Biol Psychiatry. 2008 Dec 1; 64 (11): 941-50. Epub 2008 Jul 26.
Teegarden SL, Nestler EJ, Bale TL.
izvor
Odsjek za biologiju životinja, Univerzitet u Pensilvaniji, Filadelfija, PA 19104-6046, SAD.
sažetak
OSNOVNI PODACI:
Osetljivost na nagradu je implicirana kao predisponirajući faktor za ponašanje koje se odnosi na zloupotrebu droga, kao i prejedanje. Međutim, mehanizmi koji doprinose osetljivosti nagrađivanja nisu poznati. Pretpostavili smo da bi disregulacija u dopaminskoj signalizaciji mogla biti osnovni uzrok povećane osjetljivosti nagrađivanja, pri čemu bi stimulativni stimulansi mogli djelovati na normalizaciju sistema.
METODE:
Koristili smo genetski model miša povećane osetljivosti nagrađivanja, miša preterane ekspresije Delta FosB, da bismo ispitali promene na putu nagrađivanja kao odgovor na ukusnu ishranu sa visokim sadržajem masti. Markeri signalizacije nagrađivanja kod ovih miševa ispitivani su i bazalno i nakon 6 sedmica ukusne izloženosti ishrani. Miševi su ispitivani u testu ponašanja nakon povlačenja visoke masnoće kako bi se procijenila ranjivost ovog modela na uklanjanje nagrađivanja.
REZULTATI:
Naši rezultati pokazuju izmenjenu aktivaciju puteva nagrađivanja duž kruga jezgre akumbens-hipotalamom-ventralne tegmentalnog područja koji je rezultat prekomjerne ekspresije Delta FosB u jezgru akumbensa i striatalnih regija. Nivoi vezujućeg proteina fosforiliranog cikličkog adenozin monofosfata (cAMP) (pCREB), neurotrofični faktor iz mozga (BDNF), i dopamin i ciklični adenozin monofosfat regulirani fosfoproteini sa molekulskom masom 32 kDa (DARPP-32) u nucleus accumbens su reducirani u Delta FosB miševima, što ukazuje na smanjenu dopaminsku signalizaciju. Šest sedmica izloženosti visokim udjelom masnoća u ishrani potpuno je poboljšalo ove razlike, otkrivajući moćnu sposobnost nagrađivanja ukusne ishrane. Delta FosB miševi su takođe pokazali značajno povećanje reakcija vezanih za lokomotornu aktivnost i anksioznost 24 sati nakon povlačenja sa visokim udjelom masti.
ZAKLJUČCI:
Ovi rezultati ukazuju na osetljivost na promene u nagradama koje se odnose na disregulaciju Delta FosB i dopaminske signalizacije koje se mogu normalizovati sa ukusnim dijetama i mogu biti predisponirajući fenotip u nekim oblicima gojaznosti..
Uvod
Uprkos sve većem znanju o neuronskim sistemima koji kontrolišu apetit i sitost, stope gojaznosti nastavljaju da rastu u SAD. Trenutni tretmani lekovima imaju ograničenu efikasnost, a modifikacije ponašanja pate od minimalne dugoročne usklađenosti (1). Potrošnja kalorijski gustih, ukusnih namirnica povezana je sa promjenama u stresu i putevima nagrađivanja u mozgu, sugerirajući da nagrađivana svojstva takve hrane mogu nadjačati signale energetske bilance (2-4). Hrana bogata mastima djeluje kao prirodna nagrada, aktivirajući centre za nagrađivanje mozga na način sličan drogama zloupotrebe, i kao takvi su korišteni u paradigmama samouprave (5-8). Stoga je vjerovatno da ponašanja i motivacija za prejedanje i zloupotreba droga dijele zajedničke temeljne mehanizme, potencijalno otvarajući nove načine liječenja za oba stanja.
U proučavanju odnosa između ukusnih namirnica i puteva koji regulišu nagrađivanje i stres u mozgu, prethodno smo identificirali molekularne i biokemijske markere smanjene nagrade i povećanog stresa nakon povlačenja iz ukusne dijete s visokim udjelom masti (HF). Slično kao i droga zloupotrebe, izloženost ukusnoj ishrani u našim istraživanjima rezultirala je povećanim nivoima transkripcionog faktora ΔFosB u nucleus accumbens (NAc), centralnoj strukturi nagrađivanja mozga (9, 10).. Miševi koji induktivno prekomerno izražavaju ΔFosB pokazuju povećan instrumentalni odgovor na nagradu za hranu (11), što ih čini vrijednim alatom za ispitivanje uloge senzitivnosti nagrađivanja i dugoročne disregulacije sistema nagrađivanja u molekularnim i biokemijskim odgovorima na ukusnu ishranu.
U ovoj studiji, koristili smo ΔFosB-overeksprimirajuće miševe kako bismo ispitali dugoročne promjene u markerima nagrađivanja u neurocirkuitriji NAc-hipotalamus-ventralnog tegmentalnog područja (VTA) kao odgovor na ukusnu HF dijetu. Na osnovu prethodnih studija kod ovih miševa koji su bili osetljivi na nagrađivanje, pretpostavili smo da ΔFosB-indukovane promene u osetljivosti nagrađivanja uključuju disregulaciju u dopaminskoj signalizaciji koja je rezultat NAc povratne informacije u VTA. Nadalje, pretpostavili smo da bi izlaganje prirodnoj nagradi za energetski gustu HF dijetu normaliziralo dopaminergički sistem kod ovih miševa, što je rezultiralo pretjeranim odgovorom na stres povlačenja iz ove HF dijete. Jedinstveni aspekt upotrebe ukusne ishrane kao nagrađivane supstance omogućava nam da uključimo hipotalamičke ulaze za nagrađivanje krugova u fenotipu koji može biti prediktivni za populaciju predisponiranu gojaznosti otpornoj na tretman. Da bismo ispitali ovu hipotezu, proučavali smo markere dopaminske neurotransmisije, uključujući pCREB, BDNF i DARPP-32 u NAc i tirozin hidroksilazi i dopaminskom transporteru u VTA, nakon izlaganja HF. Takođe smo ispitali specifične markere energetske ravnoteže za koje se zna da utiču na izlaz dopamina, uključujući receptore leptina i oreksina u ekspresiji VTA i oreksina u lateralnom hipotalamusu.
Materijali i metode
Životinje
Muški bitransgenični miševi koji inducibno prekomjerno eksprimiraju ΔFosB u dynorphin-pozitivnim neuronima u NAc i dorzalnom striatumu (Kelz et al., 1999) su generisani na mješovitoj pozadini (ICR: C57Bl6 / SJL) na Univerzitetskom medicinskom centru u Texasu i održavali testiran na Univerzitetu u Pensilvaniji. Svi miševi su držani na doksiciklinu (100 μg / ml u vodi za piće) do dolaska na Univerzitet u Pensilvaniji. Da bi se indukovala prekomerna ekspresija, doksiciklin je uklonjen (n = 23) (12). Kontrolni miševi (n = 26) su nastavili da primaju lek. Miševima su dodeljene grupe za ishranu osam nedelja nakon uklanjanja doksicilina, kada je pokazano da ekspresija doseže maksimalne nivoe (13). Miševi su održavani na 12: 12 ciklusu svetlo-mrak (svetla na 0700) sa hranom i vodom dostupnom ad libitum. Sve studije su sprovedene u skladu sa eksperimentalnim protokolima odobrenim od strane Institucionalnog komiteta za negu i upotrebu životinja na Univerzitetu u Pensilvaniji, a sve procedure su sprovedene u skladu sa institucionalnim smernicama.
Izlaganje dijeti
Miševi su održavani na kućnoj hrani (n = 16) ili postavljeni na HF (n = 16-17) šest nedelja. Domaća hrana (Purina Lab Diet, St. Louis, MO) sadrži 4.00 kcal / g, koja se sastoji od 28% proteina, 12% masti i 60% ugljikohidrata. HF dijeta (Research Diets, New Brunswick, NJ) sadrži 4.73 kcal / g, koja se sastoji od 20% proteina, 45% masti i 35% ugljikohidrata.
Biohemija i ekspresija gena
Miševi su analizirani nakon šest nedelja izloženosti ishrani. Mozgovi su uklonjeni iz lobanje i zamrznuti ili na suvom ledu ili NAc disecirani (približno 0.5 - 1.75 mm od bregme, na dubini od 3.5 - 5.5 mm) i zamrznuti u tečnom azotu. Tkivo je uskladišteno na -80 ° C dok se ne analizira.
Biohemijske analize
Metode za Western blot su opisane u dopunskim materijalima. Upotrebljena antitela su: Cdk5, CREB i BDNF (1: 500, Santa Cruz biotehnologija, Santa Cruz, CA) i fosfo-CREB (pCREB) (Ser 133) (1: 500, tehnologija signalizacije ćelija, Danvers, MA).
Autoradiografija receptora
Detaljne metode autoradiografije opisane su u dopunskim materijalima. Korišćeni ligandi su bili 2 nM H3 - SCH 23390 i 5 nM H3 - spiperon (PerkinElmer, Boston, MA).
In situ hibridizacija
Obrada tkiva i hibridizacija su obavljeni kao što je prethodno opisano (14). Sondu DARPP-32 ljubazno su pružili P. Greengard (Rockefeller univerzitet) i sondu oreksina J. Elmquista (Univerzitetski medicinski centar u Teksasu). Slajdovi testirani za DARPP-32 su apsorbirani u filmu za 3 dana, a slajdovi testirani za orexin su dodani filmu za 4 dana. Kvantifikacija filmskih slika provedena je kao što je prethodno opisano (10).
QRT-PCR
RNA je izolovana iz VTA i ekspresija pojedinačnih gena procijenjena pomoću TaqMan testova ekspresije gena (Applied Biosystems, Foster City, CA). Detaljne metode i statističke analize mogu se naći u dopunskim materijalima.
Analize ponašanja
Kako bi se ispitali efekti osjetljivosti na nagradu na promjene u ponašanju uzrokovane dijetom, podskup miševa je povučen iz HF nakon četiri tjedna izlaganja i vraćen u kućnu hranu (n = 9 kontrola, n = 8 ΔFosB). Dvadeset četiri sata nakon povlačenja, miševi su bili izloženi testu otvorenog polja u skladu sa našom ranije objavljenom paradigmom povlačenja iz hrane (10). Ukratko, miš je stavljen u centar aparata otvorenog polja i praćen pet minuta. Izmjereni su ukupni linijski križevi, fekalni boli, vrijeme u centru i križevi u centar.
statistika
Svi podaci, osim Western blot-a, analizirani su dvosmjernom ANOVA-om, nakon čega slijedi Fisher-ov PLSD test s liječenjem doksiciklinom (izraz ΔFosB) i prehranom kao neovisnim varijablama. Za RT-PCR analize korištena je smanjena vrijednost P kako bi se korigiralo višestruko poređenje unutar grupa srodnih gena (vidi dopunske materijale). Western blots su analizirani pomoću studentskog t-testa sa tretmanom doksiciklinom kao nezavisnom varijablom, upoređujući optičke gustine unutar istog blota. Svi podaci su predstavljeni kao srednja vrijednost ± SEM.
Rezultati
Bazalne biokemijske razlike
Da bi se razjasnili molekularni putevi koji leže u osnovi povećane osjetljivosti nagrađivanja kod miševa s prekomjernom ekspresijom ΔFosB, ispitivani su nivoi nekoliko ključnih signalnih molekula u NAc. Zabilježen je trend povećanja nivoa Cdk5 u NAc miša ΔFosB u odnosu na kontrolne životinje u leglu koje se drže na doksiciklinu (F = 5.1, P = 0.08; slika 1A). Miševi ΔFosB su izrazili značajno smanjene nivoe pCREB (F = 7.4, P <0.05; slika 1B), kao i ukupni nivoi CREB (F = 5.4, P = 0.05; slika 1C). Značajno smanjenje BDNF uočeno je i u NAc miša ΔFosB (F = 10.6, P <0.05; slika 1D).
Slika 1
Miševi koji su prekomjerno eksprimirali ΔFosB pokazali su biokemijske markere smanjene dopaminske signalizacije u NAc
Unos hrane i tjelesna težina na dijeti s visokim udjelom masti
Zatim smo ispitali efekte HF prehrane koja prirodno nagrađuje na promjene u signalnim molekulima kod miševa s prekomjernom ekspresijom ΔFosB. Nije bilo razlike između ΔFosB miševa i kontrola u unosu hrane ni u kući ni u VF. Međutim, došlo je do ukupnog smanjenja unosa kalorija normaliziranog na tjelesnu težinu kada su bili izloženi HF-u, što je specifično za ΔFosB miševe (F = 11.2, P <0.01; slika 2A). Na kraju šest tjedana dijetalne izloženosti, miševi koji su primali HF težili su znatno više od onih na chow dijeti (F = 17.2, P <0.001), a miševi ΔFosB imali su manje od kontrolnih skupina (F = 5.6, P <0.05; slika. 2B). Ovaj je učinak bio specifičan za razlike među grupama na chow dijeti (P <0.05).
Slika 2
ΔFosB miševi sa prekomjernom ekspresijom nisu pokazali razlike u unosu hrane na hranu ili hranu s visokim udjelom masti (HF)
Biohemijske razlike u ishrani s visokim udjelom masti
Da bi se utvrdilo kako se bazalne razlike u NAc signalizaciji mogu promijeniti HF dijetom, isti signalni proteini proučavani na početku bili su ispitani na životinjama koje su primile šest tjedana HF. Nije bilo značajnih razlika u nivoima Cdk5 (Sl. 3A). Nivoi pCREB i ukupnog CREB-a više nisu bili različiti nakon šest nedelja HF-a (slika 3B, C). Nivoi BDNF su značajno povišeni kod miševa ΔFosB nakon šest nedelja izlaganja HF (F = 6.5, P = 0.05; Slika 3D).
Slika 3
Hrana sa visokim udjelom masti (HF) poboljšala je signalne razlike uočene u NAc miševa s prekomjernom ekspresijom ΔFosB
Autoradiografija dopamin receptora
Koristili smo autoradiografiju receptora da bismo procijenili jesu li promjene ΔFosB inducirane dopaminske signalizacije u NAc povezane s promjenama u ekspresiji dopaminskih receptora (slika 4A). Čini se da dijeta sa visokim udjelom masti blago povećava gustoću vezivanja za D1 dopaminski receptor (P = 0.14), a ta je razlika bila veća kod ΔFosB miševa (slika 4B). Također je zabilježen trend povećanja područja vezivanja za D1 nakon HF (P = 0.06), a post hoc testiranje pokazalo je da je to značajno kod ΔFosB miševa (P <0.05; slika 4C). Za razliku od D1 receptora, nema promjena u gustini vezivanja za D2 receptor (kontrolni chow = 97.6 ± 6.9, kontrolni HF = 101.1 ± 8.2, ΔFosB chow = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) ili područje vezivanja (kontrolni chow = 47.3 ± 3.4, kontrola HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) u NAc.
Slika 4
Hrana sa visokim sadržajem masti (HF) dovela je do promena u vezivanju D1 dopaminskih receptora i ekspresiji DARPP-32 u nucleus accumbens (NAc) ΔFosB preterano eksprimirajućih miševa
DARPP-32 izraz u NAc
Hibridizacija in situ korištena je za određivanje nivoa ekspresije DARPP-32 u NAc (slika 4D). Dijeta sa visokim udjelom masti značajno je povećala ekspresiju DARPP-32 u ovoj regiji mozga (F = 5.1, P <0.05), a došlo je i do značajne interakcije između prehrane i ekspresije ΔFosB (F = 8.9, P <0.05), pri čemu su miševi ΔFosB pokazali veću promjena izazvana prehranom (slika 4E). Bazalna razlika u ekspresiji DARPP-32 između kontrolnih i ΔFosB miševa otkrivena je post hoc testiranjem (P <0.01), kao i značajan porast ekspresije DARPP-32 kod miša ΔFosB na HF (P <0.01).
Ekspresija gena u VTA
QRT-PCR korišten je za procjenu promjena u ekspresiji gena u VTA, ciljajući nekoliko ključnih gena koji su prethodno bili uključeni u regulaciju nagrade. Svi uzorci su normalizirani na β-aktin. Da bi se osiguralo da liječenje nije promijenilo ekspresiju β-aktina, proveden je zaseban test za usporedbu β-aktina s drugom internom kontrolom, GAPDH. Nije bilo značajnih razlika u ekspresiji β-aktina (vrijednosti ΔCT, β-aktin - GAPDH: kontrola chow = 2.29 ± 0.21, kontrola HF = 2.01 ± 0.04, ΔFosB chow = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
Zabilježen je trend interakcije između ekspresije ΔFosB i dijetalnog tretmana za ekspresiju tirozin hidroksilaze (F = 3.6, P <0.06; slika 5A). Šest sedmica izlaganja HF-u smanjilo je ekspresiju tirozin hidroksilaze kod kontrolnih miševa i povećalo ekspresiju kod ΔFosB miševa. Uočena je značajna interakcija između ekspresije ΔFosB i izloženosti ishrani za ekspresiju transportera dopamina (F = 6.7, P <0.03; slika 5B). Slično tirozin hidroksilazi, izloženost HF-u smanjila je ekspresiju transportera dopamina u kontrolnih miševa i značajno povećala ekspresiju u ΔFosB miševa (P <0.05). Bazalna razlika u ekspresiji transportera dopamina između kontrolnih i ΔFosB miševa nije dostigla značaj (P = 0.16), ali nakon 6 tjedana HF, ΔFosB miševi su izrazili značajno povišene nivoe dopaminskog transportera u odnosu na kontrole (P <0.05).
Slika 5
Izloženost visokim udjelom masti (HF) i ekspresija ΔFosB doveli su do promjena u ekspresiji brojnih ključnih molekula u VTA
Zabilježen je trend koji ukazuje na učinak povećane ekspresije ΔFosB na smanjenje nivoa TrkB u VTA (F = 5.7, P <0.04; slika 5C). Iako nije bilo glavnih efekata na ekspresiju κ-opioidnih receptora, postojao je trend ka smanjenoj ekspresiji kod ΔFosB miševa (P = 0.08; slika 5D). Ekspresija receptora leptina takođe je utvrđena u VTA. Utvrđen je značajan efekat izloženosti ishrani (F = 6.1, P <0.03), pri čemu HF značajno smanjuje nivoe leptinskog receptora u VTA i kod ΔFosB i kod kontrolnih miševa (slika 5E). Takođe je ispitivana ekspresija receptora oreksina 1 u VTA. Došlo je do značajnog efekta prehrane na ekspresiju oreksinskog receptora (F = 9.0, P <0.02), kod miševa izloženih HF-u koji su izražavali viši nivo VTA (slika 5F). Takođe je postojao trend da ΔFosB miševi izražavaju ukupne više nivoe oreksinskog receptora 1 u ovoj regiji mozga (P <0.05).
Ekspresija oreksina u lateralnom hipotalamusu
Izmjerili smo nivoe oreksina u lateralnom hipotalamusu, porijeklu oreksinergičke inervacije VTA, hibridizacijom in situ (slika 6A). Došlo je do značajne interakcije između ekspresije ΔFosB i izloženosti prehrani na ekspresiji oreksina (F = 9.1, P <0.01), pri čemu je HF značajno povećao nivo oreksina u kontrolnih miševa (P <0.05) i smanjio ekspresiju kod ΔFosB miševa (slika 6B). Iako nije bilo značajnih razlika u ekspresiji oreksina u bazalnom stanju, nakon 6 tjedana HF, ΔFosB miševi izrazili su značajno smanjene razine oreksina u usporedbi s kontrolama (P <0.05).
Slika 6
Hrana sa visokim sadržajem masti (HF) imala je diferencijalne efekte na ekspresiju oreksina u kontroli (Ctrl) i ΔFosB preterano eksprimirajuće miševe
Behavioralne analize
Da bi se procijenile promjene u uzbuđenju i osjećaju zbog promjene prehrane, miševi su bili izloženi testu na otvorenom terenu 24 sata nakon povlačenja HF prehrane (10). Ukupni ukrštaji linija, koji su zabilježeni kao mjera uzbuđenja, značajno su utjecali na ekspresiju ΔFosB (F = 6.6, P <0.05) i prehranu (F = 4.6, P <0.05; slika 7A). Miševi ΔFosB bili su aktivniji u novom okruženju od kontrila, a post hoc testiranje pokazalo je da su miševi povučeni iz HF-a bili znatno aktivniji od onih izloženih chow-u (P <0.05). Fekalni boli računati su kao mjera anksioznog ponašanja (10). Došlo je do glavnog efekta ekspresije ΔFosB (F = 10.2, P <0.01), kod miševa koji su prekomjerno izražavali ΔFosB, što je stvorilo više fekalnih bolija u novom okruženju, posebno u kućnim čaušima i skupinama za povlačenje HF-a (slika 7B). Miševi ΔFosB koji su bili na VF dijeti stvorili su manje fekalnih bolija od onih koji su se održavali na chow-u i onih koji su povučeni 24 sata prije testa. Čini se da prehrana nije utjecala na kontrolne miševe. Nije bilo značajnih efekata ekspresije ΔFosB niti dijete na vrijeme provedeno u središtu otvorenog polja (kontrolni chow = 14.5 ± 3.1 sek, kontrola HF = 18.0 ± 3.2 sek, kontrola W / D = 15.4 ± 1.9 sek, ΔFosB chow = 16.9 ± 2.4 sek, ΔFosB VF = 13.1 ± 3.9 sek, ΔFosB Š / D = 19.8 ± 2.6 sek).
Slika 7
Miševi koji su prekomjerno eksprimirali ΔFosB bili su osjetljiviji na efekte povlačenja visoke masnoće (HF)
rasprava
Kod tretmana gojaznosti postoji kritična potreba za identifikacijom faktora koji utiču na podložnost prejedanju i povećanju težine. Putovi za nagrađivanje mozga igraju važnu ulogu u motivaciji i odgovoru na ukusnu hranu i prehrambene promjene (6, 10, 15, 16). Kako oreksigeni i anoreksigeni signali mogu direktno uticati na signalizaciju nagrađivanja preko hipotalamus-VTA-NAc kruga, objašnjavanje gena koji reaguju na energetski bogate dijete u centrima za nagrađivanje mogu pružiti nove terapijske ciljeve u tretmanu gojaznosti (17, 18). Stoga smo ispitali biohemijske i molekularne markere signalizacije bilansne i energetske ravnoteže duž hipotalamus-VTA-NAc kruga kao odgovor na HF dijetu u ΔFosB-preekspresionim miševima kao model povećane osjetljivosti na promjene u nagradama (13, 19, 20) i osjetljivost na ponašanje nakon povlačenja dijete. Pretpostavili smo da bi se bazalna disregulacija dopaminskog signala u ΔFosB miševima normalizirala zahvaljujući efektima HF dijete, koja bi obuhvatila sjecište signala energetske bilance i dopaminskog sistema.
Da bismo ispitali markere koji ukazuju na disregulaciju u dopaminskoj signalizaciji u NAc, ispitali smo D1 receptorske nivoe i nizvodne efektore. Iako nije bilo značajnih razlika u vezivanju D1 receptora, postojao je trend izlaganja HF povećanju površine vezivanja u miševima ΔFosB. Ovo je interesantno jer indukcija ΔFosB lijekovima i prirodnim nagradama izgleda da dominira u podtipu dinorfina pozitivnih srednjih kičmenih neurona koji prvenstveno eksprimiraju D1 receptore. (9, 21). Nivoi dolaznog signala za davanje dopamina pCREB su značajno smanjeni u ΔFosB miševima, što je podržalo smanjenu aktivaciju D1 receptora u ovom području mozga (22, 23). Zanimljivo je da smo takođe otkrili značajno smanjenje ukupnih nivoa CREB u ΔFosB miševima, što ukazuje na još manji kapacitet za transmisiju dopaminskog signala koji može biti sekundaran povratnoj sprezi koja je rezultat produženog smanjenja pCREB (24). Ekspresija BDNF-a je regulisana pCREB-om, povišena je sa aktivacijom D1-a i važan je posrednik neuroplastičnosti u NAc-u (25, 26). Shodno tome, otkrili smo značajno smanjenje BDNF proteina u NAc miševa ΔFosB.
Svi srednji kičmeni neuroni u NAc ekspresuju DARPP-32 (27). Njegovi brojni nizvodni učinci čine ga vitalnim igračem u putevima nagrađivanja (28), a impliciran je u ovisnosti o drogama i drugim poremećajima koji uključuju dopaminski sistem uključujući afektivne poremećaje i šizofreniju. (27, 29). Otkrili smo duboka bazalna smanjenja ekspresije DARPP-32 u NAc miševa ΔFosB. Ekspresija DARPP-32 je regulisana BDNF-om, i stoga smanjena ekspresija može biti direktno povezana sa redukcijama BDNF nivoa otkrivenih u ΔFosB miševima (27, 29, 30). Čak i umjerene promjene fosforilacijskog stanja DARPP-32 mogu dovesti do značajnih promjena u intracelularnoj signalizaciji unutar NAc (27). Prethodne studije su izvijestile da nema promjene DARPP-32 proteina u ΔFosB miševima nakon uklanjanja 12-wk iz doksiciklina kada je provedena šira striatalna procjena (31), sugerirajući da efekti ΔFosB na DARPP-32 mogu biti specifični za vrijeme i regiju.
Pretpostavili smo da su dramatična smanjenja indeksa dopaminskog signala u NAc miševa ΔFosB vjerovatno uključivala promjene u VTA neuronima projekcije, iako ΔFosB nije prekomjerno eksprimiran unutar ovih neurona. Stoga smo ispitali ekspresiju gena povezanih s dopaminom u VTA, uključujući tirozin hidroksilazu i dopamin transporter. Nivoi transportera tirozin hidroksilaze i dopamina su pozitivno povezani sa izlazom dopamina. Došlo je do trenda da ΔFosB miševi pokazuju smanjenu tirozinsku hidroksilazu i značajno smanjenje dopaminskog transportera, u skladu sa disregulacijom dopaminskog signaliziranja u NAc.. Budući da ova bazalna smanjenja u genima povezanih s dopaminom u VTA ΔFosB miševa vjerovatno odražavaju izmijenjene povratne informacije od NAc tokom dugotrajne prekomjerne ekspresije ΔFosB, ispitali smo ekspresiju BDNF receptora, TrkB, kao mogući mehanizam povratne informacije NAc u VTA (32). Slično kao i tirozin hidroksilaza i transporter dopamina, ekspresija TrkB je takođe pokazala trend koji se bazalno smanjuje kod ΔFosB miševa koji nisu dostigli značaj kada se koriguju za višestruka poređenja. BDNF-TrkB kompleks se može retrogradno transportirati i djelovati unutar VTA-e kako bi se utjecalo na lokalnu ekspresiju gena i promovirali rast i održavanje stanica (33). Dalje, BDNF aktivacija presinaptičkog TrkB unutar NAc može direktno stimulirati neurotransmisiju dopamina (32), podržavajući smanjenje signala dopamina u ovim miševima.
Aktivacija dinorfina κ-opioidnih receptora reguliše dopaminsku signalizaciju i drugi je mehanizam kojim NAc daje povratnu informaciju VTA (34). Otkrili smo da je ekspresija κ-opioidnih receptora u VTA pokazala trend smanjenja ΔFosB miševa. Kako je pokazano da ΔFosB prekomjerna ekspresija smanjuje ekspresiju dynorphina u NAc (20), ΔFosB miševi vjerovatno imaju duboka smanjenja neto VTA κ-aktivacija opioida. Iako dynorphin signalizacija obično vrši inhibitorni učinak na dopaminske neurone (35), pacovi koji pokazuju pojačanu samo-primjenu droga zloupotrebe pokazuju smanjene nivoe dynorphin-a u NAc, ukazujući na ulogu bazično smanjenog signal-dynorphin-a u povećanju osjetljivosti na nagrade (36 , 37). Disregulacija sistema dinorfin - κ-opioida povezana je sa stjecanjem i upornošću zlouporabe droga, podržavajući kritičnu ravnotežu signalizacije opioida u normalizaciji dopaminskih puteva (38).
Na osnovu sposobnosti nagrađivanja energetski guste HF dijete, pretpostavili smo da bi disregulacija u dopaminskoj i opioidnoj signalizaciji u ΔFosB miševima predisponirala ove miševe poboljšanim odgovorima na takvu ishranu, normalizujući sistem nagrađivanja putem aktivacije hipotalamusa. -VTA-NAc krug. Tokom izloženosti ishrani od šest nedelja, nisu uočene razlike u unosu hrane između ΔFosB i kontrolnih miševa, što sugerira da promjene pronađene u biokemijskim i molekularnim markerima signala nagrađivanja u ΔFosB miševima nisu uzrokovane razlikama u potrošenim kalorijama. Kao što se očekivalo, bazalne razlike utvrđene u pCREB, ukupnim CREB, BDNF, DARPP-32 i κ-opioidnim receptorskim nivoima između ΔFosB i kontrolnih miševa su oslabljene, vjerovatno zbog povećanog izlaza dopamina u ΔFosB miševa na HF (29, 39-41) .
Ispitivanje tirozinske hidroksilaze i dopaminskog transportera u VTA pokazalo je iznenađujuće suprotne odgovore ΔFosB i kontrolnih miševa nakon HF. Kontrolni miševi su pokazali smanjenje ekspresije tirozin hidroksilaze i transportera dopamina, dok su ΔFosB miševi pokazali povećanu ekspresiju oba ova gama povezana s dopaminom. Interesantno je da se ekspresija tirozin hidroksilaze u VTA menja hroničnom primenom kokaina ili metamfetamina (42-44), što ukazuje da bi ΔFosB miševi mogli da pronađu prirodnu nagradu HF više od kontrolnih miševa.
Da bi se ispitalo kako potencijalni ulaz hipotalamusa u VTA može biti prenos signala koji odražavaju energetsku ravnotežu, ispitivana je i ekspresija receptora za leptin i oreksin receptor-1. Nivoi leptina u cirkulaciji povećavaju se HF-om, a leptin može da deluje na VTA da bi izmenio dopaminsku signalizaciju (18, 45). Ekspresija receptora leptina VTA je na sličan način smanjena sa HF u oba ΔFosB i kontrolnim miševima, u skladu sa sličnim porastom težine i unosom dijete dok je na HF. Visoka masnoća takođe povećava ekspresiju oreksin receptora-1 u VTA i ΔFosB i kontrolnih miševa. Orexin aktivira dopamine neurone u VTA, promoviše VTA plastičnost i povećava nivo dopamina u NAc (46-48). Pokazalo se da ishrana sa visokim sadržajem masti povećava ekspresiju oreksina kod miševa, u skladu sa našim zapažanjima (49, 50). Prema tome, povećana ekspresija receptora oreksina, kao i promene u leptinskoj signalizaciji u VTA može da promoviše ishranu i kod ΔFosB i kontrolnih miševa, podržavajući disocijaciju između puteva koji prenose signale energetske ravnoteže i onih koji su direktno vezani za nagradu.
Da bi se ispitali efekti koji izazivaju nagon za povlačenjem od stresa, miševi su ispitivani na testu otvorenog polja 24 sati nakon uklanjanja HF. ΔFosB miševi su bili osjetljiviji na akutne efekte povlačenja preferirane prehrane, pokazujući povećanu aktivnost uzbuđenja i proizvodnju fekalnih bolija u novoj otvorenoj areni u usporedbi sa svim drugim kontrolnim i dijetnim grupama. ΔFosB miševi su takođe pokazali zanimljiv obrazac ponašanja u ovom testu koji ukazuje na nagrađivanje i osetljivost na stres, pri čemu je HF dijeta u početku smanjila proizvodnju fekalnih bolija u odnosu na hranu, a povlačenje je opet povećalo ovaj odgovor na anksioznost. Ovo uočeno povećanje aktivnosti otvorenog polja nije koreliralo sa promjenama u ekspresiji oreksina, što upućuje na povezanost sa uzbuđenjem izazvanim stresom, što nije samo učinak promjena u signalizaciji posredovanog oreksinom. Sve u svemu, ovi podaci potvrđuju našu hipotezu da bi ΔFosB miševi bili osjetljiviji na akutne efekte povlačenja preferirane prehrane zbog njihove pojačane osjetljivosti na nagrađivanje..
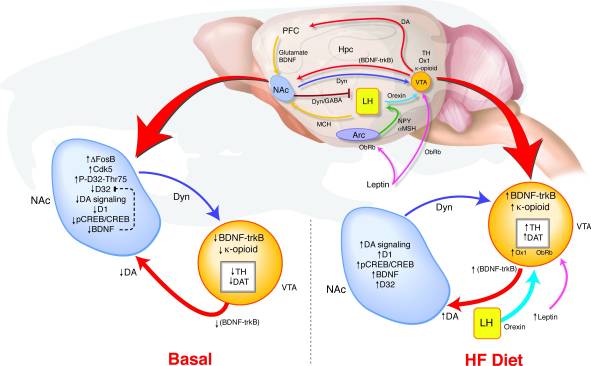
Kako dugotrajna prekomjerna ekspresija ΔFosB u NAc vodi do takvih promjena u ponašanju i nagrađivanju? Predložili smo model detektovanja slučajnih nalaza VTA u kojem izmenjena povratna sprega od NAc i releja hipotalamusa signalizira stanje nagrađivanja kako bi se odredila regulacija dopaminskog sistema koji može podržati vezu između disregulacije puta nagrađivanja i predispozicije za gojaznost (Slika 8). Tokom izlaganja visokofrekventnom zračenju, višestruki ulazi koji odražavaju i energetski bilans i stanje nagrađivanja konvergiraju se na VTA. Povećanje signala leptina i oreksina, kao i promenjena povratna sprega od NAc do lateralnog hipotalamusa može uticati na to kako ti oreksigenski signali reaguju na HF u ΔFosB miševima (17, 18, 45, 47, 51-53). Povišenja BDNF-a uzrokovana ishranom visokim udjelom masti mogu dati povratnu informaciju za VTA, dodatno promovirajući promjene u ekspresiji gena povezanih s dopaminom.
Slika 8
Hrana sa visokim udjelom masti (HF) normalizira disreguliranu nagradnu signalizaciju u ΔFosB miševima
Ovi rezultati ocrtavaju molekularne markere osjetljivosti nagrađivanja i ukazuju da dugotrajna disregulacija dopaminskog sistema može predisponirati pojedinca na ovisnost i gojaznost. Dalje, ovi podaci pružaju važan korak ka utvrđivanju potencijalnih novih terapeutskih ciljeva u tretmanu i prevenciji gojaznosti i drugih poremećaja koji mogu biti usmereni na sistem nagrađivanja. U budućnosti, bit će važno istražiti kako ovaj sistem reagira na uklanjanje HF dijete, kao i da istraži bilo kakve spolne razlike u osjetljivosti prema nagrađivanju i izloženosti ishrani s visokim udjelom masti.
Dopunski materijal
Supp. Metode
Kliknite ovdje za pregled. (61K, doc)
priznanja
Autori žele zahvaliti Cathy Steffen za pomoć u uzgoju i transferu životinja. Ovaj rad je podržan grantom Centra za dijabetes Univerziteta Pensilvanije (DK019525) i grantovima Nacionalnog instituta za mentalno zdravlje (R01 MH51399 i P50 MH66172) i Nacionalnog instituta za zloupotrebu droga (R01 DA07359).
Fusnote
Finansijska obelodanjivanja: Svi autori izjavljuju da nemaju biomedicinske finansijske interese ili potencijalne sukobe interesa.
reference
1. Wadden TA, Berkowitz RI, Womble LG, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Kaplan R, Stunkard AJ. Randomizirano ispitivanje modifikacije načina života i farmakoterapije za gojaznost. N Engl J Med. 2005; 353 (20): 2111 - 20. [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Smanjena nikotinska nagrada u gojaznosti: unakrsna usporedba kod ljudi i miša. Psihofarmakologija (Berl) 2005
3. Franken IH, Muris P. Individualne razlike u osjetljivosti nagrađivanja povezane su sa žudnjom za hranom i relativnom tjelesnom težinom zdravih žena. Apetit. 2005; 45 (2): 198 - 201. [PubMed]
4. Kelley AE, Berridge KC. Neuroznanost prirodnih nagrada: relevantnost za lijekove koji izazivaju ovisnost. J Neurosci. 2002; 22 (9): 3306 - 11. [PubMed]
5. Cagniard B, Balsam PD, Brunner D, Zhuang X. Miševi sa kronično povišenim dopaminom pokazuju pojačanu motivaciju, ali ne i učenje, za nagradu za hranu. Neuropsychopharmacology. 2006; 31 (7): 1362 - 70. [PubMed]
6. Liang NC, Hajnal A, Norgren R. Šam hranjenja kukuruznim uljem povećava akumulaciju dopamina kod štakora. Am J Physiol Regul Integral Comp. 2006; 291 (5): R1236 – 9. [PubMed]
7. Mendoza J, Angeles-Castellanos M, Escobar C. Unošenje ukusnog obroka izaziva aktivnost predviđanja hrane i ekspresiju c-Fosa u područjima mozga koja se odnose na nagrađivanje. Neuroscience. 2005; 133 (1): 293 - 303. [PubMed]
8. Schroeder BE, Binzak JM, Kelley AE. Uobičajeni profil prefrontalne kortikalne aktivacije nakon izlaganja kontekstualnim znakovima vezanim za nikotin ili čokoladu. Neuroscience. 2001; 105 (3): 535 - 45. [PubMed]
9. Nestler EJ, Barrot M, Self DW. DeltaFosB: produženi molekularni prekidač za zavisnost. Proc Natl Acad Sci SAD A. 2001; 98 (20): 11042 – 6. [PMC besplatan članak] [PubMed]
10. Teegarden SL, Bale TL. Smanjenje prehrambenih preferencija dovodi do povećane emocionalnosti i rizika za povratak hrane. Biol Psychiatry. 2007; 61 (9): 1021 - 9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR. dFosB u Nucleus Accumbens regulira instrumentalno ojačano hranu i motivaciju. The Journal of Neuroscience. 2006; 26 (36): 9196 - 9204. [PubMed]
12. Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ. Transgene životinje sa inducibilnom ekspresijom gena u mozgu. Mol Pharmacol. 1998; 54 (3): 495 - 503. [PubMed]
13. Kelz MB, Chen J, Carlezon WA, Jr., Whisler K, Gilden L, Beckmann AM, Steffen C, Zhang YJ, Marotti L, Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Duman RS, Picciotto MR, Nestler EJ. Ekspresija transkripcionog faktora deltaFosB u mozgu kontrolira osjetljivost na kokain. Priroda. 1999; 401 (6750): 272 - 6. [PubMed]
14. Bale TL, Dorsa DM. Polne razlike i efekti estrogena na ekspresiju ribonukleinske kiseline receptora oksitocina u ventromedijalnom hipotalamusu. Endokrinologija. 1995; 136 (1): 27 - 32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. Štakori koji zavise od šećera pokazuju pojačano reagovanje na šećer nakon apstinencije: dokaz o efektu deprivacije šećera. Physiol Behav. 2005; 84 (3): 359 - 62. [PubMed]
16. Will MJ, Franzblau EB, Kelley AE. Nucleus accumbens mu-opioidi regulišu unos ishrane sa visokim udjelom masti aktivacijom raspodijeljene mreže mozga. J Neurosci. 2003; 23 (7): 2882 - 8. [PubMed]
17. Zheng H, Patterson LM, Berthoud HR. Signalizacija oreksina u ventralnom tegmentalnom području je neophodna za apetit sa visokim sadržajem masti izazvan stimulacijom opioida u nucleus accumbens. J Neurosci. 2007; 27 (41): 11075 - 82. [PubMed]
18. Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, Gao XB, Thurmon JJ, Marinelli M, DiLeone RJ. Signalizacija receptora za leptin u neuronima dopamina srednjeg mozga reguliše hranjenje. Neuron. 2006; 51 (6): 801 - 10. [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. Striatalna tipska prekomjerna ekspresija DeltaFosB-a povećava poticaj za kokain. J Neurosci. 2003; 23 (6): 2488 - 93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. Bitna uloga za DeltaFosB u nucleus accumbens u morfinu. Nat Neurosci. 2006; 9 (2): 205 - 11. [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. Stvaranje dendritske kičme izazvane kokainom u D1 i D2 dopaminskim receptorima koji sadrže srednje štitne neurone u nucleus accumbens. Proc Natl Acad Sci SAD A. 2006; 103 (9): 3399 – 404. [PMC besplatan članak] [PubMed]
22. Blendy JA, Maldonado R. Genetska analiza ovisnosti o drogama: uloga vezujućeg proteina cAMP odgovora. J Mol Med. 1998; 76 (2): 104 - 10. [PubMed]
23. Nestler EJ. Molekularni mehanizmi ovisnosti o drogama. Neuropharmacology. 2004; 47 1: 24 – 32.
24. Tanis KQ, Duman RS, Newton SS. Vezivanje i aktivnost CREB u mozgu: regionalna specifičnost i indukcija elektrokonvulzivnim napadom. Biol Psychiatry. 2007
25. Kumar A, Choi KH, Renthal W, Tsankova NM, Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ. Remodeliranje hromatina je ključni mehanizam koji podupire plastičnost kokaina u striatumu. Neuron. 2005; 48 (2): 303 - 14. [PubMed]
26. Graham DL, Edwards S, Bachtell RK, Dileone RJ, Rios M, Self DW. Dinamička aktivnost BDNF u nucleus accumbens sa upotrebom kokaina povećava samo-administraciju i relaps. Nat Neurosci. 2007; 10 (8): 1029 - 37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 posreduje u djelovanju višestrukih lijekova. Aaps J. 2005; 7 (2): E353 – 60. [PMC besplatan članak] [PubMed]
28. Palmer AA, Verbitsky M, Suresh R, Kamens HM, Reed CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, Phillips TJ. Razlike u ekspresiji gena kod miševa različito odabranih za osjetljivost metamfetamina. Mamm Genome. 2005; 16 (5): 291 - 305. [PubMed]
29. Bogush A, Pedrini S, Pelta-Heller J, Chan T, Yang Q, Mao Z, Sluzas E, Gieringer T, Ehrlich ME. AKT i CDK5 / p35 posreduju indukciju DARPP-32 neurotrofnog faktora iz mozga in vitro u srednjim kičmim neuronima. J Biol Chem. 2007; 282 (10): 7352 - 9. [PubMed]
30. Benavides DR, Bibb JA. Uloga Cdk5 u zloupotrebi droga i plastičnosti. Ann NY Acad Sci. 2004; 1025: 335 – 44. [PubMed]
31. Bibb JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Efekti hronične izloženosti kokainu regulirani su proteinom neurona Cdk5. Priroda. 2001; 410 (6826): 376 - 80. [PubMed]
32. Blochl A, Sirrenberg C. Neurotrofini stimulišu oslobađanje dopamina iz mesencefalnih neurona štakora preko Trk i p75Lntr receptora. J Biol Chem. 1996; 271 (35): 21100 - 7. [PubMed]
33. Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. Bitna uloga BDNF u mezolimbičkom putu dopamina u socijalnom poraznom stresu. Nauka. 2006; 311 (5762): 864 - 8. [PubMed]
34. Nestler EJ, Carlezon WA., Jr Mezolimbički krug dopaminskog nagrađivanja u depresiji. Biol Psychiatry. 2006; 59 (12): 1151 - 9. [PubMed]
35. Ford CP, Beckstead MJ, Williams JT. Kappa opioidna inhibicija postsinaptičkih struja somatodendritičnih dopaminskih inhibitora. J Neurophysiol. 2007; 97 (1): 883 - 91. [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Sistemi dynorphina i enkefalina u mozgu u Fischer i Lewis štakorima: efekti tolerancije i povlačenja morfina. Brain Res. 1995; 683 (1): 25 - 35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Razlike između alkohola (AA) i alkohola (ANA) štakora u prodynorphin i proenkephalin sustavima. Alcohol Clin Exp Res. 1994; 18 (5): 1272 - 9. [PubMed]
38. Kreek MJ. Kokain, dopamin i endogeni opioidni sistem. J Addict Dis. 1996; 15 (4): 73 - 96. [PubMed]
39. Carlezon WA, Jr., Duman RS, Nestler EJ. Mnoga lica CREB-a. Trends Neurosci. 2005; 28 (8): 436 - 45. [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Taher M, Barczak A, Kameyama K, Huganir R, Konradi C. Receptori dopamina D1 posreduju CREB fosforilaciju putem fosforilacije NMDA receptora na Ser897-NR1. J Neurochem. 2003; 87 (4): 922 - 34. [PubMed]
41. Self DW. Regulisanje ponašanja uzimanja lekova i traženja ponašanja od strane neuroadaptacija u mezolimbičkom dopaminskom sistemu. Neuropharmacology. 2004; 47 1: 242 – 55.
42. Beitner-Johnson D, Nestler EJ. Morfij i kokain ispoljavaju uobičajene hronične aktivnosti na tirozin hidroksilazi u dopaminergičkim regijama za nagrađivanje mozga. J Neurochem. 1991; 57 (1): 344 - 7. [PubMed]
43. Lu L, Grimm JW, Shaham Y, Hope BT. Molekularne neuroadaptacije u akumbensu i ventralnom tegmentalnom području tokom prvih 90 dana prisilne apstinencije od samo-davanja kokaina kod pacova. J Neurochem. 2003; 85 (6): 1604 - 13. [PubMed]
44. Shepard JD, Chuang DT, Shaham Y, Morales M. Utjecaj samoprimjene metamfetamina na razine transportera tirozin hidroksilaze i dopamina u mezolimbičkim i nigrostriatalnim dopaminskim putevima štakora. Psihofarmakologija (Berl) 2006; 185 (4): 505 – 13. [PubMed]
45. Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, Pothos EN, Maratos-Flier E, Flier JS. Leptinska regulacija puteva dopamina u mesoakumbi. Neuron. 2006; 51 (6): 811 - 22. [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Implikacija protein kinaze C u oreksin induciranom povećanju nivoa izvanstaničnog dopamina i njegovom nagrađivanju. Eur J Neurosci. 2007; 25 (5): 1537 - 45. [PubMed]
47. Narita M, Nagumo Y, Hashimoto S, Khotib J, Miyatake M, Sakurai T, Yanagisawa M, Nakamachi T, Shioda S, Suzuki T. Direktno uključivanje oreksinergičkih sistema u aktiviranje mezolimbičkog puta dopamina i srodnih ponašanja izazvanih morfinom. J Neurosci. 2006; 26 (2): 398 - 405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, Fields HL, Bonci A. Oreksin A u VTA je kritičan za indukciju sinaptičke plastičnosti i senzibilizacije ponašanja na kokain. Neuron. 2006; 49 (4): 589 - 601. [PubMed]
49. Park ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS. Promene u ekspresiji oreksina-A i neuropeptida Y u hipotalamusu gladne i visoko-masne hrane hranjene pacovima. J Vet Sci. 2004; 5 (4): 295 - 302. [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF. Peptidi koji regulišu unos hrane: ekspresija gena za oreksin se povećava tokom stanja hipertrigliceridemije. Am J Physiol Regul Integral Comp. 2003; 284 (6): R1454 – 65. [PubMed]
51. Zheng H, Corkern M, Stoyanova I, Patterson LM, Tian R, Berthoud HR. Peptidi koji regulišu unos hrane: manipulacija akumbensima izaziva apetit aktivira neurone hipotalamusa oreksina i inhibira POMC neurone. Am J Physiol Regul Integral Comp. 2003; 284 (6): R1436 – 44. [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, Daniel RA, Landry CF, Kelley AE. Aktiviranje subpopulacije hipotalamičkih neurona koji sadrže oreksin / hipokretin pomoću inhibicije GABAA receptora posredovane inhibicije ljuske nucleus accumbens, ali ne i izlaganjem novom okruženju. Eur J Neurosci. 2004; 19 (2): 376 - 86. [PubMed]
53. Harris GC, Wimmer M, Aston-Jones G. Uloga za lateralne hipotalamičke oreksinske neurone u potrazi za nagradom. Priroda. 2005; 437 (7058): 556 - 9. [PubMed]