Esimene Behav Neurosci. 2015 mai 19, 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Abstraktne
Klassikalises konditsioneerimises seostub esialgu neutraalne stiimul (konditsioneeritud stiimul, CS) bioloogiliselt olulise sündmusega (tingimusteta stiimul, USA), mis võib olla valu (vastumeelne konditsioneerimine) või toit (söögiisu tingimine). Mõne assotsiatsiooni järel on CS võimeline algatama vastavalt kas kaitsvaid või lõpetavaid reaktsioone. Vastupidiselt aversiivsele tingimusele uuritakse inimestel söögiisu tingimist harva, kuigi selle tähtsus normaalse ja patoloogilise käitumise (nt rasvumine, sõltuvus) jaoks on vaieldamatu. Käesoleva uuringu eesmärk on tõlgendada loomade avastusi söögiisu konditsioneerimise kohta inimestele, kes kasutavad USA-s toitu. 8 osalejat uuriti ajavahemikus 10–XNUMX ilma hommikusöögita, et olla kindel, et nad tunnevad end näljasena. Kahes omandamisfaasis ennustas üks geomeetriline kuju (avCS +) vastumeelset USA-d (valus elektrilöök), teine kuju (appCS +) ennustas isuäratavat USA-d (šokolaad või soolane kringel vastavalt osalejate eelistustele) ja kolmas kuju (CS– ) ei ennustanud USA-d. Kustutusfaasis esitati need kolm kuju koos uudse kujuga (UUS) uuesti ilma USA kättetoimetamiseta. Valentsi- ja erutusreitingud ning ehmatuse ja naha juhtivuse (SCR) vastused koguti õppeindeksitena. Leidsime edukad vastumeelsed ja isuäratavad tingimused. Ühest küljest hinnati avCS + negatiivsemaks ja erutavamaks kui CS– ning indutseeris ehmatuse potentseerimise ja tugevnenud SCR. Teiselt poolt hinnati appCS + positiivsemaks kui CS- ning indutseeris ehmatuse nõrgenemist ja suuremat SCR-i. Kokkuvõttes kinnitasime edukalt (näljaste) inimeste loomade avastamist, näidates isu õppimist ja normaalset vastumeelset õppimist.
Sissejuhatus
Ohu ja toidu ennustamine on iga organismi ellujäämise seisukohalt ülioluline. Klassikalises tingimuses (Pavlov, 1927), eelneb eelnevalt aversiivsele sündmusele, nagu kerge valulik elektrilöök (aversiivne tingimusteta stiimul, USA) või isuäratav sündmus, näiteks toidupellet (isu-US), mitu korda. Hiljem on see stiimul üksi (nüüd märgistatud CS, CS +) vastavalt võimeline tekitama vastavalt kas kaitsvaid või tarbivaid vastuseid. Endist assotsiatiivset õppimist nimetatakse aversiivseks konditsioneerimiseks, viimast nimetatakse isuäratavaks konditsioneerimiseks.
Kuigi isuäralike sündmuste prognoosimine on ellujäämise jaoks sama oluline kui aversiivsete sündmuste prognoosimine, on loomadel isuäratav konditsioneerimine märkimisväärselt vähem uuritud.Bouton ja Peck, 1989; Koch et al., 1996; McDannald et al., 2011, vaadake ülevaadet Martin-Soelch jt, 2007), samuti inimestel (Klucken et al., 2009, 2013; Austin ja Duka, 2010; Delgado et al., 2011; Levy ja Glimcher, 2011). See teadustöö puudumine võib olla tingitud isuäraliku paradigma keerukusest võrreldes aversiivse paradigmaga. Näiteks peab toit olema esmane tugevdaja, kui organism on näljane, et olla rahuldav (hiljutise ülevaatuse kohta vt Dickinson ja Balleine, 1994; Clark et al., 2012). Inimiuuringutes on see raskus ületatud rahaga (Austin ja Duka, 2010; Delgado et al., 2011; Levy ja Glimcher, 2011) või erootilised pildid (Klucken et al., 2009, 2013). Neuro-kujutise uuringud on siiski näidanud, et esmane (st suupisted või joogid) ja teisese (st raha) tugevdajad aktiveerivad mõningaid tavalisi aju piirkondi (nt striatum), kuid toovad kaasa ka erinevad aktiveerimismustrid (Delgado et al., 2011; Levy ja Glimcher, 2011).
Meie teadmiste kohaselt on ainult mõned inimeste konditsioneerimise uuringud uurinud esmaste isuäraste tugevdajate nagu lõhna mõju (Gottfried et al., 2002), vesi (Kumar et al., 2008) või toit (Prévost jt, 2012). Esimeses uuringus Gottfried et al. (2002) seotud neutraalsed näod (ebameeldiv, meeldiv või neutraalne lõhn). Huvitaval kombel leidsid nad suurema aktiveerumise orbitofrontaalses ajukoores (OFC) ja ventraalses striatumis vastusena isuäratavale CS + -le võrreldes aversive CS + -ga. Nad jõudsid järeldusele, et OFC töötleb lõhna väärtust ja osaleb afektiivse väärtuse ülekandmisel lõhnast (USA) visuaalsele (CS) süsteemile. (Ventro) striatali aktiveerimist on tõlgendatud nii, et see peegeldab isuäratavat CR-i põhjustatud isuärast CR-d. Teises uuringus Kumar et al. (2008) kutsusid oma osalejaid (terved kontrollid ja raskekujulise depressiooniga patsiendid) laborisse varahommikul ja palusid neil öösel alkoholi ära hoida, et tagada enne skaneerimist janu. Fraktali pildid olid CS ja 0.1 ml vett USA-s. Huvitaval kombel näitasid tervetel osalejatel (kuid mitte depressiooniga patsientidel) teiste aktivatsioonide puhul ventraalses striatumis suuremat aktivatsiooni söögiisu CS + suhtes, mis viitab sellele, et see stiimul töödeldi rahuldust pakkuvana. Kolmandas uuringus Prévost et al. (2012) esitas osalejate eelistuste järgi ka fraktaalpilte CS-dena ja kas magusaid või soolaseid suupisteid USA-s. CS-d esitleti 6 s ja viimase sekundi jooksul esitati lisaks toidupilt. Iga kord, kui toidupilt ilmus, pani eksperimentaator osalejatele kätte toidupala, kellel lubati suupisteid kohe tarbida. Kahjuks ei teatanud autorid aju aktiveerimisest isuäratavale CS + -le klassikalises konditsioneerimisfaasis; kuid nad täheldasid vähem südame aeglustumist premeeritud CS + -ga võrreldes autasustamata CS–-ga, mis viitab diferentsiaalsele konditsioneerimisele.
Uuringute puudumine, mis kasutab hämmastavat vastust kui isuäratava konditsioneerimise indeksit, on üllatav, eriti arvestades selle laialdast kasutamist aversiivsetes tingimustes. Vastureaktsioon on esivanem ja automaatne kaitsev reaktsioon ootamatute, ootamatute ja tugevate vastumeelsete sündmuste suhtes (Koch, 1999). Seda kaitsvat vastust vahendab suhteliselt lihtne neuronaalne rada, mis hõlmab cochlearjuurseid neuroneid, retikulaarse moodustumise (PnC) caudal pontine tuuma ja seljaaju motoneuroneid (Fendt ja Fanselow, 1999; Koch, 1999). Loomkatsed on näidanud, et hämmastav võimendamine sõltub amygdala poolt PnC-le tehtud prognoosidest (Fendt ja Fanselow, 1999; Koch, 1999), samal ajal kui hämmastav nõrgenemine sõltub terviklikust tuumast \ t Koch, 1999). Selline hämmastuse vastuse moduleerimine on kasulik kaudne mõõt esitatud esiplaanil olevate stiimulite valentsuse kohta. Seega näitab võimendamine negatiivset valentsust, samal ajal kui nõrgenemine näitab positiivset valentsust, ilma et kognitiivsed protsessid seda oluliselt mõjutaksid (Hamm ja Weike, 2005; Andreatta jt, 2010). Meie teadmiste kohaselt on ainult üks loomkatse uurinud isuäratavat konditsioneerimist, mõõtes üllatuslikke vastuseid sõltuvaks meetmeks. Huvitav on see, et uuritud rottidel oli sahharoosilahuse kohaletoimetamisega seotud CS + (st kerge) nõrgenemine. Nimelt oli selline nõrgenemine nõrgenenud nendel loomadel, kellel oli NAcc kahjustus, kuid mitte amygdala kahjustustega patsientidel, mis viitab sellele, et NAcc mängib erilist rolli isuäratavate CR-de esilekutsumisel ja hämmastavate reaktsioonide leevendamisel (Koch et al., 1996).
Praeguse uuringu põhjal saame kõigepealt tõlkida see isuäratav konditsioneerimise paradigma inimestele, kasutades esmaseid tugevdajaid nagu USA, st magus (šokolaad Smarties®) või soolane (väike soolane pretzel) toit ja ärritada modulatsiooni kui CR-i mõõdet. Me ootasime, et appCS + käivitab tugevad isuäratavad CR-d, mis peegelduvad hämmastava nõrgenemise, täiustatud SCR-i ja positiivse valentsiklassiga võrreldes teiste stiimulitega, st avCS + ja CS–.
Materjalid ja meetodid
Osalejad
5 vabatahtlikku nõustus uuringus osalema ja said kursuse ainepunktid. Üheksa osalejat jäeti analüüsist välja, kaks tehniliste probleemide tõttu, kolm seetõttu, et nad olid kodeeritud mittevastavatena (keskmine ehmatuse amplituud <2 μV) ja neli, kuna neil ei olnud tingimuse kohta piisavalt ehmatavat vastust (minimaalselt = 33; vt Materjalid ja meetod). Lõpuks arvestati analüüsis 16 osalejat (22.09 meest; keskmine vanus: XNUMX aastat, SD: 2.84; vahemik: 18 – 29 aastat). Neli osalejat ei olnud kohalikud sakslased, kuus olid vasakpoolsed. Üks osaleja jäi kogu katse vältel CS-USs ühendustest teadmatuks (vt protseduuri), kuid me otsustasime seda osalejat mitte välistada, sest tema vastused olid normaalsed ega mõjutanud tulemusi.
materjalid
Tingimusteta stiimulid (USA)
Kasutati kahte liiki USA-d. Meeldiva USA-na kasutasime osalejate mitte-domineerivale käsivarrele kergelt valulikku elektrilööki. Elektrilöök saadeti kahe elektroodiga, mille läbimõõt oli 9 mm ja vahedega 30 cm. Elektriline stiimul koosnes impulss-stiimulist, mille sagedus oli 50 Hz ja 200 ms kestus, mis tekkis praeguse stimulaatori poolt (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, UK, 400 V, maksimaalne 9.99 mA). Elektrilöögi intensiivsus määrati individuaalselt eelnevalt kirjeldatud künnismenetluse abil (Andreatta jt, 2010). Lühidalt, osalejad läbisid 0.5 mA kahe järjestikuse järjestikuse kasvava ja kahaneva intensiivsusega sarja. Nad pidid iga stiimulit visuaalselt mõõtma, ulatudes 0ist (üldse mitte tunne) kuni 10ini (tõesti tugev valu) koos 4iga künnise ankurina (just märgatav valu). Elektrilise stiimuli keskmine intensiivsus oli 2.12 mA (SD = 0.56) ja see oli valus (M = 6.45, SD = 1.73). USA isu oli kas šokolaad (Smarties®) või väike soolane kringel. Isuäratava USA valik sõltus osaleja individuaalsest eelistusest, nagu esialgse intervjuu käigus teatati. Nimelt pidid osalejad teatama, kas nad söövad hommikusöögi ajal tavaliselt soolast või magusat toitu. Samuti said nad katse enda ajal vabalt valida, kas eelistavad šokolaadi või soolast kringlit. Lõpuks valis 22 osalejat šokolaadi ja 11 väikese soolase kringli.
Konditsioneeritud stimulaatorid (CS)
CS-idena esitati geomeetrilised kujundid (sinine ruut, kollane ring, roheline kolmnurk, punane kuusnurk), mille diagonaal on 8 cm. Kujud esitati 8i jaoks musta arvuti ekraani keskel. Üks kuju (avCS +) oli alati seotud aversiivse USA-ga (valus elektrilöök), üks kuju (appCS +) oli alati seotud isuäratava USA-ga (šokolaad või soolane pretzel), ühte kuju (CS–) ei seostatud kunagi aversiivse või isuäratavad USA-d ning neljas kuju (UUS) esitati väljasuremisfaasis, kuid mitte omandamisetapi ajal, et tagada selle neutraalsus.
Põgenenud sond
Ehmatusproovina kasutati valget müra 103 dB kestusega 50 ms. Akustilisi stiimuleid esitati binauraalselt kõrvaklappide kohal ja need tekkisid juhuslikult 4–6 sekundit pärast kuju tekkimist.
Küsimustikud
Enne ja pärast eksperimenti pidid osavõtjad täitma riikliku iseloomuga ärevuse inventuuri saksa versioonid (STAI, Laux et al., 1981) ja positiivse negatiivse mõjuga ajakava (PANAS, Krohne et al., 1996). STAI on nimekiri osalejate omaduste ja / või seisundiärevuse hindamiseks ning koosneb 20 tunnusest nii tunnuse kui ka olekuvariandi jaoks. Osalejate ärevuse tase enne (M = 37.06, SD = 7.80) ja pärast (M = 39.33, SD = 9.16) katse ei muutunud oluliselt [t(32) = 1.61, p = 0.117]. Praeguse proovi ärevusnäitajad olid vahemikus 20 ja 58 (M = 36.6, SD = 8.98), mis on võrreldav tavalise täiskasvanute vahemikuga (Laux et al., 1981). PANAS (Krohne et al., 1996) on positiivse ja negatiivse meeleolu indeks. Isikud, kellel on positiivse mõjutuse skaala (PAS) kõrge skoor, on altid sellistele emotsioonidele nagu entusiasm, samas kui negatiivse mõju skaala (NAS) kõrge skooriga isikud on altid sellistele emotsioonidele nagu distress. Iga üksus koosneb omadussõnast ja osalejad näitavad skaalal vahemikus 1 (väga kergelt) kuni 5 (äärmiselt), kuivõrd omadussõna peegeldab nende tundeid sellel konkreetsel hetkel. Katse tulemusena osalejate negatiivses mõjus olulisi erinevusi ei leitud [algus: M = 11.67, SD = 2.29; lõpp: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Kuidagi muutsid osalejad oma positiivset meeleolu oluliselt algusest peale (M = 26.72, SD = 4.70) lõpuni (M = 23.66, SD = 7.02)t(31) = 3.11, p = 0.004]. See osalejate positiivse meeleolu langus võis olla seotud paradigma ebameeldivustega (esitati nii valulikke elektrilööke kui ka vastumeelset valget müra).
Menetlus
Laborisse saabumisel lugesid ja kirjutasid osalejad Würzburgi ülikooli psühholoogia osakonna eetikakomitee poolt heaks kiidetud informeeritud nõusoleku vormi. Neid ei teavitatud CS-de ja USA-de vahelistest olukordadest. Pärast küsimustike täitmist kinnitati elektroodid ja valu künnise protseduur viidi läbi nagu eespool kirjeldatud.
Ajal harjumise faasneed neli geomeetrilist kuju esitati kaks korda uuringutevahelise intervalliga (ITI), mis varieerus 18 ja 25 s vahel (keskmine: 21.5 s). Selles etapis ei tarnitud ühtegi USA või hämmastavat sondi.
Enne omandamisetappi tarniti iga 7-15 s seitse valget müra, et vähendada algset hämmastavust.
Järgmised kaks omandamisetapid olid identsed (joonis. \ t 1). Iga omandamisetapp koosnes 24i katsetest: 8 CS - uuringud, 8 avCS + uuringud ja 8 appCS + uuringud. CS-järjestus oli pseudorandom, kusjuures ainus piirang oli see, et sama stiimuli ei olnud võimalik esitada rohkem kui kaks korda järjest. Eriti esitati avCS + koos välgupoldiga kui elektrilöögi sümbol ja valulik USA tarniti offsetil. AppCS + esitati ühendis, kus oli pilt Smartiesest või soolane pretzel ja osaleja võis purgist valida Smarties või pretzel. CS-i esitleti koos keelusümboliga ja USA-d ei esitatud. Iga kolme tüüpi 8 CS-i esitusviiside ajal manustati 4i ja 6i vahel pärast ärrituse algust hämmastav sond. ITI-de ajal esitati kolm täiendavat hämmastav sondi, et tagada nende ettearvamatus ja vähendada hämmastavat harjumust. Musta ekraani sisaldav ITI varieerus 18 ja 25 s vahel 21.5-i keskmisega.
JOONIS 1
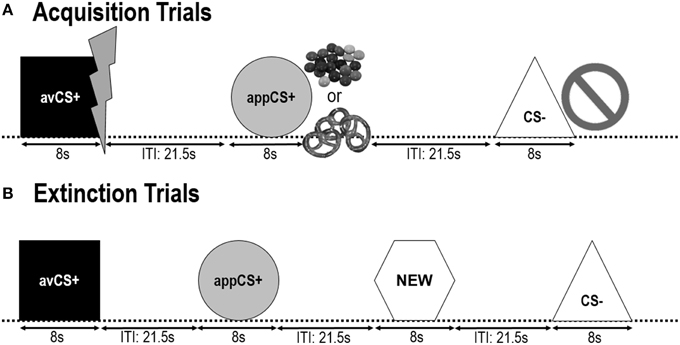
Joonis 1. Katsed kahe omandamisfaasi (A) ja väljasuremisfaasi (B) ajal. Osalejad said teada, et üks kuju (avCS +) ennustas kerget valulikku elektrilööki, üks kuju (appCS +) ennustas kas šokolaadi või väikese soolase pretzel (vastavalt nende eelistustele) ja kolmas kuju (CS–) ei ennustanud bioloogiliselt oluline sündmus. Iga kuju esitati koos pildiga, mis kujutab elektrilööki, nutikat / soolast küünt, või mitte midagi sõltuvalt sellest, milline on Ameerika Ühendriikidega. Väljasuremise faasis esitati veel kolm geomeetrilist kuju, kuid ühtegi USA-d ei esitatud. Lisaks esitati neutraalse kontrollina neljas geomeetriline kuju (NEW).
Ajal väljasuremisfaasosalejad nägid taas kolme geomeetrilist kuju (st avCS +, appCS + ja CS-) ja lisaks uut neutraalset kuju (NEW). USAd ei tarnitud ja kujundeid ei esitatud koos pilte välklampidest, šokolaadist / soolastest klotsidest või keeludest. Iga stiimul esitati kaheksakordselt pseudorandomjärjestuses (st sama stiimulit ei esitatud rohkem kui kaks korda järjest), luues 32i uuringud. 4-i ajal esitati iga CS-tüüpi 8i stiimulite esitlustest starti sondi stiimulid. Nagu omandamise ja harjumise faasis, varieerusid ITI-d 18i ja 25-i vahel ning 4-i täiendavad hämmastavad sondid saadi ettearvamatult ITI-de ajal.
Pärast iga etappi hindasid osalejad visuaalsete analoogkaaludega VAS-i valentsust (meeldivust) ja erutit (excitatory) alates 1-st 9-i. Valance skaala ulatus "1", mis näitab "väga ebameeldiv", kuni "9", mis näitab "väga meeldiv", erutuskiirus oli "1", mis näitab "rahulik", kuni "9", mis näitab "põnev". hinnatakse pärast kahte omandamisetappi ja väljasuremisfaasi. Osalejad nägid 1i jaoks geomeetrilist kuju ja seejärel pidid nad näitama, kas see kuju oli seotud elektrilöögiga, šokolaadi / soolase pretzeliga, mitte midagi või kas nad ei suutnud mingit seost teha. Eelkõige olid kõik osalejad (välja arvatud üks) teadlikud situatsioonist pärast omandamist 2.
Andmete vähendamine
Füsioloogilised reaktsioonid registreeriti V-Amp 16 võimendi ja Vision Recorder V-Amp Editioni tarkvaraga (versioon 1.03.0004, BrainProducts Inc., München, Saksamaa). Kasutati 1000 Hz ja 50 Hz sälgufiltrit. Ühenduseta analüüsid viidi läbi Brain Vision Analyzer'iga (versioon 2.0; BrainProducts Inc., München, Saksamaa).
Startle Response
Vastureaktsiooni mõõdeti elektromüograafia (EMG) abil vasakul orbicularis oculi lihaseid kahe 5 mm Ag / AgCl elektroodiga. Vastavalt suunistele (Blumenthal et al., 2005), üks elektrood paigutati õpilase alla ja teine 1 cm külgsuunas. Maandus- ja võrdluselektroodid paigutati vastavalt paremale ja vasakule mastoidile. Enne elektroodide kinnitamist hõõruti nahk kergelt ja puhastati alkoholiga, et hoida impedantsi allpool 10 kΩ. Elektromüograafiline signaal filtreeriti offline 28 Hz madala väljalülitusfiltriga ja 500 Hz kõrge väljalülitusfiltriga. Seejärel korrigeeriti EMG signaal ja rakendati 50 ms liikuvat keskmist. Me kasutasime 50 ms-i enne hätta sondi algust algtasemena (Grillon et al., 2006). Vastused hoogsatele sondidele määrati käsitsi ja uuringud ülemäärase algtaseme nihkega (± 5 μV) või liikumise esemeid jäeti edasisest analüüsist välja. 5 μV-st madalamad käivitusvastused kodeeriti nulliga ja neid kaaluti hämmastava suuruse arvutamisel (Blumenthal et al., 2005). Kokkuvõttes lükati 10.4% uuringutest tagasi ja 2i üllatusvastustest vähemalt iga 3-i reaktsioonivastuse faasis ja 4i väljalangemisfaasis 8-i puhul oli vaja iga seisundi puhul osaleja analüüsivahemikus. Sel põhjusel jäeti välja neli osalejat. Maksimaalne amplituud määratleti kui maksimaalne tipp võrreldes algväärtusega 20 – 120 ms ajaakna ajal pärast hätta sondi algust. Toorandmed normaliseeriti seejärel subjektide sees, kasutades z- skoorid ja seejärel T-skoorid, et vähendada individuaalse varieeruvuse mõju ja paremini tuvastada psühholoogilisi protsesse. T-skoorid keskmistati iga seisundi kohta (avCS +, appCS +, CS-, NEW ja ITI). Selleks, et uurida hämmastava potentsiaali või hoogu nõrgenemist, lahutati ITI hämmastavate vastuste skoorid iga seisundi hämmastavast vastusest.
Naha juhtivusreaktsioon (SCR)
Naha juhtivuse vastus (SCR) registreeriti, kasutades kahte 5 mm Ag / AgCl elektroodi, mis olid paigutatud mitte-domineeriva käe peopesale. Galvaniline vastus filtreeriti offline 1 Hz kõrge katkestusfiltriga. SCR määratleti kui erinevust (μS) vastuse alguse (1 – 3 s pärast stimulatsiooni algust) ja vastuse piigi vahel.Tranel ja Damasio, 1994; Delgado et al., 2011). SCR-i analüüsis ei võetud arvesse hätta sonde sisaldavaid katseid. Vastused allpool 0.02 μS kodeeriti nullina. SCR analüüsist jäeti välja veel viis osalejat, kuna nende keskmine SCR oli madalam kui 0.02 μS. Toores naha juhtivuse andmed olid ruutjuure transformeeritud, et normaliseerida jaotust ja skoorid keskmistati iga seisundi jaoks eraldi kahe omandamisfaasi (avCS +, appCS +, CS-) ja väljasuremisfaasi jaoks (avCS +, appCS +, CS- ja NEW ).
Statistiline analüüs
Kõiki andmeid analüüsiti SPSS for Windowsiga (versioon 20.0, SPSS Inc.). Füsioloogiliste vastuste jaoks arvutati kahe omandamisfaasi ja ekstinktsioonifaasi jaoks eraldatud mitmemõõtmelised dispersioonanalüüsid (ANOVA). Omandamise faaside ANOVA-l oli stiimul (avCS +, appCS +, CS-) ja faas (Acquisition 1, Acquisition 2) kui subjektide sisesed tegurid. Väljasuremise faasi ANOVA-l oli ainult stiimul (avCS +, appCS +, CS–, NEW) kui subjektide sisuline tegur. Valentsi-, erutus- ja situatsioonitasemeid analüüsiti eraldi ANOVA-dega, mis sisaldasid subjektide siseseid tegureid (avCS +, appCS +, CS- ja NEW) ja faasi. Sellel teguril oli valentsi- ja erutusnäitajate jaoks neli taset (T1: pärast harjutamist, T2: pärast esimest omandamisetappi, T3: pärast teist omandamisetappi, T4: pärast väljasuremise faasi), kuid kolm tasandit ettenägematute hinnangute jaoks (T1: pärast esimene omandamisetapp, T2: pärast teist omandamisetappi, T3: pärast väljasuremisfaasi).
Kõigi analüüside jaoks määrati alfa (α) tase 0.05is. Efektisuur on esitatud osaliselt η2.
Tulemused
Iga faasi valentsi- ja erutusväärtused on kujutatud joonisel fig 2; hämmastused ja SCR-id on kujutatud joonisel fig 3.
JOONIS 2
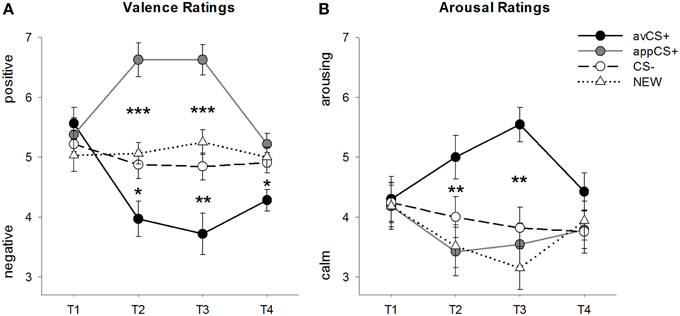
Joonis 2. Hinnang valentsile (A) ja erutusele (B). Liinid (standardvigadega) kujutavad reitinguid pärast harjutamise faasi (T1), Acquisition 1 (T2), Acquisition 2 (T3) ja väljasuremise faasi (T4). Aversiivne CS + (must tahke joon) omandas negatiivse valentsuse ja suure arousaluse pärast kahte omandamisetappi, võrreldes CS (must katkendjoon) ja NEW (must punktiirjoon). Oluline on see, et isuäratav CS + (halli tahke joon) omandas positiivse valentsuse võrreldes CS- ja UUS-ga. *p <0.05, **p > 0.01, ***p <0.001.
JOONIS 3
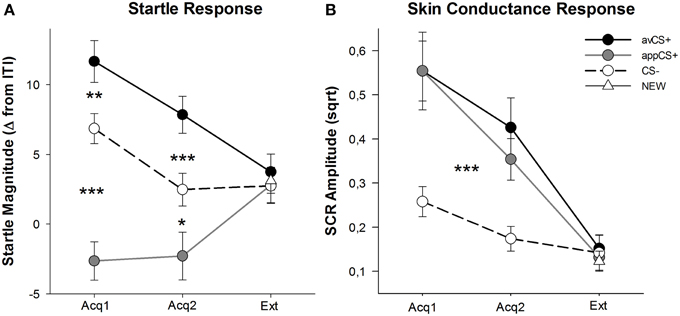
Joonis 3. Käivitusvastused (A) ja naha juhtivus (B) (standardviga) esimese omandamisetapi (Acq1), teise omandamisetapi (Acq2) ja väljasuremise faasi (Ext) puhul. Käivitusvastused tugevnesid märkimisväärselt aversiivseks CS + -ks (must tahke joon) ja nõrgendasid oluliselt omandamisfaaside ajal CS-i (must katkendjoon) suhtes isuäratav CS + (hall tahke joon). SCR oli oluliselt suurem kui avCS + ja appCS + võrreldes CS-ga. Väljasuremisfaasis ei ilmnenud erinevusi. *p <0.05, **p > 0.01, ***p <0.001.
Reitingud
ANOVA jaoks valentsireitingud omandamise ajal ilmnesid stiimulite olulised peamised mõjud [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, osaline η2 = 0.358] ja faas [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, osaline η2 = 0.096], samuti oluline interaktsioon stiimuli ja faasi vahel [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, osaline η2 = 0.235]. Järelmeetmed t-testid näitavad, et nelja geomeetrilise kuju valentsid katse alguses olid identsed (ps> 0.19), samas kui nii 1. kui ka 2. omandamise järel hinnati avCS + eriti negatiivseks ja appCS + eriti positiivseks. Täpsemalt hinnati avCS + CS-ga võrreldes negatiivsemalt hinnatud [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], UUS [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] ja rakendusCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + hinnati oluliselt positiivsemaks kui CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] ja UUS [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. CS- ja NEW-i erinevused ei olnud kunagi olulised (ps> 0.18).
Oluline on, et olulisi erinevusi ei leitud, kui võrreldi šokolaadi ja soolase pretzeliga seotud APCS + valentside reitinguid [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Pärast väljasuremise etappi hinnati avCS + endiselt CS-ga võrreldes negatiivsemaks [[t(31) = 2.40, p = 0.023], UUS [t(31) = 3.40, p = 0.002] ja rakenduste + [t(31) = 3.35, p = 0.002]. Vastupidi, appCS + valents ei erinenud enam CS-st [t(31) = 1.77, p = 0.086] või UUS [t(31) = 1.07, p = 0.293].
ANOVA jaoks arousal reitingud omandamise ajal ilmnes stiimulite oluline peamine mõju [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, osaline η2 = 0.181], kuid mitte faasist [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, osaline η2 = 0.038] ja märkimisväärne interaktsioon stiimuli ja faasi vahel [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, osaline η2 = 0.124]. Järelmeetmed t-testid ei näidanud olulisi erinevusi stiimulite vahel nende esialgse ärrituse suhtes (ps> 0.74). Kuid pärast kahte omandamisetappi hinnati avCS + -i erutavamaks kui UUT [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] ja rakendusCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] ja veidi erutavam kui CS - pärast esimest omandamisetappi [t(32) = 1.96, p = 0.058], kuid pärast teist omandamisetappi on see tunduvalt suurem;t(32) = 3.65, p = 0.001]. Vastupidiselt valentsireitingutele ei erinenud appCS + erutus CS-i ja UUU arousalist.ps > 0.13) pärast omandamisetappe.
Sarnaselt valentsireitingutega ei erinenud CS- ja NEW-i erutusest (ps> 0.07).
Nagu valentsuse puhul, ei ilmnenud erinevusi šokolaadiga seotud appCS + ja soolase pretzeliga seotud appCS + puhul [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Pärast väljasuremisfaasi ei ilmnenud olulisi erinevusi ärevusreitingutes (ps> 0.08).
Täiendav post-hoc t- uuringud, mis võrdlevad hinnanguid pärast harjumise faasi, esimest ja teist omandamisetappi ja väljasuremisfaasi, vt täiendavad materjalid.
Startle Response
Omandamisetappide ANOVA tagastas stiimuli peamise mõju [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, osaline η2 = 0.609], kuid mitte faas [F(1, 32) = 3.16, p = 0.085, osaline η2 = 0.090] ja märkimisväärne koostoime Stimulus × Phase [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, osaline η2 = 0.095]. Järelmeetmed t-testid näitasid, et avCS + on CS-ga võrreldes märkimisväärselt võimendatud, nii esimese [t(32) = 3.27, p = 0.003] ja teine [t(32) = 4.00, p <0.001] omandamisfaasid. Ehmatavad vastused avCS + -le tugevnesid ka oluliselt pärast rakenduseCSCS + vastuseid, jällegi pärast mõlemat 1. omandamist [t(32) = 8.20, p <0.001] ja omandamine 2 [t(32) = 5.74, p <0.001]. Oluline on, et rakenduse CS + ehmatuse suurus nõrgenes märkimisväärselt CS-ga - nii 1. omandamise ajal [t(32) = 6.34, p <0.001] ja omandamine 2 [t(32) = 2.91, p = 0.007]. Jällegi ja kooskõlas reitingutega ei ilmnenud appCS + puhul erinevusi, mis olid seotud šokolaadi või soolase pretzeliga [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Väljasuremise faasis ei leitud olulist mõju [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, osaline η2 = 0.008].
Naha juhtivusreaktsioon (SCR)
SCR-i ANOVA-delt kahe omandamisfaasi ajal on peamised mõjurite stiimulid [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, osaline η2 = 0.401] ja faas [F(1, 27) = 20.91, p <0.001, osaline η2 = 0.436] osutus oluliseks, kuid mitte nende koostoimeks [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, osaline η2 = 0.024]. Post-hoc t-testid näitasid oluliselt suuremat SCR-i avCS + -le [t(27) = 6.46, p <0.001] ja rakendusseCS + [t(27) = 4.84, p <0.001] võrreldes CS–-ga, samas kui osalejad näitasid SCR-i võrreldavat avCS + ja appCS + -ga [t(27) = 0.64, p = 0.527]. Eriti ei leitud erinevusi SCR-idel šokolaadiprogrammiga + ja soolase pretzel appCS + -ga [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Nagu ka reitingute ja hämmastava vastuse puhul, ei leitud väljasuremisfaasis olulist mõju [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, osaline η2 = 0.010].
Arutelu
Käesoleva uuringu eesmärk oli tõlkida loomade leiud inimestele, kasutades klassikalist isuäratavat konditsioneerimist paradigmat, mille esmane tugevdaja on tingimusteta stiimul (USA). Selleks tulid osalejad laborisse varahommikul ilma hommikusöögita, et kindlustada, et nad olid näljased, ning nende eelistuste kohaselt kasutati isuäratavaks USA-ks šokolaadi või soolase eelteksti. Omandamisetapi käigus sai üks geomeetriline kuju (avCS +) kergelt valulise elektrilöögi (aversive US), teise isu-USA-ga kuju (appCS +) ja kolmanda kuju (CS–), millel ei olnud mitte isuäratav USA ega ka mitte vastumeelne USA. Tulemused näitavad edukat vastumeelset ja isuäratavat konditsioneerimist selgesõnalisel verbaalsel tasemel (st hinnangud), kaudse käitumise tasemel (st hämmastav reaktsioon) ja füsioloogilisel tasandil (st SCR). Konkreetsemalt tõi avCS + võrreldes CS-ga rohkem negatiivseid valentsireitinguid, kõrgemaid erakordseid hinnanguid, üllatust ja suuremat SCR-i. Kõige tähtsam on see, et appCS +, võrreldes CS-ga, vallandas rohkem positiivseid valentsireitinguid, hoogu nõrgenemist ja suuremat SCR-i. Meie aversiivse konditsioneerimise tulemused olid kooskõlas ootustega, kuna varasemad uuringud on leidnud, et stiimul, mis ennustab ähvardust (avCS +), hinnatakse aversiivseks, põhjustab suuremaid hirmu vastuseid ja suurendab füsioloogilist erutust (Fendt ja Fanselow, 1999; Hamm ja Weike, 2005; Andreatta jt, 2010, 2013). Meie tulemused isuäratavale konditsioneerimisele olid kooskõlas ka eelnevate inimeste ja loomade uuringutega, mis näitavad, et tasu ennustav stiimul (appCS +) on hinnatud positiivseks, pärsib hirmu vastuseid ja suurendab füsioloogilist erutust (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin ja Duka, 2010; Prévost jt, 2012). Meie teadmiste kohaselt on see esimene uuring, mis demonstreerib, et inimestel on esmane tasu ennustav stimuleeriv hoogne nõrgenemine. Oluline on, et me võime loomkatse tulemusi üle kanda ja kinnitada.Koch et al., 1996). See loomkatse näitas, et rottide nõrgestamine sõltub NAcc (ventral striatum osa) eenditest PnC-le. Seetõttu võib meie uuringus tekkida hämmastav nõrgenemine NAcc aktiivsusele, mis on samuti kooskõlas fMRI tulemustega (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy ja Glimcher, 2011). Järelikult jõuame järeldusele, et meie isuäratav konditsioneerimise paradigma oli edukas, kuna nii selgesõnalised (hinnangud) kui ka kaudne (hämmastav sumbumine) positiivne valents on näidatud.
Lisaks sellele uuele, kuid pigem prognoositavale tulemusele väärivad mainimist veel kaks huvitavat tulemust. Esimesed, verbaalsed ja füsioloogilised erutusreaktsioonid rakendustele + dissotsieerunud. Teiseks leidsime kiiremini ära isuäratavad CR-d võrreldes aversiivsete CR-dega.
SCR näitab sümpaatilist aktivatsiooni, mida suurendatakse nii tasustamisega seotud stiimulile (appCS +) kui ka ohuga seotud stiimulile (avCS +). See tulemus on kooskõlas eelmise konditsioneerimisuuringuga, kus erootilisi pilte kasutati isuäratavaks USA-ks (Klucken et al., 2013) ja uuring, mis näitab võrreldavat SCR-i, mis näitab raha ja ennustavat müra ennustavat kihti (Austin ja Duka, 2010). Eriti on SCR orienteeruv vastus sümpaatilise süsteemi aktiveerimisega. See vastus on soovitatud kajastama käitumusliku reaktsiooni ettevalmistamist motiveerivalt olulistele sündmustele (Bradley, 2009). Selle põhjal arvame, et nii ähvardusega seotud kui ka tasustamisega seotud stiimulid tekitasid ettevalmistava vastuse järjestikustele käitumuslikele vastustele. Teisisõnu algasid toidu ja valu signaalid vastavalt lähenemise ja vältimise käitumise ettevalmistamisele. Vastupidiselt kõrgele füsioloogilisele erutusele viitasid verbaalsed vastused tasulise stiimuliga madalale erutusele. On võimalik, et verbaalset erutust mõjutab pigem USA ärritav olemus kui füsioloogiline aktiveerimine Rep. Tegelikult arvame, et appCS + vs avCS + hinnati madalaks ja väga ärevaks, sest need olid seotud madala ja väga ärevusega USA-dega. Kahjuks ei kogunud me USA-le meeleavaldusi, mistõttu on see tulevaste uuringute ülesanne seda hüpoteesi selgesõnaliselt testida.
Väljasuremise faasis ei toimetatud USA-d. See võib olla põhjustanud uue inhibeeriva õppimise, mida nimetatakse väljasuremiseks (ülevaateks vaata Milad ja Quirk, 2012), nii väheneb nii aversiivne kui ka isuäratav CR. Ilmselt ei olnud käitumuslikel (st hämmastav vastus) ja füsioloogilistel (st SCR) tasemel enam avCS +, appCS + ja CS-i suhtes erinevusi võimalik tuvastada. Samamoodi võrdsustati ekstinktsioonifaasi järel hinnatud konditsioneeritud stiimulite otsene erutus madalal tasemel, mis viitab edukale väljasuremisõppele. AvCS + hinnati siiski oluliselt negatiivsemaks kui CS-, samas kui appCS + valents ei erinenud enam CS-st. Aversiivse selgesõnalise vastuse aeglasem väljasuremine võib olla tingitud evolutsioonilisest konservatiivsusest, mis tähendab, et ähvardussignaale on eriti raske unustada, sest ohu signaali mittevastamine võib olla eluohtlik.
Jääb veel üks küsimus: miks siis kustutamise faasis (nt kaudne valents) kustutamine lõppes täielikult? Esiteks arvutati need reaktsioonid kogu faasi jooksul. Seetõttu on mõeldav, et ekstinktsioonifaasi esimestel katsetel oleksid diskrimineerivad CR-d veel tuvastatavad. Uurimuslikult järgisime seda hüpoteesi ja vaatasime nii hämmastuse vastuseid kui ka SCR-i kogu väljasuremisfaasis (vt täiendav materjal). Kuigi me ei leidnud märkimisväärseid erinevusi, täheldasime avCS + jaoks veidi suuremat hämmastavust võrreldes CS-ga ja uudsete kontroll-stiimulitega. Me täheldasime ka pisut rohkem hämmastavat sumbumist APCS + -ga võrreldes CS-ga ja uusi kontroll-stiimuleid väljasuremisfaasi alguses, mis aga kadus mõne katsega. SCR appCS + -le langes juba pärast 2nd-i väljasuremise uuringut, samas kui SCR-i avCS + -le jäi peaaegu kogu väljasuremisega võrreldes kõrgemale, võrreldes SCR-iga UUSI. Kuigi me peaksime neid tulemusi väga ettevaatlikult tõlgendama, tundus üllatav vastus paralleelselt valentsireitingutega. Veelgi enam, nii hämmastav vastus kui ka SCR toetavad veelgi ideed evolutsioonilisest konservatiivsusest ohu stiimulite suhtes.
Lõpuks peame tunnistama mõned uuringu piirangud. Esiteks, tehnilise probleemi tõttu ei saanud me teatada šokolaadi ja soolase pretzel meeldivuse (ja erutus) hindamistest. Samas viitavad isuäratavad CR-d, et osalejad kogesid kaht USA-d kui isuäratavat. Teiseks erinesid vastumeelse ja isuäratava USA kestus suuresti. Seega toimetati valus elektrilöök avCS + tasaarvestusse ja kestis täpselt 200 ms, samas kui šokolaad ja soolane pretzel esitati osalejatele umbes 2 s pärast appCS + algust ja kestus oli määramatu, sest see sõltus sellest, kui kiiresti isik sõi neid. Valik anda sellised isuäratavad USA-d, tugines varasemale uuringule inimestel (Prévost jt, 2012). Siiski oleks metoodiliselt rohkem elegantne pakkuda isuäratavamat USA-d rohkem võrreldavale USA-le, nagu näiteks mahla või ingveri ale, mis on rakenduses appCS +. Selle suure tajumise erinevuse kompenseerimiseks esitlesime geomeetrilisi kujundeid koos visuaalse stiimuliga, mis sümboliseerib USAd. Kolmandaks, me ei saa kindlasti välistada võimalust, et kiiresti kustunud füsioloogilised reaktsioonid väljasuremisfaasis tulenevad metoodilisest aspektist. Tegelikult ei esitatud selles faasis visuaalsed stiimulid koos USA sümboliga, nagu see oli omandamisfaasis. On võimalik, et CS-ide esitamine võib olla CR-d mõjutanud ja seetõttu ei olnud esimese väljasuremisuuringu ajal olulisi erinevusi. Kuid meie kiirelt kustutatud vastused on kooskõlas eelmise uuringu kustutatud vastustega, kus CS + (nägu) esitati ühendis USAga (karjumine) omandamise ajal, kuid mitte väljasuremise ajal (Lissek et al., 2008).
Kokkuvõttes leiti, et edukad aversiivsed ja isuäratavad konditsioneeritud reaktsioonid ohuga seotud stiimulile ja tasuga seotud stiimulile olid vastavalt. Huvitav on see, et selgesõnalised (reitingud) ja kaudne (hämmastav refleks) reageeringute tase töötasid sünergiliselt, kuna avCS + teatati kui negatiivset ja indutseeritud hämmastustugevust ning appCS + teatati positiivsest ja indutseeritud hämmastavast nõrgenemisest. Veelgi enam, selgesõnalised (reitingud) ja füsioloogilised (SCR) äratused rakendustest + dissotsieerusid, peegeldades kahte erinevat protsessi.
Huvide konflikti avaldus
Autorid kinnitavad, et uuring viidi läbi ilma kaubandus- või finantssuhete puudumisel, mida võiks tõlgendada võimaliku huvide konfliktina.
Tunnustused
Seda tööd toetas Collaborative Research Center “Hirm, ärevus ja ärevushäired”, SFB-TRR 58, projekt B1.
Lisamaterjal
Selle artikli lisamaterjali leiate veebisaidilt: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
viited
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. ja Pauli, P. (2013). Valu prognoositavus muudab leevendusega seotud stiimulite valentside väärtused vastupidiseks. Ees. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Abstract | CrossRef täistekst | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B. ja Pauli, P. (2010). Inimeste valu-leevendamise õpingute vaheline implantaadi ja selgesõnalise tingimuse vaheline vahe. Proc. Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Abstract | CrossRef täistekst | Google Scholar
Austin, AJ ja Duka, T. (2010). Mehhanismid tähelepanu pööramiseks suupärastele ja vastumeelsetele tulemustele Pavloviani konditsioneerimisel. Behav. Brain Res. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Abstract | CrossRef täistekst | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV ja van Boxtel, A. (2005). Komitee aruanne: juhised inimeste hämmastavate silmapiiride elektromüograafiliste uuringute kohta. Psühofüsioloogia 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Abstract | CrossRef täistekst | Google Scholar
Bouton, ME ja Peck, CA (1989). Konteksti mõju konditsioneerimisele, väljasuremisele ja enneaegse konditsioneerimise ettevalmistamisele. Anim. Õpi. Behav. 17, 188 – 198. doi: 10.3758 / BF03207634
CrossRef täistekst | Google Scholar
Bradley, MM (2009). Looduslik valikuline tähelepanu: orienteerimine ja emotsioon. Psühofüsioloogia 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Abstract | CrossRef täistekst | Google Scholar
Clark, JJ, Hollon, NG ja Phillips, PEM (2012). Pavlovia hindamissüsteemid õppimisel ja otsuste tegemisel. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Abstract | CrossRef täistekst | Google Scholar
Delgado, MR, Jou, RL ja Phelps, EA (2011). Neuraalsed süsteemid, mis põhinevad inimestel esmase ja sekundaarse tugevdajaga. Ees. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Abstract | CrossRef täistekst | Google Scholar
Dickinson, A. ja Balleine, B. (1994). Eesmärgile suunatud tegevuse motiveeriv kontroll. Anim. Õpi. Behav. 22, 1 – 18. doi: 10.3758 / BF03199951
CrossRef täistekst | Google Scholar
Fendt, M. ja Fanselow, MS (1999). Tingitud hirmu neuroanatoomiline ja neurokeemiline alus. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Abstract | CrossRef täistekst | Google Scholar
Gottfried, JA, O'Doherty, J. ja Dolan, RJ (2002). Inimeste söögiisu ja vastumeelset lõhnaõppimist uuriti sündmustega seotud funktsionaalse magnetresonantstomograafia abil. J. Neurosci. 22, 10829 – 10837. Saadaval võrgus aadressil: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. ja Johnson, L. (2006). Kontekstide ja käitumise vältimine virtuaalse reaalsuse keskkonnas: prognoositavuse mõju. Biol. Psühhiaatria 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Abstract | CrossRef täistekst | Google Scholar
Hamm, AO ja Weike, AI (2005). Hirmu õppimise ja hirmu reguleerimise neuropsühholoogia. Int. J. Psychophysiol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Abstract | CrossRef täistekst | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Konditsioneeritud seksuaalse erutuse omandamise närvi aktivatsioon: situatsiooniteadlikkuse ja soo mõju. J. Sex. Med. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Abstract | CrossRef täistekst | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). 5-HTTLPR-i polümorfism on seotud muutunud hemodünaamiliste reaktsioonidega isuäratava konditsioneerimise ajal. Hum. Aju Mapp. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Abstract | CrossRef täistekst | Google Scholar
Koch, M. (1999). Hämmastuse neurobioloogia. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Abstract | CrossRef täistekst | Google Scholar
Koch, M., Schmid, A. ja Schnitzler, H.-U. (1996). Närimiskahjustused häirivad hämmastuse leevendamist. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Abstract | CrossRef täistekst | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W. ja Tausch, A. (1996). Untersuchungen mit einer deutschen version der “Positiivne ja negatiivne mõju ajakava” (PANAS). Diagnostica 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I. ja Steele, JD (2008). Ebanormaalsed ajalised erinevused tasuvad õppimise signaale suuremas depressioonis. Aju 131, 2084 – 2093. doi: 10.1093 / aju / awn136
PubMed Abstract | CrossRef täistekst | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P. ja Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: Beltz Test.
Levy, DJ ja Glimcher, PW (2011). Õunte ja apelsinide võrdlemine: tasu-spetsiifilise ja tasu-üldise subjektiivse väärtuse esitamine ajus. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Abstract | CrossRef täistekst | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Kõrgenenud hirm konditsioneeritud sotsiaalselt oluliste tingimusteta stiimulite suhtes sotsiaalse ärevushäire korral. Olen. J. Psühhiaatria 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Abstract | CrossRef täistekst | Google Scholar
Martin-Soelch, C., Linthicum, J. ja Ernst, M. (2007). Isuäratav konditsioneerimine: närvialused ja mõju psühhopatoloogiale. Neurosci. Biobehav. Rev. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Abstract | CrossRef täistekst | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. ja Schoenbaum, G. (2011). Ventral striatum ja orbitofrontal cortex on mõlemad vajalikud mudelipõhiseks, kuid mitte mudelivabaks tugevdamiseks. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Abstract | CrossRef täistekst | Google Scholar
Milad, MR ja Quirk, G. (2012). Hirm väljasuremine kui translatsioonilise neuroteaduse mudel: kümme aastat kestnud edu. Annu. Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Abstract | CrossRef täistekst | Google Scholar
Pavlov, IP (1927). Konditsioneeritud refleksid: aju Cortexi füsioloogilise aktiivsuse uurimine. London: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM ja O'Doherty, JP (2012). Spetsiifilise ja üldise Pavlovi-instrumentaalse ülekande närvikorrelatsioonid inimese amügdalar-alampiirkondades: kõrge eraldusvõimega fMRI uuring. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Abstract | CrossRef täistekst | Google Scholar
Tranel, D. ja Damasio, H. (1994). Elektrodermiliste naha juhtivuse vastuste neuroanatoomilised korrelatsioonid. Psühofüsioloogia 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Abstract | CrossRef täistekst | Google Scholar
Märksõnad: klassikaline konditsioneerimine, tasu, karistus, hämmastav refleks, naha juhtivus
Viide: Andreatta M ja Pauli P (2015) isuäratav vs. Ees. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Vastu võetud: 11 veebruar 2015; Vastu võetud: 05 mai 2015;
Avaldatud: 19 mai 2015.
Muutis:
Niels Birbaumer, Tuebingeni Ülikool, Saksamaa
Arvustusi:
Tim Klucken, Justus Liebigi ülikool Giessen, Saksamaa
Alessandro Angrilli, Padova ülikool, Itaalia
Copyright © 2015 Andreatta ja Pauli. See on avatud juurdepääsuga artikkel, mida levitatakse vastavalt programmi tingimustele Creative Commonsi omandiõiguse litsents (CC BY). Kasutamine, levitamine või paljundamine teistel foorumitel on lubatud, kui algne autor (id) või litsentsiandja on krediteeritud ja et selles ajakirjas viidatakse algsele publikatsioonile kooskõlas aktsepteeritud akadeemilise praktikaga. Kasutamist, levitamist või reprodutseerimist ei ole lubatud, mis ei vasta nendele tingimustele.
* Kirjavahetus: Marta Andreatta, Psühholoogia osakond (bioloogiline psühholoogia, kliiniline psühholoogia ja psühhoteraapia), Würzburgi ülikool, Marcusstraße 9-11, D-97070 Würzburg, Saksamaa, [meiliga kaitstud]