YBOPI MÄRKUSED: Internetis sisalduva porno taga on Coolidge'i efekt. Coolidge'i efekt on nähtus, mida täheldatakse imetajaliikides, kus meestel (ja vähemal määral naistel) ilmnevad uued seksuaalsed huvid, kui neid tutvustatakse uutele vastuvõtlikele seksuaalpartneritele, isegi pärast seksi keeldumist varasematest, kuid endiselt kättesaadavatest seksuaalpartneritest. Seksuaalne uudsus alistab selle harjumise kõrgema dopamiini põhjustatud uuendatud elevusega. Pidev uudsuse vool muudab Interneti-porno nii erinevaks mineviku pornost.
Originaal artikkel koos graafikutega
- Dennis F. Fiorino,
- Ariane Coury ja
- Anthony G. Phillips
- Neuroteaduse ajakiri, 15. juuni 1997, 17 (12): 4849-4855;
Abstraktne
Coolidge'i efekt kirjeldab seksuaalse käitumise taaselustamist "seksuaalselt küllastunud" loomal vastuseks uuele vastuvõtlikule kaaslasele. Arvestades mesolimbilise dopamiini (DA) süsteemi rolli motiveeritud käitumise algatamisel ja säilitamisel, kasutati mikrodialüüsi, et jälgida tuuma accumbens (NAC) DA ülekannet kopulatsiooni, seksuaalse küllastuse ja seksuaalse käitumise taasalustamise ajal. Kokkuvõttes varasemate aruannetega seostati estrous naise ekraani taga ja kopulatsiooniga NAC DA effluxi märkimisväärset suurenemist. NAC DA kontsentratsioonide taastumine algväärtustele langes kokku seksuaalse küllastumisajaga, kuigi DA-metaboliitide, dihüdroksüfenüüläädikhappe ja homovanillhappe kontsentratsioonid jäid kõrgeks. Uue vastuvõtva naise esitlemine ekraani taga viis NAC DA väikese suurenemiseni, mida suurendati oluliselt uue naissoost uue kopulatsiooni ajal. Praegused andmed näitavad, et uue vastuvõtliku naise stimuleerivad omadused võivad suurendada NAC DA ülekannet seksuaalselt küllastunud isasrottidel ja see võib omakorda olla seotud seksuaalse käitumise taasalustamisega.
Sissejuhatus
Naise rott, kes on küllastunud küllastumisvõimelisusele, võib uuesti indutseerida, kui algne emane on asendatud uudse vastuvõtva naissoost. See on tuntud kui Coolidge'i efekt ja seda on täheldatud mitmetes imetajate liikides (Wilson jt, 1963). Üldised tegurid, nagu väsimus või motoorne depressioon, ei ole piisavad seksuaalse küllastuse ilmse seisundi selgitamiseks, sest uudse naise stiimulid võivad siiski tekitada kopulatsiooni. Seksuaalset küllastust saab farmakoloogiliselt, olulisel määral „farmakoloogiliselt muuta“, manustades erinevaid ravimeid, mis võivad mõjutada erinevaid neurotransmitterisüsteeme. Nende ravimite hulka kuuluvad yohimbine, 8-OH-DPAT (Rodriguez-Manzo ja Fernandez-Guasti, 1994, 1995), nalaxone (Pfaus ja Gorzalka, 1987; Rodriguez-Manzo ja Fernandez-Guasti, 1995a,b) ja apomorfiini (Mas et al., 1995c). Kuigi nende ravimite perifeerset toimet ei saa välistada (nt adrenergiline toime erektsiooni funktsioonile), on selektiivsete tsentraalsete noradrenergiliste kahjustuste katsete põhjal välja pakutud seksuaalse küllastuse aluseks olevad keskmehhanismid.Rodriguez-Manzo ja Fernandez-Guasti, 1995a) ja mikrodialüüsi eksperimendid, mis jälgisid dopamiinergilist metabolismi mediaalse preoptilise piirkonna \ tMas et al., 1995a,b).
Arvestades, et kesksed mehhanismid võivad vahendada Coolidge'i efektile iseloomulikku seksuaalse käitumise taastamist, on tõenäoliselt kandidaat mesolimbiline dopamiini (DA) süsteem, mis ulatub ventraalsest tegmentaalsest piirkonnast NAC-i. Mesolimbiline DA näib toimivat esmase modulaatorina komplekssetes integreerivates protsessides, mis hõlmavad keskkonnaalaste stiimulite, näiteks seksuaalselt vastuvõtliku naise märkide hindamist ja eesmärgipärase käitumise korraldamist, kaasa arvatud kopulatsioon (Fibiger ja Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Kuigi kesknärvi DA-neuronid reageerivad primaarsetele hüvedele ja tasudele, uudsetele või ettearvamatutele keskkonnaalastele stiimulitele, viitavad neuronite aktiveerimine kõige tugevamalt korduvate treeningute ajal (Fabre et al., 1983; Schultz, 1992; Mirenowicz ja Schultz, 1994). On palju tõendeid, mis toetavad mesolimbilise DA olulist hõlbustavat rolli rottide seksuaalse käitumise algatamisel ja säilitamisel (Pfaus ja Everitt, 1995) ja mitmed mikrodialüüsi uuringud näitavad, et isaste seksuaalse käitumise söögiisu ja tarbimise faaside ajal suureneb NAC DA väljavool (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Samas on suhteliselt vähe andmeid seksuaalse küllastumise ja seksuaalse käitumise taaselustamise neurokeemiliste korrelatsioonide kohta. Kohaldamine in vivo mikrodialüüsi Mesolimbic DA neurotransmissiooni jälgimiseks Coolidge'i efekti ajal annab ainulaadse võimaluse uurida NAC DA rolli kopulatsioonis, seksuaalses küllastuses ja kopulatsiooni taasalustamises.
Mikrodialüüsi katse viidi läbi, et määrata kindlaks järgmised asjaolud: (1), kas seksuaalse küllastuse algusega kaasneb rakuvälise DA kontsentratsiooni taastumine NAC-i eelkombinatsiooniväärtustele või alla, ja (2), kas kopulatsiooni käitumise taastamine a “ seksuaalselt küllastunud isastel rottidel, kellel on uudne vastuvõtlik naine, on korrelatsioon NAC DA väljavoolu suurenemisega.
MATERJALID JA MEETODID
Teemad. Isased Sprague Dawley rotid, kes olid saadud loomade hoolduskeskusest (Briti Columbia ülikoolis) ja naissoost Long-Evans rotid, mis saadi firmast Charles River Canada (St. Constant, Quebec, Kanada), paigutati traatvõrkpuuridesse (18 × 25 × 65 cm, viis puuri kohta eraldi kolooniaruumides. Kolooniaruume hoiti ∼20 ° C juures vastupidises 12 h valguse / pimedas tsüklis. Rottidel oli piiramatu juurdepääs toidule (Purina Rat Chow) ja veele.
Operatsioon ja käitumuslik testimine enne aju mikrodialüüsi.Naine rotid ovaritektomiseeriti kahepoolselt halotaani gaasianesteesia (Fluothane, Ayerst Laboratories) all vähemalt 4 nädalat enne testimist. Seksuaalset vastuvõtlikkust stimuleerivatel naistel indutseeris enne iga testis östradioolbensoaadi (10 μg) ja progesterooni (500 μg), 48 ja 4 hr subkutaanne süstimine. Isased rotid skriiniti seksuaalse käitumise suhtes kahel korral, 4 d peale, pleksiklaaside kambrites (35 × 35 × 40 cm) traatvõrgu põrandatega. Ainult isased rotid, kes saavutasid tulemuslikkuse kriteeriumi, mis sisaldasid embrüo ja ejakulatsiooni 5 min jooksul esimese intromissioni 15 min jooksul kahe implantaadi testimise ajal, implanteeriti mikrodialüüsi sondi juhtkanüülidega.
Isased rotid (n = 5) tuimastati ketamiinvesinikkloriidiga (100 mg / kg, ip) ja ksülasiiniga (10 mg / kg, ip) enne stereotaksilist operatsiooni. Mikrodialüüsi sondi juhtkanüülid (19 gabariit) implanteeriti kahepoolselt NAC-i kaudu (bregma koordinaadid: eesmine, + 1.7 mm; mediaalne, ± 1.1 mm; ventral, −1.0 mm; lame kolju) ja kinnitati kolju koos hambaravi akrüüliga ja juveliiride kruvid. Eduka mikrodialüüsi katse maksimeerimiseks kasutati kahepoolseid juhtkanüülimplantte. Õnneks oli käesolevas eksperimendis iga roti jaoks vaja ainult ühte kanüüli. Isased rotid paigutati eksperimendi järelejäänud ajaks individuaalselt suurtesse plastikpuuridesse, kus oli sarvkesta allapanu. Üks nädal pärast operatsiooni testiti rotte seksuaalse käitumise suhtes. Koolituse selle osa jooksul oli testimiskambris libisev pleksiklaasiga ekraan, mis jagas kambri suurteks ja väikesteks kambriteks. Isased rotid viidi suuresse kambrisse ja 15 min hiljem, emane asetati ekraani taga. Pärast 15 min ettevalmistavat perioodi eemaldati ekraan ja rottidel lasti kopeerida 30 min. Viidi läbi kolm treeningut, üks iga 4 d. Kõik rotid jõudsid tulemuslikkuse kriteeriumi igal istungil.
Coolidge'i efekti katse. Rotid implanteeriti ühepoolselt mikrodialüüsi sondidega 12-18 hr enne Coolidge'i efektiekspertiisi ja paigutati katsekambri suuresse ruumi, kus oli vaba juurdepääs toidule ja veele. Katse hommikul koguti iga 15 min. Mikrodialüüsi proovid. Katse koosnes järgmistest seitsmest järjestikusest faasist: (1) baasjoon (vähemalt 60 min); (2) emane 1 ekraani taga (15 min); (3) kopulatsioon naissoost 1'iga kuni 30 min perioodi möödumiseni ilma kinnituseta; (4) naissoost 1i taaskehtestamine ekraani taga (15 min); (5) ligipääs naissoost 1-le 15-minuti jooksul, kui ei olnud paigaldust (kui paigaldamine toimus, käsitleti seda faasi faasina 3); (6) naissoost 2i sisseviimine ekraani taga (15 min); 7) kopulatsioon naissoost 2 60 min.
Käitumine filmiti madala valgustusega JVC-videosüsteemi abil ja seda vaadati videomonitoril, mis asub väljaspool katseruumi. Seksuaalse käitumise standardmeetmed registreeriti arvuti ja sobiva tarkvara abil (Holmes et al., 1987).
Pärast mikrodialüüsi katse manustati loomadele kloraalhüdraadi üleannustamine ja perfuseeriti intrakardiaalselt soolalahusega ja formaliiniga (4%). Ajusid viilutati ja külmutati ning seejärel koronaalsed sektsioonid värviti kresüülvioletiga, et määrata mikrodialüüsi sondide paigutamine. Käitumise ja neurokeemiliste analüüside jaoks kasutati ainult NAC-sse sondidega paigutatud rottidele.
Mikrodialüüs ja HPLC-elektrokeemiline tuvastamine. Mikrodialüüsi sondid olid konstruktsioonilises kontsentratsioonis pooleldi läbilaskva õõneskiudmembraaniga (eksponeeritud 2 mm membraan, välimine läbimõõt 340 μm, molekulmassi 65000, Filtral 12, Hospal). Sondid perfundeeriti 1.0 μl / min juures modifitseeritud Ringeri lahusega (0.01 m naatriumfosfaatpuhver, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl), kasutades suitsu-süstalt (Hamilton, Reno, NV) ja süstalpumpa (mudel 22, Harvard Apparatus, South Natick, MA). Mikrodialüüsi sondi kinnitamiseks juhtkanüülis kasutati mikrodialüüsi sondi juhtkaelust. Proovitoru kaitsmiseks kasutati katsekambrile paigaldatud vedelat pööret (Instech 375s) külge kinnitatud terasest spiraal.Fiorino et al., 1993).
Mikrodialüsaadi analüütid, mis hõlmasid DA ja selle metaboliite dihüdroksüfenüüläädikhapet (DOPAC) ja homovanillhapet (HVA), eraldati pöördfaasi kromatograafiaga (Ultrasphere kolonn; Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, sisemine diameeter) ) kasutades 0.083m naatriumatsetaatpuhvrit, pH 3.5 (5% metanool). Analüütide kontsentratsioonid kvantifitseeriti elektrokeemilise (EC) tuvastamisega. Seade koosnes Bio-Radi (Richmond, CA) pumbast, Valco Instruments (Houston, TX) EC10W kaheasendisest pihustist, ESA (Bedford, MA) Coulochem II EC detektorist ja kahekanalilisest diagrammi salvestajast (Kipp ja Zonen, Bohemia, NY). Elektrokeemilise detektori parameetrid olid järgmised: elektrood 1, + 450 mV; elektrood 2, -300 mV; ja valvur, −450 mV. Tüüpilised sondide taastumised, läbi viidud vitro ja toatemperatuuril olid 22% DA, 18% DOPAC puhul ja 18% HVA puhul.
TULEMUSED
Käitumine
Coolidge'i efekti katse käitumuslikud meetmed on esitatud tabelis 1. Latentsid paigaldamiseks, sissetungimiseks ja ejakulatsiooniks, aga ka pärast esimest ejakulatsiooni toimunud postejaculatory intervall olid sarnased eelmise treeningu ajaga (andmeid ei ole näidatud). See viitab sellele, et mikrodialüüsi protseduur ei muutnud normaalset seksuaalset käitumist. Seksuaalse küllastuse areng, mida näitab ejakulatsioonide keskmine arv enne kriteeriumi täitmist (7.8 ± 0.5), igale ejakulatsioonile eelnenud intromioonide arvu järkjärguline vähenemine ja positsioonikulaarse intervalli järkjärguline suurenemine (andmeid ei ole näidatud) , oli sarnane varasemates uuringutes teatatud \ tRand ja Jordaania, 1956; Fowler ja Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo ja Fernandez-Guasti, 1994; Mas et al., 1995d). Individuaalset varieeruvust täheldati naissoost 1iga saavutatud ejakulatsioonide arvu, naissoost 1iga kopuleerimise aja ja naissoost 1 esituste arvu, mis on vajalik küllastuskriteeriumi saavutamiseks (tabel 1, alt). Mõned rotid vajasid naissoost 1i arvukalt taastamisi, kuni faas 5 oli lõppenud (n = 3). Naiste 1i ekraani taha asetamise ja partitsiooni eemaldamise toimingud võisid olla esmaste isuäratud märkidena, mis viisid kopulatsioonini. Samuti tuleb märkida, et küllastuskriteeriumiks 30 min ilma kinnituseta, kuigi seda kasutati varem (Beach ja Jordan, 1965; Mas et al., 1995b), on meelevaldne ega garanteeri, et rott ei oleks andnud rohkem aega. Sellegipoolest ei põhjustanud viivitused või eemaldamis- ja asendusprotseduurid usaldusväärset uuendatud kopulatsiooni naissoost 1iga (nt faasid 4 ja 5).

Käitumine Coolidge'i efekti katse ajal
Kõigil rottidel oli Coolidge'i efekt. Naistele 2 paigutamisega seotud aktiivsus ekraani taga ja eriti partitsiooni eemaldamine võib olla sellele tulemusele kaasa aidanud, kuid jällegi ei olnud need sündmused iseenesest piisavad kopulatsiooni uuendamiseks eksperimentis varem. Kasutati seksuaalse käitumise mõõtmisi naissoost 1i ja naissoost 2iga t Bonferroni korrektsiooniga testid. Ehkki 2-i vastusena naise 1-i ravile reageerimis- ja intromission-latentsus ei erinenud oluliselt 2i emaslooma esimese kopulatsiooniastmega, oli 0.6i emasloomade seksuaalkäitumine vähem tugev, nagu näitab oluliselt vähem ejakulatsioone (keskmine, 4.2 vs XNUMX; F = 49.86;p <0.01) ja vahekorrad (keskmine = 11.2 vs 37.0;F = 20.17; p <0.05) esimese tunni jooksul. Emaste 1 ja 2 esimesel tunnil kinnituste arv ei erinenud oluliselt.
On oluline märkida, et eksperimendi küllastumisosas (st naissoost 1) kasutatavatel naistel esines endiselt tugev proteptiivne (st hopping ja darting) ja vastuvõtlik (st lordoosi) käitumine kogu meeste kokkupuute kestusega. .
Neurokeemia
Esimesel kolmel algtasemel olid DA ja selle metaboliitide basaal nanomolaarsed kontsentratsioonid mikrodialüsaatides, mis olid keskmised ± SEM, järgmised: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; ja HVA, 234.2 ± 49.0 (sondide taastumise suhtes korrigeerimata;n = 5). Need väärtused esindasid 100% baasjoone skoori.
Neurokeemilisteks analüüsideks kasutati käitumismääratletud andmepunkte, mis vastavad igale katse faasile ja olid tavalised igale rottile. Need olid järgmised: (1) seitse proovi pärast naissoost 1i (2) esmakordset kasutuselevõttu koos 1-i emaste 3-i ja 2-i viie prooviga, pärast naissoost XNUMXi esitlemist. Joonis 1 illustreerib muutusi DA kontsentratsioonis (joon graafik, kesk-) ja DA-metaboliidid (joongraafik, \ t ülemine) paralleelselt kopulatsioonikäitumisega (tulpdiagramm, põhi) Coolidge'i efekti katse ajal.
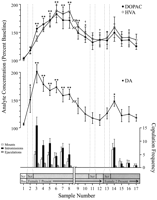
Vaata suuremat versiooni:
Joon. 1.
Nukleuse akumuleerib seksuaalse käitumise neurokeemilised korrelatsioonid Coolidge'i efekti ajal. Esimesed kaheksa proovi esindavad kronoloogiliselt pidevaid andmepunkte faasidest 1 kuni 3. Proov 1 on neljas ja viimane eelvereproovi proov.Bas). Näide 2 on naissoost 1i tutvustamine ekraani taga (Sts). Pärast 15 min eemaldati sõel ja rottidel lasti kopuleerida (proovid 3 – 8). Themurdma kohta x-axis vastab andmete väljajätmisele kolmelt rottilt, kes on kopeeritud pikema perioodi jooksul algse emasloomaga. Viimased üheksa proovi olid samuti pidevad kronoloogiliselt. Proovid 9 ja 10 vastavad faasi 3 küllastumisperioodile (st 30 min ilma kinnituseta). Seejärel sisestati emane 1 ekraani taga (proov 11) ja 15 min hiljem, ekraani eemaldati (proov 12). Pärast 15 min kopulatsiooni puudumist asetati naise 2 ekraani taha (proov 13). Proovid 14 – 17 vastavad naissoost 2iga. Iga 15 min mikrodialüüsi prooviga seotud kinnitusdetailide, intromioonide või ejakulatsioonide arv on näidatud altriba graafik. Neurokeemilised andmed on väljendatud baasjoone kontsentratsioonide protsendina. NAC DA muudatused (suletud ruudud), DOPAC (suletud ringid) ja HVA (avatud ringid) väljavool esitatakse kui rea graafikud. Tehti järgmised võrdlused: lähteproov 1 versus proovid 2 – 10; uus algnäidis 10 versus proovid 11 ja 12; uus algnäidis 12 versus proovid 13 – 17 (*p <0.05; ** p <0.01). Iseseisev t testid viidi läbi baasväärtuste vahel (proovid 1, 10 ja 12). Märkimisväärsed erinevused esimesest lähtejoontest (proov 1), †p <0.05.
Naiste 1-i (proovid 1-12) ja naissoost 2 (proovid 12 – 17) seotud neurokeemilistele andmetele viidi läbi eraldi ühesuunalised korduvad mõõtmised. A priori võrreldi Dunni mitmekordse võrdluse testi abil (Bonferroni t). Tehti järgmised kolm peamist võrdlust: (1) esialgne lähtejoone (proov 1) versus 2 – 10 (esimene kokkupuude naissoost 1iga), (2) teine lähtejoone (proov 10) võrreldes 11i ja 12i proovidega (naissoost 1) ja (3) kolmas lähtejoone (proov 12) võrreldes 13 – 17 proovidega (2-i ekspositsioon).
Vastusena naissoost 1ile esines DA-väljavoolu märkimisväärne üldine muutus [F (11,44) = 8.48; p <0.001] ja naine 2 [F (5,20) = 2.83;p <0.05]. DA väljavoolu märkimisväärne suurenemine leiti, kui emane 1 viibis ekraani taga (+ 44%,p <0.05; valim 2). Kopulatsiooni ajal suurenesid DA kontsentratsioonid veelgi, saavutades maksimaalse väärtuse (+ 95%;p <0.01) esimese kopulatsioonihoo ajal (proov 3). DA püsis kogu kopulatsiooni ajal kõrgenenud ja naasis algtaseme kontsentratsioonini alles 30 minuti jooksul, mil monteerimist ei toimunud (proovid 9 ja 10). Naise 1 taaskehtestamine ekraani taha (proov 11) ega võimalus füüsiliselt suhelda, kuid ilma monteerimiseta (proov 12) ei suurendanud DA kontsentratsiooni võrreldes teise algväärtusega (proov 10). Naise 2 olemasolu ekraani taga (proov 13) põhjustas DA väljavoolu väikese kasvu (12%) võrreldes kolmanda algväärtusega (proov 12), mis ei saavutanud statistilist olulisust. Uue kopulatsiooni naisega 2 tulemuseks oli märkimisväärne (34%) kasv (p <0.05) DA väljavoolus esimese kopulatsiooniproovi ajal (proov 14). Ehkki järgmise kolme proovi jooksul jätkus nõrk kopulatsioonikäitumine, vähenes DA kontsentratsioon algväärtuseni (proovid 15–17). Iseseisev t „algtaseme” proovide (st 1, 10 ja 12) vahel läbi viidud testid näitasid, et need väärtused ei erinenud oluliselt.
Kolmes rottis, kes naasis 1i taasalustamisel kopulatsiooni alustasid, suurenesid NAC DA kontsentratsioonid naissoost 1i kohalolekul ekraani taga (vahemik, 25-47%) ja kopulatsiooni ajal (vahemik, 13-37%), võrreldes prooviga enne naise taaskehtestamist. Need tõusud toimusid siiski ainult siis, kui seksuaalkäitumine oli jõuline ja viis ejakulatsiooni.
DOPACi olulised üldised muudatused [F (11,44) = 9.57; p <0.001] ja HVA [F (11,44) = 12.47; p <0, 001] kontsentratsioonid leiti vastusena naisele 1, kuid mitte naisele 2. Metaboliidi kontsentratsioon suurenes veidi (+ 15% mõlemal juhul) emase 1 esitamise ajal ekraani taga (proov 2), kuid see ei olnud märkimisväärne statistiliselt. DOPAC ja HVA kontsentratsioonid suurenesid aga kopulatsiooni ajal (proovid 3–8), saavutades maksimaalsed väärtused (vastavalt +80 ja + 86%; p <0.01) 60 minuti pärast (mõlemal juhul proov 6). Kuigi metaboliitide kontsentratsioonid vähenesid seksuaalse passiivsuse perioodil kokkupuutel emasega 1 (proovid 9 ja 10), püsisid kontsentratsioonid siiski algtasemega võrreldes (p <0.05 mõlemal juhul). Naise 1 taaskehtestamine ekraani taha (proov 11), juurdepääs emasele 1 pärast ekraani eemaldamist (proov 12) ja emase 2 (proov 13) sissetoomine ei põhjustanud metaboliitide kontsentratsioonides mingeid muutusi. Kerge, kuid statistiliselt ebaoluline DOPAC ja HVA kontsentratsiooni tõus (mõlemal juhul + 23%) võrreldes algtasemega (proov 12) vastas esimesele naisega 2 (valim 14) tehtud kopulatsioonihoogule. See tõus oli lühiajaline ja vähenes ülejäänud kolme valimi (15–17) algväärtusteni. Iseseisev t„baasjoone” proovide (st 1, 10 ja 12) vahel läbi viidud testid näitasid, et teise ja kolmanda lähtejoone väärtused (vastavalt 10 ja 12 proovid), kuigi need ei erine üksteisest, jäid märkimisväärselt kõrgemaks võrreldes esimese algtaseme prooviga DOPACi ja HVA jaoks (p <0.05 mõlemal juhul).
Histoloogia
Mikrodialüüsi sondid asusid NAC-s (joon.2) vahemikus, mis ulatub + 1.20 kuni + 1.70 mm kaugusele bregmist (lame kolju). Samuti oli varieeruvus ka mediolateraalsel tasandil; andmed peegeldavad NAC-i kesta ja põhiregioonide valimit.
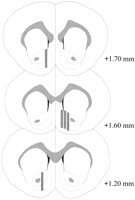
Vaata suuremat versiooni:
Joon. 2.
Mikrodialüüsi sondide asukoht Coolidge'i efekti katses kasutatud isasrottide NAC-s. Varjutatud ristkülikud vastab mikrodialüüsi sondide eksponeeritud membraanialale. Sarjast pärinevad koronaalsed aju lõigud võeti uuestiPaxinos ja Watson (1986).
ARUTLUS
Vastavalt varasematele aruannetele näitavad praegused tulemused parendatud mesolimbilise DA ülekandumist, mis on seotud isasroti seksuaalse käitumise isuäratavate ja tarbivate komponentidega.in vivo mikrodialüüs (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Lisaks annavad need tulemused neurokeemilise korrelatsiooni seksuaalse küllastuse ja sellele järgneva kopulatsiooni taasalustamiseks vastuseks uuele vastuvõtlikule naisele (Coolidge'i efekt). Käesolevad andmed näitavad, et uue vastuvõtliku naise stimuleerivad omadused võivad suurendada NAC DA ülekannet seksuaalselt küllastunud isasrottidel, mis omakorda võivad olla seotud seksuaalse käitumise taasalustamisega. See ilmneb kõigepealt NAC DA väikese suurenemise korral uue naise taga ekraani taga ja esineb kõige veenvamalt, kui naise 2iga taaskehtestatud kopulatsiooni ajal on märgatav kasv (joonis fig.1).
Esimese vastuvõtva naise olemasolu ekraani taga viis NAC DA effluxi tugevale isuäralikule tõusule (44% võrreldes algtasemega), mis oli sarnane suurusele eelmise, samasuguse konstruktsiooniga katsetes teatatud väärtusega (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992). Nende uuringutega kooskõlas oli ka tähelepanek, et NAC DA väljavool suurenes kopulatsiooni ajal veelgi (käesolevas katses> 95% kõrgem kui algväärtus). Kuigi me võime vaadata, et lõpptegevus on seotud täiustatud NAC DA vabastamisega (Wenkstern et al., 1993; Wilson jt, 1995) on oluline uurida mõisteid "isuäratav" ja "tarbiv" seksuaalse käitumise kontekstis. Kui faas, kus emane on ekraani taga, on ainult isuäratav või ettevalmistav, ei saa kopulatsioonifaasi ajal käitumist pidada puhtalt tarbivaks. Kuna "isuäratavat" saab kasutada kõigi käitumiste kirjeldamiseks, mis põhjustavad motiveeritud käitumise (kopulatsioon), siis on meeste eksponaatide esmane käitumine "tarbivas" faasis kõige paremini kirjeldatud kui isuäratav; mees veedab suurema osa oma ajast ja jõupingutustest naise poole kopuleerimiseks. Sellega seoses saame korreleerida maksimaalset NAC DA ülekannet tarbimisega sama hästi kui meeste rottide seksuaalkäitumise intensiivne isuäratav komponent.
Juurdepääs uuele naissoost naissoost tõi kaasa igaühe taasühinemise. Varasemad uuringud on näidanud, et enamus rottidest, kellel oli luba kopuleeruda küllastumisvõimele, kasutades samasugust käitumisprotokolli kui käesolevas eksperimendis kasutatud, ei jätkanud paaritumist, kui testiti 24 h hiljem (Rand ja Jordaania, 1956). On tõenäoline, et naissoost 2i uute stimuleerivate omaduste olemasolu, mis võisid sisaldada nii haistmist kui ka visuaalset ja kuuldavat märku, viisid uuendatud kopulatsiooni. Huvitav küsimus, millele tuleb veel vastata, on see, millisel mehhanismil eristab meesrott uudset naist naissoost, kellega ta hiljuti paaritas. Selle mehhanismi sait võib olla peamises haistmissüsteemis. On teatatud, et selle süsteemi terviklikkus on Coolidge'i efekti jaoks hamstrite puhul oluline.Johnston ja Rasmussen, 1984). Vomeronasali-lisandiga lõhnasüsteem, kus feromonaalse mälu protsessi kirjeldati hiirtel hiljuti (Kaba et al., 1994) on ka peamine kandidaat. Sellega seoses tuleb märkida, et NAC DA ülekande suurenemist mõõdeti kasutades in vivo voltamomeetria isastel rottidel, kes olid varustatud emasrottidele, kes olid nakatunud (Louillot jt, 1991; Mitchell ja Gratton, 1992). Lisaks kasutatakse K+ NAC DA ülekande suurendamiseks piisab otseselt haistmislampi lisatarviku vomeronaalsest närvikihist, aga ka lisatõbi pirnist.Mitchell ja Gratton, 1992).
Esimese 15 min-ga seotud kopulatsioon naissoost 2iga oli seotud NAC DA olulise suurenemisega. Erinevalt naissoost 1-ist ei tekitanud interaktsioon naissoost 2-iga sama suurusega NAC DA väärtusi nii isuäratavas (12%) kui ka tarbivas (34%) faasis. Need väikesed NAC DA suurenemised korreleeruvad hästi naissoost 2iga tähistatud seksuaalse käitumise vähenenud tasemega võrreldes naissoost 1iga. Metaboliitide kontsentratsioonid jäid küllastuse faasis kõrgeks, mille tulemusena tekkisid uued algtaseme kontsentratsioonid (proovid 10 ja 12), mis tõusid oluliselt algväärtuse väärtusest (proov 1).
DOPACi ja HVA kontsentratsiooni suurenemise ajaline lagunemine kopulatsiooni ajal on kooskõlas nende moodustumisega algühendi, DA, metaboliitidena. On oletatud, et mikrodialüüsi metaboliidi kontsentratsioonid, vähemalt loomuliku käitumise ajal, mis ei ole farmakoloogiliselt juhitud, annavad kasuliku neuronaalse aktiivsuse indeksi (Damsma et al., 1992; Fumero et al., 1994). Asjaolu, et metaboliitide kontsentratsioonid jäid kõrgeks isegi selle katse seksuaalse inaktiivsuse perioodidel, kui DA kontsentratsioonid olid tagasi algväärtuse algväärtusteks, seab selle ettepaneku kahtluse alla.
Selles katses ilmnenud DA-metaboliitide kontsentratsioonide püsiv suurenemine peegeldab DA-metaboliitide mediaalset profopunktiprofiili, mida täheldati rottidel esimesel päeval pärast seda, kui nad olid küllastunud.Mas et al., 1995a,b). DOPAC- ja HVA-kontsentratsioonide püsivat tõusu NAC-s või mPOA-s ei täheldata alati, kui paaritumisperiood on kindlaksmääratud kestusega, palju lühem kui küllastumise saavutamiseks vajalik aeg. Näiteks on paljud uuringud näidanud, et DOPAC-i kontsentratsioonid suurenesid ja jäid kopulatsiooni ajal kõrgeks, kuid langesid kohe pärast naissoost eemaldamist algtasemeni.Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Uuringus Mas et al. (1995b)DOPACi ja HVA põhilised ekstratsellulaarsed kontsentratsioonid mPOA-s jäid 4i järjestikuste päevade jooksul kõrgemaks, mis vastab seksuaalse inaktiivsuse perioodile. Neljandaks päevaks, vahetult enne loomade taasühinemist, olid metaboliitide baaskontsentratsioonid lähedal eelsoodumusele. Autorid võrdlesid neurokeemiliste muutuste mustrit nendega, mida täheldati pärast DA-retseptorite blokeerijate manustamist.Zetterström jt, 1984; Imperato ja DiChiara, 1985) ja on näidanud, et seksuaalse tegevusetuse seisund võib olla vahendatud prolaktiini vabanemise kaudu, mis võib toimida endogeense neuroleptikuna (Mas et al., 1995a,b,d). On selge, et neuroleptilise manustamisega kaasneb ekstratsellulaarse metaboliidi kontsentratsiooni suurenemine ja \ tZetterström jt, 1984; Imperato ja DiChiara, 1985). Kahjuks Mas et al. (1995a,b) ei suutnud tuvastada mPOA DA kontsentratsioone. Käesolevas uuringus taastasid DA-kontsentratsioonid NAC-s eelkombinatsiooni väärtused, samas kui DOPAC ja HVA kontsentratsioonid jäid kõrgeks. See muster on vastuolus NAC-is toimiva endogeense neuroleptiku rolliga seksuaalse küllastuse esilekutsumiseks.
Arvestades mesolimbiliste DA-neuronite osalemist motiveeritud käitumises (Fibiger ja Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) ja nende tundlikkus uute keskkonnaalaste stiimulite suhtes (Fabre et al., 1983; Schultz, 1992; Mirenowicz ja Schultz, 1994), NAC DA ekstratsellulaarsete kontsentratsioonide suurenemine vastusena uuele naissoost on kooskõlas hüpoteesiga, et aktiivsus selles DA-süsteemis on oluline seksuaalse käitumise taastamiseks. Lisaks teatatakse DA-ülekande isu ja tarbimise suurenemisest (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) ja neuronaalne aktiivsus (Shimura et al., 1994) isaste rottide mPOA-s viitavad seksuaalse käitumise põhjal, et see struktuur võib samuti kaasa aidata Coolidge'i efektile iseloomuliku uuendatud kopulatsiooni tekkele.
Kooskõlas mesolimbilise DA süsteemi üldise rolliga motiveeritud käitumises on hästi kindlaks tehtud, et ka ekstratsellulaarsed DA kontsentratsioonid on kõrgendatud enne sööki, selle ajal ja vahetult pärast söögi tarbimist, lähtudes algväärtustest ∼30 min hiljem (Wilson jt, 1995). On hästi teada, et toidu poolt põhjustatud küllastust mõjutavad selle sensoorsed omadused. Inimesed ja loomad lükkavad toitu, millele nad küllastati, ja söövad teisi toiduaineid, mida ei olnud söödud (Rullid, 1986). See tõstatab küsimuse, kas ekstratsellulaarset DA-väljavoolu NACis suurendatakse selektiivselt uue toidukoguse esitamisega, kuid mitte toiduga, mida tarbitakse hiljuti küllastumisvõimelisusele viisil, mis on analoogne käesolevas uuringus esitatud andmetega. seksuaalne motivatsioon. Kui see on kinnitust leidnud, tähendaks see looduslike hüvede, küllastustunde ja mesolimbilise DA ülekandumise sensoorsete omaduste üldine seos selle närvisüsteemi kriitilist rolli motivatsiooniprotsesside reguleerimisel, mille katkemine võib põhjustada tõsiseid söömise ja seksuaalse funktsiooni häireid .
VIITED
- ↵
- FA FA,
- Jordaania L
(1956) Meesroti seksuaalne ammendumine ja taastumine. QJ Exp Psychol 8: 121 - 133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Coolidge'i toime ajalised omadused isaste rottide kopulatsiooni käitumises. J Comp Physiol Psychiatry 65: 447 - 452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Dopamiin toimib isuäratavas ja kaitslikus käitumises. Prog Neurobiol 39: 247 - 279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Seksuaalne käitumine suurendab dopamiini ülekandumist isaste rottide tuumas ja striatumis: võrdlus uudsuse ja liikumisega. Behav Neurosci 106: 181 - 191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Neuronite aktiivsus käitava ahvi ventraalses tegmentaalses piirkonnas. Behav Brain Res 9: 213 - 235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Tasu, motivatsioon, tunnetus: mesotelentsiaalsete dopamiinisüsteemide psühobioloogia. füsioloogia käsiraamatus: närvisüsteem IV, toimetised Bloom FE, Geiger SD (Ameerika füsioloogiaühing, Bethesda, MD), lk 647 – 675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Tasuvuskohtade elektriline stimuleerimine ventralises tegmentaalpiirkonnas suurendab dopamiini ülekannet roti tuumas. Behav Brain Res 55: 131 - 141.
- ↵
- Fisher A
(1962) Stimulatsiooni varieerumise mõju seksuaalsele küllastumisele isastel rottidel. J Comp Physiol Psychiatry 55: 614 - 620.
- ↵
- Fowler H
- Whalen RE
(1961) Meeste rottide stimuleeriva stiimuli ja seksuaalse käitumise varieerumine. J Comp Physiol Psychiatry 54: 68 - 71.
- ↵
- Fumero B,
- Fernendez-Vera JR
- Gonzalez-Mora JL
- Mas M
(1994) Monoamiini käibe muutused eesnäärme piirkondades, mis on seotud meessoost seksuaalse käitumisega: mikrodialüüsi uuring. Brain Res 662: 233 - 239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) IBM-PC-põhine andmete kogumise süsteem näriliste seksuaalse käitumise salvestamiseks ja üldiseks sündmuste salvestamiseks. Physiol Behav 44: 825 - 828.
- ↵
- Hull EM,
- Eaton RC,
- Mooses J,
- Lorrain DS
(1993) Kopulatsioon suurendab dopamiini aktiivsust isaste rottide keskmises preoptilises piirkonnas. Life Sci 52: 935 - 940.
- ↵
- Hull EM,
- Jianfang D
- Lorrain DS
- Matuszewich L
(1995) Ekstratsellulaarne dopamiin mediaalse preoptilises piirkonnas: mõju seksuaalsele motivatsioonile ja kopulatsiooni hormonaalsele kontrollile. J Neurosci 15: 7465 - 7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Dopamiini vabanemine ja metabolism ärkvel olevates rottides pärast süsteemset neuroleptikut, nagu on uuritud trans-striaalses dialüüsis. J Neurosci 5: 297 - 306.
- ↵
- Johnston RE
- Rasmussen K
(1984) Naiste hamstrite individuaalne tunnustamine meeste poolt: keemiliste märkide ja lõhna- ja vomeronasaalsete süsteemide roll. Physiol Behav 33: 95 - 104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T
- Nakanishi S
(1994) Lõhnamälu esilekutsumine metabotroopse glutamaadi retseptori aktiveerimise teel. teadus 265: 262 - 264.
- ↵
- Kalivas PW,
- Sorg BA,
- Konksud MS
(1993) Psühhostimulantide suhtes sensibiliseerimise farmakoloogia ja neuraalskeem. Behav Pharmacol 4: 315 - 334.
- ↵
- LeMoal M
(1995) Mesokortikolimbilised dopamiinergilised neuronid. Funktsionaalsed ja regulatiivsed rollid. Psühhofarmakoloogias: neljanda põlvkonna edusammud, toimetused Bloom FE, Kupfer DJ (Raven, New York), lk 283 – 294.
- ↵
- Louillot A
- Gonzalez-Mora JL
- Guadalupe T,
- Mas M
(1991) Seksist tingitud lõhnaaine stiimulid indutseerivad dopamiini vabanemise selektiivset suurenemist isaste rottide tuumas. Brain Res 553: 313 - 317.
- ↵
- Mas M,
- Gonzalez-Mora JL
- Louillot A
- Ainus C,
- Guadalupe T
(1990) Suurenenud dopamiini vabanemine kopuliseerivate isaste rottide tuumaseadmetes, mida tõendab in vivo voltammeetria. Neurosci Lett 110: 303 - 308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR
- Gonzalez-Mora JL
(1995a) In vivo mikrodialüüsi abil hinnatud seksuaalse ammendumise ja taastumise neurokeemilised korrelatsioonid. Brain Res 675: 13 - 19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Voltammetriline ja mikrodialüüsi seire aju monoamiini neurotransmitterite vabanemisele sotsioseksuaalsete interaktsioonide ajal. Behav Brain Res 71: 69 - 79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Apomorfiini kaasneva käitumise esilekutsumine seksuaalsetes rottides. Eur J Pharmacol 280: 331 - 334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
- Gonzalez-Mora JL
(1995d) Seksuaalse küllastuse neurokeemia. Inhibeeritud soovi eksperimentaalne mudel. seksuaalse funktsiooni ja düsfunktsiooni farmakoloogias, ed Bancroft J (Raven, New York), lk 115 – 126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Primaadi dopamiinergiliste neuronite tasuvastuse ettearvamatuse tähtsus. J Neurofüsiol 72: 1024 - 1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Mesolimbiline dopamiini vabanemine, mis tekkis lisanduva haistmissüsteemi aktiveerimisel: kiire kronoamperomeetriline uuring. Neurosci Lett 140: 81 - 84.
- ↵
- Paxinos G,
- Watson C
(1986) Roti aju stereotaksilistes koordinaatides (2nd ed). (Academic, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Seksuaalne käitumine suurendab tsentraalset dopamiini transmissiooni isastel rottidel. Brain Res 530: 345 - 348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Seksuaalse käitumise psühhofarmakoloogia. Psühhofarmakoloogias: neljanda põlvkonna edusammud, toimetused Bloom FE, Kupfer DJ (Raven, New York), lk 743 – 758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioidid ja seksuaalne käitumine. Neurosci Biobehav Rev 11: 1 - 34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Positiivsete emotsionaalsete seisundite neurobioloogilised korrelatsioonid: dopamiin, prognoosimine ja tasu. emotsionaalsete uuringute rahvusvahelises ülevaates, ed Strongman (Wiley, New York), lk 31 – 50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Dopamiini vabanemise korrelatsioon tuuma accumbensis meheliku seksuaalse käitumisega rottidel. Brain Res 524: 160 - 163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Seksuaalse ammendumise tühistamine serotonergiliste ja noradrenergiliste ainete abil. Behav Brain Res 62: 127 - 134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Keskse noradrenergilise süsteemi osalemine seksuaalselt ammendunud rottide kopulatsioonikäitumise taastamisel yohimbiini, naloksooni ja 8-OH-DPAT abil. Brain Res Bull 38: 399 - 404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Opioidantagonistid ja seksuaalse küllastumise nähtus. Psychopharmacol 122: 131 - 136.
- ↵
- Rolls BJ
(1986) Sensoorne spetsiifiline küllastuvus. Nutr Rev 44: 93 - 101.
- ↵
- Salamone JD
(1996) Motivatsiooni käitumuslik neurokeemia: metoodilised ja kontseptuaalsed küsimused tuuma accumbens dopamiini dünaamilise aktiivsuse uuringutes. J Neurosci meetodid 64: 137 - 149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R
- Tsukamoto T,
- Kumamoto Y
(1995) Dopamiini vabanemine keskmises preoptilises piirkonnas rottide kopulatsiooni ajal. Brain Res 692: 66 - 70.
- ↵
- Schultz W
(1992) Dopamiini neuronite aktiivsus käitumisprimaadis. Semin Neurosci 4: 129 - 138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) Mediaalne preoptiline piirkond on seotud nii seksuaalse erutusega kui ka isasrottide esinemisega: neuronite aktiivsuse ümberhindamine vabalt liikuvatel loomadel. Brain Res 640: 215 - 222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Dopamiini ülekanne suureneb isaste rottide tuumaklundis nende esmakordsel kokkupuutel emasloomadega, kes on seksuaalselt vastuvõtlikud. Brain Res 618: 41 - 46.
- ↵
- Wilson C,
- Nomikos GG
- Collu M,
- Fibiger HC
(1995) Dopamiinergilised korrelatsioonid motiveeritud käitumisest: ajami tähtsus. J Neurosci 15: 5169 - 5178.
- ↵
- Wilson JR
- Kahn RE,
- FA FA
(1963) Meeste rottide seksuaalkäitumise muutmine stimuleeriva naise muutmise teel. J Comp Physiol Psychiatry 56: 636 - 644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Neuroleptiliste ravimite mõju striataalse dopamiini vabanemisele ja ainevahetusele ärkvelehes, mida uuriti intratserebraalse dialüüsi abil. Eur J Pharmacol 106: 27 - 37.
Artiklid, mis viitavad sellele artiklile
- Dopamiinergiliste neuronite endogeenne opioidide poolt indutseeritud neuroplastsus Ventral Tegmental piirkonnas mõjutab loomulikku ja opiaadi tasu Journal of Neuroscience, 25 juuni 2014, 34 (26): 8825-8836
- Tasakaalustatus patoloogiliste hasartmängude erinevat tüüpi hüvede suhtes Aju, 1 August 2013, 136 (8): 2527-2538
- Loodus- ja ravimihüvitiste seadus ühiste närviplastilisuse mehhanismide kohta koos {Delta} FosB-ga peamise vahendajana Journal of Neuroscience, 20 veebruar 2013, 33 (8): 3434-3442
- Neuronaalsed reaktsioonid Nucleus Accumbens Shell'is meessoost rotid-i seksuaalse käitumise ajal Journal of Neuroscience, 1 veebruar 2012, 32 (5): 1672-1686
- Romantiline armastus: imetajate aju süsteem paarivalikuks Royal Society filosoofilised tehingud B: bioloogilised teadused, 29 detsember 2006, 361 (1476): 2173-2186
- Hedoonilised kuumad kohad ajus Neuroteadlane, 1 detsember 2006, 12 (6): 500-511
- Dopamiinergiliste korrelatsioonide keskne ja basolateraalne amügdalaaegne söötmine rottide tuumas Accumbensis ja Medial Prefrontal Cortexis Journal of Neuroscience, 15 detsember 2002, 22 (24): 10958-10965
- Dopamiini kontsentratsiooni transientide sagedus Meeste rottide selja- ja Ventral Striatum'i suurenemine spetsiifiliste preparaatide kasutamisel Journal of Neuroscience, 1 detsember 2002, 22 (23): 10477-10486
- Raamatu ülevaade: ekstratsellulaarse dopamiini dünaamika kokaiini ägedates ja kroonilistes toimingutes Neuroteadlane, 1 August 2002, 8 (4): 315-322
- NMDA, AMPA / Kainate ja dopamiini retseptorite diferentsiaalne kaasamine tuuma Accumbens'i tuumas Pavlovia lähenemise käitumise omandamisel ja toimimisel Journal of Neuroscience, 1 detsember 2001, 21 (23): 9471-9477
- Ravimiga seotud stiimulite toimel kokaiiniga seotud käitumise kontroll rottidel: Toime pärast kustutatud operandi reageerivate ja ekstratsellulaarsete dopamiinitasemete taastumist amygdala ja tuumaklundide korral PNAS, 11 aprill 2000, 97 (8): 4321-4326
- Seksuaalse käitumise ja tõhustatud dopamiini effluxi hõlbustamine meeste rottide tuumas Accumensis pärast D-amfetamiini poolt põhjustatud käitumuslikku sensibiliseerimist Journal of Neuroscience, 1 jaanuar 1999, 19 (1): 456-463
- NEUROTUUDUS: aju tähelepanu äratamine Teadus, 3 oktoober 1997, 278 (5335): 35-37
- Sensoorse spetsiifilise küllastuse dopamiinergilised korrelatsioonid rottide keskmises eesnäärmes ja tuumasõlmes Journal of Neuroscience, 1 oktoober 1999, 19 (19): RC29
