OPMERKINGEN: As lettere stúdzjes sille iepenbiere is DeltaFosB de mienskiplike molekulêre skeakel foar sawol drugs- as gedrachsferslaving. It is in transkripsjefaktor dat betsjut dat it beynfloedet op hokker genen wurde yn- of útskeakele. Lykas earne oars sein, ferslave ferslaavjende medisinen allinich normale meganismen. Dêrom is it dom om oan te jaan dat gedrachsferslaving net kin bestean.
Proc Natl Acad Sci US 2001 septimber 25; 98 (20): 11042-11046.
doi: 10.1073 / pnas.191352698.
Eric J. Nestler *, Michel Barrot, en David W. Self
Department of Psychiatry and Center for Basic Neuroscience, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390-9070
Abstract
De longevity fan guon fan 'e gedrachsûntstekkingen dy't karakteristike drugsoerwinning tekenje hat bepaald dat regeljouwing fan neuronale geneel ekspresje kin behannele wurde yn it proses dêr't medisinen fan misbrûk in steat fan ferslaving feroarsaakje. ikFerklearjende bewiis suggerearret dat de transkriptyffaktor ΔFosB ien maatregel presintearret wêrby't drugs fan misbrûk relatyf stabige feroaringen produkt meitsje dy't it bydrage oan 'e oanfreegjen fenotype. ΔFosB, in lid fan 'e Fosfamylje fan transkripteursfaktoren, sammele binnen in subset fan neurons fan' e nucleus accumbens en dorsal striatum (hynsteegrien wichtich foar ferslavings) nei repetitive administraasje fan in protte soarten drugs fan misbrûk. Ferlykbere akkumulation fan ΔFosB komt nei compulsive rinningen, dy't bepaald dat ΔFosB akseptearje kin yn antwurd op in protte soarten twangend gedrach. Importantly, ΔFosB bliuwt yn neurons foar relatyf lange tiden fanwege syn ekstraordinêre stabiliteit. Dêrom stiet ΔFosB in molekulêre meganika dy't inisjalisearje en dan feroarings yn 'e gene ekspresje hâlde dy't langer nei de medyske eksposysje stopje. Studies yn ynfoedige transgene mûzen dy't ΔFosB of in dominante negatyf ynhibitor fan 'e proteïne eksperimintearje, jouwe direkte beweechingen dat ΔFosB in hurde sensibiliteit feroarsaket foar de gedrachske effekten fan drugs fan misbrûk en, eventueel, ferhege medisyngenoatskip. Dit wurk stipet de opfetting dat ΔFosB funksjonearret as in soarte fan "molekulêre skeakel" dy't allinich akute medisynaktive reaksjes yn relatyf stabile oanpassingen feroaret dy't in bydrage leverje oan 'e lange termynlike neuri- en gedrachske plastikaasje dy't ûnderdrukking is.
Undersykûndersyk is rjochte op it fersterkjen fan 'e komplekse wizen wêryn drugs fan misbrûk it hert feroarsaakje om gedrachsferoarings te feroarjen dy't de sukses karakterisearje. Ien fan 'e kritike útdagingen op it fjild is om relatyf stabile drug-induced feroaringen yn it harsens te identifisearjen om te rekkenjen foar dy gedrachsûntstekken dy't benammen lang libbe binne. Bygelyks, in minsklike fersoarging kin op it ferheegde risiko wêze foar it werombringen sels nei jierren fan ûnôfhinklikheid.
De stabiliteit fan dizze gedrachske abnormaliteit hat liede ta de suggestje dat se mediatisearre binne, teminsten, troch feroaringen yn 'e gene ekspresje (1-3). Neffens dizze werjefte wurdt it werhellende eksposysje fan in medisyn fan misbrûk repeatedly fertsjintwurdigje op ünnoedige synapses yn it harsens dat gefoelich binne foar it drugs. Sokke ferrifelingen úteinlik sjogge fia yntrazellulêre boadskippen kaskaden nei de kearn, wêr't se earst inisjearje en dan feroaringen yn 'e ekspresje fan spesifike genen hâlde. In primêre meganisaasje dêr't trochsykling transduksjegriningen ynfloed hawwe foar gene ekspresje is de regeling fan transkriptyfokraten, protinen dy't binden oan regeljende regio's fan genes en feroarjen fan har transkripsje.
Ien doel fan ûndersyksûndersyk is dus om transkriptiefokters te identifisearjen dy't feroare wurde yn harsensregio's dy't yn bedrige hawwe nei chronike administraasje fan drugs fan misbrûk. Ferskillende sokk transkripteurs fûnen binne yn 'e lêste desennia identifisearre (1-6). De fokus fan dizze resinsje is op in bepaalde transkripsjefaktor neamd ΔFosB.
Ynduksje fan ΔFosB troch drugs fan misbrûk
ΔFosB, kodearre troch it fosB-gen, is in lid fan 'e Fos-famylje fan transkripsjefaktoren, dy't ek c-Fos, FosB, Fra1, en Fra2 (7) binne. Dizze Fos-famyljeproteinen heterodimerisearje mei Jun famyljeproteinen (c-Jun, JunB of JunD) om aktive AP-1- (aktivator-protein-1) transkripteursfakten te bannen dy't binnen AP-1-siden biede (konsensus-folchoarder: TGAC / GTCA) yn de promotoren fan bepaalde genen om har transkripsje te regeljen.
Dizze famyljeproteinen fan Fos wurde rappe en transiental yn spesjale harsensregio's yndirekt nei acute administraasje fan in protte drugs fan misbrûk (foto 1) (8-11). Promininte regio's binne de nucleus accumbens en dorsal striatum, dy't wichtige mediators binne fan gedrachskontroleiten op 'e medisinen, benammen har loyendend en locomotor-aktivearjende effekten (12, 13). Dizze proteins komme werom nei basaalnivo binnen oeren fan drugsadministration.
Figure 1
Scheme wêrby't de graduale accumulation fan ΔFosB tsjin de snelle en transiente yndeksje fan oare Fos-famyljeproteinen as antwurd op drugs fan misbrûk. (A) It autoradiogram ferleart de ferskillende yntuksje fan dizze ferskate proteins troch akute stimulaasje (1-2 hr nei in iennichste medisynbelesting) tsjin chronike stimulearring (1 dei nei werhelle medisynbelesting). (B) Ferskillige wellen fan Fos-eauwe protten [bestiet út c-Fos (52- oant 58-kDa isoformen), FosB (46- oant 50-kDa isoformen), ΔFosB (33-kDa isoform), en Fra1 of Fra2 ( 40 kDa)] wurde yn nukleus accumbens en dorsale striatalneuronen feroarsake troch akute administraasje fan in medisyn fan misbrûk. Ek wurde induktyf biochemysk modifisearre isoformen fan ΔFosB (35-37 kDa); Se wurde ek oanwêzich (hoewol op leech nivo) nei acute drug adminisaasje, mar bestege yn langere perioaden op har stabyl. (C) By repetearje (bygelyks, twa kear deistich) drugs administraasje, draait eltse stimulus in leech nivo fan 'e stabile ΔFosB-isoformen, dy't oanjûn is troch de legere set fan oerlappende linen dy't oantsjutte dat ΔFosB troch elke akute stimulearre wurdt. It resultaat is in graduele ferheging fan 'e totale nivo's fan ΔFosB mei werhelle stimuli yn' e rin fan in chronike behanneling, dy't oanjûn wurdt troch de tanimmende stappe line yn 'e grafyk.
Sels ferskillende antwurden sjogge nei chronike administraasje fan drugs fan misbrûk (foto 1). Biochemysk modifisearre isoformen fan ΔFosB (molekulêre massen 35-37 kDa) sammelje binnen deselde harsensregio's nei repetearjende medisynbelesting, wylst alle oare Fos-famyljeleden tolerânsje sjen (dat is, reduzearre ynduksje yn fergeliking mei earste drug-eksposysjes). Sokke accumulation fan ΔFosB is beoardield foar kokaïne, morphine, amphetamin, alkohol, nicotine, en phencyclidinee (11, 14-18). Der is guon bewiis dat dizze yndeksearring selektyf is foar de dynorphine / substân P-hantlieding fan submersedelike middelste neurons yn dizze harsensregio's (15, 17), hoewol mear wurk is nedich om dit mei wissigens te fêstigjen. De 35- oant 37-kDa-isoformen fan ΔFosB dimmerize mei JunD foar in aktyf en langstige AP-1-kompleks yn dizze harsensregio's (19, 20). Dizze ΔFosB-isoformen accumulearje mei chronike medisynbelesting fanwege har úteinlik lange halve libben (21), en dêrtroch yn 'e neurons op syn minst in pear wike nei it besykjen fan drugsadministraasje ynhâlde. It is ynteressant om te notearjen dat dizze ΔFosB-isoformen tige stabile produkten fan in direkte begjingen (fosB) binne. De stabiliteit fan de ΔFosB-isoformen biedt in roman molekulêre meganis wêrby't drug-induced feroaringen yn gene ekspresje hingje kinne fan ferhâlding nettsjinsteande relatyf lange termyn fan drugswinning.
Hoewol de nucleus accumbens in krityske rol spilet yn 'e lodzjende effekten fan drugs fan misbrûk, wurdt leaude normaal funksjonearje troch regeljende reaksjes op natuerlike reinforcers, lykas iten, drinken, seks en sosjale ynteraksjes (12, 13). As gefolch dêrfan is der in protte belangstelling foar in mooglike rol fan dizze harsensregio yn oare compulsive gedrach (bygelyks pasyklike oerwinning, spielings, oefening, ensfh.). Hjirtroch hawwe wy ûndersocht oft ΔFosB regele wurdt yn in diermodel fan twangrinnen. De stabile 35- oant 37-kDa-isoformen fan ΔFosB wurde selektearre indirekt binnen de nucleus accumbens yn raten dy't twangende ferhâldingsgedrach sjen. †
Biochemyske Identiteit fan Stabile ΔFosB Isoformen
As hjirboppe neamd, binne de ΔFosB-isoformen dy't nei chronike administraasje fan in drugs fan misbrûk of compulsive rinnen sjen litte sjen in molekulêre massa fan 35-37 kDa. Se kinne differinsjearre wurde fan 'e 33-kDa-isoform fan' e ΔFosB dy't rapper, mar transiental nei ien inkelde medisynbelesting (1) (14, 19, 22) yntreekt. Aktuele bewiis leit oan dat it 33-kDa isoform is de natuerlike foarm fan it proteïn, dat feroare is om de stabile 35- nei 37-kDa-produkten te foarmjen (19, 21). De natuer fan 'e biochemyske modifikaasje dy't de ynstabele 33-kDa-isofoarm konvertearret yn' e stabile 35- oant 37-kDa-isoformen is dúdlik bleaun. It is spekulearre dat phosphorylaasje ferantwurdlik wêze kin (11). Bygelyks, yndeksearring fan ΔFosB wurdt fersmiten yn mieren dy't gjin DARPP-32 hawwe, in striatal-ferrike proteïne (23, 24). Omdat DARPP-32 de katalytyske aktiviteit fan 'e proteïne-phosphatase-1- en proteine kinase A (25, 26) regulearret, is de needsaak foar dit protein foar de normale akkumulaasje fan' e stabile ΔFosB-isoformen in mogelike rol foar fosforaasje yn 'e generaasje fan dizze stabile produkten.
Rol fan ΔFosB yn Behavioral plasticity oan drugs fan misbrûk
Ynsjoch yn 'e rol fan ΔFosB yn drugsoerwinning kaam foar in grut part út it ûndersyk fan transgenen mûzen wêryn ΔFosB selektearjend indirekt binnen de kearn accumbens en oare striatalregio's fan folwoeksen dieren (27, 28) yndirekte wurde. Importantly, dizze mice oerskriuwe ΔFosB selektearjend yn 'e dynorphine / substâns P-hanthavenige medium-spiny neuroanen, wêr't de medisinen binne leauwe dat it eauwyn oansette. De gedrachse phenotype fan 'e ΔFosB-overexprimearjende mûs, dy't op ferskate manieren as tûzen nei chronike medisynbelesting liket, is gearfette yn tabel 1. De mice sjogge opfolger fan locomotor nei kokaïne nei acute en chronike administraasje (28). Se sjogge ek fersterkende gefoeligens foar de leukende effekten fan kokaïne en morfine yn plattelânskonstruksjes (11, 28) en sille sels in koekje dosken adminisearje as littermaten, dy't gjin uterlike orefesje ‡ FosB ‡ yngeane. Dizze dieren sjogge normaal kondisearre locomotor Sensibilisaasje foar kokaïne en normaal romtlik learen yn it Morris wettermaze (28). TDizze gegevens jouwe oan dat ΔFosB de gefoelichheid fan in dier fergruttet foar kokaïne en miskien oare drugs fan misbrûk en kin in meganisme foarstelle foar relatyf langere sensibilisaasje foar de medisinen.
 striatum
striatum
| Ferhege lokomotor aktivearring yn antwurd op akute en werhelle kokaineadministration. |
| Ferhege lokkige antwurden op kokaïne en morphyn yn plakken- kondysjesûndersiken. |
| Fergrutte selsbehearsking fan lege dosinten fan kokaïne. |
| Fergrutsje motivaasje foar kokaïne yn progressive ratio-assays. |
| Ferhege anxiolytyske responsen oan alkohol. |
| Ferwachte compulsive aktive gedrach. |
Op grûn fan gegevens yn refs. 28 en 29.† ‡ §¶
Behaviorale plastikaat troch mediated troch ΔFosB yn kearn accumbens-dorsal striatum
IIn oanfolling is der foaral bewiis dat de effekten fan ΔFosB safolle as in regeljouwing fan drugsemensitivitaasje sjogge om se mear kompleksere gedrachs oangeande de ferslavingsproses. Mysen ekspresearjen ΔFosB wurkje hurder om self-administraasjeassays suFergiftigje dat ΔFosB petearje kin oan it stimulearjen fan motiveare eigenskippen fan kokaïne en dêrtroch liede ta in oanstriid foar weromreis nei medisinenl. ‡ ΔFosB-ekspresjonele messen fertsjinje ek ferbettere anxiolytyske effekten fan alkohol, § in fenotype dy't ferbûn is mei ferhege alkoholfeiligens yn 'e minske. Mei-elkoar sjogge dizze frede befiningen dat ΔFosB, neist de ferheging fan sensibiliteit foar drugs fan misbrûk, kwalitative feroarings yn gedrach makket dy't drugs sykje fan gedrach. Sa kin ΔFosB as in "molekulêre skeakel" funksjonearje dy't helpt te begjinnen en dêrnei krúsjale aspekten fan 'e fersmoarge state. In wichtige fraach ûnder aktuele ûndersiik is, oft ΔFosB akkumulaasje yn 'e medisineel-eksposysje befoarderet dat medisinen nei ferlingde perioaden, sels nei ΔFosB-nivo's normalisearre (sjoch hjirûnder).
Folwoeksen mûzen dy't erefeksje ΔFosB selektearjend binnen de nucleus accumbens en dorsal striatum ek in gruttere compulsive rinnen sjen litte yn fergeliking mei kontrôle littermaten. † Dizze beoardielingen lûke de ynteressante mooglikheid dat ΔFosB accumulation yn dizze neurons in mear algemiene rol tsjinnet yn 'e formaasje en ûnderhâld fan gewoantehears en compulsive Verzorgingen, miskien troch it fersterkjen fan 'e effektiviteit fan neurologyske circumsjiten dêr't dy neuroanen funksjonearje.
ΔFosB accumulearret yn beskate harsensregio's bûten de nucleus accumbens en dorsale striatum nei chronike eksposysje foar kokaïne. Prominint ûnder dy regio 's binne de amygdala en medial prefrontal cortex (15). In wichtich doel fan hjoeddeistich ûndersyk is om de bydragen fan ΔFosB yndeksearring yn dizze regio te begripen oan 'e oanfreegjen fenotype.
Earder wurke oan fosB-knockoutmûzen die bliken dat dizze bisten gjin sensibilisaasje ûntwikkelje foar de lokomotoryske effekten fan kokaïne, wat konsistint is mei de befiningen fan 'e hjirboppe neamde ΔFosB-overexpressing mûzen (22). De fosB-mutanten lieten lykwols ferbettere gefoelichheid foar akute effekten fan kokaïne sjen, wat net konsekwint is mei dizze oare befiningen. Ynterpretaasje fan befiningen mei de fosB-mutanten is lykwols komplisearre troch it feit dat dizze bisten net allinich ΔFosB, mar ek FosB yn folsleine lingte misse. Boppedat misse de mutanten beide proteïnen yn 't heul en fan' e ierste stadia fan ûntwikkeling. Yndie, mear resint wurk stipet konklúzjes fan 'e ΔFosB-overexpressing mûzen: induzibele overexpression fan in trunearre mutant fan c-Jun, dy't fungeart as in dominante negative antagonist fan ΔFosB, selektyf yn nucleus accumbens en dorsale striatum toant fermindere gefoelichheid foar de beleanjende effekten fan kokaïne .¶ Dizze befiningen beklamje de foarsichtigens dy't moatte wurde brûkt by it ynterpretearjen fan resultaten fan mûzen mei konstitutive mutaasjes en yllustrearje it belang fan mûzen mei induzibele en selstypespesifike mutaasjes yn ûndersiken fan plastisiteit yn it folwoeksen brein.
Target Genes foar ΔFosB
Omdat ΔFosB in transkripsjefaktor is, is it wierskynlik dat it proteïn fersteane plastykens troch feroaringen yn 'e ekspresje fan oare genes. ΔFosB wurdt generearre troch alternative splitsing fan 'e fosB-gene en fynt in diel fan' e C-terminaltransaktivaasje-dome yn folsleine FosB. As gefolch waard it oarspronklik útsteld dat ΔFosB funksjonearret as transkriale repressor (29). It wurk yn 'e sel kultuer hat lykwols dúdlik dat it ΔFosB kin slaan of útdrukke AP-1-mediïnte transkripsje ôfhinklik fan de spesifike AP-1-webside brûkt (21, 29-31). Folsleine-lingende FosB eksertearret deselde effekten as ΔFosB op bepaalde promotorfragminten, mar ferskate effekten op oaren. Fierder wurk is nedich om de meganismen te begripen dy't dizze ôfwikende aksjes fan ΔFosB en FosB te krijen hawwe.
Us groep hat twa oanpak brûkt om doelgenen te identifisearjen foar ΔFosB. Ien is de kandidaat-genoanpak. Wy beskôgen yn earste ynstânsje α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) glutamatreceptors as putative doelen, sjoen de wichtige rol fan glutamatergyske oerdracht yn 'e nucleus accumbens. Wurk oant no ta hat oanjûn dat ien bepaalde AMPA-glutamatreceptorsubunit, GluR2, in bona fide doel kin wêze foar ΔFosB (ôfb. 2). GluR2-ekspresje, mar net de ekspresje fan oare AMPA-receptor-subunits, wurdt ferhege yn nucleus accumbens (mar net dorsale striatum) by overekspresje fan ΔFosB (28), en ekspresje fan in dominante negative mutant ferswakket it fermogen fan kokaïne om it proteïne te induzearjen .¶ Derneist befettet de promotor fan it GluR2-gen in konsensus AP-1-side dy't ΔFosB bindet (28). Overekspresje fan GluR2 yn 'e nucleus accumbens, troch gebrûk fan virale mediïnteare gene-oerdracht, fergruttet de gefoelichheid fan in dier foar de beleanjende effekten fan kokaïne, wêrtroch't in diel fan it fenotype neigiet yn' e ΔFosB-ekspresjearjende mûzen [28]. Ynduksje fan GluR2 koe rekken hâlde mei de fermindere elektrofysiologyske gefoelichheid fan nucleus accumbens neuroanen foar AMPA-receptor agonisten nei chronike kokaïne-administraasje [32], om't AMPA-receptors mei GluR2 sjen litte fermindere algemiene konduktânsje en fermindere Ca2 + permeabiliteit. Reduzearre responsiviteit fan dizze neuroanen op opwinende yngongen kin dan antwurden op in drug fan misbrûk ferbetterje. De manieren wêrop dopaminergyske en glutamatergyske sinjalen yn nucleus accumbens ferslavend gedrach regelje bliuwe lykwols ûnbekend; dit sil in neuronale sirkwinsjoneel nivo fereaskje, dat is noch net beskikber.
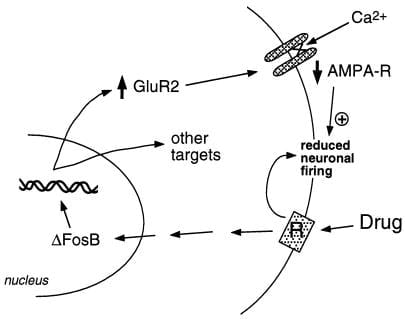
Figure 2
De AMPA glutamatreceptorsubunit, GluR2, is in putatyf doelwyt foar ΔFosB. Te sjen is hoe ΔFosB-bemiddele ynduksje fan GluR2 de fysiologyske responsens fan 'e nucleus accumbens neuroanen kin feroarje en liede ta sensibiliseare antwurden op drugs fan misbrûk. Neffens dit skema produsearje misbrûk medisinen har akute fersterkjende effekten fia remming fan neuronen fan nucleus accumbens. Mei werhelle eksposysje feroarsaakje de medisinen ΔFosB, dy't in soad doelgenen regelt, ynklusyf GluR2. Dit fergruttet it oanpart fan AMPA-receptors (AMPA-R) op nucleus accumbens neuroanen dy't de GluR2-subunit befetsje, wat feroarsake algemiene AMPA-stream en ferlege Ca2 + -stream. Dizze ferlege eksitabiliteit koe de neuroanen gefoeliger meitsje foar de akute remmende effekten fan 'e medisinen en dêrtroch foar de fersterkjende effekten fan' e medisinen.
In oar putative doel foar ΔFosB is de gene foar kodearring dynorphyn. As earder ferklearre, wurdt dynorphin yndield yn 'e subset fan nukleus accumbens medium lintsjes neuronen dy't induksje sjen litte fan ΔFosB. Dynorphyn ferskynt yn in ynterzellulêre feedback-loop: syn release ferbynt de dopaminergyske neurons dy't de medium-spiny-neuroanen, fia κ opioide-receptors yndoneare op dopaminergyske nerve-terminalen yn 'e nucleus accumbens en ek op selskippen en dendrites yn it ventral tegmental gebiet (Fig. 3) (33-35). Dit idee is konsekwint mei de kapasiteit fan in κ-reptoragonist, nei administraasje yn ien fan dizze twa harsensregio's, om drugs rewar te feroarjend (35).
REenswurk hat oanjûn dat ΔFosB de ekspresje fan dynorphin nimt, "dy kin bydrage oan it ferheegjen fan lieningsmeganismen dy't te sjen binne mei ΔFosB-yndeksje. Interestingly, another drug-regulated transcription factor, CREB (cAMP response element binding protein) (2, 3) exerts the opposite effect: it induces dynorphin expression in the nucleus accumbens and reduces the lucking properties of cocaine and morphine (4). **
BEkstra drugs-aktive activaasje fan CREB ûntlient fluch nei medyske administraasje, lykas rezjypulaasje fan dynorphin troch CREB en ΔFosB kin de omkearde gedrachsferoarings feroarsaakje, dy't yn 'e begjin en letste faze fan weromlûking foarkomt, mei negative emosjonele symptomen en fergrutting fan drugsensifisiteit yn' e frjemde fazen fan weromlûking, en sensibilisaasje oan 'e lodzjende en stimulearjende motivearjende effekten fan medisinen dy't foar spesjale punten punten binne.
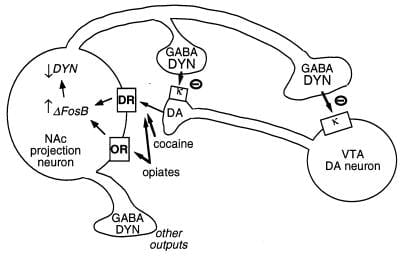
Figure 3
Dynorphyn is in putative doel foar ΔFosB. Sprekke is in ventral tegmentale gebiet (VTA) dopamine (DA) neuron dy't in klasse fan kearn accumbens (NAc) GABAergysk projektneuron útdrukt dy't dynografen foarmet (DYN). Dynorphyn tsjinnet in feedbackmeganisaasje yn dizze skeakel: dynorphine, út 'e terminen fan' e NAc neuronen, útfiert op κ opioidreptors dy't op nerve terminals en cellgenen fan 'e DA-neurons sitte om har funksjonearjen te ynsette. ΔFosB, troch ynstekking fan dyn ekspresje fan dynorphin, dizze reaksje-loop del te regeljen en de belibjende eigenskippen fan misbrûk fan drugs te ferbetterjen. Net wer te sjen is it ferspriedlike ynfloed fan CREB op dit systeem: CREB fersterket de ekspresje fan dynorphyn en dêrtroch fertsjinnet de belibjende eigenskippen fan drugs fan misbrûk (4). GABA, γ-aminobutyric acid; DR, dopaminreceptor; OR, opioïde receptor.
De twadde oanpak dy't wurdt brûkt om doelgenen te identifisearjen foar ΔFosB omfettet DNA-microarray-analyse. Yndukteare overexpresje fan ΔFosB fergruttet of fermindert de ekspresje fan ferskate genen yn 'e nucleus accumbens (36). Hoewol in soad wurk no nedich is om elk fan dizze genen te validearjen as fysiologyske doelen fan ΔFosB en om har bydrage te begripen foar it ferslavingsfenotype, liket ien wichtich doel Cdk5 te wêzen (cyclin-ôfhinklike kinase-5). Sadwaande waard Cdk5 yn earste ynstânsje identifisearre as ΔFosB-regele troch gebrûk fan mikroarrays, en letter sjen litten te wurde feroarsake yn kearn accumbens en dorsale striatum nei chronike kokaïne administraasje [37]. ΔFosB aktiveart it cdk5-gen fia in AP-1-side oanwêzich binnen de promotor fan 'e gene (36). Mei-inoar stypje dizze gegevens in skema wêryn kokaïne Cdk5-ekspresje yn dizze harsensregio's induceert fia ΔFosB. Ynduksje fan Cdk5 liket dopaminergyske sinjaasje teminsten diels te feroarjen fia ferhege fosforylaasje fan DARPP-32 (37), dy't wurdt konverteare fan in ynhibitor fan proteïne fosfatase-1 nei in ynhibitor fan proteïnekinase A op syn fosforylaasje troch Cdk5 (26).
Rol fan ΔFosB yn 'e mediatorjende "Permanente" plastysiteit oan drugs fan misbrûk
Hoewol it ΔFosB-sinjaal relatyf lange libbet, is it net fêste. ΔFosB ûntwikkelet stadichoan en kin net langer yn 't bloed fûn wurde nei 1-2 moannen fan drugswinning, ek al bepaalde gedrachsferoaringen bliuwe foar folle langere perioaden. Dêrom soe ΔFosB per se net ferskine moatte dizze semi-ferstannige gedrachsûntstekken te mediatisearjen. De swierrichheid om te finen fan 'e molekulêre oanpassingen dy't de ekstreem stabile gedrachsferoarings dy't ferbân hâlde mei sêftguod binne, fertsjinwurdigje oan de útdagings dy't yn it learen- en ûnthâldfjild binne. Hoewol binne der in elegante cellulêre en molekulêre modellen fan lear- en ûnthâld, hat it oant no ta net mooglik west om molekulêre en selleare oanpassingen te identifisearjen dy't genôch lang libbe binne om rekken te hâlden foar tige stabile gedrachsinnigens. Yndied is ΔFosB de langst libbene oanpassing dy't bekend is yn harsen foar folwoeksenen, net allinich yn antwurd op drugs fan misbrûk, mar ek op elke oare steuring (dy't gjin letsels omfetsje). Twa útstellen binne evoluearre, sawol yn 'e ferslavings- as lear- en ûnthâldfjilden, om dizze ferskil te ferantwurdzjen.
Ien mooglikheid is dat mear transiente wizigings yn 'e gene ekspresje binne, lykas dy troch Mediatisearre troch ΔFosB mediïntearre of oare transkripsjefaktoren (bygelyks CREB) Meitsje mear langere feroaringen yn 'e neuronale morfology en synaptyske struktuer. Bygelyks, In ferheging fan de tichteens fan dendrityske rintjes (benammen in ferheging fan twa-liedende wylden) begeliedt de Hegere efficacy fan glutamatergyske synapses by hippocampal pyramidale neuronen by lange termyn potensaasje (38-40), en parallels de fersterke gedrachensensitiviteit foar kokaïne dy't mediïntearre wurdt op it nivo fan medium lintsje neuronen fan 'e nucleus accumbens (41). It is net bekend oft sokke strukturele wizigingen genôch lange libbet binne om tige stabile feroarings yn gedrach te hâlden, hoewol de lêste foar minstens 1 moanne fan medisinen weromlûke. De lêste beweitsingen beweare de mooglikheid dat ΔFosB, en syn yndeks fan Cdk5, is ien mediator fan drug-induced feroaringen yn synaptyske struktuer yn 'e nucleus accumbens (Abb. 4) .. So kin in infusion fan in Cdk5 ynhibitor yn' e nucleus accumbens prevents Feardigens fan repetitive kokainebelesting om dendrityske rintensichte yn dizze regio te ferheegjen. Dit is konsekwint mei de besicht dat Cdk5, dy 't yn it harsen ferrike wurdt, reguliere neuronale struktuer en groei (sjoch refs 36 en 37). It is mooglik, hoewol net te bewizen is, dat sokke feroarings yn 'e neuronale morphology kinne it ΔFosB-sinjal sels sels útsette.
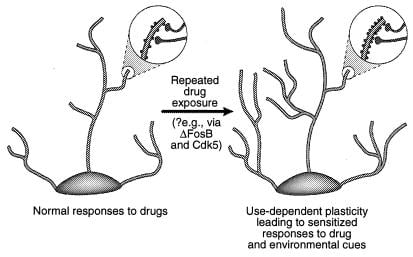
Figure 4
Regulearring fan dendrityske struktuer troch drugs fan misbrûk. Te sjen is de útwreiding fan 'e dendrityske beam fan in neuron nei chronike bleatstelling oan in drug fan misbrûk, lykas is beoardiele mei kokaïne yn' e nucleus accumbens en prefrontale cortex (41). De gebieten fan fergrutting toane in tanimming fan dendrityske stekels, dy't wurdt postulearre om te foarkommen yn kombinaasje mei aktiveare senuwterminals. Dizze ferheging fan dendrityske rintdichtheid kin wurde bemiddele fia ΔFosB en de dêrop folgjende ynduksje fan Cdk5 (sjoch tekst). Sokke feroaringen yn 'e dendrityske struktuer, dy't fergelykber binne mei dy dy't beoardiele binne yn guon learmodellen (bgl. Potensjearring op lange termyn), kinne sensibiliseare antwurden op lang libbens bemiddelje op drugs fan misbrûk as miljeutekens. [Werjûn mei tastimming fan ref. 3 (auteursrjocht 2001, Macmillian Magazines Ltd.)].
In oare mooglikheid is dat de oergeunstige yndeks fan in transkripsjefaktor (bgl. ΔFosB, CREB) liedt ta permaninte feroarings yn 'e gene ekspresje troch de feroaring fan chromatin. Dizze en in protte oare transkripsjefoksen wurde leauwe om de transkripsje fan in doelgenaat te aktivearjen of te fersprieden troch te promoatsjen fan de acetylaasje of deacetylaasje, respektivelik, fan histônes yn 'e buert fan' e gen (42). Hoewol sokke acetylaasje en deacetylaasje fan histones kinne miskien hurder fluggje, is it mooglik dat ΔFosB of CREB langer duorsume oanpassingen yn de enzymatyske masine komme dy't de histoneacetylaasje kontrolearje. ΔFosB of CREB kinne ek langere libbenswizigingen feroarsaakje troch it regeljen fan oare feroaringen fan chromatine (bygelyks DNA of histone-methylaasje) dy't yn 'e permaninte feroaringen fan' e gentrtrografie dy't yn 'e ûntwikkeling foarkomme (sjoch refs 42 en 43) . Hoewol dizze mooglikheden spekulatyf bliuwe, kinne se in meganisme leverje, dêr't transiente oanpassingen oan in medisyn fan misbrûk (of in oar ferwidering) liede ta essensjele libbenslange gedrachske gefolgen.
Referinsjes
- ↵
- Nestler EJ,
- Hope BT,
- Widnell KL
(1993) Neuron 11: 995-1006.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Berke JD,
- Hyman SE
(2000) Neuron 25: 515-532.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Nestler EJ
(2001) Nat Rev Neurosci 2: 119-128.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Carlezon WA Jr,
- Thome J,
- Olson VG,
- Lane-Ladd SB,
- Brodkin ES,
- Hiroi N,
- Duman RS,
- Neve RL,
- Nestler EJ
(1998) Science 282: 2272-2275.
- O'Donovan KJ,
- Tourtellotte WG,
- Millbrandt J,
- Baraban JM
(1999) Trends Neurosci 22: 167-173.
CrossRefliteratuerdatabases MEDLINEWeb of Science
- ↵
- Mackler SA,
- Korutla L,
- Cha XY,
- Koebbe MJ,
- Fournier KM,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210-6217.
- ↵
- Morgan JI,
- Curran T
(1995) Trends Neurosci 18: 66-67.
CrossRefliteratuerdatabases MEDLINEWeb of Science
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci USA 88: 1291-1295.
- Graybiel AM,
- Moratalla R,
- Robertson HA
(1990) Proc Natl Acad Sci USA 87: 6912-6916.
- Hope B,
- Kosofsky B,
- Hyman SE,
- Nestler EJ
(1992) Proc Natl Acad Sci USA 89: 5764-5768.
- ↵
- Kelz MB,
- Nestler EJ
(2000) Curr Opin Neurol 13: 715-720.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Koob GF,
- Sanna PP,
- Bloom FE
(1998) Neuron 21: 467-476.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Wise RA
(1998) Drug Alcohol Dependence 51: 13-22.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Hope BT,
- Nye HE,
- Kelz MB,
- Self DW,
- Iadarola MJ,
- Nakabeppu Y,
- Duman RS,
- Nestler EJ
(1994) Neuron 13: 1235-1244.
CrossRefliteratuerdatabases MEDLINEWeb of Science
- ↵
- Nye H,
- Hope BT,
- Kelz M,
- Iadarola M,
- Nestler EJ
(1995) J Pharmacol Expert 275: 1671-1680.
- Nye HE,
- Nestler EJ
(1996) Mol Pharmacol 49: 636-645.
- ↵
- Moratalla R,
- Elibol B,
- Vallejo M,
- Graybiel AM
(1996) Neuron 17: 147-156.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Pich EM,
- Pagliusi SR,
- Tessari M,
- Talabot-Ayer D,
- Hooft fan Huijsduijnen R,
- Chiamulera C
(1997) Science 275: 83-86.
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Hiroi N,
- Nakabeppu Y,
- Hope BT,
- Nestler EJ
(1995) Mol Pharmacol 48: 880-889.
- ↵
- Hiroi N,
- Brown J,
- Jo H,
- Saudou F,
- Vaidya VA,
- Duman RS,
- Greenberg ME,
- Nestler EJ
(1998) J Neurosci 18: 6952-6962.
- ↵
- Chen J,
- Kelz MB,
- Hope BT,
- Nakabeppu Y,
- Nestler EJ
(1997) J Neurosci 17: 4933-4941.
- ↵
- Hiroi N,
- Brown J,
- Haile C,
- Jo H,
- Greenberg ME,
- Nestler EJ
(1997) Proc Natl Acad Sci USA 94: 10397-10402.
- ↵
- Fienberg AA,
- Hiroi N,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Nishi A,
- Cheramy A,
- O'Callaghan JP,
- Miller D,
- Cole DG,
- en oaren.
(1998) Science 281: 838-842.
- ↵
- Hiroi N,
- Feinberg A,
- Haile C,
- Groenhof P,
- Nestler EJ
(1999) Eur J Neurosci 11: 1114-1118.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Groenhof P,
- Allen PB,
- Nairn AC
(1999) Neuron 23: 435-447.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Bibb JA,
- Snyder GL,
- Nishi A,
- Yan Z,
- Meijer L,
- Fienberg AA,
- Tsai LH,
- Kwon YT,
- Girault JA,
- Czernik AJ,
- en oaren.
(1999) Natuer (Londen) 402: 669-671.
CrossRefliteratuerdatabases MEDLINE
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Sakai N,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Duman RS,
- Nestler EJ
(1998) Mol Pharmacol 54: 495-503.
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Whisler K,
- Gilden L,
- Beckmann AM,
- Steffen C,
- Zhang YJ,
- Marotti L,
- Self SW,
- en oaren.
(1999) Natuer (Londen) 401: 272-276.
CrossRefliteratuerdatabases MEDLINE
- ↵
- Dobrazanski P,
- Noguchi T,
- Kovary K,
- Rizzo CA,
- Lazo PS,
- Bravo R
(1991) Mol Cell Biol 11: 5470-5478.
- Nakabeppu Y,
- Nathans D
(1991) Cell 64: 751-759.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Yen J,
- Wisdom RM,
- Tratner I,
- Verma IM
(1991) Proc Natl Acad Sci USA 88: 5077-5081.
- ↵
- White FJ,
- Hu XT,
- Zhang XF,
- Wolf ME
(1995) J Pharmacol Expert 273: 445-454.
- ↵
- Hyman SE
(1996) Neuron 16: 901-904.
CrossRefliteratuerdatabases MEDLINEWeb of Science
- Kreek MJ
(1997) Pharmacol Biochem Behav 57: 551-569.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Shippenberg TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449-455.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Chen JS,
- Zhang YJ,
- Kelz MB,
- Steffen C,
- Ang ES,
- Zeng L,
- Nestler EJ
(2000) J Neurosci 20: 8965-8971.
- ↵
- Bibb JA,
- Chen JS,
- Taylor JR,
- Svenningsson P,
- Nishi A,
- Snyder GL,
- Yan Z,
- Sagawa ZK,
- Nairn AC,
- Nestler EJ,
- en oaren.
(2001) Natuer (Londen) 410: 376-380.
CrossRefliteratuerdatabases MEDLINE
- ↵
- Luscher C,
- Nicoll RA,
- Malenka RC,
- Muller D
(2000) Nat Neurosci 3: 545-550.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- Malinow R,
- Mainen ZF,
- Hayashi Y
(2000) Curr Opin Neurobiol 10: 352-357.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133-141.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
Robinson, TE & Kolb, B. (1999) (1997) Eur. J. Neurosci.11, 1598-1604.
- ↵
- Carey M,
- Smale ST
(2000) Transkriysjeregeling yn Eukaryotes (Cold Spring Harbor Lab. Press, Plainview, NY).
- ↵
- Spencer VA,
- Davie JR
(1999) Gene 240: 1-12.
CrossRefliteratuerdatabases MEDLINEreach fan 'e wittenskip
 facebook
facebook- Twitter
- Google+
- CiteULike
- Hearlik
 Digg
Digg- Mendeley
HighWire-presinteare artikels dy't dit artikel befetsje
- Natuer- en Drug Rewards Act op mienskiplike neurologyske plastikaasjemeganisme mei {Delta} FosB as Key Key Mediator J. Neurosci. 2013 33 (8) 3434-3442
- Drugs, Crime and Epigenetics of Hedonic Allostasis Journal of Contemporary Criminal Justice 2012 28 (3) 314-328
- Abstract
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Abstract
- Folsleine tekst (HTML)
- Folsleine tekst (PDF)
- Morphine aktivearret de E Twenty Six-Like Transkriptfaktor-1 / Serum-Response Factor-paad fia ekstrazellulêre sinjaal-regulearre kinasen 1 / 2 yn F11-killen ûntliend fan Dorsal Root Ganglia neurons J. Pharmacol. Exp. Wetter - Agrarwetter 2012 342 (1) 41-52
- Molekulêre meganisme foar in tagongsrjochten: Epigenetyske feroarings ynsteld troch Nicotine Prime Gene ekspresje troch Cocaine Sci Transl Med 2011 3 (107) 107ra109
- Enhanced Sucrose en Cocaine sels-administraasje en cue-induced drugs sykje nei ferlies fan VGLUT2 yn Midbrain Dopamine-neurons yn messen J. Neurosci. 2011 31 (35) 12593-12603
- Chronike intermittenter hypoxia ferheget bloeddruk en ekspresje fan FosB / {Delta} FosB yn sintrale autonome regio's Bin. J. Physiol. Regul. Integr. Comp. Physiol. 2011 301 (1) R131-R139
- It ôfwêzigens fan de GPR37 / PAEL ûntfanger beheart striatal Akt en ERK2 phosphorylaasje, {Delta} FosB-ekspresje, en kondisearre plakfoarm foar amphetamin en kokaine FASEB J. 2011 25 (6) 2071-2081
- De relaasje tusken Duration of Initial Alcohol Exposure and Persistence of Molecular Tolerance Is Markedly Nonlinear J. Neurosci. 2011 31 (7) 2436-2446
- In Vivo Bioluminescence Imaging produsearret Redox-regulearre Activator Protein-1-aktivaasje yn paraventrikelige nucleus fan mice mei renovaskulêre hypertension Hypertension 2011 57 (2) 289-297
- Striatal Overexpression fan {Delta} FosB ferbrûkt chronike Levodopa-yndirekte ûnwettige bewegingen J. Neurosci. 2010 30 (21) 7335-7343
- Epigenetyske mediation fan miljeu-ynfloeden yn wichtige psychiatryske besunigingen Schizophr Bull 2009 35 (6) 1045-1056
- DNA-basearre MRI-problemen foar spesifike deteksje fan chronike eksposysje foar amfetamine yn wenjen J. Neurosci. 2009 29 (34) 10663-10670
- Alde dendritike plinyske plastytytyske yn kokaine-ôfwetterjende raten J. Neurosci. 2009 29 (9) 2876-2884
- Overexpression Screen yn Drosophila identifisearret neuronale rollen fan GSK-3 {beta} / shaggy as regulator fan AP-1-ôfhinklike ûntjouwingsplastysiteit Genetics 2008 180 (4) 2057-2071
- Transkripsje MRI: In nije werjefte fan it wenhaww Neuroscientist 2008 14 (5) 503-520
- {Delta} FosB-yndeksje yn Orbitofrontale Cortex mediares Tolerânsje nei kokaïne-yndirekte kognitive disfunksje J. Neurosci. 2007 27 (39) 10497-10507
- Fermindere kwetsberens foar it wersteljen fan methamphetamine-sykje gedrach yn glialzelline-ôflaatjende neurotropyske fakty mutantmis FASEB J. 2007 21 (9) 1994-2004
- {Delta} FosB yn 'e Nucleus Accumbens regulearret it Food-Reinforced Instrumental Behavior and Motivation J. Neurosci. 2006 26 (36) 9196-9204
- Regulearring fan {Delta} FosB Stabiliteit troch fosforaasje. J. Neurosci. 2006 26 (19) 5131-5142
- Ekspresje fan Mutant NMDA-receptors yn Dopamine D1-receptor-kontrolearde sellen ferbean kokaine-sensibilisaasje en fergruttet de kokaïnske foarkar J. Neurosci. 2005 25 (28) 6651-6657
- D1 Dopamine-receptors Modulearje {Delta} FosB-yndeksje yn rat striatum nei intermittenter Morphine-bestjoer J. Pharmacol. Exp. Wetter - Agrarwetter 2005 314 (1) 148-154
- Neurobiology of Mys Selected for High Voluntary Wheel-running Activity Integr. Comp. Biol. 2005 45 (3) 438-455
- Effekten fan wetterbeheining en rehydraasje op c-Fos en FosB finke yn 'e rat supraoptyske kearn en lamina terminalisregio Bin. J. Physiol. Regul. Integr. Comp. Physiol. 2005 288 (1) R311-R321
- Transkriale yndeks fan FosB / {Delta} FosB Gene troch mechanike stress yn Osteoblasts J Biol Chem 2004 279 (48) 49795-49803
- Ynduksje fan {Delta} FosB yn Belang-ferbûne hynstestruktueren nei chronike stress J. Neurosci. 2004 24 (47) 10594-10602
- Sim1-genodaasjemodules modulearret de homeostatyske fiedingsaksje op it fergrutsjen fan diabetesfet yn mûzels Bin. J. Physiol. Endocrinol. Metab. 2004 287 (1) E105-E113
- DNA-mikroarrayanalyse fan gene ekspresje yn minsklike optyske nervekloft astrocytes yn antwurd op hydrostatyk druk Physiol. Genomics 2004 17 (2) 157-169
- Superoxide wurdt yn 'e Sintrale Nervous System Activaasje en Sympathoexcitation fan Myocardial Infarction-Induced Heart Failure Circus. Res. 2004 94 (3) 402-409
- Adenosine A2A-receptors yn neuroadaptaasje nei werhelle dopaminergyske stimulaasje: Ymplikaasjes foar de behanneling fan dyskinesias yn 'e sykte fan Parkinson Neurology 2003 61 (90116) S74-81
- Cytoplasmyske fersus Nuclear pleatsing fan fos-ferbûne proteins yn 'e Frog, Rana esculenta, Testis: In Vivo en direkte yn Vitro-effekt fan in gonadotropine-releasinghormon Agonist Biol. Reprod. 2003 68 (3) 954-960
- Periadolesintyske messen sjen litte fergrutte Delta FosB-Upregulaasje yn Response oan kokaine en amfetamine J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB regulearret fytspaad J. Neurosci. 2002 22 (18) 8133-8138
- CREB-aktiviteit yn 'e kearn accumbens shell kontrôles fan' e gedrach fan responsueren op emosjonele stimuli Proc. Natl. Acad. Sci. USA 2002 99 (17) 11435-11440
- Psychogenomics: mooglikheden foar Understanding Addiction J. Neurosci. 2001 21 (21) 8324-8327