tikus telah diperiksa dalam ujian tingkah laku selepas penarikan diet tinggi lemak untuk menilai kelemahan model ini untuk menghilangkan rangsangan ganjaran.
KEPUTUSAN:
Keputusan kami menunjukkan pengaktifan laluan ganjaran yang diubahsuaikan di sepanjang litar kawasan accumbens-hypothalamic-ventral tegmental nukleus yang dihasilkan daripada overexpression Delta FosB di kawasan accumbens dan daerah yang striatal. Tahap unsur adenosin monophosphate (cAMP) fosforilasi kitaran yang mengikat protein (pCREB), faktor neurotropik yang berasal dari otak (BDNF), dan fosfoprotein dikendalikan dopamin dan siklik adenosin monophosphat yang dikendalikan dengan jisim molekul 32 kDa (DARPP-32) di dalam nukleus accumbens dikurangkan dalam tikus Delta FosB, yang menandakan isyarat dopamin berkurang. Enam minggu pendedahan diet tinggi lemak benar-benar memperbaiki perbezaan ini, mendedahkan keupayaan memberi ganjaran makanan yang enak. Tikus Delta FosB juga menunjukkan peningkatan yang ketara dalam aktiviti locomotor dan tindak balas yang berkaitan dengan kebimbangan 24 beberapa jam selepas penarikan lemak tinggi.
KESIMPULAN:
Keputusan ini membuktikan kepekaan yang mendasari perubahan dalam ganjaran yang berkaitan dengan disysulasikan Delta FosB dan isyarat dopamin yang boleh dinormalisasi dengan diet yang enak dan boleh menjadi fenotip predisposisi dalam beberapa bentuk obesiti.
Pengenalan
Walaupun kami meningkatkan pengetahuan tentang sistem saraf yang mengawal selera makan dan kenyang, kadar obesiti terus meningkat di Amerika Syarikat. Rawatan dadah semasa mempunyai keberkesanan yang terhad, dan pengubahsuaian tingkah laku mengalami pematuhan jangka panjang yang minimum (1). Pengambilan makanan yang kalori dan padat telah dikaitkan dengan perubahan stres dan jalur ganjaran di otak, menunjukkan bahawa ciri-ciri ganjaran makanan sedemikian boleh mengatasi isyarat imbangan tenaga (2-4). Makanan yang tinggi lemak bertindak sebagai ganjaran semulajadi, mengaktifkan pusat ganjaran otak dengan cara yang serupa dengan ubat penyalahgunaan, dan oleh itu telah digunakan dalam paradigma kendiri (5-8). Oleh itu, kemungkinan bahawa tingkah laku dan motivasi untuk makan berlebihan dan penyalahgunaan dadah berkongsi mekanisme asas yang sama, berpotensi membuka saluran baru rawatan untuk kedua-dua keadaan.
Dalam mengkaji hubungan antara makanan yang enak dan laluan yang mengawal ganjaran dan tekanan di otak, kami sebelum ini telah mengenal pasti penanda molekul dan biokimia ganjaran yang dikurangkan dan tekanan yang meningkat berikutan pengeluaran dari diet lemak tinggi (HF) yang lazat. Serupa dengan ubat-ubatan penyalahgunaan, pendedahan kepada diet yang enak dalam kajian kami menghasilkan peningkatan tahap faktor transkripsi ΔFosB dalam nukleus accumbens (NAc), struktur ganjaran otak utama (9, 10). Tikus yang menggambarkan lebih jelas ΔFosB memperlihatkan peningkatan instrumental untuk ganjaran makanan (11), menjadikannya alat yang berharga untuk mengkaji peranan kepuasan ganjaran dan disregulasi sistem ganjaran jangka panjang dalam tindak balas molekul dan biokimia terhadap diet yang enak.
Dalam kajian ini, kami menggunakan tikus ΔFosB-overexpressing untuk memeriksa perubahan jangka panjang dalam penanda ganjaran dalam neurocircuitry kawasan NAc-hypothalamus-ventral tegmental (VTA) sebagai tindak balas kepada diet HF yang enak. Berdasarkan kajian terdahulu dalam tikus-pahit yang sensitif ini, kami menghipnotiskan bahawa perubahan yang disebabkan oleh ΔFosB dalam kepekaan ganjaran melibatkan disistulasi dalam isyarat dopamin yang dihasilkan daripada maklum balas NAc kepada VTA. Selanjutnya, kami membuat hipotesis bahawa pendedahan kepada ganjaran semula jadi diet HF yang padat tenaga kemudian akan menormalkan sistem dopaminergik pada tikus-tikus ini, menyebabkan tindak balas yang berlebihan terhadap tekanan pengeluaran dari diet HF ini. Aspek unik menggunakan diet yang enak sebagai bahan yang bermanfaat membolehkan kita memasukkan input hipotalamik untuk memberi ganjaran litar dalam fenotip yang mungkin memprediksi populasi yang terdedah kepada obesiti yang tahan terhadap rawatan. Untuk mengkaji hipotesis ini, kami mempelajari penanda neurotransmiter dopamin, termasuk pCREB, BDNF, dan DARPP-32 dalam NAc dan tyrosine hydroxylase dan pengangkut dopamin dalam VTA, selepas pendedahan HF. Kami juga memeriksa tanda-tanda tertentu keseimbangan tenaga yang diketahui mempengaruhi pengeluaran dopamin, termasuk reseptor leptin dan orexin dalam ungkapan VTA dan orexin dalam hipotalamus lateral.
Bahan dan Kaedah
haiwan
Tikus bitransgenic lelaki yang secara mendadak menggambarkan ΔFosB dalam neuron positif dynorphin pada NAc dan striatum dorsal (Kelz et al., 1999) dihasilkan pada latar belakang bercampur (ICR: C57Bl6 / SJL) di Pusat Perubatan Southwestern Universiti Texas dan dikekalkan diuji di Universiti Pennsylvania. Semua tikus dikekalkan pada doxycycline (100 μg / ml dalam air minuman) sehingga tiba di Universiti Pennsylvania. Untuk mendorong overexpression, doxycycline dikeluarkan (n = 23) (12). Tikus kawalan (n = 26) terus menerima ubat. Tikus ditugaskan untuk kumpulan diet lapan minggu berikutan penghapusan doxycyline di mana ungkapan masa telah ditunjukkan untuk mencapai paras maksimal (13). Tikus dikekalkan pada kitaran 12: 12 cahaya gelap (lampu pada 0700) dengan makanan dan air yang terdapat iklan libitum. Semua kajian telah dilakukan mengikut protokol eksperimen yang diluluskan oleh Jawatankuasa Penjagaan dan Penggunaan Haiwan Institusi Universiti Pennsylvania, dan semua prosedur telah dijalankan mengikut garis panduan institusi.
Pendedahan diet
Tikus dikekalkan di rumah chow (n = 16) atau ditempatkan di HF (n = 16-17) selama enam minggu. Rumah chow (Purina Lab Diet, St. Louis, MO) mengandungi 4.00 kcal / g, yang terdiri daripada protein 28%, lemak 12% dan karbohidrat 60%. Diet HF (Diet Penyelidikan, New Brunswick, NJ) mengandungi 4.73 kcal / g, yang terdiri daripada protein 20%, lemak 45%, dan karbohidrat 35%.
Biokimia dan ungkapan gen
Tikus telah dianalisis selepas enam minggu pendedahan diet. Otak dikeluarkan dari tengkorak dan sama ada beku keseluruhan pada ais kering atau dibuang NAc (kira-kira 0.5 - 1.75 mm dari bregma, pada kedalaman 3.5 - 5.5 mm) dan dibekukan dalam nitrogen cecair. Tisu disimpan di -80 ° C sehingga diuji.
Analisis biokimia
Kaedah untuk pemecahan Barat diterangkan dalam bahan-bahan tambahan. Antibodi yang digunakan ialah: Cdk5, CREB, dan BDNF (1: 500, Santa Cruz Bioteknologi, Santa Cruz, CA) dan phospho-CREB (pCREB) (Ser 133) (1: 500, Teknologi Isyarat Sel, Danvers, MA).
Autoradiography reseptor
Kaedah terperinci untuk autoradiografi diterangkan dalam bahan tambahan. Ligands yang digunakan adalah 2 nM H3 - SCH 23390 dan 5 nM H3 - spiperone (PerkinElmer, Boston, MA).
Hibridisasi in situ
Pemprosesan dan hibrida tisu dilakukan seperti yang dinyatakan sebelumnya (14). Siasatan DARPP-32 diberikan dengan baik oleh P. Greengard (Rockefeller University), dan penyelidikan orexin oleh J. Elmquist (Pusat Perubatan Southwestern University of Texas). Slaid yang diuji untuk DARPP-32 telah dilampirkan untuk filem untuk hari-hari 3, dan slaid yang diuji untuk orexin telah digunakan untuk filem untuk hari-hari 4. Pengiraan imej filem telah dijalankan seperti yang dinyatakan sebelumnya (10).
QRT-PCR
RNA telah diasingkan dari VTA dan ungkapan gen individu yang dinilai menggunakan taifan ekspresi gen TaqMan (Applied Biosystems, Foster City, CA). Kaedah terperinci dan analisis statistik boleh didapati dalam bahan tambahan.
Analisis kelakuan
Untuk mengkaji kesan sensitiviti ganjaran terhadap perubahan tingkah laku yang disebabkan oleh diet, subset tikus telah ditarik balik dari HF berikutan pendedahan empat minggu dan dikembalikan kepada rumah chow (n = 9 control, n = 8 ΔFosB). Dua puluh empat jam selepas pengunduran, tikus terdedah kepada ujian lapangan terbuka sejajar dengan paradigma pengeluaran diet yang telah diterbitkan sebelum ini (10). Secara ringkas, tetikus diletakkan di tengah-tengah radas terbuka dan dipantau selama lima minit. Jumlah garis melintasi, biji fecal, masa di tengah, dan salib ke pusat diukur.
Statistik
Semua data kecuali Western blots dianalisis menggunakan ANOVA dua arah diikuti dengan ujian Fisher's PLSD dengan rawatan doxycycline (ΔFosB expression) dan keadaan diet sebagai pemboleh ubah bebas. Untuk analisis RT-PCR, penurunan nilai P digunakan untuk membetulkan beberapa perbandingan dalam kumpulan gen yang berkaitan (lihat bahan tambahan). Blot Barat dianalisis menggunakan ujian-t pelajar dengan perlakuan doxycycline sebagai pemboleh ubah tidak bersandar, membandingkan ketumpatan optik dalam titik yang sama. Semua data disajikan sebagai min ± SEM.
Hasil
Perbezaan biokimia asas
Untuk menjelaskan jalur molekul yang mendasari kepekaan ganjaran yang ditingkatkan pada tikus yang mengekspresikan ΔFosB, tahap beberapa molekul isyarat utama diperiksa di NAc. Terdapat kecenderungan peningkatan tahap Cdk5 pada NAc tikus ΔFosB berbanding dengan haiwan kawalan littermate yang dikekalkan pada doxycycline (F = 5.1, P = 0.08; Gambar 1A). Tikus ΔFosB menyatakan penurunan kadar pCREB (F = 7.4, P <0.05; Gambar 1B) dan tahap CREB total (F = 5.4, P = 0.05; Gambar 1C). Penurunan yang signifikan dalam BDNF juga diperhatikan pada NAc tikus ΔFosB (F = 10.6, P <0.05; Gambar 1D).
Rajah 1
Mice overexpressing ΔFosB mempamerkan penanda biokimia pengurangan isyarat dopamin dalam NAc
Pengambilan makanan dan berat badan pada diet tinggi lemak
Kami seterusnya meneliti kesan diet HF yang bermanfaat secara semula jadi terhadap perubahan molekul isyarat pada tikus yang mengekspresikan ΔFosB. Tidak ada perbezaan antara tikus ΔFosB dan kawalan dalam pengambilan makanan di rumah atau HF. Walau bagaimanapun, terdapat penurunan keseluruhan pengambilan kalori yang dinormalisasi ke berat badan ketika terkena HF yang khusus untuk tikus ΔFosB (F = 11.2, P <0.01; Gambar 2A). Pada akhir enam minggu pendedahan diet, tikus yang mendapat HF mempunyai berat badan yang jauh lebih tinggi daripada pada diet chow (F = 17.2, P <0.001), dan tikus ΔFosB mempunyai berat badan kurang keseluruhan daripada kawalan (F = 5.6, P <0.05; Gbr. 2B). Kesan ini khusus untuk perbezaan antara kumpulan pada diet chow (P <0.05).
Rajah 2
ΔFosB overexpressing tikus tidak menunjukkan perbezaan dalam pengambilan makanan pada sama ada chow atau diet lemak tinggi (HF)
Perbezaan biokimia pada diet tinggi lemak
Untuk menentukan bagaimana perbezaan dasar dalam isyarat NAc mungkin diubah oleh diet HF, protein isyarat yang sama yang dikaji pada peringkat awal diperiksa pada haiwan yang telah menerima enam minggu HF. Tidak terdapat perbezaan yang signifikan dalam tahap Cdk5 (Rajah 3A). Tahap pCREB dan jumlah CREB tidak lagi berbeza selepas enam minggu HF (Rajah 3B, C). Tahap BDNF meningkat dengan ketara dalam tikus ΔFosB selepas enam minggu pendedahan HF (F = 6.5, P = 0.05; 3D Rajah).
Rajah 3
Diet tinggi lemak (HF) perbezaan isyarat diperhatikan dalam NAc ΔFosB overexpressing tikus
Autoradiografi reseptor dopamin
Kami menggunakan autoradiografi reseptor untuk menilai sama ada perubahan yang disebabkan oleh ΔFosB dalam isyarat dopamin di NAc berkaitan dengan perubahan dalam ekspresi reseptor dopamin (Gbr. 4A). Diet tinggi lemak nampaknya sedikit meningkatkan kepadatan pengikatan reseptor dopamin D1 (P = 0.14), dan perbezaan ini lebih besar pada tikus ΔFosB (Gamb. 4B). Terdapat juga kecenderungan peningkatan kawasan pengikatan D1 setelah HF (P = 0.06), dan ujian post hoc menunjukkan ini signifikan pada tikus ΔFosB (P <0.05; Gambar. 4C). Berbeza dengan reseptor D1, tidak ada perubahan kepadatan pengikatan reseptor D2 (kawalan chow = 97.6 ± 6.9, kawalan HF = 101.1 ± 8.2, ΔFosB chow = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) atau kawasan pengikat (kawalan chow = 47.3 ± 3.4, kawalan HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) pada NAc diperhatikan.
Rajah 4
Diet tinggi lemak (HF) menyebabkan perubahan reseptor D1 receptor mengikat dan ekspresi DARPP-32 dalam nukleus accumbens (NAc) daripada ΔFosB overexpressing tikus
Ungkapan DARPP-32 dalam NAc
Hibridisasi in situ digunakan untuk menentukan tahap ekspresi DARPP-32 di NAc (Gambar 4D). Diet lemak tinggi secara signifikan meningkatkan ekspresi DARPP-32 di kawasan otak ini (F = 5.1, P <0.05), dan terdapat interaksi yang signifikan antara diet dan ekspresi ΔFosB (F = 8.9, P <0.05), dengan tikus ΔFosB menunjukkan lebih besar perubahan yang disebabkan oleh diet (Gamb. 4E). Perbezaan basal dalam ekspresi DARPP-32 antara tikus kawalan dan ΔFosB ditunjukkan oleh ujian post hoc (P <0.01), serta peningkatan ketara dalam ekspresi DARPP-32 pada tikus ΔFosB pada HF (P <0.01).
Ekspresi gen dalam VTA
QRT-PCR digunakan untuk menilai perubahan ekspresi gen di VTA, menargetkan beberapa gen utama yang sebelumnya terlibat dalam peraturan ganjaran. Semua sampel dinormalisasi ke β-aktin. Untuk memastikan bahawa ekspresi β-aktin tidak diubah dengan rawatan, ujian terpisah dijalankan untuk membandingkan β-aktin dengan kawalan dalaman kedua, GAPDH. Tidak terdapat perbezaan yang signifikan dalam ekspresi β-aktin (nilai ΔCT, β-aktin - GAPDH: chow kawalan = 2.29 ± 0.21, kawalan HF = 2.01 ± 0.04, ΔFosB chow = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
Trend untuk interaksi antara ekspresi ΔFosB dan rawatan diet diperhatikan untuk ekspresi tirosin hidroksilase (F = 3.6, P <0.06; Gambar 5A). Pendedahan enam minggu terhadap HF nampaknya menurunkan ekspresi tirosin hidroksilase pada tikus kawalan dan peningkatan ekspresi pada tikus ΔFosB. Interaksi yang signifikan antara ekspresi ΔFosB dan pendedahan diet diperhatikan untuk ekspresi pengangkut dopamin (F = 6.7, P <0.03; Rajah 5B). Mirip dengan tirosin hidroksilase, pendedahan kepada HF mengurangkan ekspresi transporter dopamin pada tikus kawalan dan peningkatan ekspresi pada tikus ΔFosB (P <0.05). Perbezaan asas dalam ekspresi pengangkut dopamin antara tikus kawalan dan ΔFosB tidak mencapai kepentingan (P = 0.16), tetapi setelah 6 minggu HF, tikus ΔFosB menyatakan peningkatan tahap pengangkut dopamin berbanding dengan kawalan (P <0.05).
Rajah 5
Pendedahan diet tinggi lemak (HF) dan ekspresi ΔFosB menyebabkan perubahan dalam ungkapan beberapa molekul utama dalam VTA
Terdapat trend yang menunjukkan kesan peningkatan ekspresi ΔFosB untuk mengurangkan tahap TrkB di VTA (F = 5.7, P <0.04; Gambar 5C). Walaupun tidak ada kesan utama pada ekspresi reseptor κ-opioid, terdapat kecenderungan penurunan ekspresi pada tikus ΔFosB (P = 0.08; Gambar 5D). Ekspresi reseptor leptin juga ditentukan dalam VTA. Terdapat kesan yang signifikan dari pendedahan diet (F = 6.1, P <0.03), dengan HF mengurangkan tahap reseptor leptin dalam VTA pada tikus ΔFosB dan tikus kawalan (Gamb. 5E). Ekspresi reseptor orexin 1 dalam VTA juga diperiksa. Terdapat kesan yang signifikan dari diet terhadap ekspresi reseptor orexin (F = 9.0, P <0.02), dengan tikus yang terdedah kepada HF menyatakan tahap yang lebih tinggi dalam VTA (Gambar 5F). Terdapat juga kecenderungan tikus ΔFosB untuk menyatakan tahap reseptor orexin 1 secara keseluruhan lebih tinggi di kawasan otak ini (P <0.05).
Ekspresi Orexin dalam hypothalamus lateral
Kami mengukur tahap orexin di hipotalamus lateral, asal usul pemeliharaan orexinergik VTA, dengan hibridisasi in situ (Gambar 6A). Terdapat interaksi yang signifikan antara ekspresi ΔFosB dan pendedahan diet pada ekspresi orexin (F = 9.1, P <0.01), dengan HF secara signifikan meningkatkan tahap orexin pada tikus kawalan (P <0.05) dan penurunan ekspresi pada tikus ΔFosB (Gbr. 6B). Walaupun tidak terdapat perbezaan yang signifikan dalam ekspresi orexin dalam keadaan basal, setelah 6 minggu HF, tikus ΔFosB menyatakan penurunan kadar orexin dengan ketara berbanding dengan kawalan (P <0.05).
Rajah 6
Diet tinggi lemak (HF) mempunyai kesan pembezaan pada ekspresi orexin dalam kawalan (Ctrl) dan ΔFosB overexpressing tikus
BeAnalisis havioral
Untuk menilai perubahan dalam gairah dan emosi disebabkan perubahan diet, tikus terdedah kepada ujian terbuka 24 jam setelah penarikan diet HF (10). Jumlah garis silang, yang dinilai sebagai ukuran rangsangan, dipengaruhi secara signifikan oleh ekspresi ΔFosB (F = 6.6, P <0.05) dan diet (F = 4.6, P <0.05; Gambar 7A). Tikus ΔFosB lebih aktif dalam persekitaran novel daripada kontil, dan ujian post hoc menunjukkan bahawa tikus yang ditarik dari HF secara signifikan lebih aktif daripada yang terdedah kepada chow (P <0.05). Boli tinja dikira sebagai ukuran tingkah laku seperti kegelisahan (10). Terdapat kesan utama ungkapan ΔFosB (F = 10.2, P <0.01), dengan tikus yang terlalu banyak mengekspresikan ΔFosB menghasilkan lebih banyak kotoran tinja di lingkungan novel, terutama pada kelompok penarikan rumah dan penarikan HF (Gbr. 7B). Tikus ΔFosB yang dikekalkan pada diet HF menghasilkan boli tinja lebih sedikit daripada yang dikekalkan pada chow dan yang ditarik 24 jam sebelum ujian. Tikus kawalan nampaknya tidak dipengaruhi oleh diet. Tidak ada kesan yang signifikan sama ada ekspresi ΔFosB atau diet pada masa yang dihabiskan di tengah lapangan terbuka (chow kawalan = 14.5 ± 3.1 saat, kawalan HF = 18.0 ± 3.2 saat, kawalan W / D = 15.4 ± 1.9 saat, ΔFosB chow = 16.9 ± 2.4 saat, ΔFosB HF = 13.1 ± 3.9 saat, ΔFosB W / D = 19.8 ± 2.6 saat).
Rajah 7
Mice over-expressing ΔFosB lebih sensitif terhadap kesan pengeluaran diet tinggi lemak (HF)
Perbincangan
Dalam rawatan obesiti, terdapat keperluan kritikal untuk mengenal pasti faktor-faktor yang mempengaruhi kerentanan untuk makan berlebihan dan peningkatan berat badan. Laluan ganjaran otak memainkan peranan penting dalam motivasi dan tindak balas terhadap makanan dan perubahan diet yang lazat (6, 10, 15, 16). Oleh sebab isyarat orexigenic dan anorexigenik secara langsung dapat mempengaruhi isyarat penghargaan melalui litar hypothalamus-VTA-NAc, penjelasan gen yang responsif kepada diet enak yang kaya tenaga dalam pusat ganjaran boleh memberikan sasaran terapeutik baru dalam rawatan obesiti (17, 18). Oleh itu, kita meneliti penanda biokimia dan molekul imbangan dan keseimbangan tenaga di sepanjang litar hypothalamus-VTA-NAc sebagai tindak balas kepada diet HF dalam tikus ΔFosB-overexpressing sebagai model kepekaan yang lebih baik kepada perubahan ganjaran (13, 19, 20) , dan kepekaan tingkah laku selepas pengeluaran diet. Kami mengandaikan bahawa penyisihan basal isyarat dopamin dalam tikus ΔFosB akan dinormalisasi oleh kesan ganjaran dari diet HF, merangkumi persilangan isyarat imbangan tenaga dan sistem dopamine.
Untuk memeriksa penanda yang menunjukkan pengasingan dalam isyarat dopamin dalam NAc, kami memeriksa tahap reseptor D1 dan effectors hilir. Walaupun tidak terdapat perbezaan yang ketara dalam reseptor D1 yang mengikat, terdapat trend pendedahan HF untuk meningkatkan kawasan mengikat dalam tikus ΔFosB. Ini adalah menarik kerana induksi ΔFosB oleh dadah dan ganjaran semulajadi nampaknya mendominasi dalam subtipe positif dynorphin-neuron berkilat sederhana yang terutamanya menyatakan reseptor D1 (9, 21). Tahap pCREB sasaran isyarat dopamin turun dengan ketara dalam tikus ΔFosB, menyokong pengaktifan reseptor D1 yang berkurang di rantau otak (22, 23). Menariknya, kami juga mengesan penurunan jumlah keseluruhan CREB dalam tikus ΔFosB, yang mencadangkan kapasiti pengurangan dopamine transduksi yang mungkin menengah kepada maklum balas berikutan pengurangan berpanjangan dalam pCREB (24). Ekspresi BDNF dikawal oleh pCREB, dinaikkan dengan pengaktifan D1, dan merupakan pengantara penting dalam neuroplasticitas berkaitan ganjaran dalam NAc (25, 26). Oleh itu, kami mengesan penurunan ketara dalam protein BDNF dalam tikus tikus ΔFosB.
Semua neuron berduri sederhana di NAc menyatakan DARPP-32 (27). Pelbagai effector hiliran menjadikannya pemain penting dalam jalur ganjaran (28), dan ia telah terlibat dalam ketagihan dadah dan gangguan lain yang melibatkan sistem dopamin termasuk gangguan afektif dan skizofrenia (27, 29). Kami mengesan pengurangan basal yang mendalam dalam ungkapan DARPP-32 dalam tikus tikus ΔFosB. Ekspresi DARPP-32 dikawal oleh BDNF, dan oleh itu ungkapan yang dikurangkan mungkin berkaitan langsung dengan pengurangan tahap BDNF yang dikesan dalam tikus ΔFosB (27, 29, 30). Walaupun perubahan sederhana dalam keadaan fosforilasi DARPP-32 boleh menyebabkan perubahan yang besar dalam isyarat intraselular dalam NAc (27). Kajian terdahulu telah melaporkan tiada perubahan dalam protein DARPP-32 dalam tikus ΔFosB berikutan penghapusan 12-wk dari doxycycline apabila penilaian striatal yang lebih luas dijalankan (31), mencadangkan bahawa kesan ΔFosB pada DARPP-32 mungkin masa dan rantau tertentu.
Kami menghipnotiskan bahawa pengurangan dramatik dalam indeks dopamin memberi isyarat dalam NAc tikus ΔFosB mungkin melibatkan perubahan dalam neuron unjuran dopamine VTA, walaupun ΔFosB tidak terlalu tertekan dalam neuron ini. Oleh itu, kita mengkaji ekspresi gen yang berkaitan dengan dopamine dalam VTA, termasuk tyrosine hydroxylase dan pengangkut dopamin. Tahap pengangkut tirosin hidroksilase dan pengangkut dopamin berkorelasi positif dengan pengeluaran dopamin. Terdapat trend untuk tikus ΔFosB untuk memperlihatkan pengurangan tyrosine hydroxylase dan pengurangan ketara pengangkut dopamin, selaras dengan disregulasi isyarat dopamine di NAc. Oleh kerana pengurangan asas dalam gen berkaitan dengan dopamine dalam VTA daripada tikus ΔFosB mungkin mencerminkan maklum balas yang diubah dari NAc semasa jangka panjang ΔFosB overexpression, kami memeriksa ekspresi reseptor BDNF, TrkB, sebagai mekanisme maklum balas NAc kepada VTA (32). Sama seperti tyrosine hydroxylase dan pengangkut dopamin, ekspresi TrkB juga menunjukkan trend yang pada dasarnya dapat dikurangkan dalam tikus ΔFosB yang tidak mencapai nilai apabila dibetulkan untuk beberapa perbandingan. Kompleks BDNF-TrkB boleh diangkut secara retrogred dan bertindak dalam VTA untuk mempengaruhi ekspresi gen tempatan dan mempromosikan pertumbuhan dan penyelenggaraan sel (33). Lebih-lebih lagi, pengaktifan BDNF TrkB presynaptic dalam NAc secara langsung boleh merangsang neurotransmitter dopamin (32), yang menyokong pengurangan dopamin pada isyarat ini.
Pengaktifan Dynorphin daripada reseptor κ-opioid mengawal isyarat dopamine dan merupakan satu lagi mekanisme di mana NAc memberi maklum balas kepada VTA (34). Kami mendapati bahawa ekspresi reseptor κ-opioid dalam VTA menunjukkan trend untuk dikurangkan dalam tikus ΔFosB. Oleh kerana overexpression ΔFosB telah ditunjukkan untuk mengurangkan ungkapan dynorphin dalam NAc (20), tikus ΔFosB mungkin mempunyai pengurangan mendalam dalam bersih VTA κ-pengaktifan opioid. Walaupun isyarat dynorphin biasanya memberikan kesan penghambatan pada neuron dopamin (35), tikus yang menunjukkan peningkatan penggunaan ubat penyalahgunaan menunjukkan penurunan tahap dinorphin dalam NAc, menunjukkan peranan untuk menurunkan isyarat dynorphin dalam meningkatkan kepekaan ganjaran (36) , 37). Disregulasi dinorphin - sistem κ-opioid telah dikaitkan dengan pemerolehan dan kegigihan penyalahgunaan dadah, menyokong keseimbangan kritikal isyarat opioid dalam normalisasi laluan dopamin (38).
Berdasarkan keupayaan ganjaran makanan diet HF yang padat, kami mengandaikan bahawa disyskulasi dalam dopamin dan ganjaran opioid dalam tikus ΔFosB akan menentu tikus ini untuk meningkatkan respon ganjaran kepada diet sedemikian, dengan itu menormalkan sistem ganjaran melalui pengaktifan hipotalamus -VTA-NAc litar. Semasa pendedahan diet enam minggu, tiada perbezaan dalam pengambilan makanan antara ΔFosB dan tikus kawalan yang diperhatikan, menunjukkan bahawa perubahan yang terdapat dalam tikus biokimia dan molekul isyarat ganjaran dalam tikus ΔFosB bukan disebabkan oleh perbezaan kalori yang digunakan. Seperti yang dijangkakan, perbezaan basal yang dikesan dalam pCREB, jumlah tahap reseptor CREB, BDNF, DARPP-32, dan κ-opioid antara ΔFosB dan tikus kawalan telah dilemahkan, mungkin disebabkan peningkatan pengeluaran dopamin dalam tikus ΔFosB pada HF (29, 39-41) .
Peperiksaan kedua-dua tyrosine hydroxylase dan pengangkut dopamin dalam VTA mendedahkan tindak balas yang menentang ΔFosB dan tikus kawalan berikut HF. Kawalan tikus menunjukkan penurunan ekspresi transporter tyrosine hydroxylase dan dopamin, manakala tikus ΔFosB memperlihatkan peningkatan ekspresi kedua-duanya yang berkaitan dengan dopamin. Menariknya, ekspresi tyrosine hydroxylase diubah dalam VTA oleh kokain kronik atau pentadbiran methamphetamine (42-44), menunjukkan bahawa tikus ΔFosB mungkin mencari ganjaran semula jadi HF yang lebih penting daripada tikus kawalan.
Untuk melihat bagaimana potensi input hipotalamik kepada VTA mungkin menyampaikan isyarat yang mencerminkan keseimbangan tenaga, ungkapan reseptor leptin dan reseptor orexin-1 juga diperiksa. Tahap leptin yang beredar meningkat oleh HF, dan leptin boleh bertindak pada VTA untuk mengubah isyarat dopamin (18, 45). Ekspresi reseptor leptin VTA juga dikurangkan oleh HF pada kedua-dua ΔFosB dan tikus kawalan, selaras dengan berat badan yang sama dan pengambilan makanan semasa pada HF. Tinggi lemak juga meningkatkan ekspresi reseptor orexin-1 dalam VTA daripada kedua-dua ΔFosB dan tikus kawalan. Orexin mengaktifkan neuron dopamine dalam VTA, mempromosikan keplastikan VTA, dan meningkatkan tahap dopamin dalam NAc (46-48). Diet tinggi lemak telah ditunjukkan untuk meningkatkan ekspresi orexin pada tikus, mengikut pemerhatian kami (49, 50). Oleh itu, peningkatan ekspresi reseptor orexin serta perubahan dalam isyarat leptin dalam VTA boleh menggalakkan ganjaran diet di kedua-dua fosb dan kawalan tikus, menyokong pemisahan antara laluan yang menyampaikan isyarat imbangan tenaga dan yang terikat secara langsung kepada ganjaran.
Untuk memeriksa kesan-kesan tekanan yang menarik dari pengeluaran ganjaran, tikus diperiksa dalam ujian lapangan terbuka 24 pada jam selepas penyingkiran HF. Tikus Ø FosB lebih sensitif terhadap kesan akut penarikan diet pilihan, menunjukkan peningkatan aktiviti rangsangan dan pengeluaran biri-biri tumbuhan di arena terbuka baru berbanding dengan semua kumpulan kawalan dan diet lain. Tikus ΔFosB juga menunjukkan corak tingkah laku yang menarik dalam ujian ini menunjukkan ganjaran dan kepekaan stres, dengan diet HF pada mulanya mengurangkan pengeluaran biri-biri fekal berbanding dengan chow, dan penarikan semula lagi meningkatkan tindak balas yang berkaitan dengan kebimbangan ini. Peningkatan ini dalam aktiviti terbuka tidak dikaitkan dengan perubahan dalam ekspresi orexin, yang menunjukkan hubungan dengan arousal yang disebabkan oleh tekanan yang bukan sekadar kesan perubahan dalam isyarat-isyarat mediasi orexin. Secara keseluruhan, data ini menyokong hipotesis kami bahawa tikus ΔFosB akan lebih sensitif terhadap kesan akut penarikan diet pilihan kerana kepekaan ganjaran yang tinggi.
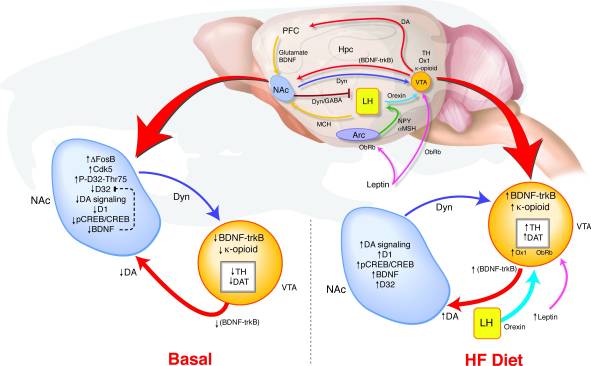
Bagaimana overexpression jangka panjang ΔFosB dalam petunjuk NAc terhadap perubahan dalam tingkah laku dan memberi isyarat balasan? Kami telah mencadangkan satu model pengesanan bertentangan VTA di mana maklum balas yang diubah dari NAc dan hypothalamus relays isyarat mengenai status ganjaran untuk menentukan peraturan sistem dopamine yang dapat menyokong hubungan antara disistulasi jalur ganjaran dan kecenderungan obesiti (Rajah 8). Semasa pendedahan HF, pelbagai input mencerminkan kedua-dua keseimbangan tenaga dan keadaan ganjaran berkumpul di VTA. Meningkatkan isyarat leptin dan orexin serta maklum balas yang diubah dari NAc kepada hipothalamus lateral boleh menjejaskan bagaimana isyarat-isyarat orenigenik ini bertindak balas terhadap HF dalam tikus ΔFosB (17, 18, 45, 47, 51-53). Ketinggian yang disebabkan oleh diet yang tinggi lemak di BDNF boleh memberi maklum balas kepada VTA, seterusnya mempromosikan perubahan dalam ekspresi gen yang berkaitan dengan dopamine.
Rajah 8
Diet tinggi lemak (HF) menormalkan isyarat imbasan yang tidak disokong dalam tikus ΔFosB
Hasil ini menggambarkan penanda molekul kepekaan ganjaran dan menunjukkan bahawa disortulasi jangka panjang sistem dopamine boleh menyebabkan seseorang menjadi kecanduan dan obesiti. Selanjutnya, data ini memberikan satu langkah penting ke arah menentukan sasaran terapi baru yang berpotensi dalam rawatan dan pencegahan obesiti dan gangguan lain yang mungkin berpusat pada sistem ganjaran. Pada masa akan datang, adalah penting untuk menyiasat bagaimana sistem ini bertindak balas terhadap penghapusan diet HF, serta menyiasat sebarang perbezaan seks dalam kepekaan terhadap ganjaran dan pendedahan makanan yang tinggi lemak.
Bahan Tambahan
Supp. Kaedah
Klik di sini untuk melihat. (61K, doc)
Penghargaan
Penulis ingin mengucapkan terima kasih kepada Cathy Steffen untuk bantuan dengan pembiakan dan pemindahan haiwan. Kerja ini disokong oleh geran dari Pusat Diabetes Universiti Pennsylvania (DK019525) dan oleh geran Institut Kesihatan Mental Negara (R01 MH51399 dan P50 MH66172) dan Institut Kebangsaan Penyalahgunaan Dadah (R01 DA07359).
Nota kaki
Pendedahan Kewangan: Semua pengarang mengisytiharkan bahawa mereka tidak mempunyai kepentingan kewangan biomedikal atau potensi konflik kepentingan.
Rujukan
1. Wadden TA, Berkowitz RI, Womble LG, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Kaplan R, Stunkard AJ. Percubaan rawak pengubahsuaian gaya hidup dan farmakoterapi untuk obesiti. N Engl J Med. 2005; 353 (20): 2111-20. [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Mengurangkan ganjaran nikotin dalam obesiti: perbandingan silang manusia dan tetikus. Psychopharmacology (Berl) 2005
3. Franken IH, Muris P. Perbezaan individu dalam sensitiviti ganjaran berkaitan dengan keinginan makanan dan berat badan relatif pada wanita yang sihat. Selera makan. 2005; 45 (2): 198-201. [PubMed]
4. Kelley AE, Berridge KC. Neuroscience ganjaran semula jadi: kaitan dengan ubat ketagihan. J Neurosci. 2002; 22 (9): 3306-11. [PubMed]
5. Cagniard B, Balsam PD, Brunner D, Zhuang X. Tikus dengan pameran dopamine meningkat secara kronik meningkatkan motivasi, tetapi tidak belajar, untuk ganjaran makanan. Neuropsychopharmacology. 2006; 31 (7): 1362-70. [PubMed]
6. Liang NC, Hajnal A, Norgren R. Sham memberi makan minyak jagung meningkatkan kena dopamin dalam tikus. Am J Physiol Regul Integr Comp Physiol. 2006; 291 (5): R1236-9. [PubMed]
7. Mendoza J, Angeles-Castellanos M, Escobar C. Entrainment oleh makanan yang enak membuat aktiviti makanan-antisipatory dan ekspresi c-Fos dalam bidang yang berkaitan dengan otak. Neurosains. 2005; 133 (1): 293-303. [PubMed]
8. Schroeder BE, Binzak JM, Kelley AE. Profil umum pengaktifan kortikal prefrontal berikutan pendedahan kepada isyarat kontekstual nikotin atau coklat. Neurosains. 2001; 105 (3): 535-45. [PubMed]
9. Nestler EJ, Barrot M, Self DW. DeltaFosB: suis molekul yang berterusan untuk ketagihan. Proc Natl Acad Sci US A. 2001; 98 (20): 11042-6. [Artikel percuma PMC] [PubMed]
10. Teegarden SL, Bale TL. Mengurangkan keutamaan diet menghasilkan peningkatan emosional dan risiko untuk kambuh makanan. Biol Psikiatri. 2007; 61 (9): 1021-9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR. dFosB di Nucleus Accumbens Mengatur Kelakuan dan Motivasi Instrumental Bertenaga Makanan. Jurnal Neuroscience. 2006; 26 (36): 9196-9204. [PubMed]
12. Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, PE Shockett, Picciotto MR, Duman RS, Nestler EJ. Haiwan transgenik dengan ekspresi gen yang disasarkan dan dijangka di dalam otak. Mol Pharmacol. 1998; 54 (3): 495-503. [PubMed]
13. Kelz MB, Chen J, Carlezon WA, Jr, Whisler K, Gilden L, Beckmann AM, Steffen C, Zhang YJ, Marotti L, Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Nestler EJ. Ekspresi faktor transkripsi deltaFosB di otak mengawal kepekaan terhadap kokain. Alam. 1999; 401 (6750): 272-6. [PubMed]
14. Bale TL, Dorsa DM. Perbezaan seks dan kesan estrogen pada ekspresi asid ribonukleik utusan reseptor oxytocin dalam hipotalamus ventrenedial. Endokrinologi. 1995; 136 (1): 27-32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. Tikus yang bergantung kepada gula memperlihatkan tindak balas yang lebih baik untuk gula selepas pantang: bukti kesan penurunan gula. Physiol Behav. 2005; 84 (3): 359-62. [PubMed]
16. Akan MJ, Franzblau EB, Kelley AE. Nukleus accumbens mu-opioids mengawal pengambilan diet tinggi lemak melalui pengaktifan rangkaian otak yang diedarkan. J Neurosci. 2003; 23 (7): 2882-8. [PubMed]
17. Zheng H, Patterson LM, Berthoud HR. Isyarat Orexin di kawasan tegegalal ventral diperlukan untuk selera makan tinggi yang disebabkan oleh rangsangan opioid nukleus accumbens. J Neurosci. 2007; 27 (41): 11075-82. [PubMed]
18. Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, Gao XB, Thurmon JJ, Marinelli M, DiLeone RJ. Isyarat reseptor leptin dalam neuron dopamine tengah mengawal makan. Neuron. 2006; 51 (6): 801-10. [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. Overprestasi jenis spesifik sel Striatal terhadap DeltaFosB meningkatkan insentif untuk kokain. J Neurosci. 2003; 23 (6): 2488-93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. Peranan penting untuk DeltaFosB dalam nukleus akrab dalam tindakan morfin. Nat neurosci. 2006; 9 (2): 205-11. [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, pembentukan tulang belakang dendritik yang diakibatkan oleh Cocaine dalam D1 dan D2 dopamin yang mengandung neuron berkilat sederhana dalam nukleus accumbens. Proc Natl Acad Sci US A. 2006; 103 (9): 3399-404. [Artikel percuma PMC] [PubMed]
22. Blendy JA, Maldonado R. Analisis genetik terhadap ketagihan dadah: peranan unsur tindak balas CAMP mengikat protein. J Mol Med. 1998; 76 (2): 104-10. [PubMed]
23. Nestler EJ. Mekanisme molekul ketagihan dadah. Neuropharmacology. 2004; 47 1: 24-32. [PubMed]
24. Tanis KQ, Duman RS, Newton SS. CREB Binding dan Aktiviti Otak: Spesifik dan Induksi Kawasan oleh Penyitaan Electroconvulsive. Biol Psikiatri. 2007
25. Kumar A, Choi KH, Renthal W, Tsankova NM, Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, DW Self, Nestler EJ. Pengubahsuaian Chromatin adalah mekanisme utama yang mendasari kepekaan cocaine-induced in striatum. Neuron. 2005; 48 (2): 303-14. [PubMed]
26. Graham DL, Edwards S, Bachtell RK, Dileone RJ, Rios M, Self DW. Aktiviti BDNF dinamik dalam nukleus yang berkait dengan penggunaan kokain meningkatkan pentadbiran diri dan kambuh semula. Nat neurosci. 2007; 10 (8): 1029-37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 mengendalikan tindakan pelbagai dadah penyalahgunaan. Aaps J. 2005; 7 (2): E353-60. [Artikel percuma PMC] [PubMed]
28. Palmer AA, Verbitsky M, Suresh R, Kamens HM, Reed CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, Phillips TJ. Perbezaan ekspresi gen pada tikus divergently dipilih untuk kepekaan methamphetamine. Mamm Genome. 2005; 16 (5): 291-305. [PubMed]
29. Bogush A, Pedrini S, Pelta-Heller J, Chan T, Yang Q, Mao Z, Sluzas E, Gieringer T, Ehrlich ME. AKT dan CDK5 / p35 memeterai induksi faktor neurotropik yang berasal dari otak DARPP-32 dalam neuron berkilat bersaiz sederhana secara in vitro. J Biol Chem. 2007; 282 (10): 7352-9. [PubMed]
30. Benavides DR, Bibb JA. Peranan Cdk5 dalam penyalahgunaan dadah dan kepekaan. Ann NY Acad Sci. 2004; 1025: 335-44. [PubMed]
31. Bibb JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Kesan pendedahan kronik terhadap kokain dikawal oleh protein neuron Cdk5. Alam. 2001; 410 (6826): 376-80. [PubMed]
32. Blochl A, Sirrenberg C. Neurotrophins merangsang pembebasan dopamin daripada neuron tikus mesencephal melalui Trk dan reseptor p75Lntr. J Biol Chem. 1996; 271 (35): 21100-7. [PubMed]
33. Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, DW Self, Nestler EJ. Peranan penting BDNF dalam laluan dopamine mesolimbi dalam tekanan kekalahan sosial. Sains. 2006; 311 (5762): 864-8. [PubMed]
34. Nestler EJ, Carlezon WA., Jr Litar ganjaran dopamine mesolimbi dalam kemurungan. Biol Psikiatri. 2006; 59 (12): 1151-9. [PubMed]
35. Ford CP, Beckstead MJ, Williams JT. Perencatan kappa opioid dopamine somatodendritic menghalang arus postsynaptic. J Neurophysiol. 2007; 97 (1): 883-91. [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Dynorphin otak dan sistem enkephalin dalam tikus Fischer dan Lewis: kesan toleransi dan pengeluaran morfin. Brain Res. 1995; 683 (1): 25-35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Perbezaan antara tikus-alkohol (AA) dan alkohol yang menghindari (ANA) tikus dalam sistem prodynorphin dan proenkephalin. Klinik Alkohol Exp Res. 1994; 18 (5): 1272-9. [PubMed]
38. Kreek MJ. Cocaine, dopamin dan sistem opioid endogen. J Addict Dis. 1996; 15 (4): 73-96. [PubMed]
39. Carlezon WA, Jr, Duman RS, Nestler EJ. Banyak wajah CREB. Trend Neurosci. 2005; 28 (8): 436-45. [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Taher M, Barczak A, Kameyama K, Huganir R, Konradi C. Dopamine D1 reseptor mengetuk fosforilasi CREB melalui fosforilasi reseptor NMDA di Ser897-NR1. J Neurochem. 2003; 87 (4): 922-34. [PubMed]
41. DW sendiri. Peraturan pengambilan dadah dan tingkah laku yang dilakukan oleh neuroadaptations dalam sistem dopamine mesolimbi. Neuropharmacology. 2004; 47 1: 242-55. [PubMed]
42. Beitner-Johnson D, Nestler EJ. Morfin dan kokain menghasilkan tindakan kronik yang lazim pada tyrosine hydroxylase di kawasan ganjaran otak dopaminergik. J Neurochem. 1991; 57 (1): 344-7. [PubMed]
43. Lu L, Grimm JW, Shaham Y, Hope BT. Neuroadaptations molekular di kawasan accumbens dan tegar ventral semasa hari 90 pertama memaksa pantang daripada pengambilan diri kokain dalam tikus. J Neurochem. 2003; 85 (6): 1604-13. [PubMed]
44. Shepard JD, Chuang DT, Shaham Y, Morales M. Kesan pentadbiran methamphetamine pada tyrosine hydroxylase dan tahap pengangkut dopamine dalam jalur dopamin mesolimbic dan nigrostriat tikus. Psychopharmacology (Berl) 2006; 185 (4): 505-13. [PubMed]
45. Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, Pothos EN, Maratos-Flier E, Flier JS. Peraturan leptin bagi laluan dopamin mesoaccumbens. Neuron. 2006; 51 (6): 811-22. [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Implikasi protein kinase C dalam ketinggian dopamin ekstraselular yang disebabkan oleh orexin dan kesannya yang memberi ganjaran. Eur J Neurosci. 2007; 25 (5): 1537-45. [PubMed]
47. Narita M, Nagumo Y, Hashimoto S, Khotib J, Miyatake M, Sakurai T, Yanagisawa M, Nakamachi T, Shioda S, Suzuki T. Penglibatan langsung sistem orexinergik dalam pengaktifan jalur dopamin mesolimbi dan tingkah laku yang berkaitan dengan morfin. J Neurosci. 2006; 26 (2): 398-405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, Bidang HL, Bonci A. Orexin A dalam VTA adalah penting untuk induksi kepekaan sinaptik dan pemekaan tingkah laku terhadap kokain. Neuron. 2006; 49 (4): 589-601. [PubMed]
49. Park ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS. Perubahan dalam ekspresi orexin-A dan neuropeptide Y dalam hipotalamus diet tikus dan pemakanan lemak tinggi. J Vet Sci. 2004; 5 (4): 295-302. [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF. Peptida yang mengawal pengambilan makanan: ekspresi gen orexin meningkat semasa keadaan hypertriglyceridemia. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1454-65. [PubMed]
51. Zheng H, Corkern M, Stoyanova I, Patterson LM, Tian R, Berthoud HR. Peptida yang mengawal selia pengambilan makanan: manipulasi akusatif yang mengundang selera mengaktifkan neuron hipotalamik orexin dan menghalang neuron POMC. Am J Physiol Regul Integr Comp Physiol. 2003; 284 (6): R1436-44. [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, Daniel RA, Landry CF, Kelley AE. Pengaktifan subpopulasi neuron-neuron yang mengandungi orexin / hipokretin yang mengandungi penghalang-penghalang reseptor yang dikuasai reseptor GABAA, tetapi tidak dengan pendedahan kepada persekitaran yang baru. Eur J Neurosci. 2004; 19 (2): 376-86. [PubMed]
53. Harris GC, Wimmer M, Aston-Jones G. Peranan untuk neuron orexin hipothalamic lateral dalam mencari ganjaran. Alam. 2005; 437 (7058): 556-9. [PubMed]