YBOP-KOMMENTAAR: It Coolidge-effekt leit efter de krêft fan ynternetporno. It Coolidge-effekt is in ferskynsel te sjen yn sûchdiersoarten wêrtroch manlju (en yn mindere mjitte wyfkes) fernijde seksuele belangstelling hawwe as se wurde yntrodusearre oan nije ûntfangende seksuele partners, sels nei wegerjen fan seks fan foarôfgeande, mar noch altyd te krijen seksuele partners. Seksuele nijichheid oerskriuwt dizze gewoante mei fernijde eksitinsje feroarsake troch hegere dopamine. In trochgeande stream fan nijichheid is wat ynternetporn sa oars makket as porn fan eartiids.
Oarspronklik artikel, mei grafiken
- Dennis F. Fiorino,
- Ariane Coury, en
- Anthony G. Phillips
- De perioade fan 'e neurology, 15 juni 1997, 17 (12): 4849-4855;
Abstract
De Coolidge-effekt beskriuwt it werynrjochting fan seksueel gedrach yn in "seksueel sattige" dier as antwurd op in roman ûntfankende mate. Op grûn fan 'e rol fan' e mesolimbyske dopamine (DA) -systeem yn 'e inisjatyf en ûnderhâld fan motivearre gedrach, mikrodialysis waard brûkt om de nukleus accumbens (NAC) DA-transformaasje te kontrolearjen yn' e ferlinging, seksueel satsje en it werynrjochting fan seksueel gedrach. Yn oerienkomst mei eardere rapportaazjes wiene de presintaasje fan in easterlike frouljus efter in skerm en kopulaasje ferbûn mei wichtige ferheging fan NAC DA efflux. De weromkomst fan NAC DA-konsintraasjes nei baselinewilden fermindere mei in perioade fan seksuele satsje, hoewol't konsintraasjes fan de DA-metabolite, dihydroxyphenylessêsäure en homovanillyske sûr, bliuwe. De presintaasje fan in roman ûntfankende frou efter in skerm liede ta in lege ferheging fan de NAC DA, dy't opmerklik waard yn 'e fernijing fan' e froulike roman. De oanwêzige gegevens jouwe oan dat de stimulearre eigenskippen fan in roman-ûntfankende froulik tsjinje kinne om NAC-DA-transformaasje te ferheegjen yn in seksueu-satsige manlike rôt, en dat kin, yn ferliking, ferhâlde wurde mei it opnimmen fan seksueel gedrach.
Ynlieding
In manlike rat dy't de satsje opslein hat kin induzearre wurde om te moardearjen as de earste froulju ferfongen binne troch in roman ûntfankende froulike. Dit is bekend as de Coolidge-effekt en is bepaald yn in oantal sûchdieren (Wilson et al., 1963). Algemiene faktoaren lykas minderheid of motoryske depresje binne net genôch om de skynbere steat fan seksueel satiaasje te ferklearjen, om't stimulearjen fan in romanfamylje noch altyd ynfloed kinne. Seksuele sêne kin ek ferkocht wurde, pharmakologysk, yn in geweldich nivo, troch it administraasje fan in ferskaat fan medisinen dy't kinne kinne op ferskate neurotransmitter-systemen. Dizze medisinen binne yhimbine, 8-OH-DPAT (Rodriguez-Manzo en Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus en Gorzalka, 1987; Rodriguez-Manzo en Fernandez-Guasti, 1995a,b), en apomorphine (Mas et al., 1995c). Hoewol it peripherale aksje fan dizze drugs kin net útsteld wurde (bygelyks adrenerike effekten op eerectilefunksje), effekten op sintraal meganisme dy't ûnderdiel fan seksueel satsje binne opsteld op grûn fan selektive sintraal noradrenergyske lysjonekseksjes (Rodriguez-Manzo en Fernandez-Guasti, 1995a) en mikrodialysis eksperiminten dy't dopaminergyske metabolism yn 'e mediale preoptyske gebiet (Mas et al., 1995a,b).
Trochdat sintraal meganismen de ferwidering fan it seksueel hâlden fan 'e Coolidge-effekt fertsjintwurdigje, is in wierskynlik kandidaat it mesolimbysk dopamine (DA) systeem, projektearje fan it ventral tegmentale gebiet nei de NAC. Mesolimbic DA liket as primêre modulator te aktivearjen yn komplekse yntegraasjeprosessen dy't de evaluaasje fan natuerlike stimulaasjes beynfloedzje, lykas toetsen fan in seksueel ûntfankende froulike, en de organisaasje fan doelstellige gedrach, ynklusyf oerwinning (Fibiger en Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Hoewol mid-drain DA neurons reagearje op primêre belestingen en foarbylden fan belesting, romantyk of unfoarriedbere omlizzende stimulearrings stimulearje de neuroanale aktivearring meast sterk oer werhelle treningssites (Fabre et al., 1983; Schultz, 1992; Mirenowicz en Schultz, 1994). Der is in protte bewiis dat in wichtige fasilitearjende rol stipet foar mesolimbic DA yn 'e ynisearring en ûnderhâld fan rykseeksuele gedrach (Pfaus en Everitt, 1995), en in oantal mikrodialysisûndersiken rapportearret yn NAC DA efflux yn 'e appetitele en folsleine faze fan manlike seksuele gedrach (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Der binne lykwols relatyf in pear gegevens op 'e neurochemyske korrelaten fan seksueel satsje en it werynjen fan seksueel gedrach. De tapassing fan yn vivo Mikrodialysis om 'e mesolimbyske DA-neurotransmission yn' e Coolidge-effekt te bewarjen biedt in unike kâns om de rol fan 'e NAC-DA te beskôgjen yn fergrutting, seksueel satsje en it werynrjochting fan' e ferwaging.
In mikrodialysis eksperimint waard dien om de folgjende te bepalen: (1) oft de komst fan seksuele satsjes begeliedt wurdt troch it weromkommen fan ekstrazellulêre DA-konsintraasjes yn 'e NAC foar prestopulaasjewearden of ûnderen, en (2) oft de werstelingen fan' seksueel satetiate "manlike rat mei in roman-ûntfankende frou is korrelearre mei ferheegingen yn NAC DA efflux.
MATERIALEN EN METOADES
Subjects. Male Sprague Dawley raten, krigen fan it Animal Care Centre (oan de Universiteit fan Britsk-Kolumbia), en froulike Long-Evans raten, krige fan Charles River Kanada (Sint Constantius, Quebec, Kanada), waarden yn wire mesh cages (18 × 25 × 65 sm; fiif per cage) yn aparte koloanjes. Koloanske-kamielen waarden bewarre by in temperatuer fan ~20 ° C op in weromreis 12 hr ljocht / tsjuster fyts. Rieten hienen unbegripend tagong ta iten (Purina Rat Chow) en wetter.
Surgery en gedrachstipe foar brain-mikrodialysis.Frouljusrotten waarden bilateraal ovaliektomisearre ûnder halotaane gasnestystia (Fluothane, Ayerst Laboratories) op syn minst 4 wiken foar testen. Seksuele yntimptaasje yn 'e stimulusfemalen waard troch subkutane ynjeksjes fan estradiolbenzoate (10 μg) en progesteron (500 μg), 48 en 4 hr, respektivelik, foardat elke test-sesje beynfloede. Guon ratten waarden skrepronearre foar seksuele gedrach op twa kear, 4 d apart, yn Plexiglas kameren (35 × 35 × 40 cm) mei wire-mesh floors. Allinne manlike raten dy't in kritearium ynfierd hawwe, wêrtroch yntromissionen yn 5 min fan 'e presintaasje fan' e froulju en ejaculationen yn 'e 15 min fan' e earste yntromissionen yn 'e beide screeningtests waarden ymplantearre mei microdialysis probe guide cannulae.
Male rats (n = 5) waarden anesthetisearre mei ketaminehydrochloride (100 mg / kg, ip) en xylazine (10 mg / kg, ip) foar stereotaksyske chirurgie. Mikrodialysis-problevel-cannulae (19 gauge) waarden bilateraal oer de NAC (koördinearre troch bregma: anterior, + 1.7 mm, mediale, ± 1.1 mm; ventral, -1.0 mm; flakke skul) en waarden befestige oan 'e skuon mei tosken acryl en Juweliers skroeven. Bilaterale guide cannulae-ymplantsjes waarden brûkt om de kâns te maximearjen foar in suksesfolle mikrodialysis eksperiment. Gelokkich, yn it hjoeddeis eksperimint, waard mar ien kaniel nedich foar elke rat. Manlike ratten waarden yndividueel yn grutte plastike kaarten mei corncob bêdden foar de rest fan it eksperimint ûnderbrocht. Ien wike nei surgery wurde ratten getten foar seksuele gedrach. Yn dit part fan training waard de testkamera mei in slide Plexiglas skerm oprjochte dat de keamer yn grutte en lytse fakma's ferdield. Manlike rotten waarden yntrodusearre yn it grutte fak en 15 minne letter, in frou waard efter it skerm pleatst. Nei in presintatuerperioade fan 15 min waard it skerm fuortsmiten, en de ratten koenen har foar 30 kompilearje. Trije treningssesjes waarden lein, ien elke 4 d. Alle ratten berikten it optredens kritearia by elke sesje.
Coolidge effekt eksperimint. Ratten waarden unilateraal ylstien mei microdialysis problemen 12-18 hr foardat de Coolidge effekt eksperimint en pleatst yn it grutte fak fan 'e testkamera mei frije tagong ta iten en wetter. Op 'e moarn fan' e eksperiminte waard mikrodialysis problemen elke 15 min sammele. It eksperimint bestie út 'e neikommende sân konsekte fazen: (1) baseline (op syn minst 60 min); (2) frou 1 efter it skerm (15 min); (3) kopulaasje mei froulik 1 oant in minne perioade fan 30 sûnder in berch trochjûn; (4) werynrjochting fan froulik 1 efter it skerm (15 min); (5) tagong ta frouljus 1 foar in minne perioade fan 15, wêrtroch't der gjin montaazjes wie (as it opnij kaam, dizze fase wie behannele as faze 3); (6) ynlieding fan froulik 2 efter it skerm (15 min); 7) kopulaasje mei froulik 2 foar 60 min.
Behavior waard ûnder loftljochting mei in JVC-video-systeem filminearre en fûn op in video-monitor dy't bûten de toetseboerd leit. Standert maatregels fan seksuele gedrach waarden opnommen mei in kompjûter en passende software (Holmes et al., 1987).
Nei it mikrodalyse eksperimint krigen de bisten in overdosis fan chlorale hydrate en pervyen yntrakarklik mei saline en formaline (4%). Brainezjes waarden ynsletten en beferzen, en letter waarden koronale dielen mei kresyl-fiolet oanbean om de pleatsing fan mikrodialysis-problemen te bepalen. Allinnich ratten mei probeplateminten yn 'e NAC waarden brûkt foar gedrachs- en neurochemyske analyzes.
Mikrodialysis en HPLC-elektrochemyske deteksje. Mikrodialysis-problemen wiene konsonrisch yn ûntwerp mei in semipermeabele hollefasermembran (2 mm membrante eksposearre, 340 μm eksterne diameter, 65000 molekulêre gewichtskoart, Filtral 12, Hospal) oan 'e distalen ein. Sesjes waarden perfoarst op 1.0 μl / min mei in wizige Ringer-oplossing (0.01 m natrium phosphate buffer, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) mei in gasdjippe (Hamilton, Reno, NV) en in syringepomp (model 22, Harvard Apparatus, Súd Natick, MA). In mikrodialysis-probe-guide-collar waard brûkt om de mikrodialysis-probe yn 'e gidehannel te befêstigjen. In stielspul, befestige oan in flechtsje (Instech 375s) dy't oan 'e top fan' e testkamera befette waard, waard brûkt om de probeproep te beskermjen (Fiorino et al., 1993).
Mikrodialysat-analyten, wêrûnder DA en metabolites dihyroxyphenylessetic acid (DOPAC) en homovanillyske sûger (HVA), waarden ôfset troch reverse-fase-chromatografy (Ultrasphere kolom; Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, ) mei in 0.083m natriumacetate-puffer, pH 3.5 (5% methanol). Analyte-konsintraasjes waarden kwantifisearre troch elektrochemyske (EC) deteksje. It apparaat bestie út in Bio-Rad (Richmond, CA) pomp, in twa-posysje ynjector fan Valco Instruments (Houston, TX) EC10W, in ESA (Bedford, MA) Coulochem II EC-detector, en in dual-channel chartrekorder (Kipp en Zonen, Bohemen, NY). Electrochemyske detectorparameters wiene de folgjende: electrode 1, + 450 mV; electrode 2, -300 mV; en wachtzelle, -450 mV. Typyske probleemferwideringen, ferdwûn in vitro en by keamertemperatuer wiene 22% foar DA, 18% foar DOPAC, en 18% foar HVA.
RESULTS
Hâlden en dragen
Behavioral maatregels út it Coolidge effekt eksperimint wurde presintearre yn Tafel 1. Latenêsten foar berch, yntroit, en ejakatoaren, lykas it postjaculatory ynterval nei de earste ejaculation wiene lykwols ferlykber mei dy yn 'e foargeande treningssitting (data net werjûn). Dit liedt oan dat it mikrodialysisproseduere gjin normale seksuele gedrach feroaret. De ûntwikkeling fan seksueel satiaasje, lykas oanjûn troch it betsjuttingsnivo fan ejakaasjes foardat it kritearium fûn waard (7.8 ± 0.5), in progressive ôfnimming fan it tal yntromisjes foar elke ejakulaasje foarôfgeand, en in progressive ferheging fan it postejaculatory ynterval (gegevens net te sjen) , wie ferlykber mei dat rapportearre yn eardere stúdzjes (Strand en Jordaanje, 1956; Fowler en Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo en Fernandez-Guasti, 1994; Mas et al., 1995d). Yndividuele fariabele wurde bepaald yn ferbân mei it tal ejaculaasjes dy't binnen it frouljus 1, de tiid dy't mei female 1 ferbruts, en it tal presintaasjes fan froulju 1 nedich om it satiatyskritrams te berikken (Tabel 1, ûndergrûn). Guon raten fereigen in soad yninigings fan froulike 1 oant faze 5 folslein (n = 3). De akten fan it pleatsen fan froulju 1 efter it skerm en it fuortheljen fan 'e dieling hawwe meast brûkt as primêre appetitele toanen dy't liede ta kopupulaasje. It moat ek oanjûn wêze dat in satetiidskriterium fan 30 min sûnder in berch, hoewol earder brûkt brûkt (Strand en Jordaanje, 1965; Mas et al., 1995b), is willekeurige en net garandearret dat in rat soe net mear tiid krije. Sels lykwols binne ferplanten of ferwidering en ferfangende prosedueres net te leverjen yn ferlingde kopulaasje mei froulik 1 (bgl. Fazen 4 en 5).

Behavior by it Coolidge effekt eksperimint
Alle ratsen eksposearren it Coolidge-effekt. De aktiviteiten dy't tawiisd binne mei it pleatsen fan froulike 2 efter it skerm en, yn it bysûnder, it fuortsmiten fan 'e partysje, kinne bydroegen hawwe oan dit resultaat, mar, opnij, binne dizze eveneminten net sels genôch om earder yn' e eksperimint kopululaasje te herheljen. Fergeliking tusken maatregels fan seksueel gedrach mei froulik 1 en froulik 2 waarden makke gebrûk t Tests mei in Bonferroni korreksje. Alhoewol't berch en yntromisaasje Lattenkomsten yn antwurd op froulike 2 net oars fanwege dyjingen yn 'e earste kopulatory bout mei froulike 1, yn it algemien, seksueel gedrach mei froulik 2 wie minder robúst, lykas oanjûn troch minder lytsere ejaculaasjes (gemeen, 0.6 vs 4.2; F = 49.86;p <0.01) en yntromysjes (gemiddelde = 11.2 tsjin 37.0;F = 20.17; p <0.05) tidens it earste oere. De oantallen mounts yn it earste oere mei wyfkes 1 en 2 wiene net signifikant oars.
It is wichtich om te notearjen dat froulju brûkt wurde yn 'e satearring fan' e eksperimint (dus de froulike 1) noch altyd sterke proefytsjen (ie, hopping en darting) en resptive (ie, lordose) gedrach foar de folsleine tiid fan har kontakt mei de manlju .
Neurochemistry
Basal-nanomolar-konsintraasjes fan DA en metaboliteiten yn mikrodialysaten, presintearre as de gemiddelde ± SEM, fan 'e earste trije baseline-samples binne: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; en HVA, 234.2 ± 49.0 (ûnkreprikearre foar probe recovery;n = 5). Dizze wearden fertsjinwurdigje 100% baseline-punten.
Behannele definiearre gegevenspunten, dy't elke faze fan 'e eksperiment en mienskiplik foar elke rat binne, binne brûkt foar neurochemyske analyzes. Dizze binne ûnder oaren de folgjende: (1) sân samples nei de earste ynlieding fan froulike 1, (2) fjouwer samples oanbean mei de ôfwêzichens fan 'e copulatory gedrach mei froulik 1, en (3) fiif samples nei presintaasje fan froulike 2. Sifer 1 jout feroarings yn konsintraasjes fan DA (line graph, midden) en DA metabolites (line graph, top) paralleling copulatory behaviors (bar graph, boaiem) by it test foar de Coolidge-effekt.
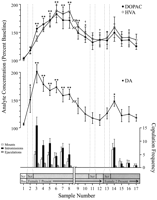
Fig. 1.
Nucleus accumbens neurochemyske korrelaten fan seksueel gedrach ûnder it Coolidge-effekt. De earste acht problemen fertsjinje chronologysk kontinuvere gegevenspunten fan 'e faze 1 nei 3. Sample 1 is de fjirde en lêste preekopulaasje baseline-samples (Bas). Sample 2 betsjut ynlieding fan froulike 1 efter it skerm (Scr). Nei 15 min waard it skerm fuortsmiten, en ratten wiene fergees te kopulearjen (samples 3-8). Debrekke op 'e x-aksis komt oerien mei it útsluten fan gegevens fan trije ratten dy't foar ferlingde perioaden mei de earste froulju kooppele binne. De lêste njoggen samples wiene ek kontinuze chronologysk. Samplijen 9 en 10 oerienkomme mei de satetiidperioade fan 'e faze 3 (dus, 30 min sûnder in berch). Fiif 1 waard doe efter it skerm ferfongen (sample 11) en 15 minne letter waard it skerm fuortsmiten (sample 12). Nei 15 min fanwege kopulaasje, waard froulike 2 efter it skerm pleatst (sample 13). Samples 14-17 oerienkomme mei kopulaasje mei froulik 2. It oantal moasten, yntrimen, of ejaculaasjes dy't ferbûn binne mei elk 15 min mikrodialysis-echte sample, binne yn 'e bottom bar graph. Neurochemyske gegevens wurde ekspresearre yn termen fan persintaazje fan baseline-konsintraasjes. Feroarings yn NAC DA (sletten fjilden), DOPAC (sletten sirkels), en HVA (iepen sirkels) efflux wurde presintearre as line grafiken. De neikommende fergelokken waarden makke: baseline sample 1 tsjin testen 2-10; nije basisprobe 10 tsjin testen 11 en 12; nije baseline-sample 12 tsjin testen 13-17 (*p <0.05; ** p <0.01). Ûnôfhinklik t Tests waarden makke tusken baseline-wearden (samples 1, 10, en 12). Foar signifikante ferskillen fan 'e earste baseline (sample 1), †p <0.05.
Separate ien-wize, werhellende maatregels ANOVA's waarden dien dien op neurochemyske data dy't ferbûn binne mei froulik 1 (samples 1-12) en froulike 2 (samples 12-17). A priori Feroarings waarden makke mei Dunn's ferskate fergelikingstest (Bonferroni t). De folgjende trije haad fergelokken waarden makke: (1) earste basisline (sample 1) fersoene samples 2-10 (earste eksposysje nei froulike 1), (2) twadde basislinen (sample 10) tsjin samples 11 en 12 (werhelling fan froulike 1) , en (3) tredde baseline (sample 12) tsjin testen 13-17 (eksposysje nei froulike 2).
Der wie in grutte feroaring yn DA efflux as antwurd op froulike 1 [F (11,44) = 8.48; p <0.001] en froulik 2 [F (5,20) = 2.83;p <0.05]. In signifikante ferheging fan DA-efflux waard fûn doe't froulike 1 efter it skerm wie (+ 44%,p <0.05; foarbyld 2). Tidens kopulaasje naam DA-konsintraasje fierder ta, en berikte in maksimale wearde (+ 95%;p <0.01) tidens de earste copulatory bout (sample 3). DA bleau de heule kopulaasje ferhege en kaam allinich werom nei baseline-konsintraasjes yn 'e perioade fan 30 min wêryn gjin montage foarkaam (samples 9 en 10). Weder opnij yntroduksje fan froulik 1 efter it skerm (foarbyld 11) noch de kâns om fysyk te ynteraksjen, mar sûnder montage (foarbyld 12), ferhege DA-konsintraasjes relatyf oan 'e twadde basiswearde (foarbyld 10). De oanwêzigens fan froulike 2 efter it skerm (foarbyld 13) resultearre yn in lytse ferheging fan DA efflux (12%) fan 'e tredde basiswearde (foarbyld 12) dy't gjin statistyske betsjutting berikte. Fernijde kopulaasje mei froulike 2 resultearre yn in signifikante (34%) ferheging (p <0.05) yn DA-efflux tidens de earste kopulaasjeprobe (sample 14). Hoewol swak copulatorysk gedrach trochgie oer de kommende trije samples, waarden DA-konsintraasjes fermindere nei basiswearden (samples 15-17). Ûnôfhinklik t Tests ûnder testen fan 'baseline' (ie, 1, 10, en 12) wiene demonstrearre dat dizze wearden net sa folle oars wiene.
Yn 'e trije ferdivedaasjes dy't de kopulaasje wûn doe't de froulike 1 werombrocht waard, waard de NAC-DA-konsintraasjes grutter as de froulike 1 efter it skerm presintearre (berik, 25-47%) en yn' e gearkomst (berik, 13-37%), foar it werynjen fan 'e froulike. Dizze ferheging lykwols pas allinich as seksueel gedrach krêftich en liede ta ejaculation.
Wichtige algemiene feroarings yn DOPAC [F (11,44) = 9.57; p <0.001] en HVA [F (11,44) = 12.47; p <0. 001] konsintraasjes waarden fûn yn antwurd op froulik 1, mar net froulik 2. Metabolite-konsintraasjes naam in bytsje ta (+ 15% yn beide gefallen) by de presintaasje fan froulike 1 efter it skerm (foarbyld 2), mar dit wie net signifikant statistysk. D'r wiene lykwols signifikante ferhegingen fan 'e konsintraasjes fan DOPAC en HVA by kopulaasje (samples 3-8), berikke maksimale wearden (respektivelik +80 en + 86%; p <0.01) nei 60 min (sample 6 yn beide gefallen). Hoewol metabolitekonsintraasjes ferminderen yn 'e perioade fan seksuele ynaktiviteit oan' e ein fan kontakt mei froulike 1 (samples 9 en 10), bleaune konsintraasjes noch altiten ferhege mei respekt foar baseline (p <0.05 yn beide gefallen). Reintroduksje fan froulik 1 efter it skerm (foarbyld 11), tagong ta froulik 1 nei ferwidering fan it skerm (foarbyld 12), en de yntroduksje fan froulik 2 (foarbyld 13) resultearre net yn feroaringen yn metabolitekonsintraasjes. Lytse, mar statistysk unbelangrike ferhegingen fan DOPAC- en HVA-konsintraasjes (+ 23% yn beide gefallen) relatyf oan basisline (sample 12) kaam oerien mei de earste oanfal fan kopulaasje mei froulike 2 (sample 14). Dizze ferheging wie lykwols koart libbe en wegere nei baseline wearden foar de oerbleaune trije samples (15-17). Ûnôfhinklik tTests ûnder testen fan 'baseline' (xNUMX, 1, 10) oanjûn dat de twadde en tredde baseline-wearden (samples 12 en 10, respektivelik), hoewol net ôfwike fan elkoar, ferhege bleaun ferlike mei de earste baseline-samling foar DOPAC en HVA (p <0.05 yn beide gefallen).
Histology
Mikrodialysis probes wiene yn 'e NAC (Abb.2) yn in range omheech te meitsjen + 1.20 nei + 1.70 mm fan bregma (flakke skul). Der wie ek variabelens yn 'e mediolaterale fleantúch; Gegevens reflektje sampling fan 'e shell en core subregionen fan' e NAC.
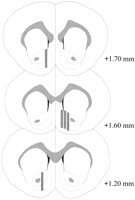
Fig. 2.
Lokaasje fan mikrodialysis problemen binnen de NAC fan manlike raten dy't yn 'e Coolidge effekt eksperimint brûkt wurde. Shaded rectangles meidwaan oan it eksponearre membraangebiet fan de mikrodialysis problemen. Serial coronal brain seksjes waarden opnommenPaxinos en Watson (1986).
DISKUSJE
Yn oerienkomst mei eardere rapportaazjes binne de oanwêzige resultaten sjen litte ferhevene mesolimbyske DA-transmoarch dy't ferbûn binne mei appetitele en folsleine komponinten fan manlike ritme seksuele gedrach as beoardielje trochyn vivo microdialysis (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Dêrnjonken jouwe dy resultaten in neurochemyske korrelat foar seksuële satsjasje en de folgjende ynstânsje fan 'e kopulaasje yn antwurd op in roman-ûntfankende frou (de Coolidge-effekt). De oanwêzige gegevens jouwe oan dat de stimulêre eigenskippen fan in roman-ûntfankende froulik tsjinje kinne om de NAC-DA-transformaasje te ferheegjen yn in seksueuze sittige manlike rôt dat bygelyks ferbân hâlde mei it opnimmen fan seksueel gedrach. Dit is foar it earst yn 'e lege ferheging fan NAC DA yn' e presintaasje fan in roman frouljus efter it skerm en komt it meast oertsjûge as in mear útwreide ferheging yn 'e fernijende kopulaasje mei froulik 2 (Fig.1).
De oanwêzichheid fan 'e earste ûntfankende frou efter it skerm liede ta in robúste appetitele groei fan NAC DA efflux (44% fan baseline) ferlykber yn maat oan wat yn in eardere eksperiminten rapporteare waard mei in fergelykber ûntwerp (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992). Ek yn oerienstimming mei dizze stúdzjes wie de beoardieling dat NAC DA efflux fierder waard fersterke by kopulaasje (oant> 95% boppe baseline yn it hjoeddeiske eksperimint). Hoewol kinne wy folsleine gedrach sjen as assosjeare mei ferbettere NAC DA-frijlitting (Wenkstern et al., 1993; Wilson et al., 1995) is it wichtich om de termen "appetitive" en "folslein" te ûndersykjen yn 'e kontekst fan seksueel gedrach. Wylst de faze wêrby't de froulju efter it skerm oanwêzich is, is allinich appetitatyf of tarieding, kin it gedrach yn 'e oerlizzende faze net allinich as folsleine ferplichting beskôge wurde. Om't "appetitich" brûkt wurde kin wurde om alle gedrachsprizen te beskriuwen dy't liede ta de folsleine gedrach fan in motivearre gedrach (oplossing), it primêre gedrach fan 'e manlike eksposysjes is aktyf yn' e "folsleine" fase better beskreaun as appetitele; De man bestjoert de measte fan syn tiid en de opdracht dy't de frouljus ferfolget om te ferpleatsen. Yn dat ferbân kinne wy de maksimale NAC DA-transmysjiaasje mei ynkommens hawwe lykas Yntinsyf oanwêzige komponinten fan manlike ritme seksuele gedrach.
Tagong nei de twadde, roman froulik levere in fernijende kopulaasje yn elke ûnderwerp. Foarige ûndersiken hawwe te sjen dat de mearderheid fan 'e raten te krijen hawwe om satsje te kopearjen, mei it brûken fan in ferlykber gedrachsprotokol oan' e ien dy't yn it tsjintwurdige eksperimint brûkt, net foltôgje doe't 24 hr letter (Strand en Jordaanje, 1956). It is wierskynlik dat de oanwêzichheid fan 'e roman stimulearre eigenskippen fan' e froulike 2, dy't mooglik olfaktoryske as ek fisuele en audiofoarsaken opnommen hawwe, feroarsake yn ferlingde kopulaasje. In nijsgjirrige fraach, dy't bliuwt te beantwurdzjen, is troch hokker meganisme in manlike rôt ûnderskiedt fan in froulike frou út in froulik dêr't er koartlyn mate hat. In side foar dat meganisme kin lizze yn it haad olfaktorysysteem. It is rapportearre dat de yntegriteit fan dit systeem essentiel is foar de Coolidge-effekt yn hamsters (Johnston en Rasmussen, 1984). It ferdylosasal-accessory-olfaktoryske systeem, lykwols, dêr't in pheromonale ûnthâldprosessen koartlyn beskreaun waard yn mûzels (Kaba et al., 1994), is ek in prime kandidaat. Yn dit ferbân is it bewiis dat ferheging fan NAC-DA-transmysjestaasje wurde brûkt yn vivo voltammetry yn manlike raten presintearre mei bedding dy't eksposearre waard oan froulike ratten yn estrus (Louillot et al., 1991; Mitchell en Gratton, 1992). Fierder is de tapassing fan K+ Direkteur oan 'e deeronasale nerveur fan' e tafoegjende olfaktorykolom, en ek oan 'e accessory olfactory bulb sels, wie genôch om NAC DA-transmission te ferheegjen (Mitchell en Gratton, 1992).
De earste 15 min bout fan kopulaasje mei froulik 2 wie ferbûn mei in signifikante groei fan NAC DA. Yn tsjinstelling mei froulik 1 hat ynteraksje mei froulik 2 gjin ferheging yn 't NAC DA fan deselde magnitude yn' e mande fan 'e appetitive (12%) of ynkommende (34%) fazen. Dizze lytse ferheegingen yn 't NAC DA lykwols goed mei it legere nivo fan seksueel gedrach werjûn mei frou 2 yn fergeliking mei froulik 1. Metabolite konsintraasjes blieken yn 'e sateaze faze blommen bleaun, wêrtroch't in nije basisline konsintraasjes (samples 10 en 12) binne fanwege it begjin fan basearre wearde (problemen 1) ferhege.
De tydlike lagen yn 'e ferheging fan DOPAC en HVA-konsintraasjes yn' e gearkomst is konsekwint mei harren formaasje as metaboliteiten fan 'e memmedei, DA. It is oanwêzich dat mikrodialysis metabolite konsintraasjes, op syn minst yn in natuerlik gedrach, dat net pharmakologysk oanwêzich is, in nuttige yndeks fan neurele aktiviteiten leverje (Damsma et al., 1992; Fumero et al., 1994). It feit dat metabolite konsintraasjes noch yn 'e perioaden fan seksuele ynaktiviteit bliuwe yn dit eksperimint, doe't DA-konsintraasjes weromkamen nei foarstelde wearden fan baseline, draaide twifel op dizze suggestje.
De persoanlike heuveling fan DA metabolite konsintraasjes dy't yn dit eksperimint sjoen sjoen spegelje it meditaal foar preoptike gebiet (mPOA) profyl fan DA metabolites dy't observearre wurde yn raten de earste dei nei't se kopieare hienen foar siedingen (Mas et al., 1995a,b). Stavere ferheveningen fan DOPAC- en HVA-konsintraasjes yn 'e NAC of mPOA wurde net altyd beoardiele as de ferlingende perioade fan in fêste tyd is, folle koarter dan de tiid dy't nedich is om satetiid te berikken. Bygelyks, in protte stúdzjes hawwe te sjen dat DOPAC-konsintraasjes ferhege waarden en bleaunen yn 'e ferwaging, mar se koenen net nei de frou weihelle wurdePfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Yn 'e stúdzje troch Mas et al. (1995b), basale ekzellekulêre konsintraasjes fan DOPAC en HVA yn 'e mPOA bleaunen op 4 opfolgjende dagen ferhege bleaun oerien mei in perioade fan seksuele ynaktiviteit. Op de fjirde dei, krekt foardat de dieren opnimme, binne de basale konsintraasjes fan 'e metaboliteiten tichtby presiaasjewilden. De skriuwers levere it patroan fan neurochemyske feroaringen oan dy sjoen nei de administraasje fan DA-receptorblokkers (Zetterström et al., 1984; Imperato en DiChiara, 1985) en hawwe beoardield dat de steat fan seksuele ynaktiviteit troch prolaktineferliening mediatisearre wurde kin, dat kin wêze as in "endogenous neuroleptysk" (Mas et al., 1995a,b,d). It is dúdlik dat neuroleptyske administraasje begeliedt wurdt troch ferheegingen yn ekstrazellulêre metabolite konsintraasjes en DA efflux (Zetterström et al., 1984; Imperato en DiChiara, 1985). Helaas, Mas et al. (1995a,b) koe de mPOA DA-konsintraasjes net fêststelle. Yn 'e hjoeddeistige stúdzje kamen DA konsintraasjes yn' e NAC werom nei precopulation wearden, wylst DOPAC en HVA konsintraasjes bliuwt. Dit patroan is net inkonsistint mei in rol foar in endogenous neuroleptyske aksje yn 'e NAC om seksueel sêdens te stimulearjen.
Mei it each op it belutsen fan mesolimbyske DA neurons yn motivearre gedrach (Fibiger en Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) en har gefoeligens oan romtlike miljeu stimules (Fabre et al., 1983; Schultz, 1992; Mirenowicz en Schultz, 1994), de beoardielde ferheging fan ekzelletellige konsintraasjes fan NAC DA yn antwurd op de roman froulik binne konsistint mei de hypoteze dat aktiviteiten yn dit DA-systeem wichtich is foar it opnimmen fan seksueel gedrach. Dêrnjonken fertsjinnet rapporten fan appetitele en ferminderingen yn DA-transmissiaasje (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) en neuroanale aktiviteit (Shimura et al., 1994) yn it mPOA fan manlike rotten ûnder seksylik gedrach suggerearje dat dizze struktuer ek bydrage kin oan fernijde kopulaasje-karakteristyk fan 'e Coolidge-effekt.
Neffens in algemiene rol foar it mesolimbyske DA-systeem yn motivearre gedrach, is it goed fêstige dat ekstrazellulêre konsintraasjes fan DA ek foar foardielen, tidens, en fuortendaliks nei konsumpsje fan in miel, mei weromkommen nei baseline wearden ~30 minne letter (Wilson et al., 1995). It is goed bekend dat sêdens troch it iten beynfloede wurdt beynfloede troch syn sensoryske eigenskippen. Minsken en bisten tekenje it iten dat se fiede oan sêd en fûgde oare fiedings dy't net iten hiene (Rolls, 1986). Dit soarget de fraach as der ek extrazellulêre DA-efflux yn 'e NAC selektearber wurde sil wurde troch de presintaasje fan in roman soarte fan iten, mar net troch iten, dat koartlyn brûkt wurdt oan sêne op in manier as analogysk oan dy dat yn' e hjoeddeiske stúdzje berikt is yn it ramt fan seksuele motivaasje. As befestige, sil dit algemiene ferhâlding tusken de sensoryske eigenskippen fan natuerlike beleanning, sattigens, en mesolimbyske DA-transformaasje in krityske rol foar dit neurologyske system ynsette yn 'e regeling fan motivierende prosessen, wêrtroch't har skea litten kin foar serieuze besunigingen fan ite en seksueelfunksje .
REFERINSJES
- ↵
- Beach FA,
- Jordan L
(1956) Seksuele ferheffing en herinnering yn 'e manlike rat. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Temporale skaaimerken fan 'e Coolidge-effekt yn' e manlike ratsje oerlibjende gedrach. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Dopamine funksjonearret yn appetitele en ferdigeningsgedrach. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Seksuele gedrach fergruttet dopamyntromming yn 'e nucleus accumbens en striatum fan manlike rotten: fergeliking mei nijsgroep en lokomotyf. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Aktiviteit fan neurons yn 'e ventral tegmentale regio fan' e behannende monkey. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Belofte, motivaasje, ferneatiging: psychobiology fan mesotelesephalyske dopamynsystemen. Yn 'e hânboek fan' e fysiology: it nervensysteem IV, eds Bloom FE, Geiger SD (Amerikaanske Physiology Society, Bethesda, MD), pp 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Elektrysk stimulearring fan belestingsplakken yn 'e ventral tegmentale gebiet fergruttet dopamyntransport yn' e kearn accumbens fan 'e rat. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Effekten fan stimulearre fariant op seksueel satsjasje yn 'e manlike rat. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Fariaasje yn stimulearre stimulus en seksueel gedrach yn 'e manlike rat. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Feroaringen yn monoamine-wikseling yn foarboarnengebieten dy't ferbûn binne mei manlik seksueel gedrach: in mikrodalysis-stúdzje. Brain Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) In IBM-PC basearre databanksysteem foar it opnimmen fan seksuele gedrach fan naget en foar algemiene eventuele opname. Physiol Behav 44: 825-828.
- ↵
- Hull EM,
- Eaton RC,
- Moses J,
- Lorrain DS
(1993) Kopulaasje fergruttet dopamynaktiviteit yn 'e mediale preoptyske gebiet fan manlike rotten. Life Sci 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Ekstrazellulêre dopamine yn 'e mediale preoptyske gebiet: gefolgen foar seksuele motivaasje en hormonale kontrôle fan' e ferwaging. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Dopamine release en metabolisme yn wakker rten nei systemyske neuroleptika as studearre troch trans-striatal dialysis. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Yndividueel erkenning fan froulike hamsters troch manlju: rol fan gemyske cues en fan de olfaktoryske en vomeronasale systemen. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Ynduksje fan in olfaktorienspeak troch de aktivearring fan in metabotropyske glutamate receptor. Wittenskip 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Hooks MS
(1993) De fermakologyske en neureleaze skeakel fan sensibilisaasje oan psychostimulâns. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Mesocorticolimbyske dopaminergyske neuronen. Funksjonele en regeljende rollen. yn Psychopharmacology: de fjirde generaasje fan progress, eds Bloom FE, Kupfer DJ (Raven, New York), pp 283-294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Seksearjende oalfaktoryske stimules stimulearje in selekteare ferheging fan dopamine frijlitting yn 'e kearn accumbens fan manlike rotten. Brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) Fergrutsje dopamine frijlitting yn 'e kearn accumbens fan kapulearjende manlike rotten as bewiisd troch yn in vivo voltammetry. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernández-Vera JR,
- Gonzalez-Mora JL
(1995a) Neurochemyske korrelat fan seksuele ferheffing en herberch as beoardiele troch yn in vivo mikrodialysis. Brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Voltammetric en mikrodialysis-kontrôle fan brain-monoamine-neurotransmitter frijlitting yn sosjeksoaze ynteraksjes. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Ynduksje fan ferlingde gedrach troch apomorphyn yn seksueel sitten ratten. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
(1995d) De neurochemyme fan seksuele satsje. In eksperimintele model fan ynsette winsk. yn The Pharmacology fan seksuele funksje en dysfunksje, ed Bancroft J (Raven, New York), pp 115-126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Beheining fan ûnfoarspelberens foar leanprizen yn prime dopaminergyske neuronen. J neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Mesolimbyske dopamine frijlitting ûntstiet troch aktivearring fan it accessory olfaktory systeem: in hege rapper chronoamperometryske stúdzje. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) It rat-brain yn stereotaksyske koördinaten (2nd ed). (Akademysk, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Seksuele gedrach ferbettert sintrale dopamine-transmission yn 'e manlike rat. Brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) De psychopharmakology fan seksueel gedrach. yn Psychopharmacology: de fjirde generaasje fan progress, eds Bloom FE, Kupfer DJ (Raven, New York), pp 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioiden en seksueel gedrach. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Neurobiologyske korrelaten fan positive emosjonele steaten: dopamine, anticipaasje en lean. yn Ynternasjonaal oersicht fan stúdzjes oer emoasje, ed Strongman (Wiley, New York), pp 31-50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Korrelaasje fan dopamine frijlitting yn 'e kearn accumbens mei manlik seksueel gedrach yn rat. Brain Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Ferfal fan seksueel ferlies troch serotonergyske en noradrenergyske aginten. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Partisipaasje fan it sintraal noradrenergysk systeem yn 'e werhelling fan' e copulatory gedrach fan seksueel ferlernde ratten troch yhimbine, naloxon, en 8-OH-DPAT. Brain Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Opioide antagonisten en it seksueel sazjittings fenomenon. Psychopharmacol 122: 131-136.
- ↵
- Rolls BJ
(1986) Sensory-spesifike sfear. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) De gedrachske neurochemystik fan motivaasje: methodologyske en konseptuele problemen yn stúdzjes oer de dynamyske aktiviteit fan 'e nucleus accumbens dopamine. J Neurosci-metoaden 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Dopamine frijlitte yn 'e mediale preoptyske gebiet yn' e manlike oerlibjende gedrach yn rat. Brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Aktiviteit fan dopamin neuroanen yn 'e behavingpriis. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) It meditaal foar preektikaal gebiet is belutsen by sawol sike-opwaaks en prestaasjes yn manlike rotten: re-evaluaasje fan neuronaktiviteit yn frijhawwende bisten. Brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Dopamine-transmysjestaasje nimt yn 'e kearn accumbens fan manlike rotten yn har earste eksposysje nei seksueel ûntfankende manlike rotten. Brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Dopaminergyske korrelat fan motivearre gedrach: belang fan rydbewiis. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Beach FA
(1963) Modifikaasje yn it seksueel gedrach fan manlike rotten dy't makke wurde troch it feroarjen fan de stimulus froulju. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Effekt fan neuroleptyske drugs op striatal dopamine frijlitting en metabolisme yn 'e wekker rat dy't troch intracerebral dialysis studearre. Eur J Pharmacol 106: 27-37.
Artikelen dy't dit artikel befetsje
- Endogenous opioïde-yndirekte neuroplastyk fan dopaminergyske neurons yn 'e Ventral Tegmental-gebieten ynfloeden Natuerlike en opiate betrekking Journal of Neuroscience, 25 juny 2014, 34 (26): 8825-8836
- Unsyklikheid yn 'e gefoeligens foar ferskillende soarten belangen yn it pasypale spiel Brain, 1 augustus 2013, 136 (8): 2527-2538
- Natuer- en Drug Rewards Act op mienskiplike neurologyske plastikaasjemeganisme mei {Delta} FosB as Key Key Mediator Journal of Neuroscience, 20 febrewaris 2013, 33 (8): 3434-3442
- Neuronale reaksjes yn 'e Nucleus Accumbens Shell yn' e seksuele gedrach yn Male Rats Journal of Neuroscience, 1 febrewaris 2012, 32 (5): 1672-1686
- Romantyske leafde: in sûchhealsysteem foar mate-keuze Philosophyske transaksjes fan 'e Royal Society B: Biologyske wittenskippen, 29 Desimber 2006, 361 (1476): 2173-2186
- Hedonyske spots yn 'e harsens De neuroscientist, 1 desimber 2006, 12 (6): 500-511
- Modulaasje troch Sintrale en Basolaterale Amygdalar Nuclei fan Dopaminergyske Korrelaten fan Fiedingen nei Sosjity yn 'e Rat Nucleus Accumbens en Medial Prefrontal Cortex Journal of Neuroscience, 15 Desimber 2002, 22 (24): 10958-10965
- Frequency of Dopamine Concentration Transients Increases in Dorsal and Ventral Striatum of Male Rats during Introduction of Conspicifics Journal of Neuroscience, 1 Desimber 2002, 22 (23): 10477-10486
- Book Review: Dynamika fan ekstrazellulêre dopamine yn 'e akute en chronike aksjes fan kokaïne De neuroscientist, 1 augustus 2002, 8 (4): 315-322
- Differinsjale ynvolvement fan NMDA, AMPA / Kainate, en Dopamine Receptors yn 'e Nucleus Accumbens Core yn' e oankeap en prestaasjes fan 'e Pavlovian Approach Behavior Journal of Neuroscience, 1 Desimber 2001, 21 (23): 9471-9477
- Kontrôle fan kokaïngewaasje fan medisinen-assosjearre stimulâns yn raten: Effekten op it hersteljen fan ferwiderjende operant-reageare en ekzelletellere dopamynnivo's yn amygdala en nucleus accumbens PNAS, 11 april 2000, 97 (8): 4321-4326
- Fergrutting fan seksuele gedrach en ferhege Dopamine Efflux yn 't Nucleus Accumbens fan mannelike raten nei D-Amphetamine-yndirekte bewuste sensibilisaasje Journal of Neuroscience, 1 jannewaris 1999, 19 (1): 456-463
- NEUROSCIENCE: De oandacht fan 'e harsens krije Science, 3 oktober 1997, 278 (5335): 35-37
- Dopaminergyske korrelaten fan sensory-spesifike saterij yn 'e mediale prefrontale kortax en nucleus Accumbens fan' e rat Journal of Neuroscience, 1 oktober 1999, 19 (19): RC29