KOMMENTAAR: Prachtige resinsje fan hast alle relevante ûndersiik oer androgen-receptors, dopamine en seksuele funksje. Fantastysk tekenjen fan 'e ynterrelaasjes fan' e hypothalamus-beleanningskrets.
Horm Behav. 2008 mei; 53(5): 647-658.
Online publisearre 2008 febrewaris 13. doi: 10.1016 / j.yhbeh.2008.01.010
Abstract
Folwoeksenheid wurdt assosjeare mei ferhegingen yn gedrachssoartige plezier, dy't op syn beurt wurde foarme troch de pubertale aktivearring fan 'e hypothalamo-hypofyse-gonadale as. Yn bistemodellen fan natuerlik beleanjend gedrach, lykas seks, drage testikulêre androgenen by oan 'e ûntwikkeling en útdrukking fan it gedrach by manlju. Om gedrachsmatigens te beynfloedzjen, ûndergiet it harsens signifikante remodeling tidens adolesinsje, en in protte fan 'e feroaringen binne ek gefoelich foar androgenen, en hannelje wierskynlik fia androgenreceptors (AR). Sjoen de delikate ynteraksje fan gonadale hormonen en harsensûntwikkeling, is it gjin ferrassing dat beheining fan hormonnivo's yn dizze gefoelige perioade de folwoeksenen en gedrach fan folwoeksenen signifikant feroaret. Yn manlike hamsters is eksposysje oan testosteron tidens adolesinsje fereaske foar normale ekspresje fan seksueel gedrach by folwoeksenen. Manlju dy't fan androgenen wurde ûntslein tidens puberteit jouwe oanhâldende tekoarten yn paring. Oarsom, androgenen allinich binne net genôch om paring te inducearjen by prepubertale manlju, ek al binne harsens AR oanwêzich foar puberteit. Yn dit ferbân is breed gebrûk fan anabole-androgenyske steroïden (AAS) tidens adolesinsje in wichtige soarch. Misbrûk fan AAS hat it potensjeel om sawol de timing as de nivo's fan androgenen yn adolesinte manlju te feroarjen. Yn hamsters fergruttet adolesinte AAS-eksposysje agresje, en feroarsake bliuwende feroarings yn neurotransmittersystemen. Derneist binne AAS sels fersterkend, lykas oantoand troch selsadministraasje fan testosteron en oare AAS. Resint bewiis suggereart lykwols dat de fersterkende effekten fan androgenen gjin klassike AR nedich binne. Dêrom is fierdere ûndersiik fan ynteraksjes tusken androgenen en beleanjend gedrach yn 'e adolesinte harsens fereaske foar in better begryp fan AAS-misbrûk.
Oersicht
Folwoeksenens wekker it brein oan sawol wille as risiko. Yn minsklike teeners nimt dit faak de foarm fan eksperimintearjen mei drugs en seks. Yn 'e Feriene Steaten is de mediaan leeftyd foar earste geslachtsferkear by manlju 16.4 jier, en 65% hawwe geslachtsferkear hân troch 12th klasse (Kaiser Family Foundation, 2005). Likemin hat dizze populaasje de heechste tariven fan yllegaal drugsgebrûk yn 'e Feriene Steaten. Neffens de 2004 Nasjonale enkête oer drugsgebrûk en sûnens brûkte 38% fan manlju fan leeftyd 18 – 25 it ôfrûne jier in yllegale drugs (SAMHSA / OAS, 2005). Boppedat brûkte 31% fan teen jonges drugs as alkohol tidens har lêste seksuele moeting (Kaiser Family Foundation, 2005). Derneist is adolesinsje in pivotale tiid yn 'e etiology fan bepaalde psychopatologen, lykas depresje, eangst, oerstallige iten, en gedrachstoornis. Wy posearje dat de pubertale sekretion fan gonadale hormonen, har aktivearring fan steroide receptors yn 't harsens, en de ynteraksje tusken hormoan en ûnderfining oer adolesinte harsensûntwikkeling bydrage oan' e gedrachferoaringen dy't sjoen wurde tidens adolesinsje.
Us doel hjir is om it bewiis te kontrolearjen dat gonadale androgenen bemiddelje mei de folwoeksenheid fan folwoeksenen en prestaasjes fan folwoeksenen fan motivearre gedrach, lykas ek de beleanjende eigenskippen fan dizze gedragingen. Wy presintearje ek bewiis dat testosteron sels beleanjend is, wat wierskynlik draacht by oan maturational feroarings yn motivearre gedrach tidens adolesinsje, doe't testosteronnivo oprint. De fokus fan dit artikel leit op ús stúdzjes fan neurale sirkwizen ûnderlizzend manlik seksueel gedrach, yn 't bysûnder yn' e Syryske hamster, mei spesjale klam op 'e ynteraksje tusken testosteron en dopamine (DA). Wy stelle foar dat pubertale androgenen sawol oergeande as lange termyn effekten hawwe op beleanningskretsen en motivearre gedrach. Wy hypotezeje fierder dat oanfolling mei exogene androgenen yn 'e foarm fan anabole-androgenyske steroïden (AAS) de normale ynfloeden fan pubertale androgenen fergruttet, wêrtroch de adolesinte ûntwikkeling fan harsens en gedrach negatyf beynfloedet.
Adolesinsje as in gefoelige perioade foar harsensûntwikkeling
Uteinlik is it brein sawol in trigger as in doelwyt foar androgeenaksje by adolesinsje. By jonge jonges (<12 jier) en jonge hamsters (<28 dagen âld) binne sirkulearjende androgenen en gonadotropinen op basale nivo's. As sekresje fan luteinisearjend hormoan út 'e foarste hypofyse rint op yn reaksje op hypothalamysk gonadotropine-frijkommende hormoan, ferheegje sirkulearjende testosteron-konsintraasjes signifikant. Dit bart troch Tanner-poadium II / III (14 jier) by jonges, en mei 28 dagen âld yn hamsters. Tsjin 'e tiid dat jonges Tanner-poadium IV / V berikke (sawat 16 jier âld) as as hamsters 50-60 dagen âld binne, leit endogeen testosteron binnen it folwoeksen manlike berik. Pubertale hormoanesekresje komt oerien mei de perioade fan adolesinsje, dy't plakfynt fan likernôch 12 oant 20 jier by minsken. Pubertale hormonen hannelje net allinich op perifere weefsels om it uterlik fan sekundêre geslachtskenmerken te feroarsaakjen dy't de iepenlike tekens fan puberteit binne, mar se hannelje ek sintraal om sawol de ferbouwing fan 'e adolesinte harsens as gedrachsryp te beynfloedzjen. Fierder liede de fysiologyske en neurologyske feroaringen ta stân brocht troch pubertale hormonen ta wichtige feroarings yn 'e ûnderfining fan in yndividu, dy't sels de rin fan harsenûntwikkeling djip kinne feroarje. Sadwaande foarmet de puberale ferheging fan geslachtssteroide hormonen, oandreaun troch ûntwikkele tydlike rijping fan 'e reproduktive neuroendokrine as, op har beurt foarming fan adolesinte gedrachsûntwikkeling fia sawol direkte as yndirekte ynfloeden op it senuwstelsel.
Minsklike adolesinsje wurdt no erkend as in wichtige en dynamyske perioade fan neurale ûntjouwing wêryn't gedrachssirkels wurde opknapt en ferfine. Hoewol it brein fan in 5-jier-âld bern al 90% fan syn folwoeksen grutte is (Dekaban, 1978), wichtige remodeling is noch te kommen. Dit konsept waard oanstutsen troch ûndersyk yn sawol minsken as bisten dy't dokuminteare dat in protte fan 'e basisûntwikkelingsprosessen dy't barre tidens perinatale harsensûntwikkeling wurde opnij sammele tidens adolesinsje. Dizze prosessen omfetsje neurogenese (Eckenhoff en Rakic, 1988; Hy en Crews, 2007; Pinos, Collado, Rodriguez-Zafra, Rodriguez, Segovia, en Guillamon, 2001; Rankin, Partlow, McCurdy, Giles, en Fisher, 2003), programmearre selstjerte (Nunez, Lauschke, en Juraska, 2001; Nunez, Sodhi, en Juraska, 2002), útwurking en snoeien fan dendrityske arborisaasjes en synapses (Andersen, Rutstein, Benzo, Hostetter, en Teicher, 1997; Huttenlocher en Dabholkar, 1997; Lenroot en Giedd, 2006; Sowell, Thompson, Leonard, Welcome, Kan, en Toga, 2004), myelination (Benes, Turtle, Khan, en Farol, 1994; Paus, Collins, Evans, Leonard, Pike, en Zijdenbos, 2001; Sowell, Thompson, Tessner, en Toga, 2001) en seksuele differinsjaasje (Chung, De Vries, en Swaab, 2002; Davis, Shryne, en Gorski, 1996; Nunez et al., 2001). Sa is de ûntjouwingsbaan fan 'e postnatale harsens net lineêr, mar wurdt ynstee karakterisearre troch in adolesinte burst fan rappe feroaring en omfettet sawol progressive as regressive barrens. Lykas elke ûntwikkelingsbiolooch wist, signalearje perioaden fan rappe ûntwikkelingsferoaring de gefoelichheid en kwetsberens foar sawol ûnderfining-ôfhinklike feroaring as oan negative gefolgen fan perturbaasje en belediging, en d'r is gjin reden om te tinken dat minsklike adolesinte harsensûntwikkeling elke útsûndering is (Andersen, 2003; Spear, 2000). Sa soe perturbaasjes yn 'e timing fan ynfloed op pubertal hormoan op' e adolesinte harsens wurde foarsizze langduorjende gefolgen te hawwen foar gedrach fan folwoeksenen.
Androgenen en neurologyske sirkels foar motivearre gedrach
Om't adolesinsje in transiente en dynamyske faze fan ûntwikkeling is, soe it lestich wêze om it adolesinte brein en gedrach yn isolemint te evaluearjen. Om it unike karakter fan 'e adolesinsje te wurdearjen, is it nuttich om it te kontrastearjen mei it harsens en gedrach fan folwoeksen folwoeksenen. Sa, mei de fokus fan dit artikel op manlik seksueel gedrach en beleanning, is it hjir wichtich om de neurale sirkwizen foar kopulaasje en seksuele motivaasje yn folwoeksen manlju yn te fieren, ynklusyf de rol fan gonadale steroïde hormonen yn gedrachaktivering en de ferdieling fan receptors foar androgenen (AR) en estrogenen (ER).
AR binne oanwêzich yn selgroepen dy't de neurale sirkwy foarmje, dy't beleanjend sosjaal gedrach bemiddelje, lykas seks. Fierder wurde harsens AR útdrukt foar puberteit yn hamsters en wurde upregulearre troch androgenen yn sawol jeugd as folwoeksen manlju (Kashon, Hayes, Shek, en Sisk, 1995; Meek, Romeo, Novak, en Sisk, 1997). Yn knaagdierhars is der substansjele oerlaap yn 'e ferdieling fan AR en ER (Houten en Newman, 1995), en aromatase (Celotti, Negri-Cesi, en Poletti, 1997), ynklusyf sawol α- as β-foarmen fan 'e estrogeenreceptor (Shughrue, Lane, en Merchenthaler, 1997). By binding oan ligand, "klassike" AR en ER funksjonearje as transkriptyfaktoaren om transkripsje en synthese fan nije proteïnen te inducearjen. Net ferrassend folgje dizze effekten in relatyf stadige tiidkursus, mei in fertrage begjin fan 'e aksje. Steroïde stimulearring fan seksueel gedrach fan manlike hamster (Edel en Alsum, 1975) is konsistint mei aksjes fia klassike genomyske aksjes. Bygelyks, 2 wiken fan eksposysje foar steroide is ferplicht om paring te herstellen yn kastraten op lange termyn. Mear resinte stúdzjes yn ratten hawwe ek rappe sellulêre effekten oantoand fan androgenen yn harsensregio's dy't in pear klassike receptors hawwe (Mermelstein, Becker, en Surmeier, 1996). Dizze steroide aksjes wurde tocht wurde bemiddele troch net-genomyske receptors. Wylst de ferdieling fan klassike AR en ER yn 'e hamsterbrein relatyf beheind is (Hout en Swann, 1999) binne de potinsjele harsensdoelen foar net-genomyske androgeenaksje folle breder.
It mediale preoptyske gebiet (MPOA) spilet in sintrale rol yn kopulaasje by manlju fan goudfisk oant minsken (besjoen yn Hull, Wood, en McKenna, 2006). Boppedat transduceert de hamster MPOA gonadale steroïdehormonen fia oerfloedige AR en ER, en testosteronimplantaten yn MPOA binne genôch om seksuele aktiviteit te herstellen yn kastraten op lange termyn (Hout en Swann, 1999). Yn manlike ratten hannelje gonadale steroïden yn 'e MPOA om basale DA-frijlitting te regeljen (Putnam, Sato, en Hull, 2003) en stimulearje mating (Hull, Du, Lorrain, en Matuszewich, 1995). Yn it begjin is d'r in beskieden ferheging fan DA as in wyfke wurdt presinteare efter in skerm. By kopulaasje nimt MPOA DA fierder ta (+ 50% fan 'e basisline), en dit effekt fereasket androgenen (Hull et al., 1995; Putnam et al., 2003). Net ferrassend, yn kastreare manlju dy't net passe, nimt MPOA DA net ta (Hull et al., 1995). It is wat lestich om dit resultaat te ynterpretearjen, om't it ûntbrekken fan DA-frijlitting ferbjustere wurdt troch it ûntbrekken fan seksuele aktiviteit. DA-frijlitting yn MPOA korreleert lykwols mei it ferlies fan paring yn koarte termyn kastraten (Hull et al., 1995), en mei testosteron-induzeare restauraasje fan seksuele aktiviteit yn kastraten op lange termyn (Du, Lorrain, en Hull, 1998; Putnam, Du, Sato, en Hull, 2001).
Binnen de knaagdier MPOA spylje de androgenyske en estrogenyske metaboliten fan testosteron spesifike rollen yn 'e regeling fan paring (Putnam et al., 2003; Putnam, Sato, Riolo, en Hull, 2005). De latency om kopulaasje (mount of intromit) te inisjearjen is ien maatregel fan seksuele motivaasje. De latency foar seksuele aktiviteit is gefoelich foar estrogenen, troch ûnderhâld fan MPOA stikstofoxide synthase, dy't op syn beurt basale DA-nivo's behâldt. Estrogeen behannele kastraten litte hege basale DA-nivo's sjen, dy't sterk korrelearje mei de mooglikheid om kopulaasje te inisjearjen. Se kinne lykwols gjin ferheging fan frou- en kopulaasje-induzeare sjen litte yn DA-frijlitting, dy't sterk korrelearje mei seksuele prestaasjes. Dêrfandinne is har seksuele prestaasjes ûnder yntakt nivo. Oan 'e oare kant, kastraten behannele mei net-aromatisearbere androgen allinich litte gjin ferhege basale DA-nivo's sjen, en se slagje net te kopiearjen. Foar normale seksuele prestaasjes binne dus sawol estrogenen as androgenen fereaske. Seksuele prestaasjes wurde normaal útdrukt as frekwinsje mjittingen fan mounts, intromissions en ejakulaasjes. Pas as sawol estrogenen as androgenen wurde ferfongen, eksposearje kastreare manlju op ferhege DA-nivo's (en koartere latencymaatregels) en froulike- en kopulaasje-induzeare DA-ferhegings (en ferhege frekwinsje-maatregels). Op dizze manier drage estrogenen yn MPOA by oan seksuele motivaasje, en sawol estrogenen as androgenen oan seksuele prestaasjes.
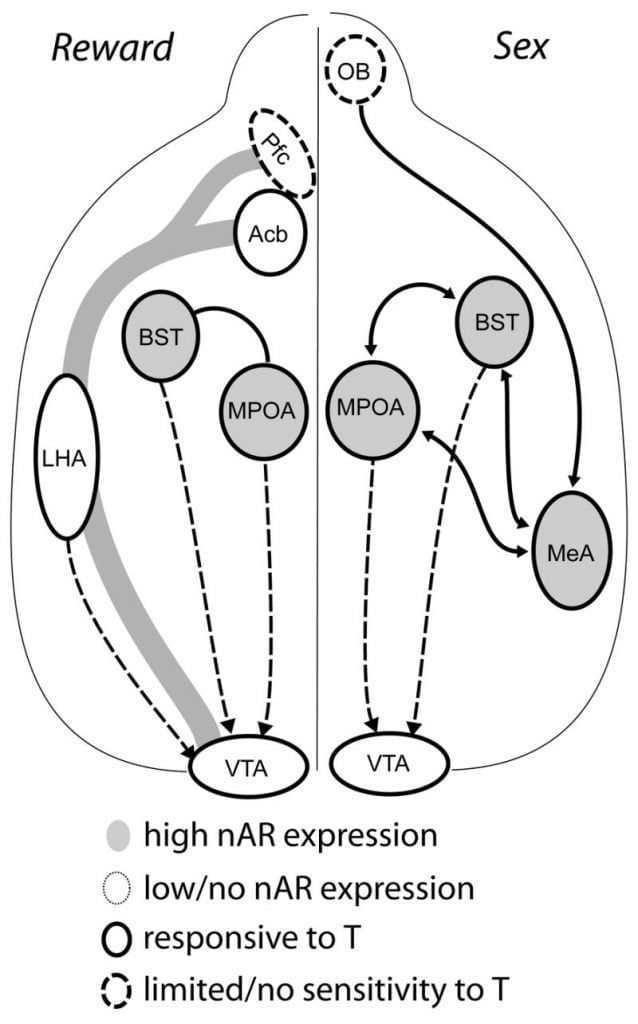
Hoewol testosteron needsaaklik is foar MPOA DA-frijlitting tidens manlik copulatorygedrach en foar it sels paarjen, kin noch testosteron noch paring allinich DA yn MPOA lokke. Ynstee dêrfan binne chemosensoryske oanwizings fan gewissige wyfkes ek nedich foar DA-frijlitting yn MPOA. Yn knaagdieren binne chemosensoryske stimuli de primêre sensoryske modaliteit om manlik seksueel gedrach te inisjearjen (Fig. 1). Chemosensoryske cues wurde oerbrocht fan 'e olfaktoryske bollen nei MPOA fia de mediale amygdaloidkern en de bêdkern fan' e stria terminalis, struktueren mei in protte AR en ER (Hout en Swann, 1999). Om de rol fan chemosensoryske signalen te bepalen yn paring-induzeare DA, mjitten wy MPOA DA tidens paring yn gonad-yntakte manlike hamsters mei unilaterale olfaktoryske bulbectomy (UBx, Triemstra, Nagatani, en Hout, 2005). Hoewol bilaterale ferwidering fan 'e olfaktoryske bollen ferwideret seksuele aktiviteit en frijlitting fan MPOA DA, unilaterale bulbektomy bemoeit de mating net. Yn dit ûndersyk feroarsake kopulaasje MPOA DA-frijlitting by it mjitten fan kontralateraal oan 'e lekteare olfaktoryske lampe, mar net yn it ipsilaterale healrûn (Fig. 2). Soartgelikense resultaten waarden waarnommen by manlike ratten mei letsels fan 'e mediale amygdala (Dominguez, Riolo, Xu, en Hull, 2001). Yn in relatearre stúdzje, gemyske stimulearring fan 'e mediale amygdala yn ratten induzeare MPOA DA-frijlitting gelyk oan dy tidens kopulaasje (Dominguez en Hull, 2001). Meiinoar suggerearje dizze gegevens dat testosteron in permissive omjouwing makket dat eksterne sensoryske stimuli MPOA kin berikke en DA-frijlitting induzearje by kopulaasje.

Fig. 2
Uteinlik aktivearje seksueel gedrach en oare natuerlike beleannings neurale beleanningswegen. It mesocorticolimbic DA-sirkwy bestiet út it ventrale tegmentale gebiet (VTA), nucleus accumbens (Acb), en prefrontale cortex (Pfc). Dopamine-sel lichems wenje yn it VTA-projekt rostrally nei de Acb en Pfc (Koob en Nestler, 1997). By rotten wurdt DA frijlitten yn Acb tidens seks (Pfaus, Damsma, Nomikos, Wenkstern, Blaha, Phillips, en Fibiger, 1990). In protte misbrûk drugs hannelje ek yn it mesolimbyske DA-systeem om DA-frijlitting (amfetaminen) te ferheegjen as DA opnij opnimmen (kokaïne, Di Chiara en Imperato, 1988), sadat har ferslaavjende eigenskippen fersterke. Op dizze wize hat testosteron it potensjeel om de frijlitting fan DA yn Acb te beynfloedzjen sawol troch har ferbettering fan seksueel gedrach, en troch syn aksjes as drugs foar misbrûk (sjoch hjirûnder).
Hjoeddeistich bewiis suggereart dat it mesocorticolimbic DA-systeem rijpt tidens adolesinsje. Acb DA-glêsdichheden ferheegje dramatysk under adolesinsje yn gerbils, wat suggerearret dat signifikante rijping fan VTA-dopaminergyske projeksjes nei de Acb plakfynt yn 'e adolesinte perioade (Lesting, Neddens, en Teuchert-Noodt, 2005). Fierder wurdt dopaminergyske ynfier nei GABA (γ-aminobutyric acid) -ergyske sellen yn 'e rat mediale prefrontale cortex ferrike en moduleare troch serotonergyske systemen tidens pubertale ûntwikkeling (Benes, Taylor, en Cunningham, 2000), en manipulaasje fan androgenen by folwoeksen ratten liedt ta feroaringen yn dopaminerge axondichtheid binnen prefrontale korteks (Kritzer, 2003). De Pfc, Acb, en de VTA hawwe net folle AR as ER, hoewol ERβ oanwêzich is yn 'e VTA (Shughrue et al., 1997). Dêrom liket it wierskynlik dat androgenen it mesocorticolimbic DA-systeem beynfloedzje fia androgen-gefoelige afferinten of fia ERβ yn 'e VTA lykas yn hypothalamus (Handa et al., Dizze útjefte). Us gegevens litte sjen dat androgen-gefoelige sellen yn manlike hamsters projektearje nei de VTA út struktueren ferbûn mei steroïde-gefoelige gedragingen. Bygelyks befetsje sawol de MPOA as de bêdkern fan stria terminalis (BST) in grut oantal AR-positive sellen dy't projeksje nei de VTA (Sato en Hout, 2006). It ventral pallidum, it wichtichste effektyf doel fan Acb (Zahm en Heimer, 1990), befettet ek in protte AR-positive sellen dy't projisearje nei de VTA. Dizze projeksjes jouwe in kâns foar androgenen om de aktiviteit fan it mesocorticolimbic DA-systeem te feroarjen.
Steroïde-ôfhinklike organisaasje fan gedrach tidens adolesinsje
De tradisjonele werjefte fan hormoanaksje op folwoeksen gedrach is basearre op aktivearjende effekten fan steroide hormonen, dy't ferwize nei it fermogen fan steroïden om gedrach yn spesifike sosjale konteksten te fasilitearjen troch aksje binnen doelzellen yn 'e neurale sirkels ûnderlizzend gedrach. Aktivearjende effekten binne foarbygeand yn 'e sin dat se komme en gean mei de oanwêzigens en ôfwêzigens fan hormoan, en se wurde typysk assosjeare mei de ekspresje fan folwoeksen gedrach. Yn tsjinstelling, ferwize organisatoaryske effekten nei it fermogen fan steroïden om nervosysteemstruktuer te byldzjen tidens ûntwikkeling. Strukturele organisaasje is permanint, bliuwt bûten de perioade fan bleatstelling oan hormoan, en bepaalt neurale en gedrachsresponsen op steroïden yn folwoeksenheid. Us begryp fan 'e ûntwikkelingsrelaasje tusken organisatoaryske en aktivearjende effekten fan steroide hormonen is yn' e ôfrûne 50-jierren evoluearre. Phoenix en kollega's stelden earst foar dat gedragings (aktivearjende) antwurden op folwoeksenen op steroïdehormonen wurde programmeare (organiseare) troch steroïdehormonen yn in maksimaal gefoelige perioade fan perinatale ûntwikkeling (Phoenix, Goy, Gerall en Young, 1959). Letter lei Scott en kollega's it teoretyske grûnwurk foar it bestean fan meardere gefoelige perioaden foar de progressive organisaasje fan it nervosysteem, en konstateare dat gefoelige perioaden it meast foarkomme yn perioaden fan rappe ûntwikkelingsferoaring (1974). Dêrnei wiisden Arnold en Breedlove op dat steroide-ôfhinklike organisaasje fan 'e harsens kin barre bûten gefoelige perioaden fan ûntwikkeling (Arnold en Breedlove, 1985). Yn 'e ôfrûne 15-jierren makket ûndersyk mei in ferskaat oan diermodellen en gedrachssystemen it dúdlik dat it adolesinte brein gefoelich is foar sawol aktivearjende as organisaasje-effekten fan gonadale steroïden (besjoen yn Sisk en Zehr, 2005). En, lykas oare perioaden fan rappe ûntjouwingsferoaring, fertsjintwurdigt adolesinsje in definieare finster fan kâns foar steroide-ôfhinklike harsensferoaring.
Us wurk mei de hamster as diermodel leveret bewiis dat manlike sosjale gedrach wurdt feroare troch steroïden tidens adolesinsje (Schulz, Menard, Smith, Albers, en Sisk, 2006; Schulz en Sisk, 2006). Foardat puberteit kin de behanneling fan testosteron gjin seksueel gedrach yn hamsters aktivearje, wat suggereart dat maturaasjeprosessen dy't neurale sirkwizen kwetsber meitsje foar aktivearring of organisaasje troch steroïde hormonen noch net binne bard (Meek et al., 1997; Romeo, Richardson, en Sisk, 2002a). Oarsom, wylst de iepen útdrukking fan manlik reproduktyf gedrach yn folwoeksenen net perfoarst de oanwêzigens fan gonadale steroïden fereasket yn 'e adolesinsje, docht de maksimale útdrukking fan gedrach. Fergelykjen fan manlik reproduktyf gedrach yn mantsjes dy't prepubertysk (NoT@P) of postpuberteel (T@P) kastrearre binne en dêrnei behannele mei testosteron yn folwoeksenen, hawwe prepubertale NoT@P-kastraten op syn minst in 50% tekoart yn manlik gedrach yn ferliking mei mantsjes dy't nei adolesinsje kastrearre binne (Fig. 3, Schulz, Richardson, Zehr, Osetek, Menard, en Sisk, 2004). Boppedat binne tekoarten yn reproduktyf gedrach lang duorjend, en kinne se net oerwûn wurde troch langere behanneling fan testosteron of troch seksuele ûnderfining yn folwoeksenheid (Schulz et al., 2004). Lykas, nei behanneling mei estrogen en progesteron, toane NoT@P-mantsjes koartere lordose-latinsjes en langere lordose-durings dan manlju kastrearre as folwoeksenen (Schulz et al., 2004), suggereare dat prepubertale kastraten minder defeminiseare binne as de manlju bleatsteld oan pubertale testosteron.

Fig. 3
It kin wêze dat NoT@P mantsjes lije oan fermindere seksuele motivaasje. Ien manier om dizze fraach oan te pakken is om de latencies te fergelykjen om diel te nimmen oan sawol ano-genital ûndersyk (AGI) as mounting tusken mantsjes gonadectomized foar (NoT@P) en nei puberteit (T@P). As seksuele motivaasje ôfhinklik is fan eksposysje foar gonadale hormoanen yn 'e adolesinsje, soene wy langere latencies foarsizze om seksueel gedrach yn NoT@P-manlju te dwaan. Yndied, mei werhelle bleatstelling oan estrus wyfkes, nimme NoT@P-mantsjes langer om AGI te begjinnen en te montearjen yn ferliking mei T@P-mantsjes (Fig. 4). Sa liket it, neist it organisearjen fan aspekten fan seksuele prestaasjes, dat pubertale hormonen ek de beleanjende aspekten fan seksueel gedrach organisearje. Yn stipe fan dizze mooglikheid herstelt sintrale administraasje fan 'e DA agonist apomorphine yn' e folwoeksenheid it berchgedrach fan NoT@P-mantsjes nei folwoeksen-typyske nivo's, wat suggerearret dat testosteron yn 'e adolesinsje normaal organisearret dopaminergyske neuronale circuits (Salas-Ramirez, Montalto, en Sisk, 2006) ). Dochs bliuwe in protte nijsgjirrige fragen oer. Soe in NoT @ P male barpress foar in estrous female of ûntwikkeljen fan in conditioned plak foarkar foar in mating lokaasje? Takomstich ûndersyk sil de rol fan pubertale hormonen ûndersykje by it organisearjen fan seksuele motivaasje en seksuele prestaasjes.

Fig. 4
Anogenitaal ûndersyk (AGI) latencies en durations útstald troch manlike hamsters gonadectomized foar puberteit (NoT@P) of nei puberteit (T@P). Alle mantsjes waarden testosteron-primed yn folwoeksenen 7 wk nei gonadectomy en ien wike foarôfgeand oan de earste gedrachstest. A. T@P mantsjes toande ferlykbere AGI latencies oer de trije testen mei in estrus female, wylst NoT @ P mantsjes ferhege AGI latencies tidens de tredde test mei in estrus female . B. T@P-manlju fermindere berch-latinsjes oer de trije gedrachstests mei in estrus-wyfke, wylst noT@P-manlju gjin feroaring yn 'e berchlatens sjen lieten oer de trije gedrachstests. Dizze gegevens suggerearje dat pubertale gonadale hormonen bliuwende, fasilitearjende effekten hawwe op folwoeksen manlike motivaasje om seksueel gedrach mei in frou te dwaan. (Net-publisearre gegevens fan dierûnderwerpen yn Schulz, KM, Richardson, HN, Zehr, JL, Osetek, AJ, Menard, TA, en Sisk, CL, 2004).
Prepubertale gedrachsreaksjes op steroïden
Ien fan 'e bliuwende puzels fan adolesinte gedrachsûntwikkeling is de reden wêrom't aktivearring fan reproduktyf gedrach yn reaksje op eksponering fan steroïden wurdt ferswakke yn prepubertale manlike hamsters. As lege nivo's fan androgenen foar puberteit de ekspresje fan manlik seksueel gedrach yn prepubertale manlju beheine, dan moat oanfoljende endogene androgenen by prepubertale manlju parse. Dit docht bliken dat it net it gefal is (Meek et al., 1997; Romeo, Cook-Wiens, Richardson, en Sisk, 2001; Romeo, Wagner, Jansen, Diedrich, en Sisk, 2002b), nettsjinsteande it feit dat it oantal en ferdieling fan AR en ER troch it heule parse-sirkwy binne gelyk yn hormoan behannele prepubertale en folwoeksen kastraten (Meek et al., 1997; Romeo, Diedrich, en Sisk, 1999; Romeo et al., 2002a). Dêrom docht bliken dat androgenen en AR needsaaklik binne, mar net genôch foar ekspresje fan manlik seksueel gedrach.
Pogingen om faktoaren te identifisearjen dy't seksuele aktiviteit beheine foar puberteit binne oant no ta mingd. Fos-antwurden op chemosensoryske signalen fan estrous froulju binne gelyk yn prepubertale en folwoeksen manlike hamsters (Romeo, Parfitt, Richardson, en Sisk, 1998). Dizze gegevens bewize dat sensoryske transduksjemechanismen matich binne foar puberteit. Sa kinne jeugdhannelingen gemososensoryske signalen fan wyfkes opspoare; wêr't se ferskille fan folwoeksenen is yn hoe't se reagearje op dy signalen. Ien potensjele ferklearring is dat prepubertale manlju net motiveare binne om seksueel gedrach te dwaan. Wy hawwe fûn dat prepubertale manlike hamsters gjin ferhege dopaminergyske responsen werjaan yn 'e MPOA yn antwurd op froulike feromonen, wylst seksueel naïve folwoeksen manlju robúste MPOA-dopaminergyske antwurden werjaan op deselde stimuli (Fig. 5, Schulz, Richardson, Romeo, Morris, Lookingland, en Sisk, 2003). Likemin slagget prepubertale manlju net de folwoeksen-typyske ferheging fan sirkulearjend testosteron sjen te litten nei bleatstelling oan froulike feromonen (Parfitt, Thompson, Richardson, Romeo, en Sisk, 1999). Sa ferskine froulike feromonen in ûnbedoelde stimulâns te wêzen foar neurochemyske en neuro-endokriene antwurden by folwoeksenen, mar net prepubertale manlju, wat suggereart dat de salience fan dizze sosjaal relevante sensoryske stimuli feroaret oer pubertale ûntwikkeling, mooglik relatearre oan it oankeapjen fan beleanjende eigenskippen en seksuele motivaasje. Derneist, hoewol testosteron AGI fasiliteart fan in wyfke yn prepubertale manlju, hinget dit effekt ôf of de manlike foarige bleatstelling hat oan in estrofyske frou. Miskien ferrassend, testosteron-behanneling fermindert de latency en fergruttet de doer fan AGI allinich by seksueel naïve prepubertale manlju (Fig. 6). Fierder, prepubertale manlju dy't ien foarige ûnderfining hawwe hienen mei in frou, werjaan folle langere AGI-latencies en koartere AGI-duorsumheden dan manlju dy't ynteraksje hawwe mei ûntfangende wyfkes foar it earst (Fig. 6). Dizze gegevens suggerearje dat ynteraksjes mei in estrous froulik aversyf binne ynstee fan beleanning foarôfgeand oan puberteit, en eliminearje dêrtroch alle fasilitearjende effekten fan testosteron op AGI tidens folgjende ynteraksjes mei in frou. It soe nijsgjirrich wêze om te witten oft de negative gedrachskonsekwinsjes fan iere eksposysje oan in estrous frou yn folwoeksenheid en folwoeksenheid oanhâlde, benammen sjoen de werhelle eksposysje oan estrous froulju tidens adolesinsje fasiliteart yn 't algemien de útdrukking fan manlik reproduktyf gedrach (Molenda-Figueira, Salas-Ramirez, Schulz, Zehr, Montalto, en Sisk, 2007).

Fig. 5
Prepubertale en folwoeksen manlike mediale preoptyske gebiet (MPOA) dopaminergyske antwurden op froulike feromonen befette yn vaginale sekresjes. Folwoeksen manlju toane ferhegings yn MPOA-dopaminergyske aktiviteit mei bleatstelling oan froulike vaginale sekresjes, wylst prepubertale manlju net ferhege MPOA-dopaminergyske antwurden werjaan op froulike feromonen. (Opnommen fan Schulz, KM, Richardson, HN, Romeo, RD, Morris, JA, Lookingland, KJ, en Sisk, CL, 2003).

Hoewol prepubertale androgeenbehanneling gjin kopulaasje kin inducearje, suggereart resint wurk fan ús laboratoarium dat it hamster nervosysteem gefoelich is foar de organisearjende aksjes fan testosteron op reproduktyf gedrach foarôfgeand oan adolesinsje (Schulz, Zehr, Salas-Ramirez, en Sisk, 2007). Kastraasje plus 19 dagen fan eksposysje foar testosteron foar of tidens, mar net nei adolesinsje, makken montagegedrach doe't testosteron waard ferfongen yn folwoeksenheid. Manlju bleatsteld prepubertaal oan testosteron werjûn ek mear intromissions yn folwoeksenheid dan manlju bleatsteld oan testosteron tidens of nei puberteit (Schulz et al., 2007). Dizze gegevens suggerearje dat it fermogen fan testosteron om gedrach neurale sirkwinen te organisearjen ôfnimt mei de leeftyd, en dat adolesinsje it ein markeart fan in langwerpige postnatale gefoelige perioade foar bleatstelling oan testosteron.
Farmakologyske androgenen
De foargeande gegevens suggerearje dat endogene gonadale steroïden motiveare gedrach ferbetterje tidens adolesinsje. No, wat bart der as men androgenen sels administraert op nivo's oant 100x normale fysiologyske konsintraasjes? Dit is it probleem fan anabole-androgenyske steroïde (AAS) misbrûk (besjoen yn Brower, 2002; Clark en Henderson, 2003). In koarte ferdjipping is hjir passend: alle AAS binne derivaten fan testosteron, alle AAS hawwe in koalstofskelet mei 4 fuseare ringen, de measte hawwe 19-koolhydraten. AAS wurde yn haadsaak brûkt foar har anabole (spierbouwende) effekten. Lykas har namme al fermoeden docht, hawwe AAS ek androgenyske eigenskippen. Testosteron is in logyske kar yn dierestúdzjes foar it ferkennen fan fûnemintele meganismen fan androgen beleanning. It bliuwt ek in populêre kar foar minsklike brûkers, faaks yn 'e foarm fan langwerkende testosteronesters lykas testosteronpropionaat. Yn 2006 wie testosteron de ienige meast foarkommende ferbeane stof ûntdutsen yn urinetests by WADA-akkrediteare laboratoaren (WADA, 2006). Testosteron stie foar de grutste fraksje (34%) fan AAS-positive urine-tests op 'e Olympyske Spullen fan 2000 yn Sydney (Van Eenoo en Delbeke, 2003). Op deselde wize testen yn urine tests fan AAS-brûkers 41% posityf foar testosteron (Brower, Catlin, Blow, Eliopulos, Beresford, 1991). By hege doses produsearje AAS wichtige gedrachsferoaringen. Yn 't bysûnder, fanwegen har nauwe relaasje mei testosteron, soe AAS-gebrûk yn' e teenjier lykje de normale steroïde milieu fan it ûntwikkeljen fan minsklik adolesint nervosysteem te ûnderbreken, ynklusyf de kwantiteit, timing, en soarte fan eksposysje fan steroïde.
Lykas by oare yllegale drugs is minske-misbrûk fan AAS in probleem fan adolesinsje. Neffens de 1994 Nasjonale húshâldlike enkête oer drugsgebrûk (SAMHSA / OAS, 1996), pykjes fan steroide gebrûk yn 'e lette adolesinsje op 18-leeftyd. Boppedat is yn 'e enkête Monitoring the Future (Johnston, O'Malley en Bachman, 2003), de libbensfrekwinsje fan steroide gebrûk ûnder senioaren op middelbere skoallen (2.7%) wie te ferlykjen mei dat foar crack kokaïne (3.5%) as heroïne (1.4%). Steroide gebrûk komt ek hieltyd faker foar op jongere leeftiden: 2.5% fan 'e studinten fan' e 8e klasse (13 – 14 jier) hawwe steroïden brûkt, gelyk oan 'e ynsidinsje fan crack (2.5%) en heroïnegebrûk (1.6%). Dizze trend nei gebrûk fan AAS yn 'e iere teeners is benammen lestich mei it each op soargen 1) dat adolesinten benammen kwetsber wêze kinne foar misbrûk fan AAS, en 2) dat eksposysje foar folwoeksenen oan AAS op farmakologyske nivo's it potensjeel hat om de normale rijping fan harsens substansjeel te feroarjen gedrach om oerdreaun morfologyske en gedrachreaksjes te produsearjen, akute en chronike.
Ongepaste agresje is de gedrachsrespons dy't meast assosjeare wurdt mei minsklik AAS-misbrûk. Yn publisearre saakrapporten is steroide gebrûk ymplisearre yn ferskate gewelddiedige moarden (Conacher en Workman, 1989; Paus en Katz, 1990; Paus, Kouri, Powell, Campbell, en Katz, 1996; Schulte, Hall, en Boyer, 1993). Yn ûndersiken fan hjoeddeistige AAS-brûkers wiene ferhege agresje en irritabiliteit de meast foarkommende gedrachseffekten fan AAS-gebrûk (Bond, Choi, en paus, 1995; Galligani, Renck, en Hansen, 1996; Midgley, Heather, en Davies, 2001; Parrott, Choi, en Davies, 1994; Perry, Kutscher, Lund, Yates, Holman, en Demers, 2003). Sjoen it berik fan androgen-eksposysjes, it ferskaat oan psychiatryske symptomen, en it potinsjeel foar pre-besteande psychiatryske dysfunksje, is it lykwols lestich de krekte rol fan AAS yn dizze gefallen fan minsklike agresje te bepalen. Resultaten fan prospective stúdzjes fan minsklike frijwilligers dy't ynjeksjes fan AAS krigen binne mingd: Tricker et al (1996) en O'Connor et al (2004) rapporteare gjin ferhegingen fan lilk gedrach, wylst oare stúdzjes ferhege agresje hawwe observearre (Daly, Su, Schmidt, Pickar, Murphy, en Rubinow, 2001; Hannan, Friedl, Zold, Kettler, en Plymate, 1991; Kouri, Lukas, paus, en Oliva, 1995; Paus en Katz, 1994; Su, Pagliaro, Schmidt, Pickar, Wolkowitz, en Rubinow, 1993). Nettsjinsteande is it wichtich om yn gedachten te hâlden dat de doses dy't wurde jûn oan minsklike frijwilligers folle leger binne as de doses foarsteld op websides foar lichemsbou, en de doer fan 'e behanneling is oer it algemien koart. Sa liket it op lykwicht earlik te konkludearjen dat AAS it potinsjeel hat om agonistysk gedrach te ferbetterjen, teminsten yn gefoelige persoanen. Pope et al (1994) fûn dat AAS psychiatryske symptomen opropt by kwetsbere persoanen.
Dierestúdzjes hawwe ek twingend bewiis levere foar AAS-induzeare agresje. Folwoeksen manlike hamsters dy't chronisch behannele binne mei steroide mei hege doasis hawwe koartere oanfallen latency en in grutter oantal oanfallen en byt nei in manlike ynkringer yn fergeliking mei unbehandele manlju (Harrison, Connor, Nowak, Nash, en Melloni, 2000; Melloni, Connor, Hang, Harrison, en Ferris, 1997). Likemin produseart in mylde provokaasje (sturtknipe) in persistinte tanimming fan agresje by adolesinte manlike ratten, ynklusyf agresje tsjin wyfkes (Cunningham en McGinnis, 2006). Fan noch gruttere soargen, eksposysje foar folwoeksenen oan AAS yn hamsters feroarsaket bliuwende ferheegingen yn agonistysk gedrach dat oanhâldt nei it stopjen fan steroide gebrûk (Grimes en Melloni, 2006). Dizze gedrachsferoaringen wurde begelaat troch bliuwende remodeling fan neurale sirkwy yn 'e anterior hypothalamus. Benammen, adolesinte AAS-eksposysje yn hamsters ferbetteret arginine vasopressine (AVP, Grimes en Melloni, 2006) en regelt serotonine en de serotonergyske 5HT1A- en 5HT1B-receptors (Ricci, Rasakham, Grimes, en Melloni, 2006). It soe as gjin ferrassing moatte komme dat AAS harsnivo's fan AR ek feroaret. Kronyske eksposysje oan testosteron as nandrolon regelet cell nucleaire AR yn manlike ratten (Menard en Harlan, 1993; Wesson en McGinnis, 2006). Sa is d'r it potensjeel foar AAS om androgen-ôfhinklike gedragingen te ferbetterjen sawol troch endogenous androgenen oan te foljen en troch androgenyske responsiviteit te ferheegjen fia ferhege AR-ekspresje.
Yn ferliking mei agonistysk gedrach hawwe AAS in minder markearre effekt op paringsgedrach yn manlike knaagdieren, en de respons hinget ôf fan it bepaalde steroide dat brûkt wurdt (besjoen yn Clark en Henderson, 2003). Yn manlike hamsters dy't testosteron konsumearje yn orale oplossingen, hawwe ejakulaasjes op in dosis-ôfhinklike manier ferhege (Wood, 2002). Noch testosteron noch nandrolone ferbettere mating yn adolesinte manlike ratten. Stanozolol, in relatyf minder potinsjele AAS mei minimale androgenyske aktiviteit, inhibearde feitlik sawol paring as agresje (Farrell en McGinnis, 2003), wierskynlik troch it ferminderjen fan endogene androgen nivo.
It is foaral wichtich om te notearjen dat adolesinte en folwoeksen hamsters ferskate gedrachsresponsen kinne werjaan op AAS-eksposysje. Wylst AAS de agonistyske gedrach markant ferbettere by adolesinte manlju, produsearre deselde behanneling yn folwoeksenen mar in beskieden tanimming fan agressyf gedrach en fermindere seksueel gedrach (Salas-Ramirez, Montaldo en Sisk, 2008). Dit is konsistint mei it konsept fan adolesinsje as in gefoelige perioade foar androgenaksje. Fierder, krekt as folwoeksen manlike hamsters fertsjinje tolerânsje foar exogene testosteron (Peters en Wood, 2005), wy leauwe dat manlju ûntwikkelje tolerânsje oannimme foar testosteron as se mature. Sa kinne de effekten fan AAS feroarje oer adolesinte ûntwikkeling, en adolesinte AAS-eksposysje kinne oermjittige agressyf en seksueel gedrachspatroanen feroarsaakje dy't kinne folhâlde oant folwoeksenheid.
It fersterkjen fan effekten fan androgenen
Paring en fjochtsjen binne elk beleanjend (teminsten as jo de striid winne). Manlike ratten sille ferskate kearen op in hendel drukke om te kopulearjen mei in wyfke (Everitt en Stacey, 1987). Op deselde manier sille manlike mûzen en froulike hamsters in betingst foar foarkar meitsje foar plak (CPP) foar lokaasjes wêr't se earder gefjochten hawwe wûn (Martinez, Guillen-Salazar, Salvador, en Simon, 1995; Meisel en Joppa, 1994). As AAS beleanjend sosjaal gedrach kin ferbetterje boppe nivo's dy't normaal waarnommen wurde yn gonad-yntakte manlju, is it logysk om te ferwachtsjen dat testosteron sels beleanjend wêze kin. Dit is test mei twa goed fêststelde diermodellen foar beleanning en fersterking: CPP en selsbehear. De resultaten fan dizze stúdzjes bewize dat testosteron fersterket yn in eksperimintele kontekst wêr't anabole effekten en atletyske prestaasjes irrelevant binne. Mei CPP wurdt de teststof hieltyd wer keppele mei in unike omjouwing (bygelyks in bepaalde keamer yn it testapparaat). As it bist ienris de fersterkende teststof assosjearet mei dy omjouwing, sil hy de omjouwing sykje, sels yn ûntbrekken fan beleanning. De earste rapporten oer androgen beleanning yn laboratoariumdieren brûkten systemyske ynjeksjes fan testosteron om CPP te inducearjen yn manlike mûzen (Arnedo, Salvador, Martinez-Sanchis, en Gonzalez-Bono, 2000; Arnedo, Salvador, Martinez-Sanchis, en Pellicer, 2002) en rat (Alexander, Packard, en Hines, 1994; de Beun, Jansen, Slangen, en Van de Poll, 1992). Dêrnei brûkte ús laboratoarium sels-administraasje fan testosteron om androgenforsterking te demonstrearjen (Johnson en Wood, 2001). Wy fûnen dat manlike hamsters frijwillich mûnlinge oplossingen fan testosteron sille konsumearje mei beide keuzetests fan 2-flesje en drinken troch iten feroarsake. Yn lettere stúdzjes demonstrearren wy sels self-administration yn manlike ratten en hamsters (Hout, Johnson, Chu, Schad en Sels, 2004). Intravenous levering elimineert potensjele ferwarrende effekten fan smaak as darmfylling op androgen-yntak.
Yn 'e kontekst fan AAS-misbrûk is it wichtich om te ûnderskieden tusken sintrale en perifeare effekten fan androgenen. Sûnt testosteron hat wiidferspraat effekten oer it lichem, koe it wurde beweare dat beleanning en fersterking mei systemyske testosteron-ynjeksjes sekundêr is foar testosteron's systemyske anabole en androgenyske aksjes. Mei oare wurden, miskien testosteron fermindert spiermoedigens en ferbettert de mienskiplike funksje, sadat bisten gewoan binne fiele better. Yndied, dizze ferklearring is brûkt yn 'e klinyske literatuer (hoewol sûnder eksperiminteel bewiis) om te arguminten tsjin it potensjeel foar ôfhinklikens en ferslaving oan AAS (DiPasquale, 1998). Packard et al (Packard, Cornell, en Alexander, 1997) liet sjen dat ynjeksjes fan testosteron direkt yn 't rat harsens CPP kinne induzearje. Likemin hat ús laboratoarium sels-administraasje intracerebroventrikulêre (icv) testosteron yn manlike hamsters (Wood et al., 2004). Intracerebral CPP en icv sels-administraasje mei testosteron argumintearje foar sintrale doelen dy't meditaasje fan androgen fersterkje.
It is opmerklik dat fersterking fan testosteron net needsaaklik folget deselde meganismen dy't earder fêststeld binne foar steroïde effekten op seksueel gedrach. Lykas earder besprutsen is de MPOA in wichtige side foar organisaasje fan seksueel gedrach fan manlike knaagdieren (Hull, Meisel, en Sachs, 2002). Yn hamsters hat de MPOA in soad steroide receptors, en testosteronimplantaten yn MPOA werstelle seksuele aktiviteit yn kastraten op lange termyn (Hout en Swann, 1999). De tiidferrin fan dizze steroïde effekten is traach: montearend gedrach bliuwt wiken nei orchidektomy, en útwreide steroide-eksposysje is needsaaklik om paring yn lange-termyn kastraten te restaurearjen (Edel en Alsum, 1975). Ynjeksjes fan testosteron yn MPOA fan manlike ratten ûntbrekke lykwols CPP (Kening, Packard, en Alexander, 1999). Dit suggerearret dat oare harsregio's wichtich binne foar androgen fersterking.
Yn tsjinstelling, manlike ratten sille in CPP foarmje foar testosteron-ynjeksjes yn Acb (Packard et al., 1997). Lykas by oare drugs fan misbrûk, is DA wierskynlik in wichtige neurotransmitter foar testosteron-fersterking: CPP induce troch systemyske testosteron-ynjeksje wurdt blokkearre troch D1 en D2 dopamine receptor antagonisten (Schroeder en Packard, 2000). Oars as oare drugs fan misbrûk suggerearje ús ûndersiken yn hamsters dat testosteron de frijlitting fan Acb DA net induceert (Triemstra, Sato, en Hout, yn 'e parse). Likemin litte stúdzjes fan manlike ratten sjen dat androgenen gjin effekt hawwe op basale DA-nivo's as op amfetamine-stimuleare DA-frijlitting (Birgner, Kindlundh-Hogberg, Nyberg, en Bergstrom, 2006; mar ek sjen Clark, Lindenfeld, en Gibbons, 1996). Fierder oefenet testosteron in relatyf lytse ynfloed op Acb DA tissue levels (Thiblin, Finn, Ross, en Stenfors, 1999). Mei-inoar suggerearje dizze gegevens dat hoewol testosteronfersterking úteinlik DA-aktiviteit yn Acb kin feroarje, kinne de meganismen ûnderskiede fan dy fan kokaïne as oare stimulanten. Wat dat oanbelanget suggerearje resinte gegevens dat chronike eksposysje oan AAS de gefoelichheid foar DA kin feroarje troch DA-metabolisme te feroarjen (Kurling, Kankaanpaa, Ellermaa, Karila, en Seppala, 2005), nivo's fan DA-receptors (Kindlundh, Lindblom, Bergstrom, Wikberg, en Nyberg, 2001; Kindlundh, Lindblom, en Nyberg, 2003) as de DA-transporter (Kindlundh, Bergstrom, Monazzam, Hallberg, Blomqvist, Langstrom, en Nyberg, 2002).
Op it stuit binne de spesifike steroïdsignalen, receptors en harsens fan aksjes foar testosteron fersterking net bekend. Op grûn fan in resinte stúdzje fan hamsters út ús laboratoarium, lykje de fersterkende effekten fan testosteron bemiddele troch sawol androgenen as estrogenen (DiMeo en Wood, 2006). Algemien misbrûkte AAS omfetsje sawol aromatisearbere as net-aromatisearbere androgenen (Gallaway, 1997; WADA, 2006). Dit ympliseart dat sawol AR en ER steroïde stimuli kinne transduce foar beleanning. D'r is de ekstra mooglikheid dat fersterking fan testosteron kin wurde bemiddele troch in kombinaasje fan klassike en net-genomyske receptors.
Ferskate rigels fan bewiis wize op 'e aksjes fan net-genomyske reseptors yn' e fersterkjende effekten fan AAS. Neist de seldsume ferdieling fan AR yn Acb en VTA, is de tiidferrin fan androgenfersterking rap (<30 min), en sinjaalferwurking fia klassike AR is miskien net rap genôch foar fersterking. Om de rol te testen fan net-genomyske AR yn AAS-fersterking, hawwe wy twa komplementêre techniken brûkt (Fig. 7). Yn ien eksperimint (Sato, Johansen, Jordaanje en Hout, 2006), wy tastiene rotten mei de testikulêre feminisaasjemutaasje (Tfm, sjoch dit probleem) om dihydrotestosteron (DHT), in net-aromatisearbere androgen, sels te behearjen. De Tfm-mutaasje ferminderet ligandbining sterk by AR. Nettsjinsteande, Tfm-rôten en har wylde soart manlike broers en susters hawwe sawat itselde bedrach fan DHT sels beheard. Dit argumint foar net-genomyske effekten fan DHT. Yn in folgjende stúdzje hawwe wy bepaald as manlike hamsters DHT sels soene beheare, konjugeare oan bovine serumalbumine (BSA, Fig. 8, Sato en Hout, 2007). DHT-BSA-konjugaten binne membraan-impermeabel; sadwaande binne har effekten beheind ta it seloverflak. Hamsters selsbehearde DHT, lykas earder oantoand (DiMeo en Wood, 2006). Se lieten in ferlykbere foarkar sjen foar DHT-BSA-konjugaten, mar slaggen net om sels BSA sels te behearjen.


Dizze gegevens wize op in sintrale rol foar sellen oerflak AR's yn androgen fersterking. Op it stuit is de krekte aard fan sokke receptors net bekend. It is suggereare dat androgenen kinne hannelje op it seloverflak troch te binen oan tawijd membraan AR (Thomas, Dressing, Pang, Berg, Tubbs, Benninghoff, en Doughty, 2006, sjoch dizze kwestje ek). Dit kin wêze yn 'e foarm fan ekstra-nukleêre klassike AR lykas rapporteare yn hippocampus (Sarkey et al., Yn dizze útjefte). As alternatyf hawwe eardere stúdzjes ek steroide-bindende siden op oare neurotransmittersystemen beskreaun. Spesifyk kinne in ferskaat oan steroide hormonen ynklusyf AAS allosterysk de GABA-A-receptor modulearje (Henderson, 2007; Lambert, Belelli, Peden, Vardy, en Peters, 2003). Likemin kinne sulfeare neurosteroïden aktiviteit feroarje fan N-methyl-D-partypeseptor-subtypes (Maleisk, Gibbs en Farb, 2002) reseptors. Dit is in wichtich gebiet foar takomstich ûndersyk.
Wêrom soe d'r in membraan AR wêze? Lykas earder besprutsen is d'r in nauwe assosjaasje tusken androgeen sekretion en beleanjend sosjaal gedrach. Wy kinne spekulearje dat de ferheging fan testosteron-sekresje dy't folget op paring as fjochtsjen tsjinnet om it gedrach te fersterkjen. As dat sa is, is it nedich om in rappe koppeling fan stimulearring (gedrach) en beleanning (testosteron) te hawwen. Dit kin it bêste berikt wurde troch bining oan membraan AR. Wat dit oanbelanget soe it ynteressant wêze om te bepalen as it klemmjen fan androgeen sekretion tidens paring de beleanjend effekten fan seksueel gedrach ferminderet.
Gearfetting
Hjir kontrolearje wy it bewiis dat androgenen potinsjele mediatoren binne fan folwoeksen motiveare gedrach, en fierders, dat de timing fan eksposysje foar androgenen tidens ûntwikkelingsprogramma's androgen-ôfhinklik motiveare gedrach yn folwoeksenheid. Anaboleus steroïden wurde rap in populêr medisyn foar misbrûk troch adolesinten yn 'e FS. Hoewol AAS de ferslaavjende potinsje fan kokaïne of heroïne miskien net hat, binne wy krekt begon it potensjeel te ferstean foar androgen fersterking en ferslaving. Yn 't bysûnder, as jeugdsport kompetitive wurdt, wurdt d'r tanimmende druk op it ûntwikkeljen fan atleten om steroïden te brûken, te begjinnen op jongere leeftiden. Dizze trend is lestich mei it each op nij bewiis foar steroïde-gefoelige neurale maturaasje by adolesinten.
Nettsjinsteande ferhege bewustwêzen troch sawol de iepenbiere as wittenskiplike mienskippen fan 'e djippe neurale feroaringen dy't folgje mei folwoeksenheid, is eksperiminteel ûndersyk fan' e ûntwikkelingsneurobiology fan puberteit beheind. Diermodellen fan adolesinte ûntwikkeling binne nedich om te ûndersiikjen hoe't de timing fan eksponysje fan hormoanen by ûntwikkeling it risiko fan in yndividu fergruttet foar psychopathology en drugsgebrûk, en hokker soarten ûnderfiningen de gedrachseffekten fan ôfwikingen yn pubertale timing ferminderje of fersterkje. Bygelyks, sosjale faktoaren lykas peer-ynfloed fergrutsje de effekten fan pubertale timing foar stof- en alkoholgebrûk (Biehl, Natsuaki, en Ge, 2007; Patton, Novy, Lee, en Hickok, 2004; Simons-Morton en Haynie, 2003; Wichstrom en Pedersen, 2001). Diermodellen fan pubertale timing sille ek ynspanningen foar minsklik ûndersyk ynformearje, en potinsjeel liede ta effektiver terapeutyske yntervinsjes tidens adolesinsje.
Acknowledgments
Wy betankje Eleni Antzoulatos, Cortney Ballard, Lucy Chu, Kelly Peters, Jennifer Triemstra, Jane Venier, Lisa Rogers, en Pamela Montalto foar help by dizze stúdzjes. Dit wurk stipe troch subsydzjes fan 'e NIH (DA12843 nei RIW, MH68764 nei CLS, en MH070125 nei KMS).
Fuotnoten
Disclaimer fan útjouwer: Dit is in PDF bestân fan in net fertsjinjen manuskript dat foar publikaasje akseptearre is. As tsjinst foar ús klanten biede wy dizze fryske ferzje fan it manuskript. It manuskript sil kopieare, sjittende, en oersicht fan 'e resultant dúdlikens wurde foardat it publisearre is yn syn definitive citable form. Tink derom dat yn 'e produksjeproses miskien fûnen ûntdutsen wurde dy't de ynhâld beynfloedzje kinne, en alle juridyske disclaimers dy't tapasse foar it tydskrift binne beskerme.
Referinsjes
- Alexander GM, Packard MG, Hines M. Testosteron hat lekkerjende affektive eigenskippen yn manlike rotten: ymplikaasjes foar de biologyske basis fan seksuele motivaasje. Behaviorale neuroanens. 1994;108: 424-8. [PubMed]
- Andersen SL. Trajektoares fan 'e harsensûntwikkeling: punt fan kwetsberens of finster fan gelegenheid? Neuroscience & Biobehavioral Reviews. 2003;27: 3-18. [PubMed]
- Andersen SL, Rutstein M, Benzo JM, Hostetter JC, Teicher MH. Seksferskillen yn dopamine-receptor-overproduksje en eliminaasje. Neuroreport. 1997;8: 1495-8. [PubMed]
- Arnedo MT, Salvador A, Martinez-Sanchis S, Gonzalez-Bono E. Belofte eigenskippen fan testosteron yn intakte manlike mûzen: in pilotstúdzje. Farmakology, biogemy en gedrach. 2000;65: 327-32.
- Arnedo MT, Salvador A, Martinez-Sanchis S, Pellicer O. Fergelykbere beleanjend effekten fan testosteron by mûzen dy't wurdearre as persoanen mei koarte en lange oanfal latency. Addiction Biology. 2002;7: 373-9. [PubMed]
- Arnold AP, Breedlove SM. Organisatoaryske en aktivearjende effekten fan geslachtssteroïden op harsens en gedrach: in reanalyse. Hormonen en gedrach. 1985;19: 469-98. [PubMed]
- Benes FM, Taylor JB, Cunningham MC. Konverginsje en plastisiteit fan monoaminergyske systemen yn 'e mediale prefrontale cortex yn' e postnatale perioade: ymplikaasjes foar de ûntwikkeling fan psychopathology. Cerebral Cortex. 2000;10: 1014-27. [PubMed]
- Benes FM, Turtle M, Khan Y, Farol P. Myelinaasje fan in kaaiferlieningsône yn 'e hippocampalfoarming fynt yn it minskemienskip by jongens, adolesinsje en adulthood. Argyf fan 'e Algemiene Psychiatry. 1994;51: 477-84. [PubMed]
- Biehl MC, Natsuaki MN, Ge XJ. De ynfloed fan pubertale timing op alkoholgebrûk en swiere drinkbanen. Journal of Youth and Adolescence. 2007;36: 153-167.
- Birgner C, Kindlundh-Hogberg AM, Nyberg F, Bergstrom L. Neuroscience Letters. 2006. Altered extracellular levels of DOPAC and HVA in the rat nucleus accumbens shell in antwurd op sub-chronike nandrolone administraasje en in folgjende amfetamine-útdaging.
- Bond AJ, Choi PY, Pope HG., Jr Beoardieling fan attintive bias en stimming by brûkers en net-brûkers fan anabole-androgenyske steroïden. Ofhinklikens fan drugs en alkohol. 1995;37: 241-5. [PubMed]
- Brower KJ. Anaboleus steroïde misbrûk en ôfhinging. Aktuele psychiatryske rapporten. 2002;4: 377-87. [PubMed]
- Brower KJ, Catlin DH, Blow FC, Eliopulos GA, Beresford TP. Klinyske testen en urinetests foar anabole-androgenyske steroïde misbrûk en ôfhinklikens. Amerikaansk sjoernaal fan drugs- en alkoholmisbrûk. 1991;17: 161-171. [PubMed]
- Celotti F, Negri-Cesi P, Poletti A. Steroïde metabolisme yn it sûchdierhars: 5alpha-reduksje en aromatisaasje. Breinûndersyk Bulletin. 1997;44: 365-75. [PubMed]
- Chung WC, De Vries GJ, Swaab DF. Seksuele differinsjaasje fan 'e bêdkern fan' e stria terminalis by minsken kin útwreidzje oant folwoeksenheid. Journal of Neuroscience. 2002;22: 1027-33. [PubMed]
- Clark AS, Henderson LP. Gedrachs- en fysiologyske antwurden op anabole-androgenyske steroïden. Neuroscience & Biobehavioral Reviews. 2003;27: 413-36. [PubMed]
- Clark AS, Lindenfeld RC, Gibbons CH. Anabole-androgenyske steroïden en harsensbeleanning. Farmakology, biogemy en gedrach. 1996;53: 741-5.
- Conacher GN, Workman DG. Geweldige kriminaliteit mooglik ferbûn mei anabole steroide gebrûk. American Journal of Psychiatry. 1989;146: 679. [PubMed]
- Cunningham RL, McGinnis MY. Fysike provokaasje fan pubertale anabole androgenyske steroïde bleatstelde manlike ratten wekket agresje nei wyfkes. Hormonen en gedrach. 2006;50: 410-6. [PubMed]
- Daly RC, Su TP, Schmidt PJ, Pickar D, Murphy DL, Rubinow DR. Cerebrospinale floeistof en gedrach feroarings nei administraasje fan methyltestosteron: foarriedige befiningen. Argyf fan 'e Algemiene Psychiatry. 2001;58: 172-7. [PubMed]
- Davis EC, Shryne JE, Gorski RA. Strukturele seksuele dimorfismen yn 'e anteroventral periventrikulêre kearn fan' e rat hypothalamus binne perinatally gefoelich foar gonadale steroïden, mar ûntwikkelje peripubertaal. Neuroendokrinology. 1996;63: 142-8. [PubMed]
- de Beun R, Jansen E, Slangen JL, Van de Poll NE. Testosteron as appetiteare en diskriminearjende stimulearring by rotten: seks- en dosis-ôfhinklike effekten. Fysiology en gedrach. 1992;52: 629-34. [PubMed]
- Dekaban AS. Feroaringen yn harsensgewichten tidens it span fan it minsklik libben: relaasje fan harsensgewichten oan lichemshichten en lichemsgewichten. Annalen fan neurology. 1978;4: 345-56. [PubMed]
- Di Chiara G, Imperato A. Drogen dy't troch minsken misbrûkt foarkomme synaptyske dopamine konsintraasjes yn it mesolimbysk systeem fan frij ferkearde ratten. Proceedings fan 'e National Academy of Sciences fan' e Feriene Steaten. 1988;85: 5274-8. [PMC fergees artikel] [PubMed]
- DiMeo AN, hout RI. Self-administraasje fan estrogen en dihydrotestosteron yn manlike hamsters. Hormonen en gedrach. 2006;49: 519-26. [PubMed]
- DiPasquale M. Anabole steroïden. Yn: Tarter RE, Ammerman RT, Ott PJ, redakteuren. Hânboek fan stofmisbrûk. Plenum Press; NY: 1998. pp. 547 – 565.
- Dominguez J, Riolo JV, Xu Z, Hull EM. Regulaasje troch de mediale amigdala fan kopulaasje en mediale preoptyske dopamine frijlitting. Journal of Neuroscience. 2001;21: 349-355. [PubMed]
- Dominguez JM, Hull EM. Stimulaasje fan 'e mediale amygdala ferbetteret mediale preoptyske dopamine-frijlitting: ymplikaasjes foar seksueel gedrach fan manlike ratten. Brain Research. 2001;917: 225-229. [PubMed]
- Du J, Lorrain DS, Hull EM. Kastraasje nimt ekstrazellulêre, mar ferheft intrazellulêre, dopamine yn mediale preoptyske gebiet fan manlike rotten. Brain Research. 1998;782: 11-17. [PubMed]
- Eckenhoff MF, Rakic P. Aard en needlot fan proliferative sellen yn 'e hippocampale dentate gyrus yn' e libbensperioade fan 'e rhesusap. Journal of Neuroscience. 1988;8: 2729-47. [PubMed]
- Everitt BJ, Stacey P. Studearjen fan ynstrumintgedrach mei seksueel fersterking yn manlike rotten (Rattus norvegicus): II. Effekten fan preoptike gebieten läsionen, kastraasje en testosteron. Journal of Comparative Psychology. 1987;101: 407-19. [PubMed]
- Gallaway S. De steroïde Bibel. Belle International Press; Sacramento, CA: 1997.
- Galligani N, Renck A, Hansen S. Persoanlikensprofyl fan manlju mei anabole androgenyske steroïden. Hormonen en gedrach. 1996;30: 170-5. [PubMed]
- Grimes JM, Melloni RH., Jr Lange feroaringen yn it neurotyske serotonine-systeem nei it stopjen fan folwoeksen anabole-androgenyske steroïde-eksposysje yn hamsters (Mesocricetus auratus) Behaviorale neuroanens. 2006;120: 1242-51. [PubMed]
- Hannan CJ, Jr, Friedl KE, Zold A, Kettler TM, Plymate SR. Psychologyske en serum homovanilline soere feroaringen yn manlju administraasje androgenyske steroïden. Psychoneuroendokrinology. 1991;16: 335-43. [PubMed]
- Harrison RJ, Connor DF, Nowak C, Nash K, Melloni RH., Jr Chronyske anabole-androgenyske steroïde behanneling tidens adolesinsje fergruttet anterior hypothalamyske vasopressine en agresje yn yntakte hamsters. Psychoneuroendokrinology. 2000;25: 317-38. [PubMed]
- Hy J, Crews FT. Neurogenesis fermindert yn 'e harsenspieling fan adolesinsje foar adulthood. Farmakology, biogemy en gedrach. 2007;86: 327-33.
- Henderson LP. Steroïde modulaasje fan GABAA-receptor-bemiddele transmission yn 'e hypothalamus: effekten op reproduktive funksje. Neurofarmakology. 2007;52: 1439-53. [PMC fergees artikel] [PubMed]
- Hull EM, Du J, Lorrain DS, Matuszewich L. Ekstrazellulêre dopamine yn 'e mediale preoptyske gebiet: gefolgen foar seksuele motivaasje en hormonale kontrôle fan' e kopulaasje. Journal of Neuroscience. 1995;15: 7465-7471. [PubMed]
- Hull EM, Meisel RL, Sachs BD. Manlik seksueel gedrach. Yn: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, redakteuren. Hormonen, harsens, en gedrach. Akademyske parse; New York: 2002. pp. 3 – 137.
- Hull EM, Wood RI, McKenna KE. Neurobiology fan manlik seksueel gedrach. Yn: Neill JD, bewurker. Fysiology fan fuortplanting. Vol. 1. Elsevier Press; New York: 2006. pp. 1729 – 1824.
- Huttenlocher PR, Dabholkar AS. Regionale ferskillen yn synaptogenesis yn minsklike cerebral cortex. Journal of Comparative Neurology. 1997;387: 167-78. [PubMed]
- Johnson LR, Wood RI. Orale testosteron selsbehear yn manlike hamsters. Neuroendokrinology. 2001;73: 285-92. [PubMed]
- Johnston LD, O'Malley PM, Bachman JG. Studinten fan middelbere skoallen (NIH-publikaasje nr. 03 – 5375) I. Bethesda, MD: Nasjonaal Ynstitút foar drugsmisbrûk; 2003. De nasjonale ûndersiikresultaten fan 'e takomst kontrolearje oer drugsgebrûk, 1975 – 2002.
- Kaiser Family Foundation. Amerikaanske teen seksuële aktiviteit. 2005. pp. # 3040 – 02.
- Kashon ML, Hayes MJ, Shek PP, Sisk CL. Oardering fan immunoreaktiviteit troch harsens androgenreceptor troch androgen yn prepubertale manlike fretten. Biology fan fuortplanting. 1995;52: 1198-205. [PubMed]
- Kindlundh AM, Bergstrom M, Monazzam A, Hallberg M, Blomqvist G, Langstrom B, Nyberg F. Dopaminergyske effekten nei chronike behanneling mei nandrolon visualisearre yn rattenbrein troch positron-emisje-tomografy. Foarútgong yn NeuroPsychopharmacology & Biologyske Psychiatry. 2002;26: 1303-8.
- Kindlundh AM, Lindblom J, Bergstrom L, Wikberg JE, Nyberg F. It anabole-androgenyske steroide nandrolon-dekanoat hat ynfloed op de tichtheid fan dopamine-receptors yn 'e manlike rattenbrein. European Journal of Neuroscience. 2001;13: 291-6. [PubMed]
- Kindlundh AM, Lindblom J, Nyberg F. Kronyske administraasje mei nandrolon-decanoate induceert feroaringen yn 'e gen-transkriptynhâld fan dopamine D (1) - en D (2) -receptors yn' e rat harsens. Brain Research. 2003;979: 37-42. [PubMed]
- King BE, Packard MG, Alexander GM. Affektive eigenskippen fan yntra-mediale preoptyske gebieten ynjeksjes fan testosteron by manlike ratten. Neuroscience Letters. 1999;269: 149-52. [PubMed]
- Koob GF, Nestler EJ. De neurobiology fan drugsbedriuw. Journal of Neuropsychiatry & Clinical Neurosciences. 1997;9: 482-97. [PubMed]
- Kuer EM, Lukas SE, Pope HG, Jr, Oliva PS. Fergrutsje agressyf reagearjen yn manlike frijwilligers nei de administraasje fan stadige ferheegjende doses fan testosteron cypionate. Ofhinklikens fan drugs en alkohol. 1995;40: 73-9. [PubMed]
- Kritzer MF. Long-term gonadectomy beynfloedet de tichte fan tyrosinehydroxylase- mar net dopamine-beta-hydroxylase-, cholineacetyltransferase- of serotonin-immunoreaktive axonen yn 'e mediale prefrontale cortizen fan folwoeksen manlike ratten. Cerebral Cortex. 2003;13: 282-296. [PubMed]
- Kurling S, Kankaanpaa A, Ellermaa S, Karila T, Seppala T. It effekt fan sub-chronike nandrolone decanoate behanneling op dopaminergyske en serotonergyske neuronale systemen yn 'e harsens fan rotten. Brain Research. 2005;1044: 67-75. [PubMed]
- Lambert JJ, Belelli D, Peden DR, Vardy AW, Peters JA. Neurosteroide modulaasje fan GABAA-receptors. Progress yn 'e neurobiology. 2003;71: 67-80. [PubMed]
- Lenroot RK, Giedd JN. Hynderûntwikkeling yn bern en adolesinten: ynsichten fan anatomyske magnetyske resonânske ôfbylding. Neuroscience & Biobehavioral Review. 2006;30: 718-29.
- Lesting J, Neddens J, Teuchert-Noodt G. Ontogeny fan 'e dopamine innervaasje yn' e nucleus accumbens fan gerbils. Brain Research. 2005;1066: 16-23. [PubMed]
- Malayje A, Gibbs TT, Farb DH. Inhibysje fan it NMDA-antwurd troch pregnenolone sulphat ûntdekt subtype selektive modulaasje fan NMDA-receptors troch sulfoaze steroide. Britsk Journal of Pharmacology. 2002;135: 901-9. [PMC fergees artikel] [PubMed]
- Martinez M, Guillen-Salazar F, Salvador A, Simon VM. Suksesfolle intermale agresje en betingst plak foarkar yn mûzen. Fysiology en gedrach. 1995;58: 323-8. [PubMed]
- Meek LR, Romeo RD, Novak CM, Sisk CL. Hannelingen fan testosteron yn prepubertale en postubertale manlike hamsters: dissosjaasje fan effekten op reproduktyf gedrach en harsens androgen receptor immunoreaktiviteit. Hormonen en gedrach. 1997;31: 75-88.
- Meisel RL, Joppa MA. Beskikbere plak foarkar yn froulike hammers nei agressyf of seksuële treffen. Fysiology en gedrach. 1994;56: 1115-8. [PubMed]
- Melloni RH, Jr, Connor DF, Hang PT, Harrison RJ, Ferris CF. Anabolearysk-androgenyske steroïde-eksposysje tidens adolesinsje en agressyf gedrach yn gouden hamsters. Fysiology en gedrach. 1997;61: 359-64. [PubMed]
- Menard C, Harlan R. Upregulearring fan ymmunoreactiviteit fan androgenreceptor yn 'e rat harsens troch androgenyske anabole steroïden. Brain Research. 1993;622: 226-236. [PubMed]
- Mermelstein PG, Becker JB, Surmeier DJ. Estradiol rediget kalsystreamen yn rat neostriatale neuroanen fia in membranreceptor. Journal of Neuroscience. 1996;16: 595-604. [PubMed]
- Midgley SJ, Heather N, Davies JB. Levels fan agresje ûnder in groep anabole-androgenyske steroide brûkers. Medisinen, wittenskip en de wet. 2001;41: 309-14.
- Molenda-Figueira HN, Salas-Ramirez KY, Schulz KM, Zehr JL, Montalto PR, Sisk CL. Folwoeksen sosjale ûnderfining herstelt ejakuleare gedrach fan folwoeksenen yn manlike Syryske hamsters sûnder pubertale testosteron. Maatskippij foar gedrachsneuroendokrinology; Pacific Grove, CA: 2007.
- Edel RG, Alsum PB. Hormonôfhinklike seksdimorfismen yn 'e gouden hamster (Mesocricetus auratus) Fysiology en gedrach. 1975;14: 567-74. [PubMed]
- Nunez JL, Lauschke DM, Juraska JM. Seldood yn 'e ûntwikkeling fan' e posterior cortex by manlike en froulike ratten. Journal of Comparative Neurology. 2001;436: 32-41. [PubMed]
- Nunez JL, Sodhi J, Juraska JM. Ovariale hormonen nei postnatale dei 20 ferminderje neuronûmer yn 'e rat primêre fisuele cortex. Journal of Neurobiology. 2002;52: 312-21. [PubMed]
- O'Connor DB, Archer J, Wu FC. Effekten fan testosteron op stimming, agresje, en seksueel gedrach by jonge manlju: in dûbelblinde, placebo-kontroleare, oerstallige stúdzje. Journal of Clinical Endokrinology en metabolisme. 2004;89: 2837-45. [PubMed]
- Packard MG, Cornell AH, Alexander GM. Belangrike affektive eigenskippen fan intra-nucleus accumbens ynjeksjes fan testosteron. Behaviorale neuroanens. 1997;111: 219-24. [PubMed]
- Parfitt DB, Thompson RC, Richardson HN, Romeo RD, Sisk CL. GnRH mRNA nimt ta mei puberteit yn it manlike Syryske hamsterbrein. Journal of Neuroendokrinology. 1999;11: 621-7. [PubMed]
- Parrott AC, Choi PY, Davies M. Anabole steroïde gebrûk troch amateuratleten: effekten op psychologyske stimmingssteaten. Journal of Sports Medicine & Physical Fitness. 1994;34: 292-8. [PubMed]
- Patton PE, Novy MJ, Lee DM, Hickok LR. De diagnoaze en reproduktive útkomst nei sjirurgyske behanneling fan 'e folsleine septate uterus, dupliseare cervix en vaginale septum. American Journal of Obstetrics & Gynecology. 2004;190: 1669 – 75. 1675 – 8. [PubMed]
- Paus T, Collins DL, Evans AC, Leonard G, Pike B, Zijdenbos A. Maturation fan wite matearje yn it minskehûs: in oersicht fan magnetyske resonante stúdzjes. Breinûndersyk Bulletin. 2001;54: 255-66. [PubMed]
- Perry PJ, Kutscher EC, Lund BC, Yates WR, Holman TL, Demers L. Maatregels fan agresje en stimmingsferoaringen yn manlike gewichtheffers mei en sûnder androgenysk anabole steroide gebrûk. Journal of Forensic Sciences. 2003;48: 646-51. [PubMed]
- Peters KD, Wood RI. Androgenôfhinklikens yn hamsters: oerdosis, tolerânsje, en potensjele opioidrike meganismen. Neuroscience. 2005;130: 971-81. [PubMed]
- Pfaus JG, Damsma G, Nomikos GG, Wenkstern DG, Blaha CD, Phillips AG, Fibiger HC. Seksueel gedrach ferbetteret de sintrale oerdracht fan dopamine yn 'e manlike rat. Brain Research. 1990;530: 345-348. [PubMed]
- Phoenix CH, Goy RW, Gerall AA, Jonge WC. Organisearjen fan aksje fan prenataal administraasje testosteronpropionat op 'e weefsels dy't paringgedrach yn' e froulike guinea pig bemiddelje. Endokrinology. 1959;65: 369-82. [PubMed]
- Pinos H, Collado P, Rodriguez-Zafra M, Rodriguez C, Segovia S, Guillamon A. De ûntwikkeling fan geslachtsferskillen yn 'e locus coeruleus fan' e rôt. Breinûndersyk Bulletin. 2001;56: 73-8. [PubMed]
- Paus HG, Jr, Katz DL. Homicide en near-homicide troch anaboleus steroïde brûkers. [Sjoch kommentaar] Journal of Clinical Psychiatry. 1990;51: 28-31. [PubMed]
- Pope HG, Jr, Katz DL. Psychiatryske en medyske effekten fan anaboleary-androgenyske steroide gebrûk. In kontrolearre stúdzje fan 160-atleten. Argyf fan 'e Algemiene Psychiatry. 1994;51: 375-82. [PubMed]
- Paus HG, Jr, Kouri EM, Powell KF, Campbell C, Katz DL. Anabolearysk-androgenysk steroide gebrûk ûnder 133-finzenen. Yntegrale psychiatry. 1996;37: 322-7. [PubMed]
- Putnam SK, Du J, Sato S, Hull EM. Testosteron restauraasje fan copulatory gedrach korreleert mei mediale preoptyske dopamine frijlitting yn kastreare manlike ratten. Hormonen en gedrach. 2001;39: 216-224. [PubMed]
- Putnam SK, Sato S, Hull EM. Effekten fan testosteronmetaboliten op kopulaasje en mediale preoptyske dopamine-frijlitting yn kastreare manlike ratten. Hormonen en gedrach. 2003;44: 419-26. [PubMed]
- Putnam SK, Sato S, Riolo JV, Hull EM. Effekten fan testosteron-metaboliteën op kopulaasje, mediale preoptyske dopamine, en NOS-immunoreaktiviteit yn kastateleare manlike rotten. Hormonen en gedrach. 2005;47: 513-522. [PubMed]
- Rankin SL, Partlow GD, McCurdy RD, Giles ED, Fisher KR. Postnatale neurogenese yn 'e vasopressine en oxytocine-befette kearn fan' e pighypothalamus. Brain Research. 2003;971: 189-96. [PubMed]
- Ricci LA, Rasakham K, Grimes JM, Melloni RH., Jr Serotonin-1A-receptoraktiviteit en ekspresje modulearje adolesinte anabole / androgenyske steroïde-induzeare agresje yn hamsters. Farmakology, biogemy en gedrach. 2006;85: 1-11.
- Romeo RD, Cook-Wiens E, Richardson HN, Sisk CL. Dihydrotestosteron aktiveart seksueel gedrach yn folwoeksen manlike hamsters, mar net yn jeugd. Fysiology en gedrach. 2001;73: 579-84. [PubMed]
- Romeo RD, Diedrich SL, Sisk CL. Estrogeenreceptorimmunoreaktiviteit yn prepubertale en folwoeksen manlike Syryske hamsters. Neuroscience Letters. 1999;265: 167-70. [PubMed]
- Romeo RD, Parfitt DB, Richardson HN, Sisk CL. Feromonen lokke lykweardige nivo's fan Fos-immunoreaktiviteit yn prepubertale en folwoeksen manlike Syryske hamsters. Hormonen en gedrach. 1998;34: 48-55. [PubMed]
- Romeo RD, Richardson HN, Sisk CL. Pubryt en de ferfeling fan 'e manlike harsens en seksueel gedrach: it werjaan fan in gedrachspotential. Neuroscience & Biobehavioral Reviews. 2002a;26: 381-91. [PubMed]
- Romeo RD, Wagner CK, Jansen HT, Diedrich SL, Sisk CL. Estradiol induceert hypothalamyske progesteronreceptors, mar aktiveart gjin paringgedrach yn manlike hamsters (Mesocricetus auratus) foar puberteit. Behaviorale neuroanens. 2002b;116: 198-205. [PubMed]
- Salas-Ramirez KY, Montalto PR, Sisk CL. Anabole androgenyske steroïden (AAS) beynfloedzje differinsjaal sosjaal gedrach yn adolesinte en folwoeksen manlike Syryske hamsters. Hormonen en gedrach. 2008 yn 'e parse.
- SAMHSA / OAS. DHHS-publikaasje nr (SMA) 1996. 1994 Nasjonaal húshâldlik ûndersyk nei drugsmisbrûk, haadfynsten 1994; pp. 96 – 3085.
- SAMHSA / OAS. Resultaten fan 'e 2004 National Survey on Drug Use and Health: National Findings. 2005. NSDUH Series H-28, DHHS Publikaasje No. SMA 05 – 4062.
- Sato SM, Johansen J, Jordan CL, Wood RI. Androgen selsbestjoer yn Tfm-ratten. 10e jierlikse gearkomste fan maatskippij foar gedrachsneuroendokrinology.2006.
- Sato SM, Hout RI. Selsadministraasje fan membraan-impermeabele anabole-androgenyske steroïden (aas) yn Syryske hamsters. 11e jiergearkomste fan maatskippij fof Behavioral Neuroendocrinology.2007.
- Schroeder JP, Packard MG. Rol fan dypamine-reseptortypes by it oankeapjen fan in testosteron betingst pleats foarkar by rotten. Neuroscience Letters. 2000;282: 17-20. [PubMed]
- Schulte HM, Hall MJ, Boyer M. Húslik geweld ferbûn mei anabole steroïde misbrûk. American Journal of Psychiatry. 1993;150: 348. [PubMed]
- Schulz KM, Menard TA, Smith DA, Albers HE, Sisk CL. Testikulêre hormoaneksposysje tidens adolesinsje organiseart flankmarkearjend gedrach en vasopressine-reseptorbining yn it laterale septum. Hormonen en gedrach. 2006;50: 477-83. [PubMed]
- Schulz KM, Richardson HN, Romeo RD, Morris JA, Lookingland KJ, Sisk CL. Mediale preoptyske gebiet dopaminergyske antwurden op froulike feromonen ûntwikkelje tidens puberteit yn 'e manlike Syryske hamster. Brain Research. 2003;988: 139-45. [PubMed]
- Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk CL. Gonadale hormonen masculinisearje en ferdigenje reproduktyf gedrach tidens puberteit yn 'e manlike Syryske hamster. Hormonen en gedrach. 2004;45: 242-9. [PubMed]
- Schulz KM, Sisk CL. Pubertale hormonen, it adolesinte harsens, en de feriening fan sosjale gedrach: lesen fan 'e Syryske hamster. Molekulêre & sellulêre endokrinology. 2006:254–255. 120–6.
- Schulz KM, Zehr JL, Salas-Ramirez KY, Sisk CL. Neuroscience Meeting Planner. Sandiego, CA: Maatskippij foar neurowetenskippen; 2007. Is adolesinsje in twadde gefoelige perioade foar it organisearjen fan effekten fan testosteron op folwoeksen manlik reproduktyf gedrach? 2007 Online.
- Scott JP, Stewart JM, De Ghett VJ. Krityske perioaden yn 'e organisaasje fan systemen. Untwikkelbedriuw Psychobiology. 1974;7: 489-513. [PubMed]
- Shughrue PJ, Lane MV, Merchenthaler I. Fergelikende ferdieling fan estrogeenreceptor-alfa en -beta mRNA yn it rat sintraal nervosysteem. Journal of Comparative Neurology. 1997;388: 507-25. [PubMed]
- Simons-Morton BG, Haynie DL. Psychososjale foarsizzers fan ferhege poadium foar smoken ûnder sechde klassers. American Journal of Health Behavior. 2003;27: 592-602. [PubMed]
- Sisk CL, Zehr JL. Pubertale hormonen organisearje it adolesinte harsens en gedrach. Frontiers yn neuroendokrinyology. 2005;26: 163-74. [PubMed]
- Sowell ER, Thompson PM, Leonard CM, Welcome SE, Kan E, Toga AW. Longitudinale mapping fan kortikale dikte en harsensgroei by normale bern. Journal of Neuroscience. 2004;24: 8223-31. [PubMed]
- Sowell ER, Thompson PM, Tessner KD, Toga AW. Mapping trochgeande brainwachtwikkeling en griente middensdichtsreduksje yn dorsale frontale cortex: Inverse relaasjes by postadolesintele harsekultuering. Journal of Neuroscience. 2001;21: 8819-29. [PubMed]
- Spear LP. De adolesinte hynder en leeftydsferoaringsbehearskings. Neuroscience & Biobehavioral Review. 2000;24: 417-63.
- Su TP, Pagliaro M, Schmidt PJ, Pickar D, Wolkowitz O, Rubinow DR. Neuropsykiatryske effekten fan anabole steroïden by manlike normale frijwilligers. JAMA. 1993;269: 2760-4. [PubMed]
- Thiblin I, Finn A, Ross SB, Stenfors C. Fergrutte dopaminergyske en 5-hydroxytryptaminergyske aktiviteiten yn manlike rattenbrein nei lange termyn behanneling mei anabole androgenyske steroïden. Britsk Journal of Pharmacology. 1999;126: 1301-6. [PMC fergees artikel] [PubMed]
- Thomas P, Dressing G, Pang Y, Berg H, Tubbs C, Benninghoff A, Doughty K. Progestin, estrogen en androgen G-protein-gekoppelte reptors yn fisk gonaden. Steroïden. 2006;71: 310-6. [PubMed]
- Tricker R, Casaburi R, Storer TW, Clevenger B, Berman N, Shirazi A, Bhasin S. De effekten fan suprafysiologyske doses testosteron op lilk gedrach yn sûne eugonadale manlju - in klinysk ûndersykssintrumstúdzje. Journal of Clinical Endokrinology en metabolisme. 1996;81: 3754-8. [PubMed]
- Triemstra JL, Nagatani S, Wood RI. Chemosensoryske signalen binne essensjeel foar paring-induzearre dopamine-frijlitting yn MPOA fan manlike Syryske hamsters. Neuropsychopharmakology. 2005;30: 1436-42. [PubMed]
- Triemstra JL, Sato SM, Houten RI. Testosteron en nucleus accumbens dopamine yn de manlike Syryske hamster. Psychoneuroendokrinology (yn parse)
- Van Eenoo P, Delbeke FT. De prevalens fan doping yn Flaanderen yn fergeliking mei de prevalens fan doping yn ynternasjonale sporten. Ynternasjonaal Journal of Sports Medicine. 2003;24: 565-570. [PubMed]
- WADA. Ungeduldige analysearjende befinings berjochten troch akkreditearre laboratoaren. 2006. http://www.wada-ama.org.
- Wesson DW, McGinnis MY. Steapelje anabole androgenyske steroïden (AAS) tidens puberteit by rotten: in neuroendokrine en gedrachsoardieling. Farmakology, biogemy en gedrach. 2006;83: 410-9.
- Wichstrom L, Pedersen W. Gebrûk fan anabole-androgenyske steroïden yn folwoeksenens: winne, goed útsjen of min wêze? Journal of Studies oer Alkohol. 2001;62: 5-13. [PubMed]
- Wood RI, Johnson LR, Chu L, Schad C, Sels DW. Testosteron-fersterking: yntravenous en yntrakebrovintrikulêre selsbehear yn manlike ratten en hamsters. Psychopharmacology. 2004;171: 298-305. [PubMed]
- Wood RI, Newman SW. Androgeen en estrogeen-reseptors libje tegearre mei yn yndividuele neuronen yn it harsens fan 'e Syryske hamster. Neuroendokrinology. 1995;62: 487-97. [PubMed]
- Hout RI, Swann JM. Neuronale yntegraasje fan chemosensoryske en hormonale sinjalen dy't manlik seksueel gedrach kontrolearje. Yn: Wallen K, Schneider JS, redakteuren. Reproduksje yn kontekst. MIT Druk; Cambridge: 1999. pp. 423 – 444.
- Zahm DS, Heimer L. Twa transpallidale paden dy't ûntsteane yn 'e rat nucleus accumbens. Journal of Comparative Neurology. 1990;302: 437-46. [PubMed]