Dev Rev. 2008; 28(1): 62-77. doi: 10.1016/j.dr.2007.08.003
Abstraktne
Noorukiea on arenguperiood, mida iseloomustavad ebaoptimaalsed otsused ja tegevused, mis põhjustavad tahtmatute vigastuste ja vägivalla, alkoholi ja narkootikumide kuritarvitamise, soovimatu raseduse ja sugulisel teel levivate haiguste sagenemist. Traditsioonilised neurobioloogilised ja kognitiivsed selgitused noorukite käitumise kohta ei ole suutnud võtta arvesse mittelineaarseid muutusi käitumises, mida täheldati noorukieas, võrreldes lapsepõlve ja täiskasvanueaga. See ülevaade annab bioloogiliselt usutava kontseptsiooni nende mittelineaarsete käitumismuutuste aluseks olevate närvimehhanismide kohta, mis on kõrgendatud reageerimisvõime stiimulitele, samas kui impulsside juhtimine on sellel perioodil veel suhteliselt ebaküps. Hiljutised inimese pildistamise ja loomkatsed annavad sellele vaatele bioloogilise aluse, mis viitab limbilise tasustamise süsteemide erinevale arengule võrreldes ülalt-alla kontrollisüsteemidega noorukieas võrreldes lapsepõlve ja täiskasvanueaga. See arengumuster võib halveneda nendel noorukitel, kellel on kalduvus riskida, suurendades halbade tulemuste riski.
Riikliku tervisestatistika keskuse andmetel sureb Ameerika Ühendriikides igal aastal üle 13,000 70 nooruki. Ligikaudu XNUMX% nendest surmajuhtumitest on põhjustatud mootorsõidukiõnnetustest, tahtmatutest vigastustest, mõrvast ja enesetapust (Eaton et al., 2006). 2005. aasta riikliku noorte riskikäitumise uuringu (YRBS) tulemused näitavad, et noorukid juhivad sõidukit pärast joomist või ilma turvavööta, kannavad relvi, kasutavad ebaseaduslikke aineid ja astuvad kaitsmata seksi. mis põhjustab soovimatut rasedust ja suguhaigusi, sealhulgas HIV-nakkust (Eaton et al., 2006). See statistika rõhutab riskantsete valikute ja tegude mõistmise tähtsust noorukite puhul.
On postuleeritud mitmeid kognitiivseid ja neurobioloogilisi hüpoteese, miks noorukid valivad ebaoptimaalset käitumist. Noorukite aju arengut käsitleva kirjanduse hiljutises ülevaates Yurgelun-Todd (2007) viitab sellele, et kognitiivne areng noorukieas on seotud kognitiivse kontrolli võimekuse järk-järgult suurema tõhususega. Seda efektiivsust kirjeldatakse sõltuvana prefrontaalse ajukoore küpsemisest, mida tõendab suurenenud aktiivsus fokaalsetes prefrontaalsetes piirkondades (Rubia et al., 2000; Tamm, Menon & Reiss, 2002) ja vähenenud aktiivsus ebaolulistes ajupiirkondades (Brown et al., 2005; Durston et al., 2006).
See üldine muster, mille kohaselt paraneb kognitiivne kontroll koos prefrontaalse ajukoore küpsemisega, viitab lineaarsele arengule lapsepõlvest täiskasvanueani. Kuid noorukieas täheldatud suboptimaalsed valikud ja tegevused kujutavad endast mittelineaarset käitumise muutust, mida saab eristada lapsepõlvest ja täiskasvanueast, nagu tõendab noorukite käitumise ja suremuse riiklik tervisestatistika keskus. Kui kognitiivne kontroll ja ebaküps prefrontaalne ajukoor olid suboptimaalse valikukäitumise aluseks, peaksid lapsed nende vähem arenenud prefrontaalset ajukoort ja kognitiivseid võimeid arvestades välja nägema märkimisväärselt sarnased või isegi halvemad kui noorukid. Seega ei saa ainuüksi ebaküps prefrontaalne funktsioon põhjustada noorukite käitumist.
Kognitiivsete ja neurobioloogiliste muutuste täpne kontseptualiseerimine noorukieas peab ravima noorukit kui üleminekuperioodi (Spear, 2000), selle asemel, et teha üksainus hetktõmmis (Casey, Tottenham, Liston ja Durston, 2005). Teisisõnu, selle arenguperioodi mõistmiseks on vaja üleminekuid noorukieasse ja sellest välja, et eristada selle arenguetapi eritunnuseid. Kognitiivsete ja närviprotsesside arengutrajektooride loomine on nende üleminekute iseloomustamiseks ja selle perioodi käitumise muutuste tõlgenduste piiramiseks hädavajalik. Kognitiivsel või käitumuslikul tasandil iseloomustatakse noorukeid kui impulsiivseid (st neil puudub kognitiivne kontroll) ja riskivalmidus nende konstruktsioonidega, mida kasutatakse sünonüümidena ja ei hinnata kummagi erinevat arengutrajektoori. Neurobioloogilisel tasandil viitavad inimese pildistamine ja loomuuringud närvisüsteemidele erinevatele neurobioloogilistele alustele ja arengutrajektoore, mis on nende impulsside kontrolli ja riskantsete otsuste konstruktsioonide aluseks.
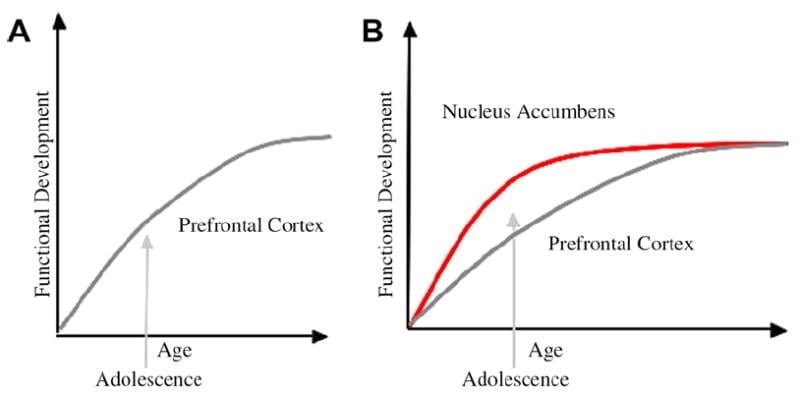
Oleme selle raames välja töötanud noorukite arengu neurobioloogilise mudeli, mis põhineb näriliste mudelitel (Laviola, Adriani, Terranova ja Gerra, 1999; Spear, 2000) ja hiljutised noorukieas tehtud uuringud (Ernst et al., 2005; Galvan, Jänes, Voss, Glover ja Casey, 2007; Galvan et al., 2006). Joon. 1 allpool kujutatakse seda mudelit. Vasakul on traditsiooniline noorukiea iseloomustus, mis on seotud peaaegu eranditult prefrontaalse ajukoore ebaküpsusega. Paremal on meie pakutud neurobioloogiline mudel, mis illustreerib, kuidas limbilisi subkortikaalseid ja prefrontaalseid ülalt-alla kontrollpiirkondi tuleb koos käsitleda. Koomiks illustreerib nende süsteemide erinevaid arengutrajektoore, kusjuures limbilised süsteemid arenevad varem kui prefrontaalsed kontrollpiirkonnad. Selle mudeli kohaselt on indiviid noorukieas rohkem kallutatud funktsionaalselt küpsete limbiliste piirkondade poolt (st limbilise tasakaalustamatus võrreldes prefrontaalse kontrolliga), võrreldes lastega, kelle puhul need süsteemid (st limbilise ja prefrontaalse) on alles kujunemas; ja võrreldes täiskasvanutega, kelle jaoks need süsteemid on täielikult küpsed. See vaatenurk annab aluse mittelineaarsetele muutustele käitumises kogu arengus, mis on tingitud selle limbilise varasemast küpsemisest võrreldes vähem küpsega ülalt-alla prefrontaalse kontrollpiirkonnaga. Tänu arengule ja kogemustele pakub funktsionaalne ühenduvus nende piirkondade vahel mehhanismi nende piirkondade ülevalt alla juhtimiseks (Hare, Voss, Glover ja Casey, 2007a). Lisaks ühitab mudel noorukieas riskikäitumise tervisestatistika vastuolulisuse ja terava tähelepanekuga. Reyna ja Farley (2006) et noorukid suudavad arutleda ja mõista nende käitumise riske. Meie mudeli kohaselt võidab limbilise süsteem emotsionaalselt silmatorkavates olukordades juhtimissüsteemide üle, arvestades selle küpsust prefrontaalse juhtimissüsteemiga võrreldes. Seda mudelit toetavate käitumis- ja inimeste pildistamise uuringute tõendid on esitatud rahuldust pakkuvas ja emotsionaalses kontekstis tehtud toimingute kontekstis (Galvan et al., 2006, 2007; Hare, Voss, Glover ja Casey, 2007b; Hare jt, 2007a). Lisaks spekuleerime selle üle, miks aju võib sellisel viisil areneda ja miks mõnel teismelisel võib olla suurem risk teha ebaoptimaalseid otsuseid, mis viivad halvemate pikaajaliste tulemusteni (Galvan et al., 2007; Hare jt, 2007b).
Arvatakse, et noorukite käitumise traditsiooniline seletus on tingitud prefrontaalse ajukoore (A) pikaajalisest arengust. Meie mudel võtab arvesse prefrontaalse ajukoore arengut koos subkortikaalsete limbiliste piirkondadega (nt nucleus accumbens), mis on seotud riskantsete valikute ja tegevustega (B).
Eesmärgile suunatud käitumise arendamine
Kognitiivse arengu nurgakiviks on võime suruda alla ebasobivad mõtted ja tegevused eesmärgipäraste kasuks, eriti mõjuvate stiimulite olemasolul.Casey, Galvan ja Jänes, 2005; Casey et al., 2000b; Casey, Thomas, David-son, Kunz ja Franzen, 2002a; Casey, Tottenham ja Fossella, 2002b). Mitmed klassikalised arenguuuringud on näidanud, et see võime areneb lapsepõlves ja noorukieas (Case, 1972; Flavell, Feach ja Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, 1970). Mitmed teoreetikud on väitnud, et kognitiivne areng on tingitud töötlemise kiiruse ja efektiivsuse suurenemisest, mitte vaimse võimekuse suurenemisest (nt. Bjorkland, 1985; Bjorkland, 1987; Case, 1985). Teised teoreetikud on oma kognitiivse arengu käsitlusse lisanud "inhibeerivate" protsesside konstruktsiooni.Harnishfeger ja Bjorkland, 1993). Selle kirjelduse kohaselt iseloomustab ebaküpset tunnetust vastuvõtlikkus konkureerivate allikate häiretele, mida tuleb alla suruda (nt. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz ja Franzen, 2002a; Dempster, 1993; Teemant, 1985; Munakata & Yerys, 2001). Seega nõuab eesmärgipärane käitumine tulemuste optimeerimiseks impulsside kontrolli või rahulduse viivitamist ning see võime näib küpsevat lapsepõlves ja noorukieas.
Noorukite käitumist on kirjeldatud kui impulsiivset ja riskantset, peaaegu sünonüümidena, kuid need konstruktsioonid tuginevad erinevatele kognitiivsetele ja neuraalsetele protsessidele, mis viitavad erinevatele konstruktsioonidele erinevate arengutrajektooridega. Täpsemalt näitab kirjanduse ülevaade, et impulsiivsus väheneb vanuse kasvades lapsepõlves ja noorukieas.Casey et al., 2002a; Casey, Galvan jt, 2005; Galvan et al., 2007) ja seda seostatakse prefrontaalse ajukoore pikaajalise arenguga (Casey, Galvan jt, 2005), kuigi on erinevusi selles, mil määral on antud indiviid impulsiivne või mitte, olenemata vanusest.
Vastupidiselt impulss-/kognitiivsele kontrollile näib riskide võtmine noorukieas suurenevat võrreldes lapsepõlve ja täiskasvanueaga ning see on seotud subkortikaalsete süsteemidega, mis on teadaolevalt seotud hüvede hindamisega. Inimese pildistamise uuringud, mis vaadatakse läbi, viitavad subkortikaalse aktivatsiooni (nt accumbens) suurenemisele riskantsete valikute tegemisel (Kuhnen & Knutson, 2005; Matthews ja teised, 2004; Montague & Berns, 2002), mis on noorukitel laste ja täiskasvanutega võrreldes liialdatud (Ernst et al., 2005; Galvan et al., 2006). Need leiud viitavad erinevatele trajektoore premeerimisel või stiimulil põhineval käitumisel, kusjuures nende süsteemide varasem väljatöötamine võrreldes juhtimissüsteemidega, mis näitavad pikaajalist ja lineaarset arengusuunda, pidades silmas ebasobivate valikute ja tegevuste ületamist eesmärgipäraste kasuks.
Inimese aju arengu neuroimaging uuringute tõendid
Hiljutised noorukite aju arengu uuringud põhinevad neuropildistamise metoodikate edusammudel, mida saab hõlpsasti kasutada arenevate inimpopulatsioonide jaoks. Need meetodid põhinevad magnetresonantstomograafia (MRI) meetoditel (vt Joon. 2) ja sisaldavad: struktuurset MRT-d, mida kasutatakse struktuuride suuruse ja kuju mõõtmiseks; funktsionaalne MRI, mida kasutatakse ajutegevuse mustrite mõõtmiseks; ja difusioontensorkujutis (DTI), mida kasutatakse valgeaine kiudude ühenduvuse indekseerimiseks. Tõendeid meie kortikaalsete ja subkortikaalsete piirkondade vahelise konkurentsi arengumudeli kohta toetab ebaküps struktuurne ja funktsionaalne ühenduvus, mida mõõdetakse vastavalt DTI ja fMRI abil.
Inimese aju arengu MRI uuringud
Mitmed uuringud on kasutanud struktuurset MRI-d normaalse aju arengu anatoomilise kulgemise kaardistamiseks (vt ülevaadet Durston et al., 2001). Kuigi aju kogumaht on kuueaastaselt ligikaudu 90% täiskasvanu suurusest, toimuvad aju halli ja valge aine alamkomponendid dünaamilistes muutustes kogu noorukieas. Hiljutiste pikisuunaliste MRI uuringute andmed näitavad, et halli aine mahul on ümberpööratud U-kujuline muster, mille piirkondlikud erinevused on suuremad kui valgel ainel (Giedd, 2004; Gogtay et al., 2004; Sowell jt, 2003; Sowell, Thompson ja Toga, 2004). Üldiselt küpsevad esimesed funktsioonid, näiteks mootori- ja sensoorsüsteemid, alad; kõrgema astme assotsiatsioonipiirkonnad, mis integreerivad need esmased funktsioonid, küpsevad hiljem (Gogtay et al., 2004; Sowell, Thompson ja Toga, 2004). Näiteks näitavad MRI-põhiseid mõõtmisi kasutavad uuringud, et koore hallainete kadu tekib kõige varem primaarsetes sensorimootori piirkondades ja hiljemalt dorsolateraalsetes prefrontaalsetes ja külgsetes ajalistes koorikutes (Gogtay et al., 2004). See muster on kooskõlas inimpäritolu primaadi ja inimjärgsete postmortemiuuringutega, mis näitavad, et prefrontaalne ajukoor on üks viimastest küpse aju piirkondadest (Bourgeois, Goldman-Rakic ja Rakic, 1994; Huttenlocher, 1979). Vastupidiselt hallainele suureneb valgeaine maht ligikaudu lineaarselt, suurenedes kogu arengu jooksul täiskasvanueas (Gogtay et al., 2004). Need muutused peegeldavad arvatavasti aksonite müeliniseerumist oligodendrotsüütide poolt, mis suurendavad neuronite juhtivust ja kommunikatsiooni.
Kuigi struktuursete muutuste uurimisel on vähem tähelepanu pööratud subkortikaalsetele piirkondadele, täheldatakse mõningaid suurimaid muutusi ajus kogu arengu jooksul just nendes piirkondades, eriti basaalganglionides.Sowell et al., 1999Vt Joon. 3) ja eriti meestel (Giedd et al., 1996). Arengu muutused struktuurses mahus basaalganglionides ja prefrontaalsetes piirkondades on huvitavad teadaolevate arenguprotsesside (nt dendriitne arboriseerumine, rakusurm, sünaptiline pügamine, müelinisatsioon) valguses, mis toimuvad lapsepõlves ja noorukieas. Need protsessid võimaldavad peenhäälestada ja tugevdada seoseid prefrontaalsete ja subkortikaalsete piirkondade vahel koos arengu ja õppimisega, mis võib kattuda suurema kognitiivse kontrolliga. Kuidas on need struktuurimuutused seotud kognitiivsete muutustega? Mitmed uuringud on seotud otsmikusagara struktuurse küpsemise ja kognitiivse funktsiooniga, kasutades neuropsühholoogilisi ja kognitiivseid meetmeid (nt. Sowell et al., 2003). Täpsemalt on teatatud seostest MRI-l põhinevate prefrontaalsete ajukoore ja basaalganglioni piirkondlike mahtude ning kognitiivse kontrolli meetmete vahel (st võime alistada sobimatu reaktsioon teise kasuks või maha suruda tähelepanu ebaolulisele stiimuli atribuudile vastava stiimuli atribuudi kasuks).Casey, Trainor jt, 1997). Need leiud viitavad sellele, et kognitiivsed muutused peegelduvad aju struktuursetes muutustes ja rõhutavad nii subkortikaalse (basaalganglioni) kui ka kortikaalse (nt prefrontaalse ajukoore) arengu tähtsust.

DTI uuringud inimese aju arengu kohta
Läbi vaadatud MRI-põhised morfomeetrilised uuringud viitavad sellele, et kortikaalseid ühendusi peenhäälestatakse, kõrvaldades sünapside ülekülluse ja tugevdades asjakohaseid seoseid arengu ja kogemustega. Hiljutised edusammud MRI tehnoloogias, nagu DTI, pakuvad potentsiaalset vahendit konkreetsete valgeaine traktide rolli üksikasjalikumaks uurimiseks aju arengus ja käitumises. Selle artikli jaoks on olulised neuroimaging uuringud, mis on seostanud kiudude arengut kognitiivsete võimete paranemisega. Täpsemalt on näidatud seoseid DTI-põhiste prefrontaalse valgeaine arengu mõõtmiste ja laste kognitiivse kontrolli vahel. Ühes uuringus oli selle võime areng positiivses korrelatsioonis prefrontaalsete-parietaalsete kiududega (Nagy, Westerberg ja Klingberg, 2004) on kooskõlas funktsionaalsete neuroimaging uuringutega, mis näitavad nende piirkondade erinevat värbamist lastel võrreldes täiskasvanutega.
Kasutades sarnast lähenemist, Liston et al. (2005) on näidanud, et valgeaine traktid prefrontaal-basaalganglionide ja tagumiste kiudude vahel arenevad edasi kogu lapsepõlves kuni täiskasvanueas, kuid ainult need traktid prefrontaalse ajukoore ja basaalganglioni vahel on korrelatsioonis impulsi kontrolliga, mida mõõdetakse jõudluse järgi liikumisel/nogo. ülesanne. Prefrontaalsed kiudtraktid määrati sama ülesande abil fMRI uuringus tuvastatud huvipiirkondade järgi. Mõlemas DTI arenguuuringus olid kiudtrakti mõõdud korrelatsioonis arenguga, kuid teatud kiudude spetsiifilisust kognitiivse jõudlusega näitas konkreetse trakti eraldamine (Liston et al., 2005) või kognitiivsed võimed (Nagy jt, 2004). Need leiud rõhutavad mitte ainult piirkondlike, vaid ka vooluahelaga seotud muutuste uurimise tähtsust, kui esitatakse väiteid vanusest sõltuvate muutuste kohta kognitiivse arengu närvisubstraatides.
Käitumise ja aju arengu funktsionaalsed MRI uuringud
Kuigi MRI ja DTI abil mõõdetud struktuurimuutusi on seostatud käitumismuutustega arengu käigus, on struktuuri ja funktsiooni seose uurimisel otsesem lähenemine aju ja käitumise muutuste samaaegne mõõtmine, nagu fMRI puhul. Võimalus mõõta MRI abil areneva aju funktsionaalseid muutusi omab arenguteaduse valdkonnas märkimisväärset potentsiaali. Käesoleva dokumendi kontekstis pakub fMRI vahendit noorukite käitumise tõlgenduste piiramiseks. Nagu varem öeldud, arvatakse, et prefrontaalse ajukoore areng mängib olulist rolli kõrgemate kognitiivsete võimete, nagu otsustusvõime ja kognitiivne kontroll, küpsemisel.Casey, Tottenham ja Fossella 2002b; Casey, Trainor jt, 1997). Nende võimete neurobioloogilise aluse hindamiseks on kasutatud paljusid paradigmasid koos fMRI-ga, sealhulgas flanker, Stroop ja go/nogo ülesandeid (Casey, Castellanos jt, 1997; Casey, Giedd ja Thomas, 2000a; Durston et al., 2003). Kokkuvõttes näitavad need uuringud, et lapsed värbavad nende ülesannete täitmisel eristatavaid, kuid sageli suuremaid, hajusamaid prefrontaalseid piirkondi kui täiskasvanud. Aktiivsusmuster aju piirkondades, mis on ülesannete täitmisel kesksel kohal (st mis korreleeruvad kognitiivse jõudlusega), muutuvad vanusega fokuseeritumaks või täpsemaks, samas kui piirkondades, mis ei ole ülesande täitmisega korrelatsioonis, aktiivsus väheneb koos vanusega. Seda mustrit on täheldatud mõlema ristlõike (Brown et al., 2005) ja pikisuunalised uuringud (\ tDurston et al., 2006) ja mitmesuguste paradigmade vahel. Kuigi neuropiltimise uuringud ei suuda selliste arengumuutuste (nt dendriitne arboriseerumine, sünaptiline pügamine) mehhanismi lõplikult iseloomustada, kajastavad leiud küpsemisega aktiveeritud ajupiirkondade arengut ja projektsioonide täpsustamist ning viitavad sellele, et need muutused toimuvad pikema aja jooksul. ajaperiood (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya ja Gabrieli, 2002; Casey, Trainor jt, 1997; Casey et al., 2002a; Crone, Donohue, Honomichl, Wendelken ja Bunge, 2006; Luna et al., 2001; Moses et al., 2002; Schlaggar jt, 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro ja Eden, 2003).
Kuidas saab see metoodika meid teavitada sellest, kas noorukitel puudub tõepoolest piisav kognitiivne kontroll (impulsiivne) või kas nad on oma valikutes ja tegudes riskantsed? Kognitiivsete kontrolliülesannetega (nt go/nogo) mõõdetud impulsside kontroll näitab lineaarset arengumustrit kogu lapsepõlves ja noorukieas, nagu eespool kirjeldatud. Hiljutised neuropiltimise uuringud on aga hakanud uurima tasuga seotud töötlemist, mis on spetsiifiline noorukite riskivõtmisele (Bjork et al., 2004; Ernst et al., 2005; May et al., 2004). Need uuringud on keskendunud peamiselt accumbensi piirkonnale, mis on osa basaalganglionidest, mis on seotud tasu ennustamisega, selle asemel, et iseloomustada selle piirkonna arengut koos ülalt-alla kontrollpiirkondadega (prefrontaalne ajukoor). Kuigi hiljutine aruanne on näidanud, et noorukite ventraalne prefrontaalne aktiivsus on väiksem kui täiskasvanutel rahaliste otsuste tegemise ajal riskikäitumise kohta.Eshel, Nelson, Blair, Pine ja Ernst, 2007).
Üldiselt on vähesed uuringud uurinud, kuidas subkortikaalsetes piirkondades (nt accumbens) muutub tasustamisskeemide areng koos ajukoore prefrontaalsete piirkondade arenguga. Veelgi enam, kuidas need närvimuutused langevad kokku tasu otsimise, impulsiivsuse ja riskivõtliku käitumisega, on suhteliselt teadmata. Meie neurobioloogiline mudel viitab sellele, et kõrgendatud reageerimisvõime hüvedele ja ebaküpsus käitumiskontrolli piirkondades võib kallutada noorukeid otsima kohest, mitte pikaajalist kasu, mis võib seletada nende riskantsete otsuste tegemise ja impulsiivse käitumise suurenemist. Otsuste tegemise subkortikaalse (nt accumbens) ja kortikaalse (nt prefrontaalne) arengu jälgimine lapsepõlvest kuni täiskasvanueani annab täiendavaid piiranguid selle kohta, kas noorukieas teatatud muutused on spetsiifilised sellele arenguperioodile või peegeldavad küpsemist, mis toimub pidevalt mõnevõrra lineaarne muster lapsepõlvest täiskasvanueani.
Hiljutise fMRI uuringu empiirilised tõendid aitavad toetada meie neurobioloogilist mudelit ja kasutavad noorukiea mõistmisel üleminekuviisi, uurides noorukieale eelnevaid ja järgnevaid muutusi. Selles uuringus (Galvan et al., 2006), uurisime käitumuslikke ja neuraalseid reaktsioone tasustamise manipulatsioonidele kogu arengus, keskendudes ajupiirkondadele, mis on seotud tasuga seotud õppimise ja loomade käitumisega (Hikosaka ja Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge ja Zhuang, 2003; Schultz, 2006) ja täiskasvanute pildistamise uuringud (nt Knutson, Adams, Fong ja Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) ja sõltuvusuuringutes (Hyman & Malenka, 2001; Volkow & Li, 2004). Tuginedes näriliste mudelitele (Laviola et al., 1999; Spear, 2000) ja eelmine pilditöö (Ernst et al., 2005), oletasime, et võrreldes laste ja täiskasvanutega ilmneb noorukitel liialdatud accumbens aktivatsioon, mis on kooskõlas ülalt-alla prefrontaalsete kontrollipiirkondade vähem küpsega. Hiljutine töö, mis näitab hilinenud funktsionaalset ühenduvust nende prefrontaalsete ja limbiliste subkortikaalsete piirkondade vahel noorukieas võrreldes täiskasvanutega, pakub mehhanismi nende piirkondade ülalt-alla kontrolli puudumiseks (Hare jt, 2007a).
Meie tulemused olid kooskõlas näriliste mudelitega (Laviola, Macri, Morley-Fletcher ja Adriani, 2003) ja varasemad pildiuuringud (\ tErnst et al., 2005), mis viitab koormuse suurendamisele noorukieas. Tõepoolest, võrreldes laste ja täiskasvanutega, näitasid noorukid tasu ootuses liialdatud accumbens-vastust. Kuid nii lapsed kui ka noorukid näitasid prefrontaalsetes kontrollipiirkondades vähem küpset vastust kui täiskasvanutel. Need leiud viitavad sellele, et nende piirkondade erinevad arengutrajektoorid võivad olla aluseks kuhjumisaktiivsuse suurenemisele, võrreldes laste või täiskasvanutega, mis võib omakorda olla seotud sel arenguperioodil täheldatud suurenenud impulsiivse ja riskantse käitumisega (vt. Joon. 4).

Mitmete arenguga seotud fMRI uuringute käigus on teatatud prefrontaalsete ja subkortikaalsete piirkondade erinevast värbamisest.Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Tavaliselt on neid tulemusi tõlgendatud ebaküpsete prefrontaalsete piirkondade mõttes, mitte ebaühtlaselt prefrontaalse ja subkortikaalse piirkondliku arengu vahel. Arvestades prefrontaalsete piirkondade tõendeid asjakohaste meetmete juhtimisel erinevates kontekstides (Miller & Cohen, 2001) ebaküpsed prefrontaalsed tegevused võivad takistada tulevaste tulemuste asjakohast hindamist ja riskantsete valikute hindamist ning võivad seega olla vähem mõjutavad tasu hindamist kui accumbens. See muster on kooskõlas eelnevate uuringutega, mis näitavad kõrgemat subkortikaalset, võrreldes kortikaalset aktiivsust, kui otsused on kallutatud otsese pikaajalise kasu tõttu (McClure, Laibson, Loewenstein ja Cohen, 2004). Lisaks on fMRI abil näidatud, et accumbensi aktiivsus on positiivses korrelatsioonis hilisema riskikäitumisega (Kuhnen & Knutson, 2005). Noorukieas, võrreldes lapsepõlve või täiskasvanueaga, ei pruugi ebaküps ventraalne prefrontaalne ajukoor tagada piisavalt ülalt-alla kontrolli tugevalt aktiveeritud tasu töötlemise piirkondade (nt accumbens) üle, mille tulemuseks on prefrontaalsete süsteemide (orbitofrontaalne ajukoor) väiksem mõju preemia saamisel akkumbenidele. hindamine.
Miks peaks aju programmeeritud nii arenema?
Noorukiea on üleminekuperiood lapsepõlve ja täiskasvanuea vahel, mis sageli esineb puberteedieas. Puberteet tähistab seksuaalse küpsemise algust (Graber & Brooks-Gunn, 1998) ja neid saab määratleda bioloogiliste markerite abil. Noorukust saab kirjeldada kui progressiivset üleminekut täiskasvanueas koos ebakindla ontogeneetilise ajaga (Spear, 2000). Evolutsiooniliselt öeldes on noorukieas periood, mille jooksul omandatakse iseseisvusoskused, et suurendada edu perekonna kaitsest lahkumisel, suurendades samas kahjulike asjaolude (nt vigastused, depressioon, ärevus, uimastite tarbimine ja sõltuvusKelley, Schochet ja Landry, 2004). Iseseisvust taotlev käitumine on liikide lõikes levinud, näiteks eakaaslastele suunatud sotsiaalse suhtluse suurenemine ning uudsuse otsimise ja riskide võtmise käitumise intensiivistumine. Psühhosotsiaalsed tegurid mõjutavad noorukite kalduvust riskikäitumisele. Riskantne käitumine on aga bioloogiliselt ajendatud tasakaalustamatuse tulemus suurenenud uudsuse ja sensatsiooniotsingute vahel koos ebaküpse "iseregulatsiooni pädevusega" (Steinberg, 2004). Meie neurobioloogilised andmed viitavad sellele, et see toimub nende kahe süsteemi (limbilise ja kontrolli) erineva arengu kaudu.
Spekulatsioonid viitavad sellele, et see arengumuster on evolutsiooniline tunnus. Peate tegelema kõrge riskiga käitumisega, et lahkuda oma perest ja külast, et leida kaaslane ja võtta riske samal ajal, kui hormoonid sunnivad noorukeid seksuaalpartnereid otsima. Tänapäeva ühiskonnas, kus noorukieas võib kesta lõputult, kui lapsed elavad koos vanematega ja neil on rahaline sõltuvus ning nad valivad hilisemas elus kaaslast, võib seda arengut pidada sobimatuks.
Liikide lõikes on tõendeid kõrgendatud uudsuse otsimise ja riskide võtmise kohta noorukieas. Samaealiste eakaaslaste otsimist ja võitlust vanematega, mis kõik aitavad noorukit paaritumiseks koduterritooriumilt eemale viia, on näha ka teiste liikide, sealhulgas näriliste, ahviliste ja mõnede lindude puhul (Spear, 2000). Võrreldes täiskasvanutega, ilmutavad noorukieas rotid vaba valiku uudsuse paradigmas rohkem uudsust otsivat käitumist (Laviola et al., 1999). Neurokeemilised tõendid näitavad, et noorukite aju tasakaal kortikaalsete ja subkortikaalsete dopamiinisüsteemide vahel hakkab noorukieas nihkuma suuremate kortikaalsete dopamiinitasemete poole.Spear, 2000). Sarnast pikaleveninud dopamiinergilist närvilisust noorukieas kuni täiskasvanueas on näidatud ka primaadi prefrontaalses ajukoores (Rosenberg & Lewis, 1995). Seega näib see kõrgendatud näiline riskide võtmine olevat liikidevaheline ja sellel on oluline kohanemisvõime.
Bioloogiline eelsoodumus, areng ja risk
Psühholoogias on juba mõnda aega tunnustatud individuaalseid erinevusi impulsside kontrollis ja riskide võtmises (Benthin, slovaki keel ja Severson, 1993). Võib-olla on üks klassikalisi näiteid individuaalsetest erinevustest nende võimete osas sotsiaal-, kognitiivses ja arengupsühholoogiaalases kirjanduses on rahulduse viivitus (Mischel, Shoda ja Rodriguez, 1989). Rahulolu viivitusi hinnatakse tavaliselt 3–4-aastastel väikelastel. Väikelapselt küsitakse, kas ta eelistaks väikest preemiat (üks küpsis) või suurt preemiat (kaks küpsist). Seejärel öeldakse lapsele, et katsetaja lahkub ruumist, et valmistuda eelseisvateks tegevusteks, ja selgitatakse lapsele, et kui ta jääb oma kohale ega söö küpsist, saab ta suure preemia. Kui laps ei oota või ei saa oodata, peaks ta helistama kella, et kutsuda eksperimenteerija ja saada seeläbi väiksem tasu. Kui on selge, et laps saab ülesandest aru, istub ta kahe auhinna ja kellaga laua taha. Tähelepanu segavad tegurid ruumis on minimeeritud, mänguasju, raamatuid ega pilte pole. Eksperimenteerija naaseb 15 minuti pärast või pärast seda, kui laps on kella helistanud, preemiad ära söönud või ärevuse märke näidanud. Mischel näitas, et lapsed käituvad tavaliselt kahel viisil: (1) nad helistavad peaaegu kohe kella, et küpsist saada, mis tähendab, et nad saavad ainult ühe; või (2) nad ootavad ja optimeerivad oma kasumit ning saavad mõlemad küpsised. See tähelepanek viitab sellele, et mõned inimesed on teistest paremad oma võimes kontrollida impulsse väga silmapaistvate stiimulite taustal ja seda eelarvamust võib tuvastada juba varases lapsepõlves (Mischel jt, 1989) ja need näivad püsivat kogu noorukieas ja noores täiskasvanueas (Eigsti jt, 2006).
Mis võiks seletada individuaalseid erinevusi optimaalses otsustusprotsessis ja käitumises? Mõned teoreetikud on oletanud, et dopamiinergiline mesolimbiline skeem, mis on seotud tasu töötlemisega, on riskantse käitumise aluseks. Individuaalsed erinevused selles skeemis, näiteks dopamiiniga seotud geenide alleelsed variandid, mille tulemuseks on liiga vähe või liiga palju dopamiini subkortikaalsetes piirkondades, võivad olla seotud kalduvusega käituda riskantses käitumises (O'Doherty, 2004). On näidatud, et nucleus accumbens aktiivsus suureneb vahetult enne riskantsete valikute tegemist rahalise riski paradigma osas (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002) ja nagu eelnevalt kirjeldatud, näitavad noorukid laste või täiskasvanutega võrreldes ülepaisutatud aktiivsust, mis on rahuldust pakkuv tulemus (Ernst et al., 2005; Galvan et al., 2006). Need andmed näitavad kollektiivselt, et noorukid võivad olla grupis riskantsemate valikute suhtes kalduvamad.Aednik ja Steinberg, 2005), kuid mõned noorukid on teistest altimad riskantsele käitumisele, mis seab nad potentsiaalselt suuremasse negatiivsete tulemuste riski. Seetõttu on arengupopulatsioonide riskide võtmise ja tasu töötlemisega seotud keeruliste aju-käitumise suhete uurimisel oluline arvestada individuaalset varieeruvust.
Et uurida individuaalseid erinevusi riskikäitumises, Galvan et al. (2007) uuris hiljuti seost tasuga seotud närvisüsteemi aktiivsuse vahel, oodates suurt rahalist tasu, isiksuseomaduste, riskide võtmise ja impulsiivsuse mõõtmise vahel noorukieas. Funktsionaalne magnetresonantstomograafia ja anonüümsed enesehinnanguskaalad riskantse käitumise, riskitaju ja impulsiivsuse kohta saadi inimestelt vanuses 7–29 aastat. Accumbensi aktiivsuse ja riskantse käitumise tõenäosuse vahel oli positiivne seos kogu arengu jooksul. See tegevus varieerus sõltuvalt üksikisikute hinnangutest sellise käitumise eeldatavate positiivsete või negatiivsete tagajärgede kohta. Inimesed, kes tajusid riskantset käitumist kohutavate tagajärgedeni viivana, aktiveerisid kõhulihaseid vähem, et premeerida. Seda seost ajendasid suuresti lapsed, kusjuures täiskasvanud hindasid sellise käitumise tagajärgi võimalikuks. Impulsiivsushinnanguid ei seostatud accumbensi aktiivsusega, vaid pigem vanusega. Need leiud viitavad sellele, et noorukieas võivad mõned inimesed olla altimad riskikäitumisele, mis on tingitud arengumuutustest, mis on kooskõlas indiviidi riskantse käitumise eelsoodumuse varieeruvusega, mitte lihtsalt impulsiivsuse muutustega (vt. Joon. 5).

Noorukite käitumist on korduvalt iseloomustatud kui impulsiivset ja riskantset (Steinberg, 2004, 2007), kuid see pildistamise kirjanduse ülevaade viitab nende käitumise erinevatele neurobioloogilistele substraatidele ja erinevatele arengutrajektoore. Täpsemalt on impulsiivsus seotud ebaküpse ventraalse prefrontaalse arenguga ja väheneb järk-järgult lapsepõlvest täiskasvanueani (Casey, Galvan jt, 2005). Negatiivne korrelatsioon impulsiivsuse hinnangute ja vanuse vahel uuringus Galvan et al. (2007) toetab seda seisukohta veelgi. Seevastu riskide võtmine on seotud kuhjumise aktiivsuse suurenemisega (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002), mis on noorukite puhul laste ja täiskasvanutega võrreldes liialdatud (Ernst et al., 2005; Galvan et al., 2006). Seega ei saa noorukite valikuid ja käitumist seletada ainult prefrontaalse ajukoore impulsiivsuse või pikaleveninud arenguga, kuna sel juhul eeldatakse, et lapsed on suuremad riskivõtjad. Leiud annavad neuraalse aluse selle kohta, miks mõned noorukid on teistest suuremas ohus, kuid annavad aluse ka sellele, kuidas noorukite käitumine erineb riski võtmisel lastest ja täiskasvanutest.
Need andmed viitavad ühiselt sellele, et kuigi noorukeid kui rühma peetakse riskivõtjateks (Aednik ja Steinberg, 2005), on mõned noorukid teistest altimad riskantsele käitumisele, mis seab nad potentsiaalselt suuremasse negatiivsete tulemuste riski. Need leiud rõhutavad individuaalse varieeruvuse arvestamise tähtsust, kui uuritakse keerulisi aju-käitumise suhteid, mis on seotud riskide võtmise ja tasu töötlemisega arengupopulatsioonides. Lisaks võivad need individuaalsed ja arengulised erinevused aidata selgitada mõnede inimeste haavatavust ainete kasutamisega seotud riskide võtmise ja lõpuks ka sõltuvuse suhtes.
Järeldused
Inimese pildiuuringud näitavad struktuurseid ja funktsionaalseid muutusi frontostriataalsetes piirkondades (Giedd et al., 1996, 1999; Jernigan jt, 1991; Sowell et al., 1999; ülevaatamiseks, Casey, Galvan jt, 2005), mis näivad paralleelselt kognitiivse kontrolli ja eneseregulatsiooni suurenemisega (Casey, Trainor jt, 1997; Luna & Sweeney, 2004; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004; Vaata ka Steinberg, 2008, see number). Näib, et need muutused näitavad aja jooksul nihet prefrontaalsete piirkondade aktivatsioonis hajusalt fokaalsemale värbamisele (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor jt, 1997; Durston et al., 2006; Moses et al., 2002) ja subkortikaalsete piirkondade kõrgemat värbamist noorukieas (Casey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Kuigi neuroimaging uuringud ei suuda selliste arengumuutuste mehhanismi lõplikult iseloomustada, võivad need mahu ja struktuuri muutused kajastada arengut nendesse ajupiirkondadesse ja nendest ajupiirkondadest küpsemise ajal ja nende projektsioonide täpsustamist, mis viitab süsteemi peenhäälestamisele koos arenguga.
Kokkuvõttes näitavad siin sünteesitud leiud, et suurenenud riskikäitumine noorukieas on seotud subkortikaalse naudingu ja kortikaalsete kontrollipiirkondade erinevate arengutrajektooridega. Neid arengumuutusi võivad süvendada individuaalsed erinevused tasustamissüsteemide aktiivsuses. Kuigi noorukiea on eristatud perioodina, mida iseloomustavad tasu otsimine ja riskide võtmine (Aednik ja Steinberg, 2005; Spear, 2000) individuaalsed erinevused neuraalsetes reaktsioonides premeerimisel soodustavad mõningaid noorukeid rohkem riske võtma kui teised, mis seab nad suuremasse negatiivsete tulemuste riski. Need leiud annavad olulise aluse, sünteesides erinevaid leide, mis on seotud riskivõtliku käitumisega noorukieas ning mõistmaks individuaalseid erinevusi ja arengumarkereid, mis näitavad kalduvust negatiivsele käitumisele.
Tunnustused
Seda tööd toetasid osaliselt riikliku uimastite kuritarvitamise instituudi R01 DA18879 ja riikliku vaimse tervise instituudi 1P50 MH62196 stipendiumid.
viited
- Benthin A, Slovic P, Severson H. Psühhomeetriline uuring noorukite riski tajumisest. Noorukuse ajakiri. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Stimuleerunud aju aktiveerimine noorukitel: sarnasused ja erinevused noortest täiskasvanutest. Journal of Neuroscience. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Kontseptuaalsete teadmiste roll organisatsiooni arengus laste mälus. In: Brainerd CJ, Pressley M, toimetajad. Põhilised protsessid mälu arendamisel: edusammud kognitiivse arengu uuringutes. New York: Springer-Verlag; 1985. lk. 103 – 142.
- Bjorkland DF. Kuidas vanuselised muutused teadmistebaasis aitavad kaasa laste mälu arengule: tõlgendusülevaade. Arenguülevaade. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenesis reesusahvide prefrontaalses ajukoores. Ajukoor. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Mälu sõltumatus ja mälu häirimine kognitiivses arengus. Psühholoogiline ülevaade. 1993;100: 42-67. [PubMed]
- Pruun TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Inimese aju funktsionaalse organisatsiooni arengu muutused sõna genereerimiseks. Ajukoor. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Ebaküpse otsmikusagara panus laste kognitiivsesse kontrolli: tõendid fMRI-st. Neuron. 2002;33: 301-311. [PubMed]
- Juhtum R. Neo-Piageti võimsuskonstruktsiooni valideerimine. Eksperimentaalse laste psühholoogia Teataja 1972;14: 287-302.
- Juhtum R. Intellektuaalne areng: sünnist täiskasvanueani. New York: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Parema frontostriataalse vooluringi mõju vastuse pärssimisele ja tähelepanupuudulikkuse / hüperaktiivsuse häirele. Ameerika lapse ja noorukite psühhiaatria akadeemia ajakiri. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Muutused aju funktsionaalses organisatsioonis kognitiivse arengu ajal. Praegune arvamus neurobioloogias. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Struktuurne ja funktsionaalne aju areng ja selle seos kognitiivse arenguga. Bioloogiline psühholoogia. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Striatoorse ja hipokampuse funktsiooni hajutamine stimuleeriva reaktsiooni ühilduvuse ülesandega. Journal of Neuroscience. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Reageerimiskonflikti, tähelepanu valiku ja funktsionaalse magnetresonantstomograafia ootuse jagunemine. Riikliku Teaduste Akadeemia menetlus. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Kliinilised, pildistamise, kahjustuste ja geneetilised lähenemisviisid kognitiivse kontrolli mudeli suunas. Arengupsühobioloogia. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Arenevate aju kujutamine: Mida oleme õppinud kognitiivse arengu kohta? Kognitiivate teadusuuringute suundumused. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN jt. Arengufunktsiooniga funktsionaalne MRI-uuring prefrontaalse aktiveerimise kohta go-no-go ülesande täitmise ajal. Kognitiivse neuroteaduse ajakiri. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Aju piirkonnad, mis vahendavad paindliku reegli kasutamist arengu käigus. Journal of Neuroscience. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Vastupidavus häiretele: arenduslikud muutused põhitöötlusmehhanismis. In: Howe ML, Pasnak R, toimetajad. Kognitiivse arengu esilekerkivad teemad 1. köide: alused. New York: Springer; 1993. lk 3–27.
- Teemant A. Tagasivõtmise võime arendamine, et juhtida tegevust, nagu näitab imikute AB-i jõudlus. Lapse areng. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Konfliktide ja reageerimisvõistluse parameetriline manipuleerimine, kasutades kiiret segatüüpi sündmustega seotud fMRI-d. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J jt. Üleminek hajusalt kortikaalsele aktiivsusele koos arenguga. Arenguteadus. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Areneva inimese aju anatoomiline MRI: mida me oleme õppinud? American Academy of Child Adolescent Psychiatry ajakiri. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R jt. Noorte riskikäitumise jälgimine – Ameerika Ühendriigid, 2005, järelevalve kokkuvõtted. Haigus- ja surmajuhtumite aruanne. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB jt. Kognitiivse kontrolli prognoosimine koolieelsest kuni hilisema noorukini ja noorukini. Psühholoogiline teadus. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala ja tuumad akumuleeruvad vastusena täiskasvanute ja noorukite kasu saamisele ja mittekasutamisele. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Täiskasvanutel ja noorukitel valitud valikuvõimalused: Ventrolateraalse prefrontaalsete ja anterioride cingulaatide väljaarendamine. Neuropsychologia. 2007;45: 1270-1279. [PMC tasuta artikkel] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Mäluülesande spontaanne verbaalne harjutus vanuse funktsioonina. Lapse areng. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Akumeenide varasemad arengud orbitofrontaalse ajukoorme suhtes võivad olla noorukite riskikäitumise aluseks. Journal of Neuroscience. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Riskivõtmine ja noorukiea: Kes on ohus? Arenguteadus. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Eakaaslaste mõju riskide võtmisele, riskieelistustele ja riskantsete otsuste langetamisele noorukieas ja täiskasvanueas: eksperimentaalne uuring. Arengupsühholoogia. 2005;41: 625-635. [PubMed]
- Giedd JN. Noorte aju struktuurne magnetresonantstomograafia. New Yorgi Teaduste Akadeemia Annals. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A jt. Aju areng lapsepõlves ja noorukieas: pikisuunaline MRI uuring. Looduse neuroteadus. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL jt. Inimese aju arengu kvantitatiivne magnetresonantstomograafia: vanuses 4–18. Ajukoor. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Inimese kortikaalse arengu dünaamiline kaardistamine lapsepõlves varases täiskasvanueas. Ameerika Ühendriikide Riikliku Teaduste Akadeemia toimingud. 2004;101: 8174-8179. [PMC tasuta artikkel] [PubMed]
- Graber JA, Brooks-Gunn J. Pubert. In: Blechman EA, Brownell KD, toimetajad. Käitumismeditsiin ja naised põhjalik käsiraamat. New York, NY: Guilford Press; 1998. lk 51–58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Nooruki aju ja potentsiaalne oht ärevuse ja depressiooni tekkeks. 2007 Esitatud avaldamiseks.
- Hare TA, Voss HU, Glover GH, Casey BJ. Prefrontaalsete ja subkortikaalsete limbiliste süsteemide vaheline konkurents on noorukieas emotsionaalse reaktiivsuse aluseks. 2007b Esitatud avaldamiseks.
- Harnishfeger KK, Bjorkland F. Inhibeerimismehhanismide ontogenees: uuendatud lähenemine kognitiivsele arengule. In: Howe ML, Pasnek R, toimetajad. Kognitiivse arengu teemad. Vol. 1. New York: Springer-Verlag; 1993. lk 28–49.
- Hikosaka K, Watanabe M. Monkey orbitaalsete ja lateraalsete eellaste hilisem aktiivsus varieerub erinevate hüvedega. Ajukoor. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Sünaptiline tihedus inimese eesmise ajukoores - arengu muutused ja vananemise mõjud. Ajuuuringud. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Sõltuvus ja aju: sundi neurobioloogia ja selle püsivus. Loodus Arvustused Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Magnetresonantstomograafilised kõrvalekalded skisofreenia lentikulaarsetes tuumades ja ajukoores. Psühhiaatriaarhiiv. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Vaimse võimekuse kognitiivse töötlemise komponentide individuaalsed ja arenguhäired. Lapse areng. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. New Yorgi Teaduste Akadeemia Annals. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Rahalise tasu suurenemise prognoosimine värbab selektiivselt tuuma accumbens'i. Journal of Neuroscience. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Finantsriskide võtmise närvisüsteem. Neuron. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psühobioloogilised riskifaktorid psühhostimulantide haavatavuse suhtes noorukitel ja loommudelitel. Neuroteadus ja bioloogiline käitumine 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstraktne riskikäitumine noorukitel hiirtel: psühhobioloogilised determinandid ja varane epigeneetiline mõju. Neuroteadus ja bioloogiline käitumine 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM jt. Frontostriaalne mikrostruktuur moduleerib kognitiivse kontrolli tõhusat värbamist. Ajukoor. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Koostööga seotud ajufunktsiooni ilmnemine: FMRI uuringud vastuse inhibeerimise arengu kohta. New Yorgi Teaduste Akadeemia Annals. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ jt. Laialt levinud ajufunktsiooni küpsemine allub kognitiivsele arengule. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC et al. Nucleus accumbens selektiivne aktiveerimine riskiotsuste tegemisel. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Mai JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA jt. Sündmusega seotud funktsionaalne magnetresonantstestimine tasulise aju ahelaga lastel ja noorukitel. Bioloogiline psühhiaatria. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Eraldi närvisüsteemid hindavad kohest rahalist tasu. Teadus. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Prefrontaalse koore funktsiooni integreeriv teooria. Neuroteaduste aastaaruanne. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Laste rahuldamise hilinemine. Teadus. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E jt. Noorukite ebaküpsus tähelepanuga seotud aju kaasamisel emotsionaalsetele näoilmetele. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neurumajandus ja hindamise bioloogilised alused. Neuron. 2002;36: 265-284. [PubMed]
- Mooses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Laste globaalse ja kohaliku töötlemise funktsionaalne MRI. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Nüüd kõik koos: kui teadmise ja tegevuse vahelised dissotsiatsioonid kaovad. Psühholoogiateadus. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Valge aine küpsemine on seotud kognitiivsete funktsioonide arenguga lapsepõlves. Kognitiivse neuroteaduse ajakiri. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstraktsed auhinnad ja karistused inimese orbitofrontaalses ajukoores. Nature Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Preemiate esitused ja tasuga seotud õpe inimese ajus: ülevaated neuropildist. Praegused arvamused neurobioloogias. 2004;14: 769-776.
- Pascual-Leone JA. Matemaatiline mudel üleminekuks Piageti arenguetappides. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hüperdopamiinergilistel hiirtel on magusate hüvede jaoks kõrgem "sooviv", kuid mitte "meeldiv". Journal of Neuroscience. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Risk ja ratsionaalsus teismeliste otsuste tegemisel: mõju teooriale, praktikale ja avalikule korrale. Psühholoogiline teadus avalikes huvides. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Monkey prefrontaalsete ja motoorse koorikute dopamiinergilise inervatsiooni postnataalne küpsemine: türosiinhüdroksülaasi immunohistokeemiline analüüs. Võrdleva neuroloogia ajakiri. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A jt. Funktsionaalne frontaliseerimine vanusega: neuroderengu trajektooride kaardistamine fMRI-ga. Neuroteadus ja bioloogiline käitumine 2000;24: 13-19. [PubMed]
- Schlaggar BL, pruun TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funktsionaalsed neuroanatoomilised erinevused täiskasvanute ja kooliealiste laste vahel üksikute sõnade töötlemisel. Teadus. 2002;296: 1476-1479. [PubMed]
- Schultz W. Käitumise teooriad ja tasu neurofüsioloogia. Psühholoogia iga-aastased ülevaated. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Welcome SE, Henkenius AL, Toga AW. Kortikaalse muutuse kaardistamine kogu inimese eluea jooksul. Looduse neuroteadus. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo tõendid noorukite ajuküpsemise kohta ees- ja striatali piirkondades. Looduse neuroteadus. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Inimese ajukoore muutuste kaardistamine kogu eluea jooksul. Neuroteadlane. 2004;10: 372-392. [PubMed]
- Spear LP. Noorte aju ja vanusega seotud käitumuslikud ilmingud. Neuroteadus ja bioloogiline käitumine 2000;24: 417-463. [PubMed]
- Steinberg L. Riskivõtmine noorukieas: millised muutused ja miks? New Yorgi Teaduste Akadeemia Annals. 2004;1021: 51-58. [PubMed]
- Steinberg L. Riskide võtmine noorukieas: aju ja käitumisteaduse uued perspektiivid. Psühholoogiateaduse praegused juhised. 2007;16: 55-59.
- Steinberg L. Sotsiaalne neuroteaduse perspektiiv noorukite riskide võtmisel. Arenguülevaade. 2008;28: 78-106. [PMC tasuta artikkel] [PubMed]
- Tamm L, Menon V, Reiss AL. Vastuse inhibeerimisega seotud ajufunktsiooni küpsemine. Ameerika lapse ja noorukite psühhiaatria akadeemia ajakiri. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y jt. Tõendid arenguerinevuste kohta kaudse järjestusõppes: laste ja täiskasvanute FMRI uuring. Kognitiivse neuroteaduse ajakiri. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Lilled DL, Zeffiro TA, Eden GF. Neuraalsete mehhanismide väljatöötamine lugemiseks. Looduse neuroteadus. 2003;6: 767-773.
- Volkow ND, Li TK. Narkomaania: käitumise neurobioloogia on viltu läinud. Loodus Arvustused Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Emotsionaalsed ja kognitiivsed muutused noorukieas. Praegune arvamus neurobioloogias. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Dopamiini ülekanne inimese striatumis rahalise tasuülesannete täitmisel. Journal of Neuroscience. 2004;24: 4105-4112. [PubMed]